Consideracions generals
Encara que sabem molt poca cosa de la vida primitiva sobre la Terra, i que moltes de les suposades evidències de vida poden ser interpretacions massa generoses d’unes dades molt minses, podem plantejar-nos-en l’estudi des de tres perspectives diverses. D’una banda, intentar d’inventariar i discutir críticament les troballes de fòssils a l’Arqueà (3800 a 2500 milions d’anys), que és l’eonotema (o divisió cronostratigràfica) més antic de què disposem com a registre rocós. En segon lloc, fer especulacions, basant-nos en els coneixements científics actuals, sobre el possible camí que ha seguit la natura per arribar a produir aquesta estranya realitat que són els éssers vius. Finalment, provar d’analitzar els condicionaments geològics de l’aparició de la vida i dels seus primers passos i narrar la història primitiva conjunta de la biosfera i dels embolcalls més superficials de la Terra on aquesta biosfera neix i es desenvolupa. La fusió d’aquestes tres perspectives en una sola i única història no és possible avui. Ni el lent, encara que ferm, avanç de la ciència en aquest camp no permet de ser gaire optimistes respecte d’aquest acompliment. Una bona dosi d’esperit crític i, àdhuc, d’escepticisme són saludables en referir-nos a aquests temes i a les conclusions temptatives dels especialistes.
Els fòssils arqueans

Esquema en forma d’espiral ascendent de les fases més antigues de la història de la vida des de l’inici de la història de la Terra.
Distrimapas, original de P. Cloud 1983.
Les roques arqueanes són molt avares en indicis que permetin de pensar en l’existència de la vida sobre la Terra; aquests indicis, però, hi són. Cal veure’n el tipus i la significació. L’estudi de les roques arqueanes fa pensar que l’atmosfera primitiva contenia molt poc oxigen lliure i que durant l’Arqueà aquest contingut anà augmentant considerablement. Deixant de banda una possible producció, relativament important, d’oxigen causada per una radiació ultraviolada molt intensa (encara que no acceptada per tothom), que s’hauria produït durant un període relativament curt dels primers temps de la història de la Terra, la font més important d’oxigen lliure és d’origen orgànic per fotosíntesi. L’evolució del contingut d’oxigen seria, doncs, una prova de l’existència d’éssers vius.
L’anàlisi de la matèria orgànica trobada en els sediments arqueans fa pensar que aquesta és fruit, si més no quasi tota, de l’activitat d’organismes vius. Sembla que una de les proves més suggerents respecte a l’existència d’éssers vius (extreta de les dades geoquímiques de què disposem) ens la dóna la composició isotópica del querogen en els sediments arqueans no metamorfitzats. Sembla clar que el material carbonàtic trobat en aquests sediments és, pel cap baix, el resultat de l’activitat biològica. Aquest material trobat en unes roques, que atenyen en certs casos els 3300 milions d’anys d’antiguitat, deriva d’organismes anaeròbics microscòpics procariotes.
Hom ha trobat a les roques arqueanes varietats diferents d’objectes que semblen microfòssils. Les varietats descrites deuen correspondre a unes formes semblants als bacteris, filiformes i esferoïdals. Una anàlisi crítica i acurada fa veure que és difícil d’assegurar la vertadera natura d’aquests fòssils hipotètics i que, sovint, anomenar-los així és, si més no, agosarat. No obstant això, el que cal estudiar amb atenció són les restes trobades en el registre rocós arqueà (quan no hi ha hagut cap contaminació) i cal, també, enregistrar el lloc i l’antiguitat dels materials que les contenen i les seves característiques, per tal d’aconseguir desxifrar-ne el significat. Cal dir en aquest punt que, de fet, hom ha trobat abundants formes esferoïdals, ja d’abans dels 3000 milions d’anys; el problema està en identificarne la natura.
Pel que fa a les formes de grandària superior, tenim unes estructures estromatolítiques d’uns 3500 milions d’anys d’antiguitat, procedents de Rhodèsia (Zimbabwe) i, també, unes de posteriors, de l’Arqueà canadenc. Aquestes estructures són compostes per columnes verticals atapeïdes i per capes hemisfèriques amuntegades. El material d’aquestes capes o làmines és calcari i orgànic. L’estructura laminar d’aquestes formacions arqueanes és indistingible dels estromatòlits posteriors, tant fòssils com actuals, dels quals es coneix l’origen, que no és altre que l’activitat de comunitats de microorganismes fotosintetitzadors. No sembla pas que cap procés sedimentan inorgànic hagi pogut produir aquestes estructures, i per això sembla ben clar que els estromatòlits arqueans són una prova de l’existència de microorganismes, sense que se’n conegui la veritable natura.
En conjunt, doncs, la vida de l’Arqueà ens és fonamentalment desconeguda, però el que sí que sembla acreditable és la seva existència i també que, en tot cas, deuria ser representada per uns microorganismes d’estructura molt senzilla i proveïts d’uns elements fotosintetitzadors, almenys molts d’ells. Aquestes conclusions s’adiuen amb la teoria de l’evolució orgànica, segons la qual l’aparició dels organismes es deuria produir, en el temps, tot passant de formes simples a formes cada cop més complexes.
L’origen de la vida
Com es pot produir la vida? Si coneguéssim perfectament la resposta d’aquesta pregunta, ens seria més fàcil de comprendre de quina manera s’ha produït realment la vida sobre la Terra. No és ben bé així. Desconeixem molts aspectes d’aquesta resposta i, per consegüent, no ens és gens fàcil d’imaginar l’origen de la vida.
Les primeres hipòtesis

Etapes hipotètiques de l’evolució cap a la vida a través de les fases evolutives nuclear, molecular, protobiològica i, pròpiament, biològica.
Distrimapas, original de J. Oró.
Fa temps que hom treballa sobre la hipòtesi de l’existència primera d’una seqüència bioquímica que devia desembocar a l’aparició dels primers organismes, és a dir, a l’inici de l’evolució orgànica. Així, hom pensa en una successió en el temps que, partint de l’atmosfera i la hidrosfera més antigues, portà a la formació de molècules orgàniques, primer molt simples i després extraordinàriament complexes. A partir d’aquí, i mitjançant un procés no gens simple, es devia arribar a la vida. Podem pensar que tot això forma part d’una evolució general o de desenvolupament negentròpic de la matèria que va de la formació de la molècula més simple, la de l’hidrogen, a la vida. Hom entén per procés negentròpic aquell que produeix un augment d’ordre i que, per tant, és de caràcter netament oposat a la majoria dels processos que observem, que són productors de desordre. Són processos ordenadors, al costat del de la producció i l’evolució biològica, el de la formació i l’evolució estel·lars i el de la producció dels elements químics.
Centrem l’atenció en les etapes que s’haurien d’acomplir en una seqüència cap a la vida. Podem resumir-les en les següents: en una primera etapa, hi hauria la producció de molècules orgàniques, relativament complexes, tals com els aminoàcids i els polipèptids, a partir de molècules senzilles (CH4, H2O, H2, NH3...); en una segona etapa, trobaríem la formació dels polímers, constituents actualment de totes les formes de vida, tals com l’ADN i l’ARN; en la tercera etapa, podem pensar en l’aparició de l’autoduplicació d’aquests polímers i, per consegüent, en el començament de l’existència d’aquest atribut fonamental de la vida, que s’associa a la mutació i a l’evolució al llarg de generacions successives; situaríem en la quarta etapa l’aparició de la cèl·lula; i en la cinquena, l’aparició d’organismes pluricel·lulars.

Esquema de l’aparell i del funcionament de l’experiment de S.L. Miller i H.C. Urey creat per reproduir, dins de les condicions probablement existents en l’atmosfera primitiva, la formació de molècules relacionades amb processos vitals. Per la clau de l’esquerra del dibuix s’introdueixen diverses mescles gasoses, semblants a les que se suposa que hi havia a l’atmosfera primitiva. En el matràs es bull l’aigua per aconseguir la conducció de gasos per tot l’aparell i, en concret, fins a la cambra on se sotmeten a una descàrrega elèctrica. Els productes finals s’acumulen al fons de l’aparell, des d’on s’extreuen dissolts. Els productes obtinguts, que tenen relació amb els processos vitals, es troben a la taula, amb indicació dels resultats quantitatius a partir de 59 000 micromols de carboni en forma de metà. 1 Elèctrode de tungstè, 2 matràs de 5 l, 3 condensador, 4 clau per a extreure mostres durant l’experiment, 5 matràs de 50 cc.
Distrimapas, original de Dickerson.
Miller, al principi dels anys cinquanta produí molècules orgàniques complexes, entre les quals hi havia quatre aminoàcids, a còpia de provocar descàrregues elèctriques sobre una barreja de molècules senzilles en un ambient no oxigenat. Més tard, cap als anys seixanta, Oró va demostrar que les molècules de l’àcid cianhídric podien passar a formar l’adenina. Aquests experiments permeten d’entendre com pot esdevenir-se la primera de les etapes esmentades. D’altra banda, el registre fòssil ens fa assistir a una visió fragmentària de l’aparició dels éssers multicel·lulars després, en el temps, de tenir evidències de l’existència dels unicel·lulars. I, també, el fet que els diversos éssers vivents actuals puguin ser arranjats en una jerarquia de complexitat progressiva que ens fa adonar del pas de l’estructura unicel·lular a la pluricel·lular.
Els problemes encara no resolts es refereixen a les etapes segona, tercera i quarta, que possiblement podrien ser formulades diversament. D’una banda, l’etapa segona i la tercera podrien coincidir amb la realitat i, de l’altra, caldria subdividir la quarta en diverses subetapes, ja que per a atènyer la complexitat de les cèl·lules més elementals actuals hi deuria haver uns passos intermedis que haurien requerit un cúmul de circumstàncies no gens fàcils de produir-se. Sense entrar ja en els darrers capítols de la formació de la cèl·lula actual, l’arribada als vertaders organismes vius, encara que molt primitius, és un gran desafiament per a la ciència.
Dins de la hipòtesi en què ens hem situat, l’experimentació de laboratori ha fracassat i les especulacions continuen apareixent sobre l’ambient possible on aquestes etapes s’haurien pogut produir. El primer model proposat fou el d’Oparin (1924), un científic rus que, ja cap als anys vint, suggerí els passos essencials dels condicionaments de l’aparició de la vida. La major part d’autors s’han adherit als seus punts de vista o a d’altres de semblants. Podríem dir que es tracta de postular l’existència d’una bona colla de substàncies inorgàniques i orgàniques en un ambient reductor, la barreja i interacció de les quals hauria permès l’avanç assenyalat de les etapes esmentades. Hom parla sovint del «brou» primordial per indicar uns acúmuls extraordinaris de molècules orgàniques que farien més planer i més possible aquest mateix avanç. Dins aquesta línia hom ha comprovat l’existència d’un gran nombre d’espècies químiques orgàniques en l’espai interstel·lar, en els cometes i en els meteorits carbonàtics. Per consegüent, hom pensa que la Terra primitiva contenia unes grans quantitats de matèria orgànica sovint en molècules força complexes, idèntiques a les que ara existeixen en els éssers vius. Com s’ha dit, no sense ironia, la Terra primitiva, abans de la vida, estava més contaminada que la major part dels centres urbans actuals.
Les hipòtesis modernes
Tots aquests fets que han aportat les investigacions astronòmiques modernes han induït a pensar en un origen extraterrestre de la vida, opinió que no sembla pas gaire defensable, deixant de banda que no fa res més que dur més enllà en l’espai, i potser en el temps, l’origen de la vida. En tot cas, com ja hem dit, la qüestió està en la manera d’aconseguir el pas de les molècules complexes als polímers, amb la propietat de reduplicació, tal com s’esdevé en els éssers vius actuals. El problema principal sempre és que caldria que es produïssin en prou quantitat perquè una part sobrevisqués i fos l’inici de les molècules fonamentals de la vida i, d’això, arribar a les primeres formes d’un organisme vivent cel·lular o «pre-cel·lular». La salvaguarda d’aquestes primeres llavors de vida és difícil. Els raigs ultraviolats les destrueixen si es formen a la superfície de l’aigua. A dins, malgrat que es troben protegides d’aquests raigs, també tenen dificultats de supervivència. Només un subministrament constant d’aminoàcids en llacunes d’aigua variable, o l’adherència d’aquestes molècules a determinades argiles o alguna altra situació peregrina i excepcional permeten d’imaginar el procés.
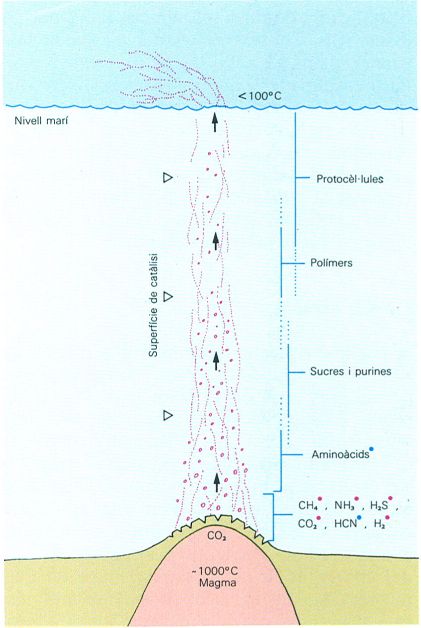
Model hipotètic d’un sistema hidrotermal que funcionaria com un flux continu de síntesi abiòtica de molècules orgàniques complexes i probionts. A la figura veiem de quina manera, en diferents llocs de la columna, ocorren fets que fan possible aquesta síntesi abiòtica. El subministrament d’energia calorífica i de CO2 té lloc a la base de l’àrea anomenada de clivellament. Més amunt es produeixen metalls i altres components deguts a condicions reductores en reaccions aigua-roca, mentre que els fluids ascendeixen per un sistema anastomòtic de fractures amb superfícies catalítiques, donant estructures més complexes i, ja en contacte amb la part inferior de la capa aquosa, sediments químics. La confirmació, almenys parcial, de la hipòtesi es dóna en els productes trobats en els sistemes hidrotermals actuals (compostos assenyalats amb un punt blau) i en les salmorres de la mar Roja (compostos assenyalats amb un punt vermell).
Jordi Corbera, original de J.B. Corliss, J.A. Baross i S.E. Hoffman.
Modernament, s’ha descobert el fet que les fonts hidrotermals submarines que funcionen sovint a les dorsals oceàniques constitueixen una àrea d’una barreja complexa de roques, gasos, calor i aigua que dóna lloc a una producció de vida exuberant amb unes comunitats bacteriològiques quimiosintètiques, i també a comunitats animals amb la presència de formacions elementals de vida. Això ha fet que hom hagi comparat els diversos elements d’aquestes fonts hidrotermals (roques ígnies i metamòrfiques, sediments químics i compostos orgànics) amb les successions de l’Arqueà, les quals presenten notables convergències en aquests tres tipus d’elements i on s’han detectat els «fòssils» més antics. Si s’acceptava el valor probatori d’aquestes analogies, hom podria pensar que l’ambient d’aquestes surgències calentes submarines amb llur flux continuat (que dóna diverses situacions físico-químiques al llarg del seu ascens) seria l’ideal perquè s’aconseguís el pas de l’estadi de molècules orgàniques complexes al de polímers reduplicables i a organismes vius.
La manca de teories o de models plenament satisfactoris ha portat a l’obertura d’unes noves vies per a explicar l’origen de la vida. Ja hem vist com l’atenció se centra en les molècules inorgàniques, con són les argiles per exemple, per a pensar que, d’una manera o altra, estan implicades en les primeres etapes de la història de la vida. En alguns treballs recents, hom fa passar uns tipus determinats de cristalls inorgànics de simples materials de protecció o ajut de les molècules orgàniques prebiològiques a protagonistes de la història primitiva de la vida. Caldria pensar, en aquest cas, que fou una seqüència geoquímica i no bioquímica allò que devia portar a l’existència dels primers organismes capaços d’evolucionar per selecció natural. Aquests primers organismes «minerals» haurien estat substituïts per uns organismes «orgànics», mitjançant l’addició de molècules orgàniques contingudes al «brou» primordial, ja que les estructures orgàniques comporten un augment de la complexitat que és inconcebible en els organismes «minerals», els quals, pròpiament parlant, cal considerar que encara no eren organismes vius. Aquesta hipòtesi és molt suggestiva perquè permet de plantejar el mateix problema des d’una perspectiva molt diferent; no es pot preveure, però, si serà el començ d’una nova visió de la història primitiva de la vida.
Fins avui, l’intent de lligar el pensament biològic, sovint molt especulatiu amb les dades deduïbles del registre rocós segueix les vies d’una seqüència bioquímica-biològica; encara que potser la narració de la història de la vida no canviaria gaire amb qualsevol altra hipòtesi.
La vida al Precambrià
La vida que coneixem, i de la qual pretenem explicar l’origen i l’evolució primitiva, es desenvolupa, en relació inextricable, amb l’atmosfera i la hidrosfera, amb l’aire i l’aigua. Per consegüent, el coneixement de l’atmosfera i la hidrosfera existents a l’Arqueà és essencial per a comprendre com hauria pogut aparèixer i perdurar la vida. D’altra banda, el coneixement de les propietats, la composició i les característiques de l’atmosfera i la hidrosfera més antigues depèn dels resultats de l’anàlisi del registre rocós.
La vida anaeròbia
Diguem, de bell antuvi, que les substàncies gasoses o fàcilment gasificables, que formen la part més gran dels elements de l’atmosfera i la hidrosfera, representen un excés dels «volàtils» que es troben a la Terra, i són el producte de l’exsudació de les parts superficials de la part sòlida del planeta. Les erupcions volcàniques són una mostra entenedora de com uns líquids i uns gasos traspuen de l’escorça terrestre i de la part superior del mantell i s’incorporen als embolcalls lleugers del nostre planeta. L’anàlisi de llurs components ens porta a pensar en una atmosfera sense gens d’oxigen lliure, ja que aquest element no es troba mai entre els productes volcànics. Sembla evident, doncs, que l’atmosfera més antiga no contenia gens d’oxigen lliure, sinó que era una barreja de gasos parcialment reduïts i oxidats, fruit de les erupcions volcàniques (o qualsevol altre tipus d’emissió eixida de la part sòlida de la Terra) i de l’impacte meteorític o col·lisió, de cossos celests amb la Terra. Aquest darrer fenomen devia ser particularment important durant els primers temps de la història de la Terra, abans de l’Arqueà. Entre aquests volàtils hi havia quantitats notables de les substàncies orgàniques o cúmuls als quals ens hem referit en el paràgraf precedent.
Així doncs, la primera fase de la vida sobre la Terra es desenvolupà en una atmosfera sense oxigen, o bé, de segur, amb una quantia escassíssima, amb una concentració per sota de l’u per cent de l’actual. Aquesta concentració no s’assoleix sinó molt més endavant, cap als 2000 milions d’anys. Sabem que l’oxigen lliure pot ser un producte de la fotodissociació de compostos que tenen l’oxigen en combinació, com és ara l’aigua i l’anhídrid carbònic. La reacció és reversible i no permet grans acumulacions, cosa que només l’activitat dels organismes fotosintetitzadors sembla explicar. Per consegüent, entre els primers éssers vius hi devia haver una gran quantitat d’organismes anaerobis fotosintetitzadors, dotats d’unes estructures molt simples. Les cianofícies actuals, encara que sovint són anomenades impròpiament algues blaves, també poden servir com a model d’aquests éssers vius primitius de l’Arqueà. Tots els «fòssils» arqueans podrien correspondre a uns organismes d’aquest tipus incloent-hi, evidentment, els responsables de la formació d’estromatòlits. Siguin o no siguin autòtrofs, es tracta d’éssers unicel·lulars procariotes, és a dir, sense una membrana nuclear, a l’estil dels bacteris i de les cianofícies.
L’enriquiment progressiu en oxigen lliure de l’atmosfera fou la causa d’unes crisis biològiques fàcilment explicables, ja que exigia uns organismes més capacitats per tal de no ser eliminats per un element tan actiu. Sembla que, en una fase de la història de la Terra, l’excés d’oxigen produït pels éssers vius, abans d’assolir una transformació adequada en què llur quimisme els permetés aprofitarlo, fou un obstacle important per al desenvolupament de la vida, llevat on la presència de substàncies oxidables eliminava aquest excés d’oxigen lliure. La presència de grans quantitats de sediments ferrífers, les anomenades BIF («banded iron formations»), o formacions de ferro en bandes, o franges, que es troben exclusivament en el Precambrià i sobretot en l’interval dels 2600 als 1800 milions d’anys, podrien haver estat un dels «desguassos» importants de l’eliminació d’aquest excés d’oxigen. Una altra part d’aquest oxigen es convertia a les capes altes de l’atmosfera en ozó (O3) tot constituint el pàmpol d’ozó que filtra l’excés de raigs ultraviolats letals per als éssers vius. Tot això ens fa pensar que la vida primerament s’establí en petits estanys d’aigües encalmades i que els primers éssers vius no vivien a la superfície per tal de no ser occits pels raigs solars enfortits amb la llum ultraviolada, però que tampoc no vivien a gaire profunditat, ja que necessitaven la llum per a realitzar la fotosíntesi. D’altra banda, la vida esdevingué més esponerosa allà on l’oxigen lliure en excés podia ser eliminat.
La vida aeròbia

Esquema dels processos i productes esdevinguts a causa de la interacció entre l’atmosfera i la hidrosfera fa 2000 milions d’anys aproximadament, amb formació de la capa d’ozó, d’estromatòlits i de les anomenades formacions de ferro en bandes.
Jordi Corbera, original de J. W. Schopf modificat per l’autor.
A partir d’aquesta vida anaeròbia calgué arribar a les transformacions biològiques que permeteren el desenvolupament de la vida aeròbia. Assolim, doncs, l’etapa oxigenada de l’atmosfera i de la vida. El registre rocós ens fa veure l’aparició d’uns sediments vermellosos («red beds»), pels volts dels 2000 als 1700 milions d’anys, que suposen una atmosfera oxigenada. És, doncs, en aquest moment de la història de la Terra que ja s’arriba a un 1 % del contingut actual d’oxigen lliure en l’atmosfera, segons el càlcul clàssic més minimista, i a percentatges de gairebé el 50 %, o més, en alguns dels plantejaments moderns. Sigui com vulgui, l’aparició d’organismes que aconsegueixen de respirar és una nova fita en el camí d’una vida més rica i amb més possibilitats evolutives. Sembla que l’aparició de la respiració en el món procariota comportà ràpidament l’aparició d’unes especialitzacions morfològiques com són els orgànuls de locomoció o de fixació.
No gaire després, i ja en aquesta atmosfera oxigènica, degué aparèixer la cèl·lula amb nucli o eucariota. El registre fòssil és tan pobre i imperfecte que fa difícil de saber exactament quan i on es produïren els primers eucariotes.
En analitzar els microfòssils hom considera que poden ser eucariotes aquells que demostren una particular complexitat morfològica i una particular grandària. Un dels autors actuals, Schopf, esmenta nou troballes de suposats organismes ecuariòtics en terrenys que oscil·len entre els 1500 i els 1600 milions d’anys. Altres autors confirmen aquesta data dels volts dels 1500 milions d’anys com la de l’aparició de la cèl·lula eucariota.

Fòssils del precambrià (A-E) i del cambrià (F-J). Els del Precambrià, trobats per primera vegada a Austràlia i després reconeguts en materials de la mateixa època (Ediacarià) en altres continents, eren animals de cos tou, sense esquelet, alguns semblants a cnidaris, com A (que recorda els pennatulacis actuals) i E (de tipus medusoide); a anèl·lids com C (Spriggina); o bé tenien formes no homologables a cap animal conegut, com B (Parvacorina) i D (Tribrachidium). Els del Cambrià tenien exosquelet; F (Opabinia), I (Hallucigenia) i J (Dinomischus) no tenen homòlegs entre els animals actuals, mentre que H (Avsheaia) recorda molt els onicòfors; de G (Pikaia), hom pensa que pot representar els primers cordats.
Jordi Corbera, originals de P.J. Wynne.
El pas subsegüent de l’evolució biològica fou l’aparició de la sexualitat, que, en l’àmbit cel·lular, suposa el procés, anomenat meiosi, de bipartició dels cromosomes. És impossible de trobar en el registre fòssil els arguments segurs del moment en què això s’esdevingué; n’hi ha prou, però, per afirmar que a partir dels 1400 milions d’anys es produeix un augment de la diversitat dels microfòssils, que sembla que provoca l’aparició de la reproducció sexual garantidora, en tot cas, de la variabilitat dels organismes. Així doncs, entre aquesta data i la de l’aparició dels primers fòssils clars d’organismes megascòpics pluricel·lulars (que es produeix vers el final del Precambrià), és quan cal situar els primers organismes sexuals; no més ençà dels 700 milions d’anys, segons les conjectures que són més plausibles.
Acabem de parlar dels primers organismes pluricel·lulars del registre fòssil. Amb això suposem que hi hagué una nova fita en la història de la vida: el pas d’estructures unicel·lulars a éssers vius, amb un nombre elevat de cèl·lules organitzades en teixits i òrgans. Atesa la inexistència de les parts dures, la possibilitat de llur fossilització fou molt minsa. Malgrat tot, ja cap a l’any 1982, hom comptava més de deu indrets on s’havien trobat uns organismes pluricel·lulars fòssils pertanyents a un període de temps comprès entre els 670 i els 550 milions d’anys. Aquest interval de temps, anterior a l’eclosió de la fauna esquelètica de la base del Cambrià, ha estat anomenat període Ediacarià (nom derivat d’Ediacara Hills, a l’Austràlia meridional), on hom ha trobat una associació de fòssils verament extraordinària. Els organismes són ja impensablement complexos i en gran part identificables amb els fílums existents actualment. A Ediacara hi ha sobretot uns cnidaris de diversos tipus, uns anèl·lids i artròpodes, a més d’organismes de fílums no identificats fins ara. En aquesta àrea han estat reconegudes 26 espècies diferents. Les altres àrees amb fòssils de la mateixa època se situen a diversos llocs on afloren materials antics. Els únics continents on encara hom no ha trobat fauna ediacariana són l’Amèrica del Sud i l’Antàrtida.
A partir, doncs, de l’Ediacarià, i segons l’opinió d’alguns estudiosos, al seu inici hi ha un límit fonamental de la història de la vida i de la Terra. És la frontera entre els temps antics, gairebé desproveïts de fòssils, i l’Eó Fanerozoic (d’animals o vida aparent o manifesta), en què el registre sedimentari es manifesta curull de restes d’organismes i on, malgrat l’ínfima proporció de restes conservades i estudiades, el seguiment del fil de la història de la vida és ja extraordinàriament més fàcil i gratificador. L’aparició de les parts dures o dels esquelets en molts grups d’organismes és un fet que, si bé, d’una banda, falseja la història de la vida (informa molt més dels grups amb esquelet que no pas dels que no en tenen), de l’altra, permet seguir de prop l’evolució biològica i, per això, fer unes generalitzacions ben fonamentades sobre aquesta mateixa evolució o història de la vida.
La vida al Fanerozoic
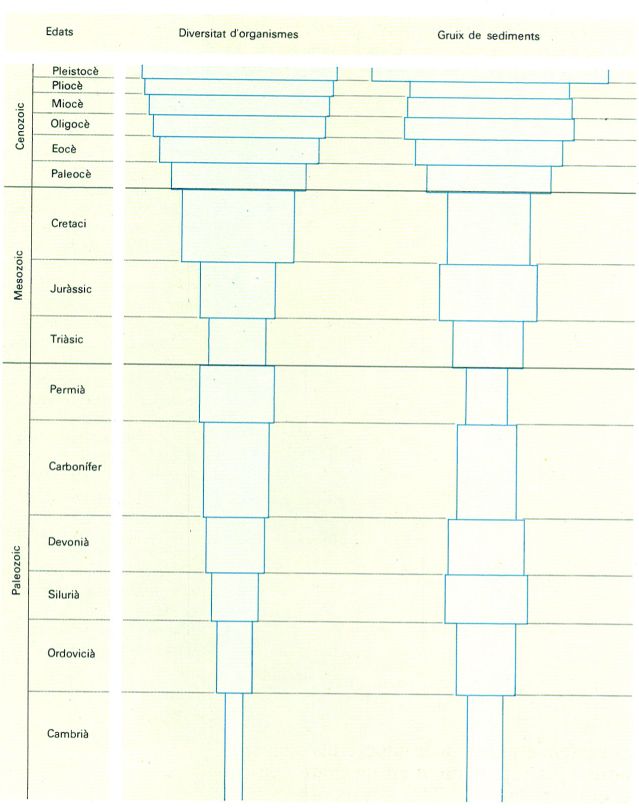
Relació entre diversitat biològica registrada en les roques i gruix de sediments dipositats per milions d’anys.
Distrimapas, original de Raup i S.M. Stanley.
El terme Fanerozoic fou creat conjuntament i en contraposició al de Criptozoic per indicar els temps geològics on les evidències de vida són clares i abundants. En realitat, quan foren creats els dos termes hom no coneixia encara els fòssils del Precambrià; el Criptozoic era, doncs, el temps de vida oculta ja que hom podia postular l’existència de la vida en aquests temps antics només d’acord amb la doctrina de l’evolució, des del punt de vista teòric, i atesos els arguments indirectes basats en el registre rocós. La situació ha canviat des que, el 1930, Chadwick creà els dos termes i donà lloc a un registre fòssil escàs però prou significatiu dins les roques precambrianes.
En tot cas, l’abundància i la varietat del registre fòssil en el Fanerozoic fa possible l’ús d’aquest terme i del seu antònim amb tota propietat. Resta només la petita discussió referent a la necessitat d’incloure-hi l’Ediacarià o, com es va fent clàssicament, de fer començar el Fanerozoic a l’inici del Cambrià.
La història de la vida al Fanerozoic, que hem de dibuixar a continuació amb unes quantes pinzellades, es basa en el registre fòssil, però tenint en compte que aquest no és pas complet i que no es tracta tampoc d’una mostra a l’atzar: una colla de factors geològics i biològics fan que es presenti esbiaixat respecte a la vida real de la qual és una representació.
Els organismes que disposen d’un esquelet consistent són més fàcils de trobar en el registre fòssil que no pas aquells que tenen totes les parts toves i bonament destructibles. També hi ha medis sedimentaris que permeten la conservació, àdhuc, d’éssers tan fràgils com les meduses, mentre que d’altres fan impossible que es conservi cap resta orgànica. D’altra banda, no totes les roques són al nostre abast en afloraments superficials, i àmplies àrees de la Terra no han estat encara explorades adequadament. Per tant, cal dir que desconeixem una gran part del registre fòssil, és a dir, dels organismes les restes dels quals es conserven a les roques i que no han arribat encara, o no podran arribar en un temps llarg en el futur, a les nostres mans.
És des d’aquesta perspectiva, doncs, que intentarem de descriure breument la vida al Fanerozoic i també de comprendre tant les estratègies i els processos generals de la vida en el curs del seu desenvolupament històric, com els condicionaments i les relacions mutus entre el món de les roques i el món de la vida. La història de la vida és inserida essencialment dins la història de la Terra.
L’explosió de la vida al Cambrià inferior
Tal com es manifesta en el registre fòssil, el Cambrià inferior representa l’aparició d’una quantitat considerable de nous tipus d’organismes, i el fet que molts d’aquests presentin parts esquelètiques que en fan fàcil la preservació (i per consegüent, la presència a les roques) constitueix una autèntica explosió en manifestacions de vida sobre la Terra. Una breu enumeració dels tipus d’animals presents a les roques del Cambrià inferior n’és una prova evident.
D’una banda, dins dels artròpodes, els que són fonamentals al Cambrià inferior són els trilobits, però també s’hi troben ostràcodes. Pel que fa als trilobits, n’hi ha més de 25 famílies conegudes, amb més de 200 gèneres al Cambrià inferior, encara que un estudi detallat mostra que l’aparició de tots els grups no és sobtada, sinó progressiva i que, fins i tot, no són presents a la base del Cambrià.
Els arqueociats, un curiós grup d’organismes format gairebé sempre per dos cons porosos encaixats, relacionats entre l’un i l’altre per unes estructures diverses, són exclusius del Cambrià inferior i la seva evolució serveix per a subdividir aquesta sèrie de l’escala geològica; hom n’ha trobat més de 600 espècies (en 140 gèneres aproximadament), en el primer estatge del Cambrià inferior, el Tommotià de Sibèria oriental, on encara no hi ha trilobits.
Els hiòlits són organismes amb closca cònica i d’atribució i afinitats difícils, que hom ha relacionat amb els cucs i també amb els mol·luscs; en tot cas, tenen una importància singular al Cambrià inferior, on n’han estat trobats més de 20 gèneres a partir de la seva mateixa base, i hom els ha emprat àdhuc com a indicadors de medi sedimentari.
Hi ha diversos tipus de mol·luscs a les roques del Cambrià inferior, alguns dels quals són d’afinitat difícil amb els mol·luscs actuals; els més importants són els gasteròpodes, dels quals hi ha més de quatre famílies dividides en uns 15 gèneres, però també en són els monoplacòfors i els bivalves.
Hi ha uns 20 gèneres de braquiòpodes primitius al Cambrià inferior que, si bé són interessants per a l’anàlisi estratigràfica, de moment no han arribat a ser tan útils com els trilobits o els arqueociats.
Malgrat que no ens han pervingut restes fòssils de celenterats al Cambrià inferior atribuïbles als representants fornits d’esquelet, sabem que algunes formes de cos tou de celenterats han deixat empremtes a les roques no sols del Cambrià inferior, sinó també del Precambrià final; d’altra banda, hi ha també força tipus de restes al Cambrià inferior que semblen afins o pertanyents als celenterats. Són molt abundants les espícules d’esponja a les roques del Cambrià inferior i també, excepcionalment, se’n troben d’enteres; sembla clar que pertanyien a dos ordres diferents. Els conodonts, petites restes d’atribució encara avui dubtosa, han estat trobats ja a la base del Cambrià inferior, i llur estudi adquireix una importància creixent gràcies al seu valor estratigràfic.
Empremtes i d’altres restes d’organismes atribuïbles a cucs, aquest grup mal definit d’animals, són presents al Precambrià i al Cambrià inferior. També hi ha restes d’organismes articulats que s’assemblen als anèl·lids o als artròpodes. S’han trobat equinoderms primitius, i hom discuteix la presència al Cambrià inferior d’uns predecessors dels graptòlits i dels briozous, entre d’altres grups. Esdevé també problemàtica la naturalesa de les petites restes que han estat atribuïdes a radiolaris. Finalment, hi ha d’altres elements del registre fòssil dels quals encara no s’ha trobat el lloc entre els fílums reconeguts fins avui.
En conjunt, doncs, la fauna fòssil del Cambrià inferior és notable per la seva varietat. Això suposa per a la història de la vida una colla de possibles formes intermediàries precedents que no han aconseguit romandre enregistrades a les roques amb prou abundància perquè les recerques sistemàtiques de la geologia moderna les hagin trobades. El canvi, en certa manera secundari, de l’adquisició d’un esquelet ha estat determinant per al nostre coneixement de la vida antiga. Mirant cap enrere, des del Cambrià inferior, els nostres coneixements només permeten de fer afirmacions molt generals i sovint amb un fonament fràgil. Per contra, a partir de la base del Cambrià i malgrat que el registre fòssil sigui esbiaixat per les causes que ja hem apuntat més amunt, coneixem amb relativa precisió les línies fonamentals de la història de la vida mitjançant l’aparició, el desenvolupament i (eventualment) l’extinció dels diversos rangs d’organismes, tant d’animals com de vegetals. Per més claredat, descriurem l’evolució dels éssers vius referint-nos per separat als invertebrats, a les plantes i als vertebrats.
L’evolució de la vida a partir del Cambrià inferior
L’evolució dels invertebrats
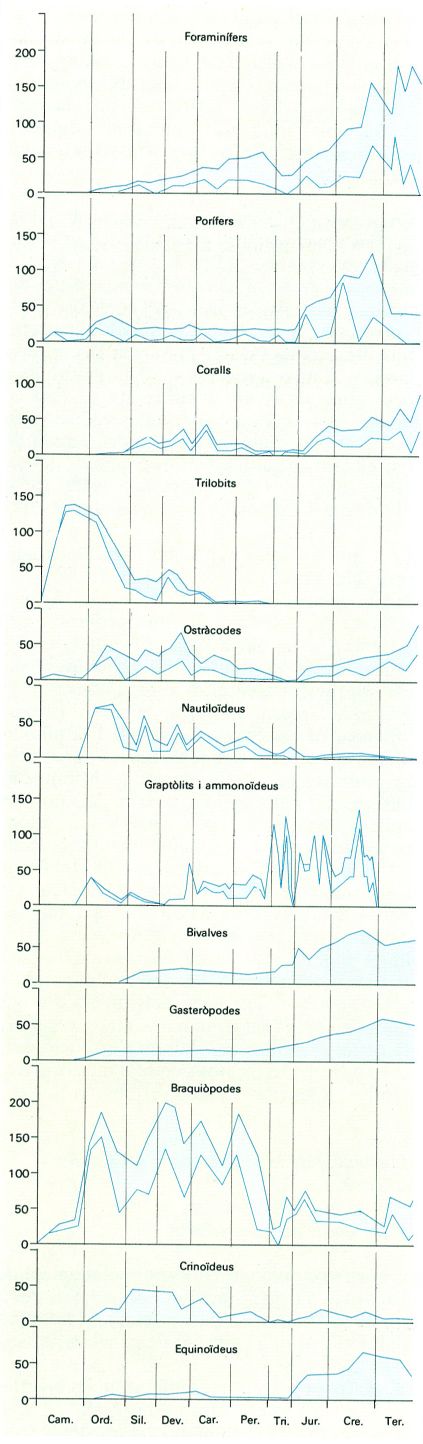
Evolució del nombre de gèneres dels grups més importants d’invertebrats marins a través dels temps geològics. Per a cada grup, la línia superior indica el nombre total de gèneres i la inferior, el nombre de gèneres nous.
Distrimapas, original de House 1967.
L’horitzó cronològic constituït pel moment de l’aparició d’organismes amb esquelet és per a nosaltres l’horitzó a partir del qual farem l’anàlisi global de l’evolució dels invertebrats.
Si agafem els grups més importants d’invertebrats i els representem en un gràfic, en podem veure la variabilitat a través del temps.
En molts casos en el gràfic hi ha una doble corba: la superior representa el total de gèneres existents i la inferior, la de gèneres nous; quan no hi ha la corba inferior això vol dir que no disposem de les dades elaborades respecte del nombre de gèneres nous de cada divisió del temps geològic.
La inevitable simplificació que comporta qualsevol representació gràfica d’aquest tipus no amaga el fet que uns grans grups d’invertebrats han desaparegut totalment i que d’altres han iniciat la seva existència després de l’inici del Fanerozoic. També posa de manifest unes oscil·lacions notables en el nombre de gèneres vivents o en l’aparició de gèneres nous en un determinat moment de la història de la Terra. Els trilobits, els graptòlits i els ammonits, entre altres grups, han desaparegut sense deixar representants en el món actual. Hi ha d’altres grups, com els coralls, en els quals hi ha espècies del Paleozoic que s’han extingit i d’altres que apareixen a partir del Triàsic i que encara formen part de la vida actual. Pel que fa a aquest punt, la gran ruptura entre el Paleozoic i el Mesozoic, és a dir, entre el Permià i el Triàsic, representa un canvi o substitució de tipus a l’interior d’alguns fílums o grans grups d’invertebrats. La majoria dels tipus dels briozous i dels braquiòpodes en són un exemple clar, deixant de banda els coralls, ja esmentats anteriorment. Situacions semblants en rangs inferiors s’esdevenen en distints moments de la història de la Terra; una observació ràpida sobre el registre fòssil d’una roca permet, sovint, a un expert, de saber aproximadament l’edat de la roca sense cap necessitat d’identificar-hi les espècies o els gèneres concrets.
Pel que fa a les oscil·lacions en el nombre dels gèneres vivents o en l’aparició de gèneres nous en un determinat moment de la història de la Terra, cada grup d’invertebrats té la seva pròpia història, encara que hi ha moments particulars de crisi que afecten la majoria dels grups. Per exemple, el nombre màxim de gèneres de trilobits existents al final del Cambrià o el descens ràpid d’aquest nombre en el moment que els ostracodes, els nautiloides, els graptòlits i els braquiòpodes experimenten una puja important. Novament cal que hom es refereixi al trànsit entre el Permià i el Triàsic per a deduir-ne un mínim generalitzat.
Finalment, l’aparició d’esquelets permet la preservació més fàcil de les restes dels éssers vius. A l’inici del Cambrià tot fa creure que la gran majoria de fílums ja devien haver existit en unes formes sense esquelet, ja que es manifesten sobtadament en el registre fòssil sense que, en línies generals, hom pugui dir que les formes complexes d’invertebrats mancaven al començament del Fanerozoic. No obstant això, és evident que una anàlisi més acurada permet en molts casos de dibuixar una evolució cap a unes formes més complexes i sovint cap a un augment de la variabilitat, sempre contrarestada pels episodis d’extinció. Aquesta pauta de l’evolució es fa palesa en construir unes gràfiques on s’indica la diversificació progressiva i les extincions a partir de l’aparició d’un fílum, o grup d’organismes.
L’evolució de les plantes

Origen i desenvolupament de les plantes terrestres vasculars al llarg dels temps geològics.
Jordi Corbera, original de Valentine 1978.
És díficil d’establir una pauta evolutiva general del món vegetal a causa de la seva pobresa en el registre fòssil, de la indefinició de les formes unicel·lulars respecte a les formes pluricel·lulars atribuïdes ordinàriament al regne animal i, també, per la situació fluctuant de la definició del mateix regne vegetal. Alguns d’aquests problemes es presenten també dins el grup heterogeni dels animals anomenats invertebrats; tanmateix els hem obviat per fer referència només als grups més importants dins del registre fòssil, grups que són perfectament definits en la sistemàtica zoològica. Semblantment, ací farem alguna referència general a l’interès del registre fòssil vegetal i després als aspectes referents a alguns grups en particular, sense pretendre d’explicar la història i l’evolució de tot el món de les plantes.
Quant a les plantes no vasculars (protòfits i tal·lòfits), podem dir que en coneixem l’existència des de temps molt antic. El fet d’haver trobat estromatòlits a l’Arqueà ens fa pensar en l’existència d’alguns d’aquests organismes en aquests temps remotíssims. També el grup dels acritarcs, d’interès en estratigrafia, ja existia al Precambrià. Sembla que també hi ha bacteris precambrians, però l’absoluta seguretat de llur presència en el registre fòssil és qüestionable, sobretot a causa de la petitesa i del perill de la contaminació. Els fongs hi són rars però se n’han detectat en el Silurià i, sobretot, a partir del Carbonífer. Hom pensa també que les dinoflagel·lades existien al Silurià, si cal fer cas de les troballes fetes a Tunísia. Així doncs, el registre fòssil de les plantes inferiors és pobre i desigual, tot i que, curiosament, en el cas dels estromatòlits, l’activitat d’algunes d’aquestes plantes ha representat les primeres formes «esquelètiques» fossilitzables del món vivent.
Els briòfits, que formen un grup encara no pròpiament pertanyent al món de les plantes vasculars, devien haver aparegut al Denonià superior, o, si més no, les primeres evidències del registre fòssil són d’aquesta època.
L’expansió dels éssers vius sobre la terra ferma la iniciaren les plantes que aconseguiren progressivament de sostreure’s a la necessitat d’estar immergides a l’aigua, gràcies a l’aparició del seu sistema vascular. Malgrat que algunes hagin retornat més tard a medis aquàtics, marins o continentals, la història de les plantes vasculars s’inscriu fonamentalment a la terra ferma a partir del Silurià superior. La diversificació progressiva dels cormòfits a través de la història geològica es tradueix en l’aparició successiva dels pteridòfits, les gimnospermes i les angiospermes, aquest darrer grup amb plantes d’una diversificació espectacular a partir del Cretaci. En concret, els pteridòfits i les pteridospermes són particularment importants i constitueixen un riquíssim component del registre fòssil en el Carbonífer d’interès econòmic primordial; les coníferes i les formes afins són predominants en el Mesozoic i, a partir del Cretaci, les angiospermes són el grup més nombrós. En línies generals, doncs, la història de les plantes superiors fa palès l’augment progressiu de complexitat a través dels temps geològics.
L’evolució dels vertebrats
El que hem dit de les plantes superiors podem dir-ho, també, dels vertebrats que, en certa manera i per analogia, poden ser anomenats animals superiors. El fet que tots els vertebrats hagin aparegut durant el Fanerozoic ens ajuda a entendre’n les pautes evolutives, les quals reflecteixen l’adquisició de caràcters que els permeten una progressiva i major independència del medi i la possibilitat d’una gamma més àmplia d’adaptacions als diferents nínxols ecològics.

Origen i desenvolupament dels peixos al llarg dels temps geològics.
Jordi Corbera, original de Valentine 1978.
Els primers invertebrats són els peixos agnats, que apareixen al Cambrià superior i no tenen mandíbules. Més endavant, al Devonià, apareixen els primers peixos pròpiament dits, és a dir, proveïts de mandíbules i amb parells d’aletes. Durant el Devonià hi ha una gran proliferació de peixos que tenen un esquelet extern i que pertanyen a grups que, o bé s’han extingit, o bé tenen avui molt poc interès. Pel que fa als teleostis, els peixos més abundants avui dia, comencen a expandir-se a partir del Triàsic. En tot cas, els peixos donen origen als amfibis i, d’ells, a tots els altres vertebrats. El grup de peixos predecessors dels amfibis són els ripidistis, peixos primitius d’aletes lobulades, extingits al Permià.
Els amfibis apareixen al Devonià, quan ja les primeres plantes havien aconseguit de dispersar-se més enllà de les zones aquoses i convertir-se en plantes de terra ferma. Així, els vertebrats, i també els invertebrats, podien conquerir la terra seca. Com diu la mateixa paraula, els amfibis no estan totalment alliberats de viure una fase de vida aquàtica i per això van ser substituïts amb avantatge, encara que no totalment, pels rèptils a partir del Carbonífer superior. Durant més de 70 milions d’anys, però, els amfibis es van propagar pels hàbitats disponibles i s’alimentaven d’allò que trobaven a les zones aquàtiques, semiaquàtiques i continentals, tot adquirint grandàries considerables. Sigui com vulgui, a més de la tímida conquesta de la terra ferma de la qual són protagonistes, aquests animals són els primers tetràpodes existents a la Terra. Tots els altres vertebrats que els succeeixen en la línia evolutiva es caracteritzen pel fet de tenir la mateixa estructura a les quatre extremitats, malgrat que s’hi donin adaptacions notables i, àdhuc, en alguns casos, desaparicions parcials o totals.

Origen i desenvolupament dels rèptils al llarg dels temps geològics.
Jordi Corbera, original de Valentine 1978.
Els rèptils s’alliberaren de l’aigua perquè foren capaços de desenvolupar un ou que es podia covar fora d’aquest element líquid. Els rèptils es diversificaren ràpidament i anaren ocupant els ambients on eren instal·lats els amfibis que declinaren ràpidament, de manera que, al Triàsic superior, els amfibis de més grandària s’extingiren definitivament. Ja abans, per això, al Permià, els rèptils predominaven i durant tot el Mesozoic eren els vertebrats dominants, malgrat que ja haguessin fet la seva aparició els mamífers al Triàsic i els ocells al Juràssic. L’expansió extraordinària dels rèptils i l’enormitat de moltes de llurs espècies en feren els animals més grans de tota la història de la vida, cosa que ha fet que l’Era Mesozoica hagi estat anomenada l’era dels rèptils. D’aquesta evolució complexa i de l’enorme diversificació, esdevinguda al Mesozoic, en queden restes en els grups actuals, que encara acompleixen un paper important en determinats ambients: les serps, els cocodrils, els llangardaixos i les tortugues.

Origen i desenvolupament dels mamífers al llarg dels temps geològics.
Jordi Corbera, original de Valentine 1978.
Els rèptils, per mitjà dels teràpsids, un petit grup extingit al Juràssic, van donar lloc durant el Triàsic als mamífers que, com el nom indica, desenvoluparen unes estratègies particulars en l’alimentació de les cries i, ràpidament, en el mateix sistema de reproducció. I a més a més, presentaren l’homeotèrmia que els allibera, en gran part, de la temperatura ambient, gràcies al manteniment de la temperatura de la sang constant. Aquesta adquisició la comparteixen amb els ocells que apareixen més tard, al final del Juràssic, provinents també dels rèptils.
Els ocells s’adapten fonamentalment al vol i presenten un domini quasi absolut del món aeri que comparteixen amb alguns invertebrats voladors i amb algun mamífer escadusser, com els rat-penats.

Etapes fonamentals del procés d’hominització, segons la interpretació més acceptada a l’actualitat.
Dades de D. Pilbeam (1984), reelaborades per l’autor
Per contra, els mamífers, que experimenten una gran diversificació a partir del Cenozoic, protagonitzen el predomini dels àmbits continentals tal com ho havien fet els rèptils al Mesozoic. Entre els mamífers destaquen els primats, dels quals sorgeix l’home, en el qual una cerebralització accentuada i la postura bípeda dreta, juntament amb la parla, el separen dels altres mamífers en comportament i realitzacions. El procés d’hominització s’inicià a partir de la radiació dels catarrins, o simis del vell món.
El pas de la vida aquàtica a la vida continental i aèria
Deixant de banda l’augment de la complexitat dels éssers vius detectada en el registre fòssil, hi ha un fenomen força curiós per a l’observador interessat en la història de la vida que, si bé té quelcom a veure amb l’adquisició de graus de més complexitat, té unes característiques pròpies remarcables. Es tracta de la conquesta de la terra ferma i, secundàriament, de l’aire. Els primers éssers vius són aquàtics, i és ben cert que la vida dins un ambient no aquàtic requereix unes estructures particulars, no gens senzilles, que evitin l’assecament, entre altres coses; per consegüent, la vida no començà per aquí.
L’associació fòssil més antiga de plantes i animals continentals conservada en el propi ambient es troba a Escòcia i és del Devonià inferior. El fet que aquesta associació correspongui a plantes i animals amb adaptacions morfològiques i fisiològiques al medi continental força avançades fa pensar que la invasió de la terra ferma pels éssers vius és anterior a aquest moment; diverses troballes i arguments basats en unes quantes evidències fan pensar que ja al Silurià havia començat a aparèixer la vegetació sobre el continent. En tot cas, l’aparició de la vegetació sobre el continent marca una nova etapa de la història, amb unes repercussions, fins i tot, sedimentàries i pedològiques; encara més, implica i provoca noves situacions en la relació atmosfera/litosfera.
Com és fàcil de comprendre, els invertebrats voladors són associats principalment a la vida en el continent. Entenem per vida continental la vida sobre la terra eixuta, encara que també hi ha àrees aquàtiques dins el continent o als seus marges. D’altra banda, la flora no aquàtica necessita unes adaptacions per a facilitar cilitar la dispersió dels elements seminals i els encreuaments. Aquests processos es desenvolupen a l’aire. És, per tant, a partir de l’existència de la cobertora vegetal sobre la terra eixuta que l’atmosfera s’omple d’éssers vius i dels seus productes. Així comença la vida aèria i, amb això, podem dir que a partir d’aquest moment la hidrosfera, juntament amb les parts més pròximes de la litosfera i l’atmosfera, constitueixen un embolcall terrestre fràgil, més o menys dens segons els llocs, que podem anomenar biosfera.
L’estratègia de l’evolució biològica
Les pautes evolutives dels grups d’organismes

Representació de la diversitat de pautes evolutives d’aparició, d’extinció i de diversificació, exemplificada en alguns dels grans grups d’animals invertebrats basat en el nombre de famílies.
Distrimapas, original de J.J. Sepkoski.
Malgrat que el registre fòssil sigui incomplet i esbiaixat, és prou ric per a l’estudi de l’evolució en el temps de molts grups d’organismes, ordinàriament d’aquells grups que tenen un esquelet. Aquesta visió, de gran interès biològic, només és possible mitjançant l’estudi paleontològic, ja que l’anàlisi dels éssers vius actuals és la d’una pura instantània en el temps, multimilionari, de la història de la vida. Així doncs, les diverses pautes d’aparició, desenvolupament i, eventualment, desaparició dels diversos grups d’organismes, assequibles mitjançant el registre fòssil, serveixen per a entendre el funcionament dels diversos fílums o d’altres divisions a través del temps. En aquest context, cal veure quines són aquestes pautes, fer-ne una classificació tipològica i intentar d’esbrinar les causes d’aquests comportaments grupals o veure-hi les successives faunes que van poblant la terra. En el primer cas, a més, cal preguntar-nos pel fet repetit, amb diverses intensitats, de les extincions en massa de grans grups taxonòmics. Cal, doncs, veure quines poden haver estat les causes d’aquestes extincions.
Pel que fa a les pautes evolutives dels grups més importants d’éssers vius podem observar uns quants grups d’invertebrats marins importants que poden esdevenir paradigmàtics. D’alguns, podem estudiar-ne l’aparició, el desenvolupament i la desaparició, ja que, com a grans grups, s’han extingit (graptòlits, trilobits), mentre que d’altres, al contrari, són en plena expansió (gimnolemats, gasteròpodes, bivalves). Pel que fa a la forma d’aparició, n’hi ha uns que presenten un començament amb una variabilitat progressiva, i d’altres en què l’aparició i diversificació màxima són molt pròxims en el temps. Del primer cas, per exemple, tenim, d’una banda, els bivalves i els gasteròpodes, amb una diversificació creixent continuada, sense canvis bruscs, mentre que de l’altre, els equinoïdeus i els gimnolemats tenen una diversificació molt escassa, fins i tot amb retrocessos durant un temps llarg i, a la fi, un augment de formes ràpid i important. Quant a les aparicions amb diversificació ràpida, podem parlar dels trilobits i dels graptolitins, la pauta de desaparició dels quals, per contra, és progressiva i relativament lenta. Els crinoïdeus, els ostràcodes, els tenolemats i els antozous presenten una disminució de la seva importància al límit entre el Paleozoic i el Mesozoic. Això obeeix al fet de l’extinció en massa d’una gran quantitat de formes al final del Paleozoic i a l’aparició progressiva de formes noves al Mesozoic-Cenozoic. En aquests grups la varietat és més gran durant l’etapa paleozoica que no pas a la més moderna; això és particularment visible en els crinoïdeus i també en els braquiòpodes articulats, els quals no mostren cap estrangulament marcat al límit Paleozoic-Mesozoic, sinó una reducció dràstica de formes que es mantenen quantitativament iguals des d’aleshores. Finalment, els cefalòpodes presenten uns escanyaments successius i marcats que evidencien també les renovacions freqüents, pràcticament totals, de tota aquesta fauna.
La història de la diversitat taxonòmica
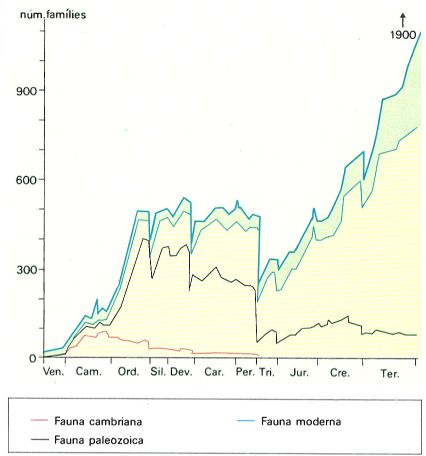
Evolució de la diversitat de famílies d’animals marins al llarg del Fanerozoic, des de fa 600 milions d’anys (en el Vendià) fins a l’actualitat. La línia superior ateny les 1900 famílies, que corresponen a les descrites o assenyalades en els oceans moderns, una part de les quals no tenen parts dures fàcilment fossilitzables; l’àrea marcada en verd correspon a aquestes famílies pobrament representades.
Distrimapas, original de J.J. Sepkoski.
Una anàlisi més detallada de l’evolució dels grups demostra, en línies generals, que hi ha un augment de la diversitat, contra el qual s’interposen les extincions en massa; d’aquestes, només les més ressaltants són capaces de produir baixades considerables en aquesta corba ascendent de la diversitat, per exemple, en l’extinció permo-triàsica. Hom sol fer aquesta anàlisi sobre famílies d’animals marins, ja que aquests animals són els més fàcils de ser conservats dins la roca i constitueixen la part més important del registre fòssil. Modernament, Sepkoski ha intentat descompondre la corba del nombre de famílies en tres corbes, corresponents a tres faunes: una fauna cambriana, que desapareix al Paleozoic superior i deixa només relíquies en faunes posteriors fins a l’actualitat (com per exemple, els braquiòpodes inarticulats); una fauna paleozoica, amb una expansió vigorosa durant el Primari i que després fa la viu-viu fins a l’actualitat; i una fauna moderna que s’inicia tímidament al Paleozoic per a créixer en diversitat a través del Mesozoic i el Cenozoic fins al moment actual. El registre fòssil del final del Terciari presenta un nombre de famílies 6,5 vegades més gran que tota la fauna paleozoica i les relíquies de forma càmbrica sumades. Així doncs, també l’anàlisi de la diversitat taxonòmica ens porta a pensar en termes d’aparició, de desenvolupament i de desaparició, sobretot si, en lloc de treballar amb agrupacions tan grans i en rangs taxonòmics tan elevats, pensem en els rangs taxonòmics inferiors, com ara espècies, gèneres i famílies.
Pel que fa a l’aparició de les formes d’éssers vius, els nostres coneixements són àmpliament hipotètics, car tot fa pensar que es dóna en uns llocs puntuals i en unes quantitats més petites, difícilment detectables atesa la incompleció del registre fòssil. Quant a la desaparició, podem considerar el cas del trànsit evolutiu a d’altres formes descendents (que correspondria, aproximadament, al mateix problema), o bé el cas més espectacular i catastròfic de les extincions en massa a les quals modernament es fa una particular atenció i que discutirem, amb brevetat, a continuació.
Les extincions en massa i llurs causes

Principals extincions marines en massa (en color verd), en el context de la història de la vida en els últims 700 milions d’anys, on s’indiquen els grups més afectats i on s’han figurat els grups totalment extingits.
Jordi Corbera, original de M. Stanley.
Des de l’inici de l’estratigrafía, la desaparició de fílums grans i l’aparició de noves formes va esdevenir un criteri útil per a subdividir el registre rocós en unes unitats que es caracteritzaven, sobretot, per aquests mateixos fòssils.
No és gens estrany, doncs, que l’anàlisi moderna de les extincions en massa faci veure que les més importants s’escauen al límit entre les unitats majors. Així, les grans extincions se situen al límit entre el Paleozoic i el Mesozoic (l’anomenada extinció permo-triàsica) i al límit entre el Mesozoic i el Cenozoic (la coneguda extinció finicretàcia), però també n’hi ha d’importants al final de l’Ordovicià, del Devonià, del Triàsic i del Juràssic. Una anàlisi més fina, només possible en temps més pròxims a nosaltres, ha detectat uns episodis d’extinció menys importants, però notables, situats en diversos nivells de l’escala estratigràfica. Fins i tot les extincions dels darrers 250 milions d’anys semblen produir-se a intervals relativament regulars, a la ratlla dels 26 milions d’anys. Aquesta repetició cíclica de les extincions indueix a cercar alguna causa que estigui sotmesa a una ciclicitat anàloga. És així que hom cerca fora de la Terra, en els estels, uns fets que s’esdevinguin cada 26 milions d’anys i que puguin influir sobre els éssers vius de la Terra. Hom ha parlat, per exemple, d’un estel petit i fred que donava voltes entorn del Sol en una òrbita molt allargada, de manera que cada 26 milions d’anys, en passar prop del Sol, hauria pogut pertorbar les òrbites dels cometes i provocar una caiguda de material estel·lar sobre la Terra, fet que hauria comportat fenòmens disparadors d’extinció, tals com els que molts autors moderns suposen per a l’extinció finicretàcica.
Si acceptem aquestes opinions, les causes de les extincions devien ser extraterrestres, derivades bé d’algun esdeveniment com el que acabem d’indicar o bé d’un canvi en la polaritat magnètica, fets diversos que, en tot cas, permetrien l’arribada a la Terra de meteorits descomunals o de radiacions nocives en quantitats exagerades que provocarien de forma directa i indirecta unes catàstrofes biològiques detectables en el registre fòssil.
Ara bé, no tothom està d’acord a cercar causes extraterrestres a les extincions o, si més no, a donar-hi una importància capital o única. En particular, hom pensa que els canvis climàtics són una causa més probable si es tenen en compte els arguments que subministra el mateix registre fòssil i la documentació derivada d’unes extincions pròximes com les ocorregudes al Quaternari. Hom ha parlat també d’extincions en massa a les mars a causa de la reducció de l’espai de la plataforma continental, del descens del nivell de la mar i també de les variacions en la salinitat de les aigües. En el cas de les anomenades crisis de salinitat, és sabut que la que tingué lloc ara fa més de 5 milions d’anys, a la fi del Miocè, al Messinià, tingué un efecte fatal per a la fauna de la Mediterrània, on es produí un dessecament; també s’ha invocat aquesta causa per a explicar l’extinció permo-triàsica, i per a d’altres extincions. Però, malgrat l’opinió d’alguns autors, no sembla que serveixi com a justificació general de les extincions.
La reducció de l’espai a la plataforma continental, amb producció d’un amuntegament biòtic, que devia produir-se a les grans regressions de la història de la Terra, sembla clar que pot contribuir a desencadenar unes extincions importants, però és molt discutible com a causa única, car el registre rocós ens demostra la no-coincidència d’extincions importants amb les fases de regressió més paleses. Pel que fa al refredament com a causa important i selectiva de les extincions, s’han aportat proves de considerable interès i, sobretot, se n’ha remarcat l’aspecte selectiu; és així, per exemple, en l’extinció del final del Cretaci i en d’altres, on les faunes d’aigües càlides en són les principals perjudicades (ja que no tenen on refugiar-se), mentre que les faunes d’àrees més fredes són menys afectades, perquè poden desplaçar-se a llocs de condicions climàtiques més bones.
Totes aquestes causes terrestres invocades fins avui, de les quals acabem d’esmentar les més importants, parteixen del punt de vista que la desaparició total o parcial de determinats medis provoca l’eliminació, per manca d’adaptació, de grups importants d’organismes. Les causes extraterrestres, d’altra banda, originen una catàstrofe a les aigües, o a l’aire, que duu també a la desaparició de grans grups d’éssers vius. Atesa la resposta diferent a un mateix episodi d’extinció dels diversos grups ecològics d’éssers vius, semblarien més explicables les extincions per causes que afecten selectivament uns diferents ambients; de cap manera, però, ningú no pot negar la coincidència d’uns esdeveniments còsmics, enregistrats dins les roques, i de determinats episodis d’extinció. Altrament, potser també cal comptar amb l’evolució pròpia, en certa manera interna, dels diversos grups d’éssers vius que, com s’ha comprovat, experimenten un desenvolupament semblant als individus vivents d’aparició, expansió, decadència i desaparició. Probablement, la conjuminació de múltiples factors interns dels éssers vius i d’altres que en són externs, siguin provocats o no per fets extraterrestres, poden explicar aquestes crisis de la història de la vida que es resolen amb la desaparició de grups majors d’éssers vius i amb l’aparició de nous, sovint ben diferents, però que acostumen a colonitzar, de bell nou, els ambients abandonats pels existents anteriorment.