El pol·len de les fanerògames i les espores de les criptògames formen part del grup de microfòssils orgànics caracteritzats per la seva bona conservació en el sediment, gràcies a la resistència de la seva paret, en part formada per esporopol·lenina. Una altra característica que els és comuna són les seves petites dimensions i la possibilitat d’ésser transportats pel vent a grans distàncies abans de sedimentar-se, cosa que permet que tinguin una àmplia dispersió. És important el fet que posseeixen una estructura i una escultura externa característiques, ja que això permet d’atribuir-los a determinades famílies, o a gèneres i fins i tot a espècies. Finalment, les seves petites dimensions fan que en una petita quantitat de sediment hom disposi de mostres en abundància.
L’estudi del pol·len i les espores és una ciència que ha suscitat un gran interès motivat per les seves aplicacions, entre les quals cal assenyalar: en botànica, principalment, com a ajut en l’estudi de la filogènia de certs grups de plantes; en medicina, a causa de les al·lèrgies produïdes pel pol·len i les espores continguts en l’atmosfera (aeropalinologia); i també perquè aporta dades aplicables a l’agronomia, com el control de la pol·linització d’arbres cultivats i la detecció i aparició de determinades espores de fongs patògens en certs conreus. L’estudi del contingut esporopol·línic de les mels és tractat per la melitopalinologia; i, finalment, la seva aplicació en arqueologia i en geologia a partir del pol·len i les espores continguts en els sediments, i obtinguts per processos químics; en arqueologia, la palinologia subministra les dades suficients per a donar-nos una imatge aproximada de l’entorn vegetal en què es movia l’home en el passat, i també dels seus conreus. En geologia, permet d’obtenir correlacions entre sediments, tant d’origen continental com marí, de vegades a grans distàncies, i conèixer la vegetació en el passat, amb el consegüent ajut a la paleoclimatologia.
Morfologia

Estructura de l’exina del gra de pol·len d’una angiosperma (Lavandula pedunculata, × 40 000), en un tall transversal. Hom hi ha indicat les parts següents: 1 tèctum, 2 columel·la, 3 capa basal, 4 endexina, 5 intina.
Lluís Centelles, a partir de materials de Maria Suàrez.
La cèl·lula d’un gra de pol·len és envoltada d’una coberta protectora (esporodermis) formada per dues capes: la intina, més interna, que limita amb la cèl·lula i és de composició semblant a les parets de les cèl·lules vegetals; i l’exina, que envolta la intina. Durant els processos de fossilització desapareixen la cèl·lula i la intina, de manera que només queda l’exina, que deu la seva resistència a la presència en la seva composició d’esporopol·lenina. En una secció de l’exina observada al microscopi electrònic es pot diferenciar una part més interna i contínua (l’endexina, típicament d’estructura lamel·lar en les gimnospermes i no lamel·lar en les angiospermes) i una part externa (l’ectexina). Hi ha, també, una diferenciació química entre ambdues parts: mentre l’endexina es tenyeix molt dèbilment amb fucsina B, l’ectexina ho fa amb un color vermell fosc. L’ectexina, alhora, es diferencia en tres parts: una capa basal contínua, una capa intermèdia (infratèctum, que pot estar formada per pilars, grànuls o alvèols) i una capa externa (tèctum, que de vegades pot ser discontínua o, fins i tot, no ser-hi). Es freqüent que l’exina presenti una escultura més o menys desenvolupada, que pot emplaçar-se per damunt del tèctum formant un relleu supratectal o, si no n’hi ha, es forma a expenses de la capa intermèdia de l’ectexina, tot modificant-la. En les espores, les dues parts de l’exina s’anomenen endospori i exospori respectivament, de manera homòloga a l’endexina i l’ectexina.

Gra de pol·len porat (× 1000) d’un arbre fòssil del mateix gènere (Alnus) que el vern actual, freqüent al Miocè i al Pliocè de Catalunya. Noteu els arcs que adornen la seva superfície, que li són característics.
Núria Solé.
A més de l’estructura i l’escultura que presenta l’exina, també és important el nombre i la forma de les obertures o zones germinatives. La majoria dels grans de pol·len presenten zones de l’exina que han sofert modificacions més o menys importants, on pot arribar a faltar per a formar les anomenades obertures o apertures, que poden ser de tres classes: circulars (porus), allargades (colps) o una combinació d’aquestes dues (colporades); en les espores només se’n presenta una d’allargada (monolet) o una de triradiada (trilet). El nombre i la distribució d’aquestes obertures és important.
La forma externa del gra de pol·len pot variar molt, des d’el·lipsoïdal o circular fins a presentar dues expansions laterals, com en la majoria de les gimnospermes. De vegades, alguns grans resten units formant tètrades o políades. Les dimensions, tant del pol·len com de les espores, presenten valors que oscil·len entre 3 i 200 µm, encara que generalment es troben compresos entre els 15 i els 60 µm. En el cas de les megàspores, aquests valors poden arribar fins a un màxim de 5000 µm.
El conjunt de detalls de l’estructura i l’escultura de l’exina, els caràcters de les obertures i les dimensions i la forma externa són característiques constants per a cada espècie vegetal. Això determina la morfologia i permet de realitzar la classificació o la determinació del pol·len i les espores fòssils, pertanyents a qualsevol període geològic.
Classificació

Morfologia dels grans de pol·len. A Tipus d’obertures i la seva posició en els grans de pol·len: 1 monocolpat, 2 dicolpat, 3 tricolpat, 4 estefanocolpat, 5 monoporat, 6 diporat, 7 triporat, 8 estefanoporat, 9 pericolpat, 10 periporat, 11 tricolporat, 12 estefanocolporat, 13 heterocolpat, 14 sincolpat, B Forma de les espores i tipus d’obertures: 15 monolet, 16 trilet. C Formes especials de pol·len: 17 i 18 vesiculat, 19 inaperturat, 20, 21 i 22 tètrada, 23 políada.
Lluís Centelles, a partir de fonts diverses
Les espores i el pol·len fòssils del Pleistocè i de part del Terciari superior es determinen per comparació amb les formes actuals. Per aquesta raó, cal disposar d’una col·lecció àmplia de pol·len de les espècies actuals, i particularment d’espècies pròpies de la regió d’estudi, que serveixi de referència per a comparar. La classificació, en aquest cas, és natural. Per al pol·len i les espores d’edat més antiga, i ja que al llarg del temps han desaparegut nombroses formes de vegetals, s’utilitza una classificació fonamentada en morfogèneres que, al seu torn, han estat reunits en categories superiors; s’adopta, així, l’esquema que proposaren Potonie i Kremp (1945), fonamentat enterament en categories morfològiques. En conseqüència, la majoria dels noms genèrics es formen per addició de sufixos, com -spora, -sporites, -pollis i -pollenites. En alguns casos, aquests noms recorden els gèneres actuals (per exemple, Ilexpollenites o Matonisporites).
Diagrames pol·línics
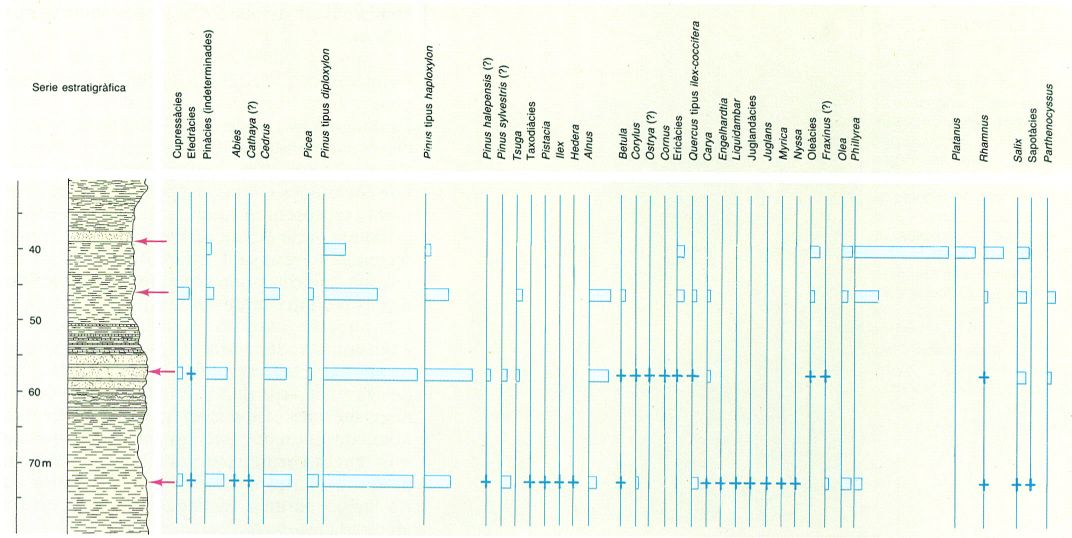
Exemple de diagrama pol·línic, realitzat a partir d’unes mostres pliocenes, on s’ha representat el pol·len arbori d’una secció de Sant Onofre, a Tortosa (Baix Ebre).
Distrimapas, original de M.F. Valle Hernández
Els diagrames pol·línics són especialment indicats en l’estudi del Pleistocè i del Terciari superior, per a la utilització de la informació obtinguda en diferents mostres de sediments. Primerament cal obtenir l’anomenat espectre pol·línic de la mostra analitzada, que consisteix a expressar en percentatges les freqüències relatives de cada espècie trobada. Amb aquests resultats hom construeix el diagrama pol·línic, és a dir, la representació gràfica dels diferents espectres pol·línics efectuats en mostres seriades procedents d’una secció geològica. La sèrie estratigràfica, és a dir, els diferents nivells, es representen en les ordenades i els espectres de cada nivell en les abscisses. D’aquesta manera es visualitzen ràpidament les variacions quantitatives que presenta cada espècie a través de la sèrie estudiada. En un principi, les anàlisis pol·líniques de sediments consideraven només el pol·len de les plantes arbòries, però en l’actualitat inclouen també plantes herbàcies i espores.
Cronostratigrafia
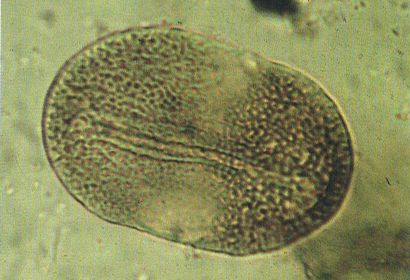
Gra de pol·len bialat d’Ovalipollis ovalis, una espècie procedent del Triàsic (Muschelkalk superior i Keuper) del Principat, del País Valencià i de Mallorca. Noteu l’obertura longitudinal, que és ben visible a la fotografia (× 900).
Núria Solé
Les primeres formes conegudes pertanyen a les espores. En el Silurià inferior es troben ja espores amb trilet, formades amb exines engruixides, procedents de plantes vasculars. En el Devonià hi ha una gran diversificació en les espores, amb una ornamentació més complexa i de dimensions més grans. En el Devonià mitjà apareix l’heterospòria en certs grups de plantes gràcies a la ràpida colonització dels hàbitats terrestres per les plantes vasculars. Els conjunts esporopol·línics més ben coneguts del Paleozoic són els del Carbonífer, que tenen una gran varietat de formes, i entre ells apareixen els de les primitives gimnospermes. Fins a l’actualitat, no hi ha cap dada procedent dels Països Catalans referent a aquests microorganismes del Paleozoic. Són coneguts, però, al N de la península Ibèrica, a Astúries i a Lleó.

Exemplars d’espora trilet del cretaci. A l’esquerra, de l’esquizeàcia Cicatricosisporites (× 1100), amb escultura estriada, molt abundant al Cretaci català; la família de les esquizeàcies, d’una gran importància cronostratigràfica, actualment té una distribució tropical. Al mig, espora de la gleiqueniàcia Gleicheniidites (× 1300), freqüent en sediments de l’Albià de les terres tarragonines; la família actualment es troba només en països intertropicals. A la dreta, Trilobosporites, també molt freqüent a l’Albià tarragoní (× 840).
Núria Solé
El començament del Mesozoic es caracteritza en les nostres regions per un canvi en la flora, que és conseqüència d’un canvi climàtic. Sembla que el clima esdevingué més àrid, cosa que s’acompanyà d’un augment de la temperatura. Això comportà una gran abundància de pol·len bialat de gimnospermes, adaptat al transport per l’aire. Formes com els gèneres Ovalipollis, Triadispora i Vallesporites són molt abundants i han estat ben estudiades al Triàsic català. De fet, els Catalànids són una de les zones més interessants i representatives del Triàsic mediterrani per la quantitat i la bona conservació de la microflora.
Durant el Juràssic continuen predominant les gimnospermes, malgrat que les espores són encara molt importants. No hi ha dades d’aquest període als Països Catalans ni tampoc a la resta de l’estat espanyol. Sembla que es tracta de flores molt cosmopolites i de poc valor cronostratigràfic.
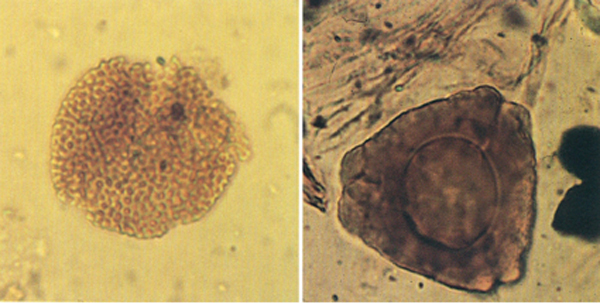
Grans de pol·len fotografiats al microscopi òptic. A l’esquerra, Tricolpopollenites (× 1000), forma que és considerada com una de les primeres angiospermes tricolpades que apareixen en el nostre registre fòssil. La mostra procedeix de l’Albià mitjà de les terres tarragonines. A la dreta, Hungaropollis, un triporat primitiu del grup dels normapol·les. El de la fotografia (× 1000) procedeix del Cretaci superior lleidatà.
Núria Solé.
Del Cretaci hi ha, en canvi, nombrosos estudis. Es produeix una gran expansió de les espores, sobretot les de la família de les esquizeàcies (Schizaeaceae), en la forma del gènere Cicatricosisporites, que s’estén des del Cretaci inferior al Terciari i és molt abundant en els sediments catalans del Cretaci inferior; l’acompanyen altres formes nombroses, com Gleicheniidites i Trilobosporites, totes amplament representades en extenses àrees de diferents parts del món. Procedents de terres tarragonines i castellonines, especialment de la zona del Maestrat, s’han obtingut conjunts esporopol·línics molt rics del Cretaci inferior. En aquesta mateixa època apareixen les angiospermes més primitives. Sembla que hi ha una evolució respecte del nombre i la forma de les obertures dels grans de pol·len de les angiospermes. El primer gra de pol·len que hom considera que pertany indubtablement a una angiosperma procedeix del Barremià; presenta una estructura columel·lar típica de les angiospermes, amb una sola obertura allargada (monocolpat), i pertany al morfogènere Clavatipollenites, que possiblement és relacionat amb la família actual de les clorantàcies (Chlorantaceae). A les nostres terres, durant l’Albià apareixen els primers grans d’angiospermes proveïts de tres obertures allargades (tricolpats), mentre que en el Cenomanià mitjà es troben grans de pol·len amb tres obertures circulars (triporats) juntament amb d’altres obertures compostes (tricolporats). És especialment important un grup de triporats, els anomenats normapol·les («normapolles»), els porus dels quals evolucionen fins a adquirir una gran complexitat i donen lloc a nombrosos gèneres morfològics d’un gran valor cronostratigràfic. Aparegueren al Cenomanià mitjà i foren molt abundants al Paleocè fins que desaparegueren ràpidament durant l’Eocè. Segons que sembla, es relacionaven amb les amantíferes o mirtàcies primitives, que formaven part d’una flora d’ambients càlids i humits, avui dia refugiada al SE asiàtic. De la zona del barranc de la Posa (Pallars Jussà) s’han trobat deu gèneres diferents de normapol·les (Plicapollis, Nudopollis, Vacuopollis, etc.), d’edat maastrichtiana, acompanyats d’una rica flora d’espores i pol·len, amb dues espècies noves: Trilobosporites herngreenii i Platycaryapollenites ferreri.
El Cenozoic inferior, a grans trets, es caracteritza per la gran diversitat d’espècies de pol·len d’angiospermes que apareixen, acompanyades d’espores, i pels baixos percentatges de gèneres encara vivents. Hom ha obtingut una rica composició esporopol·línica de sediments procedents d’àrees pròximes a Súria (Bages) i a Calaf (Anoia). A partir de l’Eocè desapareixen moltes espècies d’ambients càlids i se n’introdueixen de noves, de llocs més temperats. Aquest fet s’accentua en l’Oligocè superior, on domina aquesta flora més temperada (Alnus, Betula, Corylus, etc.), amb espècies de climes càlids (Palmae, Sapotaceae, Engelhardtia, etc.).

Exemplars de grans de pol·len. A l’esquerra, un bialat (× 750) d’una espècie extingida de pi (Pinus), concretament del tipus haploxylon, que és molt freqüent en els sediments del Miocè de Catalunya. A la dreta, un triporat de Carya (× 1000), que s’extingí a Europa durant el Quaternari inferior. És molt abundant en els sediments del Miocè i del Pliocè de Catalunya.
Núria Solé.
El refredament progressiu continua al Terciari superior. A Catalunya es coneix bé el pol·len d’aquest període ja que existeixen nombrosos estudis sobre el Miocè i el Pliocè, especialment el marí, del Principat. A més de la representació de la flora actual hi apareixen algunes formes de llocs càlids, com a relíquia d’èpoques anteriors que han suportat els canvis climàtics. Així, en el Miocè de la rodalia de Sant Sadurní d’Anoia (Alt Penedès) s’ha trobat en el sediment pol·len de manglar, que poblava aleshores aquesta zona. El Pliocè conté nombrosos pòl·lens de plantes com Taxodium, Sequoia, Tsuga i Carya, espècies que actualment viuen a l’Amèrica del Nord.
A conseqüència de la sèrie d’avenços i de retrocessos de la flora motivada pels dominis d’extensió de les geleres a Europa durant el Quaternari, aquesta flora va anar empobrintse. En cada retrocés es trobava amb les barreres naturals que representen les serralades de muntanya en direcció E-W i la mar Mediterrània; aquest fet provocà la desaparició de molts arbres.
Els diferents canvis climàtics enregistrats venen reflectits en l’anàlisi pol·línica efectuada en sediments quaternaris d’una regió determinada. Hi ha poques dades procedents dels Països Catalans, que provenen especialment de la fracció superior del Quaternari.