El fenomen vital
La vida i el planeta blau
No coneixem actualment cap evidència de l’existència de vida fora del nostre planeta. És cert que hi ha molècules orgàniques complexes a l’espai interstel·lar, i també que alguns dels meteorits que cauen a la superfície de la Terra en contenen, però els fenòmens que avui associem a la vida (capacitat de control del medi intern i dels bescanvis d’energia i aliments amb l’exterior, multiplicació i desenvolupament de nous organismes a partir d’estructures especialitzades en la conservació i la transmissió d’informació genètica, etc.) no es limiten a la simple presència o juxtaposició de determinades molècules. En una proporció gens escassa, les propietats de la matèria viva, tal com la coneixem, estan vinculades a característiques físico-químiques que, si més no al sistema solar, són específiques del nostre planeta o són un resultat de la seva història. Així, per exemple, no és imaginable la vida sense aigua; tots els éssers vius contenen aigua en proporcions elevades.
Tampoc no és imaginable la vida sense una organització cel·lular, és a dir sense l’existència d’unes unitats anomenades cèl·lules formades per un fluid aquós envoltat per una membrana semipermeable. Aquesta membrana permet isolar de l’exterior les reaccions químiques pròpies, i alhora seleccionar el pas endins o enfora de determinades molècules. En aquest fluid aquós hi ha molècules orgàniques que poden ésser utilitzades com a font d’energia (com, per exemple, l’adenosina trifosfat, o ATP), com a suport de la informació necessària per a la reproducció (els àcids desoxiribonucleics, o DNA), com a vehicle perquè aquesta informació es tradueixi en síntesis concretes d’aminoàcids i de proteïnes (els àcids ribonucleics, o RNA) o com a catalitzadors de determinades reaccions.
La diversificació dels éssers vius en un nombre elevat de formes d’organització i d’espècies, adaptades cadascuna a condicions ambientals i d’aprofitament dels recursos molt precises, és un altre tret característic de la vida actual a la Terra. L’evolució de la biosfera i la seva situació concreta en cada moment és precisament el resultat de successives diversificacions i extincions dels éssers vius dels temps precedents. Com la pròpia història dels humans, no és predictible però sí explicable a partir dels fets anteriors, més o menys aleatoris o determinats, que l’han afectat o que l’afecten.
L’obscur començament
Com va començar la vida a la Terra? En quin moment? Com van poder evolucionar a partir de precursors químics encara no vius les primeres cèl·lules, els primers organismes? Què va passar en el període anterior a la formació, ara fa 3 500 milions d’anys, de les primeres roques que comporten registre fòssil? Encara estem lluny de poder respondre de manera concloent a aquestes preguntes tot i que no falten elements per poder emetre algunes hipòtesis prou versemblants. Hem de tenir present que allò que separa els sistemes vius dels inanimats és la seva capacitat d’enregistrar, emmagatzemar i utilitzar informació al llarg de períodes dilatats de temps, i alhora obtenir rèpliques d’aquesta informació i duplicar tota la sèride de molècules i estructures que aquesta informació determina. És, en definitiva, la seva qualitat de sistemes autopoètics, és a dir, capaços d’assegurar ells mateixos la seva persistència i la seva identitat.
La majoria dels investigadors interessats per la qüestió estan d’acord a admetre que la vida sobre la Terra comença fa força més de 3 500 milions d’anys, data a partir de la qual disposem d’un registre fòssil ininterromput. El fet que els primers testimonis fòssils coneguts ja corresponguin a organismes molt semblants, si no idèntics, a alguns grups de bacteris actuals sembla indicar que l’emergència de la vida sobre la Terra ha de ser molt més antiga, potser fins i tot anterior a la formació de les més antigues roques sedimentàries conegudes. Els testimonis fòssils més antics dels inicis de l’evolució prebiòtica es poden remuntar als mateixos orígens de la Terra com a planeta. De fet, la síntesi abiòtica de compostos orgànics, que se suposa que és una condició prèvia per a l’emergència de la vida, pot produir-se en períodes de temps molt curts.
La química de la matèria viva, la bioquímica, es fonamenta en compostos de carboni reduïts, és a dir en molècules formades bàsicament per àtoms de carboni envoltats per àtoms d’hidrogen. La flexibilitat de la química del carboni és un dels secrets fonamentals de la vida sobre la Terra. Ja no es discuteix que en les molt agitades condicions dels primers cents de milions d’anys de l’eó Arqueà, els àtoms de carboni podien combinar-se amb una certa facilitat amb hidrogen, nitrogen, oxigen, fòsfor i sofre, per generar una extensa varietat de substàncies. Aquest conjunt de molècules riques de carboni s’haurien mantingut prou temps per interactuar i evolucionar, ja fos en el “brou” prebiòtic primitiu preconitzat per Oparin, Haldane i molts altres autors, ja (com indiquen hipòtesis més recents) en els bassals poc profunds o les superfícies xopes d’aigua calenta de la Terra primitiva, allí on gasos rics d’hidrogen procedents de l’interior de la Terra podien reaccionar amb els gasos rics de carboni de l’atmosfera exterior en un medi aquós en el qual podien romandre en solució prou concentrada. També podria ser que les superfícies submergides —o, si més no, humides—, d’argiles i cristalls de la Terra hadeana, haurien afavorit la polimerització de les molècules orgàniques petites en molècules més grans i complicades.
El fet que la síntesi abiòtica de compostos orgànics no hauria requerit períodes de temps gaire dilatats és confirmat tant per les dades empíriques facilitades per objectes extraterrestres com ara cometes o meteorits com per les dades de laboratori acumulades d’ençà que el 1953 s’iniciaren experiments de simulació d’aquesta síntesi: sotmetent una barreja de gasos similar a la que se suposa que hi podria haver hagut a la primitiva atmosfera de la Terra (amoníac, vapor d’aigua, hidrogen i metà) a descàrregues elèctriques durant diversos dies poden obtenir-se una colla de compostos orgànics entre els quals dos aminoàcids: alanina i glicina. A partir de barreges de gasos senzills, i emprant diferents fonts d’energia (descàrregues elèctriques, radiació ultraviolada o calor), també s’han pogut sintetitzar compostos orgànics més complexos, incloent-hi aminoàcids i bases dels àcids nucleics. No hi ha dubte, doncs, que era possible l’existència d’aquestes molècules a la Terra hadeana. Fins i tot, en algun dels experiments més recents, s’han format molècules de trifosfat d’adenosina (ATP), d’altres trifosfats precursors de nucleòtids i dels cinc nucleòtids (adenina, citosina, guanina, timina i uracil) que formen part dels àcids nucleics. Cal afegir que, tal com molt bé podia haver succeït en els ambients terrestres primitius, en aquestes mixtures hadeanes de laboratori també s’han format altres compostos orgànics desconeguts o inexistents avui en els éssers vius, la identitat i la possible funció dels quals en la vida primitiva resulten enigmàtiques; tanmateix, no és inversemblant que tinguessin algun paper en l’estructura o el metabolisme d’alguns dels microorganismes primitius desapareguts tot i que hi ha autors que no descarten que alguns d’aquests productes fossin tòxics.
Aquestes consideracions plantegen també la possibilitat teòrica que la vida s’hagi originat a la Ter-ra més d’una vegada, i totes les vegades excepte una s’hagi extingit. Tanmateix, a la pràctica, la gran semblança entre tots els organismes coneguts pel que fa als aspectes bioquímics i genètics veritablement bàsics, sembla confirmar l’origen comú de tots ells.
Les bases moleculars de la vida
És palès que en tots els organismes actuals hi ha un cert nombre de tipus de molècules sense les quals no es pot concebre l’existència de vida. La seva presència universal en totes les cèl·lules semblaria indicar que són molècules que ja eren presents en els ancessors més antics comuns a tots els éssers vius actuals als quals podem remuntar-nos. Per altra banda, aquestes molècules estan relacionades amb les característiques bàsiques comunes a tota cèl·lula, i sense elles cap organisme no podria sobreviure. Es tracta del trifosfat d’adenosina (ATP), dels àcids nucleics i dels fosfolípids.
L’ATP
En primer lloc, totes les cèl·lules posseeixen trifosfat d’adenosina (ATP), la presència del qual és necessària per a les reaccions que requereixen altes dosis d’energia, i això tant si la seva font primària d’energia és la llum del Sol o l’assimilació de molècules orgàniques procedents de la ingestió d’aliments.
Fins i tot les cèl·lules més senzilles necessiten aliment i energia per créixer i multiplicar-se. L’energia obtinguda de l’ATP permet a les cèl·lules produir àcids nucleics i proteïnes a partir de precursors presents al medi. Com que, tal com hem indicat, s’han pogut obtenir fàcilment molècules d’ATP a partir de models d’hipotètiques atmosferes arqueanes en presència de fosfats i, per altra banda, la seva presència als organismes és universal, no es pot descartar que les primeres cèl·lules obtinguessin energia pel camí més directe, utilitzant l’ATP com a aliment; encara més, possiblement usaven com a aliment tant l’ATP com algunes altres molècules d’elevat contingut energètic relacionades amb l’ATP, com ara el trifosfat de guanosina (GTP) i el trifosfat d’uricina (UTP). Amb el creixement de la població cellular el ritme de consum d’aquestes substàncies fàcilment degué superar moltes vegades el ritme de síntesi espontània, i només les cèl·lules que pogueren desenvolupar els seus propis mecanismes interns per produir-ne arribaren a sobreviure. Probablement un dels primers d’aquests mecanismes fou la fermentació, que és, encara avui, el més utilitzat i del qual ens ocuparen més enda-vant.
Els àcids nucleics
També totes les cèl·lules contenen sistemes de replicació basats en els àcids nucleics, sense els quals ni la reproducció ni el creixement serien possibles.
La replicació exacta de les molècules dels àcids desoxiribonucleics (DNA) és essencial per a la reproducció dels organismes i per al manteniment de la identitat de l’espècie. Les modificacions esporàdicament introduïdes en aquesta replicació són la base bioquímica de l’evolució i expliquen l’origen, des d’un nombre limitat de cèl·lules format fa més de 3 500 milions d’anys, dels centenars de milers d’espècies de plantes, animals, fongs protoctists i moners avui existents i també de les existents en altres períodes i avui extingides.
Les molècules de DNA són polímers, és a dir llargues cadenes d’unitats semblants: els anomenats nucleòtids. Cada nucleòtid és format per un grup fosfat, una molècula de sucre (desoxiribosa) i una base nitrogenada. El nombre de nucleòtids pot ser molt gran, fins i tot de milions, però en canvi només intervenen en la seva composició quatre únics tipus de bases nitrogenades que poden ser l’adenina, la citosina, la guanina o la timina.
Les molècules d’àcid ribonucleic també són polímers de nucleòtids però en aquest cas el sucre que en forma part és la ribosa; les bases nitrogenades poden ser adenosina, citosina, guanina o uracil, i les cadenes són generalment molt més curtes.
Els fosfolípids
Finalment, totes les cèl·lules posseeixen molècules de fosfolípids, indispensables per a la formació de membranes que les individualitzen i en regulen els intercanvis amb el medi. Els fosfolípids contenen radicals d’àcids grassos i grups fosfat, i constitueixen cadenes amb propietats molt dissemblants als seus dos extrems; un d’ells, el “cap”, té càrrega elèctrica i és hidròfil (pot interactuar amb l’aigua), i l’altre, la “cua”, és hidròfob i només interactua amb dissolvents orgànics. En medi aquós, aquestes molècules tendeixen inicialment a agrupar-se en petites estructures esfèriques amb les “cues” dirigides cap al centre i els “caps” hidròfils cap a l’exterior sempre que les “cues” siguin prou flexibles i no gaire llargues. Si són més rígides o més llargues es comencen a aplegar en una làmina que forma una doble capa amb les “cues” capiculades i els “caps” girats cap a l’exterior, la qual pot tancar-se sobre ella mateixa i formar una esfera que clogui un medi aquós interior separat de la resta del solvent.
El procés biogènic
Així doncs, si bé és cert que no es pot descartar totalment la hipòtesi que la vida actual a la Terra hagués pogut tenir avantpassats amb característiques moleculars diferents de les que mostren els organismes actuals i que no es pot excloure que moltes formes en què es manifesta la matèria viva s’extingissin abans d’originar-se la que ha arribat fins a avui, també és cert que si alguna vegada existí una forma de vida basada en uns principis químics diferents dels que ara la regeixen, desaparegué sense deixar rastre. Avui només podem retrocedir fins als avantpassats que comparteixen amb la totalitat dels organismes actuals (dels bacteris més primitius fins als humans) la maquinària de duplicació i autososteniment basada en la química dels àcids nucleics, en l’acumulació d’energia als enllaços del trifosfat d’adenosina i en la regulació del medi intern mitjançant intercanvis amb l’exterior a través de membranes semipermeables.
Però quina era la natura d’aquests avantpassats comuns? A partir de les comparacions de seqüències de gens comunes a les tres grans línies evolutives reconegudes actualment (bacteris, arqueus i eucaris), es pot concloure que l’ancessor compartit havia de ser ja un organisme força semblant als procariotes actuals, amb un genoma basat en el DNA i un aparell de transcripció i transferència basat en els RNA i en l’enzim RNA-polimerasa i amb una colla dels enzims més importants per al metabolisme cel·lular, incloent-hi alguns dels responsables del transport actiu a través de les membranes i altres implicats en diferents vies metabòliques.
Retrocedir més enrere significa endinsar-se pel camí d’hipòtesis més o menys consistents però sempre discutibles. No és inversemblant que aquell avantpassat comú de tots els organismes actuals hagués estat precedit per una llarga sèrie d’organismes més primitius en els quals les síntesis de proteïnes, els genomes a base de DNA, la biosíntesi dels components de la membrana i els sistemes de transducció d’energia haguessin aparegut, no com a resultat de processos químics prebiòtics sinó de processos biològics de caire evolutiu.
L’evolució prebiòtica
Ja als anys vint el bioquímic rus Aleksandr I. Oparin i el fisiòleg anglès John B.S. Haldane havien assenyalat que un dels requisits indispensables per a l’evolució de la matèria viva a partir de molècules orgàniques prebiòtiques era l’absència a l’atmosfera d’oxigen lliure; altrament l’oxigen hauria reaccionat activament amb elles i no n’hagués permès el manteniment. Aquests primers investigadors que plantejaren en el pla experimental hipòtesis sobre l’origen de la vida a la Terra postulaven també l’existència d’un “brou” prebiòtic primitiu al si dels oceans de la Terra encara despoblada d’éssers vius. La dificultat que representa la feble concentració de matèria orgànica que podria resultar de la dissolució al si de grans oceans de la matèria orgànica sintetitzada a l’interior de l’atmosfera ha fet que molts autors es decantin actualment per la hipòtesi d’un origen de la vida vinculat a bassals poc profunds o àrees litorals afectades per les marees o batudes per les ones.
Sense oxigen lliure que hi pogués reaccionar i destruir-les, les molècules d’aminoàcids, nucleòtids i sucres podien mantenir-se sense alteració en solució o, encara millor, en adsorció sobre alguna superfície. En aquestes condicions una molècula com la de l’ATP podia formar-se a partir de la unió de l’adenina amb la ribosa (al capdavall un sucre senzill, amb una molècula dotada de cinc àtoms de carboni) i tres grups fosfat.
Algunes molècules pogueren esdevenir catalitzadors que facilitessin i acceleressin la unió o la separació d’altres molècules sense resultar destruïdes per la reacció, ni tan sols ser alterades per ella. Els catalitzadors foren (i són) importants per a la matèria viva pel fet que introduïen ordre i organització en uns processos químics que, sense ells, es produïen a l’atzar. Gradualment, aquests catalitzadors i les reaccions que facilitaven havien de proliferar més que altres combinacions a les aigües de la Terra primitiva. Tot i que eren cada cop més complexos, no havien de requerir pas per això més energia. Avui dia, alguns grups de molècules poden autocatalitzar ordenadament o cíclicament tot un seguit de reaccions sorprenentment intricades, de tal manera que cada canvi actua sobre un altre al llarg de la cadena molecular. Algunes d’aquestes reaccions autocatalítiques “inerts” donen lloc a models cada vegada més complexos; aquesta creixent complexitat podria, amb el temps, arribar a recordar fenòmens propis de la matèria viva.
L’autoorganització hipercíclicam
Tant els càlculs teòrics com les proves de laboratori semblen suggerir que de la interacció entre dos o més cicles autocatalítics podria haver resultat un “hipercicle”. Segons alguns investigadors, però, si hagués estat així, uns catalitzadors d’aquesta mena haurien entrat en “competició” pels elements del seu medi i això mateix n’hauria limitat automàticament el nombre. Tanmateix, la idea bàsica de l’hipercicle és ben bé l’oposada: ben al contrari de destruir-se mútuament en una “lluita per l’existència” química, els compostos autoorganitzats s’havien de complementar per produir, al capdavall, per replicació, estructures semblants a la matèria viva. Aquesta mena de processos cíclics formaren versemblantment les bases no sols de les primeres “cèl·lules” sinó de tota una colla d’estructures, basades en cèl·lules i en les seves produccions, que les seguiren. Els processos cíclics són molt importants per a la vida; permeten als éssers vius de preservar elements clau de llur passat a desgrat de les fluctuacions i de la tendència al desordre del seu entorn més ampli. A més, aquest acoblament de processos permet d’aprofitar l’energia que s’allibera en reaccions exotèrmiques per a facilitar l’esdeveniment de les endotèrmiques que, d’altra manera, no serien possibles.
Com més protegides i concentrades eren les molècules orgàniques, més prolongada, complexa i autoreforçada podia esdevenir la seva activitat. Algunes podien haver estat protegides per una bombolla o enganxades a les superfícies d’argiles o cristalls. Als temps arqueans més reculats, els experiments de la natura amb llargues cadenes hidrocarbonades hagueren d’arribar a produir molècules del tipus dels fosfolípids, capaces d’encapsular una goteta de l’aigua que les envoltava, juntament amb allò que contingués aquesta, bo i permetent encara el moviment d’altres molècules endins i enfora del clos que delimitaven. Això és tant com dir la mínima expressió d’una membrana semipermeable envoltant una “cèl·lula”.
L’aparició de les membranes
És indubtable que la formació de membranes ha tingut un paper cabdal en l’aparició de les primeres cèl·lules. En els organismes actuals les membranes es componen de diverses menes de lípids, proteïnes i glúcids, i les seves funcions són tan complexes i calibrades amb tanta precisió que encara som lluny de comprendre-les del tot. Probablement, però, les primeres membranes eren molt més simples, poca cosa més que una doble capa de fosfolípids que, a diferència d’altres estructures encapsuladores possibles, aconseguia, en virtut de les seves propietats químiques, concentrar una dissolució d’altres compostos de carboni; així podia atrapar al seu entorn immediat diferents substàncies (“nutrients”) susceptibles d’interactuar al seu interior mentre impedia que sortís l’aigua interna.
L’aparició de la membrana obrí la possibilitat que es produís la primera unitat discreta del món viu: la cèl·lula procariota. Però la majoria dels investigadors pensen que, abans, les proteïnes i els lípids es combinaren en flocs translúcids de matèria encara no viva abans del començament de la vida pròpiament dita. No es coneix cap forma de vida sense una o altra mena de membrana. La membrana permet d’aïllar una porció de l’entorn i mantenir-la en unes condicions i unes propietats diferents de la matèria del seu voltant.
Probablement no ha estat una vegada sinó moltes que, sota la protecció d’una bombolla de lípids, aminoàcids, sucres senzills, fosfats i els seus derivats s’han format i s’han complexificat, utilitzant l’energia solar i absorbint ATP i altres compostos de carboni i de nitrogen de l’exterior com a “aliment”. Algunes d’aquestes bombolles de lípids s’haurien pogut dividir primer simplement a partir de la força de la tensió superficial bo i mantenint cada meitat la seva activitat interna. Més tard, molècules catalitzadores adequades poden haver començat a mantenir activament les membranes lipídiques. Potser, quan s’exhaurien els components necessaris disponibles al limitat nínxol que ocupaven, les protocèl·lules simplement dequeien i desapareixien, mentre se’n formaven d’altres en un altre bassal deixat per la marea, cada una amb un funcionament lleugerament diferent de les altres.
Membranes semipermeables
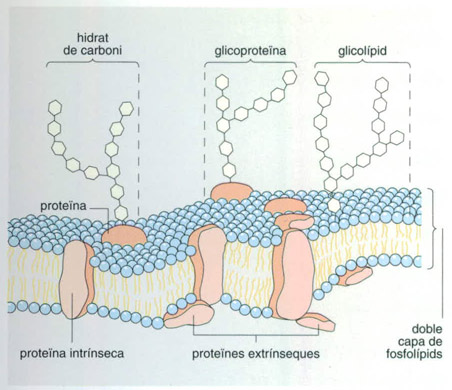
Editrònica, a partir de fonts diverses
La membrana aïlla la cèl·lula del medi. La vida tal com la coneixem comparteix unes característiques bàsiques que hem de suposar que ja existien en els primers organismes vius o, si més no, en les primeres manifestacions de la vida que no s’extingiren; acumular energia en els enllaços d’ATP, duplicar-se per mitjà dels àcids nucleics i regular el medi intern mitjançant els intercanvis amb l’exterior a través de membranes semipermeables. La formació de membranes va lligada a la mateixa naturalesa dels fosfolípids, molècules fortament polars, que per un extrem són hidròfiles i per l’altre hidròfobes, les quals, en medi aquós, tendeixen a agrupar-se en petites esferes amb les cues hidròfobes cap endins i els caps hidròfils cap enfora. Aquestes molècules poden tancar una goteta d’aigua i permetre un intercanvi de molècules entre l’espai clos a l’interior i l’espai exterior; aquest és el principi de la formació de les membranes semipermeables, una fita que determinà la formació de les primeres cèl·lules.
La tendència a l’autopoesi
Un ésser viu, per a ser-ho de debò, ha de ser autopoètic, és a dir, s’ha de mantenir ell mateix activament enfront de les agressions de l’exterior. La vida respon a les pertorbacions amb l’ús de matèria i energia per romandre intacta. Un organisme intercanvia constantment les seves parts i renova els seus components químics sense perdre mai la identitat. Els organismes vius tenen la feina d’intentar mantenir les condicions en què varen poder aparèixer i desenvolupar-se. Aquest fenomen modulador, “holístic”, de l’autopoesi, de l’activa autoconservació, és a la base de tota vida coneguda; totes les cèl·lules reaccionen enfront de les pertorbacions externes per tal de conservar aspectes clau de la seva identitat dins dels seus límits.
Una vegada capaç de mantenir-se, una estructura com la de les protocèl·lules, per tal d’esdevenir viva havia d’assolir la capacitat de reproduir-se. Els sistemes protocel·lulars eren allò que el belga d’origen rus Ilya Prigogine anomenà “estructures dissipatives”, és a dir objectes o processos que s’autoorganitzen i canvien de forma espontàniament. Amb una aportació d’energia les estructures dissipatives poden esdevenir més ordenades en comptes de menys.
A partir d’estructures dissipatives i d’hipercicles emergiren les cadenes de nucleòtids, ribosa i fosfats que d’una banda poden autoreplicar-se, i de l’altra catalitzar reaccions químiques; els RNA, la primera expressió del llenguatge de la natura. Encara no autopoètics però altament estructurats, els RNA primitius es devien acumular en petites esferes envoltades de tirallongues de lípids a les aigües càlides i riques de matèria orgànica sobre una terra benigna. Sense predadors i amb abundància d’energia disponible havia de prosseguir el procés de complexitat creixent que ha arribat a donar formes d’equilibri tan delicat i amb interaccions tan refinades.
Ja abans que emergís pròpiament la vida, a la Terra hadeana, s’havien hagut de manifestar versemblantment dues tendències químides: autoreferències i autocatàlisi. Reaccions cícliques de les substàncies químiques que produïen versions i versions d’elles mateixes que tendien a crear un ambient favorable a la repetició de les reaccions originals; un pas més i havien de poder començar a organitzar-se estructures autopoètiques capaces d’utilitzar energia per mantenir-se activament i amb èxit enfront de pertorbacions externes importants, i dotades de límits cada cop més clarament distints. En algun moment, fa més de 3 500 milions d’anys (i potser fins i tot més de 3 900) una d’aquestes estructures, delimitada per una membrana lipídica, i amb proteïnes i RNA dins seu, començà a multiplicar-se: eren la primera manifestació del que avui entenem per vida, un grapadet microscòpic de cèl·lules semblants a les dels bacteris d’avui. Un cop l’autopoesi assegurava la seva supervivència i la reproducció en garantia l’expansió, l’evolució es posava en camí. El temps dels procariotes havia començat.
L’evolució sense oxigen
Tal com ja hem assenyalat, l’evolució prebiòtica es produí en absència d’oxigen lliure a l’atmosfera de la Terra. Els primers bacteris, doncs, visqueren en un món anòxic. Això comporta una diferència molt important entre aquells primers bacteris i la majoria dels actuals, per als quals l’oxigen gasós és un element indispensable: per a aquells primeríssims éssers vius l’oxigen era un tòxic molt perillós. De fet, avui la Terra continuaria tenint molt poc oxigen lliure, com els seus veïns Mart i Venus, si no hagués estat precisament per l’acció metabòlica dels bacteris que donava oxigen com a producte resultant. L’oxigen no és un component habitual dels gasos volcànics i només se’n forma una petita quantitat a l’alta atmosfera gràcies al trencament de molècules d’aigua per l’acció que provoquen els raigs ultraviolats. Atesa la seva activitat química, l’oxigen tendeix a reaccionar no solament a l’atmosfera terrestre sinó també la de tots els planetes. Només la prodigiosa producció d’oxigen pels primers bacteris (i les algues i les plantes posteriorment) asseguren la seva presència en el planeta.
Les primeres estratègies bioquímiques
És molt probable que els primers organismes s’assemblessin als organismes més petits coneguts actualment. El DNA que contenen les cèl·lules més petites és insuficient per a poder fabricar tots els aminoàcids, nucleòtids, vitamines i enzims que necessiten per al seu manteniment i la seva reproducció; la situació dels primers organismes no devia ser gaire diferent. Els nostres ancessors bacterians portadors de DNA, probablement de menys de 0,5 µ de diàmetre, evolucionaren a partir d’un món de RNA sense DNA ni proteïnes. Sabem que les molècules de RNA catalitzen, com les proteïnes, i es repliquen, com el DNA. Aquesttes cèl·lules amb RNA probablement foren els primers éssers vius de la Terra. La majoria dels bacteris més petits que han estat ben estudiats són paràsits, i prenen el que necessiten de l’animal o la planta dins dels quals viuen. Les condicions eren igualment severes en els temps hadeans. Sense la barrera de la capa d’ozó i en absència d’oxigen, les primeres cèl·lules vives es podien proveir lliurement dels compostos orgànics acumulats al medi gràcies a l’acció de la radiació solar i les descàrregues elèctriques sobre mixtures de gasos atmosfèrics rics de carboni i nitrogen. Les primeres cèl·lules, doncs, foren probablement heteròtrofes, capaces d’alimentar-se de molècules orgàniques del mateix tipus que les que les componien: aminoàcis, sucres, àcids orgànics de cadena curta i ATP.
Però arribà el dia que aquesta mena de disbauxa química s’acabà. Aquells nutrients eren ràpidament exhaurits pels microbis que els aprofitaven per a créixer i multiplicar-se de manera incessant a cada petit espai que ocupaven. Sens dubte, en els primers milions d’anys de la implantació de la vida, cada crisi de subsistència, cada canvi de clima o cada intoxicació ocasionada per la pol·lució deguda als mateixos gasos residuals del metabolisme dels microbis, era causa de l’extinció d’algun (i probablement, alguna vegada, de la majoria) dels claps de vida existents a la Terra. La vida degué fluctuar segons la taxa a la qual el Sol podia ocasionar la síntesi de més nutrients. De fet, la vida hauria pogut desaparèixer del tot ràpidament si no hagués estat per un tret vital fonamental: la capacitat del DNA d’autoreplicar-se i de poder introduir en algunes còpies a l’atzar canvis que donen cèl·lules mutants.
Si la duplicació del DNA és necessària per a la continuïtat de la vida, la mutació, en el sentit ampli de l’expressió “canvi hereditari”, és absolutament necessària per als processos evolutius. Sense mutació no hi ha la “descendència amb modificació” que requereix la formulació darwiniana de l’evolució: sense mutació la selecció no troba variació que asseguri continuïtat si hi ha canvis ambientals. Mercès a la seva petitesa i al seu enorme nombre, el creixement de les poblacions de microbis respon amb relativa facilitat a la majoria dels canvis ambientals majors. Si en el medi tenen accés a aliment i energia, els bacteris creixen i es reprodueixen amb un ritme exponencial. Un bacteri ràpid pot trigar entorn de vint minuts a créixer prou per a dividir-se i donar dos bacteris. D’un sol bacteri, en resultarien en dos dies 2144 individus, xifra que supera el nombre d’humans que mai hagin viscut. En quatre dies de creixement il·limitat en resultarien 2288 bacteris, xifra que supera el nombre de protons (al voltant de 2266) o de quarks que els físics han estimat que hi ha a l’univers.
Aquest potencial biòtic de reproducció no es dóna en la realitat, però serveix per a recordar el poder d’una reproducció exponencial. Entorn d’un cop per cada milió de divisions apareix un descendent que no és idèntic al seu pare: és un mutant. La majoria dels mutants no poden sobreviure perquè la seva diferència resulta negativa per autoconservar-se o per reproduir-se, però alguns tenen èxit, poden créixer i reproduir-se, i es multipliquen ràpidament en el medi que ocupen. Totes les variables del medi (temperatura, qualitat i quantitat de la il·luminació, concentració de sals a l’aigua) tendien a difersificar les poblacions de microbis segons els llocs, seleccionant els mutants més ben adaptats a les seves condicions ambientals concretes.
Els processos fermentatius
Una de les primeres innovacions metabòliques seleccionada pel seu valor adaptatiu fou probablement la que capacità els bacteris per a usar els sucres per tal d’obtenir energia i emmagatzemar-la (per producció de molècules d’ATP a partir d’ADP i fosfat). Els compostos derivats del sucre (alcohols i àcids) eren excretats com a residus. És el que s’anomena fermentacions, que es produeixen en absència d’oxigen lliure, i algunes de les quals encara són funcionals en molts organismes.
Les fermentacions utilitzen com a substrat de partida sucres senzills com la glucosa o la fructosa, alcohols, àcids orgànics o fins i tot hidrats de carboni amb cadenes llargues de sucres, com ara la cel·lulosa o el midó. També poden partir de compostos senzills que continguin nitrogen, com els aminoàcids. El residu final pot consistir en anhídric carbònic i etanol, cas dels bacteris i llevats que fermenten els sucs de fruites o granes, que després de la fermentació seran begudes alcohòliques. Els productes d’excreció poden ser àcid acètic o àcid làctic, com en la fermentació del sucre per a formar vinagre (àcid acètic) o de la llet per a formar iogurts, o en la maduració de determinats formatges que contenen àcid làctic.
El procés de fermentació generalment proporciona al fermentador unes poques molècules d’ATP per cada molècula de sucre degradada. No és un procés totalment eficient, atès que els productes que en resulten (àcids i alcohols) encara contenen energia. L’evolució d’alguns bacteris fermentadors comportà una pressió de selecció per a l’aparició de nous bacteris capaços de degradar els residus dels fermentadors i d’obtenirne encara carboni i energia. Feien un paper semblant al que fan avui els organismes eucariòtics que s’alimenten d’excrements, com els escarabats piloters o els bolets de tinta. Cadenes alimentàries microbianes d’aquesta mena existeixen encara en sòls i llacs i en superfícies intermareals. També n’hi ha als intestins d’alguns insectes, com els tèrmits, i a les aigües estanyades; en definitiva a qualsevol lloc on hi hagi hidrats de carboni per fermentar i l’oxigen i la llum siguin escassos. Aquesta capacitat fermentativa ancestral és emprada com estratègia per alguns organismes força més moderns com és el cas d’alguns animals marins de respiració preferentment aeròbica. Les cèl·lules musculars de les nostres cames, quan ens fatiguem, per exemple pujant escales, abandonen momentàniament el seu metabolisme aerobi i reprenen l’antic tipus de fermentació; encara que menys eficient com a mitjà de derivar ATP, aquest metabolisme fermentatiu, llegat de la nostra història passada, es manté en tots els animals inclosos els humans.
La fixació de nitrogen atmosfèric
Uns dels fermentadors actuals que probablement s’assemblen més als primers bacteris són els clostridis (Clostridium), alguns dels quals també han desenvolupat per selecció la capacitat d’aprofitar nitrogen en forma de gas de l’atmosfera com a font de nitrogen proteínic. Utilitzant enzims, trenquen l’enllaç del nitrogen gasós (N2) de l’atmosfera, i el fixen en molècules d’aminoàcids, nucleòtids i altres compostos orgànics. Amb això, posen l’enorme font de nitrogen atmosfèrica a disposició de la resta de la biofera.
Fixar nitrogen requereix una considerable quantitat d’energia (de 6 a 18 molècules d’ATP per molècula de nitrogen). Fer-ho a gran escala, com es fa per exemple industrialment a les plantes productores de fertilitzants químics, exigeix altes pressions (300 atmosferes) i temperatures (500°C). Cap planta, protoctist, fong o animal no és capaç de fer-ho a baixa temperatura ni, en realitat, gaires bacteris. Tots els organismes depenen, per a la seva supervivència, dels bacteris fixadors de nitrogen; sense ells la vida desapareixeria de la Terra per carència de nitrogen, tot i ésser aquest el component majoritari de l’actual atmosfera terrestre. Les espècies de Clostridium, juntament amb altres grups de bacteris d’evolució més tardana, com ara les espècies d’Azotobacter i Rhizobium de les arrels de les lleguminoses o alguns cianobacteris, subministren compostos nitrogenats a tota la biosfera des de l’Arqueà.
Certament, les poblacions primitives de fermentadors (tot i que segurament alguns fermentadors) també podien usar el diòxid de carboni de l’aire) encara depenien per a la seva nutrició de compostos produïts al medi, però a mesura que els compostos orgànics produïts prebiòticament eren consumits i esdevenien escassos, s’havien d’intensificar les pressions de selecció. Potser la vida no hagués pogut continuar si no s’haguessin desenvolupat noves estratègies que fessin capaços els organismes de produir ells mateixos la seva energia i els seus propis nutrients a partir de materials del medi.
La reducció de sulfats de l’aigua
Una de les primeres vies metabòliques que es pot considerar com un pas endavant en el camí cap a l’autotròfia va sorgir en el llinatge del grup de bacteris actualment representat pel gènere Desulfovibrio, que són capaços de “respirar” sulfats i de generar ATP a partir de la conversió de sulfat en sulfur mitjançant la transferència d’electrons d’alta energia des d’alguna de les etapes d’un procés de fermentació fins al sulfat. Els desulfovibris, igual que altres bacteris semblants (Desulfotomaculum, Desulfuromonas, etc.) viuen en sediments anaeròbics que contenen matèria orgànica i l’ió sulfat (SO42–), en llots dels fons lacustres i marins. Poden fermentar compostos orgànics com l’àcid làctic i l’àcid pirúvic produint àcid acètic però obtener més ATP si simultàniament poden transformar grup sulfat en sulfur (S2–), el qual pot ser alliberat al medi en forma de sulfur d’hidrogen (H2S) o de sulfur de dimetil (H3CSCH3), gasos tots dos de notable fetidesa, d’olor semblant al d’ous podrits o algues en descomposició.
La reducció del sulfat es fa mitjançant molècules de proteïnes unides a anells de carboni característics anomenats porfirines, entorn dels quals s’organitzen cadenes proteíniques més o menys llargues; cada anell duu un àtom d’algun metall al centre. El complex de porfirina i metall rep el nom de grup hemo. Els grups hemo també són freqüents en els complexos proteínics anomenats citocroms que intervenen en la “respiració” de sulfats bacteriana. També són presents en l’hemoglobina de la sang dels animals, on intervenen en el transport de l’oxigen des dels òrgans de respiració fins a les cèl·lules.
L’accés a la fotosíntesi
Els complexos de porfirines i proteïnes com ara els citrocroms han d’haver estat una adquisició evolutiva força primerenca perquè de fet, els únics que no tenen la capacitat de sintetitzar grups hemo són els bacteris que generen ATP exclusivament per metabolisme fermentatiu. Per contra, amb l’adquisició de la capacitat de produir anells de porfirina, uns determinats tipus de bacteris pogueren desenvolupar la capacitat d’utilitzar la font d’energia més segura i abundosa: la llum del Sol.
Quan una molècula rep llum, els seus electrons són empesos a un nivell energètic superior. Normalment aquesta energia tot just és dissipada en forma de llum o de calor, i la molècula retorna al seu estat inicial d’abans de rebre la llum. Però quan aquestes molècules estan en contacte amb porfirines lligades a proteïnes encastades a membranes, com en els reductors de sulfats, l’energia lluminosa pot ser retinguda i utilitzada en forma d’energia química vinculada a l’ATP. L’energia de l’ATP pot ser usada a continuació per al moviment i per a processos de síntesi, com ara la conversió del diòxid de carboni de l’atmosfera en aliment, i també en la replicació dels àcids nucleics necessaris per a l’automanteniment i el creixement. Aquest procés bacterià d’obtenció d’aliment a partir de la llum i l’aire (la fotosíntesi) alliberà completament alguns grups de bacteris de la seva dependència de compostos orgànics preformats.
Tots els organismes fotosintetitzadors no bacterians contenen alguna de les formes de clorofil·la. Aquest pigment verd pot presentar petites variacions però està basat, fonamentalment, en un anell porfirínic que conté com a grup metàl·lic un àtom de magnesi. Així doncs, l’aparició de la fotosíntesi només va ser possible mercès a l’evolució precedent de la síntesi de les porfirines.
Els primers organismes fotosintetitzadors del planeta eren bacteris que usaven l’hidrogen gas o el sulfur d’hidrogen per a reduir el diòxid de carboni; no produïen oxigen, però feien la fotosíntesi tal com la fan avui els anomenats bacteris vermells i verds del sofre, a partir dels quals es pot reconstruir parcialment l’evolució inicial de la fotosíntesi. Es coneixen cinc tipus principals de bacterioclorofil·la diferents, els quals són sempre suplementats per unes molècules anomenades carotenoides; aquests són pigments vermells i grocs que actuen com a accessoris de les clorofil·les, ja que absorbeixen llum de longituds d’ona que les clorofil·les no poden aprofitar i els transfereixen l’energia captada del Sol. El conjunt dels diferents pigments, altres lípids i proteïnes s’aplega en unes membranes que s’anomenen tilacoides.
L’evolució de la capacitat fotosintetitzadora
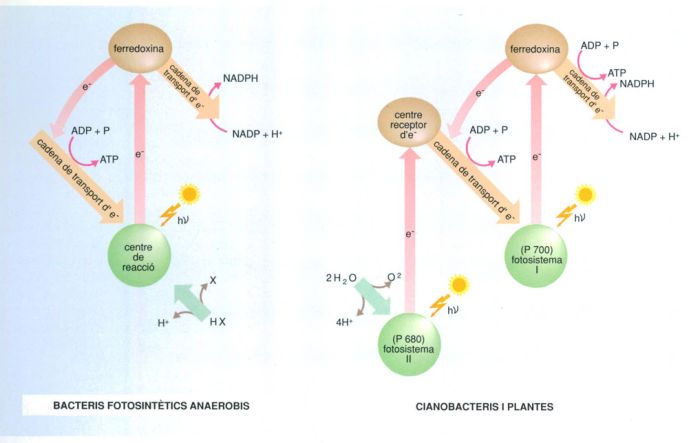
Biopunt, a partir de fonts diverses
La fotosíntesi és el mecanisme pel qual determinats organismes aprofiten l’energia despresa pels electrons excitats per la llum. Determinats bacteris existents actualment (els bacteris vermells i verds del sofre) són capaços de fer fotosíntesi anaeròbica, procés que fou seguit durant milions d’anys en el passat; la utilització de la llum solar com a font d’energia aparegué com a culminació del procés d’evolució bioquímica iniciat pels bacteris fermentadors, anaeròbics, als quals seguí l’aparició de la fotosíntesi anaeròbica de bacteris dotats només de fotosistema I (que no escindeix l’aigua i necessita donadors d’hidrogen, d’altra banda abundants en aquell moment); a continuació l’aparició de procariotes oxigènics, els cianobacteris, dotats de fotosistema II a més de l’I (capaços, per això, de trencar l’aigua i desprendre oxigen) i finalment l’aparició dels bacteris aeròbics. Aquest progrés es pogué anar produint gradualment, a mesura que augmentava la quantitat d’oxigen en l’atmosfera, com a conseqüència de l’activitat dels cianobacteris.
El procés de la fotosíntesi té dues fases diferents, cada una de les quals consisteix en un conjunt de reaccions químiques; la primera d’elles utilitza directament la llum (fase lumínica) mentre que la segona n’és independent (fase fosca). En la primera fase, els electrons de la clorofil·la són empesos fins a un nivell energètic superior. La molècula excitada de clorofil·la d’un tilacoide transmet la seva energia a una molècula adjacent i aquesta a una altra fins a assolir-ne una d’especial, present en cada unitat d’empaquetament de la clorofil·la en el tilacoide, anomenada centre d’acció: és en aquest centre d’acció on els electrons d’alta energia generen ATP. El destí final d’aquests electrons que es mouen pot ser diferent segons que tota la seva energia sigui emprada a generar ATP (cas en què retornen a una molècula oxidada de clorofil·la) o que la seva energia només sigui parcialment usada en la gènesi d’ATP; en aquest cas els electrons que encara conserven certa energia són transferits a una molècula de nicotinamida-adenina-dinucleòtid fosfat (NADP), la qual es redueix a NADPH. Aquest compost orgànic reduït serveix com a font d’energia i d’àtoms d’hidrogen a la fase fosca, en la qual es realitza la reducció del CO2 a compostos com ara la glucosa; és a partir d’aquests compostos resultants de la fotosíntesi que les cèl·lules poden formar el propi material cel·lular o acumular reserves nutritives per al seu ús posterior.
L’estudi detallat de les seqüències de nucleòtids en le molècules de RNA ribosòmic de centenars de bacteris demostra que se’n poden separar dos grups: els arqueobacteris i els eubacteris. Tots els bacteris fotosintetitzadors pertanyen al grup dels eubacteris. Els arqueobacteris no són fotosintetitzadors, però hi ha una notable i probablement antiga excepció: els halobacteris portadors de rodopsina. Aquests organismes de color taronja o vermellós utilitzen la llum del Sol com a font d’energia amb la qual generaran ATP intracel·lular, però no poden pas emprar la llum del Sol per a fabricar aliments. Al contrari, utilitzen aliments, en forma de molècules orgàniques, per a fabricar el seu sistema fotosintetitzador no clorofíl·lic.
És indubtable que la crisi de subsistència que amenaçava els fermentadors arqueans fou superada mercès a l’evolució de la fotosíntesi en aquells dos tipus de bacteris. En acumular reserves de nutrients a les seves cèl·lules, no solament es nodrien ells mateixos sinó que proporcionaven aliment a molts heteròtrofs. Comunitats pròsperes de microorganismes fotosintetitzadors i d’altres heteròtrofs que en depenien es degueren desenvolupar àmpliament durant l’Arqueà. En són testimoni les enormes quantitats d’esquists i sílexs rics de carbó, de 3 400 milions d’anys d’antiguitat com a mínim, trobades a l’Àfrica del Sud. Alguns filons de carbó d’origen bacterià trobats en aquestes roques no tenen res a envejar als formats a partir de les selves del Carbonífer 3 000 milions d’anys més tard.
La fotosíntesi no es pot fer sense llum. Per això mateix, la capacitat de detectar-la i de desplaçarse per assolir-la esdevingueren adaptacions avantatjoses per als organismes fotosintetitzadors i per als que, sense ser-ho, en depenien. De fet, és molt possible que una certa motilitat vinculada a la recerca de l’aliment mitjançant un sistema senzill de detecció química ja existís abans que haguessin aparegut els primers bacteris fotosintetitzadors. En els fangs anaeròbics de l’Arqueà uns microorganismes heteròtrofs capaços de percaçar el propi aliment havien de tenir enormes avantatges sobre els seus competidors. Però a partir de l’aparició de la fotosíntesi la presència de llum solar i d’aigua lleugerament salada començaren a implicar la presència d’aliment a la superfície del planeta arqueà.
La mobilitat i la protección contra les radiacions
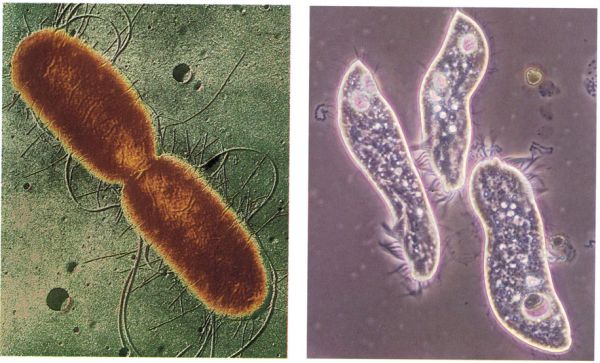
Els undulipodis, és a dir, els flagels i cilis de les cèl·lules eucariotes, com els del ciliat Chladotricha (a la dreta), sembla que deriven de bacteris similars a algunes espiroquetes actuals dotades de flagels bacterians, com aquesta Leptospira (a l’esquerra).
Biopunt, a partir de fonts diverses; fotos: Institut Pasteur (× 5 000) / CNRI i Francesc Torrella (× 420)]
L’origen del moviment en els bacteris sembla connectat amb el desenvolupament d’un dispositiu rotatori (inexistent en les cèl·lules eucariotes). Un filament en forma de fuet anomenat flagel va fixat a un “motor flagel·lar” que consisteix en un o dos corpuscles plans, un dels quals és implantat a la membrana cel·lular. Aquests discos roden mitjançant canvis de càrregues elèctriques i fan rodar el flagel; només en el cas de les espiroquetes el flagel és intern.
Tal com hem dit, els bacteris dotats de moviment gaudien d’un estimable avantatge: més facilitat d’accés a diferents llocs, a diferents fonts de nutrients; més exposició a materials genètics aliens, a productes químics d’altres organismes i per això mateix predisposats a desenvolupar simbiosis intricades en una àmplia varietat de medis. Tanmateix moure’s a la recerca de la llum també comportava un perill. La radiació solar incloïa no sols la llum que ara anomenem visible i que era la utilitzable pels bacteris fototròfics sinó també la ultraviolada, més potent i perillosa, que podia trencar els enllaços químics de proteïnes i àcids nucleics i causar la mort de la cèl·lula. Els microbis fototròfics hagueren de buscar diferents mecanismes de protecció, i els que els assoliren en la seva evolució s’asseguraren un avantatge addicional. El més immediat havia de ser procurar-se alguna mena de filtre. Viure en solucions riques en determinades sals com ara el nitrat de sodi, o sota la sorra o altres substàncies que deixessin passar la llum visible però no la radiació ultraviolada són troballes d’aquesta mena. Però alguns bacteris arribaren fins i tot a desenvolupar mecanismes semblants al nostre emmoreniment mitjançant el desenvolupament de pigments que absorbeixen les radiacions perilloses. Establiren així les condicions per poder construir colònies tan extenses que alteraren el món visible. Sobre la terra aigualosa, colònies de diferents tipus de microbis formaven en comú hàbitats compartits. Primer es devien fer visibles com a escumes tacades, claps de color porpra o siena o estranyes pastures estratificades. Els bacteris de la capa superior morien, una generació rere l’altra, a causa de l’exposició a la radiació, però les seves restes protegien els de les capes inferiors, les quals acumulaven fang, sorra i sediments orgànics per formar una mena de catifa viva. Tapissos i fangs microbians dominaven els paisatges d’aigües poc profundes de la Terra arqueana. Estructures d’aquesta mena es poden trobar actualment en moltes àrees costaneres càlides de tot el món com ara el golf Pèrsic i la costa nord-occidental d’Austràlia, i també en les grans explotacions salineres de la Baixa Califòrnia, a Mèxic, o del delta de l’Ebre, a Catalunya. Aquests tapissos bacterians actuals, d’aparença ben poc remarcable i molt de temps ignorats, són exemples vius, pràcticament fòssils vivents, d’un antic món dominat absolutament pels bacteris.
Estructura dels undulipodis
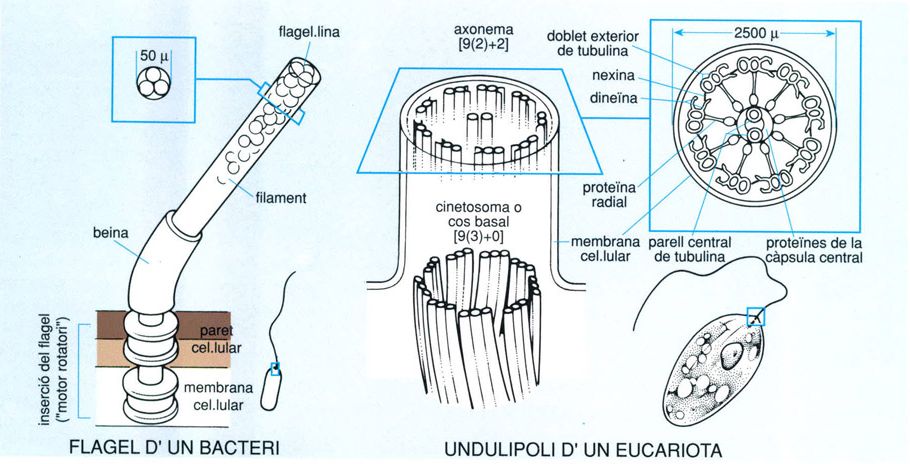
Biopunt, a partir de fonts diverses.
L’undulipodi de les cèl·lules eucariotes consta de dues parts diferenciades: una base, o cinetosoma, i un cos, o axonema. Vist en secció, el cos de l’undulipodi, que fa uns 250 nanòmetres de diàmetre, mostra un cercle de microtúbuls disposats per parelles, envoltant una parella central [9(2) + 2] i envoltat al seu torn per la membrana cel·lular. Els cinetosomes, en canvi, tenen nou triplets de microtúbuls, però manquen de la parella central [9(3) + 0]. Els flagels bacterians són molt més petits (uns 15 nanòmetres de diàmetre) i són formats per un sol filament fixat directament a la membrana cel·lular i amb una estructura mòbil giratòria a la base. Només aquesta estructura està dotada de moviment, mentre que en els undulipodis els filaments sencers poden moure’s ondulatòriament.
La reparació del DNA i la sexualitat
Els bacteris, a més de protegir-se amb una o altra mena de filtres contra l’excessiva radiació, desenvoluparen també mecanismes per reparar els possibles danys experimentats pel seu DNA, per exemple errors de duplicació entre cadenes de DNA complementàries a causa de la formació de dímers de timina; és a dir, aparellaments d’una timina amb una altra timina en comptes d’aparellar-se aquesta base amb la seva complementària, l’adenina. Aquesta mena d’accidents resultarien mortals per a qualsevol cèl·lula sense enzims reparadors que esmenessin el desgavell. Uns tipus d’enzims reparadors, els anomenats DNA polimerases, separen la part erròniament replicada i copien un nou DNA amb les bases correctament aparellades. Gairebé tots els organismes actuals tenen encara enzims reparadors, tot i que la capa d’ozó els protegeix de la radiació ultraviolada fa 2 000 milions d’anys. En molts bacteris, encara avui, aquests enzims només s’activen en presència de llum visible, tal com s’escauria si es mantinguessin les circumstàncies en què foren seleccionats com a defensa enfront dels inconvenients secundaris de l’exposició a la radiació solar.
Aquest sistema tan elaborat de protecció enfront de la radiació ultraviolada facilità una altra innovació d’una enorme importància: l’intercanvi o la fusió d’informació genètica entre diferents cèl·lules, altrament dit la sexualitat. Certament es tracta d’una sexualitat molt diferent de la que ara coneixem entre animals o vegetals de la mateixa espècie i de diferent sexe. Es tracta d’una sexualitat que no implica reproducció i en la qual només una de les fonts de gens ha de ser un organisme viu; l’altra pot ser-ho o pot ser un virus o fins i tot fragments de DNA d’una cèl·lula morta. A la Terra primitiva la sexualitat era simplement un mecanisme mitjançant el qual una cèl·lula bacteriana amb els gens alterats per la radiació ultraviolada podia obtenir-ne de nou a partir d’una altra font. La sexualitat bacteriana, en el seu origen, no condueix a la reproducció sinó a la recombinació de gens, com una carta trucada que facilita el manteniment dels bacteris en el joc de l’evolució i la supervivència d’una o altra població adaptada a qualsevol situació hostil que es donés en el medi.
L’oxigen, el primer contaminant a escala global
Al llarg dels primers temps proterozoics, fa uns 2 200 milions d’anys, la vida s’hagué d’enfrontar a la que probablement fou la primera crisi causada per la contaminació: l’emmetzinament massiu per l’oxigen. L’eó Proterozoic data del final de l’eó Arqueà (fa 2 500 milions d’anys) fins al començament del nostre eó, el Fanerozoic (fa uns 580 milions d’anys). La gran majoria dels organismes anaeròbics (tots els existents fins llavors ho eren) foren desplaçats per les formes emergents d’organismes aeròbics, capaços no sols de viure en presència d’oxigen sinó de fer-ne una de les bases del seu metabolisme mercès a la respiració.
L’origen de l’oxigen
De fet, la crisi s’inicià versemblantment a causa de la creixent escassetat de gasos lliures com hidrogen (H2) i sulfur d’hidrogen (H2S), la qual conduí a la selecció d’un grup de procariotes fotosintetitzadors capaços d’obtenir hidrogen de la seva font més abundant a la Terra: l’aigua. Eren els avantpassats del que avui anomenem cianobacteris, fins no fa gaire anomenats també equivocadament algues blaves o cianofícies. Sembla que es tractava de mutants de bacteris del sofre enfrontats a la crisi de subsistències que comportava l’enrariment de l’hidrogen i del sulfur d’hidrogen dels quals depenien.
Una descoberta feta el 1975 per un grup de microbiòlegs israelians a l’anomenat llac Solar, a la península de Sinaí, permet tenir una idea de com podien ser aquests primitius cianobacteris. En aquest petit llac artificial, caracteritzat pel fet que, a la inversa del que passa a la majoria dels llacs, l’aigua més calenta és la més profunda, descobriren un cianobacteri filamentós que es mou per lliscament al qual donaren el nom d’Oscillatoria limnetica. Es tracta d’un autèntic fòssil vivent, anella perduda entre els bacteris del sofre anaeròbics fotosintentitzadors i els cianobacteris actuals: si es troba en un ambient amb alta concentració de sulfur d’hidrogen i baixa concentració d’oxigen actua com un bacteri anaeròbic fotosintetitzador i utilitza el sulfur d’hidrogen com a subministrador principal d’hidrogen; però en ambients on la concentració de sulfur d’hidrogen és baixa i la d’oxigen és alta, actua com tots els altres cianobacteris i utilitza l’aigua com a subministrador principal d’hidrogen.
Posteriorment s’ha descobert que hi ha molts altres cianobacteris que poden revertir a la forma anaeròbica no oxigènica de fotosíntesi si hom els situa en les condicions apropiades. Molt probablement aquesta mena de jòquers de la fotosíntesi foren molt abundants abans del Cambrià, però es van veure desplaçats posteriorment pels especialistes d’una o altra forma de fotosíntesi i quedaren limitats a viure únicament en alguns medis extrems.
Els processos oxidatius
El creixement dels cianobacteris arqueans hagué d’abocar als mars i l’atmosfera quantitats insospitades d’oxigen lliure, la toxicitat del qual és molt gran a causa de la capacitat que té d’atrapar electrons i produir els anomenats radicals lliures, que són substàncies químiques de breu durada però molt reactives que poden desorganitzar els compostos orgànics. Les mateixes colònies de cianobacteris fotosintetitzadors que el produïen eren les primeres amenaçades, atès que eren les més pròximes a l’origen del gas “contaminant”.
Al començament, l’oxigen lliure que anaven produint podia reaccionar amb molts dels altres gasos presents a les aigües i a l’atmosfera arqueana, com ara l’hidrogen, el metà, l’amoníac, el monòxid de carboni o el sulfur d’hidrogen. Altrament també es podia combinar, oxidant-los, amb molts minerals elementals presents als oceans, rius i llacs com ara el ferro, el sofre, l’urani o el manganès. Precisament la presència d’òxids de ferro, manganès, urani i altres elements en roques de l’eó Proterozoic de 2 000 milions d’anys d’antiguitat (el període que va de 2 600 a 580 milions d’anys enrere) confirmaria que ja en aquelles dates hi començava a haver quantitats apreciables d’oxigen a l’atmosfera. Per contra, les roques d’edat arqueana (amb més de 2 600 milions d’anys d’antiguitat) que contenen generalment minerals de ferro, urani i sofre, tendeixen a ser més pobres d’oxigen, cosa que indicaria menor producció d’aquest element.
Un bon exemple del registre atmosfèric deixat en les roques seria la formació de Minechapecoten, a la regió de Woman River, a Ontario Occidental (Canadà), de més de 2 500 milions d’anys, molt rica en ferro. S’hi han trobat testimonis d’una gran massa d’aigua, un antic mar anaerobi, en la superfície dels marges de la qual eren dominants els desulfovibris. Els representants actuals d’aquest grup de bacteris no toleren gens l’oxigen, i per això viuen per sota de la superfície de l’aigua. En canvi, en aquesta formació dipositada al final de l’Arqueà, hi ha grans dipòsits de sulfurs, i això demostra que els desulfovibris podien reduir anaeròbicament el sulfat de l’aigua marina a sulfur utilitzant productes de la fotosíntesi dels cianobacteris com a combustible.
El cas de les formacions de ferro en bandes
Unes formacions geològiques que documenten la transició d’una atmosfera sense oxigen a una atmosfera oxigenada són les anomenades formacions de ferro en bandes (“banded iron formations”, o BIF, segons l’expressió anglesa). Aquestes formacions geològiques, que ocupen grans extensions a Austràlia, a l’Amèrica del Nord i a l’Àfrica, són molt importants perquè subministren més del 90 % de les reserves de ferro del món occidental. Daten sobretot del final de l’Arqueà i el començament del Proterozoic (fa entre 3 000 i 1 800 milions d’anys) i bàsicament consisteixen en una successió de capes que contenen ferro més oxidat (hematites) alternades amb altres que contenen ferro més reduït (magnetita) en el si d’una matriu de sílice (“chert”).
El gruix d’unes i altres bandes pot ser molt variable, n’hi pot haver de pocs micròmetres fins a altres d’uns quants metres. Les alternances entre dipòsits de ferro ferrós (reduït) i ferro fèrric (oxidat) podrien ser el resultat d’unes condicions de transport i sedimentació de ferro marcades per concentracions variables d’oxigen, conseqüència del creixement estacional de bacteris fotosintetitzadors productors d’oxigen en la superfície il·luminada de grans cubetes d’aigües somes, lacustres o marines. Els esclats estacionals de creixement i d’emissió d’oxigen produïts any rere any durant més de 500 milions d’anys haurien estat la causa de les bandes oxidades, de color més viu.
Els bacteris fotosintetitzadors no foren pas els únics que intervingueren en l’origen de les formacions de ferro en bandes. Alguns bacteris oxidadors del ferro poden captar oxigen del medi i obtenir-ne energia en aigües pobres en nutrients mitjançant la combinació d’aquest amb el ferro per a formar compostos oxidats; rovell, en una paraula. Els avantpassats arqueans d’aquests organismes devien proliferar entorn de les zones de producció d’oxigen i any rere any devien “depurar” els abocaments d’oxigen al medi precipitant-los en forma de rovell entorn de les comunitats de cianobacteris. Aquests bacteris oxidadors del ferro haurien participat en la formació de les quantitats enormes del mineral de ferro que avui s’explota a diferents formacions de ferro en bandes d’Àfrica, Amèrica i Austràlia, i les bandes alternades de minerals en diferents estats d’oxidació podrien ser un registre de les seves antigues relacions amb les colònies de cianobacteris. L’hematites s’hauria produït a l’estiu, quan els cianobacteris produïen més oxigen i els oxidadors del ferro eren més actius i abundants, mentre que les bandes de magnetita correspondrien a l’hivern, quan la producció fotosintètica d’oxigen i l’oxidació del ferro passaven per un mínim.
Al llarg d’una part del Proterozoic, entre els 2 200 i els 1 800 milions d’anys enrere, les formacions de ferro en bandes tingueren un esclat sense precedents i mai més no igualat, i sobtadament desaparegueren substituïdes pels anomenats “red beds” o bandes vermelles. Això podria ser conseqüència del fet que en aquelles dates la concentració d’oxigen a l’atmosfera ja s’havia acostat a l’actual de manera significativa.
El cas dels jaciments aurífers
Trobem un altre testimoni de les primeres activitats aeròbiques de la vida en els jaciments aurífers més importants i, en particular, els de Witwatersrand, al Transvaal (Sud-àfrica). L’or és un metall pesat rar a l’escorça terrestre; el que hi ha actualment hi va ser portat des del mantell profund en algun moment de l’eó Arqueà, i només contenen bandes de sediments aurífers una limitada sèrie de formacions d’edat arqueana.
Els treballadors que davallen pels muntacàrregues de les mines d’or del Transvaal, on es produeix el 70% de l’or de tot el món occidental, recorren una mena de túnel del temps per anar a trobar capes de cendres volcàniques i al·luvions deixats per rius arqueans sobre superfícies encara més antigues. Per localitzar nous dipòsits d’or es busquen unes capes —les anomenades “carbon leader”— de roca conglomeràtica amb un elevat contingut de carbó orgànic. Aquestes capes, atrapades entre calcàries i esquists, contenen fines venes de pirita, or i sovint també minerals d’urani. La “carbon leader” de Witwatersrand també conté estructures microscòpiques filamentoses i esfèriques que són inexplicables des del punt de vista purament mineralògic.
L’explicació més convincent és que es tracta de restes de bacteris filamentosos i coccoides que haurien atrapat els flocs detrítics d’or. Com que l’or es mou més fluidament en absència d’oxigen que en presència d’oxigen, hauria estat fàcilment erosionat de les roques per rius i torrents i transportat a la mar. Tanmateix, les partícules d’or, en presència d’altes concentracions d’oxigen i de carbó orgànic, tendeixen a deixar d’estar en suspensió i a formar flocs o grumolls que es dipositen al fons. Amb la seva producció d’oxigen i de compostos rics en carboni, qualsevol colònia de bacteris fotosintetitzadors determina una situació favorable a la floculació de l’or, de manera que podrien molt bé haver estat colònies d’aquesta mena les que haguessin determinat la floculació de quantitats considerables d’or al llarg de les lleres dels cursos d’aigua.
A més hi ha bacteris actuals, com ara Chromobacterium violaceum, capaços de produir àcid cianhídric, el mateix compost que és usat en els processos de cianuració destinats a separar l’or del mineral que l’acompanya. Tal vegada els avantpassats d’aquests microbis vivien als rius carregats de mineral de l’Arqueà ajudant a consolidar l’or floculat a partir de les partícules en suspensió mercès a l’oxigen i la matèria orgànica dels cianobacteris. Hom creu que la llera del riu al llarg del qual es va dipositar l’or de Witwatersrand fa 2 500 milions d’anys era cinc vegades més gran que l’actual delta del Mississippí; dessecada i colgada sota quilòmetres de sediments i sotmesa a plegaments, romangué ignorada fins que al segle passat els colonitzadors bòers foren atrets al Transvaal per les clapetes d’or d’unes roques fosques riques de carbó que afloraven al desert. Seguint l’aflorament cap a les profunditats del subsòl trobaren l’antic sistema fluvial enterrat del qual formava part la capa de carbó que indica la proximitat de la vena aurífera.
La informació dels estromatòlits
Fins aquí hem estat parlant sempre en termes d’hipòtesis fonamentades en dades teòriques i empíriques, en comparacions amb formes de vida actual que conserven solucions evolutives molt antigues o en testimonis indirectes de la vida. A partir d’ara uns nous testimonis s’afegeixen al coneixement que podem tenir dels inicis de la vida: els fòssils. Els registres fòssils més antics coneguts provenen de tres fonts: els microfòssils, la matèria orgànica antiga continguda en les roques i els estromatòlits.
S’anomenen estromatòlits unes estructures finament laminades, en forma de túmul o cúpula, generalment carbonatades, si bé en algun cas també poden compondre’s primordialment de sílice, de dimensions molt diverses, la formació de les quals és el resultat de l’activitat de cianobacteris en aigües poc profundes. Els estromatòlits són el que queda de les comunitats laminades de bacteris convertides en pedra. Actualment es formen estromatòlits a molts llocs i això facilita la identificació de les estructures fòssils que tenen les mateixes característiques. Procedeixen d’un tapís bacterià fibrós, format primordialment per cianobacteris, que atrapa sediments i els acumula. A mesura que els sediments es van acumulant els microorganismes o bé també queden atrapats i moren o bé es desplacen cap a la superfície i formen una nova capa per damunt de l’antiga. Si la capa antiga conté prou sediment i la taxa d’evaporació és prou alta, es pot anar endurint i mineralitzant. La repetició d’aquest procés un gran nombre de vegades condueix a la formació d’estructures de dimensió variable, entre uns quants metres quadrats i unes quantes hectàrees.
Tot i que es tracta d’estructures d’origen orgànic i que n’hi ha una formació contemporània activa, els estromatòlits han estat fins fa molt poc objecte d’interpretacions contradictòries que en dificultaven la comprensió. Ja foren reconeguts al final del segle XIX per alguns paleontòlegs, molt particularment pel nord-americà Charles Walcott, i fins i tot hom els aplica el nom de Cryptozoa pensant que eren restes d’algun grup desconegut d’animals. Però fins que no s’han estudiat els cianobacteris actuals que formen tapissos i estromatòlits en medis tan extrems com les “sebkhas” o altres estanys hiperhalins de diferents països àrids (entre ells la mateixa Austràlia occidental, on s’han trobat els més antics estromatòlits fòssils) no s’ha arribat a interpretar-los correctament com a resultat de la preservació de les comunitats dels tapissos microbians de l’activitat de remots avantpassats d’aquests cianobacteris i altres bacteris.
Els estromatòlits dominen el registre fòssil del període que va entre 3 500 i 550 milions d’anys enrere. Encara són rars a les roques de l’Arqueà, tal vegada a causa de la distribució de terres i mars en aquell període, que no facilitava l’existència de medis apropiats per a la seva formació. S’han descobert microfòssils de bacteris cap al final dels anys 60 a Fig Tree, a Zimbabwe, els quals han estat datats de 3 400 milions d’anys d’antiguitat. També s’han trobat proves de l’existència de microfòssils en roques arqueanes associades als estromatòlits de la formació de Warrawooma. Es creu que com a mínim sis tipus diferents de bacteris formaven part d’aquells tapissos bacterians.
La conformació d’una atmosfera oxidant
Al llarg de centenars de milions d’anys l’excés d’oxigen era absorbit per alguns éssers vius, pels compostos metàl·lics i els gasos reduïts dels oceans i de l’atmosfera, de manera que només localment i a batzegades començava a acumular-se oxigen a l’atmosfera. S’ha formulat la hipòtesi que la progressiva assimilació del CO2 pels primers bacteris fotosintetitzadors havia contribuït al fet que es produís la primera glaciació de la qual es conserva testimoni geològic, fa entre 2 500
i 2 300 milions d’anys; les causes de les glaciacions encara són un misteri, però l’extracció de CO2 podria haver eliminat o reduït l’efecte hivernacle d’aquest gas a l’atmosfera en un temps que la radiació solar no era tan intensa com avui.
Però amb el temps, a partir dels cianobacteris blavissos que produïen oxigen només en determinades condicions (quan hi havia escassesa d’hidrogen o de sulfats), en derivaren bacteris intensament verds que n’emetien de manera continuada. Sorgiren milers d’organismes fotosintetitzadors oxigènics adaptats a les condicions de vida de les superfícies de roca, de les aigües termals, de les escumes. Finalment, fa uns 2 000 milions d’anys l’oxigen havia ja exhaurit els minerals i els gasos reduïts amb els quals havia anat reaccionant i començà a acumular-se ràpidament a l’atmosfera i ocasionà una catàstrofe de magnitud global.
Molts tipus de bacteris desaparegueren, víctimes alhora de l’oxigen i de la llum, encara més mortífers junts que separats. La vida microbiana, llavors com ara, no tenia cap altra defensa enfront d’aquell cataclisme que la via de la duplicació del DNA, la transferència de gens i la mutació. Al preu de morts innombrables, la intensificada sexualitat que manifesten els bacteris quan es veuen exposats a les toxines reeixí a configurar un nou microcosmos totalment reorganitzat. Els nous mutants resistents a l’oxigen es multiplicaren i ràpidament substituïren els sensibles a les superfícies i el medis exposats, mentre que alguns anaerobis sobrevivien mercès a haver-se retirat a nivells anaeròbics dels fangs o dels sòls.
Des dels primers temps d’exposició, ni que fos local, a l’oxigen, s’havien desenvolupat diferents mecanismes de protecció. La informació que contenien i transmetien els gens que les determinaven, tan preciosa per a la vida en les noves condicions cap a les quals s’encaminava la Terra, es traduïa en nous fenòmens, com ara la bioluminiscència o la síntesi de la vitamina E.
L’aparició de la respiració aeròbica
Pero l’adaptació no es deturà aquí, sinó que, en una de les troballes evolutives més decisives de tots els temps, algun cianobacteri reeixí a desenvolupar un sistema metabòlic en el qual el tòxic mortal que era l’oxigen esdevenia peça indispensable del sosteniment de la vida: la respiració aeròbica. La respiració aeròbica és una via arriscada però molt eficient de canalitzar i explotar la reactivitat de l’oxigen. Essencialment és una combustió controlada que degrada molècules orgàniques i de la qual resulten diòxid de carboni, aigua i una quantitat important d’energia. Mentre que la fermentació produeix dues molècules d’ATP per cada molècula de sucre que degrada, la respiració amb oxigen en pot produir ni més ni menys que trenta-sis.
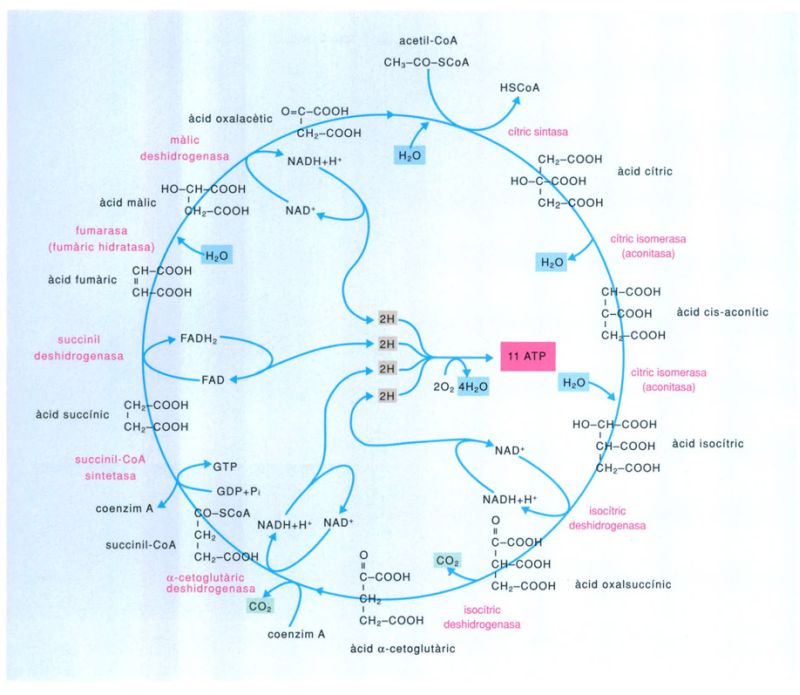
El cicle de Krebs aparegué en la història evolutiva de la vida com un pas endavant cap a l’adequació a la vida en una atmosfera rica d’oxigen. Efectivament, l’adaptació genial a la presència d’oxigen atmosfèric fou la que va inventar un sistema metabòlic que no solament suportava l’oxigen sinó que el feia imprescindible. La respiració aeròbica és una manera enginyosament eficient de canalitzar i explotar la reactivitat de l’oxigen. Es tracta essencialment d’una combustió controlada que trenca molècules orgàniques i dóna diòxid de carboni, aigua i una gran quantitat d’energia. Mentre que la fermentació dóna dues molècules d’ATP per cada molècula de sucre, la respiració pot produir-ne fins a 36.
Biopunt, a partir de fonts diverses
En aquest cas l’ATP es genera en tres etapes. La primera és idèntica a la fermentació: una molècula de sucre (per exemple de glucosa) es transforma en àcid pirúvic. La segona etapa consisteix en un cicle de reaccions, l’anomenat cicle de Krebs,que aprofita els productes resultants de la primera etapa amb producció d’energia. Part de l’energia alliberada s’aprofita per a produir més ATP, però majoritàriament es transfereix en forma d’electrons d’alta energia a molècules transportadores, com els dinucleòtids flavina-adenina (FAD) o adenina-nicotinamida (NAD), els quals passen així a les seves formes reduïdes (FADH i NADH respectivament). Aquestes molècules, que contenen, doncs, la major part de l’energia procedent dels aliments de partida, transfereixen els seus electrons d’alta energia a la tercera etapa de la respiració, una cadena de transport d’electrons en la qual participen diverses menes d’acceptors d’electrons (les ferredoxines i els citocroms entre altres), mentre que l’oxigen molecular fa el paper d’acceptor final que, en rebre els electrons juntament amb els corresponents àtoms d’hidrogen, es redueix a aigua. És en aquesta etapa que es produeix la major part de l’ATP que resulta de la respiració aeròbica.
Molt probablement els primers cianobacteris que usaren l’oxigen en la respiració ho feren amb el mateix oxigen que produïen i del qual havien de fer alguna cosa. Encara avui molts cianobacteris respiren només a les fosques, quan la fotosíntesi no és funcional, sembla que a causa del fet que utilitzen la mateixa cadena de transport d’electrons per als seus processos respiratoris i per a la fotosíntesi, i això no ho poden fer simultàniament.
La diversificació de la vida aeròbica
Els cianobacteris fotosintetitzadors i aeròbics havien trobat, literalment, el seu lloc al sòl. Només amb la llum solar, unes poques sals sempre presents a les aigües i el diòxid de carboni de l’atmosfera, podien produir tot el que necessitaven: àcids nucleics, proteïnes, vitamines i la maquinària bioquímica per fabricar-ho tot. No és gens sorprenent, ateses les grans quantitats d’energia a les quals podien accedir, que els cianobacteris es diversifiquessin en centenars de formes diferents, des de les més petites, d’uns pocs micròmetres de diàmetre, fins a les més grans, de 80 micròmetres (0,08 mm). Algunes prengueren la forma de petites esferes, altres s’envoltaren d’una matriu gelatinosa o s’agruparen en fines làmines multicel·lulars, d’altres encara formaren feixos de filaments embolcallats de gelatina, o simples filaments sense protecció, fins i tot llisos o ramificats i susceptibles d’alliberar pels seus extrems productes de fissions múltiples anomenats beòcits. Algunes, tant colonials com filamentoses, conservaren la fixació del nitrogen com a estratègia metabòlica i desenvoluparen grans cèl·lules (els heterocists) impermeables a l’oxigen i especialitzades en una fixació anaeròbica del nitrogen.
Així reeixiren a colonitzar una gran varietat d’ambients, fins i tot els més extrems, des de les aigües marines més fredes fins a les fonts termals passant per les aigües dolces d’estanys i rierols, els sols humits o fins i tot eixuts, les superfícies i les fissures de les roques. Es desenvoluparen noves relacions tròfiques, com per exemple les saprobiòtiques dels bacteris que s’adaptaren a viure del carboni, el nitrogen i altres metabòlits dels cadàvers de cianobacteris. Però el més important fou que la ininterrompuda pol·lució de l’aire provocada pels cianobacteris fotosintetitzadors forçà la selecció d’altres organismes també capaços d’utilitzar-lo en el seu propi metabolisme. Primer aparegueren bacteris tolerants, que podien viure en presència d’oxigen; més tard els qui el podien usar facultativament i finalment els pròpiament aeròbics.
L’adquisició del metabolisme aeròbic per part de diferents grups de bacteris independentment ocasionà una onada d’especiació i d’especialització, a més de l’aparició de formes i de cicles vitals molt elaborats. Alguns microbis començaren a produir per gemmació cèl·lules filles mòbils d’un aspecte molt diferent al de les cèl·lules mares; aquests bacteris flagel·lats podien desplaçar-se activament cap a llocs més favorables, fixar-se en una superfície sòlida i revertir a la forma materna per a reprendre el desenvolupament. Altres bacteris formaren xarxes de filaments semblants a fongs que es podien cargolar i penetrar entre les partícules del sòl. Altres encara, s’organitzaren en estructures ramificades que alliberaven espores. Tots aquests organismes tan elaborats eren, de fet, multicel·lulars, encara que tots ells eren encara procariotes.
Les condicions atmosfèriques modernes
De fet, entre 2 000 milions d’anys enrere i 1 800 milions, tant la concentració d’oxigen de l’atmosfera terrestre com la densitat de la pantalla d’ozó ja devien haver assolit uns valors de prop de la meitat dels actuals, i ja fa més de 1 000 milions d’anys que s’assoliren segurament els nivells actuals. En aquests punts hi ha opinions divergents, però en general sí que hi ha acord en el fet que la concentració d’oxigen a l’atmosfera mai no ha ultrapassat gaire la que hi ha actualment, entorn del 21%. Si en algun moment hagués estat gaire més gran, el registre fòssil conservaria testimonis d’un incendi generalitzat a tota la Terra. La concentració present, elevada però no gaire, sembla reflectir un compromís entre risc i profit, entre l’amenaça de combustió espontània generalitzada i els avantatges del metabolisme aeròbic.
La formació de la pantalla d’ozó posà fi, per altra banda, a la síntesi abiòtica de compostos orgànics, en impedir als raigs ultraviolats d’arribar a la superfície de la Terra. Cap a la meitat de l’eó Proterozoic, fa uns 1 600 milions d’anys, la major part de l’evolució bioquímica ja estava feta: l’ambient general de la superfície i l’atmosfera de la Terra tal com han arribat fins avui havia quedat fixat en els seus aspectes fonamentals. Quedaven pendents els enormes canvis evolutius que havien de conduir a l’aparició de plantes, animals i fongs, però els cianobacteris generadors d’oxigen ja havien preparat el terreny per a fer-ho possible. Els animals, per exemple, no haurien pogut aparèixer sense que abans ho fessin els vegetals fotosintetitzadors, productors primaris del seu aliment, ni aquests haguessin pogut evolucionar en absència d’oxigen.
Dels bacteris als protoctists
L’aparició dels eucariotes: els acritarcs
L’increment de l’energia amb possibilitats de ser aprofitada pels organismes mitjançant el metabolisme aeròbic conduí a l’aparició d’un nou tipus de cèl·lula, que hom anomena eucariota per un dels seus trets fonamentals, la presència d’un nucli separat de la resta de la cèl·lula per una membrana, el qual conté el DNA que emmagatzema la informació genètica de la cèl·lula.
La transició biològica entre la cèl·lula bacteriana i la cèl·lula eucariota sembla tan sobtada que difícilment pot ser explicada per canvis graduals en el temps. La cèl·lula eucariota és generalment força més gran que la procariota i, a més del nucli, conté orgànuls complexos i sistemes de membranes molt elaborats com mitocondris, plastidis (només en els organismes fotosintetitzadors), undulipodis i aparell de Golgi. Mitocondris, plastidis i undulipodis, per altra banda, posseeixen DNA (és a dir, gens) propis i fins i tot poden reproduir-se per divisió, com si fossin cél·lules independents.
No es pot saber amb certesa quin aspecte tenien les primeres cèl·lules eucariotes ni quan van aparèixer. El registre fòssil és molt pobre entre els 2 000 i els 1 500 milions d’anys enrere, si bé últimament ha millorat amb el descobriment de Grypania i altres eucariotes arcaics. Molt probablement va ser durant aquest interval, un cop constituïda la pantalla d’ozó, que esdevingué possible l’evolució cap a organismes més complexos que els procariotes i de vida més llarga, les cèllules eucariotes.
Probablement els primers eucariotes foren protists unicel·lulars anaeròbics, organismes aquàtics amb patrons de divisió cel·lular i cicle vital variables d’una espècie a una altra. Malauradament, és gairebé impossible seguir l’origen i les primeres passes de l’evolució dels protists en el registre fòssil pel fet que, semblantment a la majoria dels actuals, no devien formar parts dures, cosa que fa força improbable la fossilització. La majoria de les hipòtesis i de les proves relatives a les característiques i les formes de vida dels primers protists es fonamenten en l’estudi dels protists actuals. De fet, molts dels trets dels mastigotes anaeròbics moderns com els retortamonadals o els diplomonadals (per exemple, Giardia) consoliden la hipòtesi que els primers eucariotes eren protists no excessivament diferents d’alguns dels actuals; les peculiars pautes de reproducció de molts, per exemple, semblen indicar que tant la mitosi com la sexualitat biparental evolucionaren en els protists.
Potser els eucariotes més antics que existeixen són els acritarcs. Els acritarcs són microfòssils esfèrics (o, més excepcionalment, polièdrics o en forma d’estel) dotats de parets orgàniques gruixudes i complexes. Per la seva grandària i la complexitat se n’ha de descartar un origen procariòtic; probablement es tracta de formes de resistència (cists) que corresponen a alguna etapa del cicle vital d’algun protoctist, potser alguna alga, cosa que semblaria confirmar el fet que algunes microfotografies de làmines fines de roques amb acritarcs mostren, dins seu, estructures diferenciades que podrien correspondre a nuclis. Els acritarcs són força abundants en sediments dipositats en plataformes oceàniques poc profundes (esquists i carbonats) de 570 milions d’anys enrere ençà, però també se’n troben en roques més antigues; les troballes de paleontòlegs bàltics, escandinaus i nord-americans d’uns fòssils molt més antics amb aspecte d’acritarcs han fet recular fins al Proterozoic l’origen dels organismes que els produïren. Hom ha descrit acritarcs procedents dels Urals meridionals d’entre 1 650 i 1 400 milions d’anys enrere que recorden algues unicel·lulars de l’ordre de les prasinofícies o de la familia de les clorococcàcies, mentre que també s’han trobat acritarcs de parets gruixudes, juntament amb cianobacteris filamentosos i fines vesícules esferoïdals, en roques procedents de Montana d’uns 1 400 milions d’anys d’antiguitat. Siguin o no aquests antiquíssims microfòssils els més antics testimonis de vida eucariòtica, el cert és que en roques del final del Rifeà, de més de 700 milions d’anys enrere, d’Escandinàvia i d’Arizona, ja apareix un registre fòssil de plantes i d’animals planctònics i que a la formació Dal, al Canadà, en estrats d’uns 1 000 milions d’anys enrere s’han trobat làmines orgàniques macroscòpiques, que podrien pertànyer a una alga semblant a les del gènere Ulva actualment vivents.
Els cinc regnes
Biopunt, a partir de fonts diverses
La classificació dels organismes ha sofert un canvi radical gràcies als coneixements de la microstructura cellular i de les diverses hipòtesis emeses sobre l’origen de la vida i de la cèl·lula eucariota. La divisió en dos regnes, animal i vegetal, basada en la capacitat o incapacitat de moviment i l’obtenció d’aliment a partir d’altres organismes o de minerals i llum solar, atribuïdes al regne animal i vegetal respectivament, ha quedat obsoleta. I ha estat així principalment perquè hom pensa que les grans diferències entre organismes no estan en el moviment i la nutrició sinó en el fet d’ésser procariota o eucariota, és a dir, de disposar o no d’estructures cel·lulars amb nucli. El sistema de classificació dels organismes en cinc regnes, que fou proposat el 1959 per R.H. Whittaker, ha estat el més acceptat. En aquest esquema, el regne dels moners conté tots els procarionts, és a dir, bacteris i organismes relacionats, microorganismes. Els altres quatre són eucariòtics: els protoctists (o protists), que comprenen tots els unicel·lulars, siguin autòtrofs o heteròtrofs, i que per tant inclouen nombrosos gèneres abans considerats algues, protozous i fongs; els vegetals, fotosintetitzadors adaptats a la vida terrestre; els animals, pluricel·lulars i amb digestió interna; i els fongs.
L’origen de la cèl·lula eucariota

El paisatge de la Terra, d’entre 1 300 i 800 milions d’anys enrere, no devia ser gaire diferent del que mostra el dibuix. Cap al final d’aquest període (indicat pels òvals) ja hi havia probablement els col·lectius simbiòtics de bacteris antecessors de les plantes i dels animals; és a dir, cèl·lules que posseïen saquets amb clorofil·la (plastidis) que coexistien amb les subunitats capaces d’utilitzar oxigen per a la respiració aeròbia (mitocondris). Tots dos tipus de petits cossos intracel·lulars podien escindir-se en dos, malgrat formar part d’una cèl·lula eucariota. A aquests eucariotes de fa 800 milions d’anys que tenien dues formes d’aconseguir ATP, la respiració i la fotosíntesi, l’evolució els dugué a poder abandonar l’aigua i colonitzar la terra.
Biopunt
S’han proposat diverses teories sobre l’origen de les cèl·lules eucariotes. Alguns biòlegs pensen que els orgànuls cel·lulars es diferenciaren a partir del nucli. Segons aquesta teoria primer de tot les cèl·lules haurien desenvolupat membranes internes, algunes de les quals haurien envoltat el DNA i el RNA i haurien constituït així el nucli; en una segona etapa, alguns dels gens que conformaven el DNA cel·lular s’haurien escapat del nucli i, en quedar envoltats per altres membranes intracel·lulars, haurien format els altres orgànuls de la cèl·lula eucariòtica. Aquesta hipòtesi permet arribar a explicar de manera prou satisfactòria l’origen dels diferents orgànuls, però no explica com pot ser que el DNA d’aquests sigui tan diferent del nuclear i força més semblant al bacterià.
Més aviat semblaria, però, que la cèl·lula eucariòtica hagués estat el resultat de la unió simbiòtica de diferents cèl·lules bacterianes, accidental al començament, més estable després, al final permanent. El pas del temps i la pressió de selecció hauria afavorit unes relacions cada cop més integrades entre diferents organismes inicialment independents fins a constituir un nou tipus de cèl·lula. Aquesta teoria de la simbiogènesi de la cèl·lula eucariòtica, a més d’explicar d’una manera satisfactòria les diferències entre els DNA del nucli i dels orgànuls pel fet que provindrien de cèl·lules procariotes originàries diferents, es fonamenta en una comprensió més profunda del paper de les associacions biològiques derivada d’una línia de pensament biològic que arrenca d’alguns naturalistes russos i alemanys del final del segle passat.
El paper de les simbiosis cel·lulars
Una interpretació parcial de la teoria darwiniana de l’evolució ha posat l’accent tradicionalment en la importància de la competència en les relacions entre organismes i ha menysvalorat la importància de les relacions de cooperació o de simbiosi. De fet la idea de l’evolució com una sagnant “lluita per l’existència” que condueix a la “supervivència dels més aptes”, entenent per més aptes els més forts i més despietats lluitadors, no és pròpiament darwiniana sinó que fou posada en circulació pel filòsof i economista Herbert Spencer (1820-1903), un dels fundadors del mal anomenat darwinisme social. Per a Darwin l’aptesa no es mesurava en termes de força muscular o d’hàbits predadors sinó d’eficàcia reproductiva. En el joc de l’evolució la supervivència no consisteix tant a eludir la mort (que a escala individual és inevitable) com a assegurar la propagació de la vida, la continuïtat de la informació dels propis gens. Però és que a més, en aquest joc, el guanyador mai no és un individu aïllat; cap individu ni cap espècie no poden sobreviure en absència de tots els altres.
La hipòtesi de l’origen endosimbiòtic de la cèl·lula eucariota, que explica alhora l’origen dels diferents grànuls que es troben en l’interior cellular, es fonamenta en la creença que les simbiosis tingueren un paper molt més important del suposat fins ara, i s’aplica als microorganismes d’ambients pobres d’oxigen. La idea parteix d’experiments fets amb bacteris actuals, en els quals es demostra que moltes vegades es dóna aquesta simbiosi entre bacteris diferents que són molt semblants morfològicament però que metabòlicament es complementen.
Biopunt, a partir de fonts diverses
En el cas concret dels microorganismes les simbiosis són freqüents, sobretot en ambients pobres d’oxigen. Quan hi ha absència d’oxigen, la matèria orgànica es degrada molt lentament; la transformació d’un bacteri mort en els productes gasosos finals que en resulten (amoníac, sulfur d’hidrogen i metà) sovint requereix l’acció d’algun altre bacteri i, no rarament de bacteris de més d’una espècie. Així, per exemple, Methanobacillus omelianski, un suposat bacteri del sòl que és productor de metà, havia estat classificat com un microorganisme que produeix el metà a partir d’etanol, i se n’havien fet nombrosos cultius pensant que es tractava d’un únic bacteri; més tard es descobrí que en realitat no es tractava d’un bacteri sinó de dos bacteris, tan semblants morfològicament que encara molt pocs microbiòlegs han estat capaços de distingir-los, però molt diferents metabòlicament. L’un, més primitiu, fermenta l’etanol, mentre que el segon, més evolucionat, aprofita els productes de degradació del primer i els transforma en metà. S’ha pogut comprovar experimentalment que els dos bacteris junts creixen molt més de pressa que cada un d’ells per separat.
Els bacteris precursors dels mitocondris
El possible origen bacterià dels mitocondris fou el primer fenomen d’integració de procariotes en eucariotes que fou reconegut. Els mitocondris són petits cossos que estan envoltats per una membrana, es troben suspesos en el citoplasma, i són de forma ovoide o lleugerament allargada. Fan des d’uns 1,5 µm de diàmetre fins a uns 10 µm de llarg; contenen la totalitat dels enzims respiratoris de la cèl·lula i, per tant, li subministren tota l’energia necessària per als processos metabòlics. És remarcable que, gràcies a l’existència dels mitocondris, totes les cèl·lules eucariòtiques, incloent-hi les dels humans, tenen uns metabolismes prou similars. Deixant de banda la capacitat de fotosíntesi oxigènica de plantes i algues (de fet, anàloga a la dels cianobacteris), tots els trets fonamentals del metabolisme eucariòtic són els mateixos per a tots els animals, les plantes i els fongs: els de la respiració aeròbica. Això contrasta molt amb el que passa amb els bacteris, que tenen, com hem dit, models metabòlics molt diversos, entre els quals figura també el de la respiració aeròbica. El metabolisme de les cèl·lules eucariòtiques parteix de la fermentació, al citoplasma, de les molècules d’aliment fins a l’obtenció d’àcid pirúvic i la transformació d’aquest en acetilcoenzim A, el qual penetra als mitocondris i alimenta allí el cicle de Krebs. Ja hem fet referència als trets bàsics d’aquest cicle fonamental del metabolisme respiratori i a les cadenes de transport d’electrons, amb l’oxigen com a acceptor final, que constitueixen la font bàsica de formació d’ATP, el subministrador d’energia per a tots els processos que esdevenen dins les cèl·lules.
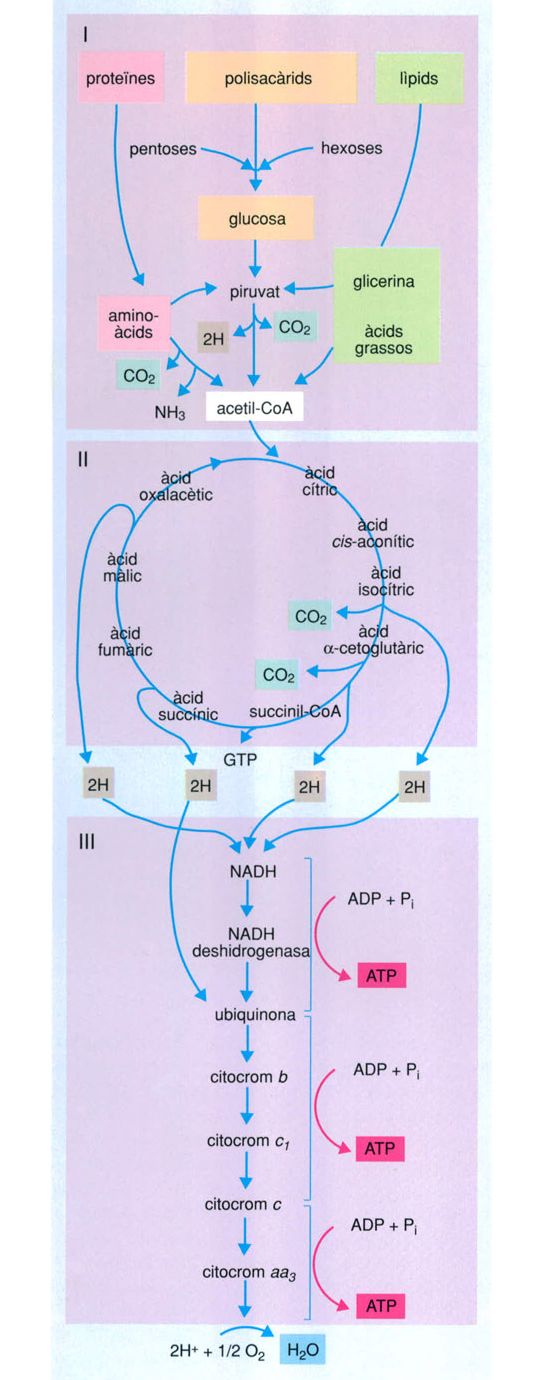
La capacitat de les cèl·lules per a tolerar oxigen fou deguda a l’evolució de cents enzims com catalases, peroxidasesi superòxid-dismutases que reaccionaren amb els radicals tòxics produïts per l’oxigen i els convertiren en compostos orgànics innocus i agua. Els microbis ben aviat trobaren una solució per a l’oxigen, solució que no tan sols els protegia d’un gas potencialment tòxic sinó que els proporcionava a més un altre mètode per a transformar l’energia mitjançant la capacitat de consumir l’oxigen produït en la totosíntesi. Amb aquest nou mètode s’incrementà la producció dAIP cellular, ja que era molt més eficient que la respiració anaeròbica en l’oxidació de molècules.
Biopunt, a partir de fonts diverses
Els mitocondris tenen al seu interior un complex sistema de membranes, en les capes interiors de les quals es localitzen els enzims que catalitzen tots els processos respiratoris. Però les seves funcions no es limiten a les de producció d’energia, sinó que contenen també alguns dels enzims necessaris per a la síntesi de molècules orgàniques complexes com ara els esteroides (lípids amb una estructura bàsica conformada per quatre anells entre els quals figuren nombroses hormones), les provitamines i aitres compostos cel·lulars. Per altra banda, com a senyal manifest del seu origen remot com a bacteris independents, els mitocondris tenen els seus propis sistemes genètic i de síntesi de proteïnes, incloent-hi DNA, RNA missatger, RNA transferidor i ribosomes. Es tracta d’un complex genètic d’estil bacterià, sense cromosomes i amb ribosomes sensibles a antibiòtics com l’estreptomicina, exactament igual que els dels bacteris aeròbics.
Els bacteris precursors dels mitocondris probablement eren semblants a bactcris predadors aeròbics com els actuals Bdellovibrio o Daptobacter. Els Bdellovibrio són petits bacteris aeròbics que es fixen a la paret cel·lular d’aitres bacteris, hi penetren girant com una barrina i s’hi multipliquen utilitzant el material genètic i les proteïnes de la seva víctima; finalment, quan ja n’han tret tot el suc, rebenten els embolcalls buits del que fou un bacteri i els bdelovibris alliberats cerquen un nou hoste i repeteixen el cicle. Daptobacter ha estat observat com a actiu predador de bacteris fotosintetitzadors com ara Chromatium en mostres procedents de l’estanyol d’en Cisó, prop de Porqueres, al Pla de l’Estany (NE de Catalunya) i es multiplica dins dels seus hostes tant en condicions aeròbiques com anaeròbiques. És versemblant que els precursors dels mitocondris fossin bacteris semblants a Daptobacter que explotaven bacteris que eren precursors de la part nucleocitoplasmàtica de la cèl·lula eucariota.
Al començament, fa potser 2 000 milions d’anys, el resultat era la mort en primer lloc dels organismes infectats i després de la majoria dels predadors, si aquests no trobaven a temps un nou hoste, per poc que el medi els fos hostil. Però alguna cèl·lula hoste degué desenvolupar una certa tolerància envers els seus predadors; els permetia viure al llarg de prolongats períodes en l’ambient, ric de nutrients, del seu propi citoplasma, mentre s’aprofitava de la capacitat dels premitocondris de respirar oxigen, no tant pels avantatges que li pogués reportar directament des del punt de vista energètic el metabolisme respiratori d’aquests, com pel fet que això la deslliurava en part del perillós tòxic en uns ambients en els quals cada cop era més present. La formació de la membrana nuclear hauria estat una condició necessària per a l’establiment d’aquesta simbiosi per tal de protegir el DNA de l’enriquiment d’oxigen, ja no sols de l’ambient extern a la cèl·lula, sinó del mateix citoplasma, a causa del funcionalisme dels mitocondris.
Per als precursors dels mitocondris també hagué de resultar avantatjós retardar o fins i tot evitar la mort de la cèl·lula hoste per tal d’assegurar la pròpia descendència, sobretot en ambients externs molt adversos (altes temperatures, acidesa o salinitat extremes, etc). La conflictiva relació entre predador i presa esdevenia a poc a poc un intercanvi d’avantatges que tampoc devia excloure de tant en tant retorns a la virulència del passat amb desavantatge de les dues parts novament en conflicte. Alguns investigadors pensen que algunes formes de càncer no són altra cosa que un retorn atàvic d’alguns mitocondris a la relació conflictiva amb aquella cèl·lula que deixen de reconèixer com a seva, una mena de rebel·lió mitocondrial enfront de la relació simbiòtica establerta.
Els bacteris precursors del citoplasma
Els bacteris hoste que aportaren citoplasma i material genètic primari a la simbiosi que descrivim devien ser probablement d’un tipus semblant al dels actuals Thermoplasma,habitants de medis extremament hostils i alhora susceptibles de tolerar la presència de petites quantitats d’oxigen. Els Thermoplasma són uns bacteris molt peculiars. Tot i que pertanyen al grup de bacteris anomenats afragmabacteris,que es caracteritzen per la incapacitat de sintetitzar els polisacàrids de la paret cel·lular rígida que envolta les cèl·lules de tots els altres bacteris, la diagnosi del seu RNA i els seus lípids mostra que són arqueobacteris (com els halobacteris i els productors de metà). La falta d’una paret cel·lular rígida, no els fa vulnerables malgrat la seva exigua grandària (algunes espècies d’aquest grup no arriben als 0,2 µm de diàmetre), perquè precisament els fa immunes als efectes de la penicil·lina i altres antibiòtics que inhibeixen el creixement de la paret cel·lular. La membrana que els delimita és constituïda per tres capes formades per molècules lipídiques i hidrocarbonades que només es dissolen en solvents orgànics.
Els Thermoplasma són tan resistents que poden créixer en medis extremament calents i àcids, amb temperatures de prop de 60°C (moren de fred a temperatures inferiors als 40°C, cosa que en dificulta enormement l’estudi) i pH del voltant d’1 o 2 (l’acidesa de l’àcid sulfúric concentrat); de fet els únics exemplars estudiats procedeixen de la superfície de piles d’escòries calentes de carbó i minerals sulfurats o de les fonts termals del parc nacional de Yellowstone, als Estats Units. Però el que en fa els perfectes candidats al parentiu amb els precursors de la part nucleocitoplasmàtica de la cèl·lula eucariòtica és el fet de ser l’únic procariota que té el DNA embolcallat per unes proteïnes semblants a les histones que són les que acompleixen la mateixa funció en el DNA cromosòmics de les plantes i els animals.
La majoria d’afragmabacteris, per a reforçar i fer més flexible la seva membrana, necessiten esteroides, tot i que gairebé cap d’ells no en sintetitza i els han d’obtenir del medi, sovint de l’animal o de la planta que parasiten. Precisament la capacitat de sintetitzar esteroides dels bacteris precursors dels mitocondris podria ser una propietat que hagués afavorit l’establiment de la simbiosi que conforma la cèl·lula eucariòtica. Les membranes dels afragmabacteris s’assemblen més a les dels eucariotes que no a les parets cel·lulars de la resta dels procariotes. Per donar flexibilitat i resistència a les seves membranes, els eucariotes empren esteroides, molècules en la síntesi de les quals intervé el metabolisme de mitocondris, encara que les primeres etapes de la síntesi d’esteroides es produeixen en el citoplasma. Podria ser que primerament la síntesi dels esteroides fos només l’aportació dels precursors dels mitocondris a la vida en comú amb el material genètic de la seva cèl·lula hoste per tal d’estalviar-li el contacte amb el perillós oxigen que ells empraven en el seu metabolisme; més tard la disponibilitat d’esteroides resultaria avantatjosa en facilitar la formació de membranes flexibles, fàcils de badar i de cloure i susceptibles de formar vesícules per a envoltar i protegir tant els mateixos mitocondris o el nucli com altres orgànuls, com també per formar altres estructures cel·lulars.
Els bacteris precursors dels plastidis
Potser uns cent milions d’anys després que els mitocondris quedessin fixats com a orgànuls cel·lulars, un nou tipus d’organisme vingué a ferli companyia en determinades cèl·lules: els precursors dels plastidis. L’origen simbiogenètic dels plastidis ja fou reconegut fa prop de cent anys, però fins fa pocs anys aquest reconeixement no ha estat generalitzat.
Els plastidis són, com els mitocondris, petits orgànuls envoltats per una doble membrana, suspesos en el citoplasma, de forma generalment ovoide o discoïdal i de mida variable, compresa entre els 3 i els 10 µm. Els plastidis solen contenir els pigments necessaris per a la fotosíntesi disposats en unes estructures laminars peculiars, si bé en alguns casos els poden perdre i llavors adquireixen funcions d’acumulació transformant-se en una mena de magatzems de substàncies de reserva.
En el cas dels precursors dels plastidis, a diferència del que succeí amb els dels mitocondris, el mecanisme d’incorporació no fou la infecció sinó la ingestió. Determinats protoeucariotes devien ingerir, entre moltes altres fonts d’aliment, alguns cianobacteris o altres bacteris fotosintetitzadors, però algunes nissagues d’aquests degueren desenvolupar alguna capacitat que els féu indigeribles, bo i permetent-los mantenir en activitat els seus valuosos pigments captors d’energia lluminosa dins i tot del citoplasma dels seus ingestors.
De fet hi ha un organisme vivent, Prochloron, que es comporta exactament com ho devien fer els precursors dels plastidis, tot i que no ho fa amb cèl·lules precursores de les eucariotes sinó amb les dels teixits de les parets cloacals de diferents espècies de tunicats del Pacífic tropical. Al final dels anys 60 hom descobrí que unes suposades algues verdes que vivien en simbiosi amb diferents gèneres de tunicats del Pacífic tropical eren en realitat procariotes fotosintetitzadors que només es diferenciaven d’un cloroplast en el fet de mantenir una paret cel·lular típicament procariota. Aquests organismes, que reberen el nom de Prochloron, constitueixen ben bé la baula perduda de la història evolutiva de la simbiosi que conduí a la cèl·lula eucariota de les algues i les plantes, perquè combinen amb èxit una fisiologia de cloroplast vegetal amb una estructura de bacteri. A diferència dels cianobacteris, que també són fotosintetitzadors però només tenen clorofil·la a i pigments verd-blavosos, Prochloron té, com les algues verdes i les plantes, clorofila a i clorofila b però no pigments verd-blavosos. No és gens inversemblant que també fossin avantpassats de Prochloron els bacteris indigestos que foren els precursors dels cioroplastidis.
Semblantment, els ancessors dels rodoplastidis de les rodofícies haurien estat cianobacteris coccoïdals. Hom comparà les seqüències de nucleòtids del RNA ribosòmic dels plastidis i del citoplasma de la rodofícia Porphyridium i observa que eren molt diferents (no tenien més d’un 15% en comú); en canvi, les primeres s’assemblen molt més a les seqüències del RNA ribosòmic dels cloroplastidis d’un protoctist fotosintetitzador, com Fuglena, o les d’un cianobacteri, com Synechococcus (les similituds són respectivament del 33 i del 42%).
Els precursors dels cilis i els flagels
També els undulipodis, és a dir, els cilis i flagels de moltes cèl·lules eucariotes i dels espermatozoides animals i vegetals, han de tenir un origen simbiòtic, encara que de moment el consens entre els biòlegs respecte a aquest punt no és tan gran com en allò que afecta la teoria simbiòtica de l’origen de mitocondris i plastidis.
Per començar, l’estructura dels undulipodis és exactament la mateixa en totes les cèl·lules que en tenen, tant si es tracta d’un ciliat infusori com d’un espermatozoide de mamífer, de les cèl·lules de la tràquea d’un humà o de les zoòspores d’un míldiu: nou parells de microtúbuls d’un diàmetre d’uns 24 nm disposats en cercle i dos microtúbuls més al centre del cercle que formen els altres nou parells i, a l’extrem basal, un petit cos anomenat cinetosoma format per nou triplets de curts microtúbuls. Les parets dels microtúbuls són formades per unes proteïnes específiques anomenades tubulines i alguns centenars d’altres proteïnes menys conegudes. Fins el 1989 només es coneixien àcids ribonucleics i això no era prou concloent respecte a l’origen simbiòtic dels undulipodis, però finalment s’ha trobat DNA als cinetosomes de Chlamydomonas, un protoctist fotosintetitzador dotat d’undulipodis.
D’altra banda, ja des de mitjan anys 60 es coneixia l’existència de microtúbuls anàlegs als dels undulipodis en les espiroquetes. Hem de considerar efectivament les espiroquetes (o bacteris semblants precursors seus) com els candidats més apropiats a haver estat els simbionts que han aportat la motilitat a la cèl·lula eucariota. Les espiroquetes més conegudes, com ara Treponema pallidum, l’agent causant de la sífilis, tenen la imatge de perillosos microbis patògens, però en realitat n’hi ha moltes altres que viuen lliurement en aigües estanyades o en tapissos microbians, i d’aitres que viuen precisament com a simbionts d’altres organismes. De fet, es coneixen espiroquetes que faciliten la motilitat a organismes eucariotes unicel·lulars sobre els quals viuen. Aquest seria el cas de les espiroquetes que viuen sobre Mixotricha paradoxa, un protoctist que viu a l’intestí del termit australià Mastodermes darwiniensis, i són les espiroquetes les que permeten als protoctists traslladar-se gràcies al seu moviment coordinat. Molt probablement l’associació entre un arqueobacteri semblant a Thermoplasma i una espiroqueta fins i tot degué precedir la incorporació a la simbiogènesi de la cèl·lula eucariota dels precursors dels mitocondris. En els fangs anòxics del principi del Proterozoic, aquests protist devien ser “atacats” per precursors dels mitocondris per a formar les bases de tota la vida subsegüent.
L’evolució dels eucariotes primigenis
L’aparició de la reproducció sexual
Els primers organismes eucariotes hagueren de ser protoctists unicel·lulars (en la mesura que puguem considerar unicel·lular un organisme resultant d’una simbiogènesi) que es reproduïen, com els bacteris, per simple divisió. Però un altre tret que acabà de separar de manera radical la naixent vida eucariòtica de la procariòtica fou l’emergència d’una forma de reproducció nova: la reproducció sexual. En efecte, tots els procariotes es reprodueixen asexualment per escissió binària o, més rarament, per gemmació. Tant en un cas com en l’altre, la descendència procedeix d’un sol progenitor i en rep tots els seu gens. Hi ha manifestacions de sexualitat —amb la formació d’un nou ésser dotat de gens procedents de més d’un progenitor (normalment, però no necessàriament, dos)— en alguns procariotes, però en ells el sexe mai no va lligat a la reproducció sinó que més aviat es relaciona amb l’adquisició d’algun gen procedent d’una altra cèl·lula, d’un virus, o del medi. Per contra, tots els animals i les plantes i la majoria dels fongs, es reprodueixen sexualment amb la intervenció de dos progenitors cada un dels quals té una contribució equivalent a la dotació genètica de la descendencia.
Els humans tendim a veure com a indestriables sexe i reproducció. Tanmateix, val la pena recordar que els tres components fonamentals que ara veiem associats a la reproducció sexual (reducció del nombre de cromosomes a la meitat, fusió dels nuclis de dues cèl·lules diferents i successió cíclica d’aquests processos), són realment tres processos distints que només tardanament han quedat vinculats a la reproducció d’organismes complexos.
Mitosi i microtúbuls
La reproducció sexual tal com la coneixem comporta dos processos diferents: mitosi i meiosi. L’un i l’altre evolucionaren en protists.
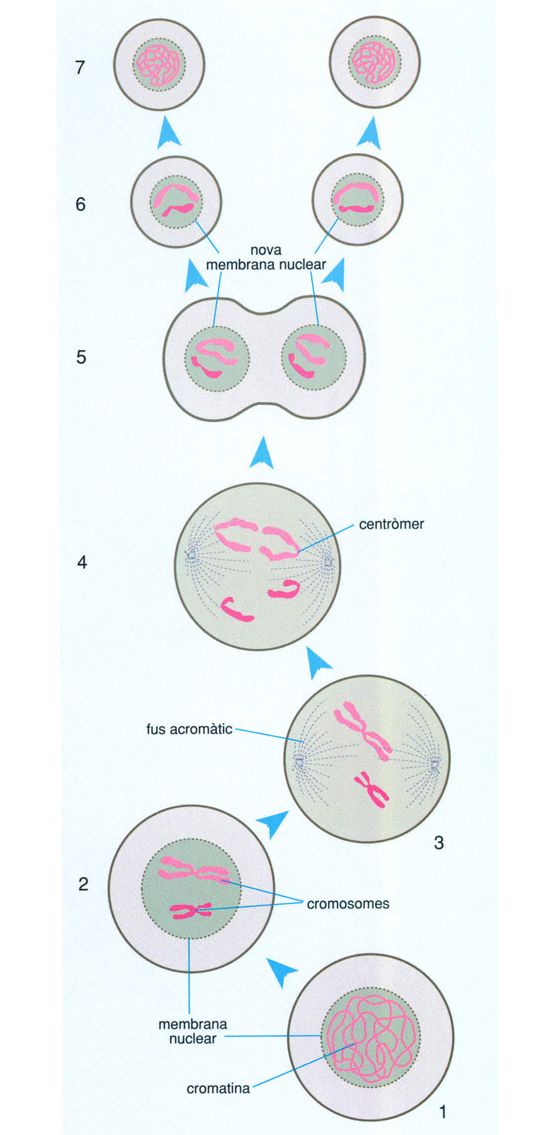
La mitosi és el procés pel qual la cèl·lula es divideix i dóna lloc a dues cèllules genèticament idèntiques a l’original, amb un nombre diploide de cromosomes. Això fa que en el cas dels organismes unicel·lulars la mitosi sigui el mecanisme de reproducció asexual. Com en la meiosi, abans de començar la divisió cel·lular, la cèl·lula ha hagut de duplicar el seu material gènetic de manera que cada cèl·lula filla rebrà una còpia idèntica de la dotació cromosòmica.
Biopunt, a partir de fonts diverses
El primer, la mitosi, es presenta també en els eucariotes unicel·lulars que es reprodueixen asexualment per divisió. Els nuclis de la majoria de les cèl·lules de plantes i animals contenen dues sèries completes, homòlogues però no idèntiques, de cromosomes. Aquestes cèl·lules que tenen dues dotacions cromosòmiques s’anomenen diploides; les que només en tenen una (generalment només les destinades a la reproducció) s’anomenen haploides. En la mitosi, cada cèl·lula filla rep una còpia de la totalitat del material genètic de la cèl·lula progenitora. Aquest material genètic es troba condensat en cromosomes, que són uns petits cossos formats fonamentalment per llargues molècules de DNA cargolades sobre elles mateixes, embolcallades en molècules de proteïnes de diferents tipus, principalment histones, i que contenen també RNA. Els cromosomes de la cèl·lula progenitora es dupliquen i les seves dues meitats idèntiques es divideixen i s’allunyen cap a pols oposats de la cèl·lula, a cada un dels quals el DNA dels cromosomes es desenrotlla, apareix una membrana nuclear que l’envolta i finalment es consuma la separació de les dues cèl·lules filles. La mitosi requereix una estreta coordinació entre dos sistemes cel·lulars: l’un dobla la quantitat de DNA cromosòmic per síntesi i l’altre forma el fus mitòtic que transporta els cromosomes als extrems oposats de la cèl·lula en divisió.
Indubtablement la mitosi s’originà durant la diversificació dels protoctists. En totes les cèl·lules d’animals, plantes i fongs, la mitosi funciona tota regularitat, però no se’n coneixen precedents entre els procariotes i, llevat d’algunes espiroquetes i d’Azotobacter, ni tan sols tenen microtúbuls, indispensables per a la formació dels fusos mitòtics. No es pot pensar, doncs, que el desenvolupament de la mitosi hagi estat un procés lineal, sinó que ha hagut de desenvolupar-se amb successives provatures i fracassos, variants, encerts i atzucacs: 250 000 especies de protoctists han estat el fruit evolutiu d’aquest procés.
Algunes amebes sense diferenciació de sexes, per exemple, posseeixen complexos de DNA amb proteïnes que mai no formen eromosomes. En alguns dinoflagel·lats, els cromosomes són anòmals, el DNA té grans quantitats d’hidroxiuracil —una base poc corrent— i mai no es desenrotllen sinó que romanen densament empaquetats, fins i tot quan no estan en mitosi. En canvi, no tenen histones. En d’altres dinoflagel·lats, el que passa és que els cromosomes s’adhereixen directament a la membrana nuclear, en una disposició que recorda la de les cèl·lules procariotes, i en algunes euglenes els cromosomes es dirigeixen cap al pol respectiu en moments diferents.
D’altra banda, la presència de microtúbuls en el fus cel·lular, els quals orienten i possibiliten els moviments dels cromosomes, fan pensar en un origen comú d’aquestes estructures i dels undulipodis. Si imaginem que algunes de les espiroquetes participants en la simbiogènesi d’alguna cèl·lula eucariota haguessin passat a l’interior de la cél·lula hoste i fossin l’origen dels microtúbuls del fus mitòtic i dels centríols, s’explicaria que alguns organismes eucariotes unicel·lulars amb sistemes mitòtics ben desenvolupats, com ara amebes i rodofícies, no tinguin undulipodis per assegurar la seva motilitat, mentre altres que sí que en tenen, com ara els amebomastigots, els perdin durant els processos mitòtics.
La reducció meiòtica
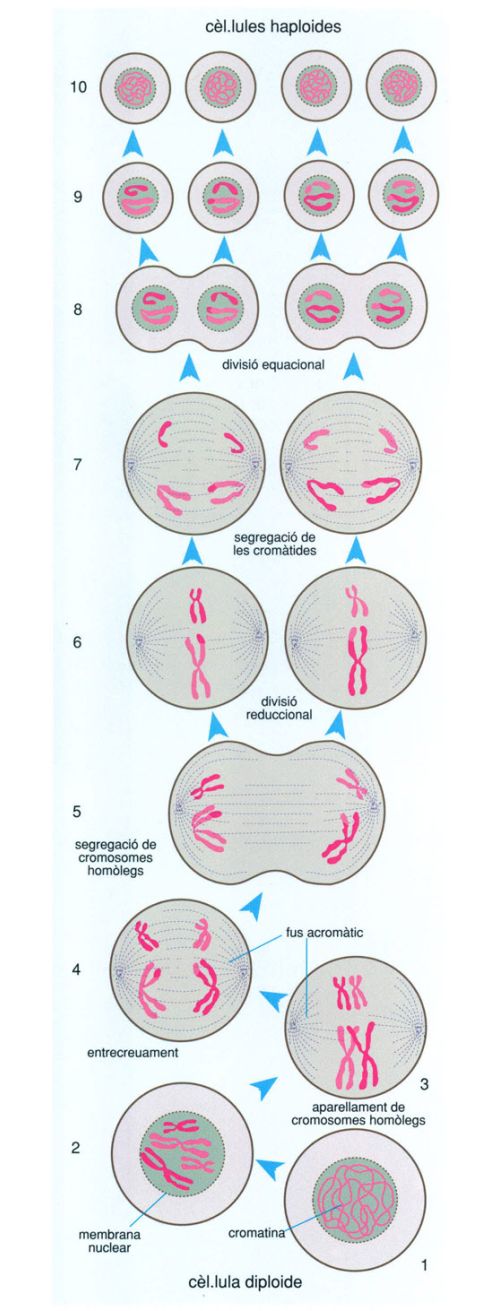
La meiosi és el mecanisme pel qual una cèl·lula diploide es dividex en quatre d’haploides. Igual que en la mitosi, cada cromosoma es condensa després de duplicarse. Les dues parts idèntiques estan unides en un punt constrenyit anomenat cinetòcor: però hi ha encara dos cromosomes similars, homòlegs, de cada tipus, provinents un de cada pare (fase 2). Els cromosomes homòlegs se situen simètricament a banda i banda del pla equatorial de la cèl·lula. Abans que la divisió cel·lular els separi, poden intercanviar seccions de DNA entre si. A continuació, la primera divisió cellular (fases 5 i 6) dóna lloc a dues cèl·lules, cadascuna de les quals duu la meitat del nombre de cromosomes que la cèl·lula original. La segona divisió cel·lular (fases 8 i 9) és semblant a la mitosi, ja que simplement se separen els cromosomes dobles pels cinetòcors. El resultat final són quatre cèl·lules amb una dotació genètica diferent.
Biopunt, a partir de fonts diverses
Però per a la reproducció sexual no n’hi ha prou amb la mitosi. Hi ha d’haver també un procés de reducció a la meitat de la dotació cromosòmica, si més no en les cèl·lules destinades a la reproducció o gametes, procés que s’anomena meiosi. La meiosi distribueix els cromosomes de manera que cada cèl·lula filla rep una meitat de cada un dels parells de cromosomes homòlegs de la seva cèl·lula progenitora. Així, les cèl·lules que resulten per meiosi d’una cél·lula diploide són sempre haploides, mentre que, en la fecundació, la fusió de dues cèl·lules haploides restableix la diploïdia. La meiosi pot incloure o no entrecreuament, és a dir, intercanvi de seccions de DNA entre cromosomes homòlegs i recombinació dels seus gens. En tots els casos, però, meiosi i fecundació condueixen a noves combinacions de gens.
Els detalls de mitosi i meiosi varien considerablement d’unes espècies a unes altres, especialment pel que fa als protoctists. Les seves funcions, per contra, són les mateixes: la mitosi assegura l’exacte repartiment en meitats del material genètic de la cèl·lula progenitora a les cèl·lules filles (qualsevol imprecisió en la segregació de cromosomes o qualsevol pèrdua de material genètic resulta letal), mentre que la meiosi produeix cèl·lules haploides a partir de céiiuies diploides. La fecundació, finalment, ha d’intervenir en el restabliment de la diploïdia entre meiosi i meiosi en qualsevol cicle vital estable. Els organismes mitòtics es poden reconèixer tant si hi ha observacions del comportament cromosòmic com si no, pel fet que l’herència genètica es transmet en ells d’acord amb les lleis de Mendel.
L’enigma de la reproducció sexual
Però hi ha un enigma en tot això. La reproducció sexual pot semblar una complicació supèrflua i innecessària. No té cap de les virtuts de la lliure transferència de material genètic de la sexualitat bacteriana i el seu “cost” (haver de produir una mena particular de cèl·lules sexuals amb la meitat de cromosomes que les altres, trobar parella, etc.), en els termes “econòmics” en què molts biòlegs l’han volgut descriure, sembla desproporcio- nat amb relació a qualsevol avantatge que en pugui resultar. Per què han d’unir-se dues meitats per a fer un tot només per obtenir d’aquest tot noves meitats? Probablement la reducció a la meitat del nombre de cromosomes en el nucli d’algunes cèl·lules podria haver estat determinada per una de les més velles amenaces que ha sofert la vida a la Terra: la fam.
L’amenaça de la fam pogué empènyer al canibalisme algun protist en més d’una ocasió. En algun cas podria ser que la presa no hagués estat totalment digerida i el nostre afamat caníbal hagués quedat dotat amb un doble joc de material genètic, és a dir que hagués esdevingut diploide. Un altre possible origen dels nuclis diploides podria haver estat alguna anomalia de la mitosi com les estudiades en diferents protoctists, sobretot d’alguns hipermastígides que viuen a l’intestí d’escarabats i tèrmits xilòfags. En aquells organismes han estat observades anomalies en la divisió dels cromosomes que condueixen al fet que, després de la duplicació, tots anessin cap a un dels dos pols de la cèl·lula entre els quals s’haguessin hagut de repartir. Però també s’ha observat en altres protoctists que la divisió nuclear es produïa, però no la cel·lular, i que els dos nuclis fills formats es fusionaven entre ells. Aquests processos, inicialment casuals, pogueren ser seleccionats per l’evolució en resultar més favorable en circumstàncies adverses tenir cèl·lules més grans (com les que tenen doble dotació de cromosomes idèntics, diploides), però es podia haver retornat a la situació anterior, amb una sola dotació (haploide), en modificar-se novament les condicions ambientals i esdevenir aquest cop més favorables a les cèl·lules petites.
L’èxit de la sexualitat
El comportament irregular de molts protists unicel·lulars en la reproducció és prou corrent; una seqüència que hagués inclòs la formació d’una cèl·lula diploide, l’aparellament dels cromosomes homòlegs d’aquesta cèl·lula i el fracàs en la divisió dels parells de cromosomes de manera que cada doble cromosoma fes cap a un dels pols de la cèl·lula, sembla condició indispensable per a la implantació de la meiosi. Una combinació de canibalisme, fusió nuclear, irregularitats en la divisió dels cromosomes i cicles de condicions del medi afavorint alternativament la diploïdia i l’haploïdia podia conduir a la mort (i segurament hi degué conduir moltes vegades), però en algun cas, per aquestes vies tortuoses, alguns organismes tingueren èxit i establiren la nova forma de reproducció.
Com altres esdeveniments evolutius, els organismes amb reproducció sexual començaren a proliferar amb una embranzida conjunta en la mesura que començaren a interactuar amb el medi i coevolucionaren plegats. Si no hi hagués hagut canvis ambientals, si les condicions climàtiques i els fenòmens geològics haguessin estat sempre regulars i predictibles potser no hagués evolucionat el sexe. De fet, hi ha organismes, com ara les esponges o alguns fongs, que viuen en ambients molt estables que alternen la reproducció sexual i l’asexual, però la majoria dels ambients del món són variables en una o altra mesura i en ells han evolucionat animals i plantes, que són organismes intrínsecament sexuals.
Les lleis de Mendel
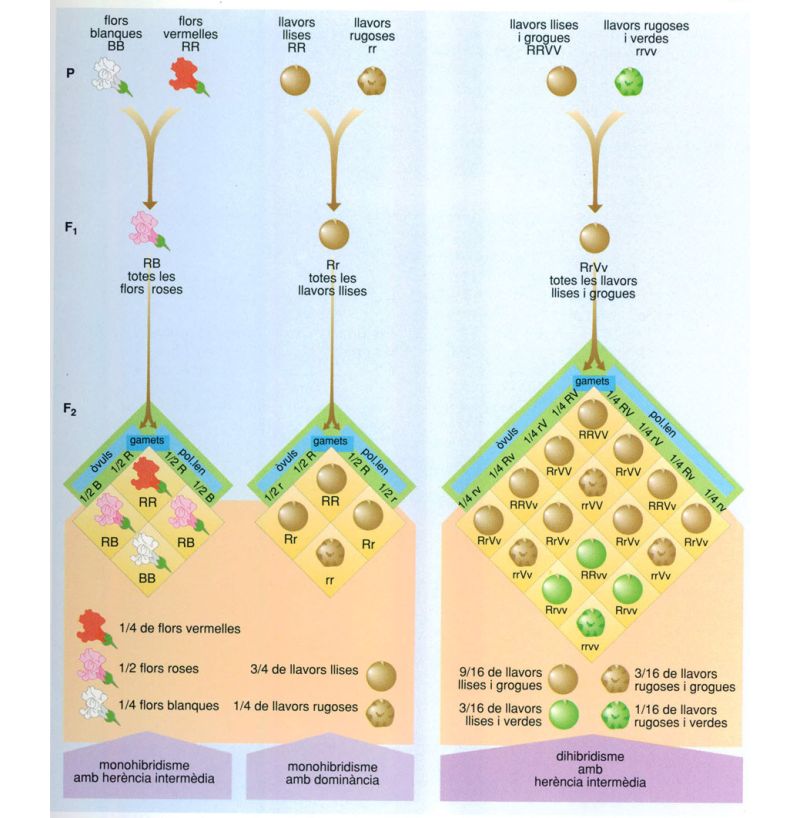
Biopunt, a partir de fonts diverses
Les lleis de Mendel expliquen la segregació i la independència dels caràcters hereditaris. En tots tres casos es pot apreciar la uniformitat de la primera generació filial; en el primer cas (esquerra) el fenotip dels individus de la F1 mostra una herència intermèdia; en el segon cas (centre) un caràcter (llavor llisa) domina sobre un altre (llavor rugosa). En el tercer cas (dreta) es representa un cas d’herència de dos caràcters: llavors llises i grogues davant de Ilavors rugoses i verdes.
L’especialització cel·lular
En qualsevol cas al Proterozoic tardà, fa uns 700 milions d’anys, tal com demostren els fòssils més antics d’animals pluricel·lulars que coneixem, ja s’havien desenvolupat els mecanismes genètics més importants que asseguraven la variació a través de la reproducció sexual. Probablement ja ho havien fet, si més no parcialment, fa uns 1 200 milions d’anys.
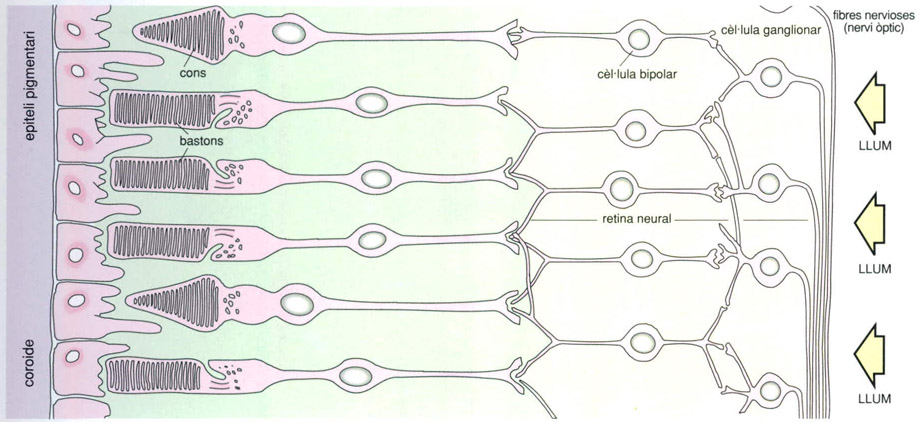
L’especialització cel·lular ha arribat a assolir uns nivells de complexitat i de diversificació extraordinaris en els animals; n’és un bon exemple l’especialització de les cèl·lules sensibles a la llum de la retina. Entre els animals, la percepció visual pot anar des de la simple capacitat de distingir la llum de la foscor, com és el cas de les planàries, fins a la formació d’imatges de gran definició, com és el cas d’alguns mamífers.
Biopunt, a partir de fonts diverses
El desenvolupament d’unes estructures (cromosomes)que permetien un empaquetament eficient de quantitats considerables d’informació genètica, l’aparició d’un procés eficaç de transmissió d’aquesta informació a les cèl·lules de la descendència (mitosi) d’una banda i d’una manera de recombinar amb rapidesa i eficàcia informació genètica de més d’una procedència (reproducció sexual) de l’altra, permeteren en algun moment del Proterozoic tardà l’aparició d’organismes eucariotes pluricel·lulars cada cop més grans, amb una estructura més complexa i diversificada, i més especialitzats. Només calia que una cèl·lula d’un protoctist flagel·lat s’enganxés amb una altra i la propulsés de manera que la segona no necessités usar els seus undulipodis i pogués aplicar a altres usos els seus microtúbuls per a iniciar la línia evolutiva de la qual derivarien els animals. Era el començament de l’especialització cel·lular, en la qual els animals han esdevingut mestres: unes cèl·lules neden, unes altres conserven la seva capacitat per a la meiosi, i altres apliquen el seu aparell espiroquetal a la percepció dels senyals del món exterior.
Hi ha un límit pràctic per a la grandària d’una cèl·lula i, per tant per a la d’un organisme unicel·lular. Les cèl·lules necessiten interactuar amb l’ambient per a absorbir materials i expulsar els productes residuals. Moltes de les reaccions vitals de la cèl·lula es produeixen a les seves membranes externes. Des d’aquest punt de vista, una cèl·lula petita és més eficient que una cèl·lula gran, i un organisme unicel·lular que augmenta molt de grandària s’enfronta amb un decreixement de la relació entre la superfície i el volum. Alguns organismes han compensat aquest inconvenient modificant la forma, per exemple replegant repetidament la membrana, cosa que permet augmentar la superfície relativa fins a un cert límit. L’única solució definitiva que han tingut els organismes unicel·lulars per a incrementar la seva grandària més enllà d’aquest límit ha consistit a esdevenir pluricel·lurars.
L’aparició dels pluricel·lulars
La pluricel·lularitat ha aparegut, d’una manera o d’una altra, moltes vegades en moltes menes d’organismes. Ja alguns procariotes, com els mixobacteris, alguns cianobacteris (nostocals) i els actinobacteris són pluricel·lulars. No es tracta, doncs, d’un caràcter que sigui exclusiu dels fongs, les plantes i els animals, encara que estiguem acostumats a associar-lo amb aquests.
Els protoctists multicel·lulars
La diferenciació cel·lular i els lligams i les comunicacions entre cèl·lules són molt escassos entre els protoctists multicel·lulars. Les volvocals, un grup d’algues verdes colònials, illustren molt bé l’esglaonament de situacions des de la pura aposicio de cèl·lules fins a organismes constituïts per cèl·lules nombroses i interdependents. Les cèl·lules individuals de les voivocals s’assemblen molt a les dels clamidomones als quals ens hem referit abans; cada una té dos undulipodis, una taca “ocular” sensible a la llum i un cloroplast.
Gonium, el gènere més senzill de voivocals colonials, consisteix en un disc format per substàncies gelatinoses que manté unides quatre, vuit, setze o trenta-dues d’aquestes cèl·lules amb els undulipodis orientats tots cap al mateix costat del disc; qualsevol de les cèl·lules pot dividir-se i iniciar la formació d’una nova colònia idèntica a la de la cèl·lula mare. Pandorina, un altre genere. consisteix en un apilotament ovalat de setze o trentadues cèl·lules en una mena de bossa gelatinosa, amb els undulipodis i les taques oculars dirigits cap a l’exterior de l’apilotament i amb una diferenciació entre les taques oculars d’un extrem de la colònia, més grans que la resta; aquest fet defineix un sentit d’avenç; qualsevol de les cèl·lules de la colònia pot originar també, per divisió, una colònia filla. En canvi, les del gènere Volvox són molt més complexes; consisteixen en una esfera buida formada per entre 500 i 600 000 cèl·lules, amb un pol anterior i un pol posterior; aquests pols defineixen un eix entorn del qual la colònia manté un moviment de rotació al mateix temps que avança, impulsada pels undulipodis, en la direcció que assenyala el pol anterior. Una matriu gelatinosa manté unides les cèl·lules, de les quals només algunes, situades prop del pot posterior, es poden reproduir. Ho fan alliberant a l’interior de la colònia, buit, cèl·lules filles que es divideixen activament i que quan ja han constituït una colònia prou gran alliberen un enzim que dissol la gelatina que manté la cohesió de la colònia mare. Algunes espècies d’aquest gènere, però, es poden reproduir sexualment, de vegades amb els dos sexes presents a totes les colònies i de vegades amb separació de papers sexuals d’una colònia a una altra. Aquí se situaria la frontera entre la vida colonial i la veritable pluricel·lularitat: en el moment en què no totes les cèl·lules poden viure independentment i hi ha especialització de papers. En les espècies de Volvox més diferenciades les cèl·lules reproductores ja no tenen undulipodis i per tant són immòbils.
Els primers animals pluricel·lulars
L’animal d’organització més simple que coneixem és Trichoplax adhaerens, només un sac de cèl·lules planes i ciliades, sense formar cap teixit i amb una especialització funcional mínima. De vida marina, és capaç de moure’s en qualsevol direcció, i no té cap diferenciació polar.
Biopunt, a partir de Margulis, 1985
També en el cas dels animals tenim un testimoni del que devien ser els animals pluricel·lulars més primitius en Trichoplax adhaerens, el més simple dels animals coneguts. Descobert inicialment per Franz Eilhard Schulze en un aquari d’aigua marina de la Karl-Franzens Universität Graz, a Àustria, el 1883, però classificat erròniament com una larva de celenterats, la veritable natura de Trichoplax no fou descoberta fins el 1965. De fet, l’única cosa que el diferencia d’una ameba pluricel·lular és el fet que se n’ha observat el desenvolupament embrionari a partir de la fecundació d’un òvul per un espermatozoide fins a formar una blàstula. L’aspecte general de l’animal adult és el d’una mena de blàstula d’uns pocs milers de cèl·lules, amb una diferenciació entre una part dorsal, formada per cèl·lules aplanades amb un petit nombre d’undulipodis, i una part ventral, formada per cèl·lules columnars amb un nombre més gran d’aquests, mitjançant els quals s’arrossega pels fons. Entremig de les dues capes de cèl·lules diferenciades hi ha una capa intermèdia que conté un fluid amb un petit nombre de cèl·lules. Es tracta clarament d’un organisme multicel·lular de nutrició heterotròfica i, per tant, és un animal, però no té cap teixit diferenciat i la seva especialització funcional és mínima. Amb tot, si veritablement, com sembla, Trichoplax és una mostra de la descendència d’algun dels primers animals diferenciats com a tals, és palès que no podem esperar trobar-ne testimonis fòssils, atesa la seva manca de parts dures.
Els testimonis fòssils de vida animal més antics coneguts fins ara foren trobats el 1947 als turons d’Ediacara, a les Flinders Ranges, uns 450 km al nord d’Adelaide, a l’Austràlia meridional, en sediments proterozoics de prop de 700 milions d’anys d’antiguitat, i ja inclouen animals força més diferenciats. Més recentment s’han trobat fòssils animals d’edat (entre 600 i 570 milions d’anys) i característiques similars al Canadà, a Rússia, a la Gran Bretanya i a Escandinàvia. Potser dels animals de les faunes ediacarianes (entre els quals figurarien esponges, cnidaris, anèl·lids i artròpodes) haurien derivat tots els fílums animals posteriors. Tanmateix, hi ha autors que interpreten la fauna ediacariana com un experiment evolutiu fallit que conduí a uns tipus animals extingits completament al final de l’Ediacarià, fa uns 570 milions d’anys. Les faunes ediacarianes, que efectivament semblen extingir-se de manera gairebé total cap al final d’aquest període, que és també el final del Proterozoic i l’inici del Fanerozoic, assenyalen probablement un límit fonamental de la història de la vida i de la Terra i, de fet, fins el 1930, assenyalaven el límit entre els primers estrats fossilífers del Cambrià i els nivells precambrians en els quals fins llavors no es coneixia cap testimoni fòssil.
De fet, l’avantatge evolutiu més important de la pluricel·lularitat rau en la possibilitat de separar funcions i de diferenciar grups de cèl·lules, organitzades en teixits i òrgans, cada un dels quals n’acompleix alguna. Això, però, exigeix també diverses connexions entre les cèl·lules d’un organisme pluricel·lular que en permetin coordinar les funcions en activitats com el creixement embrionari, la diferenciació cel·lular, el creixement i el metabolisme. Aquestes connexions són molt simples en els fongs, es compliquen més en les plantes i assoleixen una màxima complicació, molt superior a la de vegetals i fongs, en els animals.
Una altra innovació que també hagueren de desenvolupar els organismes multicel·lulars fou la d’adquirir suports físics que asseguressin el manteniment de la pròpia estructura. De fet, ja la majoria de les cèl·lules procariotes disposaven, com disposen avui llevat d’excepcions (la de Thermoplasma ola ja esmentada de Mycoplasma) d’una paret cel·lular protectora, però en els organismes pluricel·lulars la capacitat de secretar material no viu destinat a la protecció i suport tant de les cèl·lules individuals com de l’organisme en el seu conjunt, ha assolit una varietat i una sofisticació extraordinàries, en un marc de regularitats generals dins de cada grup. Així, per exemple, en les cèl·lules animals la coberta sembla consistir normalment en un complex de proteïnes i de polisacàrids que manté unides les cèl·lules immediates; les plantes tenen parets cel·lulars rígides fetes de cel·lulosa, i els fongs en tenen de quitina. Això per no parlar dels exosquelets dels invertebrats, també de quitina; dels ossos dels vertebrats, que combinen una proteïna (el col·lagen) amb sals minerals com els carbonats i fosfats de calci i de magnesi; de la fusta de les plantes llenyoses, el component principal de la qual és la lignina, un polímer orgànic derivat del fenilpropà, o de la protecció externa de la majoria de les plantes terrestres, la cutina, destinada a minimitzar les pèrdues d’aigua.
L’explosió cambriana
L’inici del Fanerozoic és assenyalat per l’aparició dels primers animals pluricel·lulars amb estructures de suport mineralitzades, exosquelets de carbonats o fosfats de calci, que s’han conservat millor que els seus antecessors de cossos tous del Proterozoic. És el que es coneix per “explosió cambriana”: l’aparició, aparentment sobtada, d’un gran nombre de tipus diferents d’animals (pràcticament la totalitat dels fílums avui existents i alguns d’extingits) que han deixat quantitats apreciables de testimonis fòssils. Tots eren aquàtics i entre ells ja hi havia representants dels dos fílums animals que després tindrien més èxit evolutiu: els artròpodes i els cordats.
No cal dir que quan ens referim a una aparició “sobtada” estem parlant en termes d’història geològica. En realitat podem diferenciar dues etapes en l’explosió cambriana. En una primera etapa, el Tommotià, descrita inicialment en una localitat de Iacútia, a la Sibèria Oriental, però present a gran part del món, hauria aparegut la primera fauna amb parts dures (l’anomenada “petita fauna de petxines”) formada per organismes de petites dimensions (uns quants centímetres com a màxim), llevat d’algunes esponges i dels anomenats arqueociàtids, un tipus d’organismes semblant a les esponges, però que s’extingí completament amb tota la resta de la fauna tommotiana fa uns 560 milions d’anys. Entre els organismes tommotians ja es poden trobar precursors indubtables d’alguns fílums actuals (esponges, mol·luscs, etc), però també hi ha fòssils d’afinitats incertes corresponents probablement a fílums extingits.
En una segona etapa, l’Atdabanià, també descrit inicialment a la Sibèria oriental però representat igualment a moltes altres parts del món com ara l’Amèrica del Nord, Grenlàndia, Austràlia i la Xina, apareixeria la veritable explosió de la fauna cambriana, que disposa d’un gran nombre de fòssils d’animals, tant tous com amb parts dures, amb innombrables formes noves que inclouen la totalitat dels fílums avui existents i uns quants d’extingits. A la famosa pedrera de Burgess Shale, prop de Field, a la Colúmbia Britànica, han estat descrits prop d’una vintena d’organismes de morfologia única que podrien correspondre a fílums extingits, al costat de nombrosos representants de tipologies més conegudes corresponents tant a fílums actuals com a fílums també extingits, com ara els trilobits.
El jaciment cambrià canadenc de Burguess Shale

Reproducció de la fauna trobada al jaciment de Burguess Shale. A l’esquerra, els dos models d’arbre evolutiu.
Jordi Corbera (esquerra) i Carles Puche, a partir d’originals de Marianne Collins, extret de Gould, 1989 (dreta)]
La gran diversitat dels models faunístics del Cambrià, la majoria dels quals no han tingut èxit evolutiu, fa pensar que no és correcte l’arbre evolutiu clàssic (a dalt a l’esquerra) basat en la permanència de la majoria dels models troncals apareguts i en l’extinció quasi únicament de les formes col·laterals inadequades.
En efecte, tots els grans models o filums (phyla) trobats per C.D. Walcott el 1909 al jaciment cambrià canadenc de Burguess Shale datat en uns 530 milions d’anys foren interpretats durant els anys vuitanta per l’equip de H. Whittington, un exemplar de la majoria dels quals és representat a la figura (els colors són especulatius). Només n’hi ha tres que subsisteixin actualment: el dels artròpodes i el dels onicòfors, representats per Marrella i Aysheaia, i el dels cordats, representats per Pikaia, els estranys filums a que pertanyien Nectocaris, Odontogriphus, Dinomischus, Opabinia, Wiwaxia, Hallucigenia o Anomalocaris nathorsti han desaparegut de manera tan desconcertant com sobtada és la seva aparició en el registre fòssil.
Això fa pensar, tal com ha suggerit S.J. Gould, en una altra mena d’arbre evolutiu (a baix a l’esquerra) en què molts dels troncs bàsics s’extingeixen, no pas necessàriament perquè eren inadequats, sinò perquè alguna “catàstrofe’’ ambiental en liquida la població. D’acord amb aquesta interpretació, la presència dels humans actuals no ha depès tant d’un llarg procés de complexificació evolutiva creixent del regne animal (interpretació clàssica i més o menys teleològica), com de la “bona sort” de Pikaia, el nostre ancestre més antic conegut.
La conquesta de la terra ferma
La vida encara havia de trigar a colonitzar els continents. Les superfícies de terres emergides resultaven tan hostils per a la majoria dels organismes proterozoics com ho pot ser per a nosaltres la superfície de la Lluna, tot i que sembla provat que alguns cianobacteris colonitzaren ja durant el Proterozoic algunes superfícies emergides humides. El camí cap a la invasió de la terra ferma probablement fou preparat per fenòmens geològics de transgressió i regressió que determinaven l’assecament periòdic de determinades àrees.
El fet de viure en terra ferma tenia tanmateix considerables atractius; d’una banda es podia accedir a molt més oxigen, i a més l’espai lliure era il·limitat. Tot i que els obstacles també eren força seriosos: fora de la protecció de l’impuls ascensional de l’aigua, els organismes es podrien col·lapsar sota el seu propi pes si no disposaven de músculs més potents i esquelets més sòlids capaços de compensar-ho. També és necessari un equipament respiratori adient, ja que calia passar d’un medi en el qual l’oxigen és present en una proporció d’unes poques parts per milió a un altre en el qual la concentració és milers de vegades superior. Així mateix, són indispensables proteccions de la superfície externa, com pells, cutícules i closques, per a evitar la intensa llum solar sense filtrar pel gruix de l’aigua que bat sobre la terra ferma. Però l’amenaça més greu era la dessecació. Només els organismes capaços de retenir i conservar l’aigua podien sobreviure en terra ferma.
Les primeres plantes terrestres
Les plantes terrestres potser són el resultat de simbiosis entre algues i fongs, igual que els líquens, però, al contrari que aquests, amb un predomini del component algal. Probablement no és pas una coincidència que el 95% de les plantes terrestres actuals tinguin a les arrels fongs simbiòtics, les anomenades micorizes. El component algal de les primeres plantes que començaren a colonitzar les riberes de la terra ferma els permetia explotar l’oxigen atmosfèric i retenir l’aigua dins d’estructures inicialment gelatinoses, com la que hem descrit en parlar dels vòlvox, més tard envoltades per una paret de lignina. El component fúngic els permetia assolir l’alteració més ràpida de la superfície de les roques que colonitzaven i extreure’n directament elements minerals, com també ajudar a retenir l’aigua i resistir amb més èxit les fases de sequera.
El resultat fou probablement plantes semblants a les hepàtiques i molses actuals, els espermatozoides de les quals encara necessiten aigua per arribar fins al gàmeta femení, l’ovocèl·lula. En tot cas les plantes terrestres més antigues que han deixat testimoni fòssil són les Cooksonia del Silurià superior del País de Gal·les i d’Austràlia (uns 420 milions d’anys enrere), de les quals no es pot afirmar amb seguretat si ja eren plantes vasculars o encara no; tenien pocs centímetres d’alt i consistien en una tija llisa, sense fulles, ramificada dicotòmicament, i amb esporangis esfèrics a l’extrem de les darreres ramificacions; s’hi observen feixos capil·lars, que podrien ser un xilema incipient, però que també podrien ser simplement vasos conductors semblants als de les setes dels esporangis de les molses actuals; en algun exemplar sembla que s’hi han observat estomes, cosa que indicaria la presència d’una cutícula protectora. Molt probablement, però, no es tracta pas de la primera planta terrestre, perquè ja en sediments de l’Ordovicià superior (uns 450 milions d’anys enrere) s’han trobat espores amb trilet, és a dir, amb una obertura triradiada, que es considera característica de les plantes vasculars o, si més no, de plantes d’hàbitat terrestre. Tot i així, la veritable conquesta del medi terrestre per les plantes s’esdevingué durant el Devonià i, sobretot, el Carbonífer, i fou protagonitzada per noves plantes vasculars, algunes de les quals assoliren ja l’estructura i les dimensions d’arbres.
Els primers animals terrestres
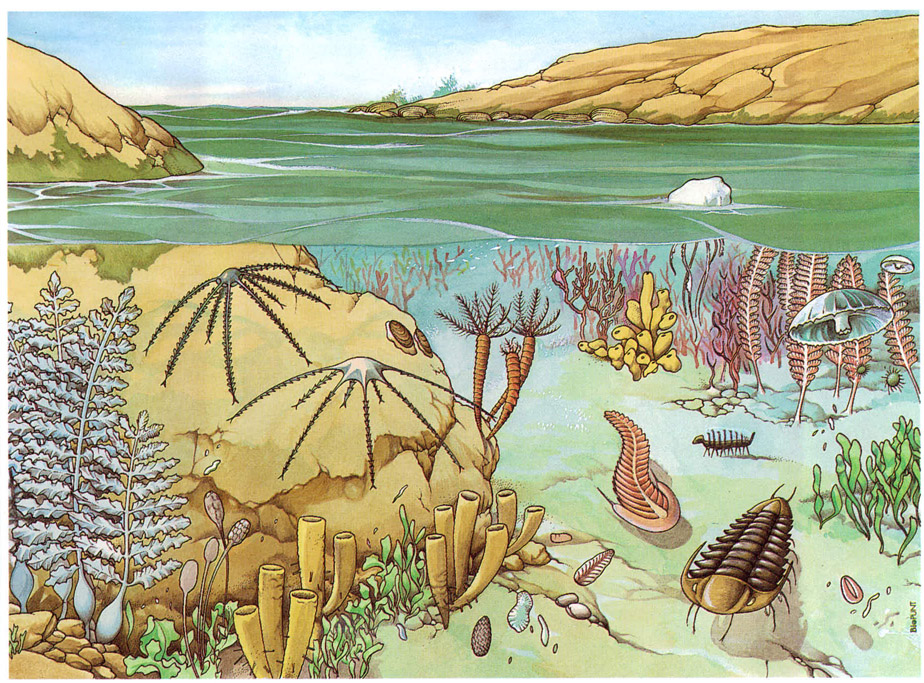
Fa 500 milions d’anys els animals desenvoluparen parts dures als seus organismes a partir de dipòsits de rebuig cel·lular. La presència d’espores fòssils en els sediments de l’Ordovicià mitjà i del Silurià, fa més de 500 milions d’anys, suggereix que la flora d’aquells temps comprenia també organismes amb l’aspecte de briòfits, semblants a les nostres molses i hepàtiques. Fa 400 milions d’anys, al final del Silurià, quan ja existien els primers peixos mandibulats i els primers insectes mancats d’ales, les plantes vasculars ja progressaven. El repte de l’escassetat d’aigua en la terra ferma va ser afrontat amb el desenvolupament de llavors, que permet a l’embrió d’esperar a créixer fins al bon moment. La fertilització i el desenvolupament a l’interior dels teixits humits parentals capacità les llavors per a sobreviure malgrat la inconstància de les pluges.
Biopunt
Els animals trigaren més a emergir a la terra ferma, entre altres coses perquè, abans de la colonització d’aiguamolls, riberes i planes inundables per les plantes, els animals que haguessin pogut sobreviure a les dificultats i inconvenients de la vida aèria no haurien trobat res per menjar. De fet molt pocs representants del regne animal han reeixit a adaptar-se plenament a la vida terrestre; ni un sol fílum animal és constituït exclusivament per organismes terrestres, i tots els animals que avui completen el seu cicle vital a terra ferma pertanyen a tres o quatre fílums predominantment aquàtics: hi ha uns quants artròpodes (principalment insectes i aràcnids), poquíssims molluscs (els cargols de terra i els llimacs), alguns anèl·lids (com els cucs de terra) i uns quants membres del nostre propi fílum (el dels cordats). De la resta de fílums animals, ni una sola espècie ha reeixit a adaptar-se a la vida terrestre en tota la història evolutiva.
I, de fet, totes les espècies animals que han reeixit a adaptar-se a la vida terrestre ho han fet, podríem dir, enduent-se amb ells el seu antic medi: cap animal no ha abandonat del tot l’aigua. La blàstula i l’embrió de rèptils, ocellis i mamífers encara es desenvolupen dins d’una cavitat (el sac embrionari) plena de líquid. La concentració de sals de la sang és pràcticament idèntica a la de l’aigua de mar. Les proporcions de sodi, potasi i clor dels nostres teixits també són sorprenentment semblants a les dels oceans.
Un factor crucial amb relació al pas dels animals al medi terrestre ha estat el metabolisme del calci. La concentració de calci en el citoplasma d’una cèl·lula eucariòtica és de prop d’una part per 10 milions, mentre que a l’aigua del mar és de prop d’una part per 10 000, o encara més alta. El calci tendeix a entrar a les cèl·lules i aquestes se n’han de deslliurar activament de manera continuada. Tanmateix el calci fa un paper central en el metabolisme de les cèl·lules nucleades; és indispensable per al moviment ameboide, la secreció cel·lular, la formació de microtúbuls, la cohesió dels teixits, la transmissió de l’impuls nerviós o la contracció muscular. Un cop usat, però, l’excés de calci ha de ser eliminat o bé acumulat fora del citoplasma cel·lular; és per això que, ja des dels temps cambrians, els organismes havien emmagatzemat les seves reserves de calci com a fosfats, fabricant dents i ossos, o com a carbonats, en forma de closques o petxines. Aparegudes inicialment en animals aquàtics, aquestes estructures han servit més tard per a resoldre la necessitat d’uns mecanismes de suport als animals que han accedit a la vida en terra ferma, per dotar-se de protecció enfront de la llum del Sol o de predadors, i com a “aquaris” individuals protectors dels perills de la dessecació.
Els fòssils d’animals terrestres més antics que es coneixen fins ara (deixant de banda hipotètics rastres de miriàpodes de l’Ordovicià, de més de 450 milions d’anys d’antiguitat) procedeixen del Devonià superior d’Escòcia (Rhynie Chert) i d’Alemanya (Alken-an-der-Mosel), uns 400 milions d’anys enrere. Rhynie Chert és una localitat escocesa que en el Devonià era ocupada per una torbera ocasionalment inundada per aigües carregades de sílice procedent de guèisers o fonts termals propers. La sílice precipitava en forma del quars criptocristal·lí (“chert”) que dóna nom a la localitat, fossilitzant el material vegetal i també un conjunt de petits artròpodes (sobretot àcars i col·lèmbols) que hi vivien i s’alimentaven de materials vegetals i alguns artròpodes predadors (milpeus i aranyes) més grans que es devien alimentar dels primers. Aiken-an-der-Mosei és una localitat renana, prop de Coblença, que correspon a un petit llac devonià al qual devien ser arrossegats alguns organismes clarament terrestres que hi han aparegut, com ara aranyes i un gran milpeus (Eoarthropleura)que, amb els seus 10 cm, és l’animal terrestre més gran conegut del Devonià. També s’hi troben altres animals que havien de ser amfibis i que potser s’alimentaven en terra ferma però vivien preferentment a l’aigua, concretament grans escorpins de fins 1 m de llarg, i euriptèrids (un grup extingit emparentat amb aquells), els uns i els altres amb brànquies dotades de proteccions ventrals que els havien de permetre incursions a) medi aeri a la recerca d’aliment.
També alguns vertebrats reeixiren durant el Devonià a accedir a la vida aèria. Els cordats semblen haver-se originat durant el Cambrià a partir de Pikaia, un dels estranys organismes de cos tou descobert en el famós jaciment de Burgess Shale, a la Colúmbia Britànica, o d’algun organisme semblant, però els primers vertebrats indubtables són els peixos àgnats que comencen a aparèixer a l’Ordovicià, el més ben conservat dels quais és l’Astraspis,descobert als gresos de Harding, a Colorado. Hom considera actualment els conodontats com els primers vertebrats. Es tracta d’uns organismes dels quals pràcticament només es coneixen unes suposades peces bucals (les més antigues es remunten al Cambrià mitjà, uns 520 milions d’anys enrere), i quatre empremtes, encara no estudiades del tot, trobades entre el 1983 i el 1986 prop d’Edimburg, a Escocia, que ja podrien ser àgnats o bé alguna forma intermedia entre cefalocordats i àgnats.
En tot cas, al començament del Devonià, fa uns 400 milions d’anys, els peixos ja s’havien diversificat en tots els grups actuals i alguns ja s’havien extingit; d’un d’aquests grups, el dels sarcopterigis —del qual encara avui sobreviuen quatre generes, el celacant (Latimeria chalumnae), que viu a l’oceà Índic, i els tres gèneres dc peixos pulmonats, que viuen un a cadascun dels continents australs (Lepidosiren paradoxa, a l’Amèrica del Sud, Neoceratodus forsteri,a Austràlia, i Protopterus, l’únic representat actualment per més d’una espècie, a Àfrica)—, sorgí el llinatge dels rifidistis, els primers peixos “amfibis”, a partir del qual evolucionarien els tetràpodes terrestres.
Els rifidistis eren peixos de forma allargada i esvelta, amb una musculatura potent i unes aletes i una cua poderoses, cosa que els havia de fer uns formidables nedadors. Tenien llargs musells, sobretot els més grans, i potser per això mateix desenvoluparen una articulació del crani que els permetia separar les dues mandíbules alhora o, indistintament, la superior o la inferior soles. Aquest fet tingué dos efectes importants, tots dos relacionats amb la vida en aigües poc fondes: en primer lloc el moviment del musell els permetia modificar el volum de la boca i, per exemple, mantenir un volum d’aigua considerable a punt per bombar cap a les brànquies, encara que el cos estigués parcialment emergit; en segon lloc, podien capturar preses en aigües poc fondes sense abaixar la mandíbula inferior, amb un ràpid moviment del musell, tal com fan encara els cocodrils. Això, i les seves poderoses aletes ventrals, els havia de permetre perseguir les preses pels aiguamolls prescindint de les possibles barreres que poguessin oferir els petits bancs de sorra o fang.
Finalment, cap al Devonià superior, fa uns 370 milions d’anys, apareixen finalment els veritables amfibis, els més antics coneguts dels quals són Ichthyostega, descobert els anys 30 al sud-est de Grenlàndia, i Tulerpeton, descobert el 1984 a Tula, al sud de Moscou, a Rússia.
El cas dels fongs
La nostra atenció per l’evolució de plantes i animals en la seva expansió cap als ambients terrestres no ens ha de fer oblidar el paper dels fongs. A més del cas de plantes i animals, el pla d’organització dels fongs representa la tercera gran estratègia evolutiva de la vida eucariòtica multicel·lular.
Els fongs es desenvolupen a partir d’espores i creixen en forma de tubs finíssims, anomenats hifes, interromputs per parets transversals, anomenades septes. Un aspecte de la seva organització alternativa consisteix en el fet que les cèl·lules fúngiques poden tenir diversos nuclis i que el citoplasma pot passar més o menys lliurement d’una cèl·lula a una altra a través dels septes. A diferència de les plantes, els fongs no obtenen l’aliment per fotosíntesi sinó per digestió i absorció dels productes resultants de la fotosíntesi, però, a diferència dels animals, aquesta digestió es produeix a l’exterior dels seus cossos. Dotats de parets cel·lulars de quitina gruixudes i consistents, aptes per a resistir la dessecació, els fongs són organismes molt adaptables, perfectes candidats a la vida terrestre, com ho confirma el fet que, de les entorn de 100 000 espècies de fong conegudes, la majoria són terrestres.
Ja hem parlat abans del probable paper simbiòtic dels fongs en l’establiment en terra ferma de la vida vegetal. Tot i així, els testimonis fòssils més antics coneguts, trobats precisament en fòssils de teixits vegetals, es remunten només al Carbonífer, fa uns 300 milions d’anys.
L’evolució del paisatge dominat per la vida
Amb l’establiment a la terra ferma de fongs, plantes i animals, tant invertebrats com vertebrats, es pot dir que comencen veritablement els temps “moderns” de la biosfera i es comencen a configurar les complexes relacions entre organismes dels diferents biomes terrestres que avui coneixem.
Tanmateix, tampoc en aquestes etapes més recents l’evolució queda lliure de sotragades. Fenòmens d’extinció massiva com els de les faunes ediacariana, tommotiana i atdabaniana es repeteixen entre la fi del Permià i l’inici del Triàsic, fa 245 milions d’anys, amb la desaparició del 52% de les famílies conegudes del Permià, o al final del Cretaci, fa uns 65 milions d’anys, amb la desaparició de “només” l’11% de les famílies, però entre elles es trobaven tots els dinosaures i moltes famílies de mamífers.
De fet, la concepció de l’evolució com un procés continuat d’avenç cap al progrés, mesurable en termes de diversitat creixent dels organismes o d’emergència d’organismes progressivament més complexos, ha de ser revisada. La història de l’evolució és més aviat la d’una successió d’episodis de diversificació seguits de fases d’extinció massiva i repetició del cicle a partir de les formes supervivents. Com un fènix naixent de les seves pròpies cendres, el DNA s’ha renovellat en noves formes llargament perdurables en superar les fases successives d’extinció massiva. Els humans podem traçar la nostra història i la dels organismes amb els quals compartim avui la biosfera, però hem de recordar que ni la nostra ni cap d’elles en particular és més necessària que les altres perquè la vida continuï després de la propera extinció massiva, que molt bé podríem ser capaços d’ocasionar pels nostres propis mitjans.
Els aiguamolls devonians

Fa 300 milions d’anys, en el període carbonífer, que abasta l’època compresa entre els 363 i els 286 milions d’anys enrere. Aquest període fou decisiu en la història de la Terra, ja que hi ocorregué l’orogènia herciniana, s’incrementà la diversitat de la flora terrestre i aparegueren els primers animals de respiració pulmonar (amfibis). És l’època dels grans cefalòpodes i dels braquiòpodes, que coexistien amb l’expansió dels artròpodes de terra ferma, afavorits pel clima càlid i l’exuberant vegetació. Paral·lelament, els microbis s’expandiren també cap a la vida aèria transportats dins l’intestí dels animals terrestres.
Biopunt
Durant el Devonià, encara sense altres animals terrestres que els artròpodes abans esmentats, la majoria de les plantes que havien començat a conquerir els medis emergits vivien en aiguamolls, on trobaven les mínimes fluctuacions estacionals pel que fa a la llum, la temperatura o la humitat, amb petites diferències de composició florística entre les comunitats vegetals que vivien a les regions més salabroses de deltes o estuaris i les d’aiguals de riberes fluvials o vores de llacs.
La frontera entre Devonià i Carbonífer, fa 360 milions d’anys, resulta una frontera força convincent pel fet que és en aquest darrer període que el món vegetal terrestre experimenta, gairebé tan “sobtadament” com la fauna marina al Cambrià, una explosió de diversitat sense precedents, que va associada amb la coevolució d’una vida animal també força rica. Cap al Devonià mitjà, fa uns 380 milions d’anys, ja hi havia moltes plantes del tipus de les falgueres, anomenades progimnospermes, amb fulles ben desenvolupades, tiges lignificades i dotades d’escorça, i port arbori de fins a una desena de metres d’alt; sembla que foren algunes d’aquestes progimnospermes les primeres plantes a desenvolupar unes estructures semblants a llavors. Això, però, passà durant el Carbonífer.
Els boscos carbonífers i permians
Els primers paisatges de les terres emergides que admeten una certa comparació amb les nostres concepcions actuals són els permians. Són els primers paisatges amb vegetals amb aspecte de planta més o menys convencional, poblats d’animals de tota casta, amb inclusió explícita dels tetràpodes. Però ja durant el Carbonífer el paisatge hauria evolucionat de manera molt espectacular.
L’aparició de la llavor i l’expansió de les plantes vasculars
Durant el Carbonífer, l’aparició de la llavor en les plantes vasculars fou un fenomen transcendent, amb grans repercussions paisatgístiques. En efecte, les llavors de les plantes vasculars representaren l’alliberament de la necessitat de l’aigua per a l’accés dels gàmetes masculins a l’òvul, i foren la innovació evolutiva decisiva que assegurà l’èxit d’aquestes plantes en la colonització de medis cada com més secs. D’altra banda, cal no descartar que l’èxit creixent de les plantes terrestres als aiguamolls comportés un increment fora de mida de matèria orgànica morta en descomposició que empobrís considerablement d’oxigen les aigües estanyades i que aquesta fos una de les pressions de selecció determinants per al pas a la respiració aèria d’alguns artròpodes i peixos. Això mateix explicaria la bona conservació dels fòssils vegetals del final del Devonià i de tot el Carbonífer, el qual deu precisament el seu nom als grans dipòsits de carbó originats per aquesta acumulació de matèria orgànica. Aquesta acumulació demostra que els boscos de licopodis arborescents i progimnospermes foren el bioma terrestre dominant tot al llarg del Carbonífer.
La primitiva fauna carbonífera

L’amfibi més antic que es coneix, de fa 370 milions d’anys, Icthyostega, va ser trobat en roques del Devonià superior de Grenlàndia. En iniciar-se el Devonià, fa uns 400 milions d’anys, els peixos ja s’havien diversificat en tots els grups actuals, i fins i tot alguns dels grups ja s’havien extingit. Al Devonià superior, fa uns 370 milions d’anys, aparegueren els veritables amfibis. Les seves característiques generals coincideixen força amb les dels ripidistis del Devonià superior, però se’n diferencia perquè té les potes més llargues i fortes, no lobulades, i pel costellam, característiques que s’interpreten com a adaptacions a caminar per l’aire lliure. Per les dents que té, suposem que caçava peixos grossos, més que no pas preses petites. Hom pensa que els primers amfibis s’assemblaven més als rèptils d’avui que no pas a les granotes que coneixem; de fet, ens el podríem imaginar com un cocodril d’un metre de llarg, tant per l’aparença com pel tipus de vida.
Biopunt
En aquests boscos carbonífers, encara sense vertebrats, regnaven els artròpodes, tant herbívors com depredadors, que anaven configurant la gran varietat de nínxols ecològics que encara avui tenen els boscos humits tropicals. Amb el creixement d’espècies arbòries, per altra banda, la vida aèria prenia tot el sentit per ais animals; la vida deixava de quedar limitada a la bidimensionalitat de la superficie del sòl i començava a ocupar tot el volum de la tridimensionalitat de la massa arbòria, si molt convenia, amb els primers assaigs de vol a càrrec d’alguns insectes. Fou en algun bosc d’aquesta mena, a la seva vorada o en alguna clariana, que els primers amfibis com ara Ichthyostega començaren a caminar. La varietat que assoliren ràpidament, prova del seu èxit evolutiu, reflecteix el fet que eren els primers animals d’una certa grandària que explotaven tant el medi del qual procedien, és a dir les aigües poc profundes de les vores de llacs i aiguamolls, com un medi nou, les terres ja del tot emergides properes als aiguals.
La majoria dels amfibis del Carbonífer eren carnívors; els més petits menjaven insectes, i els més grans també insectes a més d’altres artròpodes (cal tenir present que per alguns boscos carbonífers circulaven artròpodes de mida considerable, entre els quals podem destacar alguns milpeus de fins a tres metres i mig de llarg, i hi volaven insectes de grandària no menys respectable, com ara espiadimonis de fins a 60 cm d’envergadura), peixos, amfibis petits i, tan aviat com n’hi hagué, petits rèptils. També eren carnívors els centpeus i els espiadimonis, que s’alimentaven d’artròpodes més petits, mentre els petits artròpodes com també els grans milpeus, que s’alimentaven de fusta, eren herbívors.
L’aparició de l’ou amniòtic i l’expansió permiana dels rèptils
Els amfibis, però, mai no arribaren a alliberar-se totalment de l’aigua. Encara avui els amfibis necessiten fecundar els seus ous i desenvolupar les seves larves a les aigües dolces.
L’alliberament definitiu de la dependència del medi aquàtic havia de ser assolida per un nou grup de tetràpodes, el dels rèptils, mitjançant una innovació evolutiva particularment reeixida: l’ou amniòtic, és a dir, l’obtenció d’una estructura susceptible de mantenir un ambient aquàtic en un clos a l’entorn de l’òvul fecundat i de l’embrió en creixement fins que aquest assoleix un estadi de desenvolupament tal que pot prescindir-ne. Aquesta adaptació, comparable a la que suposà per a les progimnospermes la llavor, és molt probable que es produís en alguna espècie d’amfibi de petites dimensions del grup dels antracosaures, potser semblant a algunes salamandres arborícoles actuals de les selves d’Amèrica del Sud, que hagués començat a adaptar-se a una forma de vida d’actiu depredador d’insectes, aranyes i altres artròpodes a la capçada dels gran arbres dels boscos carbonífers. Una altra innovació transcendental vinculada al pas dels vertebrats de la vida aquàtica a l’aèria és la síntesi de la ceratina, una de les poques innovacions metabòliques que no tenen precedents en el món dels procariotes. La ceratina, la proteïna característica de les escates de la pell dels rèptils, de les plomes dels ocells i del pèl i les banyes dels mamífers, permeté als vertebrats terrestres resistir a la dessecació un cop han abandonat l’ou. Per això la capacitat de sintetitzar-la resulta decisiva.
Els primers vertebrats terrestres eren tots carnívors. La major part del material vegetal conté una gran quantitat de cel·lulosa i els vertebrats no la poden digerir si no la masteguen molt bo i no troben la manera de reclutar i mantenir una població de bacteris i protists fermentadors de la cel·lulosa que visqui simbiòticament en el seu tub digestiu, tal com fan avui els remugants al seu estómac. Només alguns productes vegetals, especialment els que constitueixen els òrgans reproductius, resulten prou rics de proteïnes i sucres amb relació al seu contingut de cel·lulosa perquè no sigui necessari mastegar i digerir un gran volum d’aliment vegetal per a satisfer les necessitats d’un animal. A la pràctica, això porta a dues vies adaptatives diferents pel que fa als herbívors. Una seleccionaria les petites dimensions, la intensa activitat i l’especialització en determinats productes alimentaris d’origen vegetal (nèctar, pol·len, sucs vegetals, fruits, llavors); aquest seria el cas avui de molts petits mamífers i ocells. L’altra seleccionaria la capacitat de coevolucionar amb poblacions bacterianes simbiòtiques capaces de digerir la cel·lulosa al seu pas per determinades parts del tub digestiu i la capacitat de desplaçament a la recerca de pastures o fulles, cosa que sovint porta associada una talla més gran. Alguns amfibis i rèptils del Carbonífer superior i del Permià superior esdevingueren herbívors seguint la colonització pionera de les terres emergides d’una certa altitud realitzada per les plantes vasculars i explotant els medis de les vorades dels exuberants boscos carbonífers d’aiguamolls o de sòls xops d’aigua. Hom es pot fer una idea de la importància d’aquesta vegetació arbustiva o herbàcia segurament força comparable a una sabana o a una sabana arbrada si es té en compte animals com l’amfibi Diadectes,un dels més antics tetràpodes herbívors, que podia arribar a mesurar 4 m; les seves necessitats alimentàries exigien considerables extensions poblades de vegetals aptes per al seu consum.
Els primers testimonis fòssils de rèptils corresponen a petits animals associats amb fòssils vegetals de grans licopodis arborescents conservats drets, tal com havien viscut fa uns 310 milions d’anys, en estrats corresponents al Pennsilvanià inferior de Nova Escòcia, al SE del Canadà. Com que de fet s’han trobat dins dels troncs podria ser que, tal com pensa Richard Cowen, de la University of California, a Davis, aquests rèptils més antics visquessin en forats dels troncs tal com fan encara avui alguns mamífers insectívors a les selves plujoses equatorials i visquessin de manera semblant a com viuen aquests.
L’èxit evolutiu dels rèptils fou gran, i al llarg dels temps permians (entre els 290 i els 245 milions d’anys enrere), en una espectacular diversificació adaptativa, desplaçaren totalment els amfibis de la immensa major part dels hàbitats terrestres i, al llarg del Triàsic, fins i tot de molts dels aquàtics, als quals s’adaptaren secundàriament alguns rèptils.
Els paisatges mesozoics
Els esdeveniments tectònics del final del Permià posaren fi als climes equatorials humits que havien afavorit el complex sistema de llacs, deltes, aiguamolls i estuaris al llarg de tota la costa meridional del continent euro-nord-americà (Euramèrica) en el qual s’havien desenvolupat els boscos carbonífers i les seves faunes.
De les gimnospermes a les angiospermes
El clima del nou continent boreal de Lauràsia, desplaçat cap al N pel seu xoc amb el continent austral de Gondwana, esdevingué més sec i les flores reflectiren el canvi. Les flores paleofítiques, dominades per licopodis i progimnospermes, donaren pas a flores mesofítiques, que durant el Triàsic cobririen ja pràcticament tota la terra ferma, dominades per les gimnospermes, principalment coníferes, ginkgos i cícades, més ben adaptades, sobretot les coníferes, per a viure en ambients més secs.
Al llarg del Cretaci s’expandeixen les angiospermes, que apareixen al registre fòssil a partir del Cretaci inferior, fa uns 100 milions d’anys. Fins al començament del Cretaci, les flores del Mesozoic havien estat dominades per les gimnospermes, que durant el Juràssic arriben a representar el 80% dels vegetals conservats al registre fòssil, però durant el Cretaci les angiospermes experimenten una ràpida expansió que, iniciada a les latituds baixes, les condueix, al final del Cretaci, a ser les plantes dominants a tot el planeta, com encara són actualment.
No es coneix amb certesa l’origen filètic de les angiospermes, tot i que hi ha raons per pensar que procedeixen de formes afins a les pteridospermes i que s’originaren en temps força anteriors als dels testimonis més antics que ens n’han pervingut en el registre fòssil. El que sí que es coneix és que una de les claus de l’èxit de les angiospermes ha estat el pas d’una pol·linització anemòfila a una pol·linització entomòfila, la qual ha comportat una coevolució entre plantes i insectes i un increment en l’eficàcia de la dispersió de les llavors a distància, sovint també mitjançant la participació d’animals, sobretot a partir de l’aparició i expansió d’ocells i mamífers.
L’esplendor reptilià
Al llarg del Permià la fauna, tal com hem dit, s’empobrí d’amfibis i s’enriquí de rèptils, ja alliberats de la dependència de l’aigua per a la reproducció i, per això mateix, més ben dotats que aquells per colonitzar medis més secs i d’ambient menys constant que les selves humides del Carbonífer. La diversitat d’ambients que colonitzaven afavorí igualment la diversificació dels mateixos rèptils. De soques reptilianes aparegudes al Permià deriven, entre altres, grups tan importants com els dinosaures (extingits al final del Triàsic), els ocells i els mamífers.
Aparegueren rèptils carnívors i herbívors, terrícoles i aquàtics, i alguns desenvoluparen diferents formes de planejar i de volar; envaïren tots els continents i tots els medis. El Mesozoic, en definitiva, fins a la fi del Cretaci (fa uns 66 milions d’anys) fou l’era del rèptils, tot i que alguns grups d’aquests, en la seva evolució, ja havien originat els primers ocells (durant el Juràssic) i els primers mamífers (durant el Cretaci), el paper dels quals encara era relativament negligible a la majoria dels ecosistemes mesozoics.
L’aparició dels ocells i dels mamífers
L’origen evolutiu dels ocells encara roman fosc, a causa sobretot de l’escassetat de testimonis fòssils. La lleugeresa i la fragilitat dels ossos dels ocells, les dimensions generalment modestes i els hàbitats que han ocupat normalment aquests animals fa que resultin mals candidats a la fossilització. Archaeopteryx,l’ocell més antic de què es coneixen testimonis fins ara, apareix d’una manera més aviat sobtada en el registre fòssil del Juràssic superior, fa entre 140 i 170 milions d’anys, a la famosa pedrera de calcària litogràfica de Solnholen, a Baviera, on fou descobert el 1861; ja posseïa alguns dels trets (plomes, presència de fúrcula, forma de la caixa craniana i de les extremitats anteriors) que caracteritzen un ocell modern, tot i que té molts més caràcters en comú amb els dinosaures teròpodes (mandíbules dotades de dents còniques, esquelet de la cua i la pelvis, etc). Més recentment, en roques del Triàsic mitjà de Catalunya d’uns 240 milions d’anys enrere i del Triàsic superior de Texas d’uns 220 milions d’anys, han estat trobats fòssils més antics que podrien correspondre també a avantpassats d’ocells relacionats amb petits rèptils bípedes, de dents punxegudes i cua llarga, però ja dotats de fúrcula i d’extremitats anteriors força llargues en les quals semblarien observar-se petits bonys, potser identificables com a insercions de plomes. En els temps posteriors a Archaeopteryx,en canvi, l’evolució dels ocells està força més ben documentada i sembla haver estat ràpida i diversificada des del Cretaci inferior.
Els cinodonts, de la soca reptiliana dels quals havien de derivar els mamífers, aparegué ja en el Permià superior, fa entre 260 i 245 milions d’anys. Com en el cas de la soca d’amfibis de la qual havien derivat els rèptils i en el de la soca reptiliana de la qual ho farien els ocells, es tractava d’animals de petites dimensions, comparables als eriçons actuals, que vivien sobretot al continent austral de Gondwana en ambients amb acusats canvis d’estacions, i foren particularment abundants i diversificats durant el Triàsic. Durant aquest període, com a mínim sis grups de cinodonts, tant carnívors com herbívors, desenvoluparen caràcters propis de mamífers, si bé sembla que fou un grup de petits carnívors el que donà origen, cap a la fi del Triàsic, fa uns 215 milions d’anys, als primers mamífers pròpiament dits.

Fa 200 milions d’anys, a l’era mesozoica primerenca, es desenvoluparen els primers mamífers, mentre que els rèptils ocupaven mars i cels. Les diferències més grans entre els rèptils i els mamífers no estan només en l’esquelet, sinó que també afecten altres caràcters. Pel que fa a la reproducció, els rèptils ponen, i ponien, grans ous amb força material de reserva per al creixement de la cria; els mamífers tenen cries de mida petita i, per tant, els nounats han depès, des de sempre, de la cura paterna.
Biopunt
Els primers mamífers eren menuts (alguns arribaven amb prou feines als 10 cm de llargada des de la punta del musell a la base de la cua) i els seus fòssils són rars i difícils de trobar si no és rentant i garbellant enormes volums de sediments. Devien ser semblants als petits insectívors actuals i, com ells, devien ser de vida nocturna, la manera més eficaç per als animals petits capaços de regular la seva temperatura de competir amb animals més grans però de sang freda i, per això mateix, menys actius a la nit. Tot i que possiblement hi hagué rèptils homeoterms, en general les baixes temperatures i l’aire fresc de la nit inhibeixen l’activitat dels rèptils. Alerta i en moviment en la fosca de la nit, amb ulls penetrants i dilatats, els primers mamífers s’apartaren de la dependència de l’escalfor del Sol i s’escamparen arreu del món. A mesura que s’allunyaven tant de l’escalfor dels tròpics com dels rèptils que hi vivien anaren perfeccionant els seus mitjans d’aïllament per mantenir la constància de la seva temperatura interna. Potser algun desenvolupà plomes com els ocells, però la majoria desenvoluparen pèls, un nou ús de la ceratina adreçat a la protecció enfront de les baixes temperatures. En la seva evolució a partir dels cinodonts, les femelles dels mamífers anaren deixant de pondre els seus ous en clots a terra a l’aire lliure; en comptes d’això els començaren a alimentar dins seu al si de l’escalfor d’un òrgan especialitzat que, en els més evolucionats, esdevindria l’úter. Després del naixement, les cries afamades podien llepar les secrecions d’algunes glàndules del ventre de les seves mares. Aquesta secreció, inicialment només suor enriquida amb calci, esdevingué el líquid orgànic més ric en aquest element: la llet.
Des del seu començament fa uns 200 milions d’anys tant ocells com mamífers es diferencien dels seus ancessors reptilians i de la majoria dels altres rèptils —llevat potser d’alguns dinosaures— per l’especial atenció i cura que dediquen a les seves cries. La majoria dels rèptils tenen abundant descendència, que abandonen després de la posta o, a tot estirar, així que ha sortit de l’ou. Ocells i mamífers, per contra, alimenten i ensinistren una descendència menys nombrosa i més vulnerable, que requereix aquesta cura per a reeixir a assolir la vida adulta.
En el Cretaci inferior apareixen els primers mamífers placentaris, és a dir amb una estructura que fa de contacte entre l’embrió en creixement i l’úter de la mare i a través de la qual s’estableix el pas de substàncies nutritives i oxigen de la mare a l’embrió i de les substàncies d’excreció i diòxid de carboni de l’embrió a la mare. El desenvolupament de la placenta ha permès als mamífers placentaris, a diferència dels marsupials, tenir temps de gestació més llargs, en els quals les cries assoleixen un desenvolupament més avançat, i uns temps de lactància més curts abans d’esdevenir independents de la mare per a llur alimentació. Tot i els avantatges que ara ens semblen evidents, durant els temps cretacis els mamífers, fins i tot els placentaris, mantingueren una presència discreta arreu. Els animals dominants eren els dinosaures, els quals, apareguts durant el Juràssic, fa uns 215 milions d’anys, conegueren al llarg del Cretaci una expansió i una diversificació sense precedents en cap altre grup d’animals que els portà a colonitzar tots els ambients, tant terrestres com aquàtics, a assajar el vol, a assolir unes dimensions mai més no igualades per cap altre animal —alhora, també hi havia dinosaures no gaire diferents per la talla dels primers ocells o els primers mamífers que hem descrit—, fins i tot a assajar el viviparisme, Simultàniament altres grups de rèptils, com els avantpassats de tots els que viuen actualment, assolien també un moment dolç de diversificació i especiació.
Els biomes cenozoics
Tanmateix, la fi del Cretaci i l’inici del Cenozoic, fa uns 66 milions d’anys, és marcat per un episodi d’extinció massiva en el qual desapareixen totalment els dinosaures i altres formes de vida, com molts grups planctònics, els ammonits i moltes famílies de plantes.
El trasbals d’una ruptura poc aclarida
Sembla que cal cercar la causa d’aquestes extincions en la col·lisió amb la Terra d’algun planetoide, segons demostraria l’estranya abundància d’iridi, un metall extremament rar a la Terra, però abundant als meteorits en els sediments de la frontera entre el Cretaci i el Cenozoic. Això fou observat per primer cop a Gubbio, a l’Úmbria, al centre de la península Itàlica, però després s’ha confirmat a més d’una dotzena d’altres punts, des de sediments del fons de l’oceà Pacífic fins a diversos punts d’Europa i de l’Amèrica del Nord; contemporàniament s’ha observat un empobriment del plàncton fòssil i de la proporció de grans de pol·len que apareixen als sediments. Sembla que el xoc degué produir un núvol de pols meteorítica que fou la causa d’un prolongat enfosquiment del conjunt —o si més no d’una gran part— de la superfície de la Terra (la simple erupció del volcà Tambora, l’any 1815, representà l’any següent l’absència d’estiu climàtic en àmplies zones de la seva perifèria). D’això s’hauria derivat un refredament important de la superfície i una minva espectacular de l’activitat fotosintètica tant a la mar com en terra ferma. Els cadàvers de bacteris i protists fotosintetitzadors i de les plantes s’haurien acumulat i la seva mort hauria ocasionat la fam de tots els organismes que depenien d’aquests productors per a la seva alimentació, amb la seva consegüent extinció.
Darrerament, arran de l’estudi de les colades de Deccan, a l’Índia, s’ha proposat una nova hipòtesi segons la qual un seguit d’erupcions generalizades també podrien explicar l’enriquiment en iridi, com també altres anomalies químiques dels sediments del límit entre el Cretaci i el Cenozoic. Ara bé, aquests dipòsits volcànics haurien requerit un període de deposició d’uns 10 000 anys, una xifra elevadíssima al costat dels mesos requerits per la hipòtesi de l’asteroide. L’escassa resolució dels sistemes actuals de datació (superior a 1’1) no permet encara de descartar cap de les dues hipòtesis. En tot cas, les grans extincions, que en principi es mostren com a desastroses, podrien haver tingut un paper decisiu en l’evolució de la vida complexa. L’extinció dels dinosaures, vista des de la perspectiva d’aquestes hipòtesis, més que no pas un afer de gens podria haver estat, simplement, un afer de mala sort. En definitiva, dinosaures i ammonits, abundants durant desenes de milions d’anys, despareixien per sempre més, juntament amb foraminífers, belemnits, rudistes i altres grups sencers. La vida a la Terra emprenia nous camins. Entre els grups supervivents hi hauria els mamífers que iniciaren una fase explosiva d’evolució.
De tota manera, aquestes imatges apocalíptiques potser són un punt exagerades. La majoria de les plantes terrestres no semblen afectades pels esdeveniments de la frontera entre el Cretaci i el Cenozoic, com ho demostra el fet que hagin deixat testimonis fòssils tant abans com després. Així i tot, és un fet que l’impacte es produí i que pogué causar importants alteracions de la temperatura, la il·luminació, les pautes del temps atmosfèric, el nivell de la mar i molts altres factors que hagin pogut ocasionar la destrucció d’una gran part (encara que, certament, no de totes) de les comunitats d’organismes, i en tot cas l’extinció de les espècies amb més dificultats d’adaptació a canvis sobtats com els que es produïen.
La flora i la fauna cenozoiques
Fa 50 milions d’anys hi hagué una expansió important de la flora i la fauna del planeta. Els animals que més s’expandiren foren els mamífers i els ocells. Els ocells colonitzaren el medi aeri mentre que els mamífers colonitzaren també altres medis: aquàtic, aeri i terrestre i, dins d’aquest últim, explotaren tots els nínxols ecològics.
Biopunt
Les plantes més tolerants a canvis estacionals i la fauna que s’hi associa reeixiren a sobreviure amb un cert èxit. Així, amb el Cenozoic s’implanta al conjunt del planeta una flora que no ha canviat substancialment des de llavors, llevat dels reajustaments d’àrees motivats pels canvis climàtics o els moviments tectònics que s’han produït en els darrers 65 milions d’anys. Amb la desaparició dels dinosaures, ocells i mamífers tenen la porta oberta a una expansió explosiva i una diversificació en la qual l’èxit dels segons fou tan gran que passaren de vuit famílies a setanta en uns 10 milions d’anys, entre el començament del Cenozoic o Terciari i la fi del Paleocè, la seva primera sèrie.
Finalment, cal afegir que coincidint amb les etapes fredes del final de l’Oligocè i del principi del Miocè es comencen a desenvolupar algunes formes de vegetació noves, riques d’espècies herbàcies i sobretot de gramínies, en latituds tropicals (sabanes) i temperades (praderies i estepes). Ofereixen recursos alimentaris a noves formes de mamífers, tant herbívors com carnívors, més grans que les existents fins llavors, de la qual cosa resultà una nova explosió evolutiva que afectà primordialment els mamífers.
L’aparició dels primats
Entre aquestes famílies aparegudes al començament del Cenozoic, fa uns 65 milions d’anys, interessa particularment la dels nostres directes avantpassats i pròxims parents: els primats primitius. Els primers primats eren petits i espavilats insectívors arborícoles, ràpids a saltar i fugir dels depredadors. Alguns fins i tot s’aferraven a les branques amb les urpes modificades que en els humans han esdevingut els dits de les mans i els peus. Saltant de branca en branca enmig de la nit afavoriren la selecció de l’agudesa i la precisió de la visió en els seus descendents i a poc a poc els seus ulls devien passar d’una posició lateral a una posició centrada frontalment que possibilita una visió estereoscòpica, indispensable per a l’apreciació de les distàncies en els moviments per les capçades dels arbres. Es devien assemblar força als actuats pro-simis, que sobreviuen únicament a Madagascar i en alguns punts del sud-est d’Àsia.
Els primats sobrevisqueren al següent episodi d’extincions massives, el tall abrupte entre l’Eocè i l’Oligocè, fa uns 38 milions d’anys, durant el qual s’extingiren no poques famílies de mamífers, en particular a l’Amèrica del Nord, i també nombrosos organismes planctònics, a causa d’un nou refredament climàtic. Durant l’Oligocè, fa uns 30 milions d’anys, els primats es diferenciaren en diverses branques (una de les quals donaria origen als homínids) que apleguen tots els antropoides actuals i l’home, juntament amb els seus avantpassats extingits i altres primats emparentats. L’evolució d’aquest grup, que interessa especialment en primer lloc perquè és el nostre, però també perquè ha esdevingut un agent nou, molt important i actiu, de modificació de la biosfera, serà objecte de la tercera part d’aquest volum.