Les regles del joc
La diversitat, l’escala i el temps
Hi ha una gran varietat d’éssers vius. L’organització de cada individu és jeràrquica. Unes entitats són formades d’altres entitats menors i aquesta organització es reflecteix en les relacions funcionals. També es descobreixen relacions jeràrquiques en els ecosistemes, formats per molts organismes individuals, generalment de diferents espècies. Hom pot suposar una relació reproductiva o de descendència entre els individus d’una mateixa espècie: entre individus de diferents espècies hi poden haver relacions de tipus molt divers. Per exemple, unes espècies serveixen d’aliment a les altres; en d’altres casos s’estableixen formes molt diverses d’interacció, directes o indirectes: els arbres fan de suport a les lianes, molts insectes pol·linitzen les flors, etc. Els ecosistemes contenen grups d’individus de diverses espècies que conviuen. La paraula ecosistema fa referència a sistema, o suggereix el concepte de sistema, quelcom fet d’elements components que són relacionats els uns amb els altres per lligams més o menys flexibles. En aquest sentit, els àtoms també són sistemes i cap sistema no és absolutament rígid. Podem dir que el món és un gran sistema fet de sistemes menors de naturalesa molt diversa, a diferents escales. Dins d’un sistema, el que anomenem jerarquia és una conseqüència del fet que no totes les connexions possibles es realitzen amb la mateixa intensitat.
Els sistemes vius són molt més rics d’estructura si hom els contempla a petita escala. Rarament les seves parts homòlogues, a totes les escales, són exactament duplicats les unes de les altres. Aquest fet fa que en certa manera sigui inapropiat l’ús d’una estadística que ha estat pensada per a ésser aplicada a unitats comparables entre les quals les diferències es poden qualificar d’aleatòries. Mai no trobarem subunitats perfectament comparables i les diferències entre unes i altres mai no es podran considerar aleatòries.
Per a caracteritzar els sistemes vius no basten ni aquestes consideracions ni d’altres de semblants de caràcter també estàtic, sinó que cal introduir el factor temps. El temps és consubstancial amb la vida: el sentim com una mena de consciència interna del món, que fa de rellotge. Certament, els sistemes vius són sistemes físics: tots els sistemes vius tenen les característiques i les limitacions del sistemes físics en general. Si hi ha alguna diferència, sembla que aquesta consisteix principalment en la capacitat per a recuperar d’una manera molt efectiva, amb molt bon rendiment, i en forma de complexitat o informació, l’equivalent d’una part molt considerable de l’augment d’entropia que ha ocorregut en l’organisme i en el seu entorn immediat.
L’entropia i la irreversibilitat dels canvis
El concepte d’entropia correspon a una manera de comptabilitzar tots els canvis físico-químics que es van realitzant. D’una manera massa simplificada acceptarem que l’entropia va sumant un component irrecuperable de tots els canvis, tant si van en un sentit com si van en el sentit invers. Té sentit perquè en tots els canvis hi ha transformacions entre treball mecànic i calor, i hi ha sempre una petita part d’aquesta calor produïda que es difon i es perd sense remei. No hi ha manera de desfer el que ja ha esdevingut, ni de seguir una trajectòria que sigui exactament inversa a la d’un canvi material realitzat abans. Aquesta irreversibilitat es pot veure i interpretar de maneres diferents.
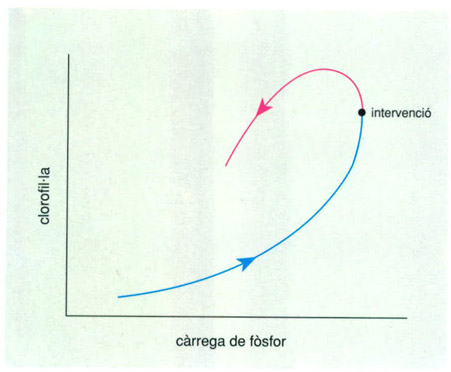
Processos d’eutrofització i de recuperació d’un llac expressats per la relació entre la càrrega de fòsfor i el contingut de clorofil·la. La via d’anada i la de tornada, després d’una intervenció exògena, mai no és la mateixa.
Editrònica, dades elaborades per l’autor
En ecologia, la irreversibilitat dels canvis és una experiència quotidiana i té un interès no solament científic i conceptual, sinó també pràctic. Per exemple, en el procés de l’anomenada eutrofització d’un llac, l’aigua s’enriqueix de materials nutritius i esdevé verda, perquè conté moltes algues microscòpiques en suspensió (i de vegades no tan microscòpiques, sinó en forma de fins cabells verds). El canvi ocorregut en el llac es pot descriure per la variació de la concentració de clorofil·la en la zona il·luminada del llac, la qual depèn de la càrrega de fòsfor que el llac rep, probablement amb les aigües de clavegueres i els sobrants del reg. Si hi posem remei i tractem de recuperar la situació inicial del llac, per exemple construint depuradores que funcionin, el camí descrit per la mateixa funció (clorofil·la en dependència del fòsfor) durant el període de recuperació és diferent del camí d’anada. L’examen de la representació gràfica del procés en un sistema de coordenades (on x és la càrrega de fòsfor, i y és clorofil·la) suggereix que la integral que correspon a la superfície del gràfic compresa entre la trajectòria associada a l’eutrofització i la que descriu el retorn a una condició acceptable, correspon al cost social de l’operació. Hom podria aquí recordar els diagrames que representen funcions associades a les propietats de la barreja de gasos en un motor d’explosió, i fins i tot reconèixer-hi les correspondències amb l’exemple dels canvis en el llac. En el cas d’un riu, el camí de tornada, de recuperació, pot ser molt més proper al d’anada, que descrigué la contaminació creixent: efectivament, el riu s’embruta aviat, però si es vol també es neteja ben de pressa. Un llac, i encara més la mar, o un aqüífer, triguen a embrutar-se, però després costen molt més de netejar.
Tot això ve del fet que en els sistemes vius i en el seu entorn immediat es realitzen contínuament transports de materials i reaccions entre materials, els quals, en la mesura que corresponen a canvis no reversibles, es poden anar apuntant en un compte d’entropia. Aquest esdevenir té una rèplica o un reflex en la informació, en la complexitat que va adquirint el mateix sistema. Els sistemes biològics, a totes les escales, aconsegueixen de recuperar una fracció important d’aquella entropia en forma del que anomenem complexitat o informació. És com si qualsevol bescanvi d’energia deixés una marca o un senyal en la ma-tèria: un cop ocorregut, ni la matèria ni l’energia no són exactament igual que abans. Una mateixa energia no es pot usar de la mateixa manera dues vegades seguides. La matèria sempre servarà algun record del bescanvi. També l’escorça terrestre preserva, en forma d’informació (plecs, falles i tota l’enorme riquesa d’estructures que la geologia estudia) un registre que conté molts indicis dels canvis materials i energètics que tingueren lloc en el passat. Però els organismes són particularment efectius en aquesta tasca, i això ho deuen en primer lloc a la complicació que tenen ja de sortida i, a més, a l’aptitud, molt gran, que tenen d’enrotllar les memòries (de diverses qualitats) sobre elles mateixes, complicant-les de moltes maneres i relacionant-les les unes amb les altres.
S’ha escrit repetidament que els sistemes són més que la suma de les seves parts; igualment es podria dir el contrari, és a dir, que són menys, perquè cadascun dels seus components, en entrar a formar part d’un sistema, veu escapçades una bona part de les potencialitats que abans tenia; el seu ulterior esdevenidor queda integrat en un sistema que limita i canalitza les possibibitats de canvi i d’evolució que tenia a priori. Solem queixar-nos que la societat limita les nostres opcions. Això és obvi en l’evolució: el fet que un organisme hagi d’emmotllar-se a les exigències de l’entorn i a tots els condicionaments que li posen els individus de la seva mateixa espècie i els de les altres espècies, fa que no s’hi puguin realitzar totes les possibilitats de canvi que oferiria el sistema genètic. És per això que l’evolució real de les espècies mai no arriba a aconseguir, ni de bon tros, la velocitat que el sistema genètic permetria a l’evolució si els fenotips no estiguessin sotmesos a tantes especificacions per passar la prova de la vida real. De fet, en situacions simplificades, sigui en el laboratori, sigui en la biosfera, després de cataclismes destructors, l’evolució ha pogut recuperar una rapidesa que s’apropa més a la possible. Desconnectar les espècies del lligam constituït per les relacions usuals en l’ecosistema, com ha esdevingut en les plantes i en els animals domesticats per l’home, ha permès una acceleració de la seva velocitat d’evolució. Darwin se sentí atret per l’estudi de la variabilitat de les plantes i els animals en domesticació, els quals li proporcionaren molts indicis sobre la variabilitat de les espècies; realment aquests animals i aquestes plantes han pogut canviar amb una rapidesa excepcional després d’haver-se alliberat dels lligams complexos que tenien en els seus respectius entorns originaris.
La complexitat i la informació
Complexitat i informació s’han usat de manera imprecisa i amb una certa indiferència. Potser sí que es poden considerar sinònims o que, almenys, presenten les mateixes dificultats pel que fa a la seva quantificació. En el llenguatge corrent apreciem la complexitat en una situació simultània o sincrònica, mentre que la informació mentalment s’associa més sovint amb el missatge que fa arribar una complexitat potencial del passat al futur. El problema que fa científicament intractables aquests conceptes és prou simple: la seva mesura. Potser la proposta més raonable és prendre com a mesura la llargada d’una expressió equivalent, formulada en un llenguatge normalitzat que descrigui la situació sense ambigüitat. Ara un hom s’inclina a veure la descripció més aviat com una corrua de zeros i d’uns, i tracta de traduir la sèrie primitiva en un altra sèrie expressada d’aquesta manera. La dificultat, molt suggestiva i inquietant, és que el valor d’un element d’informació, d’una part d’un missatge, depèn del missatge complet. Per això dos missatges molt llargs, com els codis genètics de l’home i del ximpanzé, no cal que difereixin en una extensió molt gran del seu genoma per a ser profundament divergents.
En termes de capacitat de manipular informació, un ordinador de mida doble és més que dos ordinadors de mida senzilla, i un cervell gros és més que una massa equivalent de cervells petits. Aquest és un dels avantatges que tenen els organismes grossos sobre els petits, especialment si a més viuen més temps. Però tot, és clar, depèn de com són els canvis de l’entorn. Tant el missatge com la capacitat de manipular la informació que conté, són informació; hi ha, doncs, un parallelisme amb el “software” i el “hardware” dels nostres ordinadors.
Podríem considerar que la informació és proporcional a una potència superior a 1 de la massa suport (o d’alguna expressió que combinés aquesta massa amb la seva durada), és a dir: I → Mk, k > 1. Ara bé, pensant en una quantitat (o unitat) d’informació i que s’hi afegeixi, es pot escriure I + i → (M+i)k, i → (M + i)k – Mk.
Aquestes consideracions no fan més fàcil el fet de parlar d’informació, tant més que en moltes disciplines es considera la informació com quelcom immaterial, probablement perquè una llargada semblant d’un missatge informàtic pot contenir el que cadascú pot interpretar com a sublimitats o com a bestieses. Hi entra, doncs, la interpretació de qui rep i dóna sentit al missatge. Però encara que es pugui veure la informació com una cosa “immaterial”, almenys és necessària una energia per a manejar-la i transcriure-la i revelar-la; a més, sempre hi ha un augment d’entropia associat a canvis materials, a la base de la seva generació i adquisició.
D’una manera o altra, la vida ha trobat la manera de projectar vers el futur un repertori inacabable de dades: com en les “cançons” de les balenes, el registre genètic transmet una mena de poema èpic, al qual cada generació ha pogut esmenar o afegir quelcom. L’allau d’informació és imparable, i les pertorbacions periòdiques mai no l’arriben a aniquilar totalment; com a màxim netegen una petita part de la taula per a reiniciar parcialment noves complicacions del missatge de la vida. És més ric que un inacabable castell de focs d’artifici. Això es manifesta a tots els nivells. L’evolució orgànica n’és un exemple. A l’escala de la biosfera i dels ecosistemes, l’allau d’informació sempre és present, i precisament és una de les raons perquè l’estudi de l’organització, l’evolució i la complexificació dels sistemes vius sigui sempre tan fascinant.
El sentit de la jugada
L’estratègia causal contra la il·lativa
La tradició naturalística vol que tot tingui sentit i sent una especial delectació cada vegada que reconeix alguna evidència que això és així. Abans es veia com una prova de l’acció de la providència de Déu, manifesta en la cura de les seves criatures; després s’ha vist com una prova de l’eficàcia de la selecció natural, capaç de treure el bo i millor de totes les possibilitats. Allò que tenen en comú aquestes dues visions ha estat present en tota contemplació i interpretació de la natura viva. En canvi, és notable el fet que els observadors de la natura física (de la formació dels planetes i de la Terra, posem per cas) no haguessin necessitat aquesta extrapolació, ja que es limitaren a descriure uns mecanismes i uns resultats. El fet que la història es manifesti més directament i òbviament en els éssers vius, i també la particular posició del temps en les nostres vivències, són factors que segurament són a la base de l’interès que desvetlla el desenvolupament històric dels vivents.
Hom ha interpretat les “diferents” maneres de sobreviure com l’expressió d’estratègies de la natura, usant la paraula estratègia en el sentit que li han donat els militars: un pla que se segueix amb una finalitat definida. Però la persistència d’una trajectòria orientada, que sigui quelcom més que simplement viure, rarament es manté molt temps en les suposades línies filètiques de l’evolució. Pensem, a més, que diverses estratègies explicades per polítics i militars són inventades post factum per trobar unes justificacions o vanagloriar-se d’uns fets ocorreguts i ja sense remei. Potser no val la pena entossudir-se en l’adaptació del concepte d’estratègia, encara que es pugui dir, figuradament, que la vida està jugant milions de partides d’escacs o lluitant en milions de batalles cada dia, i que es puguin recollir estadístiques de les baixes, i fins i tot dels colors dels uniformes dels guerrers.
Estratègies de supervivència

Paral·lelismes i contrastos entre estratègies de supervivència. Encara que no val la pena proposar classificacions gaire formals de les estratègies, potser pot resultar suggeridor pensar que les maneres de fer de la vida, encarnada en els organismes, es poden ordenar al llarg d’un eix segons algunes tendències que són homologables i que es poden reconèixer en moltes situacions. Els paral·lelismes que es presenten en el quadre, la major part més aplicables als animals que no a les plantes, encara que els seus trets bàsics es poden reconèixer també en els representants del regne vegetal, hi poden ajudar. En qualsevol ecosistema hi ha lloc per a molts organismes, però rarament es perd el relatiu contrast entre les espècies que poden aprofitar-se ràpidament d’una energia disponible i les espècies que es refien més de l’acumulació d’organització material i que possiblement fan de pont en el temps de manera més efectiva. Fixem-nos com aquesta classificació és independent de la que es pugui establir entre productors primaris, osmòtrofs i fagòtrofs amb relació al cicle de materials i al fluir de l’energia, però, tot i amb això, és indiscutible que els fagòtrots, per exemple, senten més vocació per les vies evolutives de la columna de la dreta.
El compromís social o ecosistema
No sabem si en algun moment de la història hi hagueren organismes-ecosistema primitius d’organització particularitzada, és a dir, organismes cadascun dels quals era fet a mida del lloc on s’originà, i en els quals es compilen totes les funcions que avui trobem diversificades: des de la fotosíntesi i l’assimilació dels components necessaris, fins a l’esmerçament de tot en la respiració. La subdivisió d’aquests hipotètics organismes-ecosistema, si existiren, en fragments complementaris i separadament reproduïbles, probablement aparegué aviat (hi ha qui no parlaria de vida fins en aquest moment) i donà lloc a ecosistemes com els actuals, integrats per moltes espècies. Cada espècie, al mateix temps que manté una successió de generacions dins l’ecosistema, pot evolucionar amb una certa independència i amb una relativa rapidesa.
Un ecosistema pot arribar a resultar difícil de definir i de delimitar. Apareix format per un conjunt d’espècies o entitats prefabricades i reproduïbles, capaces de ser mesurades quant a la seva massa o al seu nombre d’individus. Hom ha pogut comparar l’ecosistema amb una societat humana, en el sentit que dins la societat humana es podrien reconèixer diversos oficis que són complementaris, almenys en la funció d’ocupar un lloc, i que persisteixen en els punts de creuament entre els respectius cicles biogeoquímics. Un ecosistema té una organització real, formada per elements discrets, i per tant mai no pot estar en equilibri; a cada element sempre li falta o li sobra quelcom per estar perfectament encaixat, de manera que mai no s’anul·len tensions ni fluxos. Per aquesta raó, perquè sempre mantenen el sistema en tensió, la quantificació i la discontinuïtat dels components d’un ecosistema són factors considerats d’evolució.
La major part del que s’entén per teoria ecològica ha girat entorn de la qüestió de com encaixen o s’organitzen els individus de les diferents espècies per a formar un conjunt funcional (l’ecosistema), i de com la dinàmica del conjunt integra les dinàmiques poblacionals particulars de cada espècie, expressades en el seguit de naixements i de morts que en fan la història. Evidentment, la mort d’individus d’espècies que són víctimes d’altres espècies s’ha de relacionar d’alguna manera, no solament amb el naixement de nous individus de cada espècie presa, sinó també amb els naixements, la disponibihitat i la perillositat dels depredadors, més o menys a l’aguait. Una situació estacionària en la qual tot funcionés com un rellotge perfecte, seria el mític equilibri ecològic, que mai no s’ateny entre altres raons menys substancials (que el món sempre funciona a batzegades), per la profunda raó que el mateix funcionament de l’ecosistema es reflecteix en l’acumulació d’informació nova, o bé per l’establiment del que podríem qualificar de noves condicions asimptòtiques. Mai no falten pertorbacions diverses: el tempteig d’una possible aventura evolutiva, la introducció d’alguna espècie forastera, o alguna catàstrofe imprevisible motivada per agents externs a l’ecosistema (una allau, un canvi de clima) o bé nascuts en l’ecosistema (una espècie que canvia de costums i es fa més agressiva, o arriba a tenir un poder que de tan gran resulta perillós, com passa ara, i a escala mundial, amb l’espècie humana).
Els gremis ecològics
Molts ecòlegs han pogut mantenir un punt de vista que es podria qualificar d’estructuralista, en distribuir els components de l’ecosistema en diferents nínxols ecològics, o bé en gremis, que corresponien a funcions més o menys semblants o definibles, o bé a una adreça, i de tals característiques que es podien reconèixer en tots o en la majoria dels ecosistemes; per exemple, les categories de productors primaris, distribuïts per la seva mida i persistència, en plantes herbàcies anuals, arbres, etc.; els vegetarians de diverses característiques: insectes, brostejadors grossos; els carnívors petits, els carnívors grossos, els paràsits, etc. A cada nínxol ecològic li correspondria una forma peculiar de fer aquell ofici, i l’evolució, responent a la pressió de la selecció natural, s’ocuparia d’optimitzar aquesta funció ideal. La divisió i la subdivisió dels nínxols es podia portar fins on fos convenient.
El concepte de gremi s’ha aplicat generalment en sentit més restringit que el de nínxol. Els escarabats piloters serien un gremi dintre del nínxol dels artròpodes que consumeixen excrements de grans herbívors. Com que cada espècie és feta a la seva manera, reunir-les en nínxols mai no deixa de ser més o menys arbitrari. Probablement és una exageració parlar de nínxols no ocupats i de si aquests podien representar un atractiu per a la colonització o un estímul per a l’evolució que produiria un ocupant adient. Per exemple, s’ha dit que a Austràlia el nínxol representat per artròpodes que consumeixen excrements de grans herbívors era buit. La distinció i la caracterització de nínxols és tan variada i indeterminada, i fa ús de descriptors tan diversos i arbitraris, que es fa difícil, i potser no val la pena, de desenvolupar el concepte més enllà d’un ús poc definit, sense gaires pretensions.
L’organització de les relacions
Com a característiques intensives i quantificables de l’ecosistema, tenim la biomassa, l’energia que circula a través de la biomassa i el temps en què aquesta biomassa es renova. Aquest temps de renovació no és uniforme i es pot expressar en forma d’espectre, cosa que pot fer-se posant en relació la sèrie de valors decreixents de les taxes de renovació (que varien de manera contínua) amb les respectives biomasses implicades. Hi ha parts de la biomassa que es renoven més de pressa i altres que ho fan més lentament: entre l’ATP i la fusta, les taxes de renovació varien com 108. Aquestes diferències no sols existeixen entre les espècies, sinó que dins de cada individu uns compartiments materials canvien la seva matèria més ràpidament que d’altres. Hom pot assignar als ecosistemes enters valors mitjans globals: la selva amazònica es renova molt més lentament que les comunitats que es fan en el llit dels rius i les torrenteres, que només porten aigua de tant en tant. El col·lectiu d’espècies que s’aplega en cada situació reflecteix les condicions generals de perdurabilitat i constància relativa dels respectius entorns.
Sempre es pot reconèixer una organització gradual, històrica, dels ecosistemes. Amb el temps, algunes espècies s’extingeixen, d’altres entren en l’ecosistema o es trien després d’un tempteig variable. Les pressions dins l’ecosistema estimulen la selecció, que en el seu cas portarà a la formació de races noves o, fins i tot, de noves espècies. La selecció i l’evolució no es fan en el buit, sinó en el si dels ecosistemes, que són la màquina i alhora l’escenari de l’evolució.
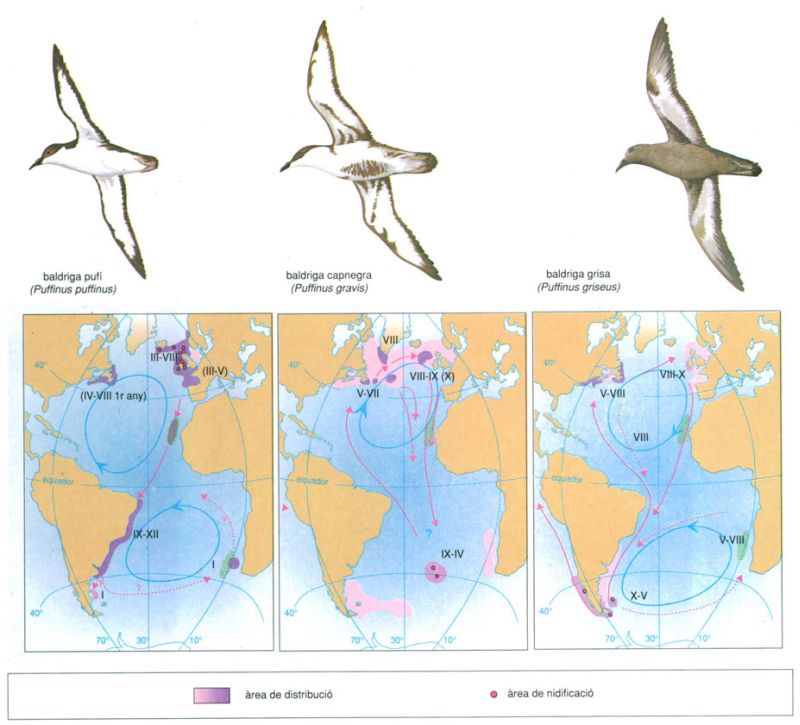
Les migracions de diferents espècies de baldrigues (Puffinus) entre punts de l’Atlàntic molt allunyats entre ells obeeix en part a l’abundància relativa dels peixos (ratllat verd: àrees d’aflorament) que els serveixen d’aliment i en part a la circulació atmostèrica i marina. La quantitat d’informacions que han d’integrar per a ajustar el seu comportament a les combinacions de situacions favorables en una àrea tan dilatada com un oceà que s’estén per tots dos hemisferis sembla enorme amb relació a les dimensions del sistema nerviós d’aquests ocells. [Sagetes blaves: circulacio oceànica; sagetes vermelles: rutes de migració dels Puffinus]. En xifres romanes els mesos de l’any.
Biopunt, a partir de fonts diverses
Moltes espècies evolucionen a cavall d’ecosistemes d’organització ben diferent. Pensem en les nombroses espècies d’insectes que tenen larva aquàtica però que són aeris en la seva forma adulta. La mateixa transmutació, no solament de forma sinó també de la manera d’integrar-se dinàmicament en el món, ocorre en molts altres organismes, des dels batracis a diversos paràsits i animals dels fons marins, les larves dels quals són nedadores. Recordem també els animals migradors, dels quals els més espectaculars i més coneguts per tothom són els ocells. Les seves poblacions dominen extensions molt grans; cada espècie té les seves peculiaritats, però en general s’ajusta a la diferenciació normal dels ecosistemes, en el sentit que cria en entorns que ofereixen una producció accelerada d’aliment: generalment és la primavera nòrdica, o bé àrees costaneres molt productives.
Aquest model és generalment vàlid: sempre els ecosistemes joves són els que ofereixen més recursos a qui va a cercar-los. Tant en el cas dels insectes com en el dels ocells, la necessitat de mudar les plomes o el tegument s’organitza i s’intercala en l’esdevenir de la vida amb solucions admirables. Molts ocells, especialment ocells marins, es mouen sobre grans extensions i és meravellós que un sistema nerviós relativament petit sigui capaç d’organitzar i de combinar la informació procedent de sensors molt variats en una mena de mapa d’un entorn molt extens. El cervell no solament recorda, relaciona i integra, sinó que a més pren decisions i oblida, funcions que no aconsegueix fer tan bé l’anomenada intelligència artificial.
La diversitat i la riquesa
Una part molt considerable de la bellesa del món i de les dificultats de l’ecologia provenen de les complicacions en l’organització i en el funcionament dels ecosistemes. Al llarg d’aquesta obra hi haurà ocasió, sens dubte, d’entretenir-se amb la presentació de detalls admirables de complementació i d’evolució. Aquí solament farem referència a dues característiques quantitatives, que són expressió externa del que podríem anomenar el “compromís social” que imposa la vida en comú: en primer lloc, la riquesa i la varietat de formes que conviuen, és a dir, la diversitat; i en segon lloc, la mena i el grau de completesa de les interaccions que es descobreixen entre les diverses espècies.
Hi ha ecosistemes que són molt rics d’espècies, com la selva amazònica o els baixos de corall; d’altres són més pobres, com la taigà o el poblament d’unes dunes. Són factors determinants d’aquesta riquesa tant la persistència i la variabilitat de l’ambient, com les condicions passades d’evolució, que han pogut fornir una reserva d’espècies més o menys rica. La diversitat té a veure amb el nombre total d’espècies, però no és només això, sinó que es relaciona també amb les proporcions entre el nombre d’individus de les espècies presents (o les seves respectives biomasses, encara que, segons aquesta mesura, l’ordre d’abundàncies pot resultar diferent). En general, per a Ni individus d’una espècie x, en trobarem un de l’espècie y, i per a Njindividus de l’espècie y, en trobarem un de la z, i en ecosistemes rics d’espècies, els valors de les N tendeixen a ser col·lectivament i en mitjana més baixos que en els ecosistemes de baixa diversitat, on el nombre d’individus d’espècies successives, ordenades per la seva importància, cauen ràpidament en passar de la més dominant a les altres.
És clar que mai ningú no ha pogut estudiar exhaustivament un ecosistema fins arribar a donar una llista quantificada i raonablement completa de totes les espècies. Sempre cal esperar trobar espècies no descrites i ara es parla sovint del risc (i més que del risc, de la certesa) que molts ecosistemes desapareixeran abans que s’arribi a tenir un coneixement raonablement acceptable de les espècies que els integren. Generalment es parla de la diversitat de grups taxonòmics limitats: diversitat d’espècies arbòries, diversitat d’espècies de papallones, etc. I sovint els valors numèrics d’aquestes diversitats estan correlacionats positivament, cosa que, fins a un cert punt, és natural, ja que les condicions d’evolució (i d’estructuració dels ecosistemes) han operat simultàniament sobre els diversos grups i han portat a un grau semblant de diversificació o d’acceptació d’immigrants.
La diversitat es pot expressar de manera convenient pel nombre d’espècies presents en una sèrie d’individus del grup corresponent agafats a l’atzar; per exemple, el nombre d’espècies que es poden comptar en 1 000 exemplars de papallones nocturnes agafades a la llum. Un altre índex força usat es basa en la distribució dels valors de probablitat pi (nombre d’individus de l’espècie i partit pel nombre d’individus total), i té la forma H = - ∑i=1 pi log2 pi. És l’expressió de Shannon-Weaver, que dóna la informació mitjana per símbol, i s’aplica a qualsevol conjunt de símbols (missatges, llenguatges). La diversitat es pot interpretar com un missatge de la naturalesa.
Una aproximació menys descriptiva i més dinàmica relaciona la diversitat amb els processos que la generen. Si es tracta d’una cursa de velocitat per omplir un espai, o d’aprofitar unes condicions excepcionals (com en un cultiu de flux), la diversitat es fa mínima; si es tracta d’un sistema menys dinàmic, sempre és més probable poder afegir-hi alguna espècie, i la diversitat creix. El valor màxim es donaria en una vitrina de museu, on cada espècie és representada per un individu; és un sistema mort, és clar. Un exemple proper n’és la mítica arca de Noè; molts dibuixants que han caricaturitzat l’entrada i la sortida dels animals no deixen de fer remarcar la poca probabilitat de la constància de nombres molt baixos en els sistemes dinàmics, i sempre hi fan sortir més conills i més rates que la parella ingressada, parella que és més probable que es conservi com a tal en el cas d’animals grossos i de vida llarga. En resum, si és més fàcil afegir un individu d’una espècie ja present que un d’una espècie nova, la diversitat baixa; si és més fàcil que la diferenciació genètica vagi endavant, és a dir, si és més fàcil afegir una nova espècie que no pas afegir un individu, la diversitat augmenta. L’home fa disminuir ineluctablement la diversitat de tots els ecosistemes, i és ben poc probable que aquesta tendència canviï de signe mentre la humanitat continuï fent pressió per accelerar la renovació de tota la resta de la biosfera (i alentir la renovació de la biomassa pròpia). La diversitat crescuda és fruit de qualsevol període d’evolució tranquil·la i continuada.
Espectres de diversitat
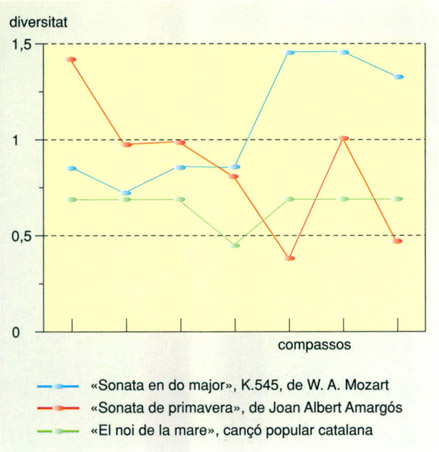
Editrònica, a partir de dades d’Andreu Roca
Una peça musical, com qualsevol conjunt de símbols, es pot interpretar en termes de diversitat. El gràfic mostra la comparació dels espectres de diversitat de la part melòdica principal dels set primers compassos de tres peces molt diferents: la “Sonata en Do major K. 545”, de W.A. Mozart (de la qual s’ha pres com a part melòdica principal la mà dreta del piano), la “Sonata de primavera” del compositor català contemporani Joan Albert Amargós (la flauta de bec contralt que inicia en solo la peça), i la cançó de bressol tradicional catalana “El noi de la mare” (el cant). S’ha pres com a símbol la nota musical, i el nombre de “presències” s’ha mesurat en temps, prenent com a unitat de presència la durada de la nota més curta que apareix en cada compàs que es considera. La diversitat s’ha calculat amb la fórmula de Brillouin. Es pot observar la diversitat baixa i l’espectre pràcticament pla d’“El noi de la mare”, expressió de la monotonia que requereix una cançó de bressol. L’elevada diversitat de la peça de Mozart a partir del cinquè compàs correspon a l’aparició d’escales de semicorxeres per graus conjunts que ornamenten els components melòdics de la peça. L’espectre de diversitat de la peça d’Amargós varia de manera més irregular.
La interacció entre les espècies
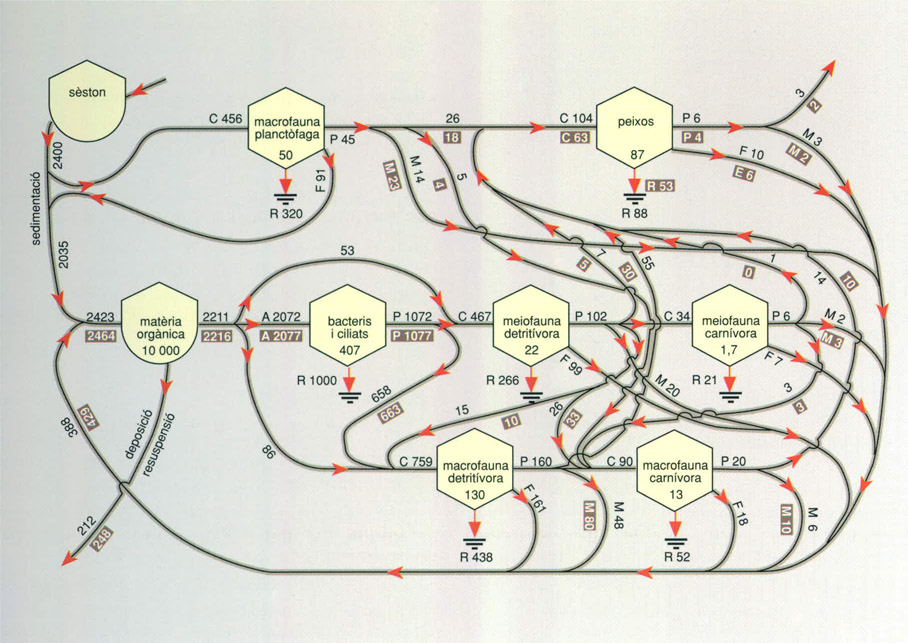
Les representacions ideals dels ecosistemes, imaginades com una base per construir models predictius, evidentment resulten molt complicades. Així i tot són un reflex molt limitat de la rica varietat de matisos que trobem en les interaccions entre uns i altres éssers vius als ecosistemes reals.
Editrònica, a partir de diverses fonts
Si hi ha poques espècies, és natural que entre elles s’estableixi una interacció forçosa. Un ecosistema format per una espècie de planta, per un animal i per algun fong o bacteri s’ha d’unir per necessitat en un circuit d’interacció. En augmentar la diversitat i haver-hi diferents espècies que fan el mateix ofici (que pertanyen al mateix nínxol, diria algú), la saturació de les connexions possibles no és tan òbvia. Cal precisar què entenem per interaccions i quines són les que compten: ens podríem limitar a veure-hi una transferència d’energia, associada a un circuit de retroalimentació que porta a una certa regulació. L’efecte de la competició (es una relació, tanmateix?) porta a tallar les relacions directes entre els competidors. Per això, si de cada relació triangular possible entre espècies se n’interromp una, com és probable que passi, ens quedem amb una connectivitat que mai no és completa o saturada, cosa que està d’acord amb el manteniment d’un grau de flexibilitat interna en els ecosistemes i en els sistemes en general, i molt notablement en els sistemes nerviosos, on la connectivitat parcial genera la jerarquia i és la clau de la seva funcionalitat. Això va en contra de nocions molt corrents, com aquella que diu que en l’ecosistema tot està unit amb tot i que n’hi ha prou que traguem la floreta més insignificant perquè tot es pugui descompondre, com un fràgil castell de nínxols ecològics.
L’ecòleg de butaca, que desitja fer models quantitatius dels ecosistemes, es veu forçat a definir on accepta enllaços i on no, i de quina mena han de ser. El naturalista de camp s’adona de la dificultat d’atrapar en models formals la rica varietat de matisos que trobem en les interaccions entre uns i altres éssers vius. Constatar aquest fet no va en detriment d’uns o d’altres: hem d’arribar a harmonitzar la percepció de la riquesa i la bellesa de la naturalesa amb el reconeixement que han d’haver-hi camins per a la seva exploració racional.
Ja hem fet referència repetidament a les relacions entre espècies lligades per una transferència d’energia. Altres relacions secundàries, encara que no menys importants, són el subministrament de vitamines, o el suport mecànic que donen unes espècies a les altres, com el que donen els arbres a les lianes, o a la munió d’epífits, com bromeliàcies i falgueres, i altres organismes menors que viuen en estricta dependència d’aquestes espècies subordinades de plantes. Els fagòtrofs, que requereixen per a la seva alimentació una percepció i una persecució actives (animals “de forquilla” especialment) són llençats a una vida més activa, i això ha representat un estímul molt poderós per a l’evolució: un perfeccionament dels òrgans dels sentits i del sistema nerviós n’ha estat la conseqüència evolutiva. El sistema nervios és fet per a captar molta informació de l’exterior, i per a dur a terme una presa de decisions i una acció immediates. Manipula informació i, com era d’esperar, s’ha projectat enfora en tota mena d’artefactes i organitzacions, com els desenvolupats per la humanitat. El sistema nerviós és, com el mateix ecosistema, un sistema jeràrquic, en el sentit que la seva connectivitat interna és parcial: cadascuna de les neurones no està connectada amb totes les altres.
Els animals representen l’esperit imprevisible dels ecosistemes. És clar que, per raons òbvies, la seva biomassa no pot representar una fracció molt gran de tot l’ecosistema, i la seva diversificació, més gran que la del component vegetal, prova una dinàmica suplementària injectada en la seva evolució. On hi ha més acció és entre espècies animals que interactuen intensament, com a depredadors molt actius, més que no pas en animals que s’alimenten de manera passiva.
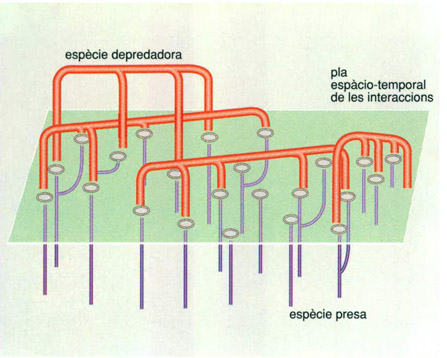
Les interaccions entre els individus de dues poblacions que comparteixen un espai, una d’una espècie depredadora (vermell) i l’altra d’una espècie presa (morat), és sempre asimètrica des del punt de vista de la termodinàmica i de la informació. Per les pèrdues que es produeixen en el pas d’un nivell tròfic a un altre, de l’energia que obté la presa pel seu aliment, només una part passa al depredador. Des del punt de vista de la informació, cada predador interacciona amb més d’una presa, mentre que, per als individus de l’espècie presa, la primera interacció també és sovint la darrera; per això la selecció natural del predador afavoreix la seva capacitat d’aprenentatge mentre que la de la presa afavoreix característiques de mimetisme o cripsi o bons reflexos de fugida.
Editrònica, dades elaborades per l’autor
Pensem ara en les interaccions entre els individus de dues espècies molt actives i imaginem un pla comú, estès en el temps i l’espai, en el qual se situen els episodis d’interacció. Un episodi pot ser que un individu d’una espècie capturi i devori un individu de l’altra espècie. El mostratge que fan del conjunt d’esdeveniments els individus que representen les respectives espècies és diferent. Suposem que es tracti d’una espècie d’insecte i d’una espècie d’ocell. Cada interacció amb molta probabilitat acabarà amb la vida de l’insecte, mentre que per a l’ocell serà un episodi més. Com ha estat dit, “el conill corre per la seva pell, la guilla solament pel seu sopar”. Cal esperar que la selecció natural porti l’insecte a manifestar característiques físiques d’efectivitat immediata (com pot ser un mal sabor associat a color advertidor, una coberta dura i espinosa, etc.) o que tendeixin a dissimular la seva presència, mentre que, per a l’ocell, la successió d’episodis de cacera té un sentit especial, ja que es poden col·locar dins d’un marc de memòria i experiència; això requereix un sistema nerviós força complex, capaç d’aprendre, o d’aprendre a aprendre.
Aquest tipus d’interacció no s’ha d’associar exclusivament amb el fet de ser caçador, ni tan sols amb el menjar, i ha tingut una gran importància en la nostra evolució. Expressa el valor positiu que té, en la selecció natural, el domini d’un segment més ampli de temps i d’espai. Tractant d’extrapolar el significat general d’aquesta dialèctica de l’evolució, resulta temptador suposar que en qualsevol situació en què s’hagués donat la vida en unitats discontínues i en condicions semblants a les que trobem a la Terra, hauria estat fatal l’aparició d’humanoides, capaços d’aprendre a aprendre a aprendre. Es comprèn, d’altra banda, que en la naturalesa no hi pot haver un nombre gaire elevat d’animals grans i ferotges.
El contracte social no exclou les grans injustícies, i els tipus d’interconnexions que descobrim en el funcionament dels ecosistemes es poden qualificar de moltes maneres. Moltes d’aquestes interconnexions no troben una analogia fàcil en la societat humana.
L’organització social
Fetes les consideracions precedents, ens adonem que un estil comparable de construcció i d’organització de l’espai en benefici propi es troba també en el que podríem considerar altres tantes cimeres de l’evolució: l’arbre de bosc n’és un exemple. Un altre cas ben evident és el dels coralls edificadors d’esculls. Els abundants dipòsits de calcària que produeixen (creixen uns quants mil·límetres l’any) configuren l’entorn i permeten que els teixits animals se situin en posició apropiada respecte de la llum i dels corrents marins. Els coralls viuen en simbiosi amb poblacions d’algues d’una gran simplificació genètica (una sola espècie es troba en els habitants més diversos dels baixos de corall) i la configuració de la part sòlida no solament apropa les algues a la superfície de la mar, és a dir, a la llum, sinó que també pot fer que es creïn formes de circulació vertical de l’aigua en la proximitat dels esculls, cosa que afavoreix l’arribada de materials nutritius provinents del fons.
La tercera cimera on l’evolució mostra un gran enginy en l’organització de l’espai és en els insectes socials i en l’home. Formiguers, termiters, les pistes adjacents, com també les ciutats i les vies de comunicació dels humans, són estructures exosomàtiques d’una gran importancia perquè canalitzen l’energia i faciliten per vies molt diverses la supervivència de l’espècie dominant. Els insectes socials són molt importants quantitativament arreu. Segons dades recents, a la selva amazònica cada hectàrea de sòl conté més de 8 milions de formigues i un milió de tèrmits. Aquests dos grups d’insectes, juntament amb alguns himenòpters com abelles i vespes, componen més del 75% de la biomassa total dels insectes. Hom esmenta xifres comparables en d’altres ambients. Dels humans no cal ni parlar-ne, i només farem notar que, igual que en el cas dels coralls, han simplificat enormement la varietat genètica del conjunt d’organismes dels quals treuen profit, tot fent-los multiplicar sota la seva “protecció”. Podem comparar les plantes de conreu, el nombre d’espècies de les quals representa una fracció molt petita de la flora natural, amb els simbionts dels coralls, que són dinoflagel·lats. També en el cas de l’home és palès el que hom assenyalava en els arbres i els coralls: les estructures molt extenses no són iguals, i en tot cas, tenen un sentit adaptatiu que fa que puguin usar de manera eficient l’energia externa o exosomàtica. Podríem dir que hi ha una part material, una estructura que serveix alhora de suport a fluxos organitzatius d’energia i que tant si és d’origen directament biològic com si ho és indirectament (a través d’una tecnologia), és de tal naturalesa que sempre es pot avaluar en termes d’informació.
La memòria i els canvis
La història que hem contat fins ara és una història de continuïtat i previsibilitat: més aviat portaria a un desenvolupament pausat sense entrebancs. Però prou sabem que ni la història humana no és així. Tota la història ha estat un seguit d’ensurts i intents de superar desgràcies. Moltes d’elles, és cert, nascudes en l’interior de la mateixa societat humana, com les guerres, i no solament les guerres. Altres, d’origen extern, incideixen una i altra vegada, sense que hi hagi responsables a qui assenyalar. Les més freqüents, com els canvis diaris i anuals, s’han assimilat dins els mateixos organismes, en forma de ritmes interns, que anticipen els canvis i predisposen l’organisme a rebre’ls positivament i a reaccionar de manera adient, tot amb relació a la duració mitjana de la vida de l’organisme. Altres canvis excedeixen la possibilitat de la memòria dels individus i tenen efectes sovint destructors, que s’estenen a segments més o menys grans de l’ecosistema.
Els canvis que colpeixen impensadament, no assimilats per la vida i que, per tant, es consideren com a catàstrofes, no han impedit que el que podem anomenar memòria evolutiva es projecti cap al futur. L’evolució ha superat potser totes les grans catàstrofes, a costa de moltes extincions, però rarament s’ha perdut alguna adquisició important de la vida, com els sistemes nerviosos complicats o la termoregulació. I potser cada vegada ha estat més planer, superades les pertorbacions d’intensitat relativament baixa, augmentar les cotes que defineixen la catàstrofe que no té remei. Actualment, l’espècie humana pot subsistir amb una quantitat d’individus considerable i amb una civilització acceptable en unes condicions que haurien estat inhòspites per a alguns dels nostres avantpassats. Ens preocupem pel clima, però fins i tot el clima ha deixat de ser un obstacle en un món dotat d’un sistema de transport i de comunicacions fortament unificador; que això passi dins d’una determinada civilització i ajudi a reforçar les diferències entre aquesta i altres grups humans menys tecnificats, ja és una altra qüestió.
La successió i les pertorbacions
A mitjan segle XIX s’expressà ben clarament una idea fonamental en ecologia: la idea de successió. La natura viva, tal com l’apreciaven els geògrafs físics i es manifestava en la coberta vegetal i en les característiques del sòl que s’hi associava, estava subjecta a un canvi històric, identificable per aspectes externs tocants a la vegetació o a les comunitats. Es fàcil retrobar antecedents d’aquesta visió almenys des d’un segle abans.
El tema de la successió ecològica s’introdueix molt fàcilment mitjançant uns quants exemples. Si s’abandona el conreu seguit d’un camp, a poc a poc i espontàniament, el camp recupera el tipus de vegetació que l’ocupava abans, la mateixa que persisteix en àrees semblants i no conreades. És un procés que demana uns quants anys, i encara més si es tracta d’arribar a reconstruir el tipus de bosc que, suposem, representa la vegetació final o estable del país. D’aquesta vegetació terminal se’n deia i se’n diu la vegetació clímax.
Aquesta paraula, clímax, ha generat una certa discòrdia: d’una banda suggeria que hi havia un camí prèviament assenyalat, com si la successió ecològica es pogués comparar al creixement d’un organisme o, almenys, al desenvolupament d’una organització; ja que, evidentment no és així, la clímax es desqualificà com a simple utopia o fantasia d’alguns ecòlegs seguidors d’una escola potser massa entusiasta. Sovint aquesta crítica s’ha intentat estendre al mateix concepte de successió. Es comprèn que sovint sigui difícil fer evidents les diferències entre dues etapes successives en un procés de canvi. Els mètodes estadístics habituals, i tal com s’acostumen a utilitzar, no fan indubtables les diferències entre etapes immediates; però hi ha una diferència considerable entre el principi i la fi. L’anàlisi instant per instant ens podria fer dubtar de si Aquil·les arribarà mai a atènyer la tortuga, però pocs dubtem del resultat final de la cursa en la paradoxa de Zenó, que és paradigmàtica de la manera com de vegades s’aplica l’estadística als temes d’ecologia.
Acceptem, doncs, el fenomen de la successió amb algunes regularitats generals. La primera és que la quantitat de clorofil·la, o la capacitat de síntesi orgànica, augmenta poc al llarg del camí, o molt menys que la quantitat total de material, i especialment de fusta. La conseqüència és que el quocient producció/biomassa (i més encara el quocient producció/[biomassa + necromassa]) disminueix al llarg de la successió. Acostuma a augmentar el nombre d’espècies diferents i també, generalment, el nombre de nivells tròfics en què es disposen: és a dir, les cadenes alimentàries esdevenen més llargues. Si no és amb els ulls del pagès que abans conreava el camp, l’organització final es pot veure com a més bonica o, almenys, més complexa.
Continuant amb l’exemple del camp, cal no oblidar que mentre dura el conreu, la successió no aconseguia avançar perquè cada anyada es retirava la collita, el material produït. Basta deixarlo perquè la natura trobi la manera d’assimilar aquell material i d’organitzar-se, i complicar-se la vida. Resulta planer descobrir algunes adaptacions comunes en les plantes que corresponen a cada una de les etapes o estadis de la successió: per exemple, les primeres invasores ho són sovint perquè produeixen un nombre elevadíssim de llavors o fruitets que el vent s’emporta, mentre que en etapes més avançades de la successió és corrent que les plantes siguin dispersades per fruits carnosos i atractius per als ocells; en aquest moment, relativament tardà, tota la comunitat biòtica ha organitzat conjuntament l’ampliació de la seva àrea.
Un altre exemple de successió es pot trobar en la vegetació d’una tartera, en la qual entren en joc els components més humils de la flora: molses i algues, i alguns animalets petits del sòl incipient. La successió és un fenomen complex, perquè va acompanyat també de la diferenciació i l’organització del sòl i de la vida que conté. En una tartera la successió es fa difícil; té poc temps per a avançar, perquè successius esllavissaments de pedruscall l’obliguen a recomençar una i altra vegada. Ens adonem també de la importància que té l’aptitud de les plantes i de les seves diàspores per a mantenir l’ocupació d’un espai. La capacitat espontània de la natura per a emportar-se en cada allau el material biòlogic produït pot ser semblant a l’efecte de l’home en retirar les collites d’un camp. La successió de les tarteres avança poc perquè es tracta d’ambients sotmesos a una explotació (natural, això sí) molt intensa.
Més llarg és el temps (dècades o segles) que una comunitat vegetal ha hagut de persistir en condicions relativament favorables per arribar a tenir la forma de bosc. Estudiosos de la vegetació han distingit tipus biològics en els vegetals, que no són més que els noms d’herba, arbust, arbre, etc., dignificats per mitjà del recurs de convertir-los en una nomenclatura que sona a grec, com no podia ser altrament: un megafaneròfit és evidentment un arbre, i un hemicriptòfit un gram (les gemmes perdurants queden arran del sòl durant dos o més anys). La successió representa una reorganització constant amb introducció de plantes de més alçada, i alhora d’una varietat creixent de tota mena d’organismes dependents, entre ells moltes espècies d’animals.
Un bosc es pot cremar de tant en tant, i això, després de la combustió de molta fusta, representa retornar a una fase equivalent a una que ja havia estat superada. La successió té algunes característiques que fan pensar en el joc de l’oca, o d’altres jocs semblants: un va avançant regularment fins que, en un moment de mala sort, es veu obligat a tornar a un estat equivalent a un que fa temps ja havia estat superat. Anotem la asimetria profunda entre l’incendi del bosc i la seva regeneració: la regeneració serà forçosament molt més lenta que la crema.
Hi ha una incertesa existencial associada amb tot el que anomenem pertorbació. L’única regularitat que s’hi pot reconèixer és que les pertorbacions més enèrgiques, és a dir, amb més energia exosomàtica associada, són menys freqüents que les més suaus. També la nostra vida comporta moltes ensopegades de menor importància i, de tant en tant, algunes garrotades fortes, la principal de les quals, certament, és única per definició. La biosfera funciona de manera semblant, podríem dir, a cops de peu. Així, doncs, la successió es podria veure, simplement, com un mecanisme de restauració o cicatrització de les ferides. Ens podem preguntar si perquè l’evolució biològica es produeixi no caldrà també que, de temps en temps, tingui lloc la necessària esporgada.
La successió en un territori marginal ocupat fonamentalment per boscos, màquies i garrigues mediterrànies prop de Sant Martinde-Londres, al N de Montpeller, al Baix Llenguadoc, mitjançant l’anàlisi dels canvis enregistrats en un seguit de punts seleccionats sobre el mapa en sèries de fotografies aèries separades per un període d’uns vuit anys, es pot observar la tendència majoritària a l’embosquinament de conreus abandonats i vegetació herbàcia, mentre que els canvis entre tipus de vegetació més complexos són molt més lents. S’hi observa també el retrocés de fases avançades de la successió cap a altres menys avançades, probable conseqüència dels incendis repetits, freqüents a la regió mediterrània.
Biopunt, extret de Rambal & Lepart, 1987
En els boscos de la zona climàtica temperada, els arbres que perden la fulla sofreixen anualment una interrupció de la vegetació, la qual cosa es pot considerar com l’equivalent d’un incendi menor (o d’una pertorbació menor, si no volem ser tan específics). La vegetació s’atura, però a diferència del que passa amb les plantes baixes, els arbres tenen l’avantatge que resten preparats, amb els borrons a punt, per a la primavera següent. En aquests boscos, el repòs hivernal permet també una retenció fàcil d’elements nutritius en el sòl. Aquesta és una qualitat dels sòls que els fa bons per a l’agricultura, qualitat que s’ha desenvolupat de manera particularment pròspera en entorns fluctuants, sigui en la regió temperada, o, en la zona intertropical, en la muntanya (conreu de la patata), o en aiguamolls (conreu de l’arròs).
En els boscos tropicals, o en boscos temperats amb aigua a disposició tot l’any i sense gelades, la vegetació esdevé més alta i sovint molt complexa. Podríem dir que, per exemple, les clímaxs de l’Amazònia són més clímax que les de la regió temperada, cosa que significa, senzillament, canviar aquella noció històrica, impressionista i estètica, per un concepte molt més quantificable, el quocient P/B (o P/[B+N]), com a expressió d’una taxa de renovació (P producció, B biomassa, N necromassa). La successió es veu com la realització d’una tendència a anar més a poc a poc. Ajuda a comprendre-ho el fet de pensar que la producció depèn de la superfície, tant per a la disponibilitat d’energia lluminosa com d’aigua del cel, mentre que l’acumulació de biomassa (i, no ho oblidem, de necromassa, que també serveix, almenys, per a donar estructura) no queda limitada de la mateixa manera. L’exemple dels boscos tropicals és particularment alliçonador, perquè mostra directament l’enriquiment d’espècies i formes de vida, amb lianes, diversos tipus d’epífits i variats animals dependents dels materials morts i dipositats sobre les branques, que fan el paper dels sòls.
La noció de successió s’aplica a tots els ecosistemes. En el desenvolupament del plàncton d’aigües dolces i salades s’hi reconeixen els mateixos trets característics, si bé les escales, i notòriament l’escala del temps, estan canviades. Normalment, la matèria assimilada pel plàncton vegetal es mou de dalt a baix, sigui per sedimentació directa o perquè el plàncton és menjat pels animals que, per sedimentació pròpia i dels seus excrements o per moviments actiu, contribueixen al mateix transport. El resultat és que a dalt es van consumint els elements químics més necessaris per a la vida. Gairebé sempre la cadena es trenca per on és més feble, és a dir pels elements més limitants, o que s’acaben més aviat, que són el fòsfor i el nitrogen, com també passa en terra. Aquells elements químics els retrobem concentrats en aigües profundes, on, a conseqüència de la falta de llum, es poden acumular perquè no hi són assimilats. Quan es destrueix l’estratificació tèrmica, generalment per l’arribada del temps fred i ventós, es barreja verticalment l’aigua i d’aquesta manera es fertilitza la superfície i s’inicia una nova successió que, com és natural, comença amb un elevat índex producció/biomassa. Per això la barreja vertical s’ha pogut comparar (encara que d’una manera no totalment correcta) amb la llaurada d’un camp.
En el plàncton el nombre d’espècies es multiplica progressivament i es desenvolupa una successió que va molt més de pressa que en els sistemes terrestres. Els productors primaris del plàncton no arriben a acumular fusta, cosa que no vol dir que no ho intentin, en el sentit que cobertes, closques i tota mena d’ornaments són més freqüents en les espècies que vénen després que no pas en les que engeguen l’inici de la successió. El joc de successions més o menys truncades se succeeix en l’aigua no solament d’una manera més ràpida que en els sistemes terrestres, sinó també més irregularment. Però el resultat és semblant: augmentar la complexitat de la vida present i frenar-se a ella mateixa per aquest principi tan general de separar els reactants potencials (on hi ha llum no hi queden nutrients, i viceversa). Dues dites populars són ben aplicables al mecanisme de la successió ecològica: “embolica, que fa fort” descriu la complexitat històrica de la natura local, així que la deixem tranquil·la, i “Déu dóna faves a qui no té queixals” expressa la necessitat de tenir separats els reactants (les faves dels queixals) per a crear i mantenir la complexitat de la natura.
Un altre exemple de successió és el que s’observa en el llit d’un riu que va descrivint meandres amb una dinàmica pròpia del riu i de les seves ribes. En aquells punts de la riba on el riu es fa enrere, la vegetació terrestre avança o s’hi desenvolupa, i ho fa disposant-se en faixes paral·leles a la vora del riu, de manera que les etapes més avançades en queden més allunyades. Atesa la geometria de la formació dels meandres, el riu, en avançar, talla quasi perpendicularment les faixes que representen etapes successives de successions anteriors. Aquest exemple és interessant per diverses raons.
El procés descrit evita que els grans boscos tropicals quedin massa temps sense renovar-se. Si interactuen amb un riu de llit mòbil, evidentment es produeix una renovació continuada de la floresta, la qual tanmateix afavoreix l’evolució de les espècies dins del seu àmbit. Aquest exemple permet veure que allò que caracteritza la successió no és una seqüència rigorosament lineal ni la visió de la clímax com a culminació final, sinó simplement la manca de simetria de qualsevol taula o matriu que expressi la distribució de les probabilitats de transició entre els diferents estadis o etapes que haguem distingit en tota la seqüència. No necessitem recórrer a cap problemàtic concepte de clímax. És tan senzill com si diem que en un any cadascun de nosaltres pot afegir un any a la seva vida, o bé morir-se; el que ningú no pot fer, però, és afegir-se un parell o més d’anys. Amb aquesta noció desapareixen molts falsos problemes i bastants discussions suscitades entorn del concepte de successió.
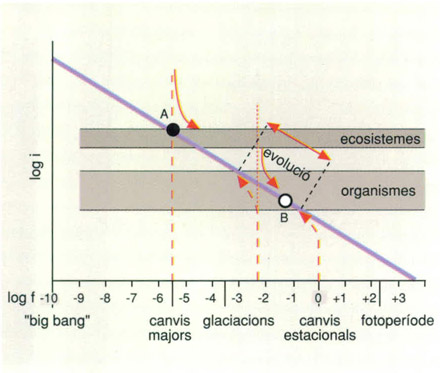
Podem traçar un catàleg de les pertorbacions conegudes que sabem que afecten determinats sistemes naturals. Veurem que apareix una relació inversa entre la freqüència d’un tipus de pertorbació i la seva violència. Aquesta intensitat pot relacionar-se amb l’energia (exosomàtica) implicada, i la freqüència s’estén entre l’esdeveniment únic, com fou el Big Bang o traca inaugural de l’Univers, fa uns 1010 anys, i allò que és tan seguit com el tic-tac del rellotge. L’escala de freqüències pot anar en logaritmes decimals del nombre d’esdeveniments per any, començant per -10.
En aquesta escala de freqüències la pertorbació anual (més o menys intensa segons les latituds) ocuparia el lloc 0, les glaciacions quedarien entre el -4 i el -6, les guerres més grans entre -2 i -1, i els canvis associats amb el dia i la nit entre el +2 i el +3.
Aquesta distribució de les pertorbacions té una importància particular com a rerefons de tots els processos de successió, és a dir, de substitució d’un sistema per uns altres al llarg del temps. Pertorbació suggereix imprevisibilitat, malgrat que assignem, de manera provisòria, una freqüència a cada tipus de pertorbació, que no vol dir gran cosa. La seva indeterminació bàsica ha estat la desesperació del ecòlegs des de sempre. Sovint s’ha pensat en cicles astronòmics, més o menys ben definits, que poden valer per al dia i per a l’any, però que més enllà d’aquests períodes tenen un valor explicatiu més dubtós. Molts fenòmens naturals d’intensitat fluctuant s’han intentat descriure per l’addició de sèries harmòniques de diferent període; però es fa difícil avaluar llur possible interacció. En els sistemes materials les ocil·lacions mai no són simètriques, és a dir, no hi ha cap fenomen (excepte les oscil·lacions usades per a descriure la llum) en el qual la baixada sigui simètrica a la pujada.
Amb relació a consideracions pràctiques i a la predicció, cal recordar que una determinada freqüència global pot permetre una distribució molt irregular en el temps dels diversos episodis. Per cridar l’atenció sobre la corresponent pèrdua de previsibilitat s’ha parlat, entre d’altres, dels efectes Noè i Josep. L’efecte Noè és aquell esdeveniment pràcticament únic, com un diluvi, o, tal vegada, la col·lisió de la Terra amb un planetoide. L’efecte Josep fa referència a la tendència que tenen alguns tipus de pertorbacions que en alguns períodes són rares o absents, a presentar-se agrupades en el temps, com passa amb els anys de pluja i els de sequera. Les construccions hidràuliques es fan per a resistir les condicions de l’aiguat del segle, però no poden tenir en consideració un esdeveniment únic de caràcter diluvial. Tot això reconeix les cartes amagades que pot jugar un món obert, que diria Popper, però deixa en evidència l’ecòleg quan es veu empès a fer prediccions: no pot dir, òbviament, que la bola de vidre li presenta un espectre de pertorbacions amb unes característiques tals que fan prudent no prendre el risc de fer de profeta. Josep, el d’Egipte, ho tingué més fàcil.
Relació entre intensitat i regularitat de les pertorbacions
Biopunt, extret de Rambal & Lepart, 1987
Una de les poques regularitats formulables respecte a les pertorbacions és que les més enèrgiques són menys freqüents que les més suaus. Aquesta regularitat es pot expressar com una relació inversa entre els logaritmes de les freqüències i els de les intensitats (mesurades per la magnitud de l'energia implicada). La pertorbació màxima, el Big Bang que inicià l'expansió de l'Univers, la veiem com un esdeveniment únic a l'escala de les desenes de milers de milions d'anys. Pertorbacions majors, com les vinculades a col·lisions de la Terra amb planetoides, són esdeveniments força més freqüents, de prop de desenes de milions d'anys. Les glaciacions són fenòmens amb una freqüència de vora desenes de milers d'anys. Les estacions de l'any es repeteixen amb una regularitat anual i la successió de dies i nits (excepte a les latituds polars) es repeteixen amb un ritme diari. La vida ha anat assimilant o interioritzant necessàriament aquells canvis que eren més freqüents i aparentment inevitables; l'evolució condueix a superar les restriccions de l'entorn i les seves fluctuacions, encara que la possibilitat d'impactes imprevistos i no assimilables es mantingui oberta. A una escala temporal més àmplia, són els ecosistemes els que interioritzen les pertorbacions de menor freqüència com ara les glaciacions o els movimetns de les plaques litosfèriques.
La successió, matriu històrica
La successió proporciona un punt de vista històric útil per a posar ordre en la imatge o el model que ens puguem formar del món, és a dir, la història, o alguns dels seus components, com la vida, els organismes i els ecosistemes, en els quals ens podem sentir particularment interessats. Val la pena examinar les relacions entre successió i evolució i fins i tot explorar les relacions entre aquest entreteixit històric i alguns aspectes de com veiem actualment la problemàtica associada al desenvolupament de la nostra civilització.
Quan es tracta de situar l’evolució en un món subjecte tot ell a un procés ininterromput de successió, hom troba relacions força directes entre tots dos conceptes. La teoria de la selecció natural estableix simplement que, en alguns casos, el genotip d’alguns dels descendents anirà suplantant el genotip més generalitzat entre els avantpassats. Quan es vol relacionar aquesta afirmació amb alguna definició de l’adequació (“fitness”), es tenen dificultats lògiques perquè no es poden caracteritzar independentment adequació i supervivència.
Seria desitjable trobar algun criteri que fos independent dels mecanismes de substitució en el temps, i que permetés de sistematitzar les diferències entre l’antecedent i el conseqüent. La teoria de l’evolució per selecció natural demana aquesta validació d’una manera externa i independent, cosa que es podria trobar tal vegada en la termodinàmica. Podríem acceptar que una sèrie de reaccions químiques, equiparables amb un augment de l’entropia, ocorregudes en el cos dels organismes d’una espècie i en l’espai immediatament perifèric en un temps donat, es podria retrobar i reconèixer com un augment d’informació en qualsevol moment posterior. Aquesta llaçada sobre el temps seria l’essència de l’evolució.
De manera inspirada per les circumstàncies socials i econòmiques, a l’estudi de les quals la idea d’evolució biològica deu probablement el seu origen, la cosa es podria formular en els termes següents. Una informació augmentada estaria en condicions d’assimilar una quantitat més gran de futurs estats probables de l’entorn i, per tant, de permetre disminuir la producció de nous individus en el tempteig incessant per ocupar una fracció més gran de l’espai disponible. La col·locació d’una quantitat més elevada d’informació comporta, encara que no necessàriament, un augment unidireccional i creixent de la quantitat d’informació que la nissaga passa del passat al futur i, molt probablement, també un augment del temps de funcionament de cada sistema individual. Algunes d’aquestes tendències en el canvi es retroben en l’estudi de la successió, cosa que prova l’existència d’una relació comuna amb principis termodinàmics o físics molt generals, els quals serien especialment preciosos com a criteris de predicció, ja que pràcticament no n’hi ha d’altres.
No sembla gaire fructífer tractar d’esgotar les possibles analogies entre un ecosistema i un organisme, però tenen propietats comunes per raó del caràcter historic que tenen tots dos: un canvi de l’augment d’entropia avui per adquirir una informació que pot facilitar la persistència demà. És en aquest sentit que la successió trobaria una explicació o una col·locació no forçada dins d’un ordre d’idees semblant.
Es pot cercar una altra connexió operativa més estreta entre successió i evolució. És possible reconèixer tal vegada certes característiques comunes entre les etapes que se segueixen en la successió, manifestes en les respectives espècies presents, i els aspectes o característiques d’individus de la mateixa espècie, quan l’àmbit de vida d’aquesta espècie s’estén sobre diverses etapes successionals. Amb relació a les formes biològiques precedents, les espècies que vénen després generalment són més corpulentes, viuen més temps, deixen menys descendents i els protegeixen millor, tenen un metabolisme més baix, etc. Ara bé, l’escenari de tota la biosfera segueix lentament el sentit general de la successió, encara que, aquí i allà, hi ha recaigudes sobtades cap a estadis que ja havien estat ultrapassats.
Qué vol dir això? Doncs que la biosfera en evolució és com si fos arrossegada damunt d’un joc d’escales mecàniques, subjecta a un vis a tergo o persuasió, suau però constant, a deixar-se emmotllar per l’etapa següent, per respondre així a un “diferencial d’evolució” que se li suggereix. El trasbals i l’agitació continus de la biosfera, com una cinta transportadora que no deixa reposar ningú gaire temps, ha de ser un motiu suficient per a induir una sèrie de característiques paral·leles i semblants en moltes línies filètiques. Augment de mida, complicació de molts dispositius orgànics, disminució i més protecció de la fillada, són característiques que apareixen paral·lelament en moltes nissagues i que s’havien considerat com a demostratives d’una suposada ortogènesi o evolució orientada o dirigida. La convergència en el canvi de molts caràcters de forma i funció pot molt bé ser simplement una conseqüència del denominador comú que admeten molts processos successionals que formen, naturalment, l’escenari en el qual té lloc l’evolució.
Val a dir que vers les etapes finals de la successió, es reconeix poca diferència d’energia entre situacions veïnes. Hi poden haver moltes realitzacions possibles de situacions dinàmicament semblants, i els ecosistemes troben poca dificultat a diferenciar-se localment. És un domini de poca desigualtat energètica entre situacions que pràcticament esdevenen equivalents i, a més, sempre resulta afavorit un allargament de la vida individual.
És un terreny propici a la diferenciació local, al joc de la vida en un domini indiferent, a fer experiments vitals, a l’aparent exageració en el disseny de tota mena de defenses, mimetismes i imitacions, a establir relacions molt complicades entre unes i altres espècies, o entre membres de la mateixa espècie. Ara, quan es parla de la conservació de les selves humides tropicals, més o menys conscientment, es té present el rerefons de complicació que sempre conserva un misteri de l’inexplicable. És ben cert que el concepte, tanmateix irreal, de clímax té una dimensió estàtica: allò que, en la seva complexitat, sembla poc funcional i té totes les característiques d’una obra d’art no forçada per les exigències del cada dia. I, malgrat les interpretacions artístiques, i per això gratuïtes, a què es presten, totes aquestes conformacions tan complicades tenen una raó de ser força utilitària, ben aparent quan hom tracta d’analitzar-les una per una.
La mandra de la natura
Un sistema és una organització no rígida, feta de diferents parts caracteritzables, sovint repetides o producte d’una fabricació en sèrie, i que estan unides les unes a les altres per lligams funcionals de naturalesa diversa. Amb una col·lecció relativament limitada de tipus de parts, i amb un joc no més extens d’interaccions, la natura produeix una gran varietat de sistemes. En són exemples els àtoms, els sistemes vius elementals unificats, és a dir, els organismes, els sistemes ecològics o ecosistemes, les societats d’insectes i les societats humanes en general, com també exemples particularitzats d’aquestes societats humanes, en forma de sistemes industrials i polítics. La definició i l’explicació d’aquests sistemes acostuma a incloure la formulació d’alguna finalitat i tendència, que molt sovint hi és posada per l’observador, el qual se situa idealment i de manera personal en el lloc del sistema en el qual pensa reconèixer aquelles característiques. Això passa segurament perquè nosaltres també som sistemes. Sovint es dóna a la tendència suposadament descoberta la forma d’una funció a maximitzar: la màxima producció de treball, la màxima eficiència, la màxima projecció de massa (biomassa) vers el futur, etc.
S’ha pogut dir que la teoria de la selecció natural ha servit per a canonitzar l’ideal de treball de la moral puritana. En molts textos de biologia s’identifica l’adequació o “fitness” amb la capacitat de deixar més descendents. L’èxit, en termes de selecció natural, no vol dir deixar més descendents, sinó passar més gens propis vers el futur, cosa que a vegades s’aconsegueix millor reduint la mortalitat dels descendents: val més que durin més i es reprodueixin just, que no pas deixar massa descendents que es malbaraten. És clar que tot depèn del món en què ens movem: si hi ha molts perills imprevisibles, el primer que s’ha de fer és deixar molts descendents per assegurar-se que algun passarà; si els riscos es van acotant i es tornen previsibles en part, el que cal és reduir al mínim la producció de descendents. Aquest és un aspecte de la mandra de la natura.
La taxa de multiplicació de les espècies no augmenta en generacions successives, al compàs de l’evolució de la nissaga, sinó que més aviat baixa per ajustar-se al mínim permissible. Això es veu arreu i podem proposar, com a exemple adient, la marxa de l’evolució de la vida en els rius. El producte del corrent del riu per la probabilitat que tenen els organismes de ser arrossegats pel mateix corrent de l’aigua (probabilitat expressable en una escala entre 0 i 1) defineix la taxa de multiplicació que és necessària per permetre que les poblacions persisteixin localment. L’estira-i-arronsa de l’evolució s’estableix entre la probabilitat d’arrossegament i la taxa de multiplicació. Els dos valors disminueixen alhora, com a síndrome del que podríem anomenar adaptació a la vida fluvial. La diferència és zero sobre períodes prou llargs, però en un riu certament tindrà fluctuacions, com a resposta a riuades o a estiatges, i les espècies han de tenir, com a previsió indispensable per a la persistència a llarg termini, un petit excés de prolificitat que els permetrà de sobreviure a aquells requeriments extraordinaris, quan s’escaiguin. O, en el cas esmentat del riu, conservar la capacitat per colonitzar l’aigua riu amunt, després que una riuada s’hagi emportat la població que vivia aigües amunt.
Les adaptacions que permeten disminuir la probabilitat o la taxa esperable d’arrossegament poden consistir en recursos més eflcaços per a subjectar-se damunt les pedres. En aquells insectes que només són aquàtics durant la fase de larva, els adults, aeris, acostumen a volar en sentit oposat al corrent del riu, per compensar, a escala de població, el resultat esperable de la deriva d’una part de les larves amb el corrent.
La natura és mandrosa: si li basta fer deu còpies no en fa cent, i la mateixa estratègia de fer còpies és una expressió de la seva mandra. Altres consideracions molt diverses acaben de confirmar aquesta tendència i aclareixen algunes possibles vies i raons per les quals es realitza. Hi ha mandra fins i tot per a morir-se: com més llarga és la vida, més possibilitats hi ha d’augmentar la mida de l’individu i d’acumular informació i, si l’entorn no és molt canviant, al mateix temps s’assegura la vida del demà. Vegeu com la mateixa tendència es continua, culturalment, en la naturalesa humana: s’utilitzen molts recursos per a allargar la vida, de tipus sanitari i de tipus mèdic, pagats amb energia exosomàtica, i, per cert, sotmesos a la llei de rendiments decreixents, ja que cada vegada és més costós, en termes d’energia exosomàtica, afegir un any a la vida mitjana estadística de les persones.
La natura, segons com es miri, no mou un dit si ho pot evitar. El premi, tanmateix ben minso, és per al qui queda en ocupació de l’espai fent el mínim de treball possible; treball en sentit físic, expressable en joules, calories o quilowatts/hora. Evidentment, un sistema que trontolla contínuament, fent aleatòria la supervivència, ha de disposar d’un nombre extra d’assaigs, la manifestació dels quals és un nombre més elevat de descendents. Però, alhora, un sistema amb aquestes característiques és l’únic que continua explorant i, per tant, l’únic que pot trobar la novetat. No és estrany que en totes les línies filètiques, les direccions que l’evolució ha reprès amb alguna novetat han començat associades amb algun trasbals, que ha servit almenys per a llançar la vida a fer noves experiències. Això és vàlid tant amb relació a la successió ecològica com amb l’evolució. Es pot pensar i creure que aquesta coincidència és el fonament d’una dependència inevitable de l’evolució respecte de la successió: aquesta dóna el bressol, la via, i aquella segueix, de manera més o menys repatània. Els arguments emprats per a mostrar la ubiqüitat de la successió, ella mateixa a l’albir dels canvis de l’entorn, semblen prou convincents.