La configuració i la disposició de la pluviïsilva
La selva plujosa equatorial o pluviïsilva és un bioma disjunt, repartit per tres continents: també en això fa notar la seva tendència a la diversificació. I és un bioma que es manifesta sota formes ben diferents en llocs relativament pròxims. Emparentades per un origen comú, les diferents selves del món han evolucionat de manera diversa al llarg dels temps geològics. En tot cas, el bioma domina una extensió considerable de les terres intertropicals.
La dispersió geogràfica de la selva plujosa
Les selves plujoses equatorials es troben dins el límit latitudinal dels 20° a banda i banda de l’equador i la majoria no ultrapassen els 10° de latitud. Tanmateix, la representació cartogràfica dels seus límits geogràfics sol variar segons els autors. D’altra banda, si bé en algunes zones la selva acaba sobtadament, en moltes altres es dóna una gradació quasi imperceptible al llarg de centenars de quilòmetres, cap a la vegetació de climes més frescos i més secs, de manera que no hi ha un punt precís on hom pugui dir objectivament que s’acaba la selva equatorial i comença un altre tipus de vegetació. En tot cas l’extensió global de la selva equatorial segueix fidelment les zones dels climes càlids i humits que es donen vora l’equador.
La pluviïsilva apareix a les tres àrees continentals o de grans illes compreses entre els dos tròpics: Amèrica, Àfrica i Àsia sud-oriental, amb l’afegit de Nova Guinea, el nord-est d’Austràlia i diverses illes del Pacífic tropical. També cal afegir el litoral oriental de Madagascar, la costa de Malabar (sud-oest de l’Índia) i el sud-oest de Sri Lanka. Arriba a ultrapassar la latitud dels tròpics en alguns punts: el sud del Brasil (on arriba a ultrapassar la latitud de 29° S al vessant atlàntic de la Serra do Mar, al límit entre els estats de Santa Catarina i Rio Grande do Sul), l’extrem sud-oriental de Madagascar, a la regió de Tölanyaro (on ultrapassa els 25° S), i a l’àrea del nord-est de l’Índia, N de Myanmar (Birmània) i territoris fronterers de Bangladesh, el Bhutan i la Xina (àrea en la qual s’acosta també als 29° N a les valls del Dibang i del Dihang, al territori d’Arunachal Pradesh, al nord-est de l’Índia). Tot plegat uns 14 milions de km2, cosa que equival aproximadament a un 10% de la superfície de les terres emergides, repartits entre més de 70 països de molt divers grau de desenvolupament (des d’Austràlia o Singapur, que figuren entre els països més desenvolupats del món, fins a alguns dels països de més baix nivell de renda d’Àfrica, Àsia o el Pacífic, incloent-hi alguns encara sotmesos a un status colonial).
El Brasil, amb el seu vast territori a la conca de l’Amazones, el Zaire, que inclou la major part de la conca del riu Zaire, i Indonèsia són els països amb més extensió de selva plujosa equatorial al seu territori. L’extensió més gran del bioma, però, es concentra en tres grans àrees pròpiament equatorials de les quals la resta de territoris de selva plujosa equatorial vénen a ser apèndixs menors, encara que no menystenibles: l’àrea sud-americana de l’Amazònia i les Guaianes, l’àrea africana del golf de Guinea i la conca del Zaire i el conjunt d’Insulíndia, Nova Guinea i l’Àsia sud-oriental.
L’herència d’un procés compartit: les memòries de Gondwana
Malgrat la considerable separació geogràfica entre unes i altres, hi ha clares similituds florístiques entre les diferents regions del bioma, que reflecteixen una herència evolutiva comuna. La majoria de les famílies i fins i tot gèneres de plantes apareixen en selves plujoses de diferents continents, separades per milers de quilòmetres d’aigües oceàniques.
Algunes de les similituds florístiques més estretes entre regions de selva plujosa de diferents continents es deuen probablement a l’excepcional capacitat de dispersió de les llavors d’algunes plantes, que han pogut ser transportades a grans distàncies per ocells o pels corrents oceànics en el decurs dels darrers milions d’anys. Aquest mecanisme podria explicar, per exemple, la presència a les selves de Madagascar d’una sola espècie de Nepenthes, gènere altament especialitzat de plantes carnívores dotades de gerricons, mentre que a l’Extrem Orient se’n troben unes 70. Tanmateix, en bona part, les causes d’aquests lligams florístics s’han d’anar a buscar en el passat geològic més llunyà, quan les diverses regions de selva plujosa del món estaven en contacte molt més estret entre elles.
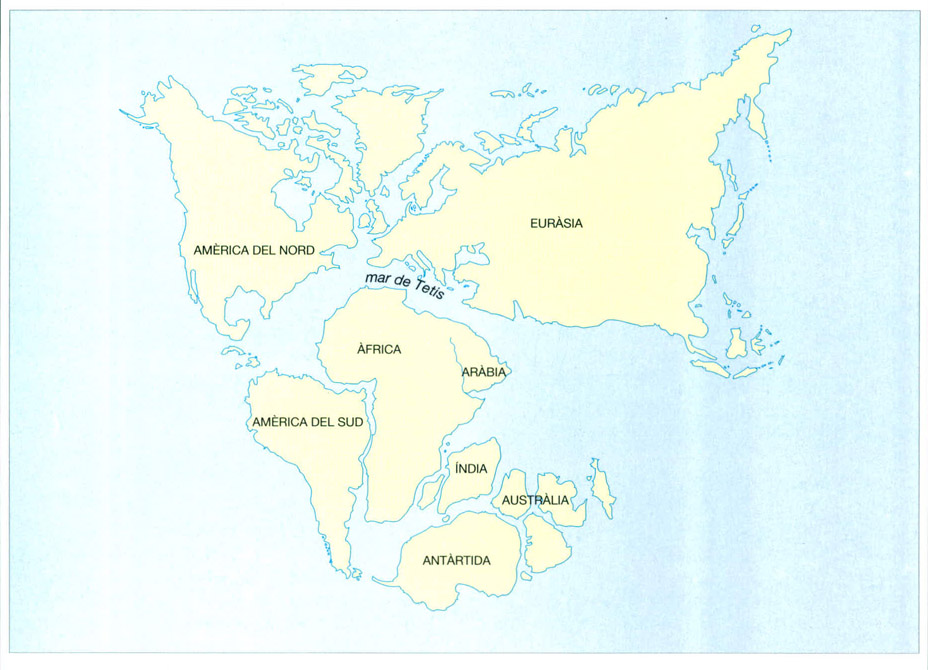
Distribució de les terres emergides durant el Cretaci inferior (fa uns 120 milions d’anys), cap al temps en què evolucionaren les primeres angiospermes o plantes amb flors. El mapa fa visible que els continents del nord ja es trobaven completament separats dels del sud, que Amèrica del Sud s’havia separat d’Àfrica, i que l’Índia ho havia fet de Madagascar. L’eixamplament de mars epicontinentals va fer separar el nord d’Àfrica i va dividir Austràlia-Nova Guinea en tres parts. La mar de Tetis circumtropical ja era evident.
Jordi Corbera, a partir de J.C. Briggs, 1987
En efecte, hi ha testimonis fòssils que indiquen que moltes de les regions de selva plujosa més separades avui, en èpoques de clima molt més humit i càlid havien estat enllaçades en unes bandes forestals contínues. Per exemple, al començament del Cenozoic, fa uns 55 milions d’anys, les selves equatorials haurien format un cinyell gairebé continu des d’Àfrica, a través d’Europa i de l’Àsia meridional, fins a Insulíndia. Tot i que aquesta faixa de selva era interrompuda de manera intermitent per braços de mar estrets i canviants, es pot suposar que les regions de selva plujosa de l’Àfrica i del sud-est d’Àsia haurien intercanviat els nous tipus de plantes en evolució amb rapidesa i eficàcia. Aquests lligams sembla que es van interrompre fa uns 30 milions d’anys, quan el clima general va esdevenir més fred i sec, però l’herència comuna més pròxima entre les regions africanes i asiàtiques de selva plujosa ha fet que s’hagin classificat conjuntament com a pertanyents al regne paleotropical (dels tròpics del Vell Món), tot diferenciant-les de les regions de selva plujosa equatorial del Nou Món (regne neotropical), separada de l’africana encara abans, durant el període Cretaci (entre 140 i 65 milions d’anys enrere), l’època en la qual les angiospermes assoliren la seva màxima diversificació i dominància. En aquell període, les masses de terra de l’hemisferi austral encara es mantenien més o menys unides en l’antic supercontinent austral de Gondwana; només Àfrica i l’Amèrica del Sud ja estaven en procés d’escindir-se, encara que romanien força properes l’una de l’altra.
Molts dels avantpassats del que ara són famílies de plantes de la selva intertropical, doncs, haurien pogut passar d’un continent a un altre, tant per ponts de terra com saltant d’illa en illa al llarg de grups d’illes volcàniques de les dorsals dels oceans que es començaven a obrir. Encara avui retrobem moltes de les mateixes famílies de plantes (com ara les euforbiàcies, les meliàcies, les moràcies o les sapotàcies), com a elements majors de la flora de totes les regions de selva plujosa dels diferents continents, encara que l’evolució subsegüent hagi donat lloc a subgrups separats, propis respectivament de les selves plujoses paleotropicals i neotropicals. Tanmateix, és difícil establir fins a quin punt s’ha produït intercanvi entre Àfrica i l’Amèrica del Sud d’ençà que es van separar. Podem suposar que la possibilitat que plantes i animals passessin de l’un a l’altre va anar decreixent a mesura que es van anar allunyant les masses continentals. En qualsevol cas, la llarga història i l’efectivitat de la separació ha deixat només una espècie comuna d’arbres (la gutífera Symphonia globulifera, l’“ehureke” de la costa ghanesa) a tots dos continents.
La regió australàsica (incloent-hi Austràlia, Nova Guinea i altres fragments menors), se separà de la part meridional de Gondwana aproximadament al mateix temps que l’Amèrica del Sud, i els grups de plantes existents efectuaren la seva pròpia evolució cap a una flora autòctona. Les condicions ambientals d’humitat tropical no existien a la regió australàsica en el moment de separar-se dels altres continents, però reaparegueren durant el Terciari, quan es desplaçaren cap al nord i s’acostaren a l’equador. En conseqüència, la regió desenvolupà la seva pròpia i peculiar flora de selva plujosa a partir de les plantes que hi havia en aquell moment, fet que atorgà un paper destacat en la flora de l’Àsia meridional a certs antics grups de coníferes que havien dominat els boscos del sud de Gondwana abans que existissin angiospermes. Així i tot, moltes de les famílies i fins i tot gèneres d’angiospermes i coníferes que evolucionaren per ocupar l’espai de selva plujosa d’Austràlia, també es poden trobar en altres continents gràcies als vincles comuns amb l’antiga flora del continent de Gondwana.
Els mateixos esquemes generals es reflecteixen en la fauna de les regions de selva, però no sempre per les mateixes raons. Així, per exemple, els simis d’Àsia i d’Àfrica formen conjuntament un grup de formes evolutivament molt properes (els catarrins), diferent del dels simis d’Amèrica (els platirrins). Però és sabut que els simis han evolucionat en temps relativament recents, i que s’han propagat des del Vell Món pels lligams d’espais forestals que existien des d’Europa fins a l’Amèrica del Nord a mitjan Terciari; després s’haurien estès vers el sud pel pont de terra de l’Amèrica Central, i finalment arribaren a l’Amèrica del Sud. A partir del trencament de la connexió per l’hemisferi nord, evolucionaren fins a diferenciar un nou grup. Trobaríem un model semblant pel que fa als rats-penats del Vell i el Nou Món, amb el nínxol que ocupen a Àsia i Àfrica els quiròpters frugívors (macroquiròpters), ocupat a l’Amèrica del Sud per un grup diferent i menys especialitzat, descendent de quiròpters insectívors (microquiròpters).
El vaivé de la història
Com que la zona contínua de bosc de clima humit que abans havia enllaçat les selves africanes i asiàtiques actualment ha desaparegut, l’evolució ha emprès també camins diferents a totes dues àrees. Així, els boscos asiàtics d’avui són dominats per arbres de la família de les dipterocarpàcies, mentre que les dipterocarpàcies africanes són arbres de sabana i de formacions llenyoses obertes que pertanyen a una subfamília força diferent de les dipterocarpàcies de l’Àsia meridional.
Durant els últims 30 milions d’anys, la història de les selves equatorials del món ha sofert un nou declivi. La tendència cap a l’aridesa a l’Àfrica i a Austràlia ha deixat només petites restes dels blocs de selva molt més extensos que hi havia abans. Sovint aquests claps aïllats de selva sobreviuen a centenars de quilòmetres lluny l’un de l’altre, arran de costa o als vessants muntanyencs on el clima es manté prou humit. Durant cada una de les fases àrides o glacials que s’han esdevingut repetidament al llarg dels darrers dos milions i mig d’anys, sembla que aquestes restes forestals encara s’havien reduït més, gairebé fins al punt de desaparèixer. Igualment, a les regions de selva plujosa d’Amèrica, hi ha la clara evidència que s’hi han produït grans períodes àrids durant aquests últims milions d’anys.
Com a dificultat addicional a aquestes vicissituds, dos dels principals dominis de selva del món, abans separats, es troben actualment en procés de fusió. Arran de la col·lisió de les plaques continentals eurasiàtica i indoaustraliana que va començar fa entre 20 i 30 milions d’anys, la dispersió de plantes i animals a través dels relativament petits estrets que actualment separen algunes de les illes pertanyents a l’una o l’altra d’aquestes dues masses continentals ha seguit un procés gradual i continu. Una herència de les seves històries prèvies diferenciades, la tenim en l’anomenada línia de Wallace (que pren el nom d’Alfred R. Wallace, el naturalista anglès contemporani de Darwin que en va fer per primer cop una acurada descripció), que és una zona de discontinuïtat de la flora i de la fauna, que segueix clarament el punt de trobada dels límits de les plaques. Així doncs, després de la diversificació produïda des de l’inici de l’era de les angiospermes, aquestes dues regions de selva es troben actualment immerses en una altra gran experiència ecològica i evolutiva.
Distribució de la vegetació durant l’últim màxim glacial
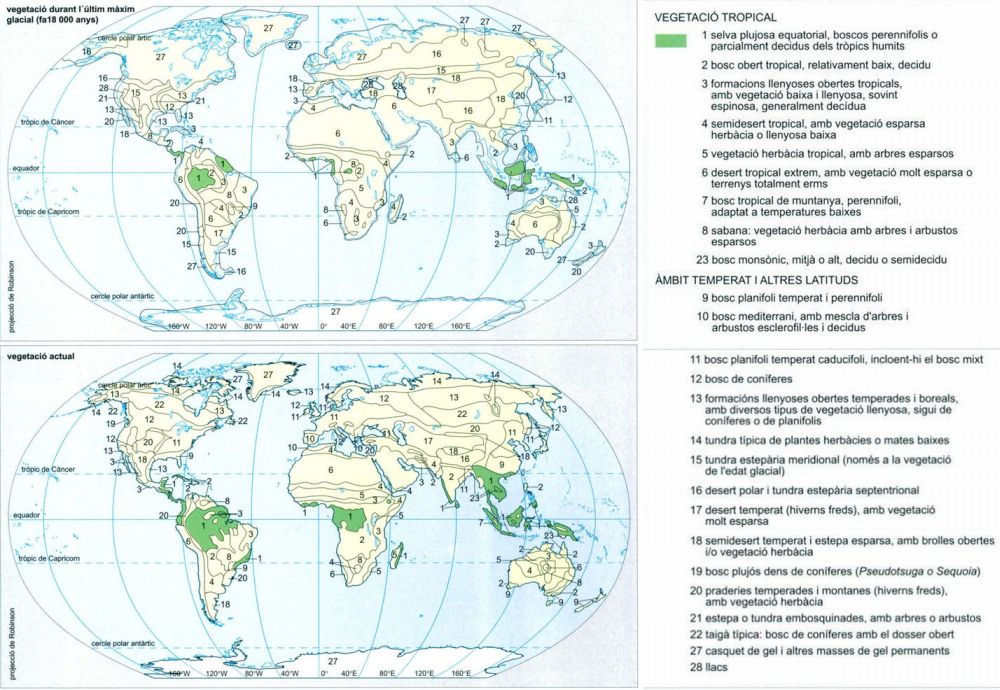
Editrònica, original de Jonathan Adams
Distribució de la vegetació durant l’últim màxim glacial i a l’actualitat (a dalt i a baix, respectivament). Fa entre 20 000 i 18 000 anys, el gel cobria gran part de l’hemisferi nord i algunes regions muntanyoses del sud, i la gran quantitat d’aigua atrapada en forma de gel feia que el nivell de la mar fos uns 120 m inferior a l’actual. L’aspecte més destacat de la vegetació de l’edat glacial és que era esparsa i oberta. A causa del fred i de l’aridesa, moltes espècies de plantes i animals es confinaren a petits refugis, on les condicions eren suficients per a sobreviure. Fa 14 000 anys, la Terra començà a escalfar-se i humitejar-se, i els boscos s’estengueren des dels refugis i pujaren fins a uns 1 000 m més amunt, a les muntanyes, on substituïren la vegetació alpina baixa. Moltes de les espècies que havien estat abundants durant l’era glacial foren forçades cap als refugis, on persisteixen actualment. La darrera edat glacial és només l’última d’una llarga seqüència de fluctuacions climàtiques que han esdevingut durant els darrers 2,5 milions d’anys. La contracció i l’expansió repetida de les àrees de distribució de les espècies que tingué lloc durant aquests cicles climàtics segurament ha tingut un paper important en l’evolució i la distribució de les plantes i els animals que veiem avui al nostre món. La distribució de la vegetació s’ha pogut reconstruir a partir de diferents fonts, com ara pol·len fòssil, ossos animals, sòls fòssils, etc. Les dades més segures són les de les terres temperades i àrtiques d’Europa i d’Amèrica del Nord, mentre que les de zones tropicals humides, ho són menys, ja que no s’hi conserven els fòssils fàcilment.
Les similituds i les diferències resultants
Com a resultat d’aquesta complexa història de canvis climàtics i de derives continentals, les selves plujoses tropicals de la Terra mostren moltes diferències estructurals i taxonòmiques regionals, que indiquen provatures evolutives. Tanmateix aquestes diferències se superposen a un remarcable fons de convergència i a un paral·lelisme entre la majoria de línies evolutives de les plantes que hi viuen, els quals confereixen una aparença general comuna a les selves plujoses tropicals de qualsevol part del món.
Així, per exemple, a Àsia, el paper d’espècie oportunista colonitzadora de les zones alterades de la selva plujosa el fan arbres de fulla gran i creixement ràpid, les euforbiàcies del gènere Macaranga, mentre que a Amèrica ho fan les properes cecropiàcies del gènere Cecropia, d’aparença força similar. Els epífits que creixen a les branques de les capçades dels arbres es troben a les selves plujoses de tots els continents, però són molt més abundants en els tròpics americans, on les bromeliàcies s’han diversificat amb enorme èxit i s’han apropiat aquest paper. Els grans arbres emergents que sobresurten per damunt del cobricel de les capçades es donen a les principals regions de selva plujosa del món, però segons els llocs hi predominen tàxons diferents: al sud-est d’Àsia, per exemple, els grans arbres de la família de les dipterocarpàcies són particularment importants com a emergents, mentre que en altres regions pertanyen, la majoria, a famílies diferents. Les palmeres formen part del dosser o cobricel forestal a totes les zones de selva, però són molt més abundants i tenen rols ecològics més diversos a les selves sud-americanes que en cap altre lloc.
La llista de semblances i diferències es podria allargar indefinidament. Observant la distribució de les selves al món, però, és obvi que hi deu haver certs factors evolutius comuns que són responsables de semblances sorprenents entre grups de plantes sense cap relació, que creixen a diferents parts del món. Tot i així, hi ha també un cert grau de desfasament que permet que certs tipus particulars de plantes amb atributs i aptituds diferencials arribin a dominar l’estructura de la selva en una àrea determinada.
Tan sorprenents com les convergències morfològiques de plantes de famílies diferents dins de cada regió de selva plujosa i entre unes i unes altres, són els resultats dels recents estudis sobre el nombre d’espècies presents a cada una d’aquestes regions. Ja s’ha esmentat que les selves plujoses tropicals tendeixen arreu del món a ser excepcionalment riques d’espècies d’animals i plantes, i encara amb algunes àrees més riques que altres, però és obvi que, dins de cada regió, alguns grups de plantes han experimentat una diversificació explosiva a causa dels atzars i de les oportunitats de la història regional. Així, per exemple, hi ha 700 espècies de “pandang” o palmera de cargol (Pandanus) a l’Àsia oriental, però moltes menys a l’Àfrica.
Tanmateix, a més gran escala, també hi pot haver una quantitat sorprenent d’ordre en el món. Uns estudis recents mostren que la riquesa d’espècies d’arbres dins de cada regió de bosc tropical tendeix a seguir el curs de la mitjana de pluviositat anual. A les parts menys humides de les regions de selva plujosa (entre 1 000 i 2 000 mm de pluviositat anual), en mostres de 0,1 hectàrees de superfície, es troben de 50 a 100 espècies, xifra que supera les 250 en les àrees d’alta pluviositat (al voltant de 6 000 mm anuals) i escassa estacionalitat. Per damunt d’aquest nivell de pluviositat (en zones on poden caure fins a 9 000 mm de pluja l’any), la corba de riquesa d’espècies d’arbres tendeix a aplanar-se clarament.
És sorprenent que, tot i les desenes de milions d’anys d’isolament i d’evolució independent, la riquesa d’espècies d’arbres entre les diferents regions tropicals de clima similar romangui força constant. Així, d’acord amb les dades acumulades fins ara, es pot esperar que una hectàrea de selva primària d’Àfrica tingui aproximadament el mateix nombre d’espècies que una superfície equivalent de selva d’Àsia o de l’Amèrica del Sud que visqui sota uns mateixos nivells de pluviositat (tot i que l’estacionalitat en la pluja també hi influeix). Aquest model sembla contradir la noció prèviament admesa que les selves africanes són relativament pobres en nombre d’espècies, a causa de la seva història geològica marcada per períodes àrids més severs que no els soferts per l’Àsia meridional o a l’Amèrica tropical. Tanmateix, l’extensa àrea de selva amb pluviositat relativament escassa, i per això mateix relativament pobres d’espècies a la regió africana de selva plujosa tropical, pot justificar el nombre globalment inferior d’espècies en aquesta regió.
Les selves plujoses africanes i malgaixes
De molt, el bloc més gran de selva a l’Àfrica és el que s’estén a l’interior i al voltant de la conca del Zaire, gran àrea de drenatge en forma de ferradura, delimitada a l’est pels escarpaments del marge occidental del Gran Rift africà. Inclou les regions d’Equador, Alt Zaire, Kivu i Kasai Oriental, a la república del Zaire, la meitat septentrional de la república del Congo i extenses regions de la República Centrafricana i del sud-est del Camerun. El seu substrat és format per roca mesozoica que recobreix un basament pre-cambrià més antic. La major part de la conca està per sota els 1 000 m d’altitud i presenta una topografia plana o suaument ondulada amb extenses zones pantanoses al seu centre. El riu Zaire drena la vall cap al golf de Guinea, a través de la plana costanera congolesa. Aquí també la roca subjacent és sobretot mesozoica, però amb roques terciàries més recents prop de la costa.
Geològicament, Madagascar és un microcontinent que es desprengué del costat oriental de l’Àfrica en algun moment del Juràssic (fa més de 140 milions d’anys). La seva fauna i flora reflecteixen la continuada proximitat geogràfica al seu origen africà, juntament amb un prolongat període d’isolament i d’evolució independent, però amb l’afegit d’un sorprenent nombre de grups de plantes i animals asiàtics. Molts d’aquests lligams entre els continents africà i asiàtic són producte de les oportunitats de propagació a través de les aigües dels oceans, en els transcurs dels últims milions d’anys. Tanmateix, certs trets característics de la biota malgaixa podrien ser herència de la distribució primitiva dels grups d’animals i plantes que hi havia a les selves terciàries de l’Àfrica oriental (o bé al subcontinent indi en moviment cap al nord), extingides de fa molt temps, que haurien reeixit a arribar a Madagascar però que haurien desaparegut dels altres llocs on vivien.
La selva africana
La selva de la conca del Zaire creix, en la seva major part, sobre sòls amb un contingut de nutrients entre moderat i alt; si més no, dins la gamma dels sòls de les selves plujoses que, en general, en són considerablement pobres. Són relativament poques les àrees d’arenes blanques molt lixiviades i pobres de nutrients, que són freqüents en zones de l’Extrem Orient i d’Amèrica. Al centre de la conca hi ha extenses àrees de bosc pantanós i de canyissar obert, algunes de les quals romanen encara deshabitades i inexplorades.
Cap al nord-oest, una franja de selva continua vers els altiplans del Camerun i al llarg de la costa nord del golf de Guinea, i inclou la totalitat del territori del Gabon i de la Guinea Equatorial (tant la seva part continental com la insular) i el sud de Nigèria. Després d’una discontinuïtat de diversos centenars de quilòmetres a la regió climàticament més seca compresa entre Cotonou, a Benín, i Accra, a Ghana, la selva reapareix i s’estén per la costa atlàntica en una amplada que oscil·la entre els 200 i els 300 km, des de Ghana fins a Sierra Leone. Cap a l’est de la conca, el clima també es torna més àrid, i la selva queda limitada a alguns enclavaments dels vessants dels relleus de la regió del Gran Rift. Cap al sud, la selva del Zaire es dilueix en un mosaic de bosc i sabana, amb els components forestals cada vegada més insignificants, a mesura que les pluges esdevenen més estacionals.
L’eixutesa relativa
En comparació amb altres regions de selva plujosa, un tret distintiu de tota la selva africana és la relativa eixutesa. Només les zones més humides dels altiplans del Camerun arriben a la pluviositat anual d’extenses àrees de l’Amazònia i de l’Extrem Orient, i encara tendeixen a tenir una molt marcada estació seca durant part de l’any. En efecte, la major part de la selva plujosa africana és situada prop dels seus límits climàtics. Amb estacions eixutes que tenen lloc una o dues vegades cada any, les capçades dels arbres del cobricel de bona part de la conca del Zaire mostren una marcada tendència a ser caducifolis, mentre que els arbres del sotabosc són de fulla perenne.
L’hàbit de caducitat de les fulles és més perceptible en els terrenys sorrencs ben drenats, en els quals, amb tota probabilitat, els efectes de la sequedat de l’estació eixuta sobre els arbres són més severs. Amb tot, la pèrdua de la fulla no és automàtica, sinó que és una reacció gradual que depèn de l’economia d’aigua de cada arbre individualment. Els anys humits per sobre de la mitjana, molts arbres de l’estrat superior conserven més temps les fulles durant l’estació eixuta, i moltes vegades les substitueixen amb fulles noves, just quan la següent estació plujosa és a punt d’arribar.
En els climes més humits de l’est del Zaire i del Camerun, els arbres de la família de les cesalpiniàcies són abundants i a vegades dominants. Les palmeres hi són rares i només prosperen en situacions particulars, com ara als llocs on hi ha hagut alteracions del bosc. En contrast amb Amèrica i amb l’Extrem Orient, hi ha molt poques plantes epífites que creixin sobre les branques dels arbres dels boscos africans. Això en part podria ser a causa del clima més sec de l’Àfrica, però potser també és una herència del fet que la important família epifítica de les bromeliàcies no ha reeixit a arribar-hi, en contrast amb els centenars d’espècies que hi ha a Amèrica.
Hi ha hagut molts debats sobre fins a quin punt la flora i la fauna actuals de la selva africana reflecteixen una història d’episodis àrids que van anihilar el bosc en algunes zones i el van deixar subsistir en altres. Aquests refugis poden haver retingut espècies que no haurien aconseguit expandir els seus efectius prou de pressa per a propagar-se i recobrir tot el seu hàbitat potencial. En efecte, hi ha senyals evidents que la coberta forestal va minvar durant la darrera glaciació, fa entre 25 000 i 12 000 anys. Si bé podem conèixer el bosc que ha subsistit en algunes àrees, com ara el Camerun, en altres àrees més a l’oest pràcticament ha desaparegut.
Hi ha indicis que la selva pot haver estat substituïda per la sabana i els prats en molts llocs de la conca del Zaire, encara que hagin sobreviscut considerables zones de bosc pantanós i de ribera. En general, els testimonis fòssils i geològics indiquen que les àrees que avui són més riques d’espècies (per exemple, el Camerun i l’est del Zaire), durant la glaciació foren refugis, i després han tornat a ser les mateixes àrees les que en el present tenen el clima més humit. Però és molt difícil de delimitar els efectes de la història glacial en el clima actual, especialment quan les dades fòssils sobre la distribució del bosc al llarg de la darrera glaciació són escasses i ambigües.
La influència humana
D’altra banda, la distribució de la selva que hom veu actualment a l’Àfrica és fins a cert punt natural i fins a cert punt l’herència d’una llarga història d’influència humana. La història humana ha estat més llarga a l’Àfrica que en cap altre indret, però es fa difícil mesurar quins efectes va exercir sobre els boscos abans que hi difongués l’agricultura fa uns 3 000 anys. El mosaic de bosc i sabana que hi ha al sud de la selva del Zaire, i que al Congo occidental s’allarga com un dit gairebé fins a l’equador, generalment es concep com el resultat recent del dens poblament agrícola que ha cremat la sabana i ha mantingut a ratlla la selva. Això no obstant, les restes fòssils indiquen que algunes parts d’aquest mosaic, si més no, han existit continuadament des de l’última glaciació, fa uns 20 000 anys. Sembla que les proporcions relatives de selva i sabana han fluctuat d’acord amb els canvis climàtics, i potser com a resposta als canvis de densitat del poblament humà. Podria ser que els humans pre-agrícoles també haguessin cremat la sabana prou sovint (per tal de fer la caça més fàcil), com per produir efectes significatius sobre l’extensió de la selva. En tot cas, d’ençà de la propagació de l’agricultura, la influència dels conreus itinerants ha estat tan profunda que ha portat molts autors a preguntar-se si ha quedat alguna selva veritablement intacta a l’Àfrica. Uns estudis ecològics sobre allò que semblaven selves primigènies han tingut com a resultat la troballa de rastres d’antics cultius i habitacions, com ara carbó, fragments de ceràmica i restes de cabanes.
Durant el segle XX, algunes parts d’Àfrica han sofert la mateixa pèrdua ràpida de la coberta forestal que experimenten altres indrets dels tròpics humits. L’Àfrica occidental ja ha perdut actualment la major part del recobriment forestal natural, a causa de l’expansió de la població agrícola, i el Camerun comença a patir el mateix problema. Per contra, la major part de l’Àfrica central es manté amb un alt grau de densitat forestal, i escassament habitada. Per imatges de satèl·lit, s’han observat senyals relativament petits dels focs provocats per activitats de cultiu, en contrast amb la situació de moltes altres àrees arreu dels tròpics humits. Tanmateix, es detecten indicis d’una xarxa reticular de plantacions i terres de cultiu que s’estenen per la selva al Zaire, que es correspondria amb la xarxa principal de carreteres. Hi ha, certament, àmplies zones de terra conreada a la costa del Gabon, i intenses activitats forestals en zones de l’est del Zaire, tot i que el seu abast real encara no ha estat mai verificat en els mapes. Amb la pressió de la població en augment, i amb les creixents inversions per part de les companyies forestals estrangeres, totes les expectatives apunten que les demandes sobre les selves centrafricanes augmentaran al llarg dels propers decennis.
La selva malgaixa
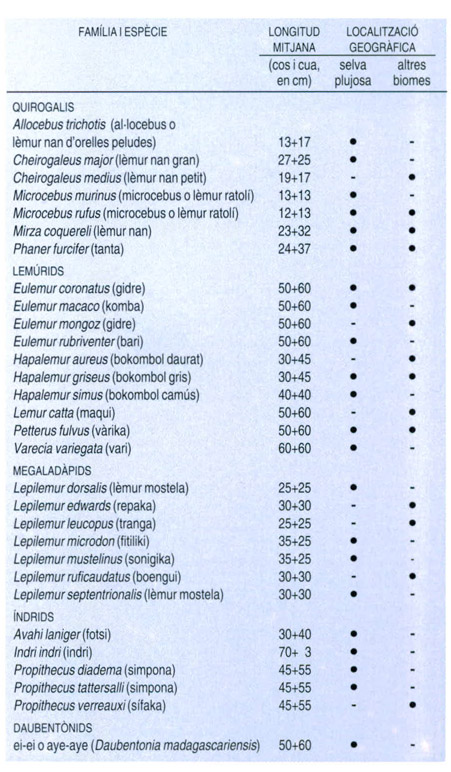
Els prosimis de Madagascar són sobretot propis de la selva plujosa oriental, tal com posa de manifest aquest quadre. Algunes espècies, tanmateix, viuen també, o fins i tot exclusivament, a les formacions forestals seques i a les bosquines xeròfiles del SW de l’illa. Algunes d’aquestes espècies han estat descobertes per a la ciència durant els anys vuitanta, com Propithecus tattersalli o Hapalemur aureus, i són moltes les que tenen un futur compromès, a causa de la destrucció accelerada a què són sotmesos els seus hàbitats. En tot cas, es tracta d’animals singulars i extremament interessants en tots els casos. Ultra els grups zoològics i les denominacions vulgars, aquest quadre també consigna les longituds mitjanes dels individus de cada espècie (cos + cua).
Dades elaborades pels autors a partir de Petter, Albignac i Rumpler, 1977
En general, Madagascar és un lloc força àrid, però al llarg de la seva riba oriental, el clima és prou humit per a sostenir una llarga faixa de selva. L’alta proporció d’endemismes d’espècies de plantes i animals (al voltant del 85% de les espècies de plantes de Madagascar són pròpies de l’illa) reflecteix la força del seu aïllament. Per això potser sorprèn que, tot i així, la selva tingui un nivell de riquesa d’espècies comparable amb el de les àrees de selva continental de clima semblant. És com si l’evolució hagués treballat “hores extres” a Madagascar per tal d’assegurar-li la seva entera quota de diversitat.Això no obstant, l’especificitat de les plantes malgaixes no va més enllà. Com que Madagascar ha viscut tan sols un període relativament curt d’aïllament, en comparació amb l’existència de les angiospermes, no ha tingut prou temps per a dur més lluny el procés de divergència evolutiva. En contrast amb l’alt nivell d’endemisme d’espècies, només hi ha una família d’angiospermes que sigui pròpia de la selva de Madagascar, les humbertiàcies. Moltes plantes malgaixes pertanyen també als gèneres que es troben arreu dels tròpics del Vell Món, que possiblement assoliren l’illa propagant-se a través del continent africà, per bé que algunes hi han diversificat un considerable nombre d’espècies. Així, per exemple, poden trobar-se cinc espècies autòctones de cafè (Coffea) en tan sols 2 km2 de selva. Atès que aquests cafès autòctons són resistents a les afeccions que pateixen els cafès cultivats procedents d’Àfrica, es pot veure que aquí hi ha un gran potencial per a la producció d’híbrids comercialment importants.
Un grup de plantes que s’ha diversificat considerablement a Madagascar són les palmeres. Hi ha dotze gèneres endèmics de palmeres a l’illa (en contrast amb només tres gèneres a l’Àfrica continental), però tots tenen fortes afinitats amb els d’Àsia. Els altres gèneres no endèmics de palmeres es troben també tots set a l’Àsia oriental, encara que tenen parents propers a l’Àfrica. Les orquídies són també diverses a l’illa, i se n’han trobat prop d’un miler d’espècies (més que en tot el continent africà), moltes d’elles a l’interior de la selva. Potser la més remarcable és l’orquídia de flor blanca Angraecum sesquipedale, que té un esperó de 35 cm de llargada. Alfred R. Wallace va predir que havia existit una papallona amb una trompa prou llarga per a arribar al capdavall de l’esperó i beure el nèctar al fons, ja que altrament l’orquídia no seria visitada pels insectes i no seria pol·linitzada. Els entomòlegs es van burlar d’aquesta hipòtesi, però el cas és que quaranta anys més tard es va descobrir una papallona que tenia una espiritrompa de 35 cm, capaç de penetrar tota la llargada de l’esperó, com havia predit Wallace. Aquesta papallona, Xanthopan morganii forma praedicta, només desplega completament l’espiritrompa quan s’apropa a flors de llarg filament, i beu immòbil sobre la flor mentre el pol·len de l’orquídia s’adhereix a la seva trompa, llest per a ser transportat i pol·linitzar la següent flor visitada.
Un altre element faunístic típic de Madagascar són els lemúrids, grup de primats primitius que en altre temps havien estat molt més estesos pel món. La selva malgaixa, certament, n’alberga diverses espècies. Probablement fou el seu aïllament en aquest continent-illa el que va fer possible que se salvessin de la competència i fins i tot de l’extinció enfront d’altres grups de primats evolucionats més recentment.
Les selves plujoses americanes
A l’interior de les conques contigües i imbricades dels rius Orinoco i Amazones, s’estén l’àrea més extensa i contínua de selva equatorial de tot el món. Aquesta àrea, unida a l’ocupada per la “mata” atlàntica brasilera, per les selves plujoses del litoral pacífic colombià, per les pluviïsilves caribenyes i per la selva plujosa mesoamericana, constitueix el ventall de bioma pluviïsílvic més impressionant del món.
La selva amazònica i orinòquica
La selva amazònica i orinòquica limita per l’oest amb la serralada andina, pels costers inferiors de la qual, fins a 1 600-1 800 m d’altitud, s’enfila, des de la Serranía de la Macarena, a Colòmbia, al nord, i al sud fins a la província boliviana de Santa Cruz. D’est a oest, formen el límit septentrional les costes atlàntiques del nord del Brasil (de l’estat de Maranhão cap al nord-oest), de la Guaiana Francesa, de Surinam, de Guyana i, a Veneçuela, la del delta de l’Orinoco; més a ponent, el límit amb els Llanos de Veneçuela i de Colòmbia ressegueix les terres baixes de les riberes meridionals de l’Orinoco i la divisòria entre el Vichada i el Guaviare. Al sud-est la delimiten les terres altes de l’oest de l’estat brasiler de Maranhão, la divisòria entre els rius Tocantins i Araguaia (Serra das Cordilheiras i Serra do Estrondo) fins a l’extrem sud-oriental de l’estat de Pará i els contraforts septentrionals de l’altiplà del Mato Grosso, de la serra dos Parecis i dels Llanos del nord-est de Bolívia. Al llarg de les valls d’alguns rius, com el Guaporé, fronterer entre el Brasil i Bolívia, o el Mamoré, el San Miguel o el Blanco, dins del territori bolivià, la selva arriba a penetrar encara més al sud. També s’estén enllà del límits aquí sumàriament descrits a les parts baixes d’algunes valls interiors dels Andes, des de Colòmbia fins a Bolívia, en particular a la vall del Napo, al Perú, d’excepcional diversitat.
El règim climàtic
L’aire humit del Pacífic que viatja en direcció est aporta la major part de la pluja de la regió, de manera que els climes més humits es troben cap a l’oest, prop dels Andes, on l’aire està carregat al màxim de vapor d’aigua. A mesura que es desplaça cap a l’est, el corrent d’aire predominant recull molt de vapor d’aigua evaporat des de la selva i l’hi torna a dipositar en forma de pluja. De tota manera, hi ha també una constant pèrdua d’aigua del sistema, amb l’abocament als rius que flueixen directament cap a l’oceà Atlàntic. En conseqüència, el clima es fa més sec d’oest a est al llarg de la conca amazònica, tot i que torna a ser més humit quan s’aproxima a la costa atlàntica pel nord-est.
En general, les selves amazòniques viuen sota un clima força més humit que les corresponents zones africanes. Malgrat que cap als marges meridionals i orientals de la regió amazònica hi ha enormes àrees de selva semiperennifòlia, hi ha també una àrea central de bosc perennifoli molt extensa. En aquestes zones més humides de la regió, amb una pluviositat anual per sobre dels 2 000 mm i sense una estació eixuta mínimament marcada, hi ha menys motius o oportunitats de sincronització dels processos biològics, i el creixement i la reproducció de plantes i animals s’esdevenen més uniformement al llarg de l’any.
Les modalitats de selva amazònica no inundable
La gran extensió dels boscos de l’Amazònia, juntament amb les peculiaritats regionals dels tipus de sòl i dels sistemes fluvials a l’interior de la conca, fan possible l’existència d’un nombre de tipus de bosc molt distints, cada un amb la seva flora especialitzada. De molt, l’àrea més gran és constituïda per la selva de “terra firme”, que és el tipus de formació característica que solem tenir a la ment quan ens referim a la selva amazònica. La selva de “terra firme” ocupa totalment els sòls ben drenats, relativament rics de nutrients disponibles. És un bosc alt (40 m o més) i normalment molt ric d’espècies d’arbres, amb un dens cobricel superior i amb un interior relativament fosc i obert.
Altres tipus de bosc es donen sota condicions que d’alguna manera són menys òptimes per al creixement de la vegetació forestal. Sobre sorres blanques lixiviades, en regions d’aigües negres, pobres de nutrients, es dóna la “caatinga” amazònica, també anomenada “campina” o “campinarana”, un bosc baix i esquifit d’arbres amb fulles petites i dures, ben diferent de la “caatinga” espinosa pròpiament dita del nord-est del Brasil. De fet, “caatinga” és un mot d’origen tupi-guaraní que significa literalment bosc blanc, és a dir sense cobricel continu que impedeixi l’entrada dels raigs del Sol. La “caatinga” es troba dispersa arreu de la regió amazònica, però particularment a la conca del riu Negro, on els sòls són formats per antigues platges marines i altres dipòsits sorrencs.
El bosc de lianes, conegut al Brasil amb el nom de “cipó” o “cipoal”, és un bosc relativament obert, en termes de cobertura forestal, però que està quasi totalment embolcallat d’enormes quantitats de plantes enfiladisses que creixen sobre els arbres i entremig d’ells. Sembla que normalment es dóna sobre ferralsòls latosolitzats, intensament erosionats i associats a importants dipòsits de ferro (com a la regió de la Serra dos Carajas) o d’alumini, encara que també es pot donar sobre els sòls, més fèrtils, anomenats al Brasil “terra roxa”. El “cipoal” tendeix a formar un mosaic amb altres tipus de bosc i és particularment abundant en la zona compresa entre Maraba i Itaituba, per on passa la carretera transamazònica.
Un altre tipus de bosc més obert molt estès a l’Amazònia és el bosc de palmeres, dominat per les palmeres babaçú (Orbignya). Sovint els boscos de babaçú, resistent al foc, són en part producte de la influència humana, com a conseqüència d’incendis freqüents. En termes d’economia indígena de les regions forestals, aquest tipus de bosc és important per les nogueres del Brasil (Bertholletia excelsa), que també hi són presents.
Les modalitats de selva amazònica inundable
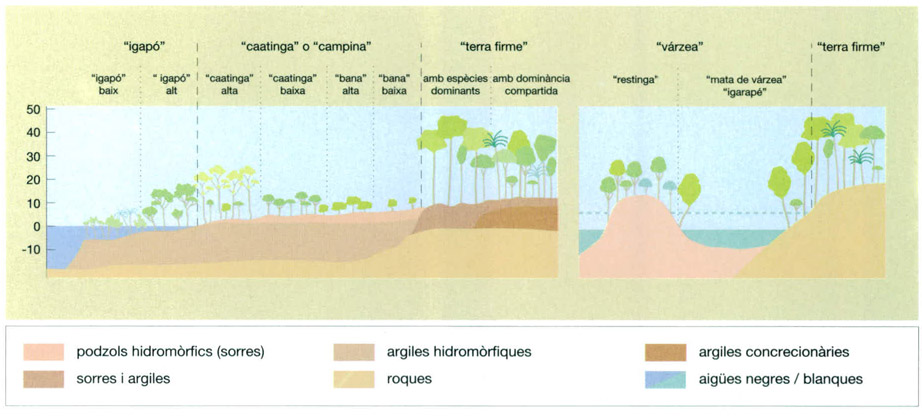
Disposició de les diferents modalitats de la selva amazònica en una conca d’aigües negres i substrats sorrencs (esquerra), i en una zona d’aigües blanques carregades de sediments que generen “restingas” (dreta).
Jordi Corbera, a partir d’E.F. Morán, 1990
Malgrat que hi ha algunes zones de bosc pantanós permanent, aquest és un component molt més escàs que no pas els boscos inundats estacionalment que s’arrengleren al llarg de les riberes i els aiguamolls de l’Amazones i els seus afluents. El caràcter altament estacional de la inundació és el resultat de les grans dimensions de la conca de l’Amazones, que amb la seva extensió al llarg de l’equador i fins als Andes en direcció oest, pot recollir la pluja estacional que cau al nord i al sud del cercle equatorial i abocar-la riu avall en immenses riuades. Els nivells d’aigua dels rius, fins i tot en les àrees relativament no estacionals més properes a l’equador, poden reflectir l’estacionalitat que es dóna en les àrees que es troben aigües amunt de la conca, on l’estacionalitat és més marcada.
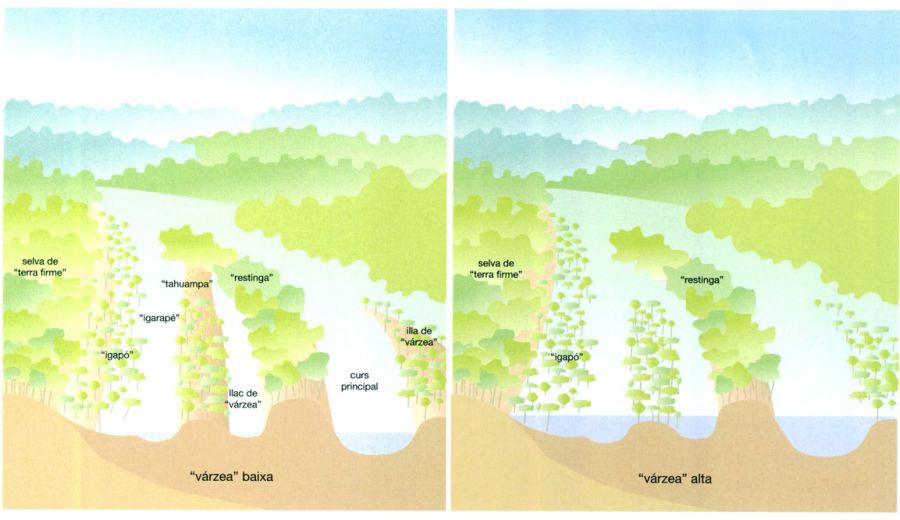
Disposició ideal de la zona de “várzea” en aigües baixes i altes, (a l’esquerra i a la dreta respectivament). A l’època d’aigües altes, tant els “igarapés” o “paranás”, que són braços secundaris, com les “tahuampas”, que són aiguamolls, com els llacs de “várzea”, que són llacunes, queden engolits per la massa d’aigua circulant, de la qual aleshores només sobresurten les “restingas” i les capçades dels arbres de les illes de “várzea”. En tot moment l’“igapó” roman més o menys inundat, i la ”terra firme” completament eixuta.
Jordi Corbera, a partir de fonts diverses
Al llarg dels sistemes fluvials de la conca amazònica hi ha diverses classes de boscos inundats estacionalment o intermitentment. Allà on el riu drena àrees de sòl argilós, els sediments en suspensió que arrossega l’aigua es van dipositant a les ribes quan es produeix la inundació. Això va bastint extensos cordons a les vores, a banda i banda del riu, coneguts com a “restingas”, que canalitzen el seu curs i limiten l’extensió de les inundacions. Aquesta àrea amb dics emergents i planes inundades s’anomena “várzea” i és recoberta per un tipus de selva —anomenada igualment “várzea”— d’estructura molt similar a la selva de “terra firme”, bé que se’n poden distingir tres modalitats ben diferents: la “várzea” sense sotabosc herbaci, densa i atapeïda, pròpia de l’alt Amazones (Solimões i Madeira), la “várzea” amb grans gramínies i més esclarissada, típica del baix Amazones (entre Manaus i l’aiguabarreig amb el Xingu), i la “várzea” d’estuari, molt rica de palmeres. Els arbres solen ser reforçats amb contraforts (potser com a adaptació d’ancoratge sobre els sòls argilosos humits), i tenen llavors amb mecanismes especials de flotació que els permeten disseminar-se quan el riu està desbordat. L’arbre del cautxú (Hevea brasiliensis) és un exemple d’espècie que està clarament adaptada a l’hàbitat de la “várzea”, amb llavors que poden surar més de dos mesos i que són una part important de la dieta d’alguns peixos. Potser a causa de la regular entrada de sediments rics de nutrients, l’estrat herbaci del sotabosc és particularment exuberant i ric d’espècies de falgueres i grans marantàcies.
A les àrees on el sediment subjacent és format per sorra lixiviada i podzolitzada, tant si són de regions de rius d’aigües clares com si ho són de regions d’aigües negres, es fa un tipus de bosc inundat molt diferent. És el cas de l’“igapó” de les regions del riu Negro i del riu Xingu, un bosc relativament baix i pobre d’espècies que creix sobre substrats magres de nutrients i que roman inundat o, si més no, embassat, més o menys permanentment. Molts dels arbres especialitzats en la vida a l’“igapó”, com Eugenia inundata, són membres de la família de les mirtàcies i es troben disposats esglaonadament segons el diferent grau d’inundació. En part a causa de la manca de cap tipus de dics de contenció, i també a causa de l’alçada relativament escassa dels arbres, les inundacions que hi ha als boscos de l’“igapó” són moltes vegades prou altes per a arribar a les capçades. Així, al període d’aigües més altes és possible poder viure l’experiència surrealista que significa fer lliscar una canoa per entre el cobricel de capçades de la selva! La inundació normalment té una durada d’unes quantes setmanes, fins i tot de mesos, i els arbres necessiten unes adaptacions bioquímiques especials per a poder sobreviure a la manca d’oxigen de les seves arrels. Durant el temps d’inundació, molts arbres deixen caure els fruits a l’aigua, on són engolits pels peixos. Sembla, doncs, que molts d’aquests arbres tenen una forta dependència dels peixos per a la propagació de les seves llavors, que han de passar pels budells d’un peix abans de poder germinar.
Els centres de diversitat: realitat o fantasia?
Dins el conjunt de la selva amazònica, no hi ha dubte que es produeix un gradient general de riquesa d’espècies d’arbres que disminueix d’oest a est, seguint la tendència de la pluviositat anual. Tanmateix, molts grups d’animals i de plantes presenten també centres d’alta diversitat dispersos, que contenen nombres relativament importants d’espècies endèmiques.S’ha apuntat que aquests centres de diversitat podrien haver estat uns refugis per a les espècies de la selva durant els períodes eixuts, coincidint amb les glaciacions, que conservarien moltes espècies que haurien estat massa lentes per escampar-se fora d’aquests refugis quan el bosc es va tornar a estendre durant els períodes interglacials més càlids. No obstant això, uns treballs recents indiquen que molts d’aquests centres de diversitat aparents podrien no ser més que una il·lusió, resultat de les diferències en la quantitat de treball de camp efectuat en diferents àrees: si s’ha fet més treball de recollida de dades en una àrea específica que en les àrees veïnes, és lògic esperar que s’hi hagin descobert més espècies! Sembla, doncs, que caldran noves recerques per establir si és cert que aquests centres de diversitat realment existeixen.
La selva o “mata” atlàntica
Al llarg de la faixa costanera atlàntica del sud-est del Brasil, de Pernambuco a Rio Grande do Sul, l’aire marítim humit dóna prou pluja als vessants orientats a mar de les serralades costaneres brasileres per a sostenir-hi una llarga franja de selva plujosa. Cap al sud, aquesta selva es dilueix imperceptiblement en el bosc temperat perennifoli, humit però més fresc, i va perdent gradualment bona part de la seva riquesa d’espècies. Els petits simis del grup dels titís daurats (Leontopithecus) en són els animals potser més emblemàtics.
Separada del bloc principal de selva amazònica per centenars de quilòmetres de brolla seca i de sabana, la “mata” atlàntica té una alta proporció d’espècies endèmiques, bé que quasi la meitat dels arbres que es troben en aquest bosc (per exemple la notable meliàcia Guarea guidonia) també es troben a la selva amazònica o en altres indrets d’Amèrica, fet que reflecteix alguna mena d’història comuna en el passat recent. Molts d’aquests arbres no endèmics també es troben dispersos per les zones profundament alterades de sabana arbrada, en forma de petites poblacions a les bandes de bosc galeria de les ribes dels rius. Així doncs, no és difícil imaginar com han pogut propagar esglaonadament les seves llavors a distàncies molt més curtes entre les àrees de bosc separades. Naturalment, també és possible que els períodes anteriors, més humits, haguessin impulsat l’extensió dels boscos galeria fins a establir continuïtat entre els diferents claps actuals, facilitant l’intercanvi gradual de plantes i animals.
La selva antillana
Al nord del bloc central de la selva amazònica, moltes de les illes escampades per la mar del Carib són parcialment o totalment cobertes de selva. Tot i que el clima de bona part d’aquesta regió és estacionalment bastant sec, l’escarpada topografia de moltes de les illes fa que l’aire marítim s’elevi, es refredi i alliberi la seva humitat en forma de ruixats. Quan això succeeix, la pluja pot excedir els 2 000 mm i les selves humides, perennifòlies o semiperennifòlies, creixen als pendents de les muntanyes i els turons.
A Trinidad, la móra (Morea excelsa) forma extensos poblaments als boscos humits però, en general, als altres llocs, aquests boscos no presenten el predomini de cap espècie en particular. A les illes de Sobrevent, a l’extrem sud-oriental del Carib, entre els arbres típics del cobricel del bosc figuren eleocarpàcies, com Sloanea caribea i altres espècies del mateix gènere, i burseràcies, com algunes espècies de Canarium. A les Grans Antilles (Cuba, Hispaniola, Jamaica i Puerto Rico) són particularment notables algunes moràcies del gènere Ficus i algunes mirtàcies del gènere Psidium, entre elles el guaiaber (P. guajava). Aquests boscos illencs són en general menys rics d’espècies que no pas els del continent sud-americà, en part segurament a causa del seu clima actual (més eixut) i passat (amb episodis de gran aridesa durant les glaciacions) i en part també pel seu relatiu aïllament de les fonts d’innovació evolutiva més àmplies del continent. És una característica general de les illes que en igualtat d’altres factors, com el clima, tendeixin a ser més pobres d’espècies que no pas les àrees continentals de les mateixes dimensions.
Entre les espècies d’arbres que es troben a les selves de les illes del Carib i a les de l’Amèrica Central i del Sud hi ha una estreta afinitat. La majoria dels gèneres són els mateixos i una extensa proporció d’espècies són comunes amb el continent (per exemple, l’esmentada Guarea guidonia, que també es troba a la selva amazònica, a la mesoamericana i a la “mata” atlàntica).
La selva pacífica litoral
Als vessants occidentals dels Andes, a Colòmbia i a l’Equador, hi ha una estreta llenca de selva que s’estén paral·lela a la costa i que per algunes valls interiors dels Andes es prolonga fins a la frontera equatoriano-peruana. Florísticament i geogràficament, aquesta selva és molt propera a l’amazònica del costat oposat de la serralada andina (amb prou feines un centenar de quilòmetres de distància en els punts de mínima separació), però probablement l’una i l’altra han estat completament aïllades des de fa milions d’anys, quan s’alçaren els Andes. Així doncs, tot i que hom hi troba els mateixos gèneres de plantes que a l’est dels Andes, sovint es tracta d’espècies diferents.
La pluviositat anual sobre els turons del peu dels Andes és extremament alta (arriba als 9 000 mm en alguns llocs) i distribuïda equilibradament durant tot l’any. En aquest clima sempre humit, la riquesa d’espècies d’arbres, lianes, arbustos i plantes herbàcies assoleix nivells gairebé increïbles. És en aquesta zona on s’ha trobat la riquesa més alta d’espècies forestals, amb 283 espècies d’arbres i lianes de tija superior a 10 cm de diàmetre, comptades en només 0,1 ha de bosc. A més, moltes espècies de plantes d’aquests boscos dels turons andins tenen una distribució molt localitzada, a vegades només un o dos turons, mentre que al turó del costat hi ha altres espècies igualment molt localitzades que ocupen el seu lloc. Això, a més gran escala, acreix encara més la riquesa d’espècies i fa d’aquests boscos una autèntica i remarcable reserva de diversitat biològica.
La selva plujosa mesoamericana
Més al nord, el bosc de l’oest dels Andes connecta amb la selva de l’Amèrica Central al Panamà, i s’estén nord enllà fins a Mèxic (on arriba, pel vessant atlàntic, fins al sud de l’estat de San Luis Potosí i, pel pacífic, fins a l’istme de Tehuantepec), excloent-ne les àrees de clima estacional més contrastat, amb estació seca prolongada com la façana pacífica de Nicaragua, El Salvador i Guatemala o el nord-oest de la península de Yucatán.
Tot i que el clima mesoamericà mostra una certa tendència a ser força més estacional que el del tròpic colombià, en aquests boscos de l’Amèrica Central encara hi ha una gran abundància d’espècies. A Costa Rica s’han trobat 233 espècies de plantes dins una àrea de mostra de només 100 m2 de bosc! En general, les selves de l’Amèrica Central són molt semblants quant a composició a les de les regions amazònica i de l’oest dels Andes, amb espècies endèmiques locals barrejades amb altres de més amplament difoses per tota la zona.
Les selves plujoses asiàtiques i indopacífiques
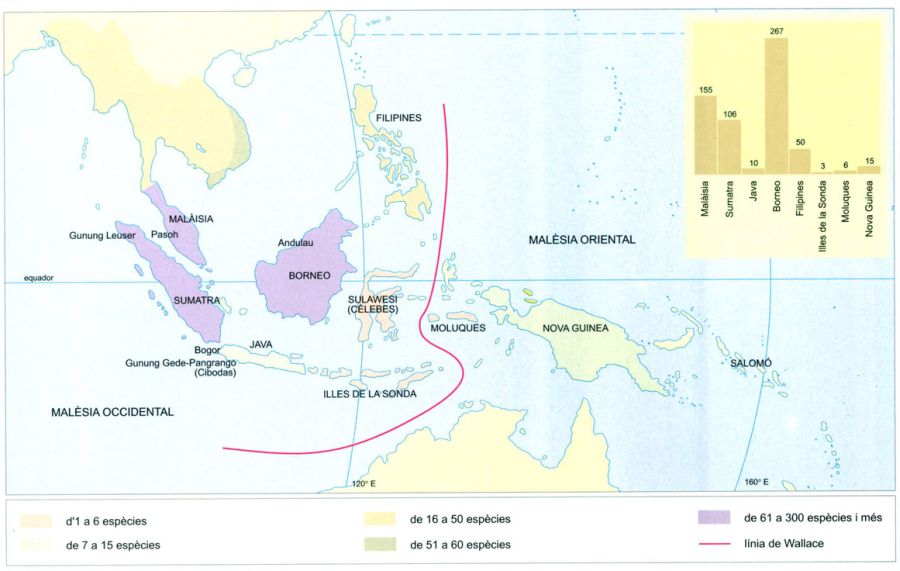
Distribució de les dipterocarpàcies a Malèsia, amb la línia de Wallace separant els dos centres principals de dispersió. Es considera que la Malèsia (regió florística amb 25 000 espècies de plantes superiors, quatre famílies endèmiques i gairebé uns 400 gèneres endèmics) inclou dos grans centres d’origen i dispersió. L’un va ser el de Borneo, Java, Sumatra i la part de la península malaia que forma part de l’antiga Sonda, des d’on la majoria de grups d’animals i plantes es van dispersar cap al continent asiàtic i les Filipines; i l’altre, la part de Nova Guinea de l’antiga Papualand, des d’on van anar cap al nord a través de Gilolo, les Moluques i les Cèlebes cap a les Filipines i l’Austràlia oriental i cap a la Polinèsia. De fet, es considera que en la flora de la Malèsia ha quedat enregistrada la clau per a desxifrar molts punts foscos de la biogeografia històrica de les plantes superiors, gràcies a la seva posició estratègica.
Editrònica, a partir de fonts diverses]
És sabut de fa temps que les selves de l’Àsia meridional i sud-oriental, d’Insulíndia, de Nova Guinea i d’Austràlia formen una unitat relativament coherent i comparteixen molts elements de la seva flora. El conjunt de terres escampades entre Indo-xina, al nord-oest, i Austràlia i Nova Guinea, a l’est, comprèn aproximadament la regió florística coneguda amb el nom de Malèsia, però és massa semblant a la regió índia per a poder ser considerada separadament. Aquestes selves són el producte de flores molt separades que van evolucionar sobre plaques continentals diferents que més tard van col·lisionar, cosa que va fer possible l’intercanvi de plantes i animals. Cada una d’aquestes col·lisions ha deixat la seva empremta en la biologia dels boscos i en els models de distribució de plantes i animals actuals.
La flora de l’Àsia tropical mostra una clara relació amb la dels tròpics africans, reflex del seu remot origen comú en la biota del gran continent boreal de Lauràsia, intermitentment connectat amb Àfrica a través d’un pont de terra, o separat d’ella només per estrets braços de mar. A l’extrem oriental, però, trobem l’herència del llarg període d’evolució independent, de la placa indo-australiana, que més tard va col·lisionar amb l’extrem sud-oriental de l’antiga placa de Lauràsia i va acollir importants aportacions de la seva flora.
La selva índia i ceilanesa
La selva equatorial índia es troba en dues àrees separades: una occidental, al llarg dels vessants marítims del sector meridional dels Ghats occidentals propers a la costa del SW de la península índia, i l’altra a l’extrem NE, prop de les fronteres amb Myanmar, la Xina i el Bhutan. A totes dues àrees, la selva semiperennifòlia del bosc monsònic, està més estesa que la selva plujosa perennifòlia, probablement a causa de la llarga història d’influència humana que ha degradat l’estructura i els sòls del bosc, fet que ha comportat un balanç d’aigua més precari.Típiques de les selves asiàtiques, les dipterocarpàcies són importants a totes dues zones de selva de l’Índia, tant al sud-oest com al nord-est, però en canvi no hi ha cap espècie comuna a totes dues zones. A la vall d’Assam, exemplars aïllats de Dipterocarpus macrocarpus i de Shorea assamica emergeixen per sobre del cobricel, tot assolint alçàries de fins a 50 m i diàmetres de tronc superiors als 2 metres. Als Ghats occidentals, robustos exemplars gegants d’altres espècies de Dipterocarpus s’enfilen fins a més de 30 m abans de presentar la primera branca.
En general, les selves dels Ghats occidentals són més diverses que les d’Assam: més de 4 000 espècies de plantes viuen en aquesta àrea de muntanya relativament petita; aquestes espècies, 1 800 en són endèmiques i la majoria limitades a la selva estricta més que no barrejades amb els altres tipus de vegetació més seca que també s’hi desenvolupa. Com ja es pot suposar d’acord amb els models generals, la màxima riquesa d’espècies es dóna en els boscos humits del sud dels Ghats occidentals, on l’estació eixuta és més curta.
A Sri Lanka, la selva plujosa de terra baixa es donaria també, de manera natural, a la major part de la meitat sud-occidental de l’illa, on la pluviositat anual és d’entre 3 500 i 5 000 mm, però en les últimes dècades ha sofert una forta regressió. Hi ha molts gèneres de plantes que es corresponen amb els de l’Àsia sud-oriental, com Mesua, de la família de les gutíferes, o Vitex, de les verbenàcies. Les dipterocarpàcies també són localment importants, amb espècies com Dipterocarpus hispidus i diferents espècies de Shorea que es troben als sòls fèrtils i humits de les planes d’inundació.
La selva malèsica occidental
Les selves equatorials de la Malèsia occidental (Indo-xina i Insulíndia) no van ser científicament conegudes fins al segle passat, i no han estat ben cartografiades fins que hom no ha disposat de dades obtingudes per satèl·lit. La totalitat de les grans illes de Sumatra, Java i Borneo, les Moluques, la major part de Cèlebes i gran part de les Filipines (tret de la façana occidental de l’arxipèlag) i de la península indo-xinesa (tret dels altiplans del centre i est de Tailàndia i de l’est de Laos i regions frontereres del Vietnam) s’inclourien en aquest grup de pluviïsilves. Cap al nord, la selva es dilueix dins de boscos monsònics caducifolis, però faixes de clima humit fan possible la continuïtat de la selva fins al seu punt més septentrional al sud de la Xina.
Els elements florístics i faunístics
Pot causar sorpresa observar que les illes disperses de selva plujosa de Malèsia occidental tenen una flora i una fauna molt similar. Moltes d’aquestes illes estan separades l’una de l’altra per centenars de quilòmetres d’aigües marines, per la qual cosa es podria esperar que l’evolució hagués emprès camins diferents a causa del seu aïllament relatiu. En canvi, tal com passa generalment arreu dels tròpics, hi ha una gran semblança entre les illes, herència de les connexions de terra que havien existit en èpoques pretèrites. En el cas de Malèsia occidental, aquests lligams van ser presents per última vegada fa menys de 10 000 anys, en el període posterior a la darrera glaciació, abans que el nivell de l’aigua pugés completament fins al nivell actual. De tota manera, hi ha un gran nombre d’endemismes i races locals que romanen confinades en illes particulars, o en parts d’algunes illes, malgrat que aquestes diferències tenen més a veure amb variacions de clima i de sòl que no pas amb la recent història d’aïllament deguda a una pujada relativament recent del nivell de la mar.
La modulació climàtica
Uns estudis sobre el capteniment dels arbres de la selva equatorial de Malàisia mostren que la lleu declinació en el grau de pluviositat que s’esdevé dos cops l’any és suficient per a estimular la renovació de la fulla o la floració de moltes espècies. Aquest efectiu estímul, que els arbres acusen clarament, es pot veure incrementat en situacions de sequera o simplement per un petit canvi de temperatura en l’estrat arbori. En algunes espècies d’arbres, sobretot de dicterocarpàcies, que no floreixen cada any sinó només esporàdicament i totes les d’una població alhora, la fructificació sembla estar sincronitzada amb les sequeres ocasionals més intenses que poden tenir lloc fins i tot a les regions més humides. Aquestes sequeres ocasionals solen estar associades amb els episodis d’El Niño. L’episodi d’El Niño de 1982-83, per exemple, comportà una important sequera a Borneo i la destrucció pel foc d’extenses zones de selva. Els focs van resultar encara més extensos i destructius a causa de la intensificació prèvia de l’activitat forestal, la qual havia escampat materials combustibles arreu del bosc; també es creu que molts d’aquest incendis van ser provocats per agricultors itinerants amb l’objectiu d’aclarir petites parcel·les de bosc, focs que ràpidament es van propagar fora de control.
Les modalitats de selva malèsica occidental
El cobricel de la selva de la Malèsia occidental normalment se situa entre els 30 i els 40 m, amb grans arbres emergents que s’eleven fins a 60 o 70 m. En la seva major part, els boscos de la regió viuen sota un clima molt poc estacional, amb una pluviositat anual superior als 2 000 mm, ben repartida al llarg de l’any. El clima més humit de tots es dóna a la regió del nord-est de Borneo, on la riquesa d’espècies també ateny els seus màxims nivells. Però en condicions particulars de sòl, causades per variacions de la geologia o del drenatge, es fan diferents tipus especials de bosc, cadascun dels quals presenta el seu propi grup característic d’espècies d’arbres, juntament amb altres espècies més àmpliament difoses.
La “keranga” és ecològicament molt semblant a la “caatinga” amazònica. És un bosc més baix que altres tipus de selva de les terres baixes, i les espècies dominants hi tenen aspecte de brucs, amb petites fulles agrupades sobre els branquillons disposats cap amunt: d’aquí ve la denominació anglesa de “heath forest” (bosc de bruguera) amb el qual han estat descrits algunes zones de Malàisia. Com la “caatinga” amazònica, aquest bosc creix sobre sòls podzolidzats pobres de nutrients. L’abundància de derivats fenòlics a les fulles, com els tanins, es creu que pot ser un mecanisme de defensa davant dels insectes herbívors i perquè així la planta no perdi els seus valuosos nutrients. Sembla que aquests derivats fenòlics són els responsables de la coloració de l’aigua dels rius negres d’aquestes àrees, que adopta un color semblant al d’un te carregat.
En els espectaculars paisatges càrstics de Borneo i de la península malaia apareixen formes silvàtiques específiques, amb espècies basòfiles. Aquestes àrees tenen una flora molt rica, amb abundants endemismes tant pel que fa als arbres com a les plantes herbàcies que s’arrapen a les esquerdes de les roques calcàries. Essent tan ben drenats de natural, els boscos que es fan a les zones de roca calcària pateixen la pressió de la secada, relativament severa durant les estacions eixutes, i moltes vegades desenvolupen colors “de tardor” abans de perdre la fulla.
Hi ha boscos pantanosos de diferents classes que ocupen àrees considerables en diferents territoris de Malèsia occidental, especialment a Borneo. El bosc pantanós de torba, per exemple, es distingeix per tenir una capa de torba d’un gruix d’entre pocs centímetres i diversos metres sota les arrels, poc profundes, dels arbres. En contrast amb el que succeeix als boscos de terra eixuta, les dipterocarpàcies rarament hi són dominants, tot i que espècies com Shorea platycarpa a vegades presenten denses poblacions. Fou precisament el bosc de torba de l’est de Borneo el que va patir amb més intensitat la secada i els incendis del principi del 1983. En aquell moment, el nivell freàtic de l’aigua va baixar tant que va provocar la dessecació d’extenses zones de torba, la qual va cremar i va causar la mort dels arbres que hi estaven arrelats.
La selva malèsica oriental i polinèsica
La regió de Malèsia oriental (Nova Guinea i nord-est d’Austràlia) és formada bàsicament per la part oriental de la placa indo-australiana, una part de l’antic continent meridional de Gondwana. Separades l’una de l’altra tan sols per una mar continental poc profunda, Austràlia i Nova Guinea van estar repetidament unides per un pont de terra en el passat, particularment durant diverses fases de baix nivell de la mar, al llarg dels dos últims milions d’anys. Tot i que aparentment el clima de selva equatorial o subequatorial era present a bona part de la meitat nord d’Austràlia durant el Terciari mitjà, la selva es troba actualment limitada a uns pocs fragments al llarg de la costa de Queensland. Per contra, Nova Guinea roman extensament coberta de selva, amb unes quantes zones de bosc monsònic a les ribes sud i est i amb boscos nebulosos a les altes muntanyes de l’interior de l’illa. Més cap a l’est les selves plujoses es retroben encara en moltes de les illes del Pacífic tropical.
Les selves d’Austràlia i Nova Guinea són dominades actualment sobretot per grups de plantes originaris de Malèsia occidental que han reeixit a difondre’s a través del límit de col·lisió de les plaques eurasiàtica i indo-australiana. En molts casos es tracta d’espècies aïllades que pertanyen a gèneres que són molt més diversos a Malèsia occidental. No obstant això, es troben gèneres endèmics o quasi endèmics de les selves d’Austràlia i Nova Guinea (per exemple el gènere Doryphora, de la família de les monimiàcies) fins i tot entre els arbres del cobricel. Es tracta d’un fort nucli de grups de plantes originaris d’aquesta regió, que en altre temps hi havien constituït una flora original de selva plujosa. Com a herència d’aquest passat, alguns gèneres de coníferes, com Araucaria, Dacrydium, Podocarpus i Agathis, encara es troben a la selva de terra baixa de la regió, de vegades en extenses formacions. L’únic eucaliptus de selva plujosa, Eucalyptus deglupta, forma poblaments purs d’arbres a la “keranga” de Nova Guinea, i té una importància econòmica considerable com a arbre fuster, fins al punt d’haver estat introduït com a arbre de repoblació a moltes regions tropicals fora de la seva àrea.