L’estructura de les plantes i la captació de la llum
Sota el sol equatorial, la selva plujosa, càlida i permanentment humida, produeix nous teixits vegetals amb més rapidesa que qualsevol altra comunitat terrestre. Aquesta alta producció s’equilibra i es manté gràcies a la mort, ja que cada dia mor tant de teixit com se’n produeix i, en descompondre’s, la matèria que fou viva genera nutrients que permeten el nou creixement. La selva plujosa és en equilibri dinàmic, i la seva taxa de reciclatge és vertiginosa comparada amb la d’altres formacions forestals de regions més eixutes o més fredes. Alimenten aquest dinamisme la velocitat de la descomposició (la selva és un niu d’organismes carronyers i capaços de digerir qualsevol cosa) i també el flux d’aigua que transporta nutrients des del sòl fins al cobricel, pouats per la succió que exerceix l’evapo-transpiració de les fulles.
De la penombra al cobricel
La vida a la selva plujosa s’estructura segons regles de domini. És a dir, gairebé tota la llum es concentra al cobricel de la selva i, amb la llum, també l’oportunitat de fotosintetitzar sucres —les peces bàsiques del creixement i del metabolisme— a partir del diòxid de carboni atmosfèric i de l’aigua. Tal com ja s’ha esmentat anteriorment, s’ha pogut observar que el 99,6% de la llum solar que arriba a la superfície exterior del cobricel d’una selva plujosa tropical és interceptat per les capçades dels arbres, mentre que l’interior de la selva roman en una densa penombra. Per tant, si una planta ha de subsistir per mitjà de la fotosíntesi (hi ha altres alternatives, és clar, com ara el parasitisme), ha de ser capaç, bé d’arreglar-se-les amb el 0,4% del total de la llum, bé d’aconseguir d’una manera o altra que les seves fulles arribin al cobricel.
Hi ha plantes fortament nemorals, adaptades a aquesta penombra, que viuen arran de terra a les selves plujoses, però representen un tant per cent molt reduït del total de la flora. Entre elles podem esmentar diverses falgueres, com ara Selaginella, i també plantes com les begònies (Begonia), diferents gramínies i ciperàcies de fulla ampla i gingebres (Zingiber i altres gèneres de la família de les zingiberàcies). Sovint, a les zones de penombra es troben petites palmeres, arbustos, plàntules i arbres joves, la majoria dels quals són, potser, destinats a arribar al cobricel, un cop hagin crescut, com a arbres gegantins o com a lianes. La selva plujosa, en efecte, és una galeria d’arbres on els troncs ofereixen als nivells més baixos una trama vertical i on les branques més altes s’entreteixeixen en un sostre.
Els arbres més alts poden superar els 60 m d’alçada i llurs troncs poden atènyer els 5 m de diàmetre al peu, on sovint són reforçats per tortuosos contraforts llenyosos. Aquests reforçaments del tronc poden partir de tres metres d’alçada o més i es reparteixen al volt de la base de l’arbre per a assegurar-ne l’equilibri. Els arbres d’aquestes dimensions, però, són una minoria. Llurs capçades solen ultrapassar les dels seus veïns i sobresortir del cobricel, i és per això que s’anomenen arbres emergents. Un dels exemples més espectaculars és el del “kempas” (Koompassia excelsa), una papilionàcia que, amb la seva suau escorça platejada, el color verd pàl·lid de les seves fulles i la seva majestuosa alçada, és un ornament de les selves plujoses de l’Àsia sud-oriental.
Sota les capçades d’aquests arbres emergents, el sostre de la selva esdevé més regular: a aquesta alçada lleugerament inferior, les capçades de la majoria dels arbres adults paren l’una al costat de l’altra formant un gruixut sostre entramat que va aproximadament dels 15 als 45 m d’alçada. És el que anomenem cobricel o dosser. Aquest estrat principal del cobricel és translúcid i estableix un gradient regular d’intensitat de la llum de dalt a baix, el qual proporciona diferents oportunitats de vida per a les plantes adaptades a uns graus específics d’il·luminació i, atès que l’aire a l’interior de la selva és més humit que a l’exterior, també a unes condicions particulars d’humitat. Dins aquesta matriu tan diversa, les diferents espècies d’arbres obren les seves capçades d’adult a diferents alçades, i cada una d’elles ho fa d’acord amb les seves necessitats fisiològiques (segons com s’hagin hagut de modificar individualment en créixer). De manera semblant, a les fulles, a l’escorça, o a tots dos llocs, solen créixer amb profusió líquens, molses, falgueres i orquídies.
L’entramat de les lianes
Tanmateix, hi ha una part important del cobricel que no és formada per les capçades dels arbres, sinó per les de les lianes. Les lianes, en efecte, constitueixen un dels elements més rellevants de la selva plujosa. És més, són quasi específiques d’aquest ambient, on tenen un paper important en la seva estructura general. A partir de les llavors caigudes o gràcies als seus propis sistemes de brotada a partir del sistema radical, les lianes projecten cap amunt una mena de rebrots exploradors que pugen en espiral, temptejant el medi que els envolta fins a trobar un suport ferm, sovint el tronc d’un arbre. Aleshores la jove liana s’aferra amb força cargolant-se al voltant del tronc o per mitjà d’unes protuberàncies especials i s’hi enfila, creixent tan ràpidament com li permeten la llum i els minerals disponibles, ja que la seva tija extensible es converteix en un feix de tubs que condueixen l’aliment i l’aigua fins a la vigorosa punta. Quan arriba dalt del cobricel, la liana projecta brots laterals al llarg de les branques del seu hoste cercant sempre la llum cap amunt i cap enfora.
Estenent-se a través de la capçada del seu arbre hoste, la liana pot acabar per connectar-se amb les branques més exteriors dels arbres adjacents. Aquests li serveixen de camí per estendre’s encara més, fins a colonitzar, sempre amb la competència d’altres lianes i dels mateixos arbres, una àmplia àrea de la selva. A mesura que creixi, la liana produirà flors i fruits i noves fulles que serviran d’aliment als animals, els quals utilitzen les múltiples tiges com a sendera per als seus trasllats diaris.
De fet, en ocupar un arbre rere un altre, una liana grossa haurà lligat les seves capçades i en molts indrets del cobricel de la selva això haurà format un veritable tapís de vegetació. Si algun dels arbres que la sostenen mor afeblit per la competència amb la liana, el seu tronc podrit es pot mantenir dret durant mesos o fins i tot anys afermat pels altres arbres. Si el tronc que mor és aquell per on ha crescut la liana, quan finalment acabi convertit en pols, els cargols que havia fet la liana en ascendir penjaran en l’aire formant una mena de gran molla estirada penjant dels altres arbres. Aquestes anelles de liana que pengen sense aparent suport del distant cobricel fins a una soca arran de terra són una característica comuna dels nivells mitjans i a peu pla de la selva plujosa.
El fet d’interferir als diferents estrats de la selva d’aquesta manera es pot considerar tant un avantatge com un desavantatge per a la liana. Una de les conseqüències menors és que les fulles que es desprenen d’una planta enfiladissa acaben a terra, lluny del seu sistema d’arrels, així que, al contrari del que succeeix amb els arbres d’arrels connectades, la liana acaba per no aprofitar els nutrients que ella mateixa produeix. Té més importància, però, el fet que sota la liana mori i arribi a caure un arbre de grans dimensions. En aquest últim cas, l’entramat que connecta la liana amb els altres arbres és un gran desavantatge, ja que, arrossegada per la força del gegant, pot caure tota la secció comunicada, i és molt probable que es destrueixi l’enfiladissa sencera.
Les palmeres enfiladisses o rotangs
A les selves plujoses del Vell Món també és normal de veure palmeres enfiladisses o rotangs (Calamus i altres gèneres del grup de les lepidocarioidees, més conegudes internacionalment per la seva denominació anglesa de “rattans”). La majoria posseeixen estructures barbades en forma de fuet de fins a tres metres de llargària, que són o bé extensions del raquis o nervadura central de les seves fulles (cirrus), o bé creixen a partir de la punta de la beina d’aquestes (flagell). Les barbes d’aquests òrgans en forma de fuet són agudes i recorbades com garfis, cosa que els permet de fermar-se a la resta de la vegetació a mesura que la palmera es fa un lloc entre les capçades. Sovint, la resta de la planta també té agullons o espines, en especial a les beines de les fulles, per a protegir la tija. Al Nou Món es troba un altre grup bastant semblant de palmeres punxants i enfiladisses, les del gènere Desmoncus i altres d’afins del grup de les cocosoidees; les seves punxes són un parell de folíols especials anomenats acantofil·les (alguns rotangs de l’Àfrica també tenen acantofil·les a més d’agulles als seus cirrus).
S’han arribat a trobar tiges principals del rotang de 186 m de llargada en un exemplar de Calamus manan a la península de Malàisia, i de 120 m en un C. albus a Java occidental. Les tiges recargolades d’aquestes plantes, ancorades fermament al cobricel gràcies a les seves frondes barbades, serveixen per a enllaçar els arbres de la selva els uns amb els altres. Algunes espècies creixen com una única tija, mentre que d’altres es ramifiquen en una colla de tiges per rebrotada arran de terra o just per sota. Aquestes espècies es troben en molts tipus de boscos de terra baixa i n’hi ha algunes, com per exemple C. trachycoleus, que estan adaptades a les inundacions estacionals. Els rotangs són força abundants a altituds mitjanes, però esdevenen poc nombrosos o quasi absents als boscos muntanyencs.
Quan encara són joves, els rotangs toleren l’ombra i, després de la germinació, poden arribar a passar uns quants anys en forma de petites palmeres. L’exposició a la llum, però, afavoreix un creixement vigorós a mesura que la petita palma cerca de fermar-se a les plàntules o a algun arbre jove proper, que constituiran el seu mode d’ascens al brillant cobricel superior. A causa de l’exposició a la llum, els marges de les selves plujoses, igual com les ribes dels rius o les clarianes pròximes obertes pels humans, sovint són infestats de rotangs que amb les seves espines formen unes barreres formidables.
Els rotangs canvien molt pel que fa al diàmetre de la tija principal i dels vims que l’acompanyen. El de la tija principal oscil·la entre els 20 cm, en alguns individus excepcionals del gènere Plectocomia, passant pels 10 cm de Calamus manan, fins a uns robustos 4 cm en algunes espècies de Laccosperma, o els prop de 2 cm d’Eremospatha i fins i tot 1 cm o menys en alguns dels Calamus, Daemonorops i els Plectocomia més petits. Els humans recullen aquestes canyes i les utilitzen amb molts propòsits com ara l’artesania o la construcció de mobles; al sud-est de l’Àsia les canyes de rotang constitueixen, després de la fusta, el segon producte de la selva més valuós.
Però els humans no són els únics que aprofiten els rotangs. Els seus fruits són una part important de l’alimentació de simis tan dispars com el dril (Mandrillus [=Papio] leucophaeus) de Nigèria, el Camerun i l’illa de Bioko (Fernando Poo) i la mona de Mentawai (Macaca pagensis) de les illes Mentawai, a la costa oest de Sumatra. En aquest darrer cas, comparant informacions de diferents conques, s’ha demostrat que les densitats dels simis són directament relacionades amb l’abundància de rotangs a la selva.
Tot i que la major part dels 13 gèneres i les quasi 600 espècies de rotangs són pròpies de l’Àsia sud-oriental, n’hi ha 9 de pertanyents a 3 gèneres diferents que també es donen a l’Àfrica occidental. En inventaris de selves primàries del sud-est de Nigèria és molt usual trobar la presència d’unes cinc espècies de rotang per hectàrea, les més comunes de les quals són Laccosperma secundiflora i les del gènere Eremospatha per més que també hi siguin presents les del gènere Oncocalamus.
Els ficus estranguladors
Però no tot aquest encordat vivent que guarneix la selva plujosa és fet de lianes que s’enfilen des de terra fins al capdamunt del cobricel. A les selves plujoses viuen també moltes espècies de ficus (Ficus), un gènere ample i divers de la família de les moràcies. Entre els tanys que pengen del cobricel de la selva, en efecte, n’hi ha uns de carnosos i d’escorça llisa, carregats de làtex, que pertanyen als ficus.
Els ficus de natura lianoide (també n’hi ha que són arbres, i no necessàriament de la selva plujosa, com la figuera mediterrània) inicien la seva vida a partir d’una llavor diminuta que ha estat dipositada a la branca d’un arbre entre els excrements d’un simi o d’un ocell. Potser els més representatius són els ficus anomenats estranguladors. Després de la germinació, la planta viu un temps com a epífita, es ferma sòlidament al seu hoste i acumula reserves d’aliment per mitjà de la fotosíntesi. Al cap d’un temps, el ficus estrangulador comença a projectar petites arrels cap avall, una o dues al començament, i moltes més tan bon punt la primera ja ha arribat a terra i accedeix a una font apropiada d’aigua. Les arrels, de 30 m de llarg o més, es van cargolant gradualment al voltant de l’hoste, i s’entreteixeixen fins a formar una xarxa que abraça l’arbre amb força. Mentre les arrels es multipliquen i es fan més gruixudes, les branques i les fulles competeixen per obtenir llum i espai amb les branques i les fulles de l’hoste, fins que aconsegueixen a poc a poc sobresortir i fer ombra a la capçada de l’arbre. Finalment, el tronc de l’hoste es troba completament revestit per l’entramat d’arrels desenvolupat pel ficus, amb poca llum i sense poder engruixir, de manera que acaba morint. Els seus teixits es descomponen i fertilitzen el ficus que, ferm sobre les seves arrels malgrat el buit deixat per l’arbre que fou el seu suport, ocupa literalment el lloc d’aquell sota el sol.
La dinàmica successional
Però els arbres que moren a causa dels ficus són una minoria. La majoria moren per altres raons. Alguns, a causa d’una estranya espècie de tèrmit, el qual, en lloc d’alimentar-se de fusta morta, s’alimenta de fusta viva; d’altres, a causa dels fongs que s’hi introdueixen quan els rosegadors o els arbres veïns que cauen els produeixen esquerdes o fissures a l’escorça; i altres, encara, moren de vells. En aquest darrer cas, les primeres branques que es mostren moribundes són les perifèriques i després les principals, fins que la capçada cau a bocins a peu mateix d’arbre. El tronc, cada vegada més necròtic, es manté dret durant un temps, tot espellifat a causa de l’escorça que li cau a trossos, fins que, de tan podrit, no pot suportar el seu propi pes i s’esfondra.
La caiguda dels grans arbres
En tot cas, la causa més comuna de la mort d’un arbre adult és la caiguda per acció del vent. La majoria dels arbres de les selves plujoses tenen un sistema d’arrels bastant precari, i el problema de l’equilibri s’incrementa amb la seva alçada. El vent, en moure les capçades, genera una forta pressió sobre els troncs dels arbres. Les tempestes tropicals solen ser precedides de fortes ventades, perquè l’aire es veu desplaçat per les gotes que martellegen la selva. Aquestes ventades sotmeten els arbres al seu pas a dures proves, fent-los oscil·lar i torçant-los i esquinçant-ne les fulles. En tals ocasions, en el silenci de la selva, se senten sons com els d’un tiroteig o com detonacions encara més fortes. Són els milions de fibres de fusta de les capçades o dels troncs que tot d’una s’esmicolen a causa de la pressió i que assenyalen la fi d’una vida a la qual havien donat suport unes quantes tones de fusta tropical. Això, posat que no sigui el sistema d’arrels tot sencer el que és arrabassat per l’empenta del vent. Sigui com sigui, esclats i espetecs precedeixen el bramul esquerdat i sibilant d’un arbre o una branca gran que cauen, o el terrabastall clamorós que fan en colpejar el terra de la selva.Després de la tempesta, quan la selva encara degota, es pot anar a veure el tronc caigut. Sol ser difícil d’arribar-hi: el lloc de la caiguda acostuma a trobar-se enmig d’un embull de plantes enfiladisses arrossegades del cobricel, lligades com es trobaven, per a la seva dissort, a la capçada de l’arbre caigut, o bé perquè s’han trobat en el camí de la caiguda. La major part de la capçada serà aixafada a causa de l’impacte i tota aquesta part de l’arbre no és més que una massa embullada de branquillons esmicolats i de fullam. Vist des d’un costat, el tronc de l’arbre caigut és com una muralla convexa amb el gra de l’escorça i les joves lianes enfiladisses encara adherides al tronc. Sovint, al costat d’aquest tronc muralla n’hi ha d’altres de disposats en la mateixa direcció. Són els troncs d’arbres més petits trencats o arrossegats pel gegant en la seva caiguda. Entre aquests i els arrossegats per les pròpies connexions de les lianes al cobricel, la caiguda d’un gran arbre pot destruir una superfície de selva de dimensions considerables.
L’obertura de clarianes
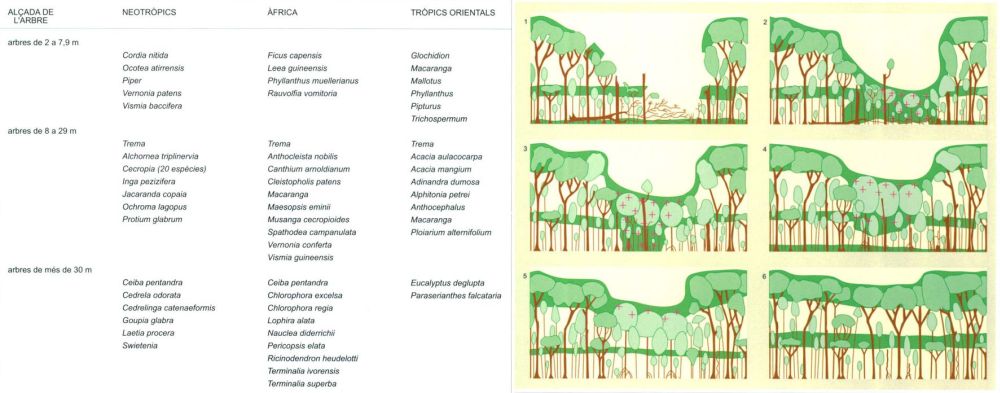
La regeneració de la selva en les clarianes té una dinàmica pròpia, que és el resultat de l’arribada de la llum als estrats inferiors on abans no accedia. La nova situació creada per la caiguda d’un arbre (1); aquest fenomen freqüent a la selva com a efecte dels ciclons, fa que s’iniciï una seqüència o successió d’espècies, que comença amb el creixement de les espècies pioneres (2), que entren en competència amb les que hi havia, i acaben essent substituïdes per les espècies més estables (3 a 6). L’esquema mostra en color verd fosc les zones on el creixement és més intens, i amb creuetes els llocs on la competència és més forta. Al quadre adjunt figuren les principals espècies pioneres d’arbres de la selva, en els tres regnes biogeogràfics del bioma. Algunes de les més grans són arbres importants per a la indústria fustera.
Editrònica, a partir de fonts diverses
Les clarianes que formen els arbres caiguts són un ingredient fonamental per a la vida a la selva plujosa. Les més grans encara poden incrementar-se durant un temps, atès que els arbres que es troben a les seves vores són més vulnerables a les ventades fortes, però la selva treballa per tancar les clarianes des del mateix moment de la seva aparició i al capdavall sempre ho aconsegueix. La clau d’aquest procés és el devessall de llum solar que a través del cobricel trencat arriba a l’abans ombrívol terra del bosc. Perquè a desgrat de l’aparent devastació que ha portat el tronc caigut, la majoria de les plàntules i dels arbres joves de l’estrat inferior sobreviuen. Creixen a partir de les llavors que poden haver germinat anys abans, però que només havien estat capaces de projectar unes quantes arrels i de fer créixer un parell de fulles a causa de la densitat de l’ombra.
En aquestes condicions, els futurs arbres sempre estan a punt per explotar l’esclat d’insolació directa. Amb la sobtada il·luminació de la clariana oberta per la caiguda de l’arbre comença una cursa: de tota la quantitat de plàntules o llavors que s’hi troben, només unes poques arribaran a la forma adulta i a tancar-la. La llum facilita a les plantes en estat de creixement l’obtenció d’una més gran quantitat d’hidrats de carboni mitjançant la fotosíntesi, i aquests, juntament amb l’aigua, serviran per a formar la major part dels seus teixits. Al mateix temps, la ràpida descomposició de la vegetació caiguda proporciona grans quantitats de valuosos nutrients, i la mort dels sistemes radiculars dels arbres caiguts o trencats significa encara més nutrients per a les plantes supervivents.
Les plàntules ja presents, però sense creixement quan es formà la clariana, són la descendència d’arbres que només es reprodueixen al cobricel madur de la selva plujosa. Però aquests no són els únics participants en la lluita per la llum. Les llavors de la majoria dels arbres del cobricel no romanen en estat latent per gaire temps una vegada han trobat un racó adient per al seu creixement. Ben al contrari, germinen, i així es posen en millors condicions que les de la llavor per a aprofitar les oportunitats que es presentin i per a protegir-se dels herbívors.
El procés de cicatrització
Hi ha moltes altres espècies d’arbres, matolls i enfiladisses que creixen segons uns models diferents. Algunes, colonitzadores especialitzades, barregen les seves llavors al llarg d’anys sencers amb la resta de matèria orgànica que arriba a terra travessant tots els estrats de la selva plujosa. Aquestes llavors han de ser molt resistents per a poder suportar llargs períodes de latència. Són estimulades a germinar per l’escalfor del sol així que s’obre una clariana i els seus plançons estan adaptats a desenvolupar-se més de pressa que no pas els dels arbres de la selva plujosa madura, fent compatible floració i fructificació amb unes estructures vegetatives de creixement més ràpid però més febles i menys durables. Si poden produir i disperar prou de pressa les seves llavors, hauran acomplert la seva funció reproductiva abans de ser definitivament sepultades a l’ombra pels arbres de la selva madura.
Altres espècies, tant del tipus oportunista com d’arbres propis d’etapes més avançades de la successió, fonamenten el seu èxit com a colonitzadores de clarianes en la germinació instantània de les seves llavors, que naveguen contínuament per la selva transportades pel vent, arrabassades en gran nombre dels arbres on han madurat per les mateixes ventades que originen les clarianes en les quals han de germinar.
Amb tants individus de tantes espècies cercant oportunitats per a assolir la maduresa i reproduir-se, les clarianes són llocs luxuriants i complexos. Tots passen per una seqüència de desenvolupament, a mesura que la clariana esdevé novament selva madura, procés que exigeix centenars d’anys per completar-se. Les diferents estratègies de creixement de les plantes que ja es trobaven en aquell indret o que hi arriben en formar-se la clariana donen com a resultat successives etapes d’avançament a càrrec de diferents espècies: l’arrencada és protagonitzada per les plàntules en latència, les llavors d’espècies de creixement ràpid o lent presents al sòl que germinen en obrir-se la clariana, i les aportades pel vent o pels animals també de creixement més o menys ràpid. A mesura que transcorren els mesos i els anys, els individus de cada espècie arriben gradualment a les seves característiques talles d’adults i comencen a reproduir-se abans no siguin ombrejats per d’altres que pertanyin a una fase més madura de la reconstrucció de la selva.
D’acord amb això, les espècies d’arbres que dominen a la clariana canvien contínuament al llarg de dècades. Naturalment, les flors, els fruits i les fulles canviants atrauen en cada fase comunitats diferents d’animals herbívors. Finalment, la clariana acaba esborrant-se quan el cobricel es tanca en quedar ocupat l’estrat superior per arbres adults entrelligats per lianes tal com és característic de la selva plujosa madura. Llevat d’unes petites prominències baixes i allargades que ocupen el lloc on van caure els arbres o d’algun clot testimoni del punt on havien estat arrelats, no resta cap traça dels grans arbres ni de les espectaculars morts que iniciaren el llarg procés de regeneració.
Mentrestant, altres arbres han caigut en altres indrets de la selva amb una freqüència que varia entre un arbre a l’any per cada 200 adults i un per cada 30, segons el tipus de sòl, el terreny i la composició florística d’unes i altres zones. La selva plujosa mai no és, doncs, uniforme, sinó que més aviat és com un mosaic de pertorbacions i de regeneració, un camp de batalla sense repòs.
Efectes de les pertorbacions a la selva
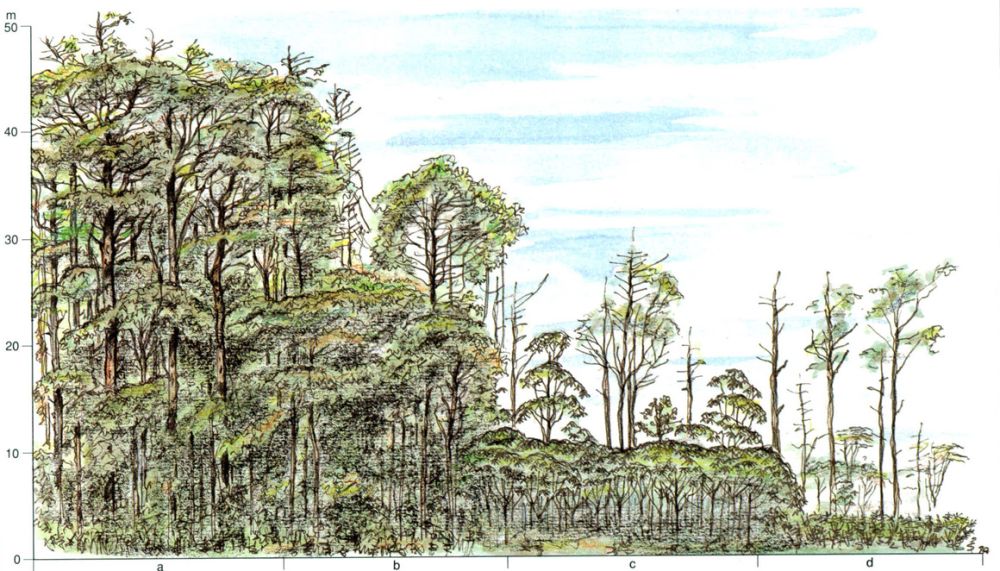
Jordi Sabater i Pi
Simplificació progressiva de l’estructura de la selva com a conseqüència de l’acció de factors pertorbadors, en aquest cas la pressió humana (apunts de camp corresponents a la selva plujosa guineana, al país fang). La selva primària (a) és, en aquest cas, un bosc multistratificat dens, d’uns 50 m d’alçada, mentre que les clarianes explotades agrícolament (d) són conreus que conserven alguns dels arbres originaris. L’abandó dels conreus porta a la instauració d’una incipient comunitat forestal o bosc terciari (c), dominada per espècies arbòries oportunistes, de creixement ràpid. L’explotació parcial del bosc primari, o el pas dels anys després de l’abandó agrícola, comporten la instal·lació d’un bosc secundari (b) menys dens i alt que el primari, amb individus arboris encara joves o amb espècies senescents del bosc terciari, si és el cas. Al domini de la “mata” atlàntica brasilera, entre d i c, solen instaurar-se comunitats arbustives o arbòries baixes (1-3 m), les anomenades “capoeira” i “capoeirão”. En cas d’accident el procés successori és similar.
Els processos coevolutius
El dinamisme permanent del medi de la selva plujosa és, paradoxalment, prou predicible pel que fa a les espècies que hi viuen. Sigui quina sigui la fase de maduresa d’una selva a la qual estigui adaptat un organisme, sempre és disponible en un punt o altre de l’entorn un espai que es troba en aquella fase. A les selves plujoses molt antigues, la competència per la llum i per l’espai ha acabat produint plantes extraordinàriament adaptades per a contramaniobrar enfront dels seus competidors. Qualsevol atribut que permeti a un arbre de créixer més ràpid o més bé que els seus veïns, encara que l’avantatge sigui mínim, donarà a l’arbre més probabilitats de reproduir-se.
Les simbiosis i els comensalismes: formigues i bartèries com a cas
A causa de l’aire càlid i humit de la selva plujosa, les fulles dels arbres són colonitzades promptament per epífits com ara les molses o els líquens. Aquests interfereixen en la captació de la llum per les fulles sobre les quals creixen, un seriós inconvenient per als arbres, que es troben permanentment amenaçats de veure’s privats de la llum que els fa viure. Les plantes enfiladisses són un problema semblant, ja que el seu pes pot limitar el creixement dels arbres joves i les seves fulles poden competir per la llum amb les de l’arbre sobre el qual s’enfilen. Un arbre que trobés la manera de mantenir netes les fulles i lliures de lianes les branques tindria un immens avantatge en la lluita per la supervivència.
Un gènere d’arbres de la família de les passifloràcies ha superat aquest repte. Els arbres del gènere Barteria han perfeccionat una tècnica per a allunyar els epífits i les enfiladisses, l’èxit de la qual es pot mesurar pel fet que són arbres molt comuns a la selva plujosa de l’Àfrica occidental, i que per a créixer i reproduir-se no necessiten arribar al cobricel superior de la selva, a desgrat de la densa ombra en la qual viuen. La clau de l’èxit de les bartèries és l’íntima relació que tenen amb colònies de formigues del gènere Pachysima. Així, doncs, les bartèries són plantes mirmecòfiles, proveïdes d’unes estructures especials que afavoreixen que les formigues visquin prop d’elles o dins mateix. En el cas de Barteria, les branques buides compleixen a la perfecció aquest paper; les seves galeries llargues i tubulars són, per a les formigues, carreteres, pastures, llocs de cria i amagatalls protegits del perillós món exterior de la selva plujosa. Quan un plançó de Barteria ja té cap a un metre d’alt comencen a créixer les seves primeres branques horitzontals.
Un bon dia aterra sobre la planta una reina alada de Pachysima, tot just fecundada durant el seu vol nupcial des de la colònia on va néixer. Ràpidament, la formiga reina escorcollarà la jove bartèria per assegurar-se que cap altre congènere no hi hagi fixat la seva residència i, confirmat això, comença a mastegar per obrir-se pas cap a la cavitat d’una de les branques laterals. Un cop dins, es desprèn de les ales, comença a pondre ous i té la seva primera progènie mentre sobreviu a base del greix prèviament acumulat i del teixit dels músculs de les seves ales. Tan aviat com surten de les pupes, una vintena de formigues obreres surten a la superfície de l’arbre i cerquen els possibles caus d’altres reines de la seva espècie. Si en troben algun, les obreres hi entren i maten sense contemplacions la reina rival i tota la seva progènie en creixement. A partir d’aquell moment es pot considerar que la bartèria ha estat adoptada per la colònia de Pachysima en creixement i que ara els destins d’una i altra han quedat entrellaçats. L’arbre, tanmateix, encara no és totalment controlat per les formigues, ja que encara no són prou nombroses. La superfície de l’arbre encara pot ser recorreguda per altres animals, incloses formigues d’altres espècies, ben capaces de matar els individus de Pachysima que troben desprotegits. Les del gènere Oecophylla, per exemple, que són formigues teixidores roges molt agressives, disputaran violentament amb les obreres de Pachysima el dret de compartir les rutes d’abastament al llarg de les branques de la bartèria.Les formigues Pachysima obren petits clots a intervals regulars dins de les branques buides de Barteria. Petites cotxinilles aplanades de la família dels còccids, probablement portades per les mateixes formigues, es fermen en aquests clots i constitueixen per a les formigues una mena de bestiar estabulat. Les cotxinilles empalmen les seves peces bucals al flux de saba que circula pels teixits de la bartèria i deixen que el fluid de la planta passi cap als seus intestins. Les cotxinilles absorbeixen una certa quantitat de sucres i minerals de la saba, però en deixen passar tanta més que al final s’escapa pels seus anus en forma de “rosada de mel”, un licor dolç i nutritiu que constitueix l’aliment bàsic de les formigues. La resta l’obtenen dels fongs que creixen en “horts” acuradament preparats també per les formigues a les branques de l’arbre. Arribat aquest punt, suposant que la bartèria creixi sense problemes, la colònia de formigues, mercès a la planta, és autosuficient pel que fa a l’aliment. Però en els foscos estrats inferiors de la selva, aquest creixement no seria possible si les negres formigues Pachysima no l’afavorissin. Això no ho poden fer de manera eficaç fins que la colònia ha pogut créixer al llarg d’un any aproximadament, que és quan ja hi ha prou obreres per a fer veritables avenços enfront dels seus propis enemics i dels de l’arbre. És aleshores que les obreres de Pachysima, armades amb agullons potents i verinosos, dirigeixen els seus esforços contra competidors, com les ja esmentades formigues Oecophylla, lluitant amb elles tant individualment com en grup i escampant els seus cadàvers al voltant de l’arbre. Assolida la victòria, les formigues negres es mouen activament per la tija i les fulles de la seva llar. Els insectes herbívors que hi volten, tant si són adults com larves, són expulsats de l’arbre tan bon punt els troben; les molses, els líquens i d’altres contaminants com les deposicions d’ocells, són netejats de la superfície de la fulla i llençats de mica en mica cap a terra. Mentrestant, les obreres ataquen qualsevol fullam que gosi tocar la capçada de la bartèria i masteguen fins a no deixar-ne res els borrons i les tiges de les plantes que creixen per sota del seu arbre. Molt aviat es forma un cercle buit de fins a 3 m de diàmetre que envolta la bartèria, la qual, lliure de lianes i amb les fulles netes, pot créixer vigorosament de 60 a 90 cm l’any.
A més de defensar el seu arbre dels efectes del creixement més ràpid d’altres plantes i de deslliurar-lo dels insectes herbívors, les formigues Pachysima també semblen ben capaces de fer recular animals més grossos que d’altra manera podrien arribar a menjar-se el petit arbre. Regularment, les obreres es llencen des de les branques més altes, formant una pluja lenta que cau sobre qualsevol mamífer que pasturi prop de l’arbre. Si n’ensopeguen algun, les formigues reaccionen agressivament: busquen un espai de pell nua tot injectant un i allí mosseguen amb força entre el pèl un virulent verí. Els humans que els han experimentat descriuen aquests atacs com a extremament dolorosos, fins al punt que poden produir discapacitacions (en alguns llocs de l’Àfrica occidental, un càstig tradicional per a les dones infidels consistia a lligar-les al tronc d’una bartèria…).
La colònia de formigues va creixent al mateix ritme que ho fa l’arbre, i es va desplaçant gradualment cap a les branques més altes, mentre que les inferiors són abandonades i cauen. Tant l’arbre com la colònia de formigues deixen de créixer al mateix temps, quan aquestes darreres han arribat a assolir el nombre d’unes 4 000 obreres. Aleshores, les plàntules de Barteria properes a un arbre gran amb una colònia ja nombrosa poden ser adoptades i protegides per les obreres d’aquesta, ja que, en produir noves reines de tant en tant, necessiten nous arbres on iniciar noves colònies.
Les metzines que alimenten
Aquesta mena de complexes dependències mútues entre espècies diferents, com les descrites entre Barteria i Pachysima, són un tret molt característic de la selva plujosa, i alhora constitueixen un tema fonamental i fascinant de la seva ecologia. Il·lustren la complexitat de la coevolució, és a dir, a l’adaptació d’una espècie a tenir un paper especialitzat respecte d’una altra espècie. Tanmateix, no totes les relacions d’aquesta mena són benignes: les relacions predador/presa també poden arribar a ser tan especialitzades o més. Així, per exemple, hi ha moltes plantes que protegeixen dels herbívors les seves fulles en creixement carregant-les de metzines com els alcaloides. Això no les fa completament immunes a la predació, però redueix el nombre d’espècies d’animals capaços de menjar-les, cosa que els proporciona moltes més possibilitats d’atènyer la maduresa abans no siguin descobertes per individus d’alguna de les poques espècies amb capacitat per a destruir-les.
Tant les plantes com els animals han d’administrar amb cura les seves reserves d’energia i de nutrients; els animals, però, com que no són capaços de produir aliment per fotosíntesi, han de viure amb reserves més aviat precàries, i destoxificar els alcaloides és prou costós. Els invertebrats en particular, per les seves reduïdes dimensions i, per tant, per les poques reserves acumulades i el seu volum corporal petit, tendeixen a especialitzar-se. Si una eruga arriba a ser capaç de descompondre l’alcaloide d’una espècie de planta en particular, la seva descendència podrà viure només dels borrons d’aquella espècie de planta. Amb el temps, l’eruga en qüestió podrà evitar la competència amb les altres erugues restringint el seu aliment a una planta hoste determinada. Aquest fenomen pot provocar l’eliminació d’algunes vies metabòliques redundants, cosa que farà esdevenir l’eruga cada cop més eficient a l’hora d’alimentar-se d’aquella planta hoste determinada, però cada cop menys eficient per menjar qualsevol altra cosa. Les papallones adultes de l’espècie cercaran borrons de la planta hoste on poder dipositar els seus ous, ja que els que siguin dipositats sobre altres plantes tindran seriosos problemes perquè es trobaran en desavantatge respecte de la resta de competidors.
Aquesta especialització esdevé més i més refinada en les generacions successives, fins que aquella espècie de lepidòpter depèn totalment de l’espècie de planta que el pot alimentar. Mentrestant, aquest mateix procés es pot haver ramificat en un procés d’especiació, ja que hi pot haver insectes dels quals hagin derivat formes diferents amb metabolismes diferents, cada una amb unes necessitats peculiars del seu llinatge. Per a cada una d’elles, qualsevol aliment del món que els envolta que no sigui prohibit és obligat.
La guerra de la diversitat
Aquesta especialització dels invertebrats, a més de generar una quantitat molt alta d’espècies animals, té, com a mínim, dues conseqüències importants per a les plantes. Totes dues resulten del fet que la predació especialitzada és potencialment molt severa, ja que els teixits de creixement més delicats d’una planta queden a l’abast d’un herbívor altament especialitzat. En primer lloc, això significa que la densitat dels individus de plantes hoste pot alhora controlar i ser controlada per la densitat dels seus predadors. Si les plantes s’agrupen densament, els insectes a punt per pondre les trobaran amb més facilitat i, per tant, n’hi haurà més d’afectades o fins i tot mortes, mentre que si les plantes són poc nombroses, la proporció afectada serà més petita. Fet i fet, per a moltes espècies, el nombre d’individus serà aquell que resulti en una densitat equilibrada amb les demandes i les capacitats dels seus hostes o predadors, un fet que aclareix, en part, per què hi ha tanta diversitat d’espècies a la selva plujosa: moltes espècies, totes prou rares, juguen entre elles un joc mortífer de fet i amagar.
L’altra conseqüència d’aquest sistema és la “cursa d’armament” evolutiva. Els insectes que siguin més capaços de trobar els seus escassos hostes, tindran una descendència amb més capacitat de supervivència, que portarà a una millora dins l’escala d’eficiència predatòria. Al mateix temps, les plantes hoste que puguin refinar les seves defenses per a invalidar les adaptacions dels seus enemics, guanyaran probabilitats de no ser consumides, i, per tant, augmentaran la seva capacitat de reproduir-se amb èxit. L’adaptació comporta la contraadaptació, i aquesta la contracontraadaptació, seguida de la contracontracontraadaptació. En aquest joc, l’avantatge més subtil pot ser crucial, tant pel que fa a la lluita entre l’hoste i el predador, com pel que fa a la competència permanent entre la planta hoste i les altres plantes, com a la pressió de la predació sobre el predador. Les dificultats de la vida a la selva plujosa són molt variades i totes les espècies lluiten en diferents fronts alhora.
La disseminació dels propàguls
El tercer gran imperatiu dels éssers vius, després d’obtenir els aliments i d’evitar la predació, és reproduir-se. La coevolució, doncs, també és important pel que fa a les estratègies reproductives de plantes i animals a la selva plujosa. Per això, en aquest àmbit, la reproducció sexual de les plantes està tan carregada de complexitat i de risc com el quotidià manteniment del creixement i de la forma.
Els sistemes de dispersió
Els individus de determinades espècies són escampats en un ampli territori, aïllats els uns dels altres per espais inhòspits. Tanmateix, aquests individus han de ser capaços d’intercanviar el pol·len a través d’aquests espais per tal de deixar llavor i, un cop acomplert això, encara cal que les llavors mateixes arribin a algun lloc adient per a la germinació. Com que les plantes adultes que produeixen pol·len i llavors són immòbils, cal que aquests elements vitals recorrin sans i estalvis distàncies considerables per tal de deixar descendència satisfactòriament.
La dispersió anemòcora
Per resoldre aquest problema, les plantes han d’utilitzar la capacitat de mobilitat del seu entorn i servir-se’n per dur a terme la seva activitat reproductiva. L’aigua, l’aire i els animals es mouen perpètuament entre les plantes de la selva plujosa. El primer element, tot i que és un medi adequat per a les cèl·lules sexuals d’algunes plantes, no sol ser utilitzat per la gran majoria. El vent, el solen utilitzar les plantes de climes temperats per a la dispersió del seu pol·len i de les seves llavors, però amb el dens desgavell de l’entramat vegetal de la selva plujosa, al transport aeri li manquen fiabilitat i abast; el problema s’agreuja encara més perquè la humitat de la selva plujosa tendeix a sobrecarregar amb aigua els grans de pol·len, cosa que en limita la mobilitat. Les llavors més petites poden recórrer distàncies importants, si disposen d’ales o plomalls que les facin arrossegables pel vent i si són alliberades per les plantes productores a prou alçada. De fet, una quarta part o més de les espècies que habiten el cobricel superior solen dispersar les seves llavors gràcies al vent. Alguns arbres produeixen immenses fructificacions de llavors lanuginoses que, amb les ventades, desfermen sobre la selva una aparent tempesta de neu.
Els boscos de camforer de Borneo (Dryobalanops aromatica) de la Malàisia peninsular, per exemple, es mantenien gràcies a la gran pressió reproductiva que suposaven les seves escampades massives, regulars i freqüents de llavors alades. Constituïen una de les rares formes de selva plujosa dominada per una única espècie d’arbre, les llavors de la qual, tanmateix, mantenien les poblacions migradores d’una subspècie particular de porc barbat (Sus barbatus oi). Malauradament, el camforer de Borneo era massa atractiu per a la indústria de la fusta i aquests boscos han estat devastats fa pocs anys.
La dispersió zoòcora
Algunes altres espècies per propagar les llavors utilitzen forces internes; els seus fruits s’obren explosivament propulsant els propàguls a certa distància. Per a la gran majoria de plantes a la selva plujosa, però, la clau de la sexualitat i de la dispersió de llavors són els animals. Aquesta relació entre els animals i les plantes ha arribat a uns nivells d’elaboració excepcionals, potenciats per la selecció natural, que actua amb gran força i precisió al llarg de les generacions. De fet, són poques les plantes que, com Barteria amb les formigues, usen els animals d’una manera sofisticada durant el creixement vegetatiu; a l’hora de reproduir-se, però, l’especificitat, la coadaptació i l’ajuda mútua entre els animals i les plantes són la regla més que l’excepció.
Les flors són les que produeixen i reben el pol·len i, per tant, si els animals han de servir els interessos de les plantes, han de sentir-se’n atrets. Les flors de la selva plujosa són tan diverses com la mateixa vegetació: de colors vius i de colors apagats, fràgils i robustes, suaus i aspres; en forma d’embut, de plat, de bol, de campana, de llaç, de raspall, de gola o de bandera. Entre la varietat d’espècies que es troben a la selva, n’hi ha que floreixen en indrets espectaculars: al mig de les fulles, per exemple, o directament a partir de l’escorça o de les branques; les flors poden brotar d’arrels enterrades, penjar oscil·lant de llargues tiges, o guarnir els branquillons més extrems del cimal de la capçada d’un arbre.
No hi ha gaire marge d’error en la pol·linització. Una planta que atregui massa pocs animals, o animals d’una espècie errònia, pagarà una penalització instantània en llavors perdudes i, de la mateixa manera, les flors més efectives obtindran beneficis immediats per a la seva espècie. Aquest passar comptes evolutiu, molt més afinat que el que pot aplicar-se en altres estructures vegetals, és el responsable de la gran diversitat de flors a la selva, les quals resulten de la complexa adaptació de les plantes als animals pol·linitzadors.
Els animals més utilitzats com a pol·linitzadors són aquells que regularment recorren amb celeritat grans distàncies de flor en flor; la majoria són alats: abelles, vespes, papallones, arnes, escarabats, ocells i rates pinyades. No és cap coincidència que les formigues, tot i ser molt corrents a la selva plujosa, no siguin quasi mai utilitzades com a pol·linitzadores: són massa sedentàries per a ser útils i més aviat sovint són perjudicials per a la pol·linització, perquè són lladres de pol·len i de nèctar.
El joc de les atraccions a l’hora de pol·linitzar
Per a poder carregar els animals voladors amb grans de pol·len, les plantes els han d’atraure, i això es pot aconseguir de diverses maneres. En alguns casos, especialment en el de les orquídies, les flors imiten objectes rellevants per a alguns insectes en particular, com ara plantes alimentàries o preses animals, i això fa que els insectes s’hi posin a sobre. De manera semblant, les flors poden adquirir la forma d’individus de la mateixa espècie de l’insecte en qüestió, temptant els membres que provin de copular, o d’atacar per defensar el seu territori. El més usual, però, és que les flors es mostrin tal com són, i que confiïn en el seu propi valor per a atreure els pol·linitzadors potencials que es trobin a prop d’elles. És per això que la majoria de les flors actuen com a elements senyalitzadors amb l’objectiu de deixar-se veure, talment com llums de neó en la nit, anunciant la seva localització amb el color i la fragància. Entre els pigments més comuns que decoren les flors hi ha els carotenoides, que són els que els proporcionen els colors groc, vermell i taronja i que reben noms tan sonors com rodoxantina i rodoviolascina. Curiosament, cap animal no és capaç de produir carotenoides sol, i tanmateix, aquests pigments són fonamentals per a la bioquímica de la visió: és com si les plantes haguessin facilitat als animals la capacitat de veure-hi, només per explotar-los…
Les dèries dels insectes
Igual que les seves formes i posicions sobre la planta, els colors i les aromes de les flors es relacionen amb els animals als quals han de temptar. A la selva plujosa es poden reconèixer diverses “síndromes” que consisteixen en combinacions de trets que suggereixen el tipus de pol·linitzador al qual s’ofereixen les flors. Les flors de color porpra cerrudes i pudents com ara l’apropiadament anomenada Amorphophallus recorren a les mosques carronyeres com a pol·linitzadors. Les abelles són atretes per flors més complexes, molt acolorides, ja que els animals són actius durant el dia, i sovint equipades amb reforços i ratlles, que reflecteixen molt vivament els raigs ultraviolats, un color molt visible per a les abelles. Curiosament, les flors vermelles i taronja més brillants atrauen els ocells, sobretot els suimangues de la família dels nectarínids, mentre que les de colors vívids però més suaus generalment en forma de trompeta són les que alimenten les papallones, les quals poden arribar fins als racons més profunds gràcies a les seves llargues probòscides.
A la nit, les flors d’olor suau i de colors pàl·lids són visitades pels borinots, i en especial per les esfinxs (esfíngids), que amb les seves llargues llengües arriben fins al fons dels tubs que formen les flors en unir els seus pètals. Moltes de les flors escarabat també s’obren de nit, i la selva plujosa a la nit és perfumada amb una mescla de les seves fragàncies. N’existeixen milions d’espècies tropicals. Són unes criatures poc àgils i, en conseqüència, les flors que volen beneficiar-se dels seus serveis han d’obrir-se en forma de bol o de plat per tal que els escarabats hi puguin accedir fàcilment.
Els rats-penats i les pàrquies
Els rats-penats també són molt diversificats i omnipresents, i omplen les nits de la selva plujosa amb el seu voleteig. Disposen de moltes flors per al seu ús particular. Algunes pengen allunyades dels entramats vegetals perquè s’hi puguin apropar volant sense problemes. Altres es troben sobre suports ferms com ara les branques o els troncs d’algun arbre. Els rats-penats són uns pol·linitzadors excel·lents, en part perquè són grans i peluts (i per tant, tenen més capacitat per a transportar més quantitat de pol·len), en part perquè s’alimenten recorrent grans distàncies, i finalment perquè estan dotats d’una memòria i una intel·ligència de mamífer. Això significa que són molt eficients per a recordar els arbres on es fan les flors que els serveixen d’aliment i també el camí que hi porta, cosa que els permet de volar sistemàticament d’arbre en arbre, nit rere nit, fertilitzant una flor rere l’altra a mesura que es van obrint.
Aquesta eficàcia és perfecta per als arbres, però exigeix una forta inversió en estructures adequades perquè el sistema de pol·linització pels rats-penats funcioni. El gènere Parkia de la família de les papilionàcies, al qual pertanyen alguns arbres àmpliament distribuïts a les selves plujoses, ho il·lustra molt bé. La seva florida consisteix en la desclosa d’una quantitat innombrable de petites flors aplegades en unes inflorescències bulboses, de manera que recorda una brotxa d’afaitar, que pengen d’uns eixos llargs i especialitzats, suspeses ben lluny de la capçada de l’arbre de manera que siguin ben accessibles des de l’aire. Les bananes (Musa), de la família de les musàcies, també produeixen una inflorescència pèndula per facilitar la pol·linització pels rats-penats.
A la selva plujosa de les costes del golf de Guinea, cap al crepuscle de la seva primera i única nit d’activitat, la inflorescència de Parkia comença a regalimar dolç nèctar per la part superior, flux que s’intensifica a mesura que cau la nit. Primer els nectarínids i les abelles encara en poden prendre una mica; el seu sucre és una font d’energia ben d’agrair de cara a la llarga nit que s’apropa, però ben aviat la foscor els allunya i els substitueixen les formigues nocturnes que cerquen aliment, i de vegades també petits rosegadors com ara els lirons (Graphiurus) o primats com el potto (Perodicticus potto). Malgrat que les inflorescències de Parkia, estiguin perillosament exposades als atacs dels rapinyaires nocturns, predadors seus, aquests mamífers se senten temptats ocasionalment a completar la seva alimentació habitual amb nèctar.
Finalment arriba un rat-penat que ha recordat el camí que porta a l’arbre on s’ha alimentat un dia precedent o que ha estat orientat per l’olor dolçament mescada de la Parkia. Després de voletejar una mica al voltant de l’arbre, el rat-penat acaba posant-se sobre la massa de flors que pengen i el seu cos, també penjant, oscil·la violentament a causa del pes. Penjant cap per avall de la inflorescència, el rat-penat comença a llepar golut les dolces gotetes mentre s’arrossega maldestrament per entre les flors, refregant el seu pit contra els petits flòsculs. Mentre fa això, el pol·len d’altres arbres de Parkia adherits a la seva pell és rebut per les petites flors femenines que així es fecunden. Quan el rat-penat finalment abandona la inflorescència aponcellada, deixa darrere seu uns quants ovaris de Parkia i s’inicia el procés de formació de llavors.
Quan apunta el dia, la inflorescència ja ha complert el seu propòsit, tot i que encara pot atreure algunes papallones i abelles per a alimentar-se dels grans de pol·len i del nèctar romanent. En arribar la llum del dia, les flors es marceixen, i en els dies que segueixen cau tota la massa de flors excepte els petits flòsculs fertilitzats. Ben aviat, a l’extrem dels llargs eixos de les capçades de Parkia només hi haurà un pàl·lid botó que porta uns sacs embrionaris en ràpid creixement. Aquests s’allargaran fins que finalment formin unes tavelles d’uns 60 cm de llargària, cada una amb una fila de llavors grans i aplanades, conegudes en alguns llocs com “bajoques de llagost”. Durant unes setmanes, aquestes tavelles pengen en uns rams que semblen mans deformes amb una colla de dits, i esperen madurar.
La compensació alimentària
Induir els animals pol·linitzadors a fer la seva feina de manera eficaç i evitar qualsevol interferència a l’encadenament d’esdeveniments que cal requereix una complexa adaptació. En la majoria dels casos, els animals han de ser compensats d’alguna manera per les seves visites a les flors i perquè aquestes es repeteixin. El “pagament” usual és l’aliment. Si hi ha una font de llum acceptable, les plantes produeixen sucres molt fàcilment mitjançant la fotosíntesi i els utilitzen com a moneda de canvi per seduir els seus pol·linitzadors. Així doncs, la majoria dels nèctars són, essencialment, solucions de sucre en aigua, per més que alguns també contenen aminoàcids, proteïnes, greixos i vitamines.
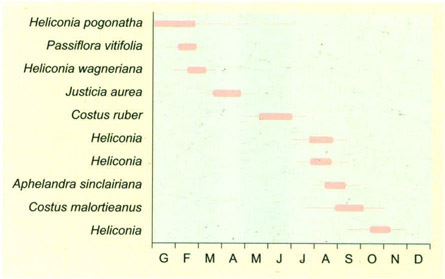
Període de floració de diverses plantes del sotabosc de la selva de terra baixa de Costa Rica que són pol·linitzades per colibrís. S’ha descobert recentment que a la selva és freqüent que les espècies que comparteixen els mateixos pollinitzadors floreixin d’una manera seqüencial. La probabilitat d’una pol·linització creuada esdevé més alta si hi ha plantes conspecífiques (per exemple, les del gènere Heliconia en el quadre) que floreixen simultàniament, i a més si les d’altres gèneres ho fan en moments diferents. Quatre de les cinc espècies d’Heliconia del gràfic comparteixen l’espècie de colibrí pol·linitzador.
Editrònica, extret de Stiles 1977
Excepte en aquells casos en què el pol·linitzador està obligat a alimentar-se de les flors d’una sola espècie (en tals casos, rars, cal que la planta floreixi ininterrompudament), les flors de cada planta han de competir entre elles i amb altres fonts d’aliment per a atreure l’atenció dels animals pol·linitzadors. Les arnes i les papallones tropicals, per exemple, sovint s’alimenten de líquids diferents que el nèctar, incloent-hi orina, suor, sang i defecacions de mamífers, i sucs de la carn en putrefacció. D’altra banda, hi ha vegades que les flors són superabundants a la selva plujosa. La necessitat d’un “suborn” competitiu exigeix que les plantes produeixin un nèctar molt nutritiu, però les solucions de sucre no poden excedir determinades concentracions sense cristal·litzar. La majoria dels nèctars que flueixen de les flors tenen per als humans un tast molt més dolç que la melassa, de manera que potser hi ha hagut una selecció per part de les plantes en el sentit d’usar sucres especials amb tast més dolç, però menys viscositat.
Els greixos contenen aproximadament el doble d’energia per molècula de sucre, i per això les gotes d’oli de nèctar ofereixen un atractiu addicional per als animals capaços d’aprofitar-los amb eficàcia. Les abelles i les mosques semblen tenir més facilitat per a fer-ho que no pas les papallones i els ocells. Cremen energia menys frenèticament i es poden permetre esperar que els olis acompleixin el seu lent procés metabòlic. De la mateixa manera, les flors que són pol·linitzades per les abelles i per les mosques solen produir un tipus de nèctar més ric de greixos que els altres, i algunes orquídies recompensen les seves abelles especialitzades únicament amb lípids.
Els aminoàcids i les proteïnes del nèctar també poden temptar insectes passavolants perquè deixin de buscar un aliment més ric en una altra banda. Algunes d’aquestes molècules són segregades directament al nèctar, però en estat de solució plantegen el problema de la cristal·lització quan la majoria d’insectes només poden prendre aliment líquid. Les flors d’algunes plantes són fetes de tal manera que és el primer insecte amb la probòscide adequada que les visita qui empeny el pol·len fins als nectaris. En altres casos, són els insectes mateixos que suquen el pol·len en nèctar i després xuclen el pollen fins deixar-lo sec; altres insectes poden menjar grans de pol·len directament.
El fascinant cas del nèctar
Un dels problemes potencials que planteja el nèctar és que es faci malbé abans que els pol·linitzadors no l’hagin trobat. Un cop exposat a l’aire, el nèctar es pot oxidar, i per a evitar-ho s’hi han d’afegir antioxidants. Malgrat que es pugui pensar que els líquids ensucrats són ràpidament colonitzats per bacteris i fongs, el cert és que la seva defensa és la mateixa dolçor. Són pocs els microorganismes que toleren una solució de sucres tan concentrada, ja que l’aigua de les seves pròpies cèl·lules resultaria extreta per osmosi. D’aquesta manera el nèctar es protegeix ell mateix, tal com passa amb l’ensucrada llet condensada, tan popular entre la gent dels tròpics perquè les llaunes obertes es poden guardar uns quants dies sense necessitat de refrigeració.
Però, com la llet condensada sobre una taula, el nèctar és vulnerable a les depredacions d’animals als quals no és destinada, sobretot de les formigues. Quan una planta ha sintetitzat sucres especials, els ha concentrat al nèctar i hi ha afegit aminoàcids, proteïnes, greixos i vitamines, no cal dir que ha hagut d’invertir una activitat metabòlica important i molts nutrients per tal de fer les seves flors atractives als ulls dels pol·linitzadors. El nèctar, doncs, és molt valuós i no s’ha de desaprofitar. Es consideren “lladres de nèctar” els animals que en prenen però són incapaços de transportar el pol·len (com per exemple les formigues sense pèls), o que ho fan de manera relativament ineficaç. Tant els uns com els altres han de ser evitats a qualsevol preu.
La major part de les flors eviten els lladres de nèctar obrint-se només durant períodes curts; així, la flor es tanca (o bé no produeix nèctar ni fa olor) a la nit o durant el dia. La pròpia estructura de la flor també pot impedir als lladres l’accés als nectaris: les corol·les llargues i tubulars, per exemple, impedeixen el pas als insectes que no tinguin la llengua adequada, mentre que barricades de pèls apuntant cap a l’entrada o altres elements similars poden protegir físicament les parts més vulnerables de la flor.
Però, precisament perquè almenys alguns insectes han de poder arribar fins al pol·len i beure el nèctar, aquests elements són intrínsecament vulnerables tot i les proteccions físiques. Per això, les plantes també utilitzen defenses químiques per allunyar els lladres de nèctar. Però el nèctar ja és un brou prou complex i, si s’hi han d’afegir elements químics protectors sense que se solidifiqui, s’ha de tractar de molècules petites. Són, doncs, més apropiades les toxines que no pas els inhibidors digestius i, de les toxines, les més utilitzades són els alcaloides. Tenen l’avantatge que són molt efectius contra els animals que no els poden metabolitzar i, per contra, són ràpidament detoxificats pels pol·linitzadors “legítims” que s’hi han adaptat.
Normalment, la defensa química dels òrgans reproductors d’una planta no es restringeix només al nèctar i al pol·len. L’estructura de la flor és important en ella mateixa, i s’ha d’evitar que els animals la facin malbé, ja sigui a causa d’una pol·linització massa efusiva com per un intent de robatori. Els tanins, efectius contra tots els animals, es dipositen sovint en grans concentracions a l’interior de les flors per aquesta raó. Un bon exemple serien les gutíferes del gènere Pentadesma, les flors grans i en forma de copa de les quals són pol·linitzades per simis.
El nèctar constitueix un aliment massa ric per als simis colobins (Colobus, Presbytis, Nasalis) perquè els interfereix la bioquímica de la digestió fermentativa als budells, però als cercopitecins els agrada d’allò més el copiós flux de nèctar de les flors de Pentadesma i van d’una flor a l’altra per beure’n a pler. A mesura que ho fan el pol·len s’adhereix a les seves barbes i va passant d’una flor a una altra. Els tanins dissuadeixen els simis, animals altrament molt destructius, de menjar-se la flor sencera; ni tan sols la mosseguen per a evitar el seu gust acerb.
La disseminació de les llavors
El mateix tipus de sistema de “pastanaga bioquímica” és utilitzat per altres plantes per a reclutar i controlar els animals que han de dispersar les seves llavors. Unes tres quartes parts dels arbres i les lianes de la selva plujosa produeixen fruits amb les llavors acompanyades d’una polpa carnosa i nutritiva, que resulta molt atractiva per als vertebrats (principalment ocells, rats-penats frugívors i simis). Els invertebrats no solen participar en la dispersió de les llavors, ja que la més petita d’aquestes resulta gran comparada amb la majoria d’ells i és difícil d’induir-los a convertir-se en portadors efectius. Després d’haver contribuït de manera molt important a la pol·linització, els insectes es converteixen en grans aniquiladors de les llavors, i aquestes s’han de protegir molt tant dels animals vertebrats com dels insectes.
Les característiques de les llavors
Les plantes necessiten dispersar les seves llavors per dues raons principals. La primera és que la producció de llavors, repetida any rere any, sol provocar una predació creixent, per tal com les poblacions predadores s’instal·len per les rodalies o són atretes de lluny estant per la magnitud del festí que esperen. Un exemple il·lustratiu seria el dels porcs barbats de Borneo (Sus barbatus barbatus), que semblen aprendre la localització dels bosquetons de bambús i roures muntanyencs que fructifiquen regularment per tornar-hi un any rere l’altre a menjar-se les glans i les llavors escampades per terra. Aquest tipus de resposta per part dels animals menjadors de llavors significa que com més a prop es trobi la llavor del seu arbre parental, o que com més temps romangui temptadora a les branques, més probabilitats hi haurà que sigui destruïda. La selecció per a evitar-ho fa que les plantes millorin la seva efectivitat per fer sortir les seves llavors de la zona de perill, ja que seran només aquestes les que aconsegueixin donar fruits al seu torn. La segona raó és que, tot i que una planta adulta productora de llavors, sens dubte viu a l’indret correcte i amb les condicions locals adequades per a la seva espècie, aquestes condicions poden canviar a causa del cicle permanent d’arbres caiguts i de regeneració de la selva plujosa. Així doncs, les llavors han de dispersar-se per a tenir l’oportunitat d’instal·lar-se en altres zones favorables, no previsibles ni en l’espai ni en el temps.
Les llavors poden arribar a uns refinaments extrems per a aconseguir ser dipositades en un indret especialment adequat per a la seva supervivència i creixement lluny de l’arbre de procedència. Un exemple clar d’això el proporcionen les lorantàcies, com els vescs (Viscum), les llavors de les quals posseeixen una capa llefiscosa que no perden ni tan sols després de passar pels budells dels ocells. Aquests, en defecar, han de refregar-se contra l’escorça de l’arbre per a lliurar-se de les llavors. Tenint en compte que les lorantàcies són plantes hemiparàsites epífites, el fet de ser dipositada a l’escletxa d’una branca resulta ideal per al creixement de la llavor.
Les plantes que utilitzen els animals per a la dispersió de les seves llavors han de fer els seus fruits atractius per a aquelles espècies que tinguin més probabilitats de distribuir-les correctament, és a dir, ben lluny i en bon estat. Només uns animals determinats s’han d’adonar de la presència dels fruits i s’han de veure recompensats per menjar-los. Els colors vius solen acomplir la primera funció; gràcies a una síntesi massiva de pigments brillants se sap que el fruit és madur i que és ple de sabors i olors que aconsegueixen que se’ls faci la boca aigua.
La polpa de la fruita madura té una funció anàloga a la del nèctar: es tracta d’una recompensa alimentària, pensada per a donar plaer, per a fer subsistir alguns animals i per a desencoratjar-ne d’altres. Les complexitats de la dispersió de llavors amb ajuda animal, però, són comparables a les de la pol·linització, i hi intervenen nombroses adaptacions i contraadaptacions. Els fruits adaptats a l’estómac dels ocells, per exemple, solen contenir metzines o productes químics nocius per als mamífers. Molts d’aquests productes produirien diarrea aguda en un simi mentre que no afecten l’ocell, solució potser beneficiosa per a les llavors amenaçades pels processos digestius dels simis.
Les característiques dels frugívors
El nombre d’animals susceptibles de dispersar llavors a la selva plujosa no és il·limitat, i tots els productors de llavors han de competir amb altres productors per obtenir les atencions dels dispersors apropiats. En termes generals, aquests darrers semblen pertànyer a dues categories diferents: els especialistes i els oportunistes. Tots dos tipus tenen necessitats diferents i, per tant, les seves activitats també tenen conseqüències diferents per a les plantes. Els especialistes frugívors, els menjadors de fruits, obtenen tots o quasi tots els sucres, els greixos i les proteïnes que necessiten de la polpa de la fruita que es mengen, mentre que els frugívors oportunistes s’interessen en els fruits bàsicament com a font de sucre i d’aigua ja que els greixos i les proteïnes els aconsegueixen menjant insectes.
Per tal d’establir una relació evolutiva íntima amb el frugívor especialista, la planta ha de presentar les seves llavors en un nutritiu paquet de polpa, cosa que té un cost important per a ella. Per això, les llavors hauran de ser tan grans com sigui possible: així tindran més reserves d’aliment i també més possibilitats d’establir-se al lloc on germinin. Les llavors grans també descoratjaran els no especialistes de menjar la fruita, ja que haurien d’omplir el seu estómac amb un llast inútil. És més, els animals petits, que no podrien empassar-se les llavors grans de cap de les maneres, tenen uns requeriments energètics diferents dels dels animals grans. Cremen energia més ràpidament i, per tant, han de menjar més aliments de digestió ràpida perquè també alliberen l’energia més ràpid. És per això que difícilment podrien permetre’s de processar els lípids i les proteïnes de la polpa de les fruites especialitzades, i de segur que no podrien afegir la despesa extra que significaria alentir el funcionament del seu estómac a causa dels bloqueigs ocasionats per les llavors grans.
Els fruits amb llavors grans i molta polpa són, doncs, del domini dels frugívors especialistes de dimensions corporals també grans. Fer aquests fruits resulta molt costós per a la planta. Per tant, se’n produeixen menys i, com a conseqüència, cada fruit té molta més importància per al futur reproductiu de l’espècie. D’altra banda, distribuint d’aquesta manera les seves inversions, la planta opta per un sistema de dispersió d’alta qualitat que implica poques llavors, però grans probabilitats d’assolir l’èxit en la dispersió.
Els frugívors especialistes als quals la planta és adaptada tendeixen a ser atents i fidels, gràcies a la generosa recompensa. Així, un elevat tant per cent dels fruits és consumit o arrencat de l’arbre abans no es podreixi o sigui aprofitable per als consumidors de llavors. Els frugívors especialistes de cos gran i intel·ligents, per exemple ocells com ara els calaus (família dels buceròtids), tenen l’avantatge addicional que abasten un territori molt extens i, per això mateix, tenen més capacitat per a distribuir les llavors a grans distàncies.
L’oferta del mercat, la demanda dels usuaris
Millorar la qualitat dels fruits no és l’única manera d’augmentar l’èxit de dispersió de les seves llavors utilitzant els especialistes que té la planta. Si hi ha pocs especialistes i, per tant, les probabilitats que els fruits (rics i valuosos) no siguin consumits resulten inacceptablement elevades, les plantes comencen a competir entre elles incrementant la qualitat nutricional de la polpa del fruit. La resta de característiques del fruit, com ara el color, el gust, la textura, l’olor i la mida també seran subjectes a una pressió selectiva semblant. És per això que la varietat de fruits en oferta als mercats tropicals és tan alta: respon a mil·lennis d’adaptació, al contrari del que passa a les àrees temperades; els fruits de la selva plujosa, certament, quasi no han sofert modificacions per part dels humans horticultors, i el seu gust i la resta dels seus atributs són encara completament naturals.
Per més que la dispersió realitzada pels frugívors especialistes sol ser prou fiable, només un volum reduït és sostingut per la petita comunitat dels animals adequats: la qualitat s’obté a costa de la quantitat. Això no sempre coincideix amb els interessos reproductius preferents de la planta, i moltes espècies de la selva plujosa adopten una estratègia alternativa basada en les activitats dels frugívors oportunistes. Si un arbre ha d’incrementar el nombre d’agents de dispersió que li siguin útils, ha de fer les llavors més petites i la polpa del fruit més ensucrada i de digestió més fàcil, a més de reforçar la coberta de les seves llavors per protegir-les dels contratemps que podria provocar un estómac oportunista massa abrasiu.
Els especialistes es veuran menys atrets per aquest fruit, però seran substituïts per una gran varietat d’animals. Els visitants, fàcilment distrets per la mateixa quantitat i varietat de fruits, seran menys fiables que els especialistes, però la quantitat total de llavors que passi pel pap del col·lectiu d’oportunistes serà igualment elevada. Cada una d’aquestes llavors s’haurà realitzat amb poca despesa i les seves reserves limitades d’aliment significaran un futur incert, però si se’n dispersa un nombre prou gran, algunes trobaran el seu camí cap a llocs apropiats per a la germinació i, amb aquestes mides, l’espècie no s’extingirà.
Els ficus (Ficus) constitueixen un exemple molt clar d’adaptació oportunista: normalment produeixen grans quantitats de fruits, cada un dels quals conté moltes llavors petites i dures. Alguns ficus produeixen diversos milions de fruits en una sola temporada i aquesta és, precisament, la seva defensa. Els fruits apareixen de sobte i el nombre de llavors amb possibilitats de germinar és prodigiós, ja que la gran quantitat de llavors produïdes evita que la planta pugui extingir-se. La producció d’aquesta mena de fruits, però, ha de ser dispersada ràpidament, si no es vol que el tret surti per la culata: cal que es desfermi una cursa contra rellotge entre els frugívors dispersors i els devoradors de llavors i lladres de fruits.
Quan el ficus de la selva plujosa produeix fruits a escala massiva és comparable a una gran cafeteria animada de dia pel voleteig i els crits dels calaus (buceròtids), dels coloms (colúmbids), dels turacs (musofàgids), dels tucans (ramfàstids), dels bulbuls (picnonòtids) i dels estornells (estúrnids), freqüentada a la nit pels rats-penats frugívors i domicili temporal de mangabeis (Cercocebus) i cercopitecs (Cercopithecus). Aquesta combinació d’oportunistes, malbaratadors i passavolants pot dispersar una proporció més petita de la producció total de llavors que la que haurien pogut dispersar els especialistes, però la quantitat de llavors en dansa és tan gran que els objectius del ficus s’acompleixen sobradament.
Les parades, l’etiqueta i les imitacions
Els fruits de la selva plujosa adaptats a grups d’animals determinats tenen trets distintius anàlegs a les síndromes de pol·linització ja esmentades. Els fruits destinats als rats-penats, per exemple, com les flors pol·linitzades per altres rats-penats, solen situar-se enfora de la capçada, sostinguts per unes branques especials que afavoreixen l’aterratge. Sovint són de colors més aviat poc vistosos, brunencs o grogosos, i tenen olors fortes, mescades. En contrast, els fruits destinats als simis i als ocells són de colors més vius, perquè destaquin entre la gamma de verds pròpia del fullam tropical i perquè resultin atractius a la vista fins i tot a distància a la llum del dia.
També aquí hi caben subtilitats i matisos. Els arbrissons dels estrats inferiors poden arribar a produir fruits específicament destinats a atreure els ocells del sotabosc, que tenen els ulls adaptats a una intensitat de llum més feble, característica del seu hàbitat. Així, rubiàcies com Lasianthus, de l’Àsia meridional, Psychotria, del Carib i de l’Àfrica occidental, o Coccocypselum, de l’Amazònia, o liliàcies com Dianella, totes elles arbustos o arbrissons del sotabosc, semblen emprar una mateixa estratègia, que consisteix a produir uns fruits d’un color blau profund. Altrament, alguns dels fruits de colors més brillants són en realitat enganyifes encaminades a encoratjar els ocells que se’ls mengen sense oferir res en canvi. La seva pellofa s’obre i es doblega, sovint fins a capgirar-se, per mostrar unes llavors cridaneres i brillants que contrasten amb el fons. Amb gran versemblança les llavors semblen estar envoltades d’una coberta carnosa, igual que la dels fruits que imiten; però en aquest cas la coberta és purament il·lusòria, una simple superfície acolorida.Dins de cada tipus de fruit, adaptat als diferents tipus d’animals, siguin simis, ocells, rats-penats, esquirols, gats mesquers o llangardaixos, sovint hi ha tot un assortit de dissenys prou diferents com perquè els animals hagin divergit segons el que mengen. Els coloms, per exemple, poden repartir-se en una extensa sèrie d’espècies diferents, cada una d’una grandària lleugerament distinta; cada una menja també fruits de mida diversa, condicionada per l’amplitud màxima de l’obertura del bec respectiu. Les peles gruixudes o les closques amb punxes poden frustrar els intents dels simis petits d’accedir al seu contingut; en aquest aspecte, els mangabeis (Cercocebus) són més forts que els cercopitecs (Cercopithecus), i els drils (Mandrillus) més que els mangabeis, cosa que té conseqüències en els fruits que trien per menjar. Però aquests grups de primats encara es poden subdividir segons l’edat i el sexe: un cercopitec mascle adult podria tenir tanta força com un dril mascle jove o com una femella adulta de mangabei, per exemple.
Els cercopitecins esmentats (cercopitecs, mangabeis i drils) es troben entre els frugívors més destacats de les selves plujoses africanes, amb equivalents a tot el Vell Món. En canvi, els colobins (còlobs, langurs, nassuts) no solen menjar fruita madura perquè la seva fisiologia digestiva no està ben adaptada a un material tan ric i ensucrat. En canvi, són importants consumidors de llavors, ja que sí que són capaços de fermentar sense problema les toxines que les protegeixen; com que se les mengen abans i tot que madurin els fruits, perjudiquen indirectament aquestes plantes, en impedir que els frugívors apropiats se’ls mengin, sencers i madurs.
Els ritmes i les sincronies
En qualsevol indret de la selva plujosa, el nombre d’arbres que estan florits, en fruit o traient noves fulles va canviant al llarg del temps. De tota manera, seria molt difícil establir un model general d’aquests canvis, ja que les nombroses espècies i individus vegetals que hi viuen segueixen cicles i ritmes de reproducció i de creixement diferents, no necessàriament sincronitzats amb esdeveniments estacionals com passa en canvi als boscos temperats. Activitat per activitat, en algunes espècies tots els individus actuen alhora, mentre que en d’altres no estan gens sincronitzats. Algunes espècies tenen cicles que tant poden ser anuals com més curts o més llargs, mentre que d’altres no en segueixen cap. Algunes espècies fructifiquen a intervals regulars, sense cap relació aparent amb el temps que fa o amb l’estació, mentre que d’altres responen a estímuls ambientals com ara la sequera.
Els cicles sense estacionalitat
Amb una observació detallada i prou llarga es poden apreciar si més no alguns models generals. El més usual a la selva plujosa és el de les plantes que treuen noves fulles dos cops l’any, amb el pic més important després del període més eixut; la floració és induïda per la manca d’aigua, de manera que els pics de floració es donen just després de l’època més seca; cinc mesos més tard, just abans que arribi l’època més humida de l’any, es produeixen els pics de la fructificació.
Aquests models generals tenen un gran impacte sobre el conjunt de la comunitat de la selva plujosa. A Borneo, per exemple, l’època de zel dels porcs barbats (Sus barbatus barbatus) coincideix amb l’època de caiguda de les flors, cosa que porta a especular amb la idea que els pètals caient com paperets de “confetti” poden ser un senyal visual al qual s’hagin adaptat els porcs. Tenint en compte que els porcells neixen uns cinc mesos després, coincidint amb el pic de fructificació, aquesta adaptació temporal sincronitza la disponibilitat d’aliment amb la demanda durant la lactància i el primer creixement dels porcells. L’abundància temporal de flors a l’estrat superior proporciona una bona font d’aliment per als rats-penats i per als insectes nectarívors, mentre que l’època de fructificació desencadena alguns canvis en el comportament dels gibons (Hylobates) i altres primats, i dels gats mesquers i els ocells frugívors, que tracten d’aprofitar la situació. Semblantment, la brotada de fulles tendres alimenta una gran varietat d’animals, que pot anar des dels simis fins a les erugues.
També es produeixen conseqüències físiques, ja que els períodes humits poden estimular la reproducció dels insectes i els amfibis que necessiten bassals per a la fresa. Aquests diferents efectes poden anar interrelacionats. Així, per exemple, molts ocells frugívors també poden menjar insectes, sobretot en circumstàncies, com la reproducció i la muda, en què els convé un aliment d’alt valor nutritiu que els ajudi a suportar l’elevat desgast que els representen aquestes activitats: si poden sincronitzar-les amb les condicions ambientals favorables als insectes, obtenen un benefici suplementari.
Per a un observador humà és temptador intentar unificar i simplificar els ritmes de la selva plujosa; però aquests parteixen d’una gran varietat d’estratègies adaptatives sovint específiques de cada espècie, i la comprensió d’això només és possible en un nivell més profund que el que cal a primera vista als ambients amb una marcada estacionalitat.
Els ritmes interns
Si considerem el comportament de la fructificació ens adonem que pagar una recompensa tan alta pels serveis dels frugívors especialistes en la dispersió de llavors de vegades no surt a compte i que hi hauria altres sortides. El fet de produir fruits permanentment seria una opció, ja que convida a l’especialització per part dels animals i també a l’ús dels fruits com a complement alimentari al llarg de tot l’any que atrauria altres especialistes quan els fruits preferits d’aquests fossin escassos. Això, però, seria una gran càrrega per a la planta, que hauria de produir fruits sempre, sense tenir en compte les fluctuacions temporals de llum, d’aigua i de disponibilitat de nutrients. De la mateixa manera, la font d’aliment fiable per a un frugívor especialista seria igualment fiable per a un consumidor de llavors, cosa que podria tenir uns resultats desastrosos per a la planta en qüestió.
La lògica dels arbres
La majoria dels arbres de la selva plujosa tendeixen, doncs, a estalviar nutrients i a produir només flors i fruits de tant en tant, segons els propis ritmes interns. D’aquesta manera, queden assegurades les reserves necessàries per a la fructificació i s’evita la manca de recursos per a la resta d’activitats, mentre la producció massiva de fruits alerta els frugívors i aclapara els consumidors de llavors, que no donen abast. Tanmateix, massa plantes fructificant al mateix temps imposen una competència innecessària per atreure frugívors especialistes, els quals poden passar fam o fins i tot morir en els períodes que separen dues fructificacions.
Per als arbres que necessiten els animals per a dispersar les seves llavors, una solució més bona és fructificar quan hi hagi el mínim possible d’altres arbres que ho facin. Això permet que els especialistes frugívors dediquin la seva atenció successivament a diferents espècies d’arbres, de manera que mai no passin fam ni les plantes quedin mancades de dispersors. Així, la majoria dels arbres i de les lianes de les selves plujoses han escalonat els períodes de fructificació al llarg de l’any de manera que no n’hi hagi ni massa ni massa pocs fructificant al mateix temps.
La sincronització de la reproducció pot ser contraproduent excepte quan el clima imposa una estacionalitat (cosa que rarament passa a les selves plujoses) o quan els fruits no són dispersats pels animals. En aquest darrer cas, és la necessitat d’evitar la predació, en comptes de la incitació dels dispersors, el que indueix uns ritmes de producció de llavors. Una de les possibles adaptacions consisteix a desbordar i afamar alternativament els predadors, i aquesta és l’estratègia dels arbres de fructificació massiva i dispersió anemòcora de la important família de les dipterocarpàcies.
El cas de les dipterocarpàcies i el porc barbat
Les dipterocarpàcies sovint són dominants als boscos de terra baixa o als pujols del sud-est asiàtic, i exerceixen una gran influència en la disponibilitat d’aliment per als animals terrestres que s’alimenten de llavors, perquè tenen la fructificació i la floració sincronitzades. Les dipterocarpàcies són molt corrents a l’illa de Borneo, on s’han realitzat estudis sobre com els animals salvatges aconsegueixen viure en aquests boscos a desgrat de la inconstància de la disponibilitat d’aliment, que han posat de relleu les conseqüències de la sincronització de la fructificació com a resposta enfront de la granivoria.
El porc barbat de Borneo (Sus barbatus barbatus) és un omnívor oportunista que consumeix arrels, fongs, invertebrats del sòl i de la fusta en descomposició, petits vertebrats, ous de tortuga, carronya de com a mínim 50 gèneres i 29 famílies de plantes. Però tot i aquesta varietat, sembla que l’element que té una influència més gran sobre la seva taxa de creixement, l’acumulació de greix i la reproducció, és la disponibilitat de fruits, mentre que la resta d’aliments habituals al bosc només permeten un creixement lent. Els fruits o les llavors de les fagàcies i les dipterocarpàcies, molt rics de lípids, són especialment importants per als porcs barbats; els roures ho són particularment perquè tenen un comportament relativament constant pel que fa a la fructificació i també perquè poden arribar a dominar els hàbitats de terra baixa. Els porcs poden sobreviure a base d’aquesta font regular d’aliment i d’altres fonts temporals, entre elles, els pics de fructificació de les dipterocarpàcies.
La fructificació sincronitzada està molt ben documentada a Sarawak, a l’illa de Borneo, perquè durant molts anys s’havien collit les llavors de les dipterocarpàcies per a exportar-les comercialment, i hi ha dades referents a l’índex de fructificació natural al bosc. Això es feia perquè les llavors contenen uns olis especials que són ingredients importants en la manufactura de peces de xocolata buides fetes amb motllo (com ara ous de Pasqua, per exemple) i altres productes. La xifra total exportada varia molt d’un any a l’altre: un terç dels anys d’ençà del 1900 ha enregistrat exportacions de menys de 10 000 t, mentre que la resta dels anys s’exportaren entre 11 000 i poc més de 28 000 t. L’índex de fructificació més important (10 000 t o més exportades al llarg de l’any) sol produir-se cada 3-5 anys, i sovint, amb una producció molt escassa o nul·la entre dos pics d’elevada producció. Aquest exemple és molt il·lustratiu de fins a quin punt varia el ritme de fructificació de les dipterocarpàcies a la selva plujosa, i alhora posa de relleu la importància en aquesta regió del bioma del problema de disponibilitat d’aliment per als mamífers que s’alimenten de llavors.
El porc barbat té unes característiques que li permeten de reaccionar vigorosament per aprofitar les oportunitats que li ofereixi la disponibilitat d’aliment i, en conseqüència, una marcada capacitat per a multiplicar-se de manera explosiva. Aquests trets, que poden ser interpretats com a adaptacions a un hàbitat amb uns recursos alimentaris distribuïts d’una manera discontínua i capriciosa, són el nombre elevat (de 4 a 12) de porcells que neixen per garrinada, un període curt de gestació (3-5 mesos) i amb la possibilitat de garrinar dues ventrades l’any, el fet de tenir el primer estre i la gravidesa a una edat tan primerenca com els 10-11 mesos, l’eficient conversió dels greixos dels aliments a greixos corporals, un elevat índex de creixement, un nombre d’individus per grup molt flexible, la sincronització dels naixements amb la caiguda dels fruits, la reducció social de l’alimentació dels mascles a favor de la de les femelles, i determinades característiques adaptades als llargs desplaçaments com ara les potes llargues i la capacitat de nedar.
Al llarg del temps, les poblacions de porcs canvien molt per immigració i emigració, esdeveniments que es correlacionen amb l’activitat de fructificació de cada petita àrea. S’interpreta que aquestes variacions reflecteixen els moviments a la recerca d’aliment dictats per l’aparició a claps i per períodes curts de fonts de fruits al si de grans àrees de la selva plujosa. Aquests canvis, però, són superats a escala regional per la variació de la talla de la població global. El 1983 hi hagué una explosió demogràfica que es repetí el 1987. Aquesta fou una repetició del que ja havia succeït a Sarawak el 1954 i el 1959. L’explicació sembla que rau en el fet que les dipterocarpàcies van fructificar massivament en una gran extensió un parell d’anys consecutius. Les xifres d’exportació dels col·lectors de llavors de dipterocarpàcies mostraren que, des del 1945, això només succeí efectivament els anys 1953-54, 1982-83 i 1986-87. A la dècada dels vuitanta, aquest esdeveniment coincidí amb les sequeres continuades a Borneo, que eren un reflex dels canvis climàtics globals. Així, es pensa que les poblacions de porcs barbats es dispararen perquè les adaptacions abans esmentades interactuaren amb la disponibilitat sostinguda de llavors oleaginoses de les dipterocarpàcies.