L’imprecís concepte de sabana
La paraula sabana és un dels primers manlleus d’una llengua ameríndia a una llengua europea. Ja apareix en la “Relació sobre les antiguitats dels indis” del monjo català fra Ramon Pané, escrita a la Hispaniola el 1498, i Gonzalo Fernández de Oviedo, a “Historia general y natural de las Indias” (1535) és el primer de donar-ne una descripció. El famós ‘cronista de Indias’ les descriu simplement com una terra sense arbres però amb molta herba, tant alta com baixa. Certament, en el context exhuberant de l’Amèrica tropical que es començava a explorar, una terra sense arbres però esponerosa d’herba mereixia un nom que la llengua de la Castella de magres pastures i terres ‘de pan llevar’ no posseïa i que hagué de manllevar del taïno, una llengua arawak que s’extingí, juntament amb els seus darrers parlants, al segle XVII, però que ha deixat innombrables manlleus al castellà i, a través seu, a moltes altres llengües.
El bioma de les sabanes és, doncs, abans de res, l’espai tropical on el triomf de l’estratègia vital de les herbes graminoides resulta absolutament palès. Allí on les pluges insuficients, els sòls massa pobres en nutrients, les roques a flor de terra, o qualsevulga condició adversa impedeixen el creixement del bosc dens però sense que l’aigua disponible sigui tan escassa que suposi una desertització. El bioma de les sabanes, tanmateix, no exclou les plantes llenyoses d’una manera absoluta: sobre un estrat herbaci dominant poden créixer mates, arbustos i fins i tot arbres, però en tot cas el seu recobriment no és tan gran com el de l’estrat herbaci definitori del bioma.
El límit entre el bosc i la sabana
Quan es fan mapes de vegetació, en els quals cal delimitar els diferents tipus de comunitats vegetals, s’estableixen de manera més o menys convencional o arbitrària separacions entre bandes o zones de vegetació. Tanmateix, les separacions entre segments contigus de la superfície terrestre no sempre resulten tan paleses sinó que la zona de contacte entre dues comunitats pot ser gradual (ecoclina) o relativament abrupta (ecotò). En la descripció de comunitats vegetals, per exemple, el principi del continuum, enunciat per l’ecòleg nord-americà Robert H. Whittaker (1924-80) el 1975, d’acord amb el qual sempre que hi hagi gradients ambientals en què la intensitat de les variables variï gradualment en l’espai, les poblacions d’espècies vegetals d’una regió varien també gradualment, sense formar discontinuïtats marcades, gaudeix d’un ampli consens. En realitat, només es donen veritables ecotons en els casos en què una variable ambiental particular varia de manera més o menys sobtada, mentre que les ecoclines corresponen a situacions en què els canvis associats a les variables ambientals es produeixen progressivament. Ambdues situacions, però, poden ser considerades com a límits o fronteres entre dues comunitats en les quals les condicions per a les plantes i els animals difereixen de manera fonamental.
Les sabanes, que ocupen prop d’un 10% de la superfície de la Terra i aporten aproximadament un 14% de la producció primària, serien un característic cas de continuum que inclouria totes les formacions tropicals amb un estrat herbaci poc o molt continu tant si tenen també un o diversos estrats de plantes llenyoses (sabanes llenyoses) com si no (sabanes herboses). Òbviament aquesta és una simplificació per tal de facilitar la unificació de textos de procedències molt diferents en els quals els especialistes usen un vocabulari molt més ric però de vegades de valor exclusivament local.
Així, dins de les sabanes llenyoses, s’inclouran tots els ecosistemes tropicals en els quals biomassa i productivitat són dominades per espècies llenyoses però sense que les capçades arribin a formar un cobricel completament tancat, permetent així l’existència d’un estrat herbaci esponerós, si més no a l’estació humida (boscos clars, sabanes boscades —incloent-hi “cerrados”, “miombos” i altres formacions similars— i sabanes arbustives). En canvi, les sabanes herboses inclouran les sabanes en la seva accepció vulgar més corrent, aquelles en què el domini de l’estrat herbaci és palès i la presència de plantes llenyoses no passa d’alguns individus aïllats (sabanes arbrades) o de petites motes de vegetació més densa (sabanes “parque”). Un criteri útil, bé que arbitrari, per a separar les unes de les altres és el de fixar-lo en un recobriment del cobricel d’un 40%, punt en el qual coincideixen sensiblement la distància mitjana entre capçades i el radi d’aquestes.
La vegetació al contacte bosc-sabana

Jordi Corbera, a partir de dades proporcionades per l’autor
Composició específica i canvis fisiognòmics de les comunitats vegetals al llarg del contacte bosc-sabana de la zona central de la Cordillera de la Costa (Veneçuela). Els inventaris corresponen a quinze transsectes paral·lels entre ells realitzats perpendicularment al gradient de vegetació sabana-ecotò-bosc perenne. Els cercles plens indiquen presència de l’espècie, i els buits, absència. A l’ecotò són presents tant algunes espècies de les dues comunitats extremes com altres d’exclusives d’aquesta comunitat de transició, com per exemple la mimosàcia arbòria Adenanthera peregrina. Els sis primers inventaris mostren una presència aclaparadorament dominant de les espècies de sabana, i una absència total de plantes ecotonals i forestals. Els cinc següents són característics de la comunitat ecotonal, ja que s’hi troben tant espècies de sabana com de bosc, bé que d’esquerra a dreta es van empobrint en les primeres i enriquint en les segones. Els quatre darrers són palesament forestals, bé que el primer dels quatre mostra encara una considerable presència d’espècies ecotonals. Els canvis fisiognòmics estan directament relacionats amb l’augment de la humitat dels sòls de la sabana, que tenen una profunditat efectiva cada vegada més gran, són menys pedregosos i presenten una proporció més elevada de partícules fines (llim) als horitzons superficials.
Les comunitats ecotonals i el procés d’arborització de la sabana
En les comunitats ecotonals, poden estar presents espècies de les dues comunitats limítrofes, però molt sovint n’hi ha un conjunt d’exclusives i característiques de l’ecotò. Tanmateix, pot resultar més útil definir el concepte d’ecotò basant-se en criteris operacionals que no per la seva composició florística. Un ecotò seria, des d’aquest punt de vista, una estreta zona de transició entre sistemes ecològics adjacents, que té un conjunt de característiques úniques definides per les escales de l’espai i el temps i per la força de les seves interaccions amb els sistemes ecològics que separa i d’aquests entre ells.
A les regions tropicals, un dels ecotons més importants és el límit bosc-sabana. El bioma de les sabanes tropicals es caracteritza per la presència de comunitats vegetals molt diverses però que tenen en comú una cobertura herbàcia contínua de gramínies i ciperàcies, la majoria de les quals presenten un metabolisme fotosintètic del tipus C4 i mostren una clara estacionalitat relacionada amb el dèficit hídric. Arbres, arbustos i palmes poden estar presents en densitats variables segons la profunditat efectiva del sòl i la disponibilitat hídrica i són ells els que, a grans trets, determinen la diferència entre les sabanes herboses i les llenyoses.
El bioma de les sabanes ocupa grans extensions de l’Amèrica del Sud, Àfrica i l’Índia i àrees més petites al sud-est asiàtic i Austràlia. S’hi manifesta un continuum des de sabanes molt seques, sobre sòls eutròfics, és a dir, rics en nutrients, no lixiviats a les vores de les zones semidesèrtiques, fins a les sabanes humides sobre sòls generalment distròfics, és a dir, deficitaris en nutrients. Tanmateix, sembla com si en el límit bosc-sabana aquell continuum s’interrompés. Es tracta d’un límit amb un accentuat dinamisme que pot romandre estable o afavorir el progrés d’una comunitat amb relació a una altra. A mesura que el nombre de mesos secs disminueix i la pluja total anual augmenta, el límit bosc-sabana es fa inestable i el bosc tendeix a avançar, a no ser que hi hagi una condició edàfica limitant com els sòls molt arenosos o els afloraments amb cuirasses ferruginoses.
En regions amb pluviositats altes en les quals hi ha àrees extenses de bosc en contacte amb la sabana, a través d’una zona ecotonal, l’absència de foc i, en menor extensió, de la pastura, pot conduir a un avenç del bosc i del recobriment dels estrats llenyosos de la sabana, per un augment en el reclutament i la supervivència de plàntules d’espècies llenyoses, tant de les de sabana com de les de bosc. Quan aquesta condició no existeix o els boscos estan molt distants, l’exclusió del foc en sabanes origina un increment gradual d’elements llenyosos provinents de l’establiment de les mateixes espècies arbòries de les sabanes. El procés d’arborització presenta una fase alentida de reclutament al llarg dels sis primers anys, per a disparar-se exponencialment després durant vint anys fins a assolir un període de saturació. En alguns casos, particularment en regions relativament humides, aquest procés pot ser autoreforçat per un mecanisme de retroalimentació positiva, a favor del procés d’arborització, sempre que no hi hagi incidència de focs com a mínim durant les dues primeres dècades.
Els tipus de sabana
S’han fet múltiples intents per tipificar les sabanes basant-se en diferents criteris, entre ells els climàtics. Des del punt de vista climàtic potser la més adequada de totes aquestes possibles classificacions és la que fou proposada per Sarmiento (1975) per a les sabanes sud-americanes. Aquest autor divideix les sabanes en tres tipus: sabanes hiperestacionals, estacionals i no estacionals, que reflecteixen com són de dràstics els canvis en el règim hídric que pateixen durant les dues èpoques contrastants de l’any.
Les sabanes hiperestacionals es desenvolupen en zones on s’acumula l’aigua de pluja o dels desbordaments dels rius. Són sòls argilosos, amb drenatge lent, que romanen inundats quasi tota l’estació plujosa, però que poden romandre secs a una profunditat de 0,5 a 1 m a causa de la presència d’una capa argilosa impermeable propera a la superfície. En començar l’època eixuta, aquests sòls es dessequen ràpidament, s’hi fan esquerdes, esdevenen molt durs i acaben patint una manca d’aigua total. A l’Amèrica del Sud, a banda de gramínies i ciperàcies, només hi creixen algunes palmeres com les “palmas llaneras” del gènere Copernicia dels “llanos” veneçolans i colombians. Si el sòl roman inundat quasi tot l’any es desenvolupa un aiguamoll permanent.
Les sabanes estacionals són les més comunes. Es troben en regions on el sòl té un bon drenatge i no hi ha acumulació d’aigua ni una capa freàtica propera a la superfície. Allà també s’hi desenvolupen importants extensions de boscos decidus. El tipus de vegetació és determinat bàsicament per les característiques del sòl. Les sabanes es troben en sòls gruixuts, ben drenats, amb poca capacitat de retenció d’aigua, pocs nutrients i una baixa capacitat d’intercanvi catiònic. Els boscos es troben en sòls més llimosos, més fèrtils, amb major capacitat de retenció d’aigua.
Les sabanes no estacionals es troben en zones de clima humit tropical Af, sense període de sequera, on la vegetació dominant són les selves plujoses tropicals. Tanmateix, localment, associats a sòls molt pobres, en sorres blanques, amb un baixíssim contingut de nutrients i una escassíssima capacitat de retenció d’aigua, també apareixen boscos esclerofil·les baixos i oberts (denominats “caatingas” al Brasil i “savanna woods” o “savanna forests” per altres autors) i sabanes no estacionals. Aquestes condicions són insuficients per a mantenir una vegetació boscosa, i les sabanes i les “caatingas” la substitueixen.
Per a les sabanes africanes i australianes, Cole (1986) ha desenvolupat una classificació similar, bé que una mica més complexa. En qualsevol cas, basar la classificació de les sabanes en criteris climàtics és només parcialment vàlid: els criteris edàfics són tan o més importants, tal com es veurà més endavant.
La sequera estacional, factor determinant: la sabana climàtica
El bioma de les sabanes abasta territoris molt amplis que envolten l’àmbit de les selves plujoses o, més rarament, s’hi intercalen. Cap límit climàtic precís, tal com ja s’ha dit, no pot ser adoptat com a frontera taxativa entre aquests dos àmbits ni, encara menys, poden diferenciar-se condicions climàtiques privatives de les sabanes. L’estacionalitat, és a dir, la successió al llarg de l’any de situacions climàtiques diferenciades que es repeteixen any rere any a la mateixa època definint unes estacions predicibles, és el tret que caracteritza el clima del bioma de les sabanes enfront del de les selves plujoses. Aquesta estacionalitat, però, varia molt d’uns punts a uns altres i, tot i que generalment s’accepta que l’existència de formacions obertes tropicals amb predomini d’un estrat herbaci requereix aquest tipus de clima, no és pròpiament aquest el factor més determinant de l’extensió ni de les característiques locals del bioma.
Els règims climàtics tropicals
Entre dos límits que segueixen aproximadament el tròpic de Càncer i el de Capricorn o latituds lleugerament més elevades s’estén un espai en què predominen els climes càlids, sense hivern tèrmic, anomenat genèricament tropical. Dins d’aquests climes es pot diferenciar tota una gamma de variants pel que fa a la pluviositat i a la humitat atmosfèrica, tant en termes absoluts com amb relació a la seva distribució al llarg de l’any. No es poden fixar uns límits precisos entre aquestes variants, sinó que hi ha una certa franja d’indefinició o de transició entre elles, com succeeix també entre el conjunt dels climes tropicals i els temperats.
Així, pel que fa al règim de pluges, arran de l’equador hi ha precipitacions abundants, però disminueixen gradualment, a mesura que hom se n’allunya; reducció, tanmateix, que no és uniforme al llarg de l’any: el clima esdevé més sec, però també més estacional, i es manifesten una època seca i una època humida ben definides. Al N de l’equador aquesta sequera és present entre desembre i març o maig, i al S, entre maig i setembre. Pel que fa al règim tèrmic, també augmenta la variació de la temperatura amb l’allunyament de l’equador. Les sabanes es localitzen en el punt mitjà de la sèrie. Sorgeixen quan la precipitació disminueix i la pluja es fa estacional, amb una estació eixuta ben definida però sense arribar a condicions subdesèrtiques o desèrtiques.
Les estacions seca i plujosa a les zones tropicals i subtropicals
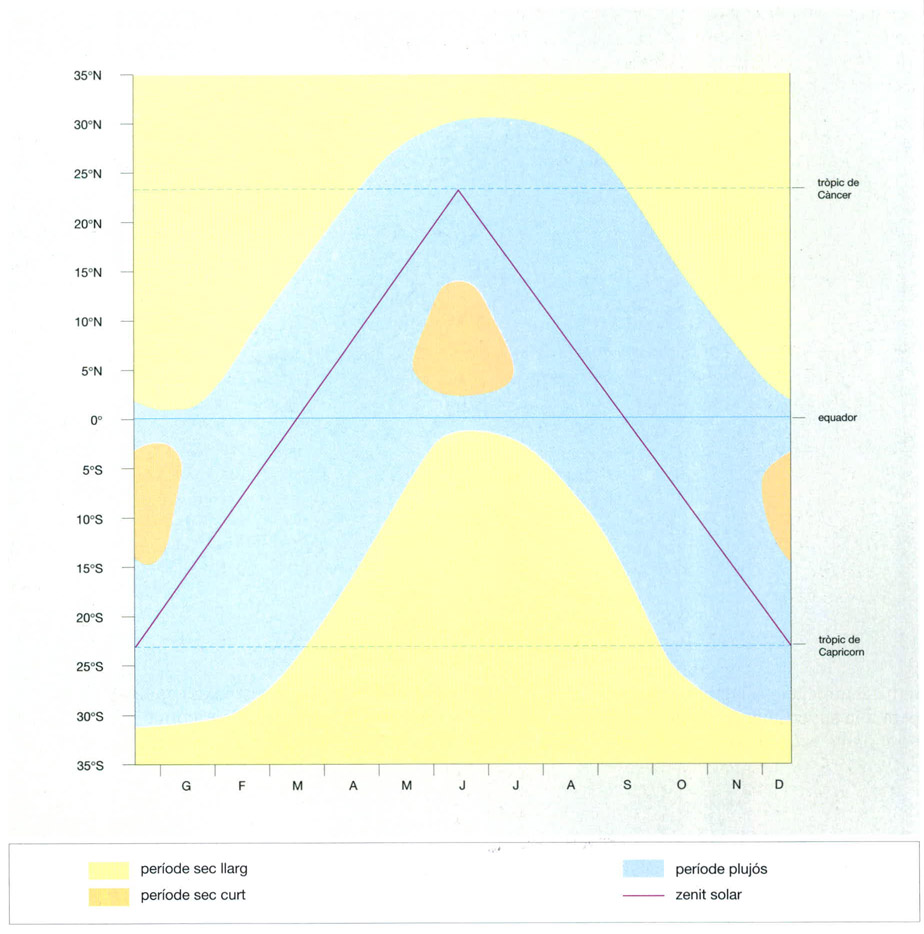
Jordi Corbera, a partir de Walter / Breckle, 1984
Repartició de les estacions seca i plujosa a les zones tropicals i subtropicals. A nivell de l’equador les pluges es reparteixen uniformement al llarg de tot l’any, de manera que no hi ha una estació seca. A mesura que hom s’apropa als tròpics, per contra, es diferencia una estació seca cada vegada més llarga. Al tròpic de Càncer les pluges es concentren de maig a setembre, mentre que en aquesta mateixa època al tròpic de Capricorn passen per l’època seca, ja que les precipitacions es donen sempre quan el Sol està al zenit al migdia. Així, el cinturó de pluges canvia segons la posició del Sol; durant l’estiu boreal, arriba fins al tròpic de Càncer, i a l’estiu austral al de Capricorn. Les dues posicions del Sol al zenit es van apropant a mesura que augmenta la distància a l’equador. Prop de l’equador, a més a més de l’època seca hivernal, hi ha un altre període sec, breu i no gaire marcat, a l’estiu, que desapareix a latituds properes als 15°. Aquests canvis climàtics graduals, a mesura que hom s’allunya de l’equador i l’època seca es va allargant, comporten canvis també en la vegetació. Les selves plujoses equatorials són reemplaçades per boscos semiperennifolis i perennifolis, i finalment per sabanes obertes.
Les regions tropicals es troben fragmentades en quatre blocs continentals (Amèrica del Sud i Mesoamèrica, Àfrica, el conjunt de l’Àsia meridional i sud-oriental i Austràlia) separades pels tres grans oceans (Atlàntic, Índic i Pacífic). Sobre aquests oceans, entre l’equador i cada un dels tròpics, se situen regularment centres anticiclònics entre els quals circula permanentment el flux dels alisis (del NE a l’hemisferi boreal i del SE a l’austral). En el veïnatge de l’equador, els alisis convergeixen en una zona de baixes pressions permanents, la zona de convergència intertropical, la localització de la qual oscil·la entre ambdós hemisferis segons la força respectiva, en cada època de l’any, dels alisis corresponents. Mai, però, no s’allunya gaire de l’equador i la intensa evaporació i la posterior precipitació de pluja que es produeix entorn d’aquesta zona de baixes pressions són determinants per a establir les condicions que permeten el desenvolupament del bioma de les selves plujoses. En canvi, en àrees més allunyades de l’equador o en aquelles en què l’amplitud de l’oscil·lació de la zona de convergència és més gran, la pluviositat esdevé més variable al llarg de l’any i s’estableix un règim de pluges estacional que és el que configura les condicions de desenvolupament dels boscos monsònics, de les formacions llenyoses obertes tropicals i de les sabanes.
Així, sobre l’equador, aproximadament entre els 5°N i els 5°S, domina l’anomenat clima equatorial humit o Af segons el sistema climàtic de Köppen (1936). Es caracteritza per unes temperatures altes i constants durant tot l’any i precipitacions elevades que molts cops superen els 2 000 mm anuals; sense cap època de sequera ni cap mes amb precipitacions inferiors als 60 mm. Una varietat de clima tropical humit però amb una estacionalitat més gran és l’anomenat clima Am, que es manifesta a les zones amb influència monsònica. En contrast extrem amb aquests climes plujosos, hi ha dues franges, centrades entorn els tròpics de Capricorn i de Càncer, on es troben els deserts més eixuts del planeta, els climes de les quals són els anomenats de subdesert (o estepa) i de desert tropicals (BSh i BWh segons el sistema de Köppen).
Entre les zones equatorials humides i les desèrtiques hi ha una franja on el clima té unes característiques intermèdies entre els dos extrems descrits. Les precipitacions són menors que a l’equador, però superen les típiques del subdesert i del desert. La temperatura és quasi uniforme durant tot l’any. Aquest clima es denomina Aw en el sistema climàtic de Köppen o també clima humit-sec tropical o de sabana. A primera vista, sembla similar al clima Am o monsònic; l’un i l’altre, per exemple, són estacionals i reben durant l’època de sequera una precipitació mensual menor de 60 mm. De fet, moltes sabanes de l’Índia i de Tailàndia es desenvolupen sota climes Am i no sota climes Aw, però es tracta de sistemes secundaris provocats per l’acció humana. Tanmateix hi ha diferències: quan la pluviositat durant els períodes de sequera és menor de 60 mm, però la precipitació total se situa per sobre d’un cert llindar (més elevat com més eixuts són els mesos de sequera) el clima es considera Am, mentre que si es troba per sota el clima serà Aw. Per exemple, a Cochin, a l’estat de Kerala, al SW de l’Índia,hi ha dos mesos que la precipitació és tan sols de 20 mm, però la precipitació anual mitjana és de 2 920 mm. És per això que es considera clarament dins la zona de climes Am. En canvi, Calabozo, a l’estat de Guárico, al centre de Veneçuela, té pràcticament tres mesos sense precipitació i un total anual de 1 300 mm, i per tant té un clima Aw clàssic. Naturalment, aquests límits són necessàriament generals i imprecisos i per això (com també per la interacció d’altres factors) poden existir sabanes sota climes Am. (Vegeu també Els tipus de clima de la Terra)
L’estacionalitat i la sabana
Un dels paràmetres fonamentals que controla la presència de les sabanes és l’estacionalitat de les precipitacions. L’estacionalitat, en efecte, determina un període prolongat d’estrès hídric a les arrels de les gramínies que condiciona la senescència de la biomassa aèria i el ritme de producció primària de dita comunitat. No obstant això, a mesura que disminueix la latitud, en les regions més tropicals la pluja total anual s’incrementa i es redueix gradualment el nombre de mesos secs. Quan aquesta supera els 2 000 mm i el període de sequera es redueix a menys de 3 mesos, apareixen successivament els boscos decidus i semidecidus tropicals i les selves plujoses. En aquestes condicions climàtiques, la presència o no de sabanes i el contacte d’aquestes amb el bosc humit tropical, depèn primordialment de les condicions edàfiques (presència o no de cuirasses ferruginoses prop de la superfície o de sòls amb alt contingut de sorra) i de la freqüència més o menys gran dels focs, sovint atribuïble a l’acció antròpica. En climes tropicals humits, en els quals el període de sequera és relativament curt, el límit bosc-sabana és inestable i afavoreix l’expansió del bosc, a no ser que es combinin l’acció periòdica del foc i les característiques edàfiques molt limitants per a l’establiment d’espècies arbòries forestals.
Tan determinant és l’estacionalitat, si més no la regular incidència d’una estació seca, per separar el bioma de les sabanes del de les selves plujoses, que no falten localitats i fins regions d’una certa extensió, sobretot a l’Àfrica, que són ocupades per sabanes o bosquets secs oberts, tot i rebre pluges que ultrapassen les que cauen en algunes regions de selva plujosa properes (més de 3 000 mm de pluja a diferents punts de les planes litorals de la república de Guinea o en algunes regions orientals del Zaire o de l’Índia). En limitar l’estació vegetativa als mesos en què hi ha aigua accessible per a les plantes al sòl, l’estació eixuta impedeix el creixement de moltes espècies pròpies de la selva plujosa que no poden suportar un estrès hídric tan prolongat.
Però, a més, la durada i la distribució en el cicle anual de les estacions seques i plujoses també pot variar considerablement al si del bioma de les sabanes, amb les consegüents repercussions en l’aspecte concret de la vegetació a cada punt del territori. Amb pluges prou abundants i un període eixut relativament breu, d’entre tres i set mesos, encara es pot donar el bosc monsònic, però així que les precipitacions dels sis mesos més plujosos resulten inferiors als 600 mm o el període de pluges s’escurça, són les sabanes llenyoses o herboses els tipus de vegetació predominants.
En general es pot dir que la pluviositat de les regions tropicals decreix a mesura que creix la distància a l’equador i que la durada del període de pluges decreix en el mateix sentit. Tanmateix, les variacions locals són nombroses al respecte i difereixen d’un continent a un altre; el sol tret comú és el fet que l’estació de pluges coincideix amb el solstici d’estiu, el període de màxima intensitat de la radiació solar (encara que, precisament a causa de la nebulositat, no coincideixi amb les temperatures anuals més elevades, sovint coincidents amb el final de l’estació eixuta). Així, per exemple, al continent africà, els boscos esclarissats de tipus “miombo” reben de 700 a 1 000 mm anuals, amb precipitacions concentrades en un període estival de pluges d’uns sis mesos de durada; als “campos cerrados” del centre del Brasil les precipitacions varien entre els 1 100 i els 2 000 mm anuals, amb un acusat eixut hivernal de tres a cinc mesos de durada; les sabanes arbrades del N d’Austràlia reben pluges inferiors, de 500 a 1 500 mm, concentrades sobretot de desembre a març; a l’Índia i a Myanmar, sotmeses al característic règim monsònic de l’Àsia meridional, les àrees de sabana reben de 800 a 1 250 mm de pluja, repartits al llarg de cinc a set mesos, amb una estació eixuta molt acusada que va de novembre-desembre fins a març-abril.
La situació geogràfica a les latituds més properes a l’equador afavoreix que el règim tèrmic tingui una estacionalitat menys marcada. La majoria de les àrees de sabana reben radiacions solars d’aproximadament entre 140 i 190 kcal/cm2 anuals amb màxims d’unes 0,5 a 0,6 kcal/cm2 diàries durant l’estació eixuta i mínims d’unes 0,3 a 0,4 kcal/cm2 durant la de pluges. Les temperatures mitjanes anuals no se situen mai per sota dels 24°C, mentre que les mitjanes de les mínimes del mes més fred se situen entre els 13 i els 18°C.
Els efectes paleoclimàtics i radiomètrics
Se sap que les glaciacions que se succeïren a les zones atemperades durant el Quaternari es traduïren a les regions tropicals de baixa altitud en climes més secs que els actuals. Aquest fenomen originà canvis marcats en els patrons regionals de la vegetació. D’aquesta manera, les selves equatorials plujoses de la conca amazònica, per exemple, quedaren limitades a les denominades àrees de refugi que, per la seva proximitat a la mar, per trobar-se en la direcció predominant dels vents carregats d’humitat i per la seva situació en determinades zones d’altitud, condicionaven un règim de pluja menys estacional que a la resta de la conca. Hom suposa que la vegetació que reemplaçà la selva equatorial plujosa fou la sabana. Els canvis climàtics no afectaren tan sols la proporció i la disposició espacial de les selves plujoses amb relació a la sabana, sinó que també quedaren reflectits en l’evolució geomorfològica dels diversos tipus de paisatges presents al tròpic. A les conques de l’Amazones i de l’Orinoco actualment existeixen petites àrees de sabanes que interrompen la matriu contínua de selves equatorials plujoses. La presència d’aquestes sota un clima no estacional es considera relictual i generalment són associades a condicions edàfiques molt limitadores per a la presència de boscos.
En una regió muntanyenca situada a la vall central de Rift, a Kenya, s’han estudiat els canvis en el límit bosc-sabana associats a l’últim canvi climàtic de l’Holocè tardà mesurant la relació 13C/12C en la matèria orgànica del sòl. Aquesta relació es considera un indicador dels canvis espacials experimentats en el límit del bosc i la sabana, atès el menor poder de discriminació isotòpica del diòxid de carboni atmosfèric per part de les gramínies C4, típiques de la sabana, comparat amb el dels arbres C3 dels boscos. La relació 13C/12C en la matèria orgànica d’una sèrie de perfils de sòl, al llarg d’un gradient altitudinal, permeté demostrar que durant l’últim període sec, a la meitat de l’Holocè, fa uns 3 400 a 3 000 anys, el límit es desplaçà 300 m més amunt de la situació actual (2 600 m sobre el nivell de la mar). Això influí en la localització dels humans pre-neolítics que, aparentment, preferien com a lloc de residència la zona ecotonal que es produeix en el contacte del bosc montà amb la sabana.
La disponibilitat de llum de les plàntules i dels exemplars joves en la zona ecotonal, altrament, ha estat identificada com un dels factors crítics que controlen la dinàmica de la sabana. Les espècies pirofítiques de sabana, fins i tot les de les sabanes llenyoses més denses, són incapaces d’establir-se i de créixer sota un cobricel tancat, mentre que els individus joves de les espècies del bosc plujós són generalment sensibles al foc i de creixement lent en les condicions més obertes de la sabana. Un dels factors que varia molt marcadament a l’ecotò és la llum, la qual en espais de pocs metres pot variar la seva intensitat, des de la que arriba al sòl en condicions obertes fins a la que s’enregistra sota el cobricel tancat de la selva plujosa.
La naturalesa dinàmica del límit del bosc plujós i la sabana llenyosa densa, dominada per Eucalyptus intermedia al NW de Queensland, origina un ecotò relativament estret entre el bosc dens, de cobricel tancat, i la sabana llenyosa relativament oberta amb un recobriment del cobricel que pot variar entre el 30 i el 70%. La transició entre les dues comunitats és acompanyada per canvis marcats en el microclima, que es reflecteixen en els gradients de temperatura, humitat, i densitat de flux fotònic. En un estudi dels canvis en l’estructura, la composició florística i el règim de llum entre la sabana d’Eucalyptus i el bosc plujós adjacent, dut a terme a la localitat de Kirrama, al NW de Queensland, s’ha observat que, durant l’estiu, la densitat de flux fotònic directe disminueix semiexponencialment a través de l’ecotò, mentre que, a l’hivern, la disminució és més lineal. Tanmateix, la disminució de la llum difusa és lineal tot l’any. El percentatge mitjà de radiació difusa varia, arran de terra, del 9,8% al bosc madur al 66,4% a la sabana, mentre que el de radiació directa ho fa del 2,9 al 38,3% en els mateixos llocs. Es trobà una correlació positiva entre la cobertura mitjana de les gramínies i la mitjana total anual de la densitat de flux fotònic a través de l’ecotò, cosa que indica la interrelació entre l’augment dels nivells de llum, la biomassa creixent de les gramínies i la susceptibilitat al foc a mesura que el recobriment i la biomassa d’aquestes augmenta a les zones més il·luminades de l’ecotò.
La sabana climàtica
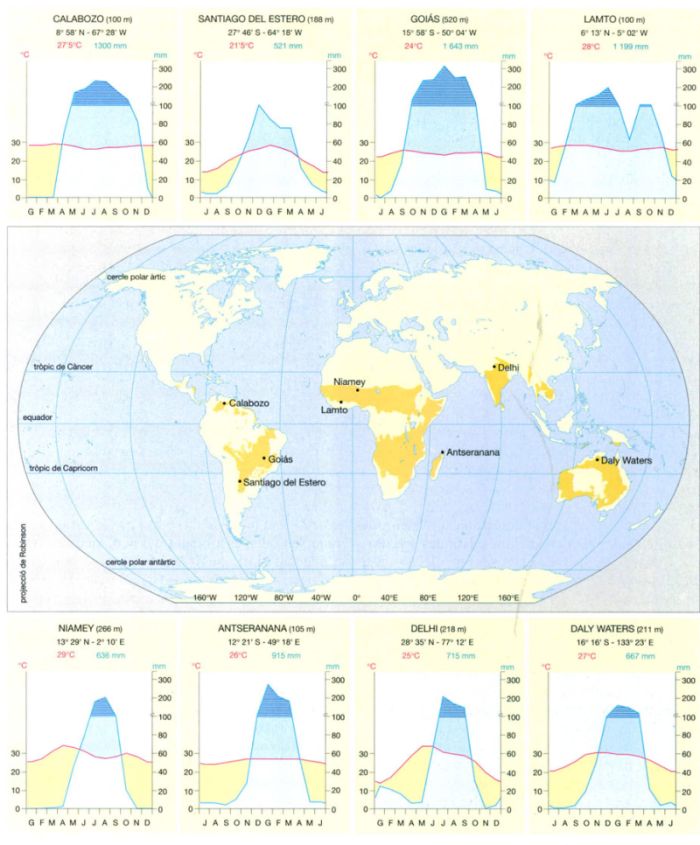
Les sabanes es distribueixen per les regions àrides i semiàrides de clima tropical i subtropical, i constitueixen un bioma adjacent al de les selves equatorials. La seva principal característica climàtica és l’alternança molt marcada d’un període sec i un d’humit, mentre la temperatura mitjana mensual es manté força constant al llarg de l’any, tal com s’aprecia als diagrames ombrotèrmics de vuit localitats representatives de les sabanes de diferents parts del món. A cada diagrama s’indiquen l’altitud, la latitud i la longitud (en negre), la temperatura mitjana (en vermell) i la precipitació mitjana anual (en blau). Les temperatures mitjanes anuals se situen entre els 21 i els 29°C, mentre que les precipitacions totals anuals prenen valors entre 500 i 1 600 mm. Les localitats situades a l’hemisferi sud tenen l’època seca al mig de l’any, mentre que a l’hemisferi nord l’estació seca comença al final d’any i es pot allargar fins al març o l’abril. D’altra banda, a les latituds més properes a l’equador, el règim tèrmic no té una estacionalitat tan marcada, tal com s’aprecia a les estacions de Calabozo, Goiás, Lamto i Antseranana, amb temperatures mitjanes mensuals que només varien uns 4 o 5°C del mes més fred al més càlid. En canvi, més lluny de l’equador, les oscil·lacions mensuals de temperatura poden arribar als 20°C. Les condicions continentals, més àrides, de les estacions de Santiago del Estero i Niamey són responsables de la pluviositat més baixa d’aquestes dues localitats. A més a més, en aquesta darrera estació les pluges es concentren en un període molt curt, ja que es troba propera al Sahel, mentre que a Lamto, al límit meridional de la sabana i tocant a la selva tropical, les pluges són abundants pràcticament durant tot l’any. Aquestes diferències en la pluviositat i la temperatura són responsables de l’aparició de formacions vegetals de sabana també molt diferents entre elles.
IDEM, a partir de fonts diverses
Les sabanes, doncs, estan generalment vinculades a climes estacionals de precipitacions mitjanes o febles, però sense arribar a les condicions d’aridesa d’un subdesert o un desert (Aw). Tanmateix, cal tenir present que les sabanes no es restringeixen a situacions amb clima Aw, car també poden aparèixer en àrees de clima humit equatorial, com passa amb les sabanes de la costa atlàntica d’Hondures i de Nicaragua, les de Trinidad, les de la costa septentrional del Brasil o les ‘illes’ de sabana al si de la selva amazònica o de la de Borneo, que apareixen en climes Af i Am. Igualment, dins de zones de clima Aw de sabana hi ha d’altres formacions vegetals com boscos decidus i boscos semiperennifolis. Així mateix, la relació sabana-clima és diferent en uns continents que en uns altres. Al continent americà no existeixen sabanes en zones amb precipitació inferior als 800 o 1 000 mm anuals i, en canvi, a l’Àfrica i a Austràlia hi ha sabanes molt seques amb precipitacions totals entre 500 i 700 mm anuals; a l’Amèrica del Sud una pluviositat que assoleixi aquests valors dóna lloc a una vegetació de bosc decidu sec o de bosquina espinosa.
Però, per què existeixen sabanes en certes parts del món i per què estan concentrades en les zones de clima Aw? No és fàcil explicar el fet que sota clima tropical es doni una vegetació natural i estable dominada per una capa contínua o quasi contínua de pastures xeromòrfiques on predominen poàcies i ciperàcies. En línies generals, els arbres, si poden establir-se en una zona determinada, haurien d’eliminar-ne competitivament aquelles plantes herbàcies amb la seva ombra i la vegetació clímax dominant seria algun tipus de bosc. Per tant, l’aparició de grans àrees de sabana només es pot justificar per l’existència de condicions locals que no permeten l’establiment dels arbres. El clima estacional Aw és una d’aquestes condicions, però ni és suficient, ni tampoc estrictament necessària en alguns casos. Hi ha boscos decidus i d’altres tipus ben desenvolupats en zones amb clima de sabana. El mecanisme, doncs, és més complex: allò que permet l’establiment i la persistència de les gramínies en algunes àrees són les seves diferències anatòmiques i fisiològiques enfront de les plantes llenyoses.
En efecte, sota un mateix clima Aw i en una mateixa zona geogràfica sovint es presenten diferents tipus de sabana. Un exemple clàssic d’això és la sèrie de “campo limpo”, “campo sujo”, “campo cerrado” i “cerradão” del Brasil; un altre seria la sèrie de sabanes de “banco”, “bajío” i “estero” de les baixes planures veneçolanes. Hi ha múltiples casos arreu del món que mostren que el que hom anomena sabana és tan sols un conjunt d’ecosistemes tropicals que tenen pràcticament una única característica comuna: la presència d’una coberta graminoide més o menys contínua. Aquests desajustaments entre el clima i la biocenosi han conduït molts autors a afirmar que la sabana no és pas el resultat d’un cert tipus de clima, sinó de la combinació d’aquest amb altres factors com el tipus de sòl, el drenatge, la freqüència de foc i les pastures. Aquesta és l’anomenada teoria holocenòtica de l’origen de la sabana.
En definitiva, a més del clima, diversos altres factors influeixen decisivament en la dinàmica del límit bosc-sabana, entre els quals es troben la llum, l’acció dels herbívors, les micorizes, la textura i la profunditat efectiva del sòl i, relacionada amb aquestes, la capacitat de reserva hídrica del substrat. Les activitats humanes afecten també aquest límit i, en la majoria de casos, afavoreixen el progrés de la sabana davant les formacions forestals tancades. En tot cas, l’acció de cap d’aquests factors no pot ser destacada en forma individual, ja que, encara que alguns, com el foc i les condicions edàfiques, poden tenir una influència més gran que la resta en el control de la dinàmica bosc-sabana, tots interactuen i cal tenir-los presents simultàniament; el que varia és el pes i la importància de cadascun en particular segons les condicions locals. No obstant això, per motius pràctics, és preferible discutir per separat la importància relativa de cada un i les respectives interaccions amb els altres a l’hora de definir i controlar el límit bosc-sabana.
Les comunitats vegetals del "cerrado"
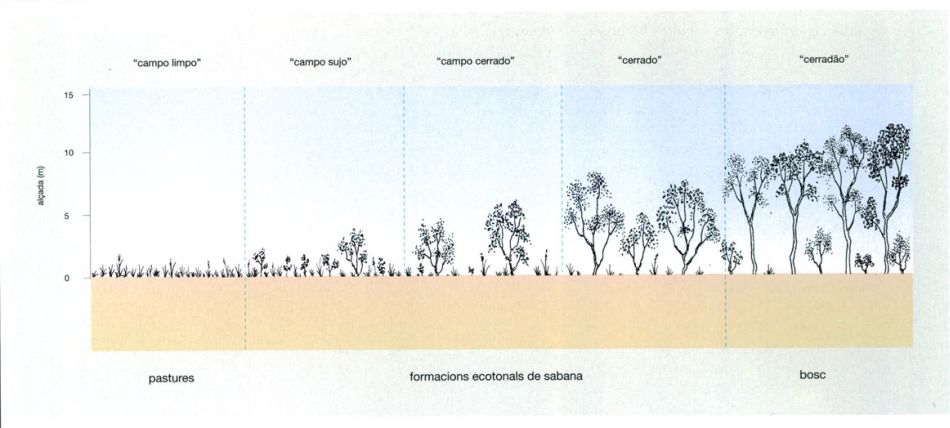
Jordi Corbera, a partir de fonts diverses
Les diverses comunitats de sabana del “cerrado” poden ser considerades, tenint en compte tant criteris florístics com fisiognòmics, formacions ecotonals entre les pastures dels “campos limpos” i el bosc xeromòrfic o “cerradão”. Si les condicions ambientals limiten l’existència del “cerradão”, apareixen formacions de sabana on predominen els components herbaci i subarbustiu, que toleren millor les condicions desfavorables. Els “campos limpos” són pastures obertes constituïdes per gramenets sense arbres, és a dir, sabanes herbàcies. El mot “limpo” significa precisament ‘net’, i es contraposa als “campos cerrados”, és a dir, amb el cobricel arbori relativament tancat. Entre uns i altres apareixen els “campos sujos”, que podrien considerar-se un tipus de sabana arbustiva, ja amb vegetació llenyosa però no tan tancats com els “campos cerrados”. El “cerrado” és la comunitat de transició entre el “campo cerrado” i el “cerradão”. Aquests diferents tipus fisiognòmics tenen una flora comuna, i entre ells existeix una continuïtat, i és per això que alguns autors utilitzen el terme “cerrado” per a referir-se indistintament a qualsevol d’ells.
Els factors geològics i edàfics: la sabana edàfica
En l’aparició i la distribució dels diferents tipus de sabana, tan importants o més que els factors climàtics ho són els relacionats amb el substrat. És per això que hom parla sovint de sabanes climàtiques i de sabanes edàfiques, per a explicar, en el segon cas, la presència de formacions de sabana en aquells indrets on per raons climàtiques potser no serien esperables.
L’efecte del substrat edàfic
Un dels factors que pot influir més decisivament en el control del límit bosc-sabana és, sens dubte, el substrat edàfic. A Veneçuela, per exemple, sota el mateix règim climàtic present en els denominats “llanos”, s’hi troben també formacions forestals, tant decídues com perennifòlies. Tanmateix, els boscos decidus estan sempre associats a sòls de menor evolució pedogenètica, originats a partir de sediments lutítics del Terciari que, pel seu origen marí o lacustre, tenen un potencial de nutrients molt més elevat, en particular pel que fa al calci. Això contrasta amb els sòls originats a partir de la formació “Mesa”, que constituí un dipòsit sedimentari d’origen terrestre, dipositat sobre aquells sediments fa més de dos milions d’anys, procedent en bona part de la meteorització de roques amb elevat contingut de quars (arenisques i granits de l’escut de Guaiana).
Aquest origen i la llarga exposició als factors pedogenètics a què ha estat sotmès després de la seva deposició, ha condicionat sòls de caràcter distròfic amb un alt contingut de sorra als horitzons superiors del sòl. El gruix d’aquesta formació és més gran en els “llanos” orientals, mentre que en els centrals i centreorientals ha estat en gran part desmantellada per l’erosió hídrica, relacionada amb l’ascens de la Serranía del Interior. L’erosió total de la formació “Mesa” i l’aflorament de substrats edàfics derivats dels sediments del Terciari, determina la presència de boscos decidus. Això no obstant, en gran part dels casos, les sabanes herboses distròfiques de Trachypogon no presenten un contacte directe amb aquests, sinó amb boscos semiperennifolis, que poden estar presents com a illes enmig d’una matriu predominant de vegetació de sabana herbosa. De fet, quan s’avança de la sabana a l’ecotò i d’aquest al bosc semiperennifoli, els sòls passen a ser relativament profunds i ben drenats, sense cuirasses ferruginoses en superfície, amb un contingut de sorra decreixent, mentre augmenta simultàniament el de partícules fines de llims i d’argiles, amb el consegüent augment de la disponibilitat d’humitat edàfica.
El límit bosc-sabana, segons estudis duts a terme en diferents localitats de la Cordillera de la Costa i de la Serranía del Interior de Veneçuela, pot ser gradual o abrupte, segons la freqüència més gran o més petita del foc i del temps passat des de l’abandó de les activitats relacionades amb l’agricultura itinerant i la ramaderia de tipus extensiu. A la majoria de localitats estudiades, les sabanes herboses amb Trachypogon com a espècie dominant estan associades a les zones més alteroses de les muntanyes, els cims i en les parts superiors o mitjanes dels vessants. A mesura que les línies de drenatge convergeixen cap a parts més baixes del vessant, la densitat dels elements llenyosos de la sabana augmenten gradualment, fins a limitar amb una comunitat ecotonal, la qual, en el cas de la secció central de la Cordillera de la Costa, és constituïda per dos estrats llenyosos: un d’arbori poc dens, dominat per la mimosàcia Adenanthera peregrina i, en menor proporció, per l’annonàcia Xylopia aromatica, i un d’arbustiu, en el qual es presenten força sovint les asteràcies Eupatorium odoratum i E. amigdalinum. A la zona frontal, més il·luminada, en contacte amb la sabana de Trachypogon, generalment hi creix un estrat graminoide, constituït per Andropogon bicornis i, en menor proporció, Panicum maximum.
L’amplada d’aquesta comunitat ecotonal és variable, però en general és estreta i no supera els 5 m. En la majoria de casos coincideix amb una disminució del pendent del vessant i una densitat més gran de línies o eixos de drenatge de caràcter intermitent. Els resultats obtinguts indiquen que el límit és controlat per l’acció combinada de foc, la profunditat efectiva del sòl, la disponibilitat d’humitat edàfica i la pedregositat superficial. A mesura que s’avança en el gradient sabana-ecotò-bosc, disminueix la pedregositat i augmenta simultàniament la profunditat efectiva del sòl, el contingut de llims i la humitat edàfica. Així mateix, es va trobar una major concentració de carboni orgànic i de fòsfor disponible, possiblement relacionats amb l’aportació més gran de virosta per part del bosc. Ambdós substrats, tant el del bosc com el de la sabana, són de caràcter distròfic (ultisòls), i sembla que la fertilitat edàfica no està involucrada en el canvi de vegetació.
S’han estudiat casos semblants en altres punts de Veneçuela, del Brasil o de Ghana. Textura, condició hídrica i profunditat efectiva del sòl comptarien entre els factors primaris que controlarien la separació entre sabanes i boscos. La presència de sòls amb una textura més fina a la banda del bosc sembla particularment determinant en molts casos.
L’efecte de la fertilitat
Encara que sovint és perceptible una fertilitat més gran en els sòls del bosc que en els de la sabana, una bona part dels estudiosos consideren que aquesta és de caràcter secundari i no determinant en el control del límit bosc-sabana. Més aviat es relacionaria amb els mecanismes de circulació de nutrients i amb la producció més gran de virosta dels boscos en comparació amb la sabana. Tanmateix, altres estudiosos han trobat resultats que interpreten en el sentit que el control del límit bosc-sabana estaria regulat pels canvis en la fertilitat edàfica.
Alguns d’aquests resultats, en contradicció amb els d’altres autors en zones semblants, indicarien que al Brasil, en el gradient de sabana herbosa (“campo limpo”), arbustiva (“campo sujo”), arbrada (“campo cerrado”), densament arbrada (“cerradão”) i bosc semiperennifoli, existeix un augment gradual i paral·lel a la fertilitat del sòl. Aquests resultats contradictoris potser podrien explicar-se per l’existència, justificable en termes de composició florística i de presència d’espècies indicadores, de dos tipus de “cerradão”, un d’ells associat a sòls mesotròfics, amb un contingut de calci més alt que el més generalitzat, present en substrats edàfics distròfics.
L’efecte de la geomorfologia
Els canvis climàtics que es produïren durant el Quaternari, juntament amb modificacions d’origen tectònic, han comportat canvis marcats en el paisatge de les regions tropicals. Aquests s’han expressat en modificacions de les formes originals del relleu i en l’escapçament dels horitzons superficials del sòl per acceleració de fenòmens d’erosió relacionats amb desequilibris del substrat. Durant els períodes pluvials del Quaternari s’han donat preferentment fenòmens de ferralitització dels sòls, amb pèrdua de silici i bases i concentració diferencial d’òxids de ferro a l’horitzó B (biostàsia) mentre que, per una altra banda, generalment coincidint amb els interpluvials secs, durant els quals la disminució de la cobertura de la vegetació ha afavorit l’erosió accelerada dels sòls (rexistàsia), s’han donat fenòmens d’encuirassament units a la destrucció dels horitzons superiors del sòl. L’acumulació gradual d’òxids de ferro in situ, per oscil·lacions anuals del nivell freàtic al llarg de períodes considerables de temps i la posterior desaparició del mal drenatge estacional és una de les vies de formació de les cuirasses ferruginoses conegudes amb el nom de laterites o plintites. Un cop ha desaparegut el nivell freàtic penjant, comença un lent i gradual procés de deshidratació dels òxids de ferro del subhoritzó mencionat, que origina la formació de laterita a una determinada profunditat en el perfil del sòl. Això es pot donar per canvis relacionats amb l’activitat tectònica (falles) i l’evolució geomorfològica del paisatge.
La presència de cuirasses ferruginoses a la superfície del sòl o uns pocs centímetres per sota dificulta considerablement la penetració de les arrels dels arbres i en fa un substrat de difícil colonització per part d’espècies forestals. A l’estació biològica de Los Llanos, a la regió de Calabozo, a Veneçuela, es presenta una vegetació de sabana herbosa de Trachypogon arbrada, interrompuda per petits claps de bosc semiperennifoli (“bosquetes”). El tipus de sòl més generalitzat a l’àrea és relativament profund i presenta textures franco-arenoses en els seus horitzons superiors; un segon tipus de sòl, caracteritzat per presentar afloraments de laterites o cuirasses ferruginoses tant en superfície com a pocs centímetres de profunditat, és menys freqüent i es presenta a claps discontinus. La vegetació de l’estació s’ha protegit dels incendis i de la pastura durant més de 25 anys, cosa que ha condicionat un procés d’arborització de la sabana herbosa tant per espècies llenyoses de la sabana com dels boscos. Tanmateix, la taxa de colonització de les espècies llenyoses i el progrés dels boscos és molt baixa en aquelles àrees on aflora la laterita o on es troba a pocs centímetres per sota de la superfície del sòl, encara que aquestes àrees són lentament i gradual colonitzades per la liana llenyosa Serjania atrolineata, de la família de les sapindàcies, que en la seva forma juvenil presenta caràcter arbustiu.
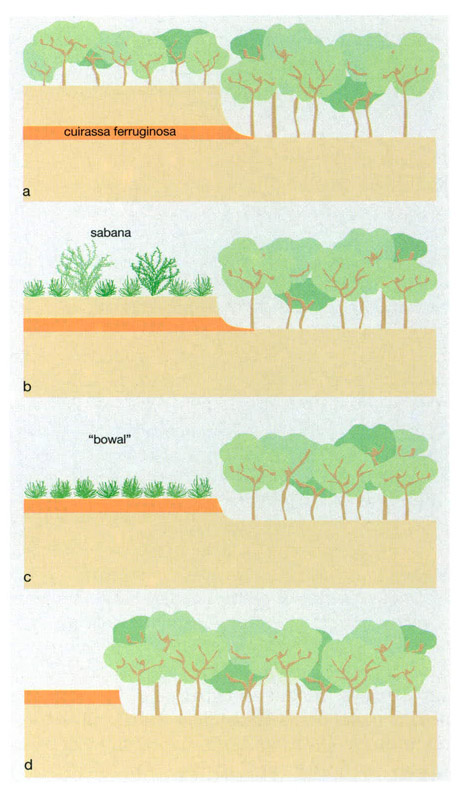
El cicle pedobotànic a les sabanes guineanes en contacte amb el bosc mostra els efectes que els factors edàfics poden tenir sobre les comunitats vegetals. En una primera etapa (a), el bosc s’estén tant per les valls com pels planells elevats; una cuirassa ferruginosa no posa cap impediment al creixement de les plantes, ja que es troba a certa profunditat. Posteriorment (b), el bosc és destruït per la intervenció humana i el substitueix una formació de sabana. La subsegüent erosió dels sòls per les activitats antròpiques (c) determina l’aflorament de la cuirassa ferruginosa, que dificulta considerablement la penetració en profunditat de les arrels dels arbres. Aleshores apareix una sabana sense arbres, anomenada “bowal”, on predominen les gramínies de caràcter xeromòrfic. Finalment l’erosió regressiva (d) del planell fa retrocedir la cuirassa, de manera que el bosc s’estén sobre l’antic “bowal”.
IDEM, a partir de Schnell, 1972
A l’Àfrica occidental, els afloraments de cuirasses a les planes amb laterites determinen un límit abrupte entre bosc i sabana. La prolongada intervenció antròpica en aquestes àrees durant milers d’anys hauria conduït a una erosió accelerada dels horitzonts superficials dels sòls. Així s’originaren canvis considerables en la vegetació: el bosc fou substituït per una sabana llenyosa a mesura que la profunditat efectiva del sòl es reduïa, i aquesta fou substituïda, al seu torn, per una sabana herbosa (denominada “bowal” a Guinea), constituïda per gramínies xeromòrfiques quan la laterita aflora a la superfície.
La cuirassa ferruginosa no és indestructible i també es meteoritza gradualment per l’acció mecànica, cosa que afavoreix la infiltració de l’aigua i la seva ressurgència en el tal·lús que limita la sabana herbosa i el bosc. Això provoca un fenomen d’erosió regressiva, que fa que hi hagi un desmantellament progressiu de la laterita i una progressiva colonització del front erosiu per part del bosc en detriment de la sabana, fenomen que completaria el cicle pedobotànic.
En general, l’acció dels agents geomorfològics que condicionen canvis en el paisatge es produeix en períodes de milers d’anys. Tanmateix, de vegades, per exemple a la regió centreoriental de l’estat veneçolà de Guárico, es presenten fenòmens d’erosió regressiva que originen la formació ràpida d’escorrancs a les planes cobertes per sabanes herboses. Els escorrancs, que s’originen a la part més baixa d’aquest paisatge, actuen un cop formats com a col·lectors d’aigües d’escolament tant superficials com subsuperficials. Les taxes d’avenç d’aquests escorrancs s’han estimat d’entre 8 i 10 m anuals, i les seves capçaleres són colonitzades per espècies llenyoses de les sabanes tals com la dil·leniàcia anomenada curatel·la (Curatella americana), la fabàcia Bowdichia virgilioides i la malpighiàcia Byrsonima crassifolia. A mesura que passa el temps, amb la millora de les condicions hídriques en eixamplar-se l’escorranc, s’estableixen gradualment espècies típiques dels boscos en galeria de la regió com la papilionàcia Andira scandia, l’annonàcia Xylopia aromatica, la cesalpiniàcia Cassia moschata o la rubiàcia Genipa caruto, entre d’altres. D’aquesta manera, en períodes de temps inferiors a cent anys, un nou bosquetó interromp la continuïtat de la vegetació de sabana herbosa original.
El decisiu paper del foc
El foc, juntament amb la presència de sòls distròfics, un clima estacional i la presència d’herbívors, es considera un dels factors fonamentals en la presència de sabanes. En general, les espècies associades a boscos semiperennifolis que limiten amb sabanes no estan dotats de mecanismes de tolerància al foc, ben a l’inrevés de les espècies llenyoses de la sabana. Així, el gruix de l’escorça de les espècies típicament forestals es mesura en millímetres, mentre que el de l’escorça de les espècies llenyoses de les sabanes pot assolir alguns centímetres i, a més, poden tenir suberificada la capa més externa.
Quan es discuteix sobre el factor foc, cal tenir en compte el tipus de material combustible, el seu contingut en humitat, la biomassa i la presència o absència de substàncies com terpens, que podrien augmentar encara més les temperatures assolides per les flames. Igualment cal considerar la freqüència dels incendis i si aquests es produeixen al principi o al final del període eixut. En general, si la sabana es crema cada any, la biomassa que s’acumula al final d’aquest no és gaire elevada. Quan no hi ha limitacions en la profunditat efectiva del sòl, la producció anual de l’estrat herbaci dependrà de la durada del període de pluges. Les temperatures que s’han mesurat en focs de sabanes varien entre els 70 i els 800°C a nivell del sòl i entre els 200 i els 800°C un metre per sobre. Unes temperatures tan elevades poden donar-se en comunitats herbàcies quan la quantitat de combustible fi excedeix les 7,80 t/ha, que és un valor força proper a la taxa anual de producció primària estimada de Hyparrhenia rufa a l’estació biològica de Los Llanos, a Calabozo, a l’estat veneçolà de Guárico.
Evolució en el temps de la coberta llenyosa en diferents tipus de sabana
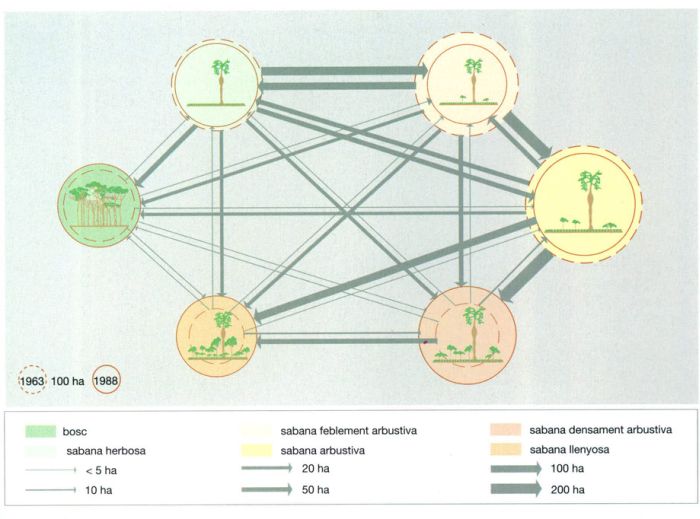
Els cercles delimitats per una línia discontínua representen la superfície en hectàrees que ocupava l’any 1963 cada tipus de sabana i el bosc semiperennifoli; les circumferències contínues indiquen el mateix per a l’any 1988. Les fletxes indiquen el pas d’un determinat tipus d’ecosistema a un altre; la seva amplada és proporcional al nombre d’hectàrees que en resultaren afectades.
IDEM, a partir de Gautier, 1990
Evolució de la coberta llenyosa a la reserva de Lamto, a la Costa d’Ivori al llarg de 25 anys, del 1963 al 1988. A l’esquema es veu clarament com la coberta llenyosa de les sabanes de Lamto augmentà considerablement durant el període esmentat. Les sabanes densament arbustiva i llenyosa triplicaren la seva extensió. El 1963, entre totes dues ocupaven una dotzena part de la superfície total de la reserva, i el 1988 ja en representaven la quarta part. Aquests dos ecosistemes i el bosc augmentaren en extensió, mentre que la sabana herbosa i les menys densament llenyoses disminuïren. La disminució és força important, sobretot, en aquestes dues darreres. La sabana arbustiva es reduí en una quarta part, i la feblement arbustiva en dues cinquenes parts. La sabana herbosa tan sols perdé una desena part de la seva superfície, bàsicament a favor del bosc, tal com indiquen les fletxes, molt més ampla la que passa de la sabana herbosa al bosc que la que marca el sentit invers. En canvi, els intercanvis amb els altres tipus de sabana, o són molt petits, o estan equilibrats. Les transferències més grans són les que es donen entre la sabana feblement arbustiva i l’arbustiva, i entre aquesta darrera i la densament arbustiva. En conjunt, durant aquests 25 anys, la coberta forestal augmentà unes 60 ha, cosa que confirma la hipòtesi que els incendis que es donen periòdicament no impedeixen la reforestació.
El foc i la sabana africana
A la regió central de la Costa d’Ivori hi ha una extensa falca de sabanes herboses amb claps de bosc semiperennifoli que penetra ben cap al S al si de la regió de selva equatorial plujosa. Sembla que deu el seu origen als canvis climàtics del Quaternari (períodes interpluvials secs), l’últim dels quals, fa 12 000 anys, conduí a l’expansió de la sabana en detriment del bosc. Actualment, tot i la millora del règim de pluges, la falca de sabanes encara es manté i alguns investigadors pensen que ha estat l’acció humana propagant el foc el que ha impedit la reforestació de la regió. La zona ecotonal és formada per una mena de sabana llenyosa que rep localment el nom de “bodga”, constituïda per un estrat, de més de 7 m d’alçada, d’arbres típics de sabana com la combretàcia Terminalia glaucescens. L’estrat arbustiu, d’entre 3 i 7 m d’alçada, és format per espècies sabanícoles com ara l’euforbiàcia Bridelia ferruginea i la cesalpiniàcia Bauhinia thonningii. Es reconeix una altra zona subarbustiva d’1 a 3 m constituïda per una barreja d’espècies sabanícoles i forestals el recobriment de les quals a la zona ecotonal depèn de la qualitat de l’ombra i de l’exposició al foc. Les espècies forestals com ara l’annonàcia Uvaria ovata o l’euforbiàcia Mallotus oppositifolius es concentren a la part interna de l’ecotò, menys exposada a l’acció del foc i amb més ombra. L’estrat graminoide està ben desenvolupat i és dominat per Pennisetum uniseta i Andropogon macrophyllus, que són particularment abundants a l’ombra dels arbustos ja mencionats. A la vora d’aquesta comunitat es troben també gramínies grans com la xisca (Imperata cylindrica) i altres espècies com ara la zingiberàcia Aframomum latifolium.
A l’estació experimental de Lamto, situada a l’extrem meridional de la falca (6°13’N; 5°02’W), s’ha estudiat l’evolució de l’estrat arbori de la sabana en parcel·les protegides de l’acció del foc durant un període de 18 anys. S’hi ha observat que, durant els primers sis anys, l’increment d’espècies llenyoses fou bastant lent, però que a partir d’aquell moment s’accelerà ràpidament i als 15 anys s’havia multiplicat per cinc el nombre d’espècies presents amb relació a les que hi havia al començament de l’experiment. La primera etapa (els primers sis anys) d’aquesta successió secundària es caracteritza per un ràpid creixement en alçada i una notable expansió de les capçades de les espècies llenyoses de sabana, amb la consegüent reducció de l’estrat graminoide per sota d’aquests bosquetons per efecte de l’ombra. La segona etapa (els sis anys següents, és a dir entre sis i dotze anys després del darrer incendi) es distingeix per un augment considerable en la riquesa d’espècies, particularment de les forestals, que veuen facilitades la seva implantació i la seva colonització dels bosquetons pel tancament del cobricel. En cloure’s aquest, els bosquetons comencen a ser freqüentats per un conjunt d’espècies d’animals, principalment ocells, rats-penats i simis que ajuden a dispersar les llavors de les espècies implantades en aquesta segona fase, el 85% de les quals presenta propàguls amb disseminació per zoocoria. La tercera etapa (dels 12 als 18 anys després del darrer foc) es caracteritza per una saturació de l’espai potencialment disponible per al reclutament de noves espècies, cosa que es pot traduir en una aturada de l’enriquiment de la biodiversitat, encara que continuï augmentant la densitat de la formació. Passats 18 anys s’observa un canvi en la jerarquia de les espècies, ja que comencen a declinar en la seva abundància relativa les espècies arbòries de sabana com ara la ja esmentada Terminalia glaucescens o la bixàcia Cochlospermum planchonii i també les espècies forestals de creixement ràpid de caràcter pioner com ara la sapindàcia Allophylus spicatus o la cesalpiniàcia Afzelia africana. Paral·lelament, es detectà un increment d’espècies més lligades en relació a la presència d’un cobricel forestal, que implica més ombra i més humitat, tals com la rubiàcia Psychotria obscura i l’eritroxilàcia Erythroxylum emarginatum.
A partir de fotografies aèries a escala 1:5 000, l’any 1990 es va fer un estudi dels canvis d’àrees que han experimentat les comunitats tant de bosc com de sabanes durant un període de 25 anys (del 1963 fins al 1988) a la regió. Els resultats indiquen que en aquest temps s’han produït canvis considerables, que es tradueixen en un augment del recobriment forestal en detriment de la sabana. Les comunitats arbòries no sols s’han expandit a partir dels boscos presents a les parts més alteroses de turons i altiplans, sinó també per un progressiu increment de les espècies llenyoses a les diferents comunitats de sabana. Aquest fenomen no solament es manifesta en les àrees protegides del foc, sinó també en aquelles en les quals hi ha focs amb certa freqüència. Cal remarcar que a les regions de sabana de l’Àfrica occidental hi ha dos règims d’incendis, ambdós relacionats amb activitats antròpiques: un de primerenc afecta el començament de l’època de sequera (entre desembre i gener), mentre que l’altre es produeix al final d’aquest període (març). Els efectes sobre l’estructura de la vegetació de l’un i l’altre són diferents, ja que la biomassa acumulada al final de l’època seca és més gran i, per tant, les temperatures assolides pel foc són també més elevades en els focs tardans. L’acció recurrent del foc en règim primerenc no ha impedit a la regió fins aquí considerada l’avenç gradual del bosc enfront de la sabana ni l’enriquiment progressiu de les diferents comunitats ecotonals en espècies llenyoses d’origen forestal i pròpies de les sabanes.
El foc i la sabana veneçolana
A l’estació biològica de Los Llanos, a Calabozo, a l’estat veneçolà de Guárico, s’ha exclòs una àrea de 390 ha de l’acció del foc i de la pastura i se n’han estudiat els canvis en l’estructura i la composició florística, tant en l’estrat graminoide com en el llenyós al llarg del temps. Durant els nou primers anys de protecció, la fisonomia de la sabana s’ha mantingut essencialment similar a les sabanes cremades anualment; tanmateix, superats 16 anys de protecció, la densitat d’elements llenyosos s’incrementà exponencialment. Al cap de 25 anys el nombre d’exemplars llenyosos a la sabana era 131 vegades més gran que al començament, mentre als boscos el factor multiplicador només era de 20 vegades. Pel que fa a l’estrat herbaci, la protecció respecte al foc i a la pastura ha anat canviant gradualment l’estructura i la composició florística de la sabana. Així, en una sabana dominada originàriament per Trachypogon plumosus començà a guanyar importància Axonopus canescens mentre que, a partir del 1977, la invasió per la gramínia africana Hyparrhenia rufa de les àrees amb sòls de profunditat efectiva més gran de l’estació ha comportat un canvi en la fisiognomia d’aquest estrat, ja que la gramínia introduïda és més alta.
L’augment en la densitat dels elements llenyosos en les sabanes de l’estació biològica de Los Llanos s’ha originat per processos similars als ja assenyalats a la Costa d’Ivori, relacionats amb la disminució de les gramínies presents en la perifèria de les petites agrupacions de plantes llenyoses de la sabana constituïdes fonamentalment per Curatella americana, Bowdichia virgilioides i Byrsonima crassifolia. En absència de foc, l’abundància i el creixement de C. americana s’han incrementat considerablement, la qual cosa determina canvis en el microclima que, units a la producció i l’acumulació de virosta, afavoreixen l’establiment, a les vores d’aquelles agrupacions, d’espècies llenyoses heliòfiles típiques de la zona ecotonal dels boscos semiperennifolis, tals com la bignoniàcia Goldmania macrocarpa, la bixàcia Cochlospermum vitifolium o la sapindàcia Allophylus occidentalis. Aquestes espècies tant poden establir-se solitàriament a l’estrat graminoide de la sabana com constituir conjuntament amb altres espècies llenyoses ecotonals tals com l’eritroxilàcia Erythroxylum orinocensis, la bignoniàcia Arrabidaea coralina, l’annonàcia Xylopia aromatica o la rubiàcia Genipa caruto, un front d’avenç del bosc enfront de la sabana. A més, en absència de foc, s’ha posat de manifest que un creixement més gran en l’alçada i en el recobriment de les agrupacions de plantes llenyoses de sabana poden millorar la fertilitat del sòl i reduir les temperatures, els nivells de llum i les taxes d’evapo-transpiració a nivell del sòl de les àrees que queden sota la seva influència i facilitar d’aquesta manera el reclutament d’espècies forestals d’ombra o esciòfiles que no poden instal·lar-se a la sabana oberta.
S’han cartografiat les agrupacions llenyoses presents a la vegetació de sabana de l’estació biològica de Los Llanos utilitzant fotografies aèries a la mateixa escala, però fetes amb 17 anys de diferència. Les dades indiquen que en aquest període d’exclusió del foc i de la pastura, les agrupacions llenyoses han augmentat considerablement tant pel que fa al nombre com al recobriment, que ha passat de l’11 al 59%. Desafortunadament, la vegetació de l’estació biològica de Los Llanos es cremà totalment l’any 1991 i parcialment l’any 1993; tot i així Hyparrhenia rufa es recuperà ràpidament a partir del banc de propàguls en el sòl i dels claps de sabana no cremats, de manera que no es va produir un canvi en la fisiognomia ni en la composició florística de la sabana. L’efecte consecutiu dels dos incendis unit a l’acumulació del material llenyós mort, junt amb l’alta producció de biomassa de la gramínia africana mencionada (8 t/ha any), produí, l’any 1993, la destrucció total d’un petit bosc dominat per Cassia moschata.
Els resultats obtinguts a les sabanes veneçolanes coincideixen amb els d’altres experiències d’exclusió del foc realitzades a l’Àfrica occidental (Nigèria i Ghana), al N d’Austràlia i al Brasil. Tots els casos indiquen que la protecció del foc afavoreix el reclutament d’espècies arbòries. Si el clima és relativament humit i la sabana es troba adjacent als boscos, l’exclusió del foc i, en menor grau, de la pastura, origina una seqüència successional de sabana cap a bosc, sempre que no hi hagi limitacions relacionades amb la profunditat efectiva del sòl. Quan el clima és més sec i no es troben boscos propers a la sabana protegida, l’augment en la densitat dels arbres prové solament del reclutament d’individus de les mateixes espècies llenyoses de la sabana.
El foc i la sabana australiana
Al N d’Austràlia, on ja no existeix una clara estacionalitat climàtica, el bosc monsònic es troba en contacte amb les sabanes d’Eucalyptus, que cobreixen grans extensions de la regió nord-occidental d’Austràlia. Un arxipèlag de petites ‘illes’ d’aquell bosc, inferiors a 5 ha, interrompen la continuïtat de les sabanes; un grup d’investigadors australians ha estudiat els factors que regulen l’establiment de plàntules d’espècies del bosc monsònic a les sabanes d’Eucalyptus que han estat protegides de l’acció del foc durant molt de temps. En aquestes es desenvolupa un sotabosc dens arbustiu que fa ombra i redueix el dèficit d’humitat del sòl. S’ha demostrat que és el canvi de microclima el que provoca l’augment de la supervivència i el reclutament de plàntules de bosc en aquesta zona ecotonal.
Alguns autors consideren poc probable que, en absència de foc, siguin la fertilitat i la humitat edàfica les causes de l’avenç del bosc a despeses de la sabana i suggereixen un possible paper dels fongs de les micorizes en l’establiment i el creixement de les plàntules d’espècies boscanes. Uns estudis duts a terme al N d’Austràlia suggereixen que l’interval de temps sense presència de foc perquè es pugui donar un avenç del bosc monsònic és d’algunes dècades; la lentitud del procés successional seria deguda a sistemes poc eficients en la dispersió de les espores d’aquests fongs i els seus requeriments d’arbres hostes. En l’actualitat, l’alta freqüència de focs tardans no controlats dificulta l’avenç del bosc monsònic, atès que aquest règim afecta el límit d’aquest bosc amb la sabana.
La influència de la fauna i dels humans
És obvi que la fauna forma part dels biomes i és incoherent, per tant, parlar de la influència dels animals sobre la sabana, perquè en són part constituent. Això no obstant, l’acció humana ha interferit molts sistemes sabànics tot afavorint la presència de determinats herbívors, de forma que aleshores té sentit considerar alguns dels elements faunístics com agents pertorbadors. En aquest sentit, els mateixos humans en són distorsionadors.
L’acció dels grans herbívors
Amb relació a l’herbivoria, resulta molt gran el contrast entre les sabanes africanes, les australianes i les de la zona tropical de l’Amèrica del Sud. Les sabanes orientals i sud-orientals d’Àfrica tenen una gran varietat de grans mamífers herbívors salvatges (i també de carnívors depredadors). A Austràlia només hi ha cangurs, que pertanyen a diverses espècies dels gèneres Macropus i Thylogale, mentre que a les sabanes de l’Amèrica tropical pràcticament només es troba el cérvol de cua blanca (Odocoileus virginianus), a les sabanes ben drenades, i la capibara, “chigüire” o “carpincho” (Hydrochaeris hydrochaeris), a les periòdicament inundables; els consumidors primaris més importants són les formigues. A més de la fauna silvestre, les sabanes també suporten molts herbívors domèstics, més nombrosos que els salvatges a Austràlia, a l’Amèrica del Sud i en algunes regions de l’Àfrica.
Els mamífers herbívors, siguin salvatges o domèstics, poden afectar la dinàmica del límit bosc-sabana, segons el nombre d’individus per unitat de superfície, de les seves preferències alimentàries (pasturadors, brostejadors o mixtos) i d’acord amb els seus hàbits socials gregaris o solitaris. El seu paper en aquesta dinàmica ha estat estudiat detalladament a l’Àfrica oriental, on s’ha observat, per exemple, que l’elefant africà (Loxodonta africana) que habita tant en boscos com en sabanes, pot afectar la dinàmica del límit de boscos baixos, bosquines semiperennifòlies i sabanes, a causa dels danys que produeixen en les espècies arbòries. L’elefant pot modificar els seus hàbits alimentaris en funció de la disponibilitat de gramínies o de fulles i fruits dels arbres, cosa que pot ser un factor de mortalitat accelerada per a aquests últims, ja que en molts casos abaten l’arbre per poder accedir a les fulles de la capçada. També poden excavar el sòl per a buscar arrels i eliminar l’escorça dels arbres, particularment la d’aquells que tenen escorces grogues o verdes, com ara diferents espècies d’Acacia o de Sterculia. L’alteració del límit bosc-sabana pels elefants i altres brostejadors com la girafa (Giraffa camelopardalis), aparentment està relacionada amb els augments en la seva densitat en àrees on els animals estan protegits de la cacera. En els últims anys s’han donat increments accelerats en la població dels elefants, per exemple, als parcs nacionals de Tsavo, a Kenya, i de Serengueti, a Tanzània.
En la interacció que es produeix entre el foc i la pastura per part d’elefants i girafes, és el primer el que té l’efecte més gran en la dinàmica de l’ecotò. L’eliminació dels arbres i l’augment dels nivells de llum certament afavoreixen l’establiment de gramínies heliòfiles, que durant l’època seca faciliten la penetració del foc, el qual impedeix la regeneració de les plàntules i inhibeix el creixement d’arbres joves. El sistema és interactiu i cíclic; la destrucció accelerada de boscos i bosquines disminueix una de les fonts de recursos alimentaris dels elefants, cosa que du com a conseqüència la reducció de les seves poblacions. Això inicia un nou cicle d’increment gradual en les poblacions d’arbres.
Els ungulats que pasturen poden influir en la dinàmica del límit pels efectes nocius que causen en les diferents etapes de regeneració de les plantes llenyoses. S’han observat episodis d’establiment d’espècies del gènere Acacia a les sabanes de Tanzània, associats a epidèmies d’àntrax que havien delmat les poblacions d’impales (Aepyceros melampus). Aquest antílop, d’hàbits alimentaris mixtos, s’alimenta de les plàntules d’acàcies. Els remugadors brostejadors, com els impales i les cabres, també poden afavorir l’augment en la densitat dels individus de les espècies arbòries dispersant les seves llavors per la sabana. Un efecte semblant ha estat observat a les sabanes pròximes a l’estació biològica de Los Llanos, que cada any es cremen, i també pasturades per herbívors introduïts com per exemple els bous (Bos taurus). El nombre d’individus adults i subadults de la cesalpiniàcia Cassia moschata per unitat de superfície, és significativament més elevat que a l’àrea adjacent de l’estació mencionada, on s’ha protegit la vegetació de l’acció del foc i de la pastura. A l’àrea on hi ha pastura i també es produeixen incendis, els individus de C. moschata tendeixen a ocupar els llocs més amagats dels bosquetons on es troben. En aquests casos, l’acció dels herbívors, a més de dispersar les llavors, també en facilita la germinació, per causa de l’escarificació química de les cobertures de les llavors, quan passen per les vies digestives d’aquests. En aquest tipus d’interacció mútua, aquelles cesalpiniàcies retribueixen els animals amb fruits dotats de llavors cobertes de polisacàrids i disacàrids de fàcil assimilació.
La pressió antròpica
Les activitats humanes també poden afectar la dinàmica del límit bosc-sabana, i molt. Principalment en promoure incendis i augmentar-ne la freqüència, en destruir la cobertura llenyosa per incrementar les àrees de pastura o per conrear la terra de manera itinerant. En aquest darrer cas, la incidència s’accentua allí on a causa del creixement demogràfic s’han hagut d’escurçar els cicles de guarets. De fet, un gran nombre de sabanes actuals tenen un origen clarament antròpic.
Tret d’alguns casos accidentals, el foc és usat pels humans a la sabana com a eina de treball. En aquests casos, les seves activitats alteren la dinàmica del límit bosc-sabana i generalment n’afavoreixen el progrés. Només a la Costa d’Ivori s’ha observat un cas en què els baules, habitants de la regió de sabana del centre-sud del país, que practiquen una agricultura itinerant de subsistència, afavoreixen l’avenç del bosc, desenvolupant les seves activitats agrícoles a la comunitat ecotonal (“bodga”), que posteriorment abandonen.
Els baules, en efecte, estableixen preferentment els seus cultius en aquesta comunitat de transició, entre el bosc semiperennifoli i la sabana densament arbrada, aparentment a causa de la fertilitat edàfica superior d’aquesta comunitat. L’artiguen, la sembren amb diverses espècies de nyam (Dioscorea) i d’altres cultius secundaris tals com el blat de moro (Zea mays), el pebrer (Piper nigrum) i el bananer (Musa) i durant tres anys consecutius l’agricultor manté lliure de males herbes l’àrea cultivada. En particular, elimina les gramínies grans, com ara la xisca, cosa que evita l’acció del foc. Un cop l’agricultor, passat aquest temps, abandona el lloc, s’hi estableix una seqüència successional, que al principi és dominada per espècies típicament ruderals, tals com la composta Mikania cordata, malvàcies com ara Urena lobata i diferents espècies del gènere Sida o algunes rubiàcies del gènere Borreria. Aquest primer estadi és substituït quatre anys més tard per un matoll secundari dens d’una alçada que no supera els quatre metres, que és format per espècies forestals del límit bosc-sabana, com la sapindàcia Paullinia pinnata o l’euforbiàcia Mallotus oppositifolius. La tercera fase es caracteritza per la implantació d’un estrat arbustiu secundari, constituït per exemplars joves de les espècies arbòries típiques de boscos, tals com la moràcia Chlorophora excelsa o l’esterculiàcia Cola gigantea. La darrera etapa és representada per un bosc baix caracteritzat per un estrat arbori, en el qual es troben Cola. gigantea, Chlorophora excelsa i l’anacardiàcia Spondias mombin.
La regió de la Gran Sabana, al SE de Veneçuela, malgrat una pluviositat anual que supera els 2 000 mm, presenta actualment una vegetació amb predomini de la sabana, bé que interrompuda per claps de bosquines i de boscos baixos i mitjans perennifolis. L’escurçament dels cicles de guaret lligat a un procés de sedentarització i instal·lació dels indígenes en petits nuclis de població fixos, ha condicionat un desequilibri en el cicle de nutrients de les comunitats forestals mitjançant la destrucció pel foc dels seus mecanismes de conservació, associats a l’acció de les arrels fines i les micorizes. Els indígenes utilitzen el foc tant en les seves activitats d’agricultura itinerant, com en algunes pràctiques de cacera. La mort de les arrels fines per l’acció del foc duu com a conseqüència la pèrdua accelerada de nutrients (calci, potassi, magnesi), alliberats per l’incendi. Tanmateix, l’oxidació de la matèria orgànica a causa del mateix agent produeix una forta pèrdua de nitrogen, tant en forma gasosa com mineral (nitrat) que, en unir-se amb l’aigua, l’acidifica i augmenta la seva capacitat de rentat. Tot aquest procés, unit a l’acció contínua del foc, va degradant la vegetació, de bosc primari perenne a bosc secundari, després a bosquina i finalment a sabana.
Uns estudis realitzats al límit bosc-sabana de la secció central de la Cordillera de la Costa, a Veneçuela, han posat de manifest que, quan es combina un material geològic parental ric en quars amb un paisatge amb grans desnivells, sòls d’escassa profunditat efectiva, distròfics, i un clima amb una forta sequera estacional, l’equilibri bosc-sabana és ràpidament desplaçat a causa de l’explotació ramadera de tipus extensiu, combinada amb l’acció del foc periòdic. L’efecte sinèrgic dels dos factors es tradueix en una ràpida erosió del sòl, mitjançant la qual l’horitzó superior és substituït per un paviment de graves més o menys fines, de naturalesa quarsosa, que provenen de la meteorització mecànica de blocs de les quarsites que afloren a la regió i de la ràpida alteració dels esquists rics en alumini i en fragments de quars. Els boscos queden relegats als llocs de menor pendent, on la confluència de línies de drenatge i l’acumulació de material edàfic més fi provinent de l’erosió dels vessants, condiciona una millora de les condicions hídriques del sòl.