Les sabanes americanes
Segurament la disponibilitat de nutrients per a les plantes és, a l’Amèrica tropical, com a la resta dels tròpics, un limitador ecològic tan important com la disponibilitat d’aigua, de manera que aquests factors es conjuguen, juntament amb el foc, per a conformar un marc de condicions ambientals que afavoreix la persistència de formacions obertes. D’altra banda, encara que la seva major expressió se situï als tròpics, a l’Amèrica del Sud les sabanes llenyoses i els boscos secs ultrapassen àmpliament les latituds tropicals, tot penetrant a les regions subtropicals i prolongant-se cap a les terres temperades del centre i el sud d’Argentina i de Xile.
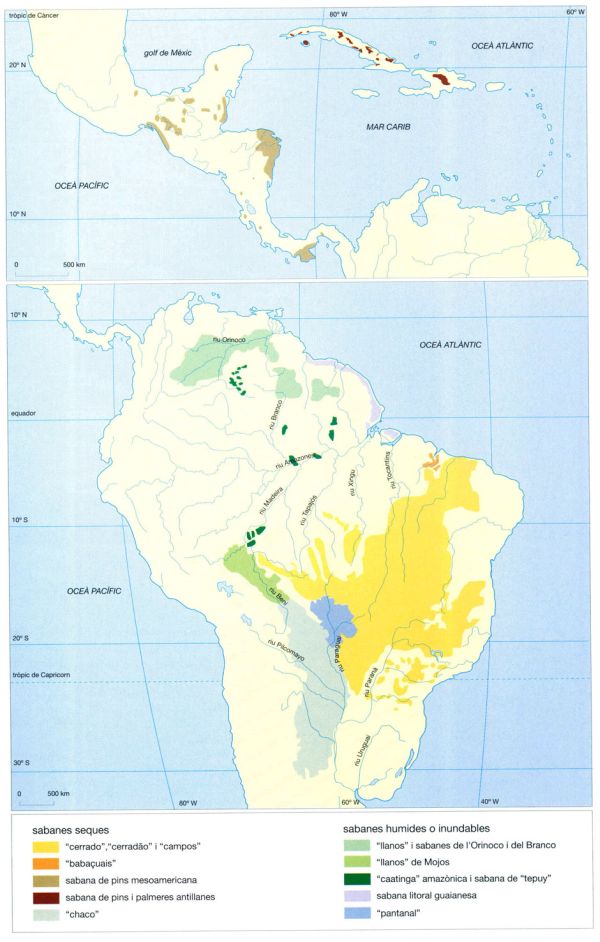
Distribució de les sabanes meso i sud-americanes. En algunes àrees, per exemple als “cerrados” del Brasil i als “llanos” de Colòmbia i Veneçuela, el seu monòton paisatge només queda interromput de tant en tant per les estretes franges dels boscos en galeria que voregen els cursos fluvials. En d’altres regions, com per exemple a l’Amazònia o a l’Amèrica Central, es dóna la situació inversa; petites clapes aïllades de vegetació de sabana s’escampen entre la coberta contínua de les selves plujoses. La distribució i les relacions de les sabanes americanes amb els factors ambientals, bé que no són senzilles, no arriben al grau de complexitat de les sabanes africanes i australianes. La composició florística és remarcablement uniforme al llarg de tota l’àrea de distribució, i existeix una continuïtat des de les sabanes arbrades, passant per les arbustives, i fins a les herbàcies. Això no vol dir, però, que cada tipus de sabana no mostri unes característiques ecològiques i fisiognòmiques pròpies. Així, es pot fer una primera distinció entre sabanes seques, certament herbàcies, però amb una sensible presència de vegetació llenyosa, que passen anualment per un període de sequera, i sabanes humides, amb un clima més humit i una estacionalitat menys marcada que afavoreix el creixement d’espècies llenyoses amb afinitats selvàtiques. Les condicions ambientals pròpies de cada regió són responsables de l’existència dels diferents subtipus dins de cadascun d’aquests dos grans grups.
IDEM, a partir de fonts diverses
Aquest ampli univers de condicions ambientals, com també la seva dispersió per tota la geografia meso i sud-americana, suggereix l’existència de molts tipus de sabanes i molt diferents, alguns relacionats amb les selves plujoses equatorials, altres, ja en els límits de la seva àrea de distribució, emparentats clarament amb els boscos i les bosquines xerofítics subtropicals i amb les estepes de climes freds. En primer lloc es consideraran les sabanes llenyoses d’ambients més humits, amb afinitats selvàtiques, a continuació les que poden ser relacionades amb diferents tipus de sabanes herboses, de les quals molts cops resulta arbitrari separar aquelles atès que apareixen formes transicionals, tal com es veurà en tractar dels “cerrados” brasilers. Finalment s’analitzaran els boscos i les boscanes (“parques”) del Chaco i les formacions transicionals amb les estepes i els prats temperats a l’Argentina i amb els boscos i les bosquines mediterranis del centre de Xile. Una característica comuna a totes, que les diferencia molt de les sabanes africanes o asiàtiques, és la pobresa relativa de la fauna de grans mamífers herbívors. El paper d’aquells grans herbívors és substituït en part a moltes sabanes americanes per invertebrats, en particular per les formigues talladores de fulles de la subfamília dels atinins.
Les “caatingas” amazòniques i els “tepuys” guaianesos
La immensa regió amazònica, coberta en la seva major part per selves plujoses, presenta nombrosos enclavaments d’ecosistemes no forestals, entre els quals destaquen aiguamolls, sabanes herboses i sabanes llenyoses. Quelcom de semblant es produeix en les selves guaianeses, que s’estenen des de la zona sud-oriental de Veneçuela fins a confondre’s amb la selva amazònica pràcticament a la mateixa desembocadura d’aquest riu gegantí. Les sabanes llenyoses ocupen hàbitats molt particulars dins d’aquest continu incommensurable de les selves plujoses.
Efectivament, sobre els materials sorrencs poc compactes, constituïts majoritàriament per quars, anomenats “arenas blancas” o “areias brancas”, la selva obre pas a formacions llenyoses baixes i obertes que han rebut diferents noms a mesura que han estat conegudes i analitzades en diferents parts de l’Amazònia. Aquestes extensions de sorres blanques, encara que mai no ocupen superfícies contínues gaire considerables, són particularment freqüents en la conca alta del riu Negro, on afloren les velles estructures geològiques de l’escut guaianès, tant en territori del Brasil, com de Veneçuela i de Colòmbia, i també caracteritzen una franja de la plana costanera de les Guaianes. Al Brasil aquest tipus de vegetació s’anomena “caatinga” amazònica o també “campinarana”, a Veneçuela es coneix amb el nom de “bana”, a Colòmbia com a “varial”, i a Guayana i Surinam es considera com un tipus particular de sabana llenyosa o com a una bosquina esclerofil·la (no té res a veure amb la “caatinga” espinosa del NE brasiler àrid). L’extrema pobresa dels sòls sorrencs o podzols amazònics sobre els quals es fa, desenvolupats sobre un substrat de quars quasi pur, comporta una vegetació estructural i florísticament pobra, amb clares reminiscències fisiognòmiques de les sabanes llenyoses del “cerrado” brasiler. Les espècies dominants són arbres o també palmes, que en els llocs de màxim desenvolupament poden assolir de 10 a 15 m d’alçada, mentre que en les formacions més baixes i obertes no ultrapassen els 6 o 8 m. L’estrat inferior, sovint molt dens, és dominat per arbustos i per plantes herbàcies de famílies de les bromeliàcies, les aràcies i altres latifoliades, però amb una escassa representació de les gramínies. En canvi, acostumen a ser abundants les molses, els líquens i les algues, que entapissen el terra i la base dels troncs. La fauna en aquests ambients queda empobrida de la mateixa manera que la vida vegetal.
Els sistemes que caracteritzen els “tepuys” guaianesos d’alguna manera s’assemblen a les “caatingas” amazòniques. Relictes estranys d’un món desaparegut, sobre aquests grans blocs tabulars de gresos pre-cambrians, erosionats i aïllats els uns dels altres, anomenats “tepuys”, han evolucionat unes biotes excepcionalment riques en endemismes, però pobres pel que fa a estructura, ja que aquests sistemes, poden assolir com a molt (segons les característiques més o menys favorables del concret substrat rocós) el port d’un bosc baix i obert encara que normalment es limiten a una bosquina, tanmateix vinculada a la selva plujosa. Una bosquina, doncs, d’una extraordinària riquesa i diversitat pel que fa als elements herbacis i subarbustius, amb predomini d’espècies de famílies com les bromeliàcies, les rapatàcies i altres de relacionades. La vegetació característica dels “tepuys” dels altiplans guaianesos assoleix la seva extensió més gran a la Guaiana veneçolana, al SE de l’estat de Bolívar, on es troben els extensos altiplans d’AuyánTepuy, Duida, Jagua, Roraima i molts d’altres, però aquests altiplans s’estenen també per les zones adjacents de Guyana, Surinam, Brasil i Guaiana francesa i penetren, bé que amb alçades força menors, enllà de l’Amazònia colombiana (Sierra del Chiribiquete, Cerro de la Campana i d’altres) per acabar en la Serranía de la Macarena, tocant els Andes.
Aquests dos tipus de formacions, les “caatingas” amazòniques i els “tepuys” dels altiplans guaianesos comparteixen, com a similaritat bàsica determinant de les seves estructures, l’extrema adversitat ecològica representada per un substrat constituït per arenes quarsítiques quasi totalment desproveïdes de nutrients, cosa a la qual cal afegir un sòl orgànic, molt àcid, en què la matèria vegetal es descompon i es recicla molt lentament. L’aigua, en canvi, mercès a l’altíssima precipitació d’aquests llocs (entre 3.500 i més de 5.000 mm anuals), no és un factor limitador encara que el sòl sigui molt permeable. Aquestes formacions tropicals sobre gresos es poden comparar basant-se en les característiques tròfiques amb una altra mena d’ecosistemes: les torberes de l’alta muntanya o de la tundra.
Els “cerradões” i “cerrados” brasilers i les formacions afins
Un tipus diferent de sabanes llenyoses, per cert molt característics del tròpic sud-americà, són els “cerradões” (singular, “cerradão”), boscos esclerofil·les baixos i oberts, típics dels altiplans de l’interior del Brasil anomenats “cerrados”. El nom portuguès de “cerrado” designa tant una regió natural molt extensa del centre del Brasil com una província biogeogràfica caracteritzada per una flora i fauna pròpies o, encara, un conjunt de formacions vegetals que mostren proporcions variades d’elements llenyosos i herbacis. Com a regió natural, els “cerrados” cobreixen gairebé dos milions de km2 de l’antiquíssim escut brasiler, principalment als estats de Minas Gerais, Bahia, Goiás i Mato Grosso.
El relleu de la zona de “cerrado” es caracteritza per altiplans profundament dissecats, que formen dilatats paisatges esgraonats amb alçades que van des dels 300 fins gairebé els 2.000 m. En alguns casos la dissecció d’aquestes antigues superfícies ha transformat les peneplanes en serralades i se les coneix per aquest nom (Serra do Espinhaco, Serra de Canastra, etc). Com a regió natural d’extensió continental, els “cerrados” s’interposen com una unitat amb personalitat pròpia entre les planes amazòniques més plujoses, les formacions seques de la “caatinga” espinosa del NE, i els boscos subtropicals meridionals on l’hivern és més marcat. Allò que confereix unitat a aquest paisatge és, a més a més del relleu, la cobertura vegetal conformada per formacions de sabana herbosa o de “cerrado” sensu lato, que abasten des de pures pastures fins a boscos oberts. Encara que es consideri la seva forma més densa (el “cerradão”) el cobricel arbori mai no es tanca del tot, cosa que permet l’arribada de llum suficient al sotabosc i, per tant, el consegüent desenvolupament d’una significativa cobertura herbàcia rica en gramínies. Per això ha estat considerat, segons diferents punts de vista, de vegades com un bosc, i de vegades com la forma més atapeïda i amb un major desenvolupament arbori d’una sabana llenyosa. Tanmateix, les semblances florístiques i faunístiques amb les sabanes circumdants és tan gran que resulta difícil comprendre el “cerradão” sense considerar-lo en tant que membre més forestal d’una seqüència de formacions sabanoides que començaria amb pastius gairebé purs (“campos limpos”), que seguiria amb altres pastius més o menys rics en arbustos i arbres baixos (“campos sujos”), seguits per les típiques sabanes llenyoses (“campos cerrados” o “cerrados” sensu stricto), per arribar a la seva màxima riquesa en elements arboris i al màxim desenvolupament de la biomassa en el “cerradão”.
La Serra do Roncador, a la divisòria entre les capçaleres dels rius Xingu i Araguaia, a l’estat brasiler de Mato Grosso, és una de les zones on s’ha estudiat més bé el “cerradão”. L’estrat arbori, constituït per arbres que rarament ultrapassen els 12 o 15 m d’alçada, té un recobriment al voltant del 50% de la superfície del sòl i permet així el desenvolupament d’un estrat de gramínies important que facilita la propagació periòdica del foc. L’existència de taques discontínues de “cerradões” en el “planalto” central brasiler ha estat interpretada de dues maneres contraposades. Una primera hipòtesi és que representen relictes d’una coberta forestal abans contínua, que l’acció humana, ja des de l’arribada dels primers pobladors i particularment com a conseqüència dels incendis sovintejats amb finalitats de cacera, reduïren a aquestes ‘illes’ de boscos degradats i oberts, immerses en una ‘mar’ de sabanes. La segona interpretació, per contra, suposa que la sabana és la formació estable en equilibri amb les condicions ambientals del centre del Brasil (clima, geologia, sòls i incendis), mentre que la seva forma més rica, el “cerradão”, tan sols prospera en alguns ambients més favorables per a la vegetació arbòria pel fet que presenten millors condicions pel que fa a humitat i a nutrients edàfics.
Els “campos cerrados”, en canvi, són sabanes llenyoses més obertes que el “cerradão”, el recobriment arbori i arbustiu de les quals assoleix valors d’entre el 10 i el 30%. La diversitat de les espècies llenyoses és molt alta i, conjuntament amb la rica flora herbàcia, configura les formacions tropicals sabanoides amb més elevada diversitat florística conegudes arreu del món. Així, per exemple, en una parcel·la protegida de “campo cerrado” dels voltants de Brasília, s’han arribat a inventariar unes 300 espècies de plantes per hectàrea. A part el nucli central del “cerrado”, aquestes formacions obertes es prolonguen cap al NE i cap al SE en una sèrie d’‘illes’ que, en el primer cas, destaquen enmig de les formacions subdesèrtiques i arbustives de la “caatinga” nord-oriental, mentre que en el segon interrompen la continuïtat original dels boscos subtropicals austrobrasilers. Així, es troben àrees insulars més o menys extenses de “cerrado” tant al NE brasiler, als estats de Bahia, de Pernambuco i fins i tot de Rio Grande do Norte, com al SE, als estats de São Paulo i de Paraná. Aquestes formacions obertes ocupen sempre altiplans ben delimitats de les planes circumdants, anomenats “chapadas” o “tabuleiros”. D’altra banda, una important extensió de sabanes herboses i llenyoses, equivalent als “cerrados” del “planalto” però a l’altre costat de l’Amazònia, s’estén per l’estat de Roraima, a les cotes més baixes de l’escut guaianès. Aquestes sabanes de la conca del riu Branco es continuen a Guyana (Rupununi) i a l’anomenada Gran Sabana, a l’estat veneçolà de Bolívar.
Els “llanos” de l’Orinoco i els de Mojos
A Veneçuela mateix, com també a Colòmbia, es troba la segona gran regió de sabanes de l’Amèrica del Sud: els “llanos” de l’Orinoco que, en conjunt, cobreixen quasi mig milió de quilòmetres quadrats. Igual que els “cerrados”, els “llanos” presenten un variat mosaic d’ecosistemes, des de boscos decidus tropicals fins a selves perennifòlies en galeria, al llarg de rius i torrents, però predominen en el paisatge els diferents ecosistemes de sabanes herboses i llenyoses, com també els “esteros” i altres tipus d’aiguamolls.
També aquí apareix el gradient de densitat i riquesa d’arbres (des de gramenets purs fins a sabanes llenyoses de tipus “cerrado”) assenyalat al “planalto” brasiler, encara que als “llanos” la diversitat d’espècies arbòries és molt menor. Alguns autors havien anomenat “chaparrales” a les sabanes amb una coberta important d’arbres baixos de diferents espècies (i fins i tot famílies) anomenats “chaparros” i, de vegades, també “paralejos”, principalment la dil·leniàcia Curatella americana, la curatel·la, però l’ús consagrat fa preferible reservar el nom de “chaparrales” a les formacions arbustives així anomenades de les mediterrànies californianes, molt diferents ecològicament a les sabanes. L’extensió més important d’aquestes sabanes arbrades apareix a la regió dels “llanos” centrals de Veneçuela, als estats de Guárico i d’Anzoátegui, així com també a la zona septentrional de l’estat de Bolívar, tot just al S de l’Orinoco. Als “llanos” colombians apareix un tipus molt particular de formació llenyosa oberta sobre sòls estacionalment inundats, que són els “saladillales”, dominats pel “saladillo” (la gutífera Caraipa llanorum), que pel seu aspecte i fisiognomia s’assembla a alguns alzinars de la conca mediterrània.
A la regió de Mojos, a Bolívia, hi ha sabanes semblants als “llanos” veneçolans i colombians. Es tracta de formacions dominades sovint per palmeres, amb una especial propensió a la inundació. Aquestes formacions són tractades amb més detall a propòsit de la reserva de biosfera de Beni.
El “chaco” subtropical
Si hom es desplaça des dels tròpics cap al subtròpic sud-americà, les formacions netament tropicals de selves humides, sabanes herboses i sabanes llenyoses són substituïdes per un univers biòtic completament diferent. Són les formacions de la regió del Chaco, la sabana progressivament embosquinada o “chaco” (terme quítxua que significa ‘lloc de cacera’), que en una extensió de quasi un milió de km2 imprimeix la seva fisiognomia a les planes subtropicals de Bolívia, Paraguai i Argentina.
El “chaco” conforma un bioma apassionant i multifacètic, dominat sovint pels vegetals espinosos. En primer lloc el caracteritzen boscos de diferent port segons les condicions climàtiques, que es tornen més seques i continentals a mesura que es creua la regió des dels rius Paraguai i Paraná cap al centre-oest de l’Argentina (s’hi atenyen temperatures de fins a 48°C). A les ribes d’aquests dos grans rius, el bosc txaqueny assoleix el port i la diversitat d’una selva subtropical (Chaco oriental humit, popularment conegut per l’eloqüent nom d’“el impenetrable”), mentre que a mesura que l’hivern es prolonga i es fa més fred i sec, el bosc s’empobreix fins a acabar convertit, al seu extrem sud-occidental, en una bosquina xermòrfica (“chaco” sec) que es confon gradualment amb el subdesert del “monte” argentí que el delimita cap als contraforts muntanyosos i les valls interiors occidentals de la base dels Andes.
Aquest gradient de formacions forestals cada cop més baixes, obertes i esclerofil·les queda interromput quan els contraforts muntanyencs (per exemple, a les Sierras Pampeanas del NW de l’Argentina) retenen la humitat i indueixen així la presència d’un bosc txaqueny més ric, l’anomenat “chaco serrano”, o quan les precipitacions assoleixen valors més elevats, que comporten l’aparició de sabanes i boscos subtropicals encara més humits. Antigament s’hi feia un bosc obert amb arbres majestuosos com la tipa (Tipuana tipu), de la família de les fabàcies, o el “timbó”, “pacará” o “oreja de negro” (Enterolobium contortisiliquum), de la de les mimosàcies, com també els boscos anomenats de transició, rics de fustes valuoses com el “cebil” (Piptadenia macrocarpa), que és una mimosàcia, la cedrela mexicana o “cedro amargo” (Cedrela mexicana), que és una meliàcia, i la juglandàcia anomenada noguera austral (Juglans australis), però l’avenç impetuós de la frontera agro-pecuària pels vessants i les parts baixes de les serres del N de l’Argentina i de Bolívia fan perillar els últims relictes d’aquestes formacions originals.
Però, a més de la seva diversitat de sistemes forestals, el “chaco” també és ric i divers en comunitats amb predomini de l’estrat herbaci: sabanes inundables, gramenets, una extraordinària diversitat d’ambients aquàtics i semiaquàtics a la seva part oriental, extenses pastures salines o alcalines en els seus confins amb la Pampa i, tant en moltes valls orientals com en algunes depressions del Chaco occidental, grans palmerars del carandaí Copernicia australis. També, en una àmplia franja de transició entre les formacions humides del Chaco oriental i les subhumides i seques de l’occidental, apareix, al Chaco central, una particular fisiognomia de parc que recorda molt les sabanes herboses estacionals dels “cerrados” o dels “llanos”, però amb una flora i una fauna completament diferents. La porció nord-occidental del Chaco, finalment, presentava fins fa poques dècades un mosaic de pastures i sabanes subtropicals on s’establí una ramaderia bovina que superà la capacitat de càrrega d’aquests sistemes herbacis i n’induí ràpidament l’embosquinament secundari amb espècies arbustives espinoses, de les quals el “viñal” (Prosopis ruscifolia) constitueix l’exemple més clar i més trist. Aquestes bosquines espinoses quasi monospecífiques, anomenades “viñalares”, ja que són dominades per aquesta mimosàcia, no tenen cap interès econòmic, sigui forestal o pastoral, i la seva extensió constitueix un exemple palès de les conseqüències negatives de l’ús no sostenible dels recursos naturals.
Cap als seus límits meridionals, les formacions txaquenyes s’empobreixen gradualment d’elements llenyosos amb el consegüent augment d’importància de l’estrat herbaci, adquirint així un aspecte de sabana. Aquestes formacions de gramenets amb arbres baixos, sovint espinosos, han estat anomenats a l’Argentina “parque mesopotámico” o “espinal” i envolten com un gran semicercle les praderies de la pampa. Entre els arbres més característics de l’espinal s’ha de mencionar el “caldén” (Prosopis caldenia), que es desenvolupa en sòls arenosos de les províncies argentines de Córdoba i de San Luis i que ha estat explotat per la qualitat de la seva fusta. També són abundants altres espècies de mimosàcies del mateix gènere Prosopis i del gènere Acacia, però tenen menor interès econòmic ja que són de menor port.
Les formacions sabanoides del Con Sud
A la zona temperada de Xile central, sota un clima mediterrani de pluges hivernals, el típic bosc esclerofil·le original d’aquesta regió climàtica ha estat reemplaçat per l’acció humana per sabanes molt similars a l’“espinal” argentí, però on les gramínies dominants són en la seva major part espècies anuals i l’“espino” que les caracteritza és Acacia caven. La pastura abusiva i els incendis freqüents han estabilitzat formacions secundàries de bosquines on es conserven algunes espècies del bosc original però dins d’una estructura vegetal molt més baixa i oberta, similar a la vegetació primària que es pot donar sota climes més secs.
Ja a la regió temperada-freda dels Andes australs a Xile i a l’Argentina es troben altres formacions llenyoses obertes que conformen amplis ecotons entre les estepes seques i els boscos humits de faigs australs (Nothofagus). Entre les formacions obertes de tipus sabanoide més interessants d’aquestes àrees es poden destacar els espectaculars boscos de coníferes australs, com els de “pehuen” (Araucaria araucana), i els més oberts de xiprer xilè (Libocedrus chilensis). Les dues formacions apareixen com a l’última avançada dels arbres sobre les estepes patagòniques.
Les sabanes de Mesoamèrica i del Carib
A la regió de l’istme centramericà i a les illes del Carib, les sabanes llenyoses no assoleixen la importància que tenen a l’Amèrica del Sud, ja que en aquesta regió no són tan freqüents alguns dels ambients típics on prosperen aquests ecosistemes. Els principals tipus de formacions obertes en aquesta regió són, per una banda, les sabanes herboses i, per una altra, les pinedes, que es podrien considerar com un tipus molt particular de sabana. S’han descrit sabanes a Mèxic, a Cuba i a Costa Rica, on formen nombroses taques de vegetació oberta encara que en superfícies limitades. Són similars en composició, estructura i ecologia a les sabanes estacionals d’Amèrica del Sud, amb espècies dominants com la ja esmentada curatel·la (Curatella americana) i el “nancé” (Byrsonima crassifolia), un altre “chaparro” o “peralejo”, l’un i l’altre d’àmplia distribució a totes les sabanes neotropicals. A aquestes s’afegeixen, a l’estrat dominant, algunes espècies d’arbres de llinatge boreal com ara alzines (Quercus) i pins (Pinus). Les pinedes són formacions molt característiques a determinats tipus d’hàbitats i arriben a cobrir àrees relativament extenses a Belize, Hondures, Nicaragua, Cuba i Santo Domingo. Aquestes formacions obertes estan generalment associades a substrats sorrencs, com a la Misquitia de la costa atlàntica a banda i banda de la frontera que separa Hondures i Nicaragua, o fins i tot a dipòsits d’arenes blanques, molt similars als descrits per a les “caatingas” amazòniques, com passa en el cas de Cuba. A Belize es troben pinedes de pi d’Hondures (Pinus caribaea var. hondurensis) en sòls arenosos però amb un subsòl impermeable, tant a la costa com als turons més cap a l’interior. En realitat, en aquestes àrees es troben mosaics de formacions obertes que inclouen des de pinedes fins a alzinars de Quercus oleoides, sabanes arbrades i aiguamolls amb palmes.
Les anomenades Sabanas de Misquitos són pinedes de pi cubà mascle (Pinus caribaea) que s’estenen formant una franja d’uns 50 km d’amplada pel litoral del Carib als dos costats de la frontera entre Hondures i Nicaragua. Només apareixen sobre materials grollers-graves i sorres del Plistocè inferior que s’estenen entre el nivell de la mar i una altitud de 50 m. Les pinedes ocupen els sòls alts i ben drenats mentre que en els sòls baixos i inundables els substitueixen sabanes i aiguamolls de diferent naturalesa. El límit amb les selves altes que continuen cap a l’interior és ben net, tal com correspon a un límit geològic i edàfic precís, ja que tan bon punt el substrat arenós és desplaçat per materials de textures menys grolleres, la selva desplaça les pinedes. Aquestes formacions tropicals dominades per coníferes estan emparentades amb les pinedes de Louisiana o de Florida, al SE dels Estats Units, i, com aquelles, també són periòdicament escombrades per incendis de sotabosc, que afecten poc els pins adults però incideixen en la renovació del poblament. El pi ha estat molt explotat comercialment però es regenera naturalment assegurant el manteniment del sistema, a densitats més baixes que les originals, en un fenomen de ‘sabanització’ de l’ecosistema original.
A Cuba i a l’illa de Pinos també apareixen pinedes de pi cubà mascle (Pinus caribaea var. caribaea) sobre substrats sorrencs. Així, per exemple, a la província de Pinar del Río, a l’W de l’illa de Cuba, una formació llenyosa oberta caracteritza els dipòsits d’arenes quarsítiques, sobre els quals es forma una vegetació baixa d’on sobresurten els pins cubans mascles (P. caribaea var. caribaea) i els pins cubans femelles (P. tropicalis), conjuntament amb diverses espècies de palmes i un sotabosc arbustiu relativament diversificat. El que crida l’atenció d’aquestes formacions és l’escassesa de gramínies i l’accentuat esclerofil·lisme de quasi totes les espècies, un altre cas notable d’escleromorfil·lisme causat per la limitació de nutrients. Unes altres formacions llenyoses obertes que a Cuba ocupen hàbitats molt específics són les que apareixen sobre afloraments de serpentines, on es desenvolupen sòlsrics en metalls pesants tòxics per a la majoria de les plantes, i les que caracteritzen els sòls recoberts per cuirasses laterítiques més o menys desmantellades. En tots dos casos es tracta de formacions obertes d’arbres baixos i amb una gran diversitat de palmes. A Santo Domingo, sabanes arbrades amb pins de l’espècie P. occidentalis o amb els típics arbrissons (“chaparros” o “peralejos”) de les sabanes neotropicals, com Byrsonima o Curatella, cobreixen un relleu relictual d’altiplans molt dissecats. També a moltes de les Antilles menors existeixen comunitats semblants, encara que en aquestes illes no hi ha arribat cap espècie de pi.
Les sabanes africanes i malgaixes
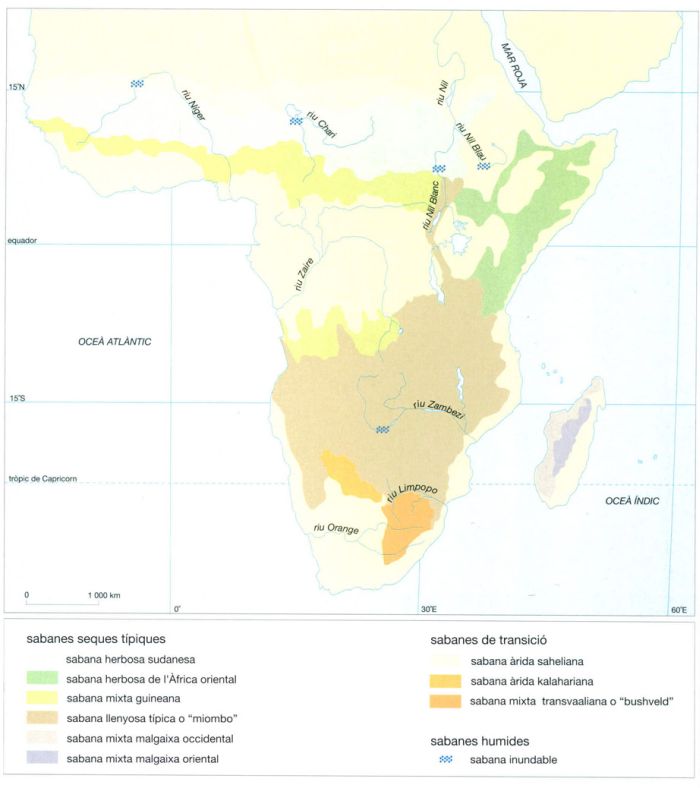
Distribució de les sabanes africanes continentals i malgaixes. A l’Àfrica, les diferents comunitats vegetals de sabana es distribueixen segons un model força complex, resultat tant dels canvis climàtics importants que sofriren les terres d’aquest continent, com d’una història geomorfològica també força complexa que donà lloc a un relleu i uns sòls molt diversificats. En conseqüència, el continent africà es caracteritza per una gran diversitat de sabanes, principalment arbustives i arbrades, de composició florística molt diferent. A més a més, les extenses àrees de selva plujosa de la zona central del continent separen les sabanes més septentrionals de les més meridionals, introduint un nou factor de diversificació. Si bé hi ha espècies vegetals que apareixen a totes les sabanes del continent, altres presenten una distribució unipolar, és a dir, es troben o bé al N o bé al S del cinturó de selves tropicals. Els canvis fisiogràfics i climàtics, que en temps geològics recents (des de principi del Quaternari) tingueren lloc a l’Àfrica oriental, marcaren una nova diferència entre les comunitats vegetals d’aquesta regió i les de la zona occidental. Per això hi ha espècies de plantes que són exclusives de l’E del continent, i d’altres d’àmpliament esteses que en canvi manquen en aquesta zona.
IDEM, a partir de fonts diverses
Sens dubte és a l’Àfrica on el bioma de les sabanes pren la màxima extensió i es manifesta amb la màxima varietat de tipus de vegetació i de poblament animal. Com a l’Amèrica del Sud, els extensos altiplans interiors acullen sabanes llenyoses, sovint qualificables de boscos clars mentre que les àrees més deprimides i de vegades mal drenades més aviat acullen sabanes herboses, però aquest model general admet, com ja s’ha posat de relleu en parlar de les sabanes americanes, una amplíssima variació de tipus, fins i tot en àrees molt properes.
Les sabanes guineanes
En contacte amb les selves plujoses equatorials de la regió guineana (de vegades a dins i tot, en claps de sòls pobres de nutrients o que han estat explotats en el passat per agricultors itinerants), formant una àmplia aurèola al marge septentrional de la selva plujosa, des de la costa atlàntica compresa entre la baixa Casamance i l’W de Libèria fins a l’alt Nil, apareixen poblaments de sabanes, tant llenyoses com herboses, amb nombroses afinitats florístiques amb aquelles selves. Una situació semblant es dóna al S de l’àrea de selva plujosa equatorial, tant a la baixa conca del riu Zaire com en algunes àrees de Gabon, i comporta també la presència de claps menors de vegetació predominantment herbàcia (“ésôbé”) que s’intercalen al si de la selva plujosa adjacent.
Són característiques d’aquestes sabanes la presència freqüent de petits claps de selva plujosa i de boscos en galeria al llarg dels cursos d’aigua i l’absència d’algunes espècies pròpies de les sabanes sudaneses, amb les quals confinen pel nord, com l’arbre de la mantega o “karité” (Vitellaria [=Butyrospermum] paradoxa, família de les sapotàcies). Els arbres més comuns són la combretàcia Anogeissus leiocarpus, l’ocnàcia Lophira lanceolata i diferents cesalpiniàcies del gènere Isoberlinia; també és freqüent la palmera de Palmira africana o “rônier” (Borassus aethiopum). L’estrat herbaci sol ser alt, d’1,5 a 3 m; hi predominen espècies dels gèneres Andropogon, Hyparrhenia, Loudetia, Panicum (com ara P. maximum, l’herba de Guinea) i Pennisetum. A les de la perifèria meridional de la selva plujosa de la conca del Zaire, en contacte amb el “miombo”, apareixen nombroses espècies llenyoses nanes de gèneres ben representats entre els arbres de la selva plujosa africana, com la crisobalanàcia Parinari pumila, l’apocinàcia Landolphia lanceolata, o l’anisofil·leàcia Anisophyllea poggei.
Pel que fa a la fauna, val la pena remarcar entre els mamífers una certa tendència a la melanosi, és a dir un cert enfosquiment del pèl, generalment associada a l’hàbitat forestal. Entre els herbívors destaquen el gran èland (Taurotragus derbianus), l’antílop equí (Hippotragus equinus), el cob comú (Kobus kob) i el cob aquàtic (K. ellipsiprymnus); també s’hi fan la majoria dels grans carnívors de la fauna africana sabanícola i molts dels més petits, com mangostes i civetes.
Les sabanes sudaneses
Al N de les sabanes guineanes, des de l’Atlàntic fins a Etiòpia, s’estén una zona de sabanes que confina en els seus límits septentrionals amb els subdeserts sahelians. Són probablement, amb les de l’Àfrica oriental, les més típiques i conegudes de les sabanes herboses africanes, encara que en aquest espai se’n presenten també de llenyoses, tant arbòries com arbustives, que en alguns casos prenen l’aspecte de bosquets clars en els quals predominen algunes de les espècies arbòries presents també de manera més escadussera a les herboses.
El límit meridional de l’arbre de la mantega o “karité” (Vitellaria [= Butyrospermum] paradoxa) constitueix a grans trets, tal com s’ha dit, el límit entre el domini sudanès i el guineà. Pel nord, el trànsit cap a la zona saheliana de subdeserts d’espinosos amb recobriment herbaci discontinu es fa per un gradient gairebé imperceptible que, a la part més occidental, més directament sotmesa a la influència atlàntica (Senegal i Mali), se situa al voltant del paral·lel 13°N per davallar després gradualment cap al sud fins a arribar al voltant dels 10°N als confins entre la República Centrafricana i el Sudan, i encara més a les regions meridionals d’aquest país; probablement és el fet de l’existència de poblacions sedentàries lligades a l’agricultura cerealícola el tret que assenyala amb més claredat el límit entre sabana i subdesert, límit que les sequeres persistents de les darreres dècades han tendit a desplaçar més cap al S. Entre les espècies llenyoses més freqüents predominen mimosàcies (Acacia, Prosopis, Albizia, Parkia), combretàcies (Anogeissus, Combretum, Terminalia) i cesalpinàcies (Isoberlinia, Cassia, Afzelia), com també algunes espècies d’altres famílies, com l’euforbiàcia Uapaca togoensis, la meliàcia Khaya senegalensis o la dipterocarpàcia Monotes kerstingi. A aquestes cal afegir, a les regions sotmeses a explotació agrícola, algunes espècies arbòries no cultivades, però sí preservades pels agricultors pel profit que n’obtenen, entre les quals el ja esmentat arbre de la mantega, el “cad” o “gao” (Acacia [= Faidherbia] albida), usat com a farratge i aprofitat també en múltiples altres usos, o el famós baobab Adansonia digitata.
La fauna de grans herbívors de les sabanes sudaneses és encara més rica i variada que la de les guineanes. Inclou tots els antílops citats abans i també l’antílop jeroglífic (Tragelaphus scriptus), la gasela de front roig (Gazella rufifrons), l’oribí (Ourebia ourebi) i diferents duikers (Sylvicapra grimmia, Cephalophus rufilatus), als quals cal afegir l’elefant africà (Loxodonta africana), el búfal africà (Syncerus caffer), la girafa (Giraffa camelopardalis), el facoquer (Phacochoerus aethiopicus) i el babuí anubis (Papio anubis) i altres simis. No cal dir que tots els seus predadors estan també ben representats, des del lleó (Panthera leo) fins al licaó (Lycaon pictus), passant pel lleopard (P. pardus), el guepard (Acinonyx jubatus) i per tota la varietat de hienes (Crocuta, Hyaena) i xacals (Canis adustus, C. aureus, C. mesomelas).
Les sabanes de l’Àfrica oriental
A l’Àfrica oriental hi ha les sabanes per excel·lència, semblants fisiognòmicament a les precedents, i han estat divulgades a bastament per la literatura i el cinema de safaris o, actualment, per la promoció turística dels grans parcs de la regió, segurament els més ben equipats de tot Àfrica. S’estenen, a grans trets, entre els grans llacs i l’Índic, des del llac Victòria al nord fins al baix Zambezi al S. En la seva forma més típica consisteixen en un estrat herbaci elevat en el qual predominen les gramínies Themeda triandra i Heteropogon contortus, al si del qual creixen, més o menys esparsos, arbres de diferents espècies, fonamentalment mimosàcies (Acacia) i combretàcies (Combretum), però també cesalpiniàcies de les que en altres àrees de l’Àfrica austral donen lloc a les sabanes llenyoses i els boscos clars del “miombo” (Brachystegia, Julbernardia). Localment, a les parts més alteroses dels altiplans que envolten el Gran Rift, apareixen precisament aquests “miombos”, més característics de les regions més interiors del continent africà.
Es tracta de les sabanes africanes amb una mastofauna més rica i variada, tant pel que fa als herbívors com als carnívors. Alguns herbívors, com els nyus blaus (Connochaetes taurinus), les zebres de Burchell (Equus burchelli) o les gaseles de Thomson (Gazella thomsoni), viuen en grans ramats que migren segons les estacions, o en petits grups familiars, com els impales (Aepyceros melampus), que formen ‘harems’ amb un mascle dominant, o els elefants africans (Loxodonta africana), que formen petits grups de femelles amb les cries, liderats per la femella de més edat, mentre els mascles se separen d’aquests grups familiars en arribar a la pubertat i viuen sols o s’apleguen en grups coetanis.
La sabana llenyosa típica o “miombo”
A la zona meridional de les selves plujoses de la conca del Zaire i de les sabanes florísticament afins que han estat tractades conjuntament amb les guineanes, s’estén, fins al veïnatge de les riberes de l’Okawango i del Zambezi, una vasta regió d’altiplans poblada fonamentalment d’una sabana llenyosa, sovint qualificada de bosc clar, caracteritzada pel predomini de les cesalpiniàcies dels gèneres Brachystegia, Julbernardia i Isoberlinia, sobretot dels dos primers, que reben a la regió el nom de “miombo”.
Els “miombos” més típics es fan en altitud (entre 1.000 i 1 850 m) i tenen un estrat herbaci dens de gramínies perennes. A més baixa altitud, als fons de les àmplies valls dels gran rius de la regió (Okavango, Zambezi, Luangwa, Limpopo, Cunene), les sabanes llenyoses són representades per boscos clars de “mopane” (Colophospermum mopane, família de les cesalpiniàcies). Al nord, a la perifèria meridional de la vall del Congo, en particular en àrees relativament deprimides de la província zairesa de Shaba (Katanga) i de la zona septentrional de Zàmbia, apareix una altra variant en la qual dominen la dipterocarpàcia Marquesia macroura i la cesalpiniàcia Berlinia giorgii, sovint considerada com una formació secundària resultant de la destrucció d’un bosc dens perennifoli. Localment, en àrees mal drenades, apareixen sabanes herboses semblants a les de les planes de Kenya i de Tanzània.
El més característic de la fauna de grans herbívors és la presència d’espècies de bosc, de pelatge ratllat, com el bongo (Tragelaphus euryceros) o el niala (T. angasi).
Els mosaics sud-africans de sabanes llenyoses i herboses
Al S de la regió del “miombo”, fins als confins australs del continent (excloses les regions més àrides, de desert i subdesert, de Kalahari i del Namib) s’estén una zona de relleu més complex, en la qual sabanes llenyoses i herboses, boscos afromontans, praderies temperades, “fynbos” mediterranis, aiguamolls o boscos en galeria es barregen de manera inextricable en un complex mosaic, del qual cal inisitir aquí en els components del bioma de les sabanes: la sabana herbosa occidental del Kalahari i la sabana llenyosa, coneguda a Sud-àfrica com a “bushveld”.
El Kalahari, si més no a la part central i la septentrional, és en realitat, malgrat la seva reputació de desert, el límit sud-occidental del bioma de les sabanes (que molts autors fan coincidir amb el límit austral de la combretàcia Terminalia sericea). Només les seves parts meridionals i occidentals, en els confins del Karoo i del Namib, són un veritable subdesert. És l’absència de cursos d’aigua superficials a causa de la permeabilitat del substrat sorrenc el que ha fomentat la imatge de Kalahari com a desert. En realitat, tot i que a les dunes s’hi fan només poblaments esclarissats de gramínies de llargs rizomes i algun “witgat” (Boscia albitrunca), una petita caparidàcia semblant a la taperera, la major part de l’espai és ocupat per sabanes herboses molt obertes. A l’estrat herbaci hi predominen Themeda triandra, coneguda a Sud-àfrica com “rooi grass”, diferents espècies de gramínies del gènere Aristida, i algunes compostes del gènere Pentzia; a l’estrat arbori hi ha acàcies de diverses espècies, principalment Acacia giraffae, el “camel’s-thorn” o “kameeldoorn” dels sud-africans, però també A. heteracantha, A. mellifera, “cross-berries” (Grewia flava, família de les tiliàcies), ginjolers de búfal (Ziziphus mucronata, família de les ramnàcies) i gegantins baobabs Adansonia digitata escampats isoladament o en petites motes.
El “bushveld” és l’extensió meridional de les sabanes llenyoses de tipus “miombo” que es prolonga ben enllà del tròpic de Capricorn, al llarg dels costers de mitjana altitud dels vessants marítims encarats a l’oceà Índic de les muntanyes de Natal i de l’extrem oriental de la província del Cap. En altitud, a l’E i al N de Zimbabwe, al veí Moçambic i al N del Transvaal (entre 1 600 i 1 800 m d’altitud), en contacte amb els boscos afromontans dels estatges culminals de les muntanyes, predomina el bosc clar de Brachystegia spiciformis, amb Julbernardiaglobiflora, la proteàcia Faurea speciosa, la meliàcia Ekeber-gia capensis, la crisobalanàcia Parinari curatellifolia i diverses combretàcies dels gèneres Terminalia i Combretum, bé que sovint aquesta vegetació ha estat transformada per la intervenció humana en una sabana herbosa amb P. curatellifolia, sovint preservada per aprofitar-ne els fruits, i combretàcies o en boscos secundaris d’Uapaca kirkiana.
A cotes inferiors, entre 1 400 i 1 600 m, es troben poblaments gairebé purs de Brachystegia boehmii amb proteàcies, la dipterocarpàcia Monotes glaber i, de vegades, Parinari curatellifolia, amb un estrat herbaci graminoide; molt més extenses en el passat, aquestes sabanes llenyoses han estat reemplaçades per sabanes més obertes amb poblaments arbustius, principalment de proteàcies (Protea angolensis, P. gaguedi). Més avall, entre 1 200 i 1 400 m, des de la regió de Bulawayo, a Zimbabwe, fins als confins meridionals del bioma, sobre sòls sorrencs procedents de la meteorització de granits, apareixen els poblaments que assenyalen els confins meridionals del bioma de les sabanes: les sabanes llenyoses o boscos esclarissats de lilà bord africà (Burkea africana, família de les cesalpiniàcies) i terminàlia argentada (Terminalia sericea). A les mateixes cotes, sobre sòls de textura més pesant són sabanes herboses amb acàcies les que predominen i, en sòls mal drenats, sabanes llenyoses de “mopane” (Colophospermum mopane), que ocupen també una gran extensió a cotes inferiors, tal com passa a les regions més septentrionals.
La gran fauna de les sabanes de l’Àfrica austral repeteix a grans trets la ja descrita per altres àrees de sabana africanes. Són característics d’aquesta regió el petit i gràcil “springbok” (Antidorcas marsupialis), l’amenaçat “blesbok” de front blanc (Damaliscus dorcas) i les zebres de Burchell i de muntanya (Equus burcheli, E. zebra). També s’hi conserven algunes de les poblacions més nombroses d’antílop sabre (Hippotragus niger); les poblacions d’elefants estan creixent a la majoria d’espais protegits de la regió.
Les sabanes malgaixes
Madagascar havia estat cobert en la seva major part de boscos i bosquines. Actualment, el 80% de la superfície de l’illa pel cap baix és coberta per vegetació secundària. La destrucció i la degradació de boscos i bosquines han donat lloc a tota una varietat de tipus de vegetació secundaris, des de gramenets poc densos, passant per altres d’embosquinats, sabanes herboses i llenyoses, fins a boscos secundaris.
L’activitat humana és la principal responsable de la destrucció del bosc primari, i com que es creu que l’illa de Madagascar només ha estat poblada pels humans els darrers 1.500 a 2.500 anys, es pot considerar que les sabanes herboses i llenyoses han de ser un fenomen relativament recent. Tanmateix, la troballa de fòssils d’animals, ara extingits, adaptats a la pastura així com també les anàlisis pollíniques de sediments antics, que han demostrat que moltes de les espècies de plantes que creixen a praderies i sabanes de Madagascar ja hi vivien abans de l’arribada dels primers humans, han plantejat seriosos dubtes a alguns científics. En tot cas, però, tot i que és possible que alguns tipus de vegetació de sabana existissin abans de l’arribada dels humans, el cert és que actualment l’activitat humana és un factor que té un paper important en la dinàmica i l’expansió d’aquests hàbitats. Les espècies llenyoses de les sabanes malgaixes són, per una banda, les que sobreviuen a la crema dels boscos, relíquies d’aquelles que ocupaven inicialment l’àrea i, per una altra, les oportunistes que han envaït la vegetació secundària. S’ha proposat que els límits actuals entre les àrees de sabana i les àrees de bosc corresponen a límits entre tipus diferents de substrats i de geomorfologia i que això seria a causa de la diferent susceptibilitat al foc que presenten els diferents tipus de boscos. Per exemple, els boscos amb sotabosc poc desenvolupat, com ara els que es fan sobre els afloraments de calcàries que es troben a l’oest de l’illa, resisteixen força bé el foc i generalment es conserven bé, mentre que algunes àrees veïnes, amb sòls sorrencs més profunds, són ocupades ara per sabana. Les sabanes es mantindrien en aquest cas a causa dels focs repetits regularment i l’explotació pels ramats.
La regió oriental de Madagascar, que rep les pluges més abundants i comprèn l’escarpat penya-segat oriental, l’altiplà central i les serralades muntanyoses, té característicament una coberta de boscos perennifolis. En contrast, la regió occidental, que presenta un hivern llarg i sec i que davalla gradualment des de l’altiplà central fins a les costes de l’W i del S, es caracteritza per tenir boscos caducifolis i bosquines. A ambdues regions es troben espais amb vegetació que es pot classificar com a sabana herbosa o llenyosa, més extensos i diversificats a la regió occidental. Madagascar té un nombre total d’espècies molt elevat, i s’estima que 10.000 espècies corresponen a plantes vasculars. El nivell d’endemisme és en conjunt molt alt, tant a la regió oriental com a l’occidental (aproximadament el 80% de les espècies i entre un 15 i un 20% dels gèneres son endèmics). Tanmateix, els hàbitats de sabana tenen uns nivells d’endemisme molt més baixos, com també una menor diversitat d’espècies en comparació amb els boscos i bosquines climàcics.
Les sabanes de la regió oriental
A la regió oriental només es fan sabanes als costers més secs, orientats a ponent, de les terres altes de l’altiplà central, entre 800 i 1 600 m, on es troba, a les àrees menys pertorbades, un tipus de bosc característic, el bosc de “tapia” (Uapaca bojeri), un arbre de la família de les euforbiàcies, nuós i de fulla perenne, que arriba a fer 12 m d’alt, dominant la vegetació d’aquestes àrees. De fet, el bosc mai no és dens, i en la majoria de les àrees, les pertorbacions externes l’han reduït a sabanes llenyoses o a gramenets on tan sols sobreviuen alguns arbres escampats. Altres espècies comunament associades amb U. bojeri són Leptolaena pauciflora, L. bojerana i Sarcolaena oblongifolia, sarcolenàcies endèmiques de Madagascar, com també l’ericàcia Agauria salicifolia, la teàcia Asteropeia densiflora, la sapindàcia Dodonaea madagascariensis, les asteràcies Brachylaena microphylla i Dicoma incana, rubiàcies dels gèneres Alberta i Tarenna, la proteàcia Faurea forticuliflora, les anacardiàcies Protorhus buxifolia i Rhus taratana, l’araliàcia Schefflera bojeri i diferents cunoniàcies del gènere Weinmannia; la palma Chrysalidocarpus decipiens es troba a les àrees més humides.
Mentre que un cert nombre d’espècies d’arbustos i d’herbes poden créixer sota la lleugera ombra dels arbres, en moltes àrees obertes predominen les gramínies: a les cotes més elevades les endèmiques, típiques de les terres altes centrals, com Loudetia simplex stipoides i Aristida rufescens, mentre que a altituds més baixes prenen importància espècies no endèmiques molt difoses, com ara Heteropogon contortus, Hyparrhenia rufa i Hyperthelia dissoluta. Upaca bojeri és molt resistent al foc, i persisteix en aquells indrets on les flames han eliminat les altres espècies llenyoses. No obstant això, existeixen grans àrees a Madagascar que antigament foren poblades per “tapia” i on aquesta espècie no ha sobreviscut. Upaca bojeri és un arbre important per als malgaixos, ja que els seus fruits són comestibles i posseeix propietats medicinals.
Les sabanes de la regió occidental
Les sabanes de l’W malgaix, tant les herboses com les llenyoses, són molt més extenses i diverses que les de la regió oriental. S’hi poden diferenciar bàsicament quatre grups de components llenyosos. En primer lloc, les espècies relictes dels boscos preexistents, generalment grans arbres que han resistit els focs repetits o que, gràcies a la seva mida costen massa de tallar, per la qual cosa han pogut sobreviure; aquest seria el cas de diversos baobabs (Adansonia, en particular A. grandidieri), del tamarinde (Tamarindus indica) i de la gran palmera Bismarckia nobilis. En segon lloc, les espècies endèmiques de la sabana, que es reduirien a tres arbres: la malpighiàcia Acridocarpus excelsus i les asteràcies Dicoma incana i D. oleifolia. En tercer lloc, les espècies autòctones invasores, en particular les espècies adaptades als sòls empobrits, com les que creixen als afloraments rocosos, per exemple l’eritroxilàcia Erythroxylum platycladum, la bignoniàcia Fernandoa madagascariensis, com també les del gènere Stereospermum, l’apocinàcia Mascarenhasia lisianthiflora i la combretàcia Terminalia seyrigii. Finalment, les espècies invasores exòtiques, introduïdes a Madagascar en data relativament recent des del continent africà o des d’altres indrets, com la mimosàcia Acacia farnesiana, les celastràcies Cassine aethiopica i Maytenus linearis, la palma Hyphaene coriacea, l’anacardàcia Sclerocarya birrea subspècie caffra o les ramnàcies del gènere Ziziphus. A cada àrea concreta, els components llenyosos de la sabana dependran de factors locals, ambientals i històrics. Cal remarcar que en certes localitats predominen espècies llenyoses determinades, per exemple en les extenses planúries a prop del parc nacional d’Isalo, on Bismarckia nobilis forma una sabana de palmeres molt notable.
Les espècies herbàcies son predominantment gramínies, i inclouen tant espècies endèmiques, com ara Loudetia simplex subspècie stipoides, L. filifolia subspècie humbertiana o Aristida rufescens, com espècies no endèmiques, d’amplia distribució: A. congesta, Heteropogon contortus, Hyparrhenia cymbaria, H. rufa i H. schimperi, la xisca (Imperata cylindrica), l’herba de Guinea (Panicum maximum) i Themeda quadrivalvis. També són comunes algunes espècies anuals.
Les sabanes austroasiàtiques i insulíndiques
Les diferents condicions climàtiques del subcontinent indi i de la península indo-xinesa i les illes insulíndiques configuren dos panorames força diferents pel que fa a l’extensió i la distribució del bioma de les sabanes.
La imprecisa condició de la sabana asiàtica
La major part del subcontinent indi, que es troba situat a cavall de les zones tropical i subtropical (entre els 8 i els 38°N), està sotmesa a un clima monsònic. Les precipitacions que cauen sobre la totalitat de la regió provenen primordialment del monsó del sud-oest i es concentren al llarg d’un període relativament breu, de finals de maig al setembre, si bé el monsó en retirada (del nord-est) també porta pluges a algunes zones, principalment a la part meridional del país. Així, la major part del territori té una estació de pluges després d’un llarg període sec i calorós. El resultat és que el tipus de vegetació propi de la major part del subcontinent indi són els boscos caducifolis secs, la composició específica i altres característiques biòtiques dels quals varien d’una zona a una altra en funció de la quantitat i la durada de les precipitacions, com també d’altres factors geològics, edàfics i biòtics. El subcontinent indi, per altra banda, té una història molt antiga de poblament humà (ha estat el bressol d’algunes de les civilitzacions més antigues) i per això els boscos de la regió han sofert un esclarissament extensiu per tal de fer-los habitables i conreables i han estat recorreguts per incendis i bestiar des de temps immemorial, cosa que ha donat com a resultat un tipus de vegetació de sabana herbosa sotmès a una intensa pressió biòtica, a la major part del territori potencial dels boscos caducifolis secs.
Per la seva banda, el sud-est asiàtic és com un continent que hagués lliurat el seu cor a la mar, i resulta una de les regions més humides del món. A causa d’aquesta humitat intrínseca, la distinció entre boscos, sabanes llenyoses i sabanes herboses està especialment desdibuixada en aquest territori, i tant sabanes llenyoses com herboses són les més regularment poblades d’arbres del món. Els boscos esclarissats són abuntants a la part continental del sud-est asiàtic, on constitueixen la formació més important de Myanmar, Tailàndia, Laos, Cambodja i Vietnam (sovint fins a un 45% del total de coberta forestal) i també es troben a les illes més orientals d’Indonèsia i a Papua Nova Guinea mentre, en realitat, les sabanes pròpiament herboses són molt rares a la regió i generalment només s’hi troben en condicions ecològiques força particulars. La vella idea segons la qual totes aquestes sabanes s’havien originat en temps recents a partir d’espais forestals artigats ja no és, però, sostenible, atès que cada cop hi ha més evidències que al sud-est asiàtic hi ha hagut, si més no, petits nuclis de sabana des de temps molt antics, la majoria probablement associats amb determinats trets de la topografia i del sòl. Tanmateix, no és menys evident que l’extensió de les formacions llenyoses obertes i les sabanes ha fluctuat constantment a partir d’aquests espais nuclears sota la influència de canvis climàtics i accions humanes.
A l’últim període glacial, quan els tròpics eren més secs que no pas ara, els boscos oberts i la sabana probablement ocupaven extensions molt més grans del sud-est asiàtic que no pas avui dia, fins i tot a la península de Malaca, avui clarament infeudada al bioma de la selva plujosa en tota la seva extensió, sembla que una mena de ‘passadís’ de sabana enllaçava el continent i les regions insulars actuals. Quan es va restablir el monsó després de la retirada del gel a les latituds septentrionals, les sabanes llenyoses i herboses s’haurien tornat a encongir fins a quedar restringides als espais nuclears als sòls sorrencs, als pendents escarpats i pedregosos i als sòls pobres amb una capa de laterita (plintita) prop de la superfície. Però a continuació, amb l’arribada dels primers pobles neolítics al sud-est asiàtic, fa aproximadament 12.000 anys, les formacions llenyoses obertes i les sabanes s’havien de tornar a expandir, en aquest cas sota la influència de l’ús humà del foc i de les eines de tall; si més no a les àrees on un clima estacional permetia escampar extensament el foc durant l’estació seca.
A les àrees nuclears de sabana també hi ha foc, però no és el factor determinant de l’existència mateixa de la sabana; mentre que a les noves extensions d’aquesta, el foc és un element essencial ja que, si es controla o s’evita, la formació a poc a poc torna a esdevenir més atapeïda i acaba prenent caràcter forestal. Actualment continuen estenent-se les sabanes, tant llenyoses com herboses, en llocs pròxims a l’ecotò entre sabana i bosc en els quals es provoquen focs regularment. A l’estació experimental de Sakaerat, al NE de Tailàndia, per exemple, es poden observar molts fronts ben definits en els quals s’ha deturat algun foc, que indiquen la progressió de sabanes i formacions llenyoses obertes al si del bosc monsònic i fins i tot de la selva plujosa. També val la pena remarcar que molts dels jaciments neolítics del sud-est asiàtic s’associen a àrees actuals de sabana, encara que és difícil decidir si aquestes àrees són un producte de la població humana o si van ser triades originàriament pels seus sòls lleugers i la seva accessibilitat. Les sabanes del sud-est asiàtic serien, doncs, alhora, tant una formació climàtica com una formació edàfica, com fins i tot una formació antròpica, originada sota la influència de les pressions humanes. Les mateixes sabanes, tant llenyoses com herboses, poden haver-se produït a diferents llocs per obra de diferents combinacions de processos.
>Les sabanes de l’Índia subhumida
A les zones subhumides del Deccan, amb precipitacions de 800 a 1 250 mm, distribuïdes al llarg de nou mesos (de març-abril a novembre-desembre), igual com succeeix en moltes àrees de l’Àfrica sotmeses a una pressió humana comparable, es donen formacions sabanoides amb un estrat llenyos més o menys dens, però de composició florística no excessivament allunyada de la de les sabanes africanes amb combretàcies. Sobre un estrat herbaci dominat per “daabsuli” (Heteropogon contortus) o per “pavna” o “sedwa” (Sehima nervosum) i diferents espècies de Dichanthium, creix un estrat llenyós en el qual predominen combretàcies Anogeissus latifolia, el “bakla” o “dawra”, i Terminalia tomentosa, juntament amb la burseràcia Boswellia serrata, el “salai” o “luban”, la papilionàcia Pterocarpus marsupium, el “bijasar” o “pitasara”, i l’ebenàcia Diospyros melanoxylon, el “tendu” o banús coromandel. Cap al N, en sòls sorrencs, aquestes espècies comencen a ser desplaçades pel sal (Shorea robusta) en una transició cap als boscos monsònics amb predomini dels estrats llenyosos.
Els boscos espinosos de l’Índia semiàrida
De llevant a ponent, a mesura que disminueixen les precipitacions, tant en quantitat com en durada, les condicions passen de ser semiàrides a àrides. A l’Índia peninsular, el vessant de sotavent dels Ghats Occidentals rep una precipitació molt baixa i té un clima semiàrid. A causa de la gran variabilitat de la precipitació d’un any per l’altre, es fa difícil de determinar un límit precís per a la regió semiàrida; les zones amb una precipitació anual mitjana d’entre 200 i 800 mm poden ser considerades típicament semiàrides. Els boscos i bosquines espinosos (“thorn forests”) amb un estrat herbaci de tipus sabana són característics de les regions semiàrides de l’Índia i es donen en algunes àrees del Panj¯ab i de Hary¯amna, a la major part de Raj¯astha (a l’est dels monts Ar¯aval¯a), en àrees de la part occidental d’Uttar Prad¯esh i de Madhya Prad¯esh, al sud de Gujar¯at i a una estreta franja a sotavent dels Ghats Occidentals a Mah¯ar¯ashtra, Karn¯ataka i Tamil Nadu.
La vegetació arbòria és dominada per les mimosàcies, principalment dels gèneres Acacia i Prosopis. Les àrees rocalloses dels turons són dominades per una vegetació arbustiva, amb eufòrbies suculentes. Durant l’estació seca, el sòl roman pràcticament nu o amb algun arbust escadusser, però tan bon punt cauen les primeres pluges, les gramínies i les altres espècies herbàcies comencen a créixer de manera exuberant i els arbres treuen un fullatge verd esponerós. Així, durant l’estació de les pluges, els boscos espinosos constitueixen un exemple ben característic de sabana llenyosa. Malgrat l’elevat nombre d’espècies de gramínies i altres plantes herbàcies presents en aquests ‘boscos’, la diversitat és molt baixa comparada amb la dels boscos caducifolis. Cap espècie vegetal no és endèmica d’aquests boscos espinosos indis.
Els boscos espinosos acostumen a agrupar-se en dues categories: els septentrionals i els meridionals, relativament més ben coneguts els primers que els segons. Els típics boscos espinosos septentrionals, al N del massís de S¯atpur¯a, es componen principalment de dues mimosàcies, el “kumta” (Acacia senegal) i el “bordi” (Prosopis cineraria [=P. spicigera]). Altres espècies arbòries comunes són: el “ronj” (Acacia leucophloea), el “babul” (A. nilotica) i el “khair” (A. catechu), que són mimosàcies, i l’“hingota” (Balanites aegyptiaca, família de zigofil·làcies), el “vurtuli” o “kunlai” (Dichrostachys cinerea, família de les mimosàcies), el “khatai” o “bilangra” (Flacourtia indica, família de les flacourtàcies) i la celastràcia Maytenus spinosa. Els “pilu”, com el “barapilu” o “khanjar” (Salvadora oleoides) i el “pilu” pròpiament dit o “chhota pilu” (S. persica), són típics de les àrees salines. Hi ha una gran varietat d’espècies arbustives tant disseminades entre els arbres com en forma de bosquina contínua, les més comunes de les quals són el “tainti”, també anomenat “kair” o “karel” (Capparis decidua, família de les caparidàcies), l’“aak” o “madar” (Calotropis procera, família de les asclepiadàcies) i el “jharberi” (Ziziphus nummularia, família de les ramnàcies). Altres arbustos, com per exemple el “phog” (Calligonum polygonoides, família de les poligonàcies), el “ghorak boonti” o “chaya” (Aerva javanica, família de les amarantàcies), el “jhamo” o “khip” (Crotalaria burhia, família de les papilionàcies), el “jangli karonda” (Carissa opaca, família de les apocinàcies) i l’asclepidàcia Leptadenia pyrotechnica, també són freqüents en aquests boscos, especialment en els més degradats i amb més gran quantitat d’herbàcies. A les zones muntanyoses, el “thor” (Euphorbia nivulea) és l’espècie arborescent més comuna, encara que més aviat sol presentar-se en forma de gran matoll. L’acantàcia Justicia vasica és un altre arbust comú de les zones rocalloses i pedregoses.
La coberta herbàcia és dominada per gramínies com els “anjan” (Cenchrus ciliaris) i altres congèneres, com “kala anjan” (C. setigerus), el “sewan” (Lasiurus hirsutus), el “pavna” o “sedwa” (Sehima nervosum), el “dhaab” (Desmostachya bipinnata), el “daabsuli” (Heteropogon contortus), el “makra” (Dichanthium annulatum), el “bhanjuri” (Apluda mutica), Dactyloctenium scindicum i espècies d’Aristida, Cymbopogon, Eragrostis, Bothriochloa, Eleusine, etc. Les espècies resistents al foc, com el “kans” o canya dolça espontània (Saccharum spontaneum) i S. bengalense, també són força comunes.
A les àrees salines, especialment les regions de Saur¯ashtra i del Kutch, a Gujar¯at, les espècies llenyoses predominants són la ja esmentada Salvadora oleoides i el “jhau” (Tamarix ericoidea), que és una tamaricàcia. La mimosàcia americana Prosopis chilensis [=P. juliflora], un mesquite mexicà, coneguda localment com a “kabuli kikar”, introduïda fa més d’un segle, s’ha naturalitzat a la zona semiàrida i s’ha estès a grans àrees de sòls relativament salins, fet que també ha estat afavorit pels programes de repoblació forestal. Cal destacar que un altre Prosopis, el “bordi” (P. cineraria [= P. spicigera]), espontani a la regió, és un arbre molt apreciat, ja que, a més de proporcionar fusta i farratge, els seus fruits verds també són emprats com a aliment; per contra, P. chilensis no és tan gustós per al bestiar i la seva fullaraca produeix substàncies al·lelopàtiques que inhibeixen el creixement d’altres plantes sota la seva capçada, i per això moltes comunitats locals, per exemple els bishnois del Rajasthan, protegeixen activament el “bordi”. Al llarg dels corrents efímers i estacionals, a les àrees periòdicament inundades, és dominant la gramínia Vetiveria zizanioides, el “khus”, amb presència ocasional de palmes del gènere Phoenix.
Els boscos espinosos més meridionals són gairebé idèntics, llevat d’algunes diferències en les espècies menors. El “khair” (Acacia catechu) i el “babul” (A. nilotica) són les espècies arbòries dominants. A les zones muntanyoses es poden trobar eufòrbies com Euphorbia tirucalli i E. antiquorum.
S’ha estimat que la biomassa dels boscos espinosos oscil·la entre 5 t/ha fins a més de 20 t/ha, segons l’estat de protecció. L’estrat llenyós representa del 30 al 50% de la biomassa total, mentre que l’herbaci presenta una variació considerable en la proporció entre biomassa subterrània i aèria, segons la composició en espècies i d’altres factors ecològics, però en general, la biomassa subterrània és superior a la corresponent a les parts aèries. La producció de les gramínies ha rebut una gran atenció i diversos estudis han demostrat que els gramenets semiàrids de les àrees de bosc espinós, amb una gestió adient, poden produir anualment fins a 20 t/ha de farratge. Tanmateix, tant si s’apliquen pràctiques silvipastorals adients com si no, les espècies arbòries tenen una gran influència sobre la producció de les gramínies; per exemple, sota Prosopis cineraria [= P. spicigera] s’obté un major rendiment d’herba que no pas sota Acacia senegal. De la mateixa manera, alguns arbustos (espècies de Butea, Mimosa, Ziziphus, Cassia) també redueixen el creixement herbaci.
Les sabanes del sud-est asiàtic
Al sud-est asiàtic, com arreu dels tròpics, qualsevol tall en el continuum que va des de les sabanes llenyoses atapeïdes de boscúria fins a les herboses que consisteixen en gramenets gairebé desproveïts d’arbres és ben arbitrari, i encara més si es pensa que el tant per cent de recobriment d’una mateixa capçada pot variar entre el 50 i el 77% simplement en funció de l’estat fenològic de l’arbre. També varia la visibilitat a través del brancam, que pot variar entre (-1/4D +10)2 a les sabanes llenyoses més denses, i (-1/7D +10)2 a les sabanes herboses més obertes (essent D = la distància en m).
Les sabanes llenyoses
Les sabanes llenyoses del sud-est asiàtic són molt semblants en estructura general i fisionomia al “miombo” de l’Àfrica austral, a les sabanes amb eucaliptus de l’Austràlia septentrional i als boscos de sal (Shorea robusta) de l’Índia peninsular.
En concret, a la part continental del sud-est asiàtic, la formació clau és el bosc sec caducifoli de dipterocarpàcies, conegut en birmà com a “indaing” (terra inculta, amb Dipterocarpus tuberculatus) i en siamès com a “pà tengrang” (bosc natural de Shorea). A causa del cobricel força tancat que tenen aquests boscos, l’alsacià Andreas Franz Wilhelm Schimper (1856-1901), un dels estudiosos pioners de la seva ecologia, classificà inicialment aquesta formació com ‘bosc sabanoide’. És àmpliament distribuïda en àrees sotmeses als climes estacionals Am (monsònic) i Bw (de sabana) de la classificació de Köppen, però més aviat a les sotmeses a aquest segon. La pluviositat hi oscil·la generalment entre els 1.000 i els 1.500 mm, amb una estació eixuta de cinc a set mesos (d’octubre/novembre a març/abril). En alguns llocs, l’aigua evaporada pot superar la pluja caiguda nou mesos l’any. El valor anual del coeficient d’humitat d’Angström (H) normalment és menor de 240, cosa que qualificaria el clima de semiàrid. La temperatura mitjana del mes més fred poques vegades davalla per sota dels 20°C.
La formació creix generalment per sota els 1.000 m d’altitud, encara que alguns tipus poden aparèixer en algunes àrees de muntanya seques fins a 1.300 m. Són comuns els focs de baixa o mitjana intensitat (entre 250 i 450 kW/m), tant naturals com provocats pels humans, entre novembre i abril, i tan sols són greus quan hi ha hagut un creixement poc usual de fullaraca i herba seca (principalment gramínies i, en particular, els bambús nans). Això pot passar quan els tècnics forestals intenten erròniament excloure totalment el foc d’aquestes formacions i permeten que s’acumuli el combustible al llarg d’un període d’uns quants anys. Llavors, quan inevitablement s’incendia per causes naturals o accidentals, dóna com a resultat molta destrucció, conseqüència de focs extremadament intensos (fins a 2.000 kW/m), amb temperatures del foc superiors a 900°C.
El bosc sec caducifoli de dipterocarpàcies és dominat per diverses barreges de sis espècies de fulla caduca d’aquesta família: Dipterocarpus intricatus, D. obtusifolius, D. tuberculatus, Shorea obtusa, S. roxburghii (= S. talura) i S. siamensis (= Pentacme suavis i les seves varietats). Altres arbres molt característics són algunes dil·leniàcies del gènere Dillenia, la simarubàcia Irvingia malayana, l’arbre de la laca de Birmània (Gluta usitata, família de les anacardiàcies), el pi Pinus merkusii, les fabàcies Pterocarpus macrocarpus i Sindora siamensis, alguna combretàcia del gènere Terminalia o la mimosàcia Xylia kerrii.
La caiguda de les fulles normalment es produeix de novembre a març, però varia molt d’una espècie a una altra i d’un any a l’altre. La fructificació també es produeix abans dels focs estacionals o just després que els focs han passat pels boscos. El creixement màxim en diàmetre té lloc al setembre, cap a la fi de l’estació de les pluges. Als boscos sabanoides més alts, les capçades de l’estrat superior poden assolir més de 30 m d’alçada però és més comú que oscil·lin entre 15 i 20 m, mentre que, en els casos més extrems d’empobriment del sòl i de sequera, poden no arribar a 10 m. La majoria d’arbres mostren una àmplia varietat d’adaptacions tant als focs moderats com a l’estrès hídric, entre elles fulles i brots terminals tomentosos i escorces gruixudes i aspres; les arrels primàries i laterals són protegides d’igual manera.
L’estrat herbaci comprèn un espectre de geòfits, hemicriptòfits i anuals no menys adaptats, cadascú a la seva manera, al foc i a la sequera. Cycas siamensis, una cícade àmpliament distribuïda, o les palmeres nanes del gènere Phoenix, per exemple, es protegeixen amb una veritable armadura de bases de fulles mortes encavalcades, mentre molts geòfits, com ara les orquídies terrestres del gènere Habenaria, aconsegueixen sobreviure als focs mercès als seus òrgans subterranis. L’estrat herbaci, però, és dominat principalment per bambús nans del gènere Arundinaria i altres gramínies, com Hyparrhenia, les xisques (Imperata), o les dels gèneres Arundinella, Dichanthium, Eulalia, Heteropogon, Polytoca o Themeda. L’acumulació de les restes mortes d’aquestes gramínies tendeix a assolir el seu màxim durant els mesos de febrer i març, i alimenta de combustible els focs de mitjan i final de l’estació seca.
Les sabanes herboses
Algunes de les variants més obertes de les formacions suara descrites, com per exemple els boscos oberts més clars dominats per l’“ingyin” (Shorea siamensis) i S. obtusa, podrien haver estat igualment incloses en aquest apartat, sense cap inconvenient. De fet, el ventall que va dels boscos sabanoides a les sabanes herboses pròpiament dites és molt ampli i, a més dels boscos de dipterocarpàcies i tots els seus derivats sabanoides, s’hi inclouen formacions molt diverses i força localitzades, com els boscos de l’ebenàcia Diospyros burmanica, anomenada ‘te’ a Myanmar, o els bosquetons amb “khair” (Acacia catechu), terminàlia d’Oliver (Terminalia oliveri), i tec de Hamilton (Tectona hamiltoniana). Cal remarcar que les formacions de sabana herbosa pròpies del sud-est asiàtic penetren fins a la vall de Kabaw a l’estat indi de Manipur, fronterer amb Myanmar, encara que sigui en forma de variants prou peculiars. De la mateixa manera, hi ha exemples de sabanes herboses a les illes insulíndiques, especialment a les de la part oriental d’Indonèsia i també a Nova Guinea, allí on el clima predominant és més marcadament estacional, del tipus Am i Aw de la classificació de Köppen. La majoria de les sabanes herboses, nogensmenys, ocupen superfícies petites i localitzades i no sovintegen tant com les llenyoses; són generalment enclavaments peculiars dins de les sabanes llenyoses de la regió, més dominants.
Tanmateix, sí que hi ha sabanes herboses pròpiament dites al sud-est asiàtic, com les anomenades ‘turons pelats’ de la serralada Petchabun i el parc nacional Khao Yai, a l’escarpament sud-oest de l’altiplà de Korat, a l’est de Tailàndia. Aquestes sabanes són petits claps dins d’una àrea fonamentalment boscosa i sovint confinen, no sols amb altres tipus de sabana amb més presència d’arbres, sinó també amb boscos monsònics semicaducifolis. Aquests claps són molt aprofitats per la megafauna d’herbívors del parc, com els sambars (Cervus unicolor) i els muntjacs (Muntiacus). Altres àrees on apareixen sabanes com aquestes són les rodalies de Mondolkiri, a Cambodja, i a les de Ban Me Thuot, a Vietnam.
Els orígens d’aquestes petites ‘illes’ de gramenets incloses dins de boscos i sabanes llenyoses més extensos semblen el resultat d’un conjunt de factors que ocasionen una forma o altra d’estrès ecològic. A Sakon Nakhon, al NE de Tailàndia, per exemple, una esplèndida sabana herbosa avui gairebé desapareguda sembla haver estat el resultat d’inundacions fortes i regulars durant les estacions de pluges. Els “turons pelats” de les muntanyes de Petchabun han estat originats, en canvi, per un règim de tala, pastura i focs repetits i continuats al llarg de molts anys. A la regió on es donen alguns d’aquests darrers enclavaments de sabana herbosa, la pluviositat anual és inferior a 800 mm i, en alguns casos, a 500. El foc és freqüent en totes aquestes sabanes, encara que es trobin en espais protegits com parcs nacionals o santuaris faunístics, i els focs intensos poden obrir clarianes en un bosc i impedir-ne així la regeneració. Els focs molt intensos i freqüents acostumen a afavorir generalment les herbes en detriment del component llenyós. Al parc nacional Wasur d’Irian Jaya, per exemple, fa poc s’han transformat algunes àrees de selva plujosa en sabana a causa del foc.
Algunes àrees particulars, molt localitzades, de sabana també poden estar relacionades amb jaciments arqueològics i, per tant, amb pautes anteriors d’assentament, com passa en algunes àrees frontereres entre Tailàndia i Myanmar i a certs enclavaments neolítics del primer d’aquests països. Les mateixes sabanes llenyoses també són lluny de ser uniformes en les seves estructures, de manera que acostumen a mostrar una heterogeneïtat, tant horitzontal com vertical, força marcada, amb un disseny en mosaic de claps més oberts o més tancats que reflecteixen la seva història ecològica, la seva dinàmica general, la topografia local, la pedregositat del sòl, la història de l’aprofitament humà del sòl, i les pautes locals d’incidència i intensitat del foc. No tots aquests factors i processos són encara prou ben compresos.
En llengua siamesa, les sabanes obertes es diuen “pàa yâa”, que vol dir ‘herba borda’ o fins i tot ‘jungla d’herba’, i aquests hàbitats són molt importants en alguns parcs nacionals o santuaris faunístics, incloent-hi les cèlebres sabanes obertes del santuari de Thung Yai, que fa poc s’han salvat de l’amenaça d’una gran intervenció de ‘desenvolupament’. Les herbes més característiques són la xisca (Imperata cylindrica), la panissola repent (Panicum repens), la canya dolça espontània (Saccharum spontaneum), la canyota (Sorghum halepense), Vetiveria zizanioides i Eupatorium odoratum, i entre les plantes llenyoses són freqüents el pataner (Careya arborea, família de les barringtoniàcies), el “khair” (Acacia catechu), l’acàcia de Siam (A. siamensis) i la bellíssima fabàcia Pterocarpus macrocarpus. Tanmateix, la composició florística de cada espai de sabana sol reflectir fonamentalment la de la formació llenyosa original de la qual ha derivat.
Les sabanes australianes i pacífiques
Pràcticament una quarta part del territori australià, uns 2 milions de km2 disposats en un arc que travessa el nord tropical d’Austràlia del NW d’Austràlia Occidental, a través del N del Territori Septentrional i de Queensland fins al SE d’aquest estat, correspon a àrees de sabana. La sabana, en efecte, és el bioma dominant en el conjunt d’aquesta regió, en la qual hi ha petites illes de selva equatorial plujosa, praderies i brugueres. Al llarg de l’Austràlia nord tropical hi ha un fort gradient de pluges de N a S. Al Territori Septentrional, per exemple, la pluviositat a Darwin, a uns 12°S de latitud, és de 1 491 mm/any, a Katherine, a uns 15°S, és de 980 mm/any, i a Tennant Creek, a uns 19°S, de 450 mm/any, circumstància que condiciona les diverses menes de sabana existents. Es retroben sabanes similars a les australianes en moltes illes del Pacífic sud-occidental, des de Nova Guinea i Timor, immediates a Austràlia, fins a les allunyades Marianes i Fiji.
La consolidada presència de la sabana a Austràlia
La biota de les sabanes australianes manté una relativa integritat i una elevada diversitat pel que fa a tots els seus components més vistents, és a dir plantes, invertebrats, rèptils i mamífers. No s’ha produït l’extinció de mamífers petits i mitjans, tan vulnerables a les pràctiques europees d’ús de la terra arreu del centre i del S d’Austràlia. La flora té un baix percentatge relatiu de plantes alienes (generalment inferior al 10%), i al N d’Austràlia no hi ha guineus ni conills, que han causat un fort impacte en la biota nativa de l’Austràlia central i meridional.
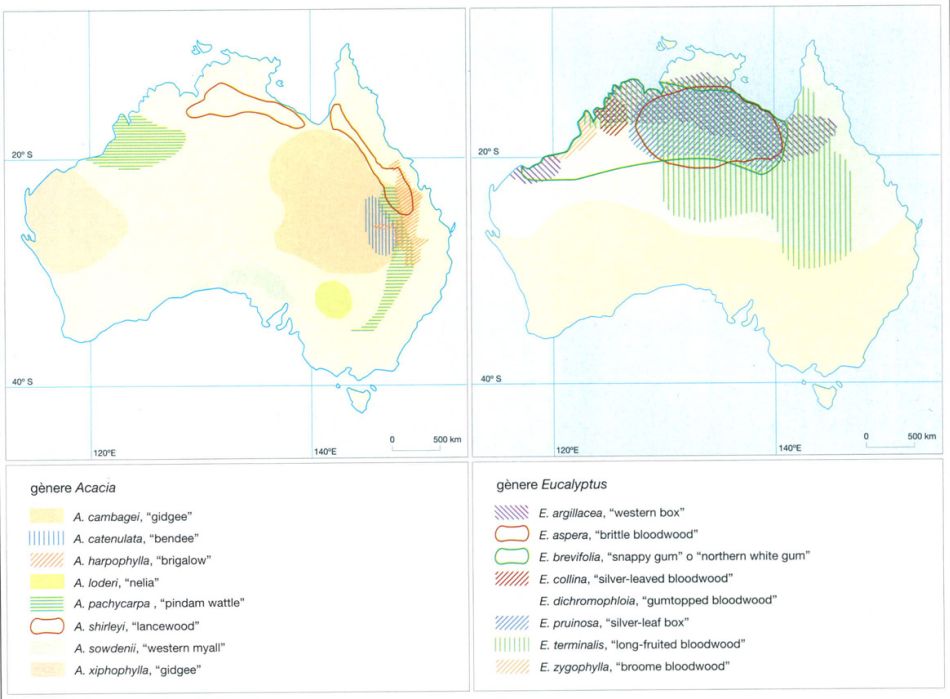
Distribució de les principals espècies d’acàcies i eucaliptus dels ecosistemes de sabana australians. La precipitació mitjana anual i la textura i la composició del sòl determinen que en cada àrea creixin unes o altres espècies d’eucaliptus o acàcies. Eucalyptus argillacea, per exemple, creix en sòls de textura fina, en àrees amb una precipitació mitjana d’entre 500 i 750 mm anuals, mentre que E. dichromophloia es fa en afloraments rocallosos i en sòls sorrencs, en zones on la mitjana de pluges anuals supera aquests valors. A les zones més àrides, les acàcies, tant arbòries com arbustives, reemplacen els eucaliptus.
IDEM, a partir de Beadle, 1981
La sabana australiana sembla que és un fenomen antic, el producte de molts cicles de canvis climàtics durant les eres Terciària i Quaternària, més que no pas un fenomen recent. Tot i que sembla que la majoria de les espècies de la sabana són endèmiques d’Austràlia, les sabanes llenyoses amb eucaliptus de l’Austràlia tropical contenen una proporció molt més alta de gèneres pantropicals que els boscos d’eucaliptus d’Austràlia meridional. Així doncs, molts dels gèneres de les sabanes i formacions llenyoses obertes de l’Austràlia tropical ja havien d’existir al continent australià abans de la separació de la placa tectònica d’Australàsia de l’antic continent de Gondwana fa uns 50 milions d’anys. Almenys 37 gèneres de gramínies (Aristida, Eragrostis, Panicum, Chloris, Sporobolus, Bothriochloa, etc.) i uns quants de fabàcies són compartits amb Àfrica, com també 10 espècies d’acàcies del subgènere Acacia, marcadament diferents de la majoria de les espècies pròpiament australianes d’acàcia, que pertanyen a un altre subgènere.
Uns trentena d’aquests gèneres més de gramínies, per altra banda, són compartits, no tan sols amb Àfrica, sinó també amb la regió indomalaia, fins a la qual degueren arribar transportats per la placa índia, separada al seu torn de Gondwana. Això significa que totes aquestes espècies de gramínies possiblement evolucionaren a l’estrat herbaci de la sabana fins a adquirir els trets ecomorfològics que les caracteritzen abans de l’aparició dels marsupials a Austràlia o de la dels ungulats a Àfrica. Tan sols alguns gèneres de gramínies són endèmics del continent australià, és a dir, evolucionaren després que la placa tectònica australiana s’hagués separat de la resta de l’antic continent de Gondwana. Entre aquests destaquen Astrebla, per exemple, que forma extensos gramenets de “tussock” sobre els sòls argilosos molt esquerdats de les zones més àrides dels tròpics, i Triodia i Plectrachne, que amb els seus “hummock” cobreixen les grans extensions sorrenques i pobres en nutrients del centre d’Austràlia.
L’estrat inferior està format majoritàriament per aquestes gramínies de metabolisme C4, tant anuals com perennes, i plantes herbàcies no graminoides (també tant anuals com perennes) i mates, tots de metabolisme C3. De les deu formes de vida majoritàries, les més comunes són les espècies anuals i les perennes amb fullatge anual. La biomassa herbàcia es troba majoritàriament en l’interval d’entre 2 i 6 t/ha, però pot arribar fins a valors d’entre 8 i 10 t/ha en hàbitats particulars. L’abundància de gramínies anuals decreix, i la de les perennes augmenta al llarg de gradients de pluja decreixent i d’increment del contingut d’argiles del sòl. Els estrats llenyosos són dominats per nombroses espècies del gènere Eucalypthus. Els arbres perennifolis són la forma vital més representada a l’estrat superior, seguits pels arbres i grans arbustos caducifolis, que representen una quarta part de les espècies.
Els patrons fenològics varien marcadament entre els diferents estrats, formes de vida i espècies. En l’estrat inferior la major part del creixement i la floració s’esdevé durant el període de l’estació humida, i normalment s’ha completat a l’abril. Les espècies anuals, sobretot gramínies com per exemple les melques (Sorghum), poden començar a assecar-se al març, poc després de la caiguda de les llavors. El contingut d’humitat declina gradualment al llarg de l’estació seca, i al setembre les espècies de l’estrat inferior normalment s’han assecat del tot.
Les sabanes llenyoses i els gramenets
Dins de les nombroses variants de formacions llenyoses obertes australianes convé remarcar-ne algunes per la seva extensió i representativitat. Seria el cas dels boscos i les bosquines sabanoides d’eucaliptus, les sabanes arbrades, els gramenets de “tussock” i els de “hummock”.
Les sabanes llenyoses en forma de boscos i bosquines sabanoides ocupen les àrees més humides. En els sòls laterítics de la part més humida de la península del cap York i de la zona nord-oriental de la terra d’Arnhem, per exemple, predominen sabanes llenyoses amb un estrat herbaci dominat per Heteropogon triticeus i Sorghum leiocladum, sobre el qual s’enlairen poblaments oberts d’eucaliptus. En canvi, en els sòls sorrencs i pedregosos de la part humida del nord i nord-oest d’Austràlia (terra d’Arnhem i regió de Kimberley) l’estrat herbaci de la sabana llenyosa amb eucaliptus el formen diverses melques anuals (Sorghum intrans, S. stipoideum i S. australiense) i Aristida holathera.
Les àrees subhumides corresponen a les sabanes herboses amb arbres aïllats, de les quals es troben exemples tant al NW (regió de Kimberley, al nord d’Austràlia Occidental, i regions frontereres del Territori Septentrional) com al NW (regions del sud de la península de cap York). En aquests casos l’estrat inferior dels poblaments d’eucaliptus, força més esclarissats que en els boscos sabanoides, és dominat per Chrysopogon latifolius, Heteropogon triticeus, Mnemesia rottboellioides i Sorghum leiocladum.
En ambients més eixuts, com ara els sòls argilosos, negres i molt clivellats que es troben des de Victoria River Downs, al nord-oest del Territori Septentrional, fins al centre de Queensland són característics els gramenets de “tussock” dominats per Chrysopogon fallax, Dichanthium fecundum, Heteropogon contortus i Themeda triandra amb algun peu d’eucaliptus ocasional. També dominen les mates de gramínies en “tussock” als sòls negres inundables estacionalment de les planes costaneres del N del continent, en aquest cas amb predomini d’espècies dels gèneres Eleocharis i Oryza, com també els sòls argilosos de la zona àrida, en els quals les espècies dominants són les “Mitchell grasses” (Astrebla), un dels gèneres de gramínies endèmic d’Austràlia esmentat abans.Finalment, els sòls menys fèrtils de l’àrid interior d’Austràlia estan poblats de gramenets de “hummock” (generalment amb arbustos i arbrissons dels gèneres Eucalypthus i Acacia escampats per ci per lla) on predominen els gèneres Triodia i Plectrachne que formen els característics coixinets de “hummock” amb fulles esclerofil·les, que en el seu creixement poden arribar a formar anells de més de 10 m de diàmetre, conegudes a Austràlia com a “spinifex”.
Dins de l’espai de les sabanes australianes apareixen localment altres tipus de vegetació. Formacions complexes com el bosc monsònic i el “vine thicket” o bosquina de lianes poden aparèixer en situacions topogràfiques en què la humitat del sòl sigui relativament favorable o en què hi hagi protecció del foc.
Les sabanes de les illes del Pacífic
Les sabanes ocupen, a les illes del Pacífic tropical, superfícies generalment petites i que en sovint van lligades a l’acció humana. A Nova Guinea i a Timor predomina el tipus de sabana llenyosa amb eucaliptus, acàcies o melaleuca, encara que a les terres altes de Nova Guinea apareix també una sabana herbosa d’altitud. A les illes menors la sabana s’associa a l’estacionalitat del clima o a condicions topogràfiques a sotavent dels vents que porten les pluges.
A les sabanes de terra baixa de Nova Guinea predominen les gramínies Themeda australis i T. novoguineensis amb un estrat arbori de diferents espècies d’eucaliptus (Eucalyptus alba, E. confertiflora, E. papuana), de vegades amb un estrat llenyós més baix amb acàcies (per exemple Acacia leptoclada), bànksies (per exemple Banksia dentata) i altres arbrissons. També hi ha sabanes inundables amb un estrat herbaci de grans gramínies, com ara Phragmites karka, Leersia hexandra o diferents espècies d’arrossos (Oryza) i un estrat arbori en el qual predominen diferents espècies de Melaleuca (M. cajuputi, M. leucadendron), alguns altres arbres com ara la barringtoniàcia Barringtonia tetraptera, la fabàcia Erythrina fusca o la rubiàcia Mitragyna speciosa com també alguns Pandanus de port arbori. Les formacions herbàcies d’altitud de Nova Guinea poden considerar-se sabanes per la seva fisiognomia i, en part, per la seva composició florística, però en realitat són més aviat praderies orotropicals similars als “páramos andins” o a altres formacions herbàcies de l’alta muntanya tropical; són particularment característiques les sabanes herboses d’altitud amb falgueres arborescents (Cyathea, Dicksonia) i un estrat herbaci de diferents espècies de gramínies en “tussock”.
També són particularment originals les sabanes llenyoses amb “niaoulí” (Melaleuca quinquenervia), de Nova Caledònia. Actualment les diferents variants d’aquest tipus de sabana, que es diferencien tant per la densitat i l’alçada més o menys gran de l’estrat arbori d’aquesta mirtàcia com pels estrats llenyosos i herbacis inferiors, és present en una gran varietat de condicions climàtiques i topogràfiques arreu d’aquestes illes.