L’aprofitament d’uns recursos hídrics insuficients o inabastables
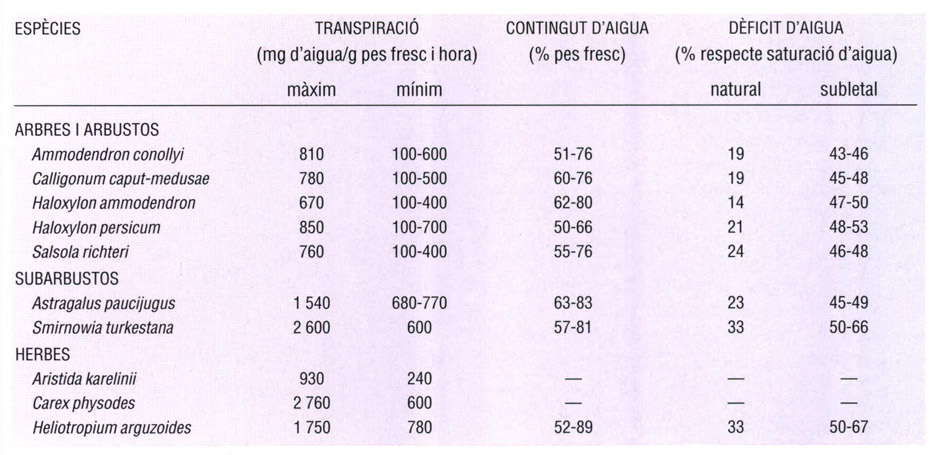
Balanç hídric d’algunes espècies vegetals del desert de Karakum. Les plantes del Karakum tenen sempre un dèficit d’aigua molt baix, i molt allunyat dels valors determinats experimentalment com a letals. El potencial osmòtic és elevat només en les halòfites (Haloxylon, Salsola), adaptades a medis salins, mentre que les altres espècies presenten un valor alt únicament durant els períodes de secada. Les taxes de transpiració són molt variables; tal com s’esperaria, prenen valors més baixos en les plantes llenyoses com arbres i arbustos que en les petites mates i les herbes. Els valors disminueixen mentre dura la sequera.
Goodall, 1983
Els deserts freds presenten uns sistemes ecològics complexos amb plantes, microorganismes i animals altament especialitzats i ben adaptats a les condicions extremes de l’entorn. L’alta especialització i la interacció activa entre ells i el medi que els envolta els permet aprofitar de manera efectiva els pobres recursos que ofereix el desert i assegurar un funcionament estable i força productiu de tot l’ecosistema. L’existència d’un període hivernal amb temperatures baixes i evaporació feble fan que, amb la mateixa quantitat de precipitacions, l’aigua disponible augmenti substancialment en comparació amb la disponible als deserts càlids. Aquest fet caracteritza particularment el funcionament de tots els ecosistemes desèrtics freds i en primer lloc les peculiaritats del seu balanç hídric, és a dir, entre l’entrada i el consum d’aigua. Durant l’hivern, l’aigua de les precipitacions no s’evapora ni és consumida per la vegetació, com succeeix als deserts tropicals o als mateixos deserts freds a l’estiu, sinó que s’acumulen i es retenen fins que comença l’estació càlida. A les regions meridionals, amb hiverns més temperats, aquestes precipitacions són generalment absorbides pel sòl i s’hi acumulen com a reserves hídriques. Als deserts més septentrionals, amb temperatures constantment negatives durant l’hivern, les precipitacions es dipositen a la superfície del sòl en forma de neu i, d’aquesta manera, l’aigua es conserva fins a l’entrada del període càlid.
La importància de la textura dels sòls
Durant la fosa de la neu a la primavera, una gran quantitat d’aigua del desglaç es troba a la superfície del sòl. La seva penetració posterior, el caràcter de la humectació i les particularitats del consum depenen de les propietats del sòl, i principalment de la seva permeabilitat i del modelat del relleu. El substrat més favorable per a la conservació de la humitat és el sòl arenós. A causa de l’alta permeabilitat i la baixa capacitat de retenció, l’aigua penetra amb facilitat al subsòl, fins al nivell freàtic si aquest es troba prou a prop de la superfície, entre els 6 i els 10 m, i d’aquesta manera s’assegura una recàrrega de l’aqüífer. A més a més, l’amplitud de la porositat minimitza els efectes de capil·laritat, de manera que pràcticament no hi ha cap corrent capil·lar ascendent significatiu i les pèrdues d’aigua improductives, per evaporació física, es limiten als primers 50 cm del sòl. Segons valors mesurats a la reserva natural de Repetek, al desert de Karakum, per evaporació física només es perd entre el 30 i el 40% del total anual de precipitacions.
Els deserts amb substrat argilós presenten un quadre lleugerament diferent. Els sòls d’aquests deserts destaquen per una baixa permeabilitat. El coeficient de permeabilitat als sòls franco-argilosos del subdesert de la Ciscàspia septentrional, (és a dir, la velocitat de filtració de l’aigua) és només de 0,1 mm/min, molt baix si es compara amb els sòls franco-arenosos, on el coeficient de permeabilitat és aproximadament d’1,5 mm/min, o sigui, 15 vegades superior. És evident, doncs, que la reserva d’aigua que prové del desglaç (que als deserts septentrionals correspon a uns 150 mm) no pot ser absorbida gaire de pressa pel sòl. En el cas dels espais drenats per la inclinació del pendent es produeix un escolament superficial i la major part de l’aigua s’escapa. En terrenys plans, tanmateix, l’aigua es redistribueix per la superfície i una part considerable fa cap a les depressions del terreny i penetra gradualment al sòl fins al nivell freàtic. En aquest cas, la major part de l’aigua queda preservada de l’evaporació física. Si el nivell freàtic és prou profund, entre 3 i 4 m, l’evaporació només afecta l’aigua dels horitzons superficials i, per altra banda, la humitat d’aquests horitzons superiors és aprofitada ràpidament per les plantes, fet que en redueix també la pèrdua per evaporació física. Als subdeserts argilosos de la Ciscàspia septentrional, la pèrdua d’aigua per evaporació física, tenint en compte fins i tot la de l’estació càlida, és només del 46% del total de precipitacions anuals, i la major part prové, a més, de les precipitacions de l’època estival. Les pèrdues varien segons la posició microtopogràfica: el consum més gran d’aigua (d’un 50 a un 55%) es dóna en els llocs elevats (microelevacions) i als terrenys plans, mentre el consum més baix (d’un 30 a un 40%), té lloc a les fondalades, ja que la humitat penetra més profundament al sòl i queda més ben preservada de l’evaporació.
Així, doncs, els deserts freds posseeixen un avantatge significatiu respecte als càlids: l’existència d’un període fred permet que la humitat de les precipitacions atmosfèriques es conservi i n’impedeix la pèrdua improductiva, que no supera, generalment, la meitat del total de les precipitacions anuals. Cal tenir en compte que el que s’ha dit pot aplicar-se només a les regions que gaudeixen del màxim de precipitacions a l’hivern o, en tot cas, les tenen distribuïdes regularment al llarg de tot l’any. Als deserts on les precipitacions cauen principalment durant l’estiu, com passa a la major part de deserts centrasiàtics, aquest avantatge es perd. Al Gobi, per exemple, on el màxim de precipitacions és indubtablement a l’estiu, al voltant del 90% del total anual de precipitacions es perd per evaporació física i no és possible, per tant, que sigui aprofitada pels organismes que viuen al desert. Aquest fet caracteritza també els deserts argilosos ben drenats, amb un relleu de pendents accentuats i una baixa permeabilitat del sòl. En aquest cas, el paper més important el fa l’escolament superficial, com a resultat del qual la major part d’aigua del desglaç surt del territori on havia caigut, per rius i rierols temporals.
Per tant, la majoria de deserts i subdeserts freds acumulen al sòl, durant el període fred, més de la meitat del total de precipitacions atmosfèriques anuals, aigua que, durant la primavera, amara un gruix prou considerable del sòl. Així, doncs, com a mínim durant un cert període de temps, aquests deserts no tenen dèficit hídric. Tota aquesta aigua és aprofitada per les plantes, que la consumeixen enla transpiració. Les plantes no aprofiten solament lahumitat dels horitzons del sòl que arriben a la superfície; n’hi ha moltes que, gràcies a un sistema radical profund, es beneficien de les aigües de la capa freàtica. Per això, el nivell de les aigües freàtiques disminueix considerablement durant l’estació vegetativa. És interessant remarcar que en el cas dels terrenys plans amb un relleu poc drenat, la profunditat del nivell freàtic és determinada per la profunditat de penetració del sistema radical de les plantes. El consum d’aigües freàtiques i el descens subsegüent del seu nivell continua fins que la zona del sòl impregnada d’humitat capil·lar provinent de les aigües subterrànies, perd el contacte amb les arrels de les plantes.
La disbauxa primaveral de les efímeres
La saturació d’humitat que experimenta el sòl durant la primavera, com també l’abundància de calor i llum, permeten un vigorós creixement de les plantes. És ben sabut que els deserts amb un dèficit hídric marcat es caracteritzen per la presència de plantes amb molts mecanismes morfològics i fisiològics per aprofitar la humitat amb eficàcia. No obstant això, als deserts freds, durant la primavera, les plantes que hi predominen no disposen d’aquests mecanismes: la tija herbàcia és fràgil i les fulles, d’un verd brillant, es marceixen ràpidament. S’assemblen més a les plantes pròpies dels terrenys humits que a les més típiques del desert. Manifestament aquest tipus de plantes no sofreixen cap manca d’humitat al llarg del seu període de creixement, i no requereixen, per tant, cap mecanisme especial per a economitzar el consum d’aigua. L’única particularitat que posseeixen és la capacitat per créixer i desenvolupar-se ràpidament. Aquestes plantes corresponen a una forma biològica peculiar dins de la vegetació del desert: la de les efímeres, és a dir, les plantes anuals (efímeres pròpiament dites) o amb parts persistents subterrànies (efemeroides) que inicien i completen el cicle vital amb gran rapidesa quan les condicions ambientals els són favorables. Les plantes efímeres estan amplament representades en la vegetació del desert i pertanyen a famílies molt diverses. En alguns deserts se’n poden trobar centenars d’espècies; als deserts de l’Àsia mitjana i el Kazakhstan, per exemple, n’hi ha de 300 a 350. Cal remarcar que aquesta forma de vida només es troba en aquells deserts en què el màxim de precipitacions o d’acumulació d’aigua té lloc durant l’estació freda. Als deserts amb el màxim de precipitacions a l’estiu, com els centrasiàtics, pràcticament no hi ha espècies efímeres.
Les plantes efímeres cobreixen el sòl del desert com una espessa catifa i aprofiten sobretot les capes superiors del sòl (mai més enllà dels 40 cm de fondària, normalment fins a uns 10) on hi ha la major part d’aigua no salina més accessible. El volum principal d’arrels (més del 95%) es troba cap als 10 cm de profunditat. El ràpid creixement, amb una transpiració intensa, permet el màxim aprofitament efectiu de la humitat d’aquestes capes del sòl. Com més de pressa aprofiten la humitat d’aquests horitzons les efímeres, menys aigua es perd en evaporació física, ja que són precisament els horitzons superiors del sòl els que hi estan més exposats. Així, en els deserts freds, les efímeres tenen un paper ecològic fonamental, impedeixen l’evaporació física i disminueixen substancialment el consum improductiu d’humitat del sòl durant la primavera.
Cal remarcar que el breu període d’abundància d’aigua no és només molt beneficiós per a les plantes del desert, sinó també ho és per als animals. En particular, durant aquest període, es dóna un excedent d’aliments altament nutritius i de fàcil accés per als herbívors. Gràcies al fet que els teixits de les efímeres retenen molt bé la humitat, aquests animals no necessiten altres fonts d’aigua, i el temps que no perden per buscar aigua i abeurar-se el poden aprofitar per a altres tipus d’activitats. És precisament durant aquest període que té lloc la reproducció de la majoria d’espècies d’invertebrats. Els ocells nien i les cries es desenvolupen en poc temps. Les tortugues es reprodueixen també en aquesta època, com també els rosegadors i els ungulats. L’activitat dels insectes i altres invertebrats és extremament elevada. Però tota aquesta explosió de vida s’acaba ràpidament. Ja al mes de maig, quan comença la secada, s’atura bruscament l’activitat de la majoria d’animals. Moltes espècies de vertebrats, com les tortugues i els suslics, comencen la hibernació, de la qual no despertaran fins a la primavera següent. Els invertebrats s’endinsen a la terra, a gran profunditat, i s’hi queden també fins a la primavera, realitzant una activitat mínima. Només algunes espècies romanen actives: aquelles que són capaces de subsistir en condicions de sequedat extrema.
Les arrels profundes i les fulles estalviadores
El començament del període de sequera estival es caracteritza pel gran dèficit d’humitat del sòl, però una part molt significativa de plantes del desert, al contrari dels efímers, continuen vegetant, és a dir, vivint activament. Aquestes plantes estan obligades a superar unes condicions absolutament desfavorables. Aprofiten al màxim les reserves d’humitat del sòl accessibles i el sòl s’asseca fins que assoleix l’anomenat punt de marcidesa permament. Encara hi ha una petita quantitat d’aigua al sòl, però està tan estretament retinguda al voltant de les partícules sòlides que resulta pràcticament inaccessible per a les plantes. En els sòls arenosos la quantitat d’aigua retinguda per les partícules és més petita i per això el punt de marcidesa permament correspon a una quantitat d’aigua al sòl pràcticament negligible: del 0,5 a l’1,5% del pes del sòl. Als sòls argilosos, l’aigua inaccessible està molt més lligada i el punt de marcidesa permanent arriba a la majoria de les plantes quan encara hi ha al sòl d’un 10 a un 12% d’aigua i, en el cas dels sòls argilosos salins, d’un 14 a un 15%. Així, doncs, és en els sòls arenosos i no salins on hi ha més aigua disponible per a les plantes. La utilització de tècniques agrícoles adreçades a dessalar els sòls (que sovint consisteixen a rentar-ne el perfil) incrementa considerablement la quantitat d’aigua disponible per a les plantes i augmenta la productivitat d’aquests sòls.
La insuficiència de l’aigua del sòl per a les plantes durant el període estival s’agreuja amb les altes temperatures de l’aire i els incessants vents abrusadors, secs i calents, que fan més gran la descàrrega d’aigua i produeixen la deshidratació dels teixits. En aquestes condicions, únicament poden existir formes especialitzades de plantes, amb mecanismes morfològics i fisiològics que evitin o redueixin la deshidratació; són formes de vegetació aptes per a extreure amb una màxima efectivitat l’aigua del sòl més profunda o de més difícil accés i de retardar-ne l’evaporació dels seus teixits, és a dir, d’economitzar-ne el consum. Els vegetals que s’inclouen dins aquest grup s’anomenen xeròfits, entre els quals hi ha també espècies amb un sistema d’arrels profundes, que arriben a les capes del sòl humides a tocar de les aigües subterrànies. En aquests casos, les arrels principals baixen verticalment com un fil de plomada i es ramifiquen només quan arriben al nivell freàtic. A aquest grup de plantes pertanyen formes tan típiques del desert com l’arç de camell (Alhagi persarum), que és una fabàcia, les arrels de la qual penetren a profunditats superiors a 5 o 6 m, o el repetidament esmentat sacsaül negre (Haloxylon aphyllum), amb arrels fins a 10 m de profunditat. L’aprofitament de les aigües freàtiques permet a aquestes plantes d’abastar-se d’aigua durant tot el període d’eixut. Un consum massa intensiu d’aquestes reserves subterrànies, tanmateix, pot arribar a ser perjudicial per a les mateixes plantes que les aprofiten en rebaixar-ne en excés el nivell i fer insuficient la recàrrega amb les precipitacions d’hivern i primavera, cosa que pot comportar la degradació i la transformació de l’estructura de la coberta vegetal en què predominen aquestes plantes d’arrels profundes.
Tot i això, a la majoria de deserts freds el sistema d’arrels de moltes de les plantes no arriba a les aigües subterrànies i per això es veuen forçades a proveir-se únicament amb les reserves d’aigua acumulades al sòl provinents de les precipitacions.
Durant la primavera, aquestes espècies, igual que les efímeres, es troben prou ben proveïdes d’aigua fàcilment accessible, però a diferència d’aquestes, el seu sistema d’arrels arriba a horitzons del sòl més profunds. Cap a l’estiu, aquestes plantes aprofiten fins al màxim les reserves d’humitat existents i assequen el sòl fins al punt de marcidesa permament. Això no obstant, com que continuen vives, és evident que els és imprescindible ni que sigui una mínima quantitat d’aigua per a mantenir-se. La majoria de xeròfits utilitzen petites quantitats d’aigua extretes de les retingudes al voltant de les partícules sòlides del sòl, de difícil accés, o bé es limiten a la petita quantitat d’humitat condensada que penetra al sòl en forma de vapor, a partir de l’aire o de les aigües freàtiques. Per a poder sobreviure en aquestes condicions, les plantes han de reduir al màxim l’evapo-transpiració. Molts xeròfits de fulles toves han de reduir severament l’àrea de les fulles. En aquest cas, les fulles grans i tendres, que perden fàcilment la humitat, són substituïdes per fulles més petites i rígides. Quan augmenta el dèficit d’aigua, les plantes es desprenen gairebé de totes les fulles, per tal de conservar la humitat dels brots. Altres xeròfits dels deserts freds, davant la insuficiència d’humitat, clouen els estomes, per tal de reduir o d’evitar l’evaporació de les fulles. Això, és clar, implica una reducció de la fotosíntesi i en el cas de secades molt llargues, les plantes comencen a passar gana: les fulles s’esgrogueeixen però generalment no arriben a morir.
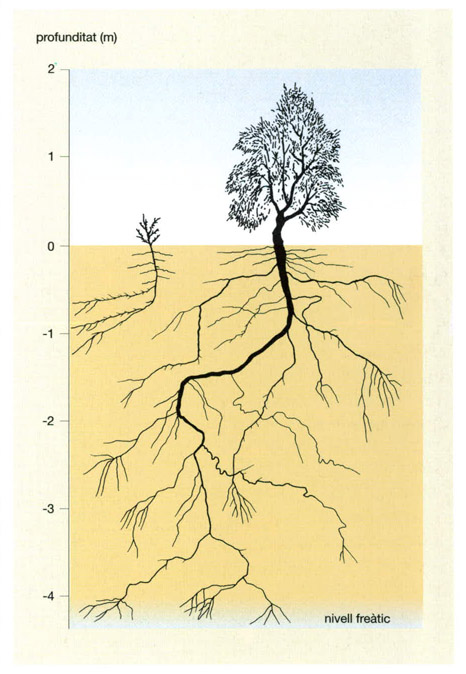
El sistema radical de dos exemplars de zaisan (Haloxylon ammodendron) del desert de Karakum, un d’entre 10 i 15 anys d’edat (a la dreta) i l’altre de només dos anys (a l’esquerra), mostra el grau de desenvolupament i la ramificació que assoleixen les arrels de les espècies xeròfites, les úniques que tenen una biomassa subterrània superior a l’aèria. El zaisan pot ser de port arbori o bé arbustiu, segons les condicions concretes en què la planta es desenvolupi. Quan les seves arrels arriben al nivell freàtic, normalment a una profunditat de 4 o 5 m, la planta pot créixer i fer-se grossa ràpidament. La competència que es fan entre ells els zaisans és, sobretot, en l’àmbit subterrrani, per tal d’accedir a l’aigua escassa. Per això, l’exemplar més jove dirigeix les seves arrels cap a la zona no ocupada pel sistema radical del seu germà més gran.
Jordi Corbera, a partir de Goodall, 1983
Una altra propietat molt comuna de les plantes del desert, que els permet disminuir l’evapo-transpiració, és la possibilitat de transformar completament alguns dels seus òrgans, en especial les fulles i els brots caulinars. A aquest tipus de plantes pertanyen les anomenades formes afil·les, que habitualment no tenen fulles i en les quals la funció d’aquestes és realitzada pels brots caulinars, per les tiges o per fulles modificades com si es tractés de rebrots d’origen caulinar, per exemple els sacsaüls (Haloxylon) o els “kandyms” (Calligonum). Moltes plantes del desert desenvolupen diferents tipus de cutícules que les protegeixen de l’evapo-transpiració excessiva; la majoria tenen també les fulles recobertes d’una pilositat espessa que embolcalla literalment les parts de la planta més exposades a l’evapo-transpiració. En altres casos el paper dels teguments protectors és a càrrec de diferents secrecions impermeables, com ceres, gomes o resines, que cobreixen aquestes parts de la planta.
En altres plantes, les halosuculentes, la retenció d’aigua es fa als teixits en forma de solució salina. S’inclouen en aquest grup moltes espècies de quenopodiàcies com les barrelles (Salsola) o les cirialeres (Salicornia). Però la sal, sobretot en concentracions altes, és perjudicial per a la planta i, per això, les halosuculentes gaudeixen d’una estructura interior que els permet minimitzar el dany causat per la sal. Aquestes plantes poden protegir-se parcialment de les solucions salines amb unes membranes cel·lulars especials i gràcies a la transpiració, en la qual expulsen la sal juntament amb l’aigua. Les plantes halosuculentes poden obtenir aigua pura, imprescindible per a la seva activitat vital, de les solucions salines, isolant una part de la sal en teixits especials. La reestructuració del règim hídric que duen a terme les halosuculentes és extraordinàriament notable, i la majoria són plantes que tenen un tipus especial de fotosíntesi, el vinculat al metabolisme C4.
Producció escassa, productivitat discreta
Així, doncs, gràcies a diferents mecanismes, les plantes del desert fred s’adapten a la sequedat i a la salinitat. La diversitat d’espècies i formes de vida que presenten els permet aprofitar al màxim la humitat de les reserves del sòl, fins i tot allí on aquestes són de difícil accés, com els horitzons profunds o els sòls salins. Es mantenen així vives durant els períodes de sequedat extrema i altes temperatures, i poden vegetar força temps, independentment del dessecament del sòl a l’època estival. Això no obstant, des del punt de vista de la producció de les plantes, aquest tipus de funcionament no és efectiu, ja que el període de vegetació és molt curt, inferior al de les plantes efímeres. Els mecanismes destinats a economitzar la humitat, per norma general, limiten la fotosíntesi de la planta i, consegüentment, el creixement de la massa orgànica és petit. Les plantes guanyen en resistència però la seva producció és escassa.
La biomassa i la producció
A la majoria de deserts freds, amb les precipitacions repartides al llarg de tot l’any o concentrades majoritàriament a les temporades d’hivern i de primavera, les plantes aprofiten sovint més de la meitat del total de precipitacions anuals (fins al 70%), per a la pròpia producció de biomassa; la pèrdua d’aigua en evaporació física improductiva és relativament petita. Això permet una productivitat força considerable, en termes relatius. Així, al subdesert de la Ciscàspia septentrional, amb una pluviositat anual de 250 a 300 mm, només la producció de la biomassa aèria representa al voltant de 1 600 kg/ha de massa seca, i alguns anys arriba als 2 000 kg/ha. Al desert del Karakum, amb una pluviositat de 100 a 150 mm, la producció mitjana de fitomassa aèria en diferents comunitats arbustives, subarbustives i herbàcies oscil·la entre 450 i 510 kg/ha, amb un màxim de 1 230 kg/ha. Si es prenen en compte les arrels i altres òrgans subterranis de les plantes, la producció total als diferents deserts de l’Àsia mitjana (Karakum, Kizilkum, petit Barsuki), pot variar de 200 a 1 400 kg/ha, segons el tipus de planta i la quantitat de precipitacions, i en aquests casos la part subterrània de la planta representa del 65 al 85% de la biomassa total. Els deserts amb la màxima quantitat de precipitacions a l’estiu es caracteritzen per una productivitat de la vegetació molt inferior. És el cas del Gobi que, amb una mitjana de precipitacions anuals de 130 a 150 mm, el 70% de les quals es produeixen a l’estiu, té una producció de fitomassa aèria no superior a 150-250 kg/ha en cap associació. Com s’ha assenyalat, en aquests casos, fins al 90% del total de precipitacions anuals es perd en evaporació física i no és a l’abast de la vegetació.
La biomassa i la producció
A la majoria de deserts freds, amb les precipitacions repartides al llarg de tot l’any o concentrades majoritàriament a les temporades d’hivern i de primavera, les plantes aprofiten sovint més de la meitat del total de precipitacions anuals (fins al 70%), per a la pròpia producció de biomassa; la pèrdua d’aigua en evaporació física improductiva és relativament petita. Això permet una productivitat força considerable, en termes relatius. Així, al subdesert de la Ciscàspia septentrional, amb una pluviositat anual de 250 a 300 mm, només la producció de la biomassa aèria representa al voltant de 1 600 kg/ha de massa seca, i alguns anys arriba als 2 000 kg/ha. Al desert del Karakum, amb una pluviositat de 100 a 150 mm, la producció mitjana de fitomassa aèria en diferents comunitats arbustives, subarbustives i herbàcies oscil·la entre 450 i 510 kg/ha, amb un màxim de 1 230 kg/ha. Si es prenen en compte les arrels i altres òrgans subterranis de les plantes, la producció total als diferents deserts de l’Àsia mitjana (Karakum, Kizilkum, petit Barsuki), pot variar de 200 a 1 400 kg/ha, segons el tipus de planta i la quantitat de precipitacions, i en aquests casos la part subterrània de la planta representa del 65 al 85% de la biomassa total. Els deserts amb la màxima quantitat de precipitacions a l’estiu es caracteritzen per una productivitat de la vegetació molt inferior. És el cas del Gobi que, amb una mitjana de precipitacions anuals de 130 a 150 mm, el 70% de les quals es produeixen a l’estiu, té una producció de fitomassa aèria no superior a 150-250 kg/ha en cap associació. Com s’ha assenyalat, en aquests casos, fins al 90% del total de precipitacions anuals es perd en evaporació física i no és a l’abast de la vegetació.
El cicle dels nutrients
La productivitat relativament alta dels deserts freds és possible, no sols per l’efectiu aprofitament que fan els vegetals de la humitat del sòl, sinó també per la riquesa en nutrients dels sòls desèrtics, especialment nitrogen, fòsfor i potassi. La gran reserva d’aquests elements és determinada per la ràpida mineralització de les restes de vegetació morta; com a resultat, les substàncies nutritives de la massa vegetal retornen al sòl de forma accessible a les plantes, i són aprofitades per a la síntesi de nova producció orgànica. Ja s’ha esmentat que el sòl del desert és ric en microorganismes resistents a les altes temperatures i la sequedat, que permeten la ràpida mineralització de les substàncies vegetals orgàniques, i que és ric també de bacteris fixadors de nitrat. Aquests microorganismes es caracteritzen per funcionar amb més intensitat quan les èpoques de sequera i humitat s’alternen periòdicament. Precisament durant els períodes humits que segueixen la temporada seca té lloc un inusual i ràpid augment del nombre de microorganismes i un sobtat esclat de la seva activitat. Una explicació a aquest fet pot ser que durant el període sec i calorós són destruïdes les substàncies inhibidores, que sovint frenen el creixement dels microorganismes. Cal dir, a més, que els microorganismes reaccionen a la humitat més ràpidament que els vegetals; només algunes hores després de la pluja comencena desplegar una activitat vital extraordinària i a multiplicar-se veloçment. Aquesta alta activitat delsmicroorganismes, inesperada al desert, es manifesta particularment en la ràpida mineralització que duen a terme de les restes vegetals mortes.
A més dels microorganismes, també els vertebrats tenen un important paper en la ràpida mineralització de la matèria orgànica vegetal. Com ja és sabut, al desert habiten molts vertebrats herbívors, sobre tot rosegadors i ungulats. Tots ells consumeixen una gran quantitat de massa vegetal; durant el període vegetatiu de les plantes, se’n mengen bàsicament les parts verdes vives, però durant la temporada seca, en consumeixen també els brots secs, morts. La matèria vegetal orgànica consumida es redueix a molècules orgàniques i minerals senzilles durant el procés de digestió i intercanvi de substàncies en l’organisme de l’animal, i reapareix a l’exterior en forma accessible a les plantes. Només una petita part, del 10 al 40%, queda a l’exterior en forma de restes de plantes no digerides ni mineralitzades. En els llocs on els rosegadors i els ungulats són abundants, aquests animals mineralitzen directament cap al 30% de tota la producció vegetal aèria dels deserts freds. D’aquesta manera, tant els vertebrats herbívors com els microorganismes tenen un paper fonamental en els processos bioquímics dels ecosistemes d’aquest bioma, ja que realitzen la mineralització de la matèria orgànica i permeten reciclar els nutrients; com a resultat de tot aquest procés, les plantes reben la substància nutritiva imprescindible per al seu creixement.
L’alta eficiència dels consumidors
Per a completar el panorama del funcionament eco-lògic dels deserts freds, cal parlar especialment de la funció dels consumidors, és a dir de la fauna, una funció lligada a l’estructura específica dels animals que hi viuen que, en els deserts freds, és força diferent que en altres biomes. Als biomes més humits, com estepes o boscos, els animals dominants són els anomenats sapròfags, els invertebrats del sòl que s’alimenten de matèria orgànica morta i representen del 80 al 98% de la massa animal total d’aquests ecosistemes. En els deserts i els subdeserts freds, en canvi, predominen els fitòfags, que s’alimenten bàsicament de les parts vives de les plantes. Els sapròfags no constitueixen en deserts i subdeserts més enllà d’un terç de la massa animal total.
L’elevada proporció de vertebrats
Cal remarcar que, entre els fitòfags de deserts i subdeserts, el lloc fonamental l’ocupen els vertebrats herbívors, i especialment els mamífers. La magnitud de la biomassa total de vertebrats no és inferior a la que es troba en boscos i estepes, més rics en vida, i pel que fa a la relació entre la biomassa animal i la producció de biomassa vegetal, és considerablement més alta. Així doncs, si en els deserts i els subdeserts d’argila, la biomassa de vertebrats assoleix de valors 3,7 a 6,1 kg/ha, en els boscos mixtos de coníferes i planifolis és tot just de 2,2 a 5,5 kg/ha, i únicament en els ecosistemes de boscos esteparis més rics i de boscanes decídues (com per exemple les rouredes) arriba a 12,9 kg/ha. Al mateix temps, la relació d’aquesta biomassa de vertebrats amb la producció, és a dir, el creixement anual de la biomassa vegetal als deserts i els subdeserts és força més alta: de 0,9 a 1,2; mentre que als boscos i els boscos esteparis no puja de 0,2.
Aquestes xifres donen testimoni de la considerable pressió dels animals herbívors sobre la vegetació, i del seu important paper en els ecosistemes dels deserts i els subdeserts freds. Els animals herbívors consumeixen, efectivament, una gran part de la biomassa orgànica produïda per la vegetació. Si es té en compte el consum, no sols de la massa vegetal viva, sinó també dels brots morts durant els períodes de secada, en què el procés de vegetació de les plantes s’alenteix o es detura del tot, llavors la quantitat de producció de plantes superficials consumida pels animals a molts deserts i subdeserts arriba al 60 o 70%, i de vegades més.
La importància de la brostejada
La influència dels animals té, doncs, un gran significat ecològic per als ecosistemes desèrtics. Ja s’ha assenyalat l’important paper d’aquesta activitat en la mineralització de la massa vegetal i el cicle biològic dels nutrients, però encara és més important per al creixement i el desenvolupament de les plantes mateixes. És ben sabut que una brostejada excessiva pot ser mortal per a les plantes, disminuir-ne la productivitat i portar a la degradació total de la capa vegetal; aquest fet es pot observar sovint quan el nombre d’animals salvatges augmenta o hi ha un excés de ramats pasturant. Un augment de la pressió en els animals fitòfags fa que les plantes absorbeixin menys humitat del sòl. A les pastures dels deserts, on la vegetació és consumida amb intensitat pels animals, el sòl gaudeix d’humitat superficial i de reserves d’aigua no aprofitades, fins i tot durant l’època de secada. Aquestes reserves estan protegides de l’evaporació física per una disposició força profunda de les capes humides i per l’absència de lligams capil·lars amb la superfície; la humitat aprofitada per les plantes no és completa perquè és explotada per la pressió dels animals. Per exemple, al subdesert de la Ciscàspia septentrio-nal, la brostejada selectiva per part dels mamífers herbívors de la barrella terrera (Bassia [=Kochia] prostrata), l’arrel de la qual sovint penetra el sòl a una profunditat de 50 cm a 2 m, hi determina la preservació de la humitat del sòl a un nivell del 17%, és a dir, força per sobre del punt de marcidesa permament. Al mateix temps, en els llocs protegits d’animals, amb vegetació intacta, el contingut d’humitat en el gruix del sòl baixa sovint fins al 14-15%, arriba al punt de marcidesa permament i indica clarament que tota la humitat accessible és aquí totalment consumida per les plantes.
Per altra banda, és sabut que la biomassa vegetal no resulta perjudicada per un brosteig moderat dels animals. A més, quan la biomassa vegetal disminueix a causa dels animals que la pasturen o la brostegen, en condicions de dèficit extrem d’humitat i de sequedat intensa de l’aire, disminueix considerablement la capacitat d’evapo-transpiració de les plantes, les protegeix de la deshidratació i en redueix el risc de mortalitat. Ja s’ha assenyalat que, en condicions de secada, moltes plantes desenvolupen mecanismes per a reduir l’àrea de les fulles i la massa dels teixits vius, amb la qual cosa aconsegueixen un menor consum d’humitat. Això, tanmateix, no sempre resulta suficient i són els animals els qui separen mecànicament els teixits vius i permeten que la planta sobrevisqui. Aquest fenomen es dóna molt sovint a la naturalesa i, els anys particularment eixuts, que sobrevenen periòdicament, poden salvar les plantes de la mort. Al subdesert dela Ciscàspia septentrional, per exemple, l’any 1972, en què la secada va ser particularment intensa, allí on la vegetació era protegida de la pastura, van morir gairebé totes les plantes autòctones, fins i tot xeròfits altament adaptats als deserts, com el donzell pauciflor (Artemisia pauciflora), la barrella ter-rera (Bassia [=Kochia] prostrata), etc. D’altra banda, aquest mateix tipus de plantes situades als llocs pasturats, brostejades a fons i amb una biomassa considerablement menor, es van conservar en condicions de secada i van sobreviure amb èxit.
El paper del trepig
Al desert, adquireixen també un important significat les formes mecàniques de l’impacte animal sobre la capa vegetal. És prou coneguda la influència dels ungulats, que provoquen la compressió dels sòls argilosos, en redueixen la permeabilitat i, consegüentment, n’augmenten l’escolament superficial. Els sòls arenosos i francs també són sovint malmesos per l’acció de les peülles. Tanmateix, l’impacte causat pels ungulats no es pot interpretar de manera simplista. És sabut que quan les peülles dels animals estoven moderadament la superfície del sòl, les llavors s’hi poden fixar més profundament, i això les protegeix dels impactes negatius externs. És també important el fet que aquests impactes destrueixen sovint la crosta d’algues, líquens i molses que sovint creixen a la superfície del sòl i, consegüentment, milloren les condicions perquè germinin les llavors, augmenten la permeabilitat del sòl i creix la productivitat de les plantes altes. En els deserts i els subdeserts argilosos, la destrucció de la pel·lícula d’algues a les zones de pastura fa que la permeabilitat del sòl sigui de bon tros superior que a les zones protegides de la pastura. D’acord amb les mesures que s’hi han fet, a les zones del subdesert de la Ciscàspia septentrional on els prats van ser tancats als ramats durant un període de 30 anys, el coeficient de filtració dels sòls francs era només de 0,05 mm/min, a causa de la pel·lícula d’algues desenvolupades a la superfície; a les zones de pastura, en canvi, on la pel·lícula és destruïda constantment per les peülles dels animals, la filtració era de 0,24, és a dir, cinc vegades superior.
Una dinàmica reforçadora de la desertificació
La dinàmica dels ecosistemes dels deserts freds, igual que la de qualsevol altre ecosistema, depèn de factors molt diversos. Hi ha, per exemple, les fluctuacions de curta durada, lligades a les oscil·lacions climàtiques i atmosfèriques, que abasten la durada d’una estació o un any i es manifesten en les desviacions de la humitat (la quantitat de precipitacions) o en la quantitat de calor normal (la mitjana anual). També hi ha canvis de llarga durada, que són els que es produeixen a una escala de temps geològica. No es tracta únicament, com podria pensar-se, dels canvis associats amb les variacions globals del clima, sinó també dels determinats pels processos tectònics, ja que en la dinàmica i la formació dels deserts, hi té un paper particularment important la formació de muntanyes, que aïlla certes regions de les influències oceàniques i les converteix en zones àrides. Finalment, també hi intervenen factors intrínsecs a la mateixa dinàmica ecològica del bioma, concretament a processos de successió d’alguna manera mancats o que no poden ultrapassar uns llindars de producció o d’acumulació de biomassa que són els compatibles amb els recursos locals.
Els canvis vinculats a factors extrínsecs
En la llarga durada, però, la dinàmica dels ecosistemes dels deserts està lligada principalment amb els canvis cíclics del clima terrestre. Sol considerar-se que al llarg del Quaternari i fins al present ha tingut lloc un notable augment de l’aridesa del clima, que encara es manté actualment i que s’associa a la dessecació progressiva dels deserts. Existeixen nombrosos factors que testimonien un grau d’humitat més alt dels deserts en èpoques passades no gaire allunyades. Per exemple, a les zones d’aridesa extrema del desert de Gobi on actualment no viu ningú, s’han trobat una gran quantitat de restes d’assentaments humans neolítics. Per les dades que es posseeixen, l’àrea dels llacs de la zona àrida d’Àsia s’ha fet de 6 a 10 vegades més petita durant el Quaternari. Aquest i altres factors permeten parlar d’una dessecació natural progressiva de les regions desèrtiques.
Malgrat tot, cal tenir en compte que els processos de dessecació no estan necessàriament lligats als canvis climàtics. De fet, no hi ha evidències d’un augment global de l’aridesa del clima durant el Quaternari. L’increment de la sequera als deserts podria ser resultat d’una simple dessecació local dels territoris desèrtics, sense cap canvi climàtic global. Actualment, els processos de dessecació s’intensifiquen a causa de l’acció antròpica. L’excés de pastura provoca la degradació de la vegetació, ja que els ramats destrueixen amb les peülles els febles sòls arenosos, o bé, al contrari, provoquen la compressió dels durs sòls d’argila; tot això comporta la destrucció dels ecosistemes dels deserts, i és la causa d’una desertització creixent. L’indicador principal és el descens de la productivitat biològica i la degradació de l’ecosistema.
Els canvis vinculats a factors intrínsecs
No són únicament els impactes externs els responsables de la dinàmica dels ecosistemes dels deserts freds. Els mateixos impactes antròpics no poden ser considerats extrínsecs si procedeixen de les poblacions residents. En tot cas, són també fonamentals els canvis en l’ecosistema associats al seu propi desenvolupament, és a dir, a la dinàmica successional pròpia de tot ecosistema.
Molses abassegadores
En els deserts freds, una fase successional molt característica associada a l’absència d’explotació pels herbívors és la que consisteix en la formació d’una capa de molses i líquens o de molses i algues a la superfície del sòl. Generalment l’espècie dominant en aquesta mena de crostes és la molsa Tortula caninervis (=T. desertorum) o alguna de propera. Un antic experiment de camp al desert arenós de Karakum, que va consistir a excloure els ramats, va demostrar que en llocs on originàriament hi havia una absència completa de molsa, al cap de 8 anys se n’havia format una capa que, segons les condicions del lloc concret, podia ocupar d’un 10 a un 40% de la superfície del sòl; al cap de 15 anys podia ocupar d’un 20 a un 50% i, passats 25 anys, cobria ja del 25 al 70% de tota l’àrea. La massa de molsa, en aquest temps, havia assolit els 980 kg/ha, i el gruix podia oscil·lar entre 0,5 i 2 cm.
Aquesta crosta de molsa actua negativament sobre les plantes vasculars: la seva productivitat baixa gradualment i es conserven només en un estat molt poc ufanós. Per exemple, la producció de les bosquines de sacsaül blanc (Haloxylon persicum), es redueix de dues a dues vegades i mitja per l’efecte de la crosta de molsa, mentre que les brolles i gramenets la redueixen en un 30 o un 40%. Les plantes llenyoses i les herbàcies perennes es marceixen, i la ciperàcia Carex physodes, una de les espècies sovint dominants als deserts arenosos, és probablement la més afectada.
El mecanisme de l’impacte de la crosta de molsa consisteix, bàsicament, a interceptar les precipitacions atmosfèriques. La crosta de molsa (o de molsa i liquen), gràcies a la seva elevada higroscopicitat, pot retenir del 30 al 40% de tota la humitat de les precipitacions i, en alguns casos, fins al 80%. La molsa aprofita una part d’aquesta humitat, però la major part s’evapora i es perd per a les plantes. A més, la crosta de molsa canvia substancialment les condicions de temperatura a les zones on creix vegetació herbàcia. La molsa reté la irradiació de calor del sòl i, consegüentment, augmenta de manera considerable la temperatura de les capes d’aire immediates a la superfície d’aquest, fet que provoca el sobreescalfament de les fulles basals o fins i tot de la planta sencera en el cas de moltes plantes herbàcies, amb totes les conseqüències negatives que això comporta.
La molsa, a diferència de les plantes vasculars, es caracteritza per una resistència molt gran als factors extrems, les temperatures altes i baixes i la dessecació, i suporta més fàcilment la deshidratació. Un dels avantatges fonamentals de la molsa és la capacitat de créixer amb temperatures ambientals baixes i fins i tot negatives, amb la qual cosa acompleix el seu desenvolupament abans que altres plantes. Al desert de Karakum, el procés vegetatiu de la molsa comença 50 o 60 dies abans que el de les espècies herbàcies. És molt important la disposició superficial dels rizoides de la molsa, que li permeten aprofitar fins i tot les precipitacions més insignificants. Tot això confereix a la molsa una significativa superioritat davant d’altres espècies, i li permet guanyar en la lluita per la humitat. En poques paraules, la crosta de molsa suplanta les plantes vasculars, ocupant tot l’espai dels deserts d’arena; sota la seva influència té lloc una transformació de la coberta vegetal, que veu disminuïda considerablement la productivitat. La funció de la crosta de molsa consisteix, sobretot, a activar els processos de dessecació dels ecosistemes, que s’esdevé com a resultat d’una utilització menys eficient, per part de les plantes, de l’aigua de les pluges; provocat, principalment, pel considerable increment de les pèrdues en evaporació física improductiva. El resultat de tot aquest procés és la desertització de l’ecosistema.
Sacsaüls assedegats
Una altra forma de successió ecològica pròpia dels ecosistemes del deserts, està vinculada amb l’estat del nivell freàtic. Es dóna principalment en les bosquines de sacsaül negre (Haloxylon aphyllum) dels deserts de l’Àsia mitjana i el Kazakhstan. El sacsaül negre és un gran matoll arborescent que fa de6 a 10 m d’alçada. Molt sovint, les bosquines de sacsaül es troben a les depressions (per exemple, ales de rereduna del desert arenós de Karakum), on les aigües subterrànies es troben relativament a prop de la superfície (de 6 a 10 m) i són aprofitades per aquesta planta. Quan floreixen, els sacsaüls formen bosquines compactes amb capçades atapeïdes que arriben a un recobriment del 40 al 45%, i una elevada productivitat. En aprofitar les aigües subterrànies, el sacsaül negre no depèn de les precipitacions, i manté l’activitat vegetativa durant l’època de secada estival. Això, però, comporta una gran despesa d’aigües subterrànies i es reflecteix en la davallada del nivell freàtic. Al desert de Karakum (a la reserva de natura de Repetek), una bosquina de sacsaüls consumeix, durant un any, fins a 320 mm d’aigua, una quantitat diverses vegades més gran que la de la pluviositat. El nivell freàtic davalla de 10 a 12 cm durant el període vegetatiu. Durant l’hivern i la primavera s’omplen de nou gràcies als torrents d’aigua de les àrees circumdants, tot i que mai no acaben d’assolir el nivell anterior, de manera que el de cada primavera és inferior al de la primavera precedent. A les bosquines de sacsaül blanc de la reserva natural de Repetek, el nivell de les aigües subterrànies bai-xa cada any 1 cm de mitjana; del 1930 al 1970, el nivell de les aigües subterrànies va baixar fins a30 cm, tot i que la mitjana de precipitacions atmosfèriques anuals va ser relativament estable.
El descens del nivell freàtic comporta un augment de la mineralització. Tot plegat es reflecteix negativament en l’estat de la bosquina de sacsaüls. A mesura que creix la dificultat d’accedir a l’aigua, a causa del descens del nivell freàtic i l’augment de la concentració de sals minerals, de 5-6 g/l fins a 7-8 g/l, els boscos de sacsaül comencen a esclarissar-se, i quan la concentració és de 12 a 14 g/l té lloc la mort en massa del sacsaül blanc. A les bosquines de sacsaül negre (Haloxylon aphyllum), quan la mineralització de les aigües subterrànies arriba a 16 g/l, el nombre d’exemplars de sacsaül morts és del 40 al 60%. Per acabar, es pot afirmar que aquest procés finalitza amb la completa degradació de les bosquines de sacsaül i una reestructuració de la comunitat vegetal.
Els ecosistemes dels deserts freds es caracteritzen,doncs, per processos successionals, lligats a la inter-acció interna dels seus components que condueixen a una reorganització substancial de l’estructura i el funcionament de l’ecosistema i, com a norma general, comporten la dessecació, l’increment del dèficit hídric, la caducitat i la degradació de les comunitats vegetals desèrtiques.