El truc de l’esclerofil·lisme
Cada combinació de factors ambientals pot ser afrontada amb diferents estratègies, però una d’elles sol ser l’òptima i normalment acaba dominant. En el cas dels vegetals, els factors climàtics condicionen l’estructura i la durada de les fulles, caràcters bàsics de resposta en front de la distribució de temperatures i precipitacions al llarg de l’any. En ambients en què les temperatures no són limitadores en cap moment i en què l’aigua abunda sempre, els vegetals són perennifolis i tenen fulles relativament grans i poc endurides, anomenades malacofil·les. Aquest tipus domina en biomes tropicals humits i temperats humits en què es desenvolupen, per exemple, les pluviïsilves i les larisilves. Quan el fred o la secada actuen com a limitadors en algun període de l’any, apareixen vegetals malacofil·les però caducifolis, és a dir, que perden les fulles durant el període desfavorable. Aquest és el cas dels caducifolis d’estiu de climes subdesèrtics i dels caducifolis d’hivern dels climes temperats freds. Quan el període desfavorable és molt llarg, l’estratègia anterior no funciona, ja que no hi ha temps de fabricar fulles noves i amortitzar-les en el curt període d’activitat vegetativa, i cal recórrer a una estratègia de resistència. En aquests casos, dominen els vegetals perennifolis i aciculifolis com les coníferes, amb fulles capaces de resistir inactives les llargues èpoques de fred excessiu, tal com succeeix en la taigà. En la major part del bioma mediterrani es donen un conjunt de factors que han provocat l’aparició d’una altra estratègia diferent, la dels perennifolis esclerofil·les. Aquesta estratègia fa front a un ambient mai clarament limitador però molt sovint proper al límit, amb una irregularitat i una impredicibilitat considerables.
Les característiques funcionals
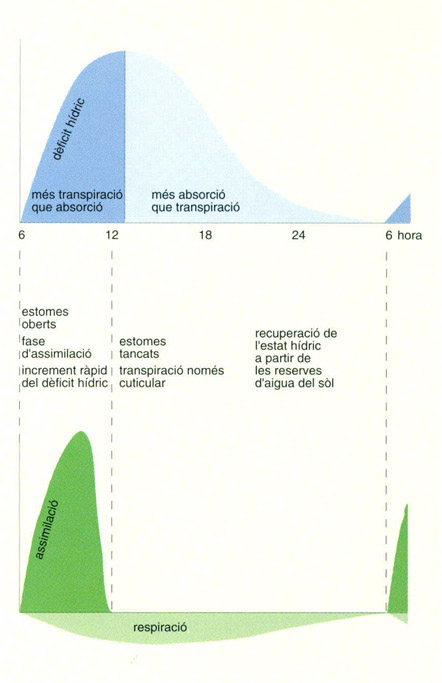
Evolució diària de l’activitat vegetal d’una planta esclerofil·la, un dia d’estiu. En avançar l’estació eixuta, la fase d’assimilació s’escurça perquè les condicions hídriques esdevenen limitants cada cop més de pressa. En les condicions més adverses, la respiració total pot ser superior a l’assimilació; aquesta pot arribar, fins i tot, a anullar-se.
Editrònica
Els factors ambientals limitadors en el bioma mediterrani són la secada estival i el fred hivernal. Això vol dir que, excepció feta de les èpoques humides i fresques de primavera i tardor, els ecosistemes mediterranis s’enfronten de manera quasi constant a un ambient que imposa unes restriccions que els mantenen quasi sempre al ralentí, ja que en pocs casos són prou fortes per a provocar una aturada total de l’activitat biològica. El caràcter perennifoli de la vegetació mediterrània típica respon bàsicament a aquest tipus de funcionament, ja que permet iniciar l’activitat tan bon punt les condicions ambientals deixen de ser limitadores, i això pot succeir en qualsevol moment de l’any a causa de la irregularitat climàtica inherent al bioma. En experiències de laboratori, s’ha demostrat la ràpida capacitat de resposta dels vegetals esclerofil·les, alguns dels quals comencen a fotosintetitzar 10 minuts després d’una irrigació precedida d’un període de secada realment sever que les mantenia en total inactivitat. Aquesta afirmació, de validesa general, admet matisacions derivades de les característiques dels diferents subclimes del bioma, ja que tant la durada del període eixut com la intensitat del fred tenen una gamma de variació considerable i només en determinades regions de marcada continentalitat, com per exemple a les parts centrals de Síria, ambdós factors actuen de manera rigorosa.
Sota aquestes condicions no hi ha més remei que mantenir les fulles tot l’any per anar aprofitant els curts períodes favorables que poden aparèixer en determinats moments, cosa que explica el caràcter perennifoli de la majoria de la vegetació del bioma. D’altra banda, les fulles han de suportar freds i secades, cosa que obliga a un disseny estructural especialment resistent. Les fulles esclerofil·les es caracteritzen per cutícules gruixudes, un parènquima en palissada especialment desenvolupat, un alt contingut en materials estructurals forts com la cellulosa i la lignina, i un nombre reduït d’estomes situats en criptes parcialment ocloses per pèls. Aquestes fulles són poc propenses a perdre aigua per evaporació cuticular, controlen de manera molt efectiva la transpiració per obertura i el tancament d’estomes, resisteixen bé mecànicament la pèrdua d’aigua i suporten temperatures inferiors als 0°C sense patir danys. Són, diríem, unes fulles tot terreny, més semblants a un 4x4 que a un utilitari o a un cotxe de luxe, amb tots els avantatges i inconvenients que això implica.
La transpiració i la captació de la llum
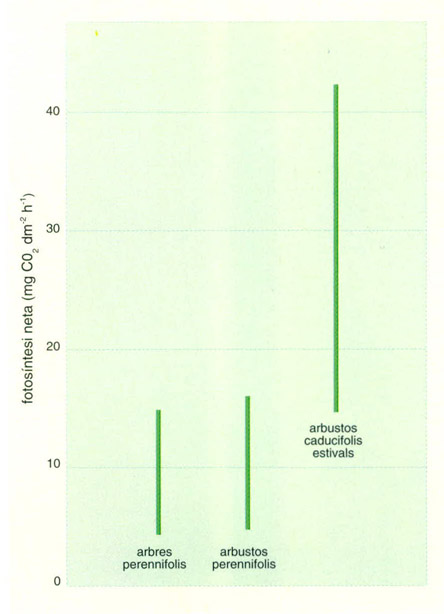
Capacitat fotosintetitzadora de les plantes de clima mediterrani mesurada principalment sobre la planta viva (al centre, espècies arbustives perennifòlies; a l’esquerra, arbres perennifolis; a la dreta, caducifolis estivals).
Editrònica
El control de les pèrdues d’aigua, tal com ja hem mencionat, és doble. La cutícula gruixuda actua com a aïllant i es combina amb una baixa densitat estomàtica. En una fulla malacofil·la, l’àrea ocupada pels estomes representa del 0,5 a l’1,5% d’àrea foliar total, mentre que en una esclerofil·la només representa del 0,2 al 0,5%. La duresa estructural permet pèrdues d’aigua importants sense que el marciment provoqui danys mecànics irreversibles, i així, mentre que en molts vegetals malacofil·les la pèrdua del 30% de l’aigua foliar comença a produir danys, molts esclerofil·les suporten pèrdues de fins al 70%. Aquesta mateixa estructura els permet resistir temperatures properes als -10°C. Ara bé, amb una estructura tan resistent no es pot anar de pressa, cosa que es tradueix en unes capacitats fotosintetitzadores relativament baixes. Les mateixes barreres que limiten la pèrdua d’aigua també posen dificultats al diòxid de carboni per a arribar als cloroplasts, i l’alt contingut en materials estructurals representa una proporció baixa de substàncies proteiques actives com els enzimes fotosistetitzadors. Així, mentre que una fulla malacofil·la té unes taxes fotosintètiques que oscil·len entre 15 i 40 mg de diòxid de carboni per decímetre quadrat i hora i el seu sistema fotosintetitzador no se satura per il·luminacions altes, una fulla esclerofil·la s’ha de contentar amb taxes de 4,5 a 16 mg de diòxid de carboni per decímetre quadrat i hora, i arriba a la saturació per il·luminacions inferiors a la màxima ambiental.
En el mateix bioma, però, la fixació total anual de diòxid de carboni dels caducifolis estivals i els esclerofil·les és semblant, si bé els primers exhibeixen taxes de fotosíntesi clarament superiors i els segons ho compensen amb una activitat més perllongada. En conjunt, el problema per als vegetals mediterranis és el mateix: els sobra llum i els manca aigua.
Per a entendre els avantatges derivats de la morfologia esclerofil·la, es pot comparar el comportament de dos arbres típicament mediterranis com l’alzina (Quercus ilex), perennifoli de fulles esclerofil·les, i el roure martinenc (Quercus pubescens), caducifoli hivernal. El segon comença el tancament estomàtic quan el contingut foliar d’aigua baixa per sota del 90% respecte de la saturació, i efectua un tancament total quan aquest valor arriba al 70%, però continua perdent aigua per transpiració cuticular a un ritme que equival al 50% de la transpiració màxima. L’alzina, en canvi, comença el tancament estomàtic quan el contingut hídric arriba al 85% de la saturació, i fa un tancament total quan arriba al 70%, de manera molt semblant a com ho fa el roure però amb la diferència que la transpiració cuticular representa, només, el 3% de la total. Això, juntament amb la capacitat per a suportar pèrdues d’aigua superiors sense sofrir danys, permet a l’alzina resistir períodes de secada 15 vegades més llargs que els que pot resistir el roure martinenc.
El cost estructural
El caràcter robust de les fulles esclerofil·les es tradueix en un elevat pes específic, és a dir, un alt contingut de matèria orgànica per unitat de superfície. Així, mentre que molts vegetals malacofil·les tenen un pes específic menor de 10 mg per centímetre quadrat, els esclerofil·les poden arribar als 20-40 mg per centímetre quadrat. Una estructura resistent pot ser més duradora, però també ha de durar temps llargs perquè és més cara de produir i més difícil d’amortitzar, sobretot si tenim en compte que la mateixa resistència estructural imposa restriccions a la màxima taxa de fotosíntesi que es pot assolir. Construir 1 g de fulla esclerofil·la representa un cost energètic equivalent a 1,7 g de glucosa, molt superior al necessari per a construir 1 g de fulla malacofil·la i fins i tot més elevat que el de construcció d’una fulla aciculifòlia de la taigà, preparada per a suportar les baixíssimes temperatures hivernals i que només arriba a 1,3 g de glucosa.
Quan una cosa és molt cara i poc productiva s’ha de fer durar, i és per això que els vegetals esclerofil·les són perennifolis, és a dir, a l’hivern mantenen les fulles i la longevitat foliar és així superior a 1 any, la mínima necessària per a assegurar la perennifòlia. L’índex d’esclerofília, o sigui, la relació ponderal entre lignina i cel·lulosa amb relació a les proteïnes, té valors de 230-450 per als esclerofil·les, enfront dels 80-180 dels caducifolis hivernals mediterranis.
Les fulles esclerofil·les són, doncs, relativament pobres de nutrients essencials ja que tenen un contingut proteic baix i són, per tant, poc costoses de construir en l’aspecte nutricional, però també són difícils de descompondre, de tal manera que es resisteixen a alliberar els pocs nutrients que contenen. Aquesta característica pot derivar-se del seu origen evolutiu, situat per molts autors en ambients oligotròfics anteriors a l’aparició del clima mediterrani. Sigui quin sigui el seu origen, aquest conjunt de grandeses i misèries associades a l’esclerofil·lisme l’han convertit en una solució òptima als problemes ambientals que planteja el clima mediterrani i han esdevingut el tret més característic dels paisatges del bioma.
Mai parats, quasi sempre al ralentit
Els ecosistemes del bioma mediterrani són, en conjunt, poc esponerosos i productius comparats amb d’altres de climes més càlids com els tropicals, o més freds com els temperats, ja que acusen aquesta constant restricció ambiental que, bé a causa de la manca d’aigua o de la baixa temperatura, els obliga a funcionar a un ritme lent.
La biomassa
La quantitat de matèria orgànica acumulada per un ecosistema, anomenada biomassa, és un paràmetre que indica el grau de complexitat que assoleix. Els factors ambientals, bàsicament els climàtics com temperatura i disponibilitat d’aigua, però també els edàfics com la disponibilitat de nutrients, són els que determinen el grau màxim de desenvolupament dels sistemes terrestres. En el cas del bioma mediterrani, amb les limitacions ambientals ja comentades, aquest desenvolupament és inferior al dels sistemes forestals temperats i superior al dels deserts, tal com correspon a la seva situació de transició entre tots dos biomes.
En general, quan la pluviositat ultrapassa els 400 o els 500 mm anuals, es desenvolupen ecosistemes dominats per vegetació arbòria, mentre que en cas contrari se’n troben d’altres dominats per arbustos. En aquest segon cas, la biomassa acumulada és relativament modesta, i ateny des de valors inferiors a 1 t ha–1 (tona per hectàrea) en el cas de les formacions arbustives de la zona de transició cap als deserts, fins a les 50 o 60 t ha–1 (tona per hectàrea) en les més desenvolupades, que correponen a la transició cap a les formacions forestals. Es fa molt difícil assignar valors a cadascun dels tipus d’ecosistemes arbustius del bioma, ja que, tot i que els factors climàtics i edàfics tenen un pes molt considerable, d’altres com la recurrència d’incendis o la història d’explotació poden tenir un paper encara més determinant. En cap cas, però, i suposant ecosistemes arbustius no afectats pel foc ni per l’explotació, no s’assoleixen biomasses molt superiors a les 100 t ha–1.
En el cas dels boscos mediterranis es donen valors superiors als esmentats, tot i que si han estat sotmesos a explotació o es troben en fase de regeneració després d’un foc, es poden trobar valors inferiors als màxims esmentats per als arbustius. És molt rar, en el bioma mediterrani, que hi hagi boscos que no hagin estat sotmesos a una explotació continuada que els manté en un estadi ben allunyat del seu màxim desenvolupament possible, i això contribueix a enfortir la idea que es tracta d’un bioma amb boscos magres. De tota manera, en els pocs casos de boscos ben conservats, s’ha comprovat que poden assolir-se valors de biomassa propers a les 300 t ha–1, semblants als de molts ecosistemes forestals d’altres biomes. Aquest seria el cas d’alguns magnífics alzinars situats en àrees poc poblades de la conca mediterrània com les muntanyes sardes, i que ens han arribat pràcticament verges d’explotació forestal, o de les restes dels esponerosos boscos australians de “jarrah”, amb arbres que poden assolir els 40 m d’alçada i que es mantenen actius durant l’estiu gràcies a la seva profunda arrelada. Les restriccions ambientals es tradueixen, doncs, en la lentitud de renovació d’aquesta biomassa més que no pas en el seu valor. Dit d’una altra manera, els boscos mediterranis poden assolir valors elevats de la biomassa però tenen una producció baixa que es tradueix en unes taxes de renovació i acumulació relativament baixes i clarament inferiors a les que es poden trobar, per exemple, en un bosc temperat o en una pluviïsilva.
Tots els valors de biomassa esmentats fins ara fan referència a tones de matèria orgànica seca, és a dir sense comptar l’aigua, acumulades en les parts aèries dels vegetals. Les dades de biomassa animal són escasses, i en cap cas no representen una fracció considerable, quantitativament parlant, de la total, tot i que la seva funció pot ser fonamental. Quant a la biomassa vegetal subterrània, les dades són poc abundants a causa de les complicacions metodològiques del seu estudi, tot i que quantitativament poden representar una fracció important de la biomassa total. En un bioma on la secada estival constitueix un factor ambiental determinant, hom pot esperar un bon desenvolupament de les parts subterrànies encaminat a aconseguir un subministrament d’aigua adequat i constant. La biomassa subterrània pot ser superior a l’aèria en el cas de les comunitats arbustives dels llocs més secs del bioma. En la majoria de les formacions forestals, però, la relació biomassa subterrània/biomassa aèria és inferior a 1. La biomassa subterrània no és, en cap cas, menyspreable, ja que fins i tot en boscos ben desenvolupats com un alzinar, amb una biomassa aèria de 170 t ha–1, les arrels poden arribar a les 70 t ha–1, cosa que representa el 40% de la biomassa aèria i el 30% de la total de l’ecosistema.
L’economia hídrica
Les baixes temperatures hivernals imposen restriccions al funcionament dels ecosistemes mediterranis, però el factor amb més pes i que ha donat lloc a adaptacions més marcades és el derivat de la combinació de secada i altes temperatures durant l’època estival. Estalviar l’aigua, fer-ne un ús que sigui l’òptim, és el problema que han hagut de resoldre les espècies d’aquest bioma. Els animals, en general, que obtenen l’aigua dels aliments o la ingereixen directament, no pateixen problemes seriosos de subministrament ja que tenen prou mobilitat per aconseguir-ne. De fet, la majoria dels animals són més independents de les condicions climàtiques que els vegetals, a causa de la seva estructura, del seu funcionalisme molt regulat i de la seva mobilitat. Excepció feta dels menys adaptats a la vida terrestre, com determinats crustacis, mol·luscs o amfibis, la manca d’aigua en el bioma mediterrani no arriba als extrems que es donen en els deserts i que han afavorit l’aparició d’adaptacions morfològiques, etològiques i fisiològiques en la majoria dels animals que hi habiten. La fauna edàfica és, segurament, la que acusa d’una manera més marcada la manca estacional d’aigua, i en els seus components trobem diferents estratègies per a fer front a aquest problema, que es poden agrupar en tres grans tipus. Una primera estratègia consisteix a mantenir-se permanentment en fondària, en capes del sòl que no arriben a assecar-se mai i en les quals les arrels profundes dels vegetals, abundants en el bioma mediterrani, mantenen el sòl esponjat i asseguren el subministrament de matèria orgànica. Una segona estratègia és la dels animals que viuen sempre prop de la superfície, tot fent front a la secada per dues vies diferents. La letargia o la producció d’estructures reproductives de resistència permet evitar la manca d’aigua. Els que romanen actius, en canvi, han desenvolupat estructures protectores, com ara fortes cutícules complementades amb mecanismes fisiològics que permeten estalviar aigua i consumir els materials que constitueixen les dures fulles esclerofil·les. Finalment, una tercera estratègia consisteix a realitzar migracions, ja sigui en sentit vertical cap a capes inferiors del sòl, o bé obliquament en direcció a les fondalades, de sòl més profund i humit.
Els vegetals, però, són molt més sensibles al problema de la manca d’aigua i, dominants com són en els ecosistemes terrestres, aquest factor esdevé essencial per a entendre’n el funcionalisme. En el bioma mediterrani es troben, bàsicament, vegetals malacofil·les caducifolis hivernals, esclerofil·les perennifolis i malacofil·les caducifolis estivals. Els primers, restringits a les parts més humides i fresques del bioma, tenen una estratègia derivada més d’una adaptació al fred hivernal que no de la manca d’aigua estival, molt amortida i poc determinant en aquests ambients. Els altres dos tipus corresponen a estratègies relacionades amb l’economia hídrica i, secundàriament, amb la temperatura. En condicions de secada estival i fred hivernal moderat, que podríem considerar com a mitjanes o típiques del bioma, els ecosistemes mediterranis estan dominats per vegetals esclerofil·les perennifolis, i quan la secada és més accentuada i el fred hivernal disminueix, en les zones més càlides del bioma, els vegetals dominants corresponen al tipus dels caducifolis estivals. En el primer cas, la secada pot afrontar-se amb un seguit d’adaptacions morfològiques i fisiològiques, mentre que en el segon cas, el rigor de la secada combinat amb la suavitat de les temperatures hivernals afavoreix la concentració de l’activitat en aquesta última època, amb la qual cosa l’estiu esdevé un període de repòs.
En els ecosistemes dominats per vegetals esclerofil·les perennifolis, l’estalvi d’aigua durant l’època eixuta s’aconsegueix per un efectiu mecanisme de tancament estomàtic que permet regular el flux d’aigua. Llavors l’activitat queda limitada a les hores més fresques del dia, i s’interromp durant les més caloroses per tal d’evitar pèrdues d’aigua excessives i no compensables per les reserves edàfiques. D’aquesta manera s’aconsegueix que la transpiració al llarg de l’època estival sigui fins al 20% inferior a la que es dóna al llarg de l’època hivernal, sense necessitat de reduir l’àrea foliar sinó gràcies als mecanismes fisiològics del control de la transpiració.
En el cas dels caducifolis estivals, la transpiració esdevé gairebé nul·la a ple estiu per una reducció dràstica o total de la superfície foliar; l’activitat es concentra durant la resta de l’any. Hi ha estratègies intermèdies que consisteixen bé en una reducció parcial de l’àrea foliar per pèrdua d’una part de les fulles o bé en una reducció per substitució de les fulles hivernals per d’altres amb característiques més xeromòrfiques. Aquest dimorfisme estacional permet, doncs, combinar les dues formes de regulació: la transpiració es redueix per la disminució de la superfície foliar i per un augment del control fisiològic d’aquesta. La reducció de la massa transpirant durant el període eixut pot arribar, en aquests casos, a deixar-la en valors de prop del 15% de la massa transpiradora hivernal.
Els ecosistemes mediterranis tenen, doncs, mecanismes que els permeten d’aprofitar exhaustivament l’aigua de què disposen: cal optimar l’ús d’aquest factor limitador. Un alzinar, per exemple, amb una precipitació anual d’uns 500 mm, evapotranspira el 80% de l’aigua de precipitació, és a dir, uns 464 mm, mentre que sota una precipitació de 870 mm n’evapotranspira el 54%, que correspon a 469 mm, tot posant de manifest l’aportació mínima d’aigua que necessita per a funcionar. Amb precipitacions inferiors a 400 mm, o bé amb períodes de secada molt llargs, el paisatge és dominat per formacions arbustives, amb unes necessitats hídriques més modestes. En una zona de “chaparral” californià, per exemple, el consum d’aigua per evapotranspiració d’una mateixa comunitat arbustiva oscil·la entre els 250 i 350 mm anuals, que corresponen respectivament a valors de pluviositat de 300 i 1 300 mm anuals, cosa que fa palesa tant la irregularitat del clima mediterrani com la importància dels valors mitjans i baixos de precipitació a l’hora de determinar les característiques del paisatge. Sobrats de llum i mancats d’aigua, els boscos i les bosquines mediterrànies en deixen molt poca per als magres rius del bioma.
La producció
El factor limitador lligat a la disponibilitat d’aigua imposa limitacions als ecosistemes mediterranis, que es tradueixen en una productivitat global baixa i unes taxes d’increment o renovació de la biomassa també baixes. Els boscos i les bosquines mediterranis funcionen lentament, com motors de baix consum i baix rendiment.
La producció primària neta aèria d’una pluviïsilva tropical és del voltant de les 20 t/ha any en pes sec, i la d’un bosc temperat de coníferes o caducifolis ateny les 12 t/ha any. En el cas dels ecosistemes mediterranis, es troben valors mitjans de producció de prop de les 5 t/ha any per als boscos i de 3 t/ha any per a les formacions arbustives, tot i que en determinades circumstàncies especialment favorables es poden atènyer valors propers a les 10 t/ha any i en condicions de secada extrema pot no superar-se el valor d’1 t/ha any. Cal tenir en compte, doncs, que molts boscos mediterranis tenen produccions inferiors a les de la taigà, que ateny per terme mitjà les 8 t/ha i any sota unes condicions climàtiques especialment dures a causa de les baixes temperatures, la qual cosa posa de manifest el paper determinant de la disponibilitat hídrica en el cas dels ecosistemes terrestres.
La descomposició
La descomposició de la matèria orgànica morta representa una part molt important en els fluxos d’energia i els cicles de nutrients dels ecosistemes, ja que determina el retorn dels elements a la forma inorgànica, és a dir, a l’estat en què estan disponibles novament per a la vegetació. La descomposició és un procés molt complex en el qual intervenen en un grau molt important els fongs i els bacteris del sòl, com també tota la fauna edàfica que s’alimenta d’aquestes restes. En els ecosistemes mediterranis, el procés de descomposició és relativament lent ja que s’enfronta a dos problemes bàsics: la manca d’aigua, que limita l’activitat dels descomponedors, i la resistència estructural de les restes vegetals provinents dels vegetals esclerofil·les.
La secada estival dóna lloc a una dessecació intensa de les capes superiors del sòl, que limita fortament o atura del tot l’activitat descomponedora dels fongs i els bacteris, com la de molts components de la fauna edàfica, els quals tenen, també, un paper molt important en el procés de descomposició, ja que en fragmentar la fullaraca faciliten enormement l’atac fúngic i bacterià. Aquesta fauna presenta, en les parts més àrides del bioma mediterrani, un màxim de densitat hivernal i un mínim estival relacionat amb una disminució de les poblacions i amb una migració cap a capes més profundes del sòl on es conserva una certa humitat.
En les zones més humides i fresques del bioma es donen dues màximes anuals, a la primavera i tardor, ja que el fred hivernal actua també com a limitador. En el cas dels bacteris, es dóna una successió o substitució progressiva d’unes espècies per unes altres a mesura que el procés de dessecació del sòl avança. De tota manera, l’activitat microbiana es veu fortament reduïda al pic de l’estiu, cosa que atura de manera gairebé total el procés de descomposició. L’activació de les poblacions bacterianes pot ser hídrica, tot donant lloc a màxims de densitat i activitat un mes després de les primeres pluges que segueixen al període sec, o bé tèrmica, coincidint amb la pujada primaveral de les temperatures. La descomposició, doncs, queda limitada a les èpoques humides de l’any, que són, també, les més fresques, tot donant lloc a un procés de mineralització discontinu amb dos màxims anuals, separats per una època massa seca i una altra de relativament freda. La coincidència de calor i humitat, que tant afavoreix els processos de descomposició, ja que la majoria dels fongs i bacteris tenen un màxims d’activitat amb humitats elevades i temperatures entre 25 i 35°C, no es dóna sota condicions climàtiques mediterrànies.
El caràcter esclerofil·le d’una bona part dels vegetals mediterranis representa un segon inconvenient que alenteix el procés de descomposició. Els continguts de lignina i de nitrogen de les restes vegetals condicionen fortament la seva velocitat de descomposició. L’esclerofil·lisme va acompanyat d’una característica especial de les fulles: una proporció particularment alta de materials estructurals, com ara la lignina, combinada amb una baixa proporció de nutrients essencials com el nitrogen, cosa que fa que aquestes fulles s’assemblin més a les de les coníferes que no pas a les dels planifolis. Les fulles d’un vegetal esclerofil·le típicament mediterrani com l’alzina (Quercus ilex) contenen poc més de l’1% de nitrogen, mentre que les d’un planifoli de clima temperat humit com el faig (Fagus sylvatica) assoleixen el 2%. Això fa que en el mateix període (18 mesos) en què les fulles de la virosta d’una roureda de clima temperat perden un 40% del seu pes, les d’una comunitat dominada per esclerofil·les com el “chaparral” perden només un 20%. Com que les fulles, que es renoven ràpidament comparades amb les parts llenyoses, representen més del 70% de la virosta produïda anualment per la vegetació, les seves característiques determinen la velocitat de descomposició de la virosta en conjunt. El caràcter esclerofil·le dóna lloc, doncs, a un alentiment del procés de retorn dels nutrients a la fracció inorgànica, endarreriment que pot provocar un empobriment relatiu del sòl, compensat per l’aparició recurrent del foc que mineralitza la matèria orgànica acumulada.
La circulació dels nutrients
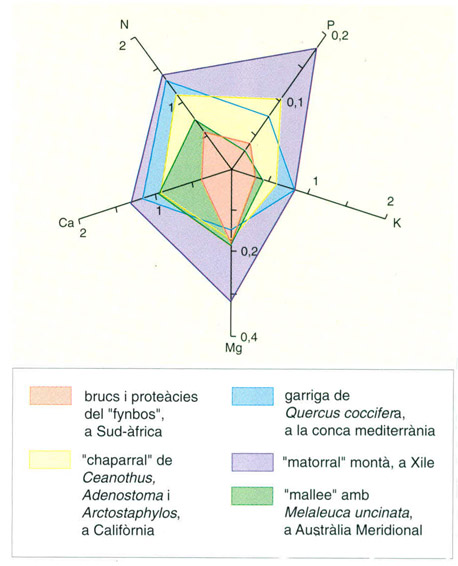
Contingut foliar de nutrients (N nitrogen, P fòsfor, K potassi, Mg magnesi, Ca calci) en els quatre tipus de bosquina més representatius del bioma mediterrani, expressats en percentatge sobre la matèria seca total de la fulla. La pobresa relativa del “fynbos” acidòfil capenc respecte del “matorral” calcícola xilè és ben palesa.
Editrònica, a partir de Kruger 1983
La circulació global de nutrients en un ecosistema es produeix en part dins mateix de l’ecosistema i en part en un àmbit extern. El cicle intern dels nutrients consisteix bàsicament en l’absorció d’aquests nutrients pels vegetals, la seva retranslocació o mobilització des dels òrgans vells cap a les parts noves de la planta, la seva retenció en les noves estructures sintetitzades i el seu retorn a la virosta i subsegüent descomposició. El cicle extern engloba les entrades i les sortides de nutrients del sistema, la major part de les quals estan associades a la precipitació i a les aigües de drenatge, tot i que poden ser importants les derivades de la deposició seca o l’absorció directa de gasos com el nitrogen. La velocitat de circulació dels nutrients està molt associada a la de la matèria orgànica, ja que una elevada productivitat o una ràpida descomposició donen lloc a una mobilització intensa dels nutrients i a una acceleració dels seus cicles.
En el cas dels sistemes mediterranis, ja hem esmentat la seva baixa productivitat associada a la baixa disponibilitat d’aigua, cosa que determina un alentiment general dels cicles. D’altra banda, la baixa velocitat de descomposició reforça aquesta lentitud, de manera que la velocitat general de renovació dels nutrients en aquests ecosistemes és baixa.
En determinades condicions, la disponibilitat dels nutrients, o d’algun element essencial, com el fòsfor o el nitrogen, pot actuar com a limitadora. Això passa en tots els biomes en condicions de pobresa edàfica, però és relativament comú a les zones mediterrànies i sobretot a les regions australianes i capenca mediterrànies. En aquestes condicions, els ecosistemes tendeixen a posar en marxa mecanismes que assegurin el subministrament i el control de nutrients reforçant algunes parts del cicle com la retranslocació. D’aquesta manera, en retirar una bona part dels nutrients essencials de les estructures que han de ser eliminades, com per exemple les fulles velles, s’evita que aquests nutrients s’acumulin en la virosta o es perdin per lixiviació. Els vegetals mediterranis adaptats a aquestes condicions, normalment esclerofil·les de fulla petita com les ericàcies o les epacridàcies, tenen fulles amb una baixa proporció de nutrients i arriben a reciclar per retranslocació fins el 70% dels nutrients essencials durant el procés de senescència foliar, rebaixant encara més la velocitat de circulació dels nutrients. Alguns, com Banksia ornata, que creixen sobre sòls pobres, arriben a retirar fins el 90% del fòsfor i el 30% del nitrogen de les fulles senescents. Els teixits i les estructures de vida llarga, com ara les tiges i les arrels, poden actuar com a magatzems d’aigua i nutrients, cosa que augmenta el control dels cicles de nutrients pels vegetals. Les voluminoses soques constituïdes per un teixit especialitzat anomenat lignotúber, presents en la majoria dels arbres i arbustos mediterranis amb capacitat per a rebrotar, s’han interpretat com una adaptació al foc, tot i que també tenen un paper molt important com a magatzems de nutrients i aigua, i estan molt desenvolupades en espècies pròpies de sòls oligotròfics, com les ericàcies o els “jarrah” australians, capaços de formar boscos molt desenvolupats sobre els que són considerats com uns dels sòls forestals més pobres del món. En aquests casos de marcada oligotròfia i descomposició lenta, el foc actua com a mobilitzador de nutrients en mineralitzar la capa de virosta acumulada, i sembla un factor essencial per a dinamitzar, periòdicament, el funcionament d’aquests sistemes.
Les condicions d’oligotròfia edàfica no són generals en el bioma mediterrani, però cal no oblidar que la majoria de la flora del bioma procedeix d’espècies adaptades a ambients pobres de nutrients i que, per tant, poden exhibir adaptacions relacionades amb l’economia dels nutrients aparentment sobreres en els ambients actuals. D’altra banda, les condicions de secada determinen una pobresa relativa de nutrients, ja que aquests no poden ser absorbits del sòl a causa de la manca d’aigua per a vehicular-los. No és estrany, doncs, que la majoria de vegetals mediterranis presentin característiques relacionades amb l’estalvi de nutrients —per exemple les baixes proporcions de nutrients en les fulles— i el control del cicle de nutrients —com per exemple una elevada taxa de retranslocació—, una coincidència de l’època de caiguda de les fulles velles amb la de producció de les noves i una elevada concentració de nutrients en estructures com la fusta o l’escorça, des de les quals poden ser mobilitzats per cobrir necessitats puntuals.
Biogeografia de l’esclerofil·lisme
Les formacions esclerofil·les de la conca mediterrània
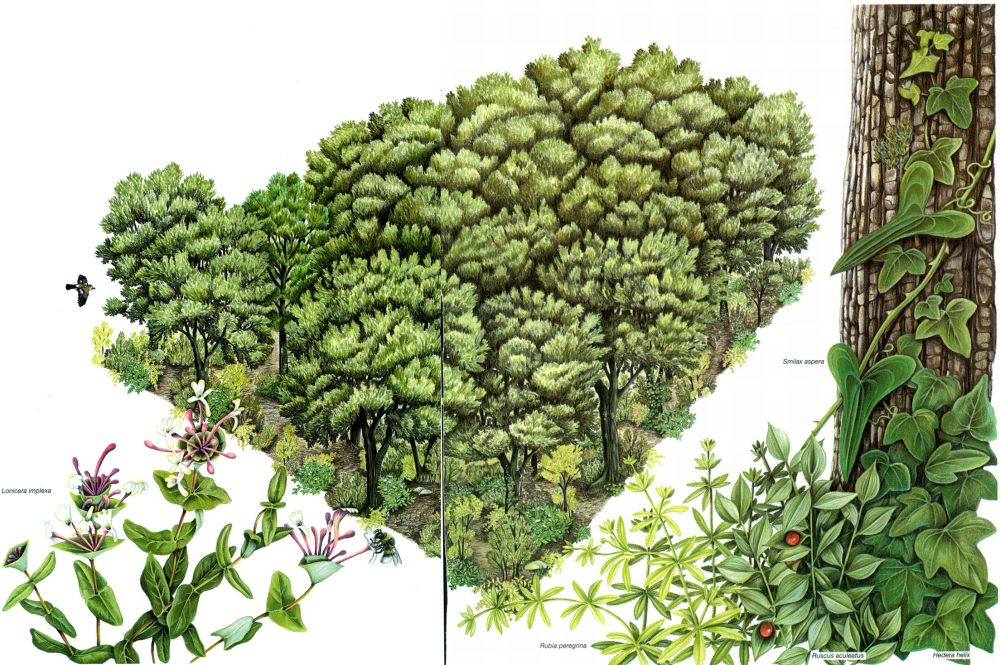
Bloc esquemàtic de l’estructura d’un alzinar típic que permet d’apreciar el port modest i la simplicitat de l’estrat arbori —integrat per alzines (Quercus ilex) i alguns pins (Pinus halepensis)—, la pràctica absència d’estrat herbaci, i la riquesa de l’estrat arbustiu i lianoide; a primer terme, justament, apareixen algunes de les plantes enfiladisses o agafatoses més representatives, com el lligabosc (Lonicera implexa), la roja (Rubia peregrina), l’arítjol (Smilax aspera) o l’heura (Hedera helix), com també el galzeran (Ruscus aculeatus).
Anna M. Ferrer
A la conca mediterrània hi ha diverses menes de formacions esclerofil·les, dominades per arbustos o per arbres segons quines siguin les característiques climàtiques i edàfiques de cada lloc. Quan la precipitació ultrapassa els 400 mm anuals, el paisatge sol estar dominat per boscos, mentre que per a valors inferiors de precipitació trobem dominància de les comunitats arbustives. Les condicions edàfiques matisen aquest esquema tan senzill, ja que sobre substrat calcari o sobre sòls magres i de fort pendent, una pluviositat superior als 400 mm anuals pot ser insuficient per a mantenir una comunitat arbòria, i llavors s’hi instal·la una formació arbustiva. La frontera entre les formacions arbòries i arbustives no és pas clara, ja que es poden trobar estadis intermedis com ara boscos poc desenvolupats o comunitats arbustives amb un estrat arbori poc dens.
Els alzinars i les pinedes
Els boscos esclerofil·les més típics i desenvolupats són els formats per diverses espècies perennifòlies del gènere Quercus. En les zones més humides de l’àrea mediterrània dominen els alzinars, que són boscos esponerosos amb una estructura exhuberant i amb un estrat arbori constituït per l’alzina típica (Quercus ilex). Aquests boscos poden atènyer dimensions considerables, ja que els arbres assoleixen els 25 m d’alçada. Les formacions d’alzinar més denses i riques es troben en les zones costaneres, ja que en les àrees muntanyoses es desenvolupen boscos florísticament més pobres, mancats de les espècies arbustives termòfiles.
En les àrees de clima més continental, com a l’interior de la Península Ibèrica i en les àrees de transició cap a subclimes més àrids, es fan boscos dominats per una espècie molt propera a l’alzina típica, l’anomenada alzina carrasca o simplement carrasca (Quercus rotundifolia). Els carrascars, sotmesos a condicions climàtiques més dures, tenen un aspecte menys esponerós que els alzinars litorals, ja que l’estrat arbori no arriba a desenvolupar-s’hi tant i els estrats arbustiu i herbaci tenen molt poca entitat.
A la part occidental de la conca mediterrània prospera, encara, un tercer tipus de bosc dominat per la surera (Quercus suber). Les suredes ocupen les àrees silícies i humides, i arriben a altituds menors que els alzinars muntanyencs. L’arbre dominant és diferent, però el conjunt d’espècies arbustives i herbàcies que l’acompanyen no difereix gaire de les que es poden trobar en els alzinars litorals típics.
A la part oriental de la conca apareixen formacions arbòries dominades per Q. calliprinos, un arbre molt semblant al garric (Q. coccifera) però de talla més gran, que es presenta acompanyat d’altres espècies perennifòlies com el llentiscle (Pistacia palaestina) i la savina (Juniperus phoenicea). Aquestes formacions arbòries, sovint laxes, són molt abundants a Síria, el Líban, Israel i Jordània, on ocupen un espai ecològic semblant al dels alzinars occidentals.
En les zones de clima submediterrani, és a dir, amb un eixut estival menys acusat i amb hivern lleugerament més fred, els alzinars i els carrascars deixen pas a boscos caducifolis o semicaducifolis dominats també per espècies del gènere Quercus. Es tracta, generalment, d’arbres marcescents, és a dir, que mantenen les fulles seques agafades a les branques durant una bona part de l’hivern, en un comportament a mig camí dels caducifolis i els perennifolis. Hi ha diverses espècies del mateix gènere que ocupen àrees geogràfiques diferents: el roure martinenc (Quercus pubescens), el roure de fulla petita (Q. faginea), el roure cerrioide (Q. cerrioides), el roure reboll (Q. pyrenaica) i el roure africà (Q. canariensis) a la mediterrània occidental, i Q. macrolepis,Q. ithaburiensis,Q. infectoria, Q. boissieri, Q. frainetto, Q. trojana, Q. aegilops i el roure martinenc, a la part més oriental. En moltes situacions poden formar-se masses mixtes en les quals coexisteixen espècies perennifòlies i marcescents.
També es poden trobar masses arbòries dominades per coníferes, per exemple de pi blanc (Pinus halepensis) i pi de Calàbria (P. brutia), aquest últim relegat a la part oriental. El paper d’aquestes espècies és molt variat, ja que poden formar masses forestals pures o bé mixtes, barrejades amb diferents espècies de Quercus, o constituir un estrat arbori lax en comunitats arbustives. Sovint el seu paper es considera secundari, en el sentit que són masses forestals que substitueixen boscos de perennifolis o semicaducifolis alterats. Hi ha altres espècies de coníferes amb una importància paisatgística menor, com ara el pi pinyer (P. pinea), el pinastre (P. pinaster), el xiprer (Cupressus sempervirens), la savina articulada o de Cartagena (Tetraclinis articulata) i la savina pròpiament dita (Juniperus phoenicea), que formen masses arbrades relativament poc extenses.
En els ambients de muntanya, encara sota condicions climàtiques clarament mediterrànies, i compartint el territori amb els boscos de planifolis de fulla perenne o decídua, es poden trobar boscos constituïts per les diferents subspècies d’un pi típicament mediterrani, la pinassa (P. nigra), que en algunes zones deixa pas a una altra espècie clarament extramediterrània, el pi roig (P. sylvestris). Les abundants subspècies de la pinassa són producte de l’aïllament de les poblacions en els ambients orogràfics mediterranis, fenomen que ha donat lloc, en d’altres casos, a espècies clarament diferenciades. Aquest és el cas dels cedres (Cedrus atlantica, C. libani i C. brevifolia) i dels avets (Abies maroccana, A. pinsapo, A. nebrodensis, A. cephalonica, A. borisi-regis, A. equi-trojani, A. bormulleriana i A. cilicica), totes elles amb àrees de distribució molt reduïdes però amb un gran interès, que es deriva del seu caràcter endèmic i relicte. Algunes d’aquestes espècies es troben en perill d’extinció a causa de la migradesa dels seus efectius; el cas més accentuat és el de l’avet de Nebrodi (A. nebrodensis), del qual queden entre 10 i 20 exemplars en estat silvestre.
Les màquies, brolles i timonedes
Les formacions arbustives presenten característiques fisiognòmiques molt variades, segons els condicionants climatològics i edàfics. Les més desenvolupades reben el nom de màquies: és també la “macchia” italiana, el “maquis” francès, el “xerovuni” grec, el “choresh” hebreu. Es tracta de formacions molt denses, gairebé impenetrables, que atenyen alçades superiors a 2 m, dominades per espècies arbustives que poden assolir talla gairebé arbòria, com ara l’ullastre (Olea europaea varietat sylvestris), el garrofer (Ceratonia siliqua), els llentiscles (Pistacia lentiscus, P. palaestina), la murta (Myrtus communis), els arboços (Arbutus unedo, A. andrachne), entre d’altres. En el cas de formacions arbustives més baixes (al voltant d’1 m) i dominades pel garric (Quercus coccifera), la comunitat rep el nom de garriga. En molts casos, aquestes formacions arbustives poden presentar un estrat arbori lax de coníferes, sobretot de pi blanc (Pinus halepensis) i pi de Calàbria (P. brutia), que els dóna un aspecte vagament forestal. La seva importància paisatgística és molt gran, ja que ocupen moltes zones forestals a causa de la degradació dels boscos.
Trobem, encara, comunitats arbustives d’estructura més simple, que no sobrepassen els 2 m d’alçada i amb una densitat clarament menor, cosa que permet que la radiació solar arribi fàcilment fins a terra. Es tracta de les anomenades brolles, dominades per diferents espècies d’estepes (Cistus), brucs (Erica) i argelagues i ginestes (Genista) i per d’altres de la família de les labiades com ara el romaní (Rosmarinus officinalis). Les espècies dominants sovint no són pròpiament esclerofil·les, car presenten característiques malacofil·les o bé tenen fulles molt petites o absents.
Les formacions arbustives de menys volada, típiques de llocs molt secs o degradats, són les anomenades timonedes (els “tomillares” ibèrics, els “phrigana” grecs, els “batha” palestins), riques de labiades com farigoles, espígols i ventolloses (Thymus, Lavandula, Phlomis), entre d’altres, i de cistàcies com les del gènere Helianthemum. La degradació d’aquestes comunitats porta a la implantació de prats secs que constitueixen la mínima expressió de la vegetació mediterrània.
Finalment, les àrees culminants de l’alta muntanya mediterrània, amb un clima clarament més fred però plenament afectades per la secada estival, presenten unes formacions vegetals arbustives molt característiques, l’anomenada vegetació oromediterrània, dominada per coixinets espinosos, genèricament anomenats eriçons (Erinacea anthyllis, Vella spinosa), de morfologia semblant als que ocupen algunes àrees costaneres molt batudes pel vent, adaptats a suportar l’elevada insolació, el fred i la secada, accentuada aquesta última pel poder dessecant del vent.
Les formacions esclerofil·les californianes
A la mediterrània californiana es reprodueix un esquema semblant de distribució de les formacions, amb boscos esclerofil·les dominats per arbres del gènere Quercus a les parts més humides i formacions arbustives en les zones de clima més sec.
El “savanna oak” i altres boscos
Les formacions boscoses presenten una gran varietat d’espècies, tal com passa en les formacions arbustives a causa de les baixes o nul·les taxes d’extinció associades a les glaciacions. S’hi poden trobar espècies caducifòlies o marcescents, com els roures coneguts popularment com a “valley oak” (Quercus lobata), “blue oak” (Q. douglasii), “Engelman oak” (Q. engelmanii), “black oak” (Q. kelloggii) i “Oregon white oak” (Q. garrayana), alternats amb coníferes com ara els pins Pinus sabiniana i P. coulteri. En llocs més secs, dominen les espècies arbòries de Quercus perennifolis, com el “canyon live oak” (Q. chrysolepis), el “coast live oak” (Q. agrifolia), el “scrub interior live oak” (Q. wislizenii) i Q. tomentella. Aquestes formacions boscoses presenten una morfologia molt variada, ja que les diferents espècies es barregen formant masses mixtes. En molts casos, les masses arbòries no són denses sinó laxes, i donen lloc a un paisatge de pastures arbrades conegut com “savanna oak”, d’origen clarament antropogènic, causat per la pastura i els focs recurrents de baixa intensitat que no afecten els grans arbres. Les formacions dominades per espècies perennifòlies poden presentar estructura arbustiva sota condicions ambientals limitadores, tot configurant una frontera imprecisa entre boscos i bosquines.
Entre els boscos de la zona mediterrània californiana cal esmentar un cas excepcional: el dels boscos de sequoia (Sequoia sempervirens) que es troben en alguns punts de la costa, en el cor de la zona de clima mediterrani. Aquests boscos són formats per arbres que assoleixen més de 50 m d’alçada, i constitueixen un cas excepcional en un bioma on els boscos tenen un aspecte molt més modest. Tenen una certa semblança amb els boscos de la sequoia gegant (Sequoiadendron giganteum) de les muntanyes del nord de l’àrea mediterrània nord-americana, però mentre que en aquest cas el gran desenvolupament és fruit d’unes condicions climàtiques clarament diferents, en el cas de les sequoies costaneres el factor determinant és l’abundància de boires que es formen per la incidència dels vents oceànics, cosa que dóna lloc a una reducció de les pèrdues hídriques i a una aportació extra d’aigua per condensació sobre la vegetació.
Les coníferes, en conjunt, també tenen un paper molt variable, tal com passa a la conca mediterrània, ja que poden ser boscos de substitució o constituir estrats arboris sobre comunitats arbustives. Són molt característics Pinus attenuata, P. muricata i el pi insigne o de Monterey (P. radiata), i també diverses espècies del gènere Cupressus, com ara el xiprer de Monterey (C. macrocarpa). Moltes d’aquestes espècies són clarament pirofítiques, cosa que les afavoreix en ambients on els incendis són freqüents.
El “chaparral” californià
Les més famoses de les formacions arbustives mediterrànies californianes tenen caràcter arbustiu i són genèricament conegudes sota la denominació de “chaparral”. El “chaparral” prospera a les zones amb precipitacions inferiors als 400 o 500 mm anuals, i malgrat això la seva riquesa d’espècies és molt gran. Hi predominen espècies dels gèneres Adenostoma, com ara el “chamizo” (A. fasciculatum), Arctostaphylos, com la “manzanita” (A. glauca), Ceanothus, com la “lila silvestre” (C. gregii), Heteromeles i Rhus (R. diversiloba), juntament amb algunes del gènere Quercus, com ara l’“encinillo” o “californian scrub oak” (Q. dumosa).
Es diferencien uns quants tipus diferents de “chaparral” segons les condicions ambientals i les espècies dominants. Es troben formacions boscoses baixes o “woodland chaparral” dominades per l’“encinillo” (Quercus dumosa), comunitats arbustives denses amb dominància de Ceanothus, conegudes per això amb el nom de “chaparral de Ceanothus”, les de més altitud, amb dominància d’espècies del gènere Arctostaphylos, o “chaparral de manzanita”, o les de les zones més seques i de sòls pobres dominades per Adenostoma que formen el “chaparral de chamizo”.
En les zones més seques i pedregoses, exposades a la influència marina o desèrtica, es troben formacions arbustives baixes dominades per petits arbustos caducifolis dels generes Artemisia i Salvia, anomenades “coastal sage shrub”, que tenen un aspecte molt semblant al de les brolles i les timonedes de la conca mediterrània.
Finalment, els prats secs representen els estadis de degradació de comunitats de vegetació arbòria o arbustiva principalment en els casos que han estat afectades pel foc.
Les formacions esclerofil·les xilenes
La zona mediterrània xilena ofereix, també, una gran varietat de paisatges esclerofil·les. S’hi fan diverses formacions boscoses i arbustives que van des dels “encinales” fins al “matorral costero” passant pels “espinales”.
Els “encinales”
Les formacions boscoses, que poden arribar fins a uns 10-15 m d’alçada i ocupen les parts més humides, com obacs i fons de valls, són dominades per espècies esclerofil·les com el “litre” (Lithraea caustica), la quil·laia (Quillaja saponaria), el “boldo”(Peumus boldus), i el “peumo” (Cryptocarya alba), entre d’altres. La palmera endèmica, Jubaea chiloensis creix en una àrea molt limitada al nord-est de Valparaíso. De fet, els “encinales” ocupen poca extensió en el paisatge actual.
Els “matorrales” i l’“espinal”
En llocs més eixuts, o per degradació d’aquestes formacions arbòries, apareix una formació arbustiva alta, l’anomenat “matorral”. Sovint la composició específica del “matorral” és molt semblant a l’esmentada abans, i la diferència més clara és deguda al grau de desenvolupament. A les espècies acabades d’esmentar cal afegir el “guayo bollen” (Kageneckia oblonga) i Colliguaya odorifera, arbustos esclerofil·les. Enmig d’aquestes espècies en trobem d’altres, algunes malacofil·les i caducifòlies estivals com ara Baccharis linearis, B. rosmarinifolius, Colletia spinosissima, Proustia pungens i el “trevo” (Trevoa trinervis). En els llocs més eixuts també apareixen cactàcies columnars com ara el “quisco” (Trichocereus chiloensis) i bromeliàcies com el “chagual” o “puya cardón” (Puya berteroniana) i P. coerulea. En les zones costaneres, les espècies esclerofil·les perden importància i passen a ser dominants les crasses i malacofil·les, les darreres de les quals es comporten com a caducifòlies estivals; és el cas de la fúcsia Fuchsia lycioides, el “huanil” (Proustia pungens) i Adesmia arborea, a més de les ja esmentades anteriorment, i donen lloc a formacions conegudes amb el nom de “matorral costero”.
A les valls centrals es fa una comunitat característica amb aspecte sabanoide, dominada per l’“espino” (Acacia caven), i anomenada per això “espinal”. Es tracta d’arbrets que poden arribar als 3 o 4 m d’alçada i desenvolupar capçades de 5 o 6 m de diàmetre. Tenen una gran plasticitat, ja que es comporten com a caducifolis hivernals, perennifolis o caducifolis estivals segons les característiques climàtiques. El recobriment dels arbustos pot oscil·lar entre el 5 i el 15%.
En la transició entre els “espinales” i el “matorral”, l’estrat arbustiu es fa més dens i s’enriqueix amb espècies típiques d’aquest darrer. L’estrat herbaci d’aquestes comunitats és dominat per espècies introduïdes d’origen eurasiàtic que suporten molt millor la pastura i que han desplaçat les espècies indígenes. Aquestes formacions sabanoides són considerades com a estadis de degradació del “matorral”, tot i que en determinats indrets massa secs per suportar una formació arbustiva densa poden representar-hi la vegetació climàcica.
En els vessants de la serralada andina, per damunt els 1 800 m d’altitud, el “matorral” típic és substituït per una formació arbustiva de caràcter també xèric dominada per espècies com Colliguaya integerrima, Kageneckia angustifolia i Guindilia trinervis. A més altitud apareixen formacions arbustives espinoses en les quals abunden les espècies arbustives amb forma de coixinet. Les coníferes, que tenen molt poca importància en la regió mediterrània xilena, apareixen en aquesta àrea on trobem alguns boscos de “ciprés de la cordillera” (Austrocedrus chilensis).
Les formacions esclerofil·les capenques
A la zona mediterrània sud-africana manquen les formacions arbòries, ja que les poques espècies de port arbori, com l’anomenat “silvertree” o “silverboom” (Leucadendron argenteum) tenen àrees de distribució reduïdíssimes. El paisatge és dominat per les formacions arbustives. Només queden unes petites restes dels antics boscos temperats humits, que van patir una greu regressió d’ençà de la colonització europea.
Hom distingeix quatre grans tipus de vegetació arbustiva, anomenats “stranveld”, “fynbos”, “renostervosveld” i “karoo”. Aquesta classificació es fonamenta en criteris derivats dels usos agrícoles i ramaders que poden dur-se a terme en cadascun dels tipus considerats, ja que inclouen condicionaments edàfics i climàtics, i reflecteix d’una manera clara les característiques de la vegetació natural.
A grans trets, les formacions de “stranveld” se situen en les zones àrides costaneres; el “coastal fynbos” i el “mountain fynbos” ho fan en zones intermèdies quant a aridesa i amb sòls relativament pobres; els “coastal renostervosveld” i el “mountain renostervosveld” sobre sòls molt pobres i en la transició cap a les zones més àrides; finalment, les formacions de “karoo” es troben en les zones més àrides, de transició cap als deserts.
El “fynbos” i el “strandveld”
El “fynbos”, que pot atènyer fins a 2 o 3 m d’alçada, és dominat per ericàcies i restionàcies, tot i que també hi trobem com a acompanyants espècies d’arbustos esclerofil·les (dels gèneres Protea, Leucadendron, Olea, Euclea, Rhus), malacofil·les caducifolis estivals (Zygophyllum) o suculents (Euphorbia). L’abundància relativa de les espècies àfil·les (del tipus de les restionàcies), les esclerofil·les de fulla petita (del tipus de les ericàcies) i les esclerofil·les de fulla ampla (del tipus de les proteàcies), depèn de la riquesa del sòl, cosa que dóna lloc a un mosaic de vegetació en el qual trobem un espectre que va des de les formacions del tipus landa o bruguerar fins a les que recorden més una màquia.
En el cas del “strandveld”, que ocupa preferentment les dunes costaneres i els sòls granítics més rics, les espècies dominants són vegetals esclerofil·les que donen a la formació un aspecte molt semblant al de les màquies de la conca mediterrània, i que corresponen a les mateixes espècies arbustives acompanyants mencionades en el tipus anterior.
El “renosterveld” i el “karoo”
En les zones costaneres degradades i en les interiors de transició cap al semidesert del Karoo, trobem el “renosterveld”, formacions arbustives laxes amb un aspecte molt semblant al “coastal sage” californià, dominades per la composta “renosterbos” (Elytropappus rhinocerothis), amb acompanyants que sovint són arbustos caducifolis estivals, tot i que en les zones costaneres hi podem trobar alguns esclerofil·les.
Les formacions de “karoo”, instal·lades en els llocs més àrids, són dominades per plantes suculentes, cosa que els confereix un aspecte molt peculiar, tal com correspon al seu caràcter de transició cap als deserts interiors.
Les formacions esclerofil·les australianes
Les regions mediterrànies australianes presenten unes característiques molt semblants a les de la regió capenca, ja que a les condicions climàtiques mediterrànies s’afegeix una pobresa edàfica de nutrients molt marcada, amb sòls de caràcter també silici.
La vegetació és dominada per espècies d’eucaliptus (Eucalyptus), els noms vulgars dels quals també designen les formacions on predominen, tot i que també s’hi poden trobar abundants proteàcies en l’estrat arbustiu i epacridàcies que fan el mateix paper que les ericàcies europees, és a dir, ocupen els sòls més pobres en nutrients. Hi ha algunes altres peculiaritats florístiques, com les monocotiledònies arborescents dels gèneres Kingia i Xanthorrhoea i la cicadàcia Macrozamia.
El “jarrah”, el “wandoo” i el “mallee” occidentals
A les zones més plujoses de l’àrea mediterrània australiana es fan els boscos de “jarrah” (Eucalyptus marginata) que, com tots els altres arbres del seu gènere, no és pròpiament esclerofil·le, sinó més aviat de fulla coriàcia. Aquests boscos sempre verds assoleixen fàcilment els 20 o 25 m d’alçada i ocupen les àrees de precipitació superior als 600 mm anuals.
Els boscos de “jarrah” australians són substituïts per boscos extramediterranis de “karri” (E. diversicolor) allà on les pluges superen els 1 200 mm anuals, tal com passa a l’extrem meridional de l’oest australià.
En la zona de transició entre els boscos de “jarrah” i els boscos de “karri” trobem formacions boscoses dominades pel “marri” (Eucalyptus calophylla), un altre eucaliptus que s’endinsa també en la zona del “jarrah” donant lloc a boscos mixtos.
En zones lleugerament més seques, amb precipitacions entre 500 i 600 mm anuals, domina el “wandoo” Eucalyptus redunca, que forma un bosc més clar. En les zones que només reben de 300 a 500 mm de precipitació a l’any, tal com succeeix en la major part de la zona mediterrània meridional australiana, apareixen formacions arbustives laxes anomenades “mallee”, un nom de procedència indígena que designa unes comunitats dominades per més d’un centenar d’espècies diferents d’eucaliptus (Eucalyptus). Tant el “wandoo” com el “mallee” han estat profundament transformats i convertits en pastures i conreus de cereals.
Als llocs més secs (menys de 400 mm anuals de pluja i 7 mesos de secada) i els de sòls sorrencs extremament pobres, apareixen formacions arbustives dominades per proteàcies, amb algunes espècies d’eucaliptus (Eucalyptus) i de casuarina o “bull oak” (Casuarina). Tot i que són comunitats molt poc productives, la seva riquesa florística és remarcable.
El “mallee” sud-oriental
L’esquema anterior és vàlid per a la regió d’Austràlia Occidental, però no pas per a l’altra àrea mediterrània, situada a Austràlia Meridional. Només l’1% d’aquesta regió rep una pluviositat superior als 600 mm anuals, cosa que fa que hi manquin els boscos i que estigui dominada per formacions arbustives o arbredes poc denses.
Les formacions arbòries obertes presenten alguna de les 18 espècies d’eucaliptus de port arbori pròpies d’aquesta regió, com per exemple Eucalyptus obliqua, el conegut “sugar gum” (E. cladocalyx) o l’eucaliptus blau (E. viminalis). Però també s’hi poden trobar algunes àrees amb masses arbòries de casuarina (Casuarina luehmannii) o de “cypress pine” (Callitris columellaris).
Aquí, les formacions arbustives de “mallee” són dominades per un gran nombre d’espècies d’eucaliptus (Eucalyptus) i d’acàcies (Acacia), acompanyades d’altres que pertanyen als gèneres Callitris i Casuarina.
Es distingeixen set grans tipus de “mallee”, agrupats en dues subcategories i dominats per espècies diferents, segons les característiques climàtiques i edàfiques. Un primer grup en reuneix tres que tenen plantes arbustives de fulla petita al sotabosc, dominades respectivament pel “mallée” blanc costaner (Eucalyptus diversifolia) la primera, Eucalyptus incrassata i “broombush” (Melaleuca uncinata) la segona, i per E. incrassata i Triodia irritans la tercera. Un segon grup reuneix les de sotabosc dominat per herbàcies o quenopodiàcies: l’una amb el mateix “mallée blanc costaner” (E. diversifolia), l’altra amb el “mallée” blanc (E. behriana), l’altra amb el “mallée” roig E. socialis i E. dumosa, i l’última amb E. oleosa i Triodia irritans. El primer grup correspon als típics dels sòls més pobres i presenten més similituds ecològiques amb els sistemes arbustius de sòls oligotròfics d’altres biomes que no pas amb els pròpiament mediterranis. De fet, al mateix continent australià hi ha comunitats del tipus “mallee” instal·lades en sòls pobres de les zones climàtiques temperada i tropical.
En els llocs més àrids o de sòls més pobres apareixen formacions arbustives baixes (menys de 2 m) amb diverses espècies de proteàcies (dels gèneres Banksia, Hakea, Melaleuca, Leptospermum) acompanyades d’algunes espècies dels gèneres Eucalyptus, Casuarina i Xanthorrhoea, entre d’altres. Hi ha, també, algunes àrees dominades per vegetació herbàcia, amb espècies de gramínies com ara el “cutting grass” (Gahnia trifida) i el “thatching grass” (G. filum), o de liliàcies com les “iron grass” (Lomandra dura i L. effusa).