L’ample ventall de les coníferes
Els boscos característics de les selves temperades són les laurisilves, selves formades majoritàriament per arbres perennifolis, de fulla verda brillant i coriàcia (semblant a la fulla dels arbres de les selves tropicals), però també hi ha selves formades majoritàriament per coníferes, i selves mixtes d’espècies perennifòlies, caducifòlies i coníferes. Les coníferes, en efecte, són abundants a les selves temperades, a diferència del que passa a les selves tropicals. En tot cas, totes les selves temperades comparteixen un origen comú remot, arctoterciari, que explica la importància que hi tenen les falgueres, les coníferes i algunes de les famílies més antigues d’angiospermes, com les magnoliàcies i altres famílies de laurifolis. Les diferents selves temperades de l’hemisferi austral, malgrat les grans distàncies que separen els seus territoris respectius, comparteixen també l’herència de la flora anterior a la fragmentació del continent de Gondwana. L’auge de les angiospermes i la davallada de les coníferes —i altres gimnospermes— és un dels canvis biogeogràfics més interessants que han tingut lloc en la història de la Terra. Actualment, tan sols hi ha unes 550 espècies de coníferes enfront de les aproximadament 250 000 espècies d’angiospermes que han estat descrites. La família de les araucariàcies, juntament amb la de les lebaquiàcies, representada només per formes fòssils, és la més antiga del grup.
Les coníferes són un grup de vegetals, generalment de port arbori, que si bé estan àmpliament distribuïdes pel planeta, són especialment abundants a les zones temperades i fredes d’ambdós hemisferis, on constitueixen la principal font de fusta. Entre les coníferes es troben els vegetals més grans del món i els més longeus; és el cas de l’“ahuehuete” o xiprer de Moctezuma (Taxodium mucronatum), arbre de tronc enormement gruixut (a Mèxic hi ha un exemplar amb un tronc de 42 m de diàmetre, un volum de 700 m3 i un pes aproximat de 550 t que s’estima que té uns 2 000 anys d’edat), i de les colossals sequoies, com la sequoia costanera (Sequoia sempervirens) i la sequoia gegant o de muntanya (Sequoiadendron giganteum), que poden fer més de 100 m d’alçada i assoleixen igualment longevitats mil·lenàries. La diversitat de formes i colors de les seves capçades, juntament amb seu el fullatge sempre verd, han afavorit l’ús freqüent de les coníferes com a plantes ornamentals. A més a més, han demostrat tenir qualitats plàstiques per a aquests propòsits, ja que un nombre considerable d’espècies que en condicions naturals tenen port arbori es poden fer créixer amb port arbustiu en jardins. Si bé la quasi totalitat de les coníferes són perennifòlies, algunes espècies perden la fulla a la tardor i a l’hivern, com és el cas d’alguns Larix, Pseudolarix i dels taxodis (Taxodium).
En llocs on les condicions són adequades per a un creixement ràpid dels arbres, les pinàcies i la resta de coníferes solen ser desplaçades per les angisopermes. Tot i això, les coníferes persisteixen en aquells indrets on les angiospermes no poden competir avantatjosament. Algunes de les característiques que les afavoreixen són la seva gran longevitat i el fet que assoleixen dimensions més grans, trets que els permeten, d’una banda, la permanència com a components de la vegetació durant segles, o fins i tot mil·lennis, i, de l’altra, superar pertorbacions a les quals les angiospermes no sobreviuen. Així, de vegades poden mantenir fàcilment les seves poblacions i la seva àrea de distribució, i fins i tot estendre-la, tal com succeeix amb els boscos de la costa NW de l’Amèrica del Nord.
Les araucariàcies, les més antigues
La família de les araucariàcies és considerada la més antiga de les famílies vivents de coníferes; se’n coneixen fòssils des del Mesozoic, i alguns corresponen sense cap mena de dubte a espècies actuals. Només n’hi ha dos gèneres vivents, Araucaria i Agathis, i si bé ambdós són molt importants a les selves temperades, també se’n troben algunes espècies en àrees tropicals dels països riberencs o insulars del Pacífic. Les araucàries sud-americanes estan actualment confinades en àrees molt limitades dels Andes xilenoargentins (Araucaria araucana) i de la regió del Paranà (A. angustifolia), que constitueixen els darrers refugis testimonials d’una vegetació forestal més antiga que durant el Terciari dominava el paisatge de la major part dels continents australs. El “kauri” (Agathis australis) és el gegant de les selves neozelandeses; pot arribar a assolir els 60 m d’alt i un gruix de 7 m de diàmetre a la base del tronc. En el passat, aquesta espècie havia arribat a ocupar 1,5 milions d’ha en els sòls més pobres de les regions septentrionals de l’Illa del Nord, al N del paral·lel 38°S, però aquesta extensa àrea de distribució ha quedat reduïda a unes 50 000 ha, confinades sobretot a les cotes de més altitud.
Les pinàcies, les més abundants i difoses

Les pinàcies són les coníferes més difoses a les selves temperades, ja sigui com a arbres dominants o bé com a acompanyants. Entre les més abundants hi ha els avets (Abies), les pícees (Picea) i els pins (Pinus). A la il·lustració apareixen representats l’avet de Colorado (Abies concolor), l’avet roig americà (A. amabilis), l’avet gegant americà (A. grandis), l’avet de l’Himàlaia (A. spectabilis), l’“oyamel” (A. religiosa), la pícea de l’Himàlaia (Picea smithiana), la pícea ploranera (P. spinulosa), la pícea de Sitka (P. sitchensis), el pi d’Elliot (Pinus elliottii), el “southern pine” (P. palustris), el pi de Moctezuma (P. montezumae), el pi blau indi (P. wallichiana) i el pi canari (P. canariensis). Altres gèneres de pinàcies també força corrents són les tsugues o hemlocks (Tsuga), de les quals són una mostra el hemlock de l’Himàlaia (T. dumosa) i l’occidental (T. heterophylla), i les Pseudotsuga, gènere al qual pertany l’avet de Douglas (P. menziesii).
Jordi Corbera
Els gèneres de coníferes més comuns a la major part del planeta, sobretot a l’hemisferi nord, són les pinàcies. Els pins (Pinus), les pícees (Picea), els avets (Abies) i les tsugues i hemlocks (Tsuga) tenen un paper considerablement important en boscos boreals, muntanyencs, temperats i fins i tot tropicals. Sense anar més lluny, el bioma dels boscos boreals està dominat en la seva totalitat per pinàcies; a més, aquesta família domina també i està altament diversificada en diferents regions de selves temperades, com les costes del NW de l’Amèrica del Nord o les muntanyes mesoamericanes.
Les selves plujoses temperades del NW de l’Amèrica del Nord estan dominades per l’avet de Douglas (Pseudotsuga menziesii), una espècie que pot viure més de 1 000 anys i atènyer 90 m d’alçada i un diàmetre de 3 m. La diversitat de coníferes, però, i més particularment de pinàcies, és espectacular en aquests boscos plujosos temperats. Es tracta sovint d’altres espècies grans i també molt longeves, com el hemlock occidental (Tsuga heterophylla) i l’avet roig americà (Abies amabilis), que són codominants amb l’avet de Douglas (el primer fonamentalment a les altituds baixes i mitjanes i el segon més muntanyenc) en tota l’àrea que s’estén des del N de l’illa de Vancouver fins als 55°N, al SE d’Alaska, o la pícea de Sitka (Picea sitchensis), espècie molt comuna al llarg de tota la costa, sobretot a les ribes de rius i llacs. Amb aquestes, però, es barregen també altres espècies de dimensions més modestes com l’avet de Colorado o avet blanc americà (Abies concolor), el pi groc o pi ponderosa (Pinus ponderosa) o el pi de Lambert o pi gegant californià (P. lambertiana). L’avet gegant americà (Abies grandis) recolonitza les àrees desforestades dels indrets més humits, mentre que la varietat glauca de l’avet de Douglas (Pseudotsuga menziesii var. glauca) fa el mateix en indrets més secs, al N de les Muntanyes Rocalloses d’Idaho. El hemlock californià o avet de Mertens (Tsuga mertensiana), per la seva banda, es troba a 1 200 m d’altitud a Oregon, però al S d’Alaska se situa al nivell de la mar, on fa el paper de codominant amb la pícea de Sitka.
Les pinàcies, més concretament els pins, són també característics de les planes costaneres del SE dels Estats Units, on els sòls són pobres i sorrencs i els incendis força freqüents. Les espècies més abundants són el pi de fulla llarga o “southern pine” (Pinus palustris), el pi d’Elliot (P. elliottii var. elliottii), el pi “loblolly” (P. taeda) i el pi cedre (P. glabra).
Les muntanyes mesoamericanes, principalment les de Mèxic, amb set espècies d’avet i més d’una trentena de pins, són un gresol de biodiversitat pel que fa a les pinàcies, que hi acostumen a tenir una distribució molt localitzada. De fet, algunes espècies només es coneixen en petites àrees de determinades serralades muntanyoses. Entre les més comunes o més típiques destaquen el pi blanc mexicà o “ayacahuite” (Pinus ayacahuite), el pi de Moctezuma (P. montezumae) i el pi de Nicaragua (P. oocarpa), que estén la seva àrea de distribució des de les muntanyes de Mèxic fins a les del N de Nicaragua, i entre els avets, l’“oyamel” (Abies religiosa).
Al seu límit superior, les laurisilves himalaianes deixen pas a boscos temperats de pinàcies dominats per arbres que poden assolir els 30 o 40 m d’alçada, com la pícea de l’Himàlaia (Picea smithiana), la pícea ploranera (P. spinulosa), el hemlock de l’Himàlaia (Tsuga dumosa), l’avet de l’Himàlaia (Abies spectabilis), i pins com el pi blau indi (Pinus wallichiana) o el pi de tres fulles de l’Himàlaia (P. roxburghii), tots ells arbres que també apareixen sobresortint del dosser barrejats amb la laurisilva. Un cas paral·lel es dóna a les Canàries amb les pinedes de pi canari (P. canariensis)— un arbre que sembla estar estretament emparentat amb el pi de tres fulles de l’Himàlaia—, les quals, a les illes de relleu més alterós, formen un estatge altitudinal per damunt del de la laurisilva.
Les cupressàcies i les taxodiàcies, les més diversificades
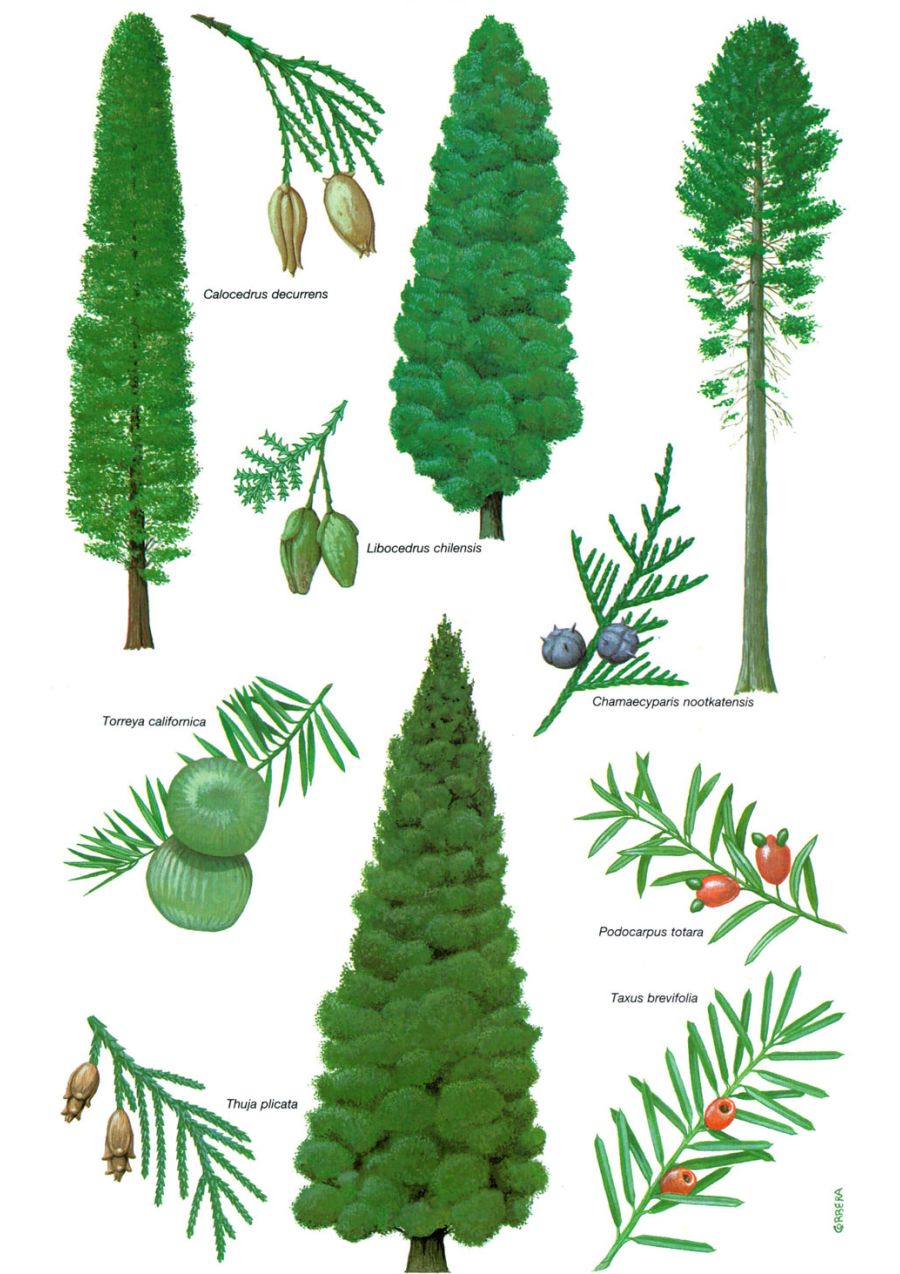
Les cupressàcies (cedres i xiprers) i les taxodiàcies (taxodis) són les coníferes més diversificades dels boscos temperats, amb representants a la majoria d’àrees del bioma. Entre les primeres hi ha el xiprer de les Guaitecas o “ten” (Pilgerodendron uvifera), un arbre o arbust alt de capçada cònica i fulles esquamiformes i perennes que creix a les selves mixtes valdivianes, a les nord-patagòniques i a les magallàniques; es converteix així en la conífera que assoleix les latituds més australs. Una altra cupressàcia pròpia de les selves temperades de l’extrem més meridional del continent americà, que tanmateix no s’estén tan al S com l’anterior, és el xiprer xilè (Libocedrus [=Austrocedrus] chilensis), un arbret de tronc recte, de com a molt 20 m d’alçada, i de capçada estreta i densa. Als boscos temperats de la meitat N del continent americà també hi ha representants de les cupressàcies, entre els quals la tuia gegant (Thuja plicata), l’“Alaska yellow-cedar” (Chamaecyparis nootkatensis) i el cedre d’encens californià (Calocedrus decurrens). A la làmina es representen també el “t¯otara” (Podocarpus totara), una podocarpàcia neozelandesa, i dues taxàcies dels boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord, Taxus brevifolia, el teix del Pacífic, i Torreya californica, la nou moscada californiana.
Jordi Corbera
Cupressàcies i taxodiàcies són dues famílies amb considerables afinitats (fulles en forma d’esquames o d’acícules curtes, pinyes o gàlbuls arrodonits o ovoides, llavors no carnoses), fins al punt que en el passat havien estat considerades com una única família. Malgrat aquestes similituds, però, acullen en conjunt vegetals molt diferents que viuen en medis força dispars; inclouen des de les coníferes més gegantines, com les sequoies (Sequoia, Sequoiadendron, Metasequoia) i alguns taxodis com l’“ahuehuete” o xiprer de Moctezuma (Taxodium mucronatum), fins a les que no solen passar d’arbustos i només excepcionalment assoleixen el port arbori, com càdecs i ginebres. Comprenen també des de plantes d’aiguamoll, com el taxodi de Virgínia o “bald cypress” (T. distichum), fins a altres que viuen en els ambients més secs de les muntanyes mediterrànies o dels subdeserts nord-americans.
A les selves temperades de l’extrem austral del continent americà s’hi fan dues cupressàcies força singulars: el làrix americà o “alerce” (Fitzroya cupressoides) i el “ten” o xiprer de les Guaitecas (Pilgerodendron uvifera). El làrix americà es troba a la Cordillera de la Costa, al S de Valdivia, i també a l’illa de Chiloé i als Andes, a cotes situades entre els 700 i els 1 400 m. És considerada una espècie relicta, confinada a hàbitats on altres espècies no es poden establir i, a més, és l’espècie més longeva de l’Amèrica del Sud, amb exemplars que arriben als 3 600 anys i atenyen grans dimensions. Però la sobreexplotació a què ha estat sotmesa per la seva valuosa fusta ha fet disminuir dràsticament el nombre d’arbres vells, que es troben cada cop més rarament, i ha calgut protegir l’espècie. El xiprer de les Guaitecas és la conífera que presenta l’àrea de distribució més austral de tot el planeta, ja que arriba fins a la Terra del Foc. A la selva valdiviana es troba principalment amb el làrix americà, però més cap al S, als arxipèlags del S de Xile i a la Terra del Foc, s’associa més aviat amb els faigs australs (Nothofagus). Una tercera cupressàcia d’aquesta regió que de vegades es dóna a l’E dels Andes, sota precipitacions que oscil·len entre els 1 000 i els 1 500 mm, i s’associa al “pehuén” (Araucaria araucana), és el xiprer xilè (Libocedrus [=Austrocedrus] chilensis), que al llarg de la vora occidental del subdesert patagònic forma l’ecotò entre aquest i les selves temperades de la banda més plujosa de la serralada dels Andes.
La tuia gegant o “western red-cedar” (Thuja plicata), la cupressàcia més apreciada dels boscos del NW de l’Amèrica del Nord, alterna amb el hemlock occidental (Tsuga heterophylla) al bosc de la conca del Colúmbia i al N de les Muntanyes Rocalloses, on esdevé dominant als indrets més humits. Al llarg de la costa de la Colúmbia Britànica es presenta en associació amb l’“Alaska yellow-cedar” (Chamaecyparis nootkatensis), una altra cupressàcia àmpliament distribuïda per aquesta regió. A Alaska, la tuia gegant forma boscos densos, tant en indrets humits com al llarg dels cursos d’aigua. És un arbre de grans dimensions, i la seva fusta imputrescible era molt utilitzada com a material principal de construcció pels pobles autòctons, tant en les cases com en les embarcacions. El cedre d’encens californià (Calocedrus decurrens), en canvi, és una cupressàcia de dimensions més modestes i de fusta tova (molt apreciada, tanmateix, per fer-ne llapis), que creix en indrets més secs i es troba al llindar mateix del bioma. Aquest és també el cas de les gegantines sequoies costaneres (Sequoia sempervirens), que són l’espècie dominant a l’extrem meridional dels boscos de la costa NW i que assoleixen aquí les seves dimensions més grans. Als aiguamolls del SE dels Estats Units, les taxodiàcies estan representades pel taxodi de Virgínia o d’aiguamoll (Taxodium distichum), també anomenat “bald cypress” (‘xiprer calb’).
Al Japó, és remarcable el “kohyamaki” o pi parasol japonès (Sciadopitys verticillata), una espècie que només es fa espontàniament en dues localitats del centre de Honshū, però que al Terciari era força comú a gran part del continent eurasiàtic; avui, malgrat la seva raresa com a planta espontània, és freqüent trobar-la plantada al voltant dels temples sintoistes. També és endèmic, en aquest cas de les illes macaronèsiques, el ginebre de les Açores (Juniperus brevifolia), que acostuma a formar ginebredes característiques en aquestes illes.
Les podocarpàcies i les filocladàcies, les més tropicals
Podocarpàcies i filocladàcies són coníferes que no presenten pròpiament els cons que donen nom a la família, sinó que tenen les llavors situades a l’extrem d’una branca i rodejades —ja sigui a la base o bé tot al voltant de la llavor— per un embolcall carnós; sembla que aquest conjunt és atractiu per als ocells que fan la funció disseminadora. Aquestes famílies tenen una distribució principalment austral, però a les latituds tropicals, sobretot a l’Àsia meridional, on estan àmpliament difoses, arriben a penetrar a l’hemisferi nord; una d’elles, Podocarpus macrophyllus, arriba fins al SW del Japó, on es coneix amb el nom d’“inumaki”. Les filocladàcies comprenen un únic gènere, Phyllocladus, els “celery pines”, que tenen representants tant a Nova Zelanda i Tasmània com a Nova Guinea i Borneo, i que es caracteritzen per la reducció extrema de les fulles veritables, però també per presentar uns eixamplaments de les rametes (fil·locladis) amb l’aspecte i la funció de fulles. Alguns autors no reconeixen aquest gènere com a família separada de les podocarpàcies.
La majoria dels podocarps (Podocarpus) són grans arbres de distribució principalment tropical, però també són presents a les selves temperades de l’hemisferi austral i, tant al sotabosc d’aquestes com a l’alta muntanya de Nova Zelanda i en altres punts de la seva àrea de distribució, fins i tot hi ha representants de port arbustiu. A les selves temperades de Tasmània, entre els 1 000 i 1 500 m d’altitud, creix el “plum pine” (P. lawrencii), el nom del qual fa referència al característic embolcall de la llavor, que té l’aspecte d’una petita pruna. A les selves temperades de Nova Zelanda, d’altra banda, s’hi fan principalment el “tōtara” (P. totara) i el “tōtara” de muntanya o de Hall (P. hallii). De fet, moltes de les selves de l’Illa del Nord de Nova Zelanda estan dominades per podocarpàcies com el “rimu” (Dacrydium cupressinum), el “kahikatea” (Dacrycarpus dacrydioides), el “mataī” (Prumnopitys taxifolia) i el ja esmentat “tōtara”. A la selva valdiviana predominen el “mañío” de fulles punxants (Podocarpus nubigenus) i el “mañío” de fulles llargues (P. salignus).
Les taxàcies i les cefalotaxàcies: les més atípiques
Taxàcies i cefalotaxàcies tindrien, segons alguns autors, un origen diferent que la resta de coníferes, aparegudes entre el Carbonífer superior i el Permià, aparentment a partir de les lebaquiàcies. Taxàcies i cefalotaxàcies, en canvi, derivarien d’ancessors diferents i tindrien un origen més antic. Com les podocarpàcies, en comptes de cons tenen òvuls terminals que donen llavors protegides d’un embolcall carnós. En el cas de les taxàcies, aquest embolcall rep el nom d’aril.
El teix europeu (Taxus baccata), difós tant pel bioma de les boscanes decídues com per la muntanya mediterrània, té a les selves temperades euxíniques el límit oriental de la seva àrea de distribució. A l’estrat arbori inferior dels boscos d’avet de Douglas (Pseudotsuga menziesii) i hemlock occidental (Tsuga heterophylla) de la costa NW de l’Amèrica del Nord, el teix del Pacífic (Taxus brevifolia) és l’arbre dominant, i també apareix una mica més cap al S, en els boscos californians de sequoies, juntament amb una altra taxàcia, la nou moscada californiana (Torreya californica). Dues taxàcies més, el cedre pudent o “stinking-cedar” (T. taxifolia) i el teix de Florida (Taxus floridana), són endèmiques d’una petita àrea de la regió sud-oriental dels Estats Units, del curs inferior del riu Apalachicola, al NW de Florida i de les àrees limítrofes del SW de l’estat veí de Geòrgia.
Les cefalotaxàcies són pròpies de les selves temperades asiàtiques, des de l’E de l’Himàlaia fins al Japó. Algunes espècies, com la japonesa “inugaya” (Cephalotaxus harringtonia) o la xinesa C. fortunei sovint són cultivades, tant pels seus fruits com pel seu caràcter ornamental.
Els arbres laurifolis i altres planifolis
El que marca veritablement el caràcter de la majoria de les selves temperades, però, no són les coníferes, sinó els planifolis de fulles amples, perennes, lleugerament coriàcies, d’un verd lluent, que se solen conèixer amb el nom de laurifolis. Encara que l’adjectiu laurifoli deriva del nom llatí dels llorers (Laurus), els membres de la família de les lauràcies no són els únics que presenten aquest tipus de fulles. Ben al contrari, els vegetals laurifolis pertanyen a un gran nombre de famílies, moltes de les quals compartides per selves temperades i selves tropicals. Un cas paradigmàtic seria el de les selves plujoses de l’E d’Austràlia, on el trànsit de tropicals a temperades es fa sense solució de continuïtat i unes i altres comparteixen plantes de famílies molt primitives, algunes endèmiques del continent australià o compartides només amb la veïna Nova Guinea. Una de les més notables és l’Austrobaileya scandens, l’única espècie de la família de les austrobaileiàcies, endèmica de les selves tropicals del NE d’Austràlia i de Nova Guinea. Una altra és l’idiosperm (Idiospermum australiense), l’únic representant de la família de les idiospermàcies, acantonat en un nombre molt limitat de localitzacions de les selves tropicals del NE de Queensland. Altres plantes d’aquestes famílies primitives d’angiospermes ja es troben tant a les selves tropicals com a les temperades; aquest és el cas dels tres gèneres endèmics de monimiàcies (Doryphora, Atherosperma i Daphnandra), que alguns autors situen en una família independent, la de les aterospermatàcies, o també el d’Eupomatia laurina, una de les dues espècies de la família de les eupomatiàcies, que es fa a les selves plujoses de l’E d’Austràlia, des del NE de Nova Gal·les del Sud fins al NE de Queensland, i també a Nova Guinea.
A diferència del que s’esdevé a la majoria de les boscanes decídues i tal com passa a les selves tropicals, a les selves temperades no es manifesta cap dominància clara d’una determinada espècie sinó més aviat d’una combinació d’espècies, i una gran varibilitat local en l’abundància relativa de cada una d’elles. En qualsevol cas, cal destacar i remarcar el pes específic d’algunes famílies, sobretot perquè sovint es tracta de famílies molt antigues que conserven els trets més arcaics de les angiospermes: presència de traqueides en comptes de vasos, disposició de les peces florals en espiral i nombre indefinit (i sovint elevat) de peces en cada verticil, i flors grans, vistoses i fragants. La presència de fulles amples, lanceolades o ovals, poc retallades, perennes i lleugerament endurides constitueix també un tret arcaïtzant que té molt a veure amb el fet que les condicions climàtiques de les actuals selves temperades reprodueixen les de les selves subtropicals, que és on s’originaren moltes d’aquestes famílies durant el Terciari.
Les magnòlides: magnòlies, llorers, canyellers i altres angiospermes arcaiques
Un nombre considerable d’aquestes famílies, com les magnoliàcies, les winteràcies, les esquisandràcies, les austrobaileiàcies, les idiospermàcies, les eupomatiàcies o les il·liciàcies, que s’apleguen en la subclasse de les magnòlides, figuren entre les més antigues de les angiospermes. De totes elles, les que tenen un paper més destacat en les selves temperades són les lauràcies i les magnoliàcies, mentre que winteràcies, il·liciàcies, monimiàcies i piperàcies tenen un paper més secundari. Gairebé totes presenten una distribució tropical i subtropical.
Les lauràcies i les famílies afins

Les lauràcies i algunes famíles afins són les angiospermes més característiques de les selves temperades. A la làmina s’han representat sis lauràcies —el “loro”, llorer canari o llorer de les Açores (Laurus azorica [=L. canariensis]), el “redbay” (Persea borbonia), el “lingue” (P. lingue), el “tabunoki” (Persea [=Machilus] thunbergii), el til (Ocotea foetens) i el camforer (Cinnamomum camphora)— i una monimiàcia, el llorer xilè (Laurelia sempervirens) Els laurifolies, originats molt probablement al Triàsic (tot i que s’ha trobat pol·len fòssil del Carbonífer que podria pertànyer a alguna espècie primitiva d’angiosperma), durant molt temps foren simples elements secundaris de les formacions vegetals del planeta, dominades per gimnospermes. A partir del Cretaci mitjà, però, les angiospermes començaren a expandir-se ràpidament i acabaren convertint-se en els arbres dominants en molts boscos. El factor responsable d’aquesta ràpida expansió podria ser tant un canvi ambiental que hagués afavorit les angiospermes com la possibilitat que aquestes haguessin evolucionat fins a assolir unes característiques que els donessin certs avantatges davant les gimnospermes. Entre aquestes característiques hi ha un cicle de vida curt, una producció abundant de llavors i una pol·linització altament eficaç ajudada per insectes i altres animals.
Jordi Corbera
Així, les lauràcies es troben distribuïdes per tots els continents, principalment en àrees tropicals i subtropicals. Els seus centres de màxima diversitat se situen al SE d’Àsia i a l’Amèrica tropical, però es conserven espècies relictes en diferents punts d’una àrea força més dilatada, a latituds superiors que van des de la regió mediterrània o el NW de l’Amèrica del Nord fins al S de Xile o Nova Zelanda. A les laurisilves macaronèsiques, les de la regió del Paranà i les sinojaponeses, les lauràcies hi estan particularment ben representades.
El “loro”, llorer canari o llorer de les Açores (Laurus azorica [=L. canariensis]) és l’espècie dominant a les laurisilves de les Canàries i Madeira, i l’únic endemisme compartit per tots els arxipèlags macaronèsics. A les laurisilves macaronèsiques s’hi fan, a més, altres lauràcies, com el til (Ocotea foetens), el “barbusano” (Apollonias barbujana) i el “viñátigo” (Persea indica). Diferents espècies del gènere Persea apareixen també al SE dels Estats Units —el baguer vermell o “redbay” (P. borbonia) i el baguer d’aiguamoll o “swamp bay” (P. palustris)—, al S de Xile —els “lingues” (P. lingue i P. meyeniana)— i al Japó, on el “tabunoki” (P. [=Machilus] thunbergii) és un dels arbres més importants de les laurisilves i el “hosobatabu” (P. [=Machilus] japonica) el seu destacat acompanyant.
A les laurisilves de la regió del Paranà les lauràcies estan representades per una colla de gèneres, principalment Ocotea, Cryptocarya i Nectandra. A la part brasilera d’aquesta àrea, la majoria d’aquests arbres reben el nom de “canelas”: la “canela sassafras” (Ocotea pretiosa) i les “canelas amarelas” (Nectandra nitidula, N. lanceolata i N. puberula). A la selva missionera argentina es coneixen més aviat com a “laureles”: el “laurel negro” o “aju’y hu”, en guaraní (N.angustifolia [=N. saligna]), i el “laurel de río” (N. falcifolia). Al sotabosc dels boscos de sequoia (Sequoia) i als d’avet gegant americà (Abies grandis) i tuia gegant (Thuja plicata) del N de Califòrnia té un important paper el llorer californià (Umbellularia californica), mentre que a les laurisilves del N de Nova Zelanda el tenen el “taraire” (Beilschmiedia tarairi) i el “tawa” (B. tawa).
Particularment diversificades estan les lauràcies de les laurisilves asiàtiques (sovint anomenades boscos laurofagacis pel predomini que hi tenen lauràcies i fagàcies). Ja s’ha fet esment del gènere Persea, que les laurisilves asiàtiques tenen en comú amb les d’altres zones de selves temperades, i aquest també és el cas d’Apollonias (compartit amb les laurisilves macaronèsiques) o de Lindera (compartit amb les del SE dels Estats Units). També hi ha gèneres comuns a les laurisilves asiàtiques i a les selves temperades d’Austràlia i Nova Zelanda, i fins i tot a les primeres i les selves tropicals de la regió indomalaia (Actinodaphne, Alseodaphne, Cinnamomum, Cryptocarya, Dehaasia, Litsea, Neolitsea, Phoebe). Al Japó, les espècies més destacades a l’estrat arbori són probablement el camforer (Cinnamomum camphora) i el “tabunoki” (Persea [=Machilus] thunbergii), però altres arbres com el camforer japonès o “yabunikkei” (Cinnamomum japonica) —més resistent al fred que el seu congènere, el “hosobatabu” (Persea [=Machilus] japonica)— o el “kuromoji” (Lindera umbellata) són destacats acompanyants.
La família de les monimiàcies és afí a la de les lauràcies, i a un dels seus gèneres, Laurelia, pertanyen molts dels llorers australs, com el “laurel” o llorer xilè (L. sempervirens [=L. serrata]), el “huahuán” o “tepa” (L. philippiana) i altres arbres i arbustos de les selves valdivianes, així com el “pukatea” (L. novae-zelandiae) dels fons de les valls i indrets humits de l’Illa del Nord i de l’extrem septentrional de l’Illa del Sud de Nova Zelanda. El sassafràs de Tasmània (Atherosperma moschatum) és un dels arbres característics dels boscos de faigs australs d’aquella illa i del S de l’estat australià de Victòria, en els quals forma un segon cobricel —a uns 20 m d’alçada— amb les seves capçades, per sota del dels faigs australs; de vegades fins i tot esdevé l’espècie dominant d’alguns boscos secundaris.
Les magnoliàcies i les famílies afins
La de les magnoliàcies és l’altra gran família de magnòlides de les laurisilves. N’hi ha de perennifòlies i de caducifòlies i la seva distribució es limita a les regions càlides i temperades d’Amèrica i Àsia. La magnòlia comuna (Magnolia grandiflora), és una de les espècies perennifòlies emblemàtiques de l’estrat arbori superior del bosc planifoli mixt meridional, propi de la Plana Costanera Atlàntica del SE dels Estats Units, mentre que la magnòlia d’aiguamoll (M. virginiana) es fa als rodals de cedre blanc atlàntic (Chamaecyparis thyoides), en cursos d’aigua lents i al voltant de les basses permanents d’aquella plana costanera, des del N de Florida fins a Maine, i sobretot a la costa de Carolina del Nord, de Virgínia i del S de Nova Jersey, i en altres comunitats d’aiguamoll. Al continent asiàtic i al Japó les magnoliàcies estan força més diferenciades en gèneres distints que a Amèrica i, a més de magnòlies (Magnolia), s’hi fan magnoliàcies dels gèneres Elmerrillia, Manglietia i Michelia, com el “champak” o “sapu” (M. champaca) dels vessants meridionals de l’Himàlaia.
De totes les famílies d’angiospermes, la de les winteràcies és possiblement, junt amb la de les magnoliàcies, la que presenta més trets arcaics. Les winteràcies són un dels pocs grups d’angiospermes que tenen el teixit conductor format de traqueides —com el de les gimnospermes—, i les seves flors mostren caràcters que les acosten a allò que devien ser les angiospermes més primitives. La seva distribució és bàsicament austral (Amèrica del Sud, Austràlia, Nova Zelanda, Nova Caledònia i Madagascar), però tant a la regió indomalaia com a la neotropical tenen alguna presència a l’espai tropical al N de l’equador. El gènere Drimys, americà, és potser el més representatiu. Hi pertanyen tant el “canelo” xilè o “boighe” (D. winteri), de les selves valdivianes, com D. brasiliensis, de les selves de la regió del Paranà. També en forma part el gènere Pseudowintera, un dels representants del qual és l’“horopito” (P. colorata) dels boscos montans mixtos de l’Illa del Sud de Nova Zelanda.
La de les il·liciàcies és una família estretament relacionada amb les de les winteràcies i les magnoliàcies i que comprèn un únic gènere amb dues espècies. Una, l’anís estrellat xinès (Illicium verum), viu a les laurisilves de la Xina i el Japó meridionals; l’altra, l’anís estrellat groc (I. parviflorum), a les ribes del golf de Mèxic.
També es relaciona amb les magnoliàcies la família de les annonàcies, a la qual pertanyen espècies com el “pawpaw” (Asimina triloba) dels boscos mixtos del SE dels Estats Units.
Les hamamèlides: liquidàmbars, roures, alzines i faigs australs
Les hamamèlides, també anomenades amentíferes, apleguen principalment plantes arbòries o arborescents de fulla caduca; estan especialitzades en la pol·linització anemòfila i per això tenen les flors reduïdes, unisexuals i reunides en inflorecències denses característiques anomenades aments. Moltes hamamèlides són arbres forestals de gran importància a les boscanes decídues, però també n’hi ha que tenen un paper cabdal a les selves temperades mixtes de tots els continents, principalment a l’hemisferi boreal, i a les mediterrànies.
Les hamamelidàcies i les famílies afins
Tres famílies monotípiques (és a dir, formades per un sol gènere que compta amb una única espècie), trocodendràcies, eupteleàcies i tetracentràcies, les més primitives de les hamamèlides, comparteixen amb les winteràcies —de les quals probablement derivaren— i altres famílies afins el tret de no posseir vasos conductors, sinó només traqueides. Relíquies de la flora arctoterciària, aquestes tres famílies són pròpies de les selves temperades sinojaponeses i himalaianes. Trochodendron aralioides, únic representant vivent de les trocodendràcies, és un arbre laurifoli l’àrea de distribució del qual es prolonga des del Japó fins a Taiwan. El tetracèntron xinès (Tetracentron sinense), únic representant vivent de les tetracentràcies, creix a la Xina meridional i arriba, pel vessant S de l’Himàlaia, fins al riu Arun, a l’E del Nepal. Euptelea polyandra, únic representant vivent de les eupteleàcies, té una distribució semblant.
Liquidàmbars (Liquidambar) i avellaners de bruixa (Hamamelis) són arbres caducifolis mitjans i petits de la família de les hamamelidàcies que es presenten principalment a les boscanes decídues de les regions orientals d’Àsia i Amèrica, i als sotaboscos de les selves temperades d’aquestes regions, on els acompanyen un gran nombre d’espècies d’altres gèneres de la mateixa família. La “rasamala” (Altingia excelsa) i Exbucklandia populnea, per exemple, són espècies arbòries característicament acompanyants dels boscos perennifolis humits sinojaponesos i himalaians, mentre que la parròtia persa (Parrotia persica), que és un arbre endèmic de les selves hircanianes i en domina una petita part, es troba més sovint en diferents variants de les selves dominades pel roure de fulla de castanyer (Quercus castaneifolia).
La fusta de suro americana (Leitneria floridana), un arbrissó característic de la terra baixa humida de les terrasses de les planes d’inundació del baix Mississipí i les costes del golf de Mèxic, de fusta blana i lleugera (més lleugera que el suro, cosa que explica el seu nom), és l’únic representant de la família de les leitneriàcies, una família monotípica endèmica del SE dels Estats Units. Ben al contrari de les leitneriàcies, d’àrea de distribució tan reduïda, les miricàcies són pràcticament cosmopolites i només falten a les regions àrides subtropicals del Vell Món i del continent australià. La “faya” (Myrica faya) és una de les espècies dominants del “fayal-brezal”, una de les formacions característiques de les laurisilves macaronèsiques; al mateix gènere pertanyen arbustos com la “wax myrtle” o murta de cera (M. cerifera), l’“evergreen bayberry” (M. heterophylla) o l’“odorless bayberry” (M. inodora) de les pinedes i aiguamolls del SE dels Estats Units.
Les fagàcies i les betulàcies

Les fagàcies tenen una notable presència a les selves temperades, baldament siguin les boscanes decídues el bioma on presenten la seva màxima difusió. A l’hemisferi sud les més típiques són els faigs australs (Nothofagus), àmpliament distribuïts per l’Amèrica del Sud i Oceania, especialment per Nova Zelanda. Es diferencien dels faigs septentrionals (Fagus) per presentar les flors masculines aïllades, o com a molt, agrupades de dues en dues o de tres en tres, però mai formant grans masses. Les seves fulles petites, de color verd violaci, creixen molt atapeïdes sobre pecíols curts, i poden caure a la tardor o bé mantenir-se damunt l’arbre durant tot l’any. Sovint els faigs australs són els arbres dominants del bosc, com passa, per exemple, amb el “coihue” comú (Nothofagus dombeyi), molt estès a les selves temperades de l’extrem meridional d’Amèrica. A l’altra punta del planeta, a Nova Zelanda, una altra espècie que sovint és dominant és el “hard beech” (N. truncata), un arbre que pot viure fins a 400 o 500 anys, mentre que el “myrtle beech” (N. cunninghamii), de fullatge dens de color verd fosc, ocupa en exclusiva l’illa de Tasmània. A l’hemisferi nord, les fagàcies més abundants solen ser diverses espècies de roures i alzines (Quercus). Alguns també constitueixen boscos on són l’espècie més abundant, com per exemple el roure de fulla de castanyer (Q. castaneifolia), l’arbre més abundant a molts boscos temperats hircanians. Altres, en canvi, són espècies acompanyants, com el roure castanyer d’aiguamoll (Q. michauxii), l’“over-cup oak” (Q. lyrata) i l’alzina llorer d’aiguamoll (Q. laurifolia) dels boscos del SE dels Estats Units.
Jordi Corbera
Les fagàcies i les betulàcies són dues famílies constituïdes principalment per arbres caducifolis propis de les boscanes decídues, encara que una part de les fagàcies siguin arbres o arbustos perennifolis propis de les mediterrànies i de les selves temperades. Tanmateix, tant algunes d’aquestes espècies perennifòlies com algunes de les caducifòlies tenen el seu paper en les laurisilves. Els roures (Quercus), per exemple, amb més d’un centenar d’espècies diferents, són els principals protagonistes dels boscos mixtos mesoamericans de pins i “encinos” (denominació local dels roures). Ben lluny d’aquests boscos, a les laurisilves euxíniques, el roure de Hartwiss (Q. hartwissiana) és un dels arbres dominants, juntament amb dues fagàcies més, el castanyer comú (Castanea sativa) i el faig oriental (Fagus orientalis), mentre que el roure de fulla de castanyer (Quercus castaneifolia) ho és de les hircanianes. A les laurisilves dels vessants meridionals de l’Himàlaia, els roures i altres fagàcies, principalment dels gèneres Castanopsis i Lithocarpus, comparteixen protagonisme amb les lauràcies. El mateix succeeix a les laurisilves del S de la Xina i del S del Japó, en les quals apareixen tant roures com alzines (aquestes darreres aplegades per alguns autors en el gènere Cyclobalanopsis, considerat per altres un subgènere dins de Quercus) i també diferents espècies dels gèneres Castanopsis i Lithocarpus, que tenen el seu centre de màxima diferenciació justament a les muntanyes del SW de la Xina. Entre les alzines més comunes al Japó destaquen l’“akagashi” o alzina japonesa de fulla estreta (Quercus acuta) i l’“urajirogashi” (Q. salicina) i, a l’Himàlaia, el “kharsu” (Q. semecarpifolia).
Als boscos de la costa NW de l’Amèrica del Nord apareixen nombroses fagàcies, tant caducifòlies com perennifòlies, i també alguna betulàcia. Entre les primeres figuren el roure blanc d’Oregon (Quercus garryana) i el roure negre californià (Q. kelloggii) i entre les perennifòlies el “tan oak” (Lithocarpus densiflora), comú al sotabosc de les muntanyes Klamath i, més cap al S, als boscos de sequoies i fins i tot a les mediterrànies californianes, diferents espècies d’alzines, com la “canyon live oak” (Quercus chrysolepis) i l’alzina californiana o “encina” (Q. agrifolia). A les pinedes i, sobretot, a les selves planifòlies mixtes del SE dels Estats Units també són nombrosos els roures i les alzines. En aquestes selves planifòlies mixtes apareixen també altres fagàcies com el faig americà (Fagus grandifolia), que és un dels arbres dominants a l’estrat arbori superior; entre els roures predominen espècies com el roure roig meridional (Quercus falcata), el roure castanyer d’aiguamoll (Q. michauxii), el roure d’aigua (Q. nigra), el roure blanc americà (Q. alba), el “post oak” (Q. stellata) i també, restringits a les terres més baixes i humides, el roure d’Arkansas (Q. arkansana), el “turkey oak” (Q. laevis), el roure d’escorça de cirerer (Q. falcata var. pagodifolia), el roure salze (Q. phellos) o l’“over-cup oak” (Q. lyrata). A les pinedes es troben el roure de Maryland o “blackjack oak” (Q. marilandica), el “sand post oak” (Q. margaretta) i el “bluejack oak” (Q. incana), i entre les alzines, l’alzina de Virgínia o “southern live oak” (Q. virginiana), l’alzina de Darlington o alzina llorer comuna (Q. hemisphaerica) i l’alzina llorer d’aiguamoll (Q. laurifolia).
A les laurisilves de l’hemisferi austral els protagonistes són els faigs australs (Nothofagus), tant caducifolis com perennifolis. Al S de Xile i l’Argentina els caducifolis estan representats per la lenga (N. pumilio), el “roble pellín” (N. obliqua), el “ñirre” (N. antarctica) i el raulí (N. procera), mentre que les espècies perennifòlies són els “coihues” o “coigües”, és a dir, el “coihue” comú (N. dombeyi), el “guindo” o “coigüe” magallànic (N. betuloides) i el “coigüe” de Chiloé (N. nitida). Tots ells són arbres dominants en un o altre dels sistemes forestals de l’extrem austral d’Amèrica.
A Nova Zelanda els faigs australs dominen els boscos de les àrees més seques o més fredes, encara que són presents en diverses proporcions en la majoria de selves en què es barregen coníferes i planifolis. El “hard beech” o faig austral de terra baixa neozelandès (Nothofagus truncata) apareix principalment per sota dels 1 000 m a l’Illa del Nord i a l’extrem N de l’Illa del Sud; tolera els sòls ben drenats i pobres en nutrients. El “red beech” o faig austral roig neozelandès (N. fusca), el faig austral neozelandès que assoleix més alçada (més de 40 m), se situa, en canvi, als pendents col·luvials. A l’E de l’Illa del Sud, les fagedes de “mountain beech” o faig austral muntanyenc neozelandès (N. solanderi var. cliffortioides) formen un límit forestal pràcticament ininterromput al llarg de moltes de les serralades axials i dels Alps Meridionals. A les regions septentrionals i nord-occidentals de l’Illa del Sud sovint domina el “silver beech” o faig austral argentat neozelandès (N. menziesii). Més cap al S, els faigs australs platejat, muntanyenc i roig, coexisteixen al N i a l’E de Fiordland (al SW de l’Illa del Sud). El platejat domina les valls de la regió de Fiordland, on les pluges són molt abundants, i en algunes zones, allà on els sòls són porosos i descansen sobre roques pobres en nutrients, s’hi poden trobar formes nanes, genèticament diferenciades, de faig austral platejat neozelandès.
A Austràlia (i sobretot a Tasmània) el més important dels faigs australs és el “myrtle beech” o faig austral de fulla de murta (Nothofagus cunninghamii). És un arbre de considerable port, el més alt dels faigs australs (fins a 50 m als boscos més densos i més alts de l’W de Tasmània), i forma boscos al S de Victòria i a Tasmània, des del nivell de la mar fins als 1 000 m a Tasmània i fins als 1 300 a l’estat de Victòria. També és l’arbre dominant de l’estrat arbori inferior en boscos de grans eucaliptus de les mateixes regions. Més restringida és l’àrea del “negrohead beech” o faig austral de Moore (N. moorei), que només se sol trobar en emplaçaments rics en nutrients dels confins septentrionals de Nova Gal·les del Sud amb Queensland. El bosc subalpí de “tanglefoot” o faig austral subalpí tasmanià (N. gunnii), d’altra banda, és característic del límit superior del bosc d’algunes zones de Tasmània.
Tal com passa a les boscanes decídues, les betulàcies tenen un paper més secundari que les fagàcies a les selves temperades de tota llei. Només tenen un paper dominant, de vegades, en formacions permanents de les vores de cursos d’aigua o en les formacions secundàries resultants d’alguna pertorbació. És el cas del carpinus del Caucas (Carpinus caucasica) i del vern barbat (Alnus barbata) a les laurisilves euxíniques, o del carpinus de Carolina (Carpinus caroliniana) i l’òstria americana comuna o de Virgínia (Ostrya virginiana) al SE dels Estats Units. El vern roig americà o vern d’Oregon (Alnus rubra [=A. oregana]) es troba als boscos del NW de l’Amèrica del Nord, però només en els estadis inicials de la successió, després que s’hagi produït alguna pertorbació.
Les dil·lènides: camèlies, brucs i arboços
Les dil·lènides representen un nivell evolutiu intermedi en el conjunt de les angiospermes. Generalment són espècies de fulles simples amb el marge enter o dentat i les seves flors solen ser bisexuals i tenir els pètals separats (tret d’algunes famílies, entre elles les ericàcies, que els tenen poc o molt soldats). Comprenen unes 25 000 espècies, tant llenyoses com herbàcies, aplegades en prop de 80 famílies. Entre les que inclouen espècies llenyoses n’hi ha unes quantes que tenen representants conspicus als estrats arboris de les selves temperades.
Les teàcies
Una família de dil·lènides que té nombrosos arbres i arbustos d’importància remarcable a les selves temperades asiàtiques, i també en algunes d’americanes, és la de les teàcies. Algunes espècies com el te (Camellia sinensis) han assolit una importància econòmica considerable. Són arbres i arbustos —també alguna liana— típicament laurifolis, de flors generalment bisexuals, grans i vistoses, cosa que ha contribuït a fer que algunes espècies, per exemple la camèlia (C. japonica), siguin molt apreciades com a plantes de jardí en climes temperats lliures de gelades. Entre altres, es poden esmentar com a espècies de teàcies de l’estrat arbori inferior de les laurisilves asiàtiques el “sakaki” (Cleyera ochnacea), arbre que al Japó es planta tancant els perímetres de molts recintes sagrats, el “hamahisakaki” (Eurya emarginata) de les laurisilves sinojaponeses, el te d’oli (Camellia sasanqua), que es troba des d’Assam fins al Japó, o les diverses espècies dels gèneres Eurya i Schima de les selves temperades d’altitud d’Assam i el SW de la Xina.
Les teàcies també estan representades en l’àmbit de les selves temperades del SE dels Estats Units per arbres com la gordònia americana o “loblolly-bay” (Gordonia lasianthus) i les camèlies americanes (Stewartia). La franklínia (Franklinia alatamaha), un altre arbre de la família de les teàcies propi d’aquesta regió, només sobreviu com a arbre de jardí, ja que fou completament exterminat en el seu medi natural, les riberes del riu Altamaha, a Geòrgia, cap al final del segle XVIII. A les laurisilves macaronèsiques és present el “mocán” (Visnea mocanera), espècie endèmica de Canàries i Madeira.
Les ericàcies i les cletràcies
Ericàcies i cletràcies són famílies estretament relacionades, fins al punt que molts autors consideren que el gènere Clethra, l’únic de la família de les cletràcies, pertany a les ericàcies. En un i altre cas es tracta de famílies d’arbustos i petits arbres, la primera quasi cosmopolita (absent només de les regions àrides d’Austràlia) i la segona restringida a les regions tropicals i subtropicals d’Àsia i Amèrica.
Algunes ericàcies que a la regió mediterrània o a l’Europa central tenen port arbustiu assoleixen port arbori als arxipèlags macaronèsics. És el cas de les diferents varietats o subespècies macaronèsiques de bruc boal (Erica arborea) i de bruc d’escombres (E. scoparia), components bàsics de les formacions de “fayal-brezal”. Altres espècies són endèmiques, com l’arboç canari (Arbutus canariensis), que és un endemisme de les Canàries occidentals, mentre que l’arbre de santa Maria (Clethra arborea) ho és de Madeira.
En molts dels boscos mixtos de pins i roures de la muntanya temperada mesoamericana, arboços (Arbutus) i boixeroles (Arctostaphylos) de port arbori, coneguts allí amb el nom genèric de “madroños”, tenen un paper destacat. Alguns, per exemple l’arboç californià (Arbutus menziesii), fins i tot arriben més al N i formen part d’algunes de les selves plujoses de coníferes del NW de l’Amèrica del Nord. També és una ericàcia la liònia arbòria (Lyonia ferruginea) del SE dels Estats Units.
Les ròsides: llorers cirers, boixos, grèvols i afins
Les ròsides apleguen un conjunt força heterogeni de famílies amb representants arboris, arbustius i herbacis. Solen tenir fulles pinnades i rarament simples, i flors cícliques amb els pètals lliures o, més rarament, soldats a la base. Formen part del grup de les ròsides famílies tan importants com les rosàcies o les lleguminoses (fabàcies o papilionàcies, cesalpinàcies i mimosàcies). Les primeres estan representades a les laurisilves macaronèsiques pel llorer cirer portuguès (Prunus lusitanica), amb dues subespècies endèmiques, una de les Açores i l’altra de les Canàries i Madeira; les segones tenen una presència nombrosa, sobretot a les selves subtropicals de la regió del Paranà, on cesalpinàcies i mimosàcies fan ostentació d’un alt nivell de diversitat d’espècies, entre les quals destaquen alguns arbres emergents del cobricel, com la “grapia” o “yvyra pere” (Apuleia leiocarpa), una cesalpinàcia, o el “timbó”, “timbaúva” o “oreja de negro” (Enterolobium contortisiliquum), una mimosàcia. Però a les selves temperades les ròsides tenen també altres representants més destacats.
Les cunoniàcies i les eucrifiàcies
També són ròsides les cunoniàcies i les eucrifiàcies, dues famílies de distribució austral que són herència de l’antiga flora de Gondwana. La de les eucrifiàcies és una família monotípica i el seu únic gènere, Eucryphia, comprèn cinc espècies d’arbres i arbustos laurifolis: dues del Xile meridional —una arbòria, l’“ulmo” (E. cordifolia), i l’altra arbustiva, el “guindo santo” (E. glutinosa)—; dues de Tasmània, E. lucida i E. millinganii; i una, E. moorei, de Nova Gal·les del Sud. Pel que fa a les cunoniàcies, estan àmpliament distribuïdes per les regions tropicals i subtropicals d’Amèrica, Àsia i Oceania, per Madagascar i altres illes de l’Índic i per l’Àfrica meridional; són uns 250 arbres i arbustos planifolis, més de la meitat dels quals pertanyen al gènere Weinmannia. El “tineo” (W. trichosperma) és una de les espècies més notables de la selva valdiviana; el “kāmahi” (W. racemosa) i el “tawhero” (W. silvicola) ho són de les selves neozelandeses, i W. rubifolia creix als boscos de faigs australs de l’extrem meridional d’Austràlia.
Les mirtàcies
La família de les mirtàcies està àmpliament representada a les laurisilves, en particular a l’hemisferi austral, on nombroses espècies arbòries d’eucaliptus (Eucalyptus) dominen en diferents selves temperades australianes. Els eucaliptus confinats als boscos oberts humits d’Austràlia estan entre els arbres més alts del món. Per exemple, al segle passat a Victòria s’inventarià un exemplar de freixe de muntanya de Tasmània (E. regnans) que feia 110 m d’alçada, el més alt trobat fins al moment, i actualment el més alt que es coneix fa 98,75 m; també s’ha amidat un eucaliptus blau de cintes o “manna gum” (E. viminalis) de 89,9 m. No obstant això, molts dels boscos on creixien aquests arbres tan alts ja no existeixen, i avui només queda un mer vestigi dels del segle passat, en els quals hi havia probablement exemplars d’una alçada encara superior. Durant els primers anys, els increments anuals en el creixement es poden produir molt ràpidament (2 m durant els primers deu anys no és infreqüent), i en el cas d’E. regnans la meitat de l’altura final de l’arbre madur es pot assolir en els primers 25 a 30 anys. En menys de cent anys un d’aquests eucaliptus pot atènyer uns nivells de biomassa increïbles.
Encara que a Nova Zelanda no hi ha eucaliptus, a les selves temperades d’aquell arxipèlag és important la presència d’espècies arbòries de mirtàcies, en particular dels “rātā” (Metrosideros), per exemple el “northern rātā” (M. robusta). A les espècies que creixen al llarg dels estuaris protegits i braços de mar i a les parts més baixes de les valls costaneres de l’extrem septentrional de l’arxipèlag hi predomina sovint el “pōhutukawa” (M. excelsa [=M. tomentosa]); i a les maresmes i selves muntanyenques de l’Illa del Sud el “southern rātā” (M. umbellata).
També hi ha mirtàcies a la selva valdiviana, on el “tepú” (Tepualia stipularis), un arbre de tronc múltiple d’uns 8 m d’alçada, forma poblaments densos en sòls mal drenats, prop de cursos d’aigua; també s’hi fan la pitra (Myrceugenia planipes) i altres congèneres, i l’“arrayán” o “palo colorado” (Lumaapiculata). Fins i tot es troben mirtàcies (Blepharocalyx longipes i Myrciaria rivularis) a les selves subtropicals de la regió del Paranà.
Les nissàcies i les proteàcies
La de les nissàcies és una família pròpia de les façanes orientals de l’Amèrica del Nord i d’Àsia, des de les latituds temperades fins a les tropicals. Hi pertanyen la davídia (Davidia involucrata), dels boscos mixtos del SW de la Xina, i els “tupelos” —el “tupelo” aquàtic (Nyssa aquatica), el “tupelo” de l’Ogeechee o llima de Geòrgia (N. ogeche) i el “tupelo” negre (N. sylvatica)—, que són arbres de fulla decídua del SE dels Estats Units.
Tant pel nombre i la varietat dels seus representants com per la seva vistositat, la de les proteàcies és una de les famílies més remarcables de l’hemisferi austral, encara que té representants al N de l’equador, a l’Amèrica central i a la Xina i el Japó. No són pas exclusives de les selves temperades, sinó que es troben també en medis encara més plujosos, com les selves tropicals, i en medis més àrids, com mediterrànies i sabanes. Són presents a les selves temperades del S de Xile, on es fan el notru o “ciruelillo” (Embothrium coccineum), d’espectacular floració vermella, el “gevuín” o “avellano” (Gevuina avellana) i diferents espècies del gènere Lomatia: l’“avellanillo” (L. dentata), el “fuinque” (L. ferruginea) i el radal (L. hirsuta). En algunes de les selves de terra baixa de l’Illa del Nord de Nova Zelanda s’hi fa una altra proteàcia, el “rewarewa” (Knightia excelsa).
Les aquifoliàcies, les celastràcies i afins
La família de les aquifoliàcies és coneguda sobretot per la vistositat de les fulles i fruits dels grèvols, distribuïts àmpliament per totes les regions tropicals i temperades del món, encara que amb una presència menor a Àfrica i a Austràlia que als altres continents. Tanmateix, el representant més important del gènere Ilex, al qual pertanyen els grèvols, és el mate (I. paraguariensis) de les selves de la regió del Paranà, no sols per la seva significació biogeogràfica sinó més particularment per l’econòmica. Els grèvols són força variats, sobretot a les regions orientals dels Estats Units, tant en l’àmbit de les boscanes decídues com en el de les selves temperades, en el qual creixen, per exemple, el grèvol americà comú (I. opaca) i també I. cassine, I. vomitoria, I. decidua o I. myrtifolia. A les illes macaronèsiques el grèvol canari o “acebiño” (I. canariensis), de les Canàries i Madeira, i el “naranjero salvaje” (I. perado [=I. platyphylla]), de Tenerife i La Gomera, es diversifiquen en diverses subespècies i varietats segons les illes.
Afí a la de les aquifoliàcies, la família de les celastràcies també abasta les regions tropicals i temperades d’arreu del món. A les selves del S de Xile és representada per la “leña dura” o “maitén negro” (Maytenus magellanica). També hi pertanyen els evònims (Euonymus) arboris o arbustius, tan freqüents a les laurisilves himalaianes i sinojaponeses —per exemple el “masaki” (E. japonica) i el “mayumi” (E. sieboldiana)—. També mostra afinitat amb aquifoliàcies i celastràcies la família de les buxàcies, a la qual pertanyen el boixos (Buxus); a l’estrat arbori inferior de les selves euxíniques, per exemple, sobre substrat calcari, hi abunden els poblaments de boix còlquic (B. colchica), un arbrissó perennifoli molt semblant al boix comú (B. sempervirens). A les selves hircanianes s’hi fa una espècie vicària de tots dos, B. hyrcana.
Les corinocarpàcies, una petita família d’arbres i arbustos d’Austràlia, Nova Zelanda i altres illes del Pacífic meridional, també es relacionen amb aquest grup. A Nova Zelanda, les representa principalment el “karaka” (Corynocarpus laevigata) de les selves costaneres de les regions septentrionals i d’algunes illes properes com l’illa Chatham i l’illa Pitt, un arbre que els maoris han contribuït històricament a propagar i que sempre han apreciat força pels seus fruits (encara que, pròpiament, no es pot considerar que hagi estat mai cultivat).
Les meliàcies, les rutàcies i afins
La família de les meliàcies té una distribució principalment tropical i subtropical i inclou un gran nombre d’arbres de fustes precioses, en particular les caobes americanes (Swietenia) i africanes (Khaya, Entandrofragma). A l’àmbit de les selves temperades, les meliàcies prenen singular importància en l’anomenada ‘selva de meliàcies’ de les laurisilves de la regió del Paranà, amb l’estrat arbori superior dominat per l’“ygary” o “cedro” (Cedrela fissilis) i l’inferior pel “cedro ra” o “canjerana” (Cabralea canjerana), l‘“yrupe rupa” o “cancharana macho” (Guarea kunthiana) i diferents espècies de Trichilia, totes elles meliàcies. El “kohekohe” (Dysoxylum spectabile), que també és una meliàcia, és un dels arbres més característics de les selves de terra baixa de l’Illa del Nord de Nova Zelanda.
La mateixa distribució presenta, a grans trets, la família de les rutàcies, a la qual pertanyen tots els cítrics, que tenen un paper particularment important a les laurisilves sinojaponeses. Les rutàcies, però, també són presents, per exemple, a les selves de la regió del Paranà i d’Austràlia. Afins a meliàcies i rutàcies són algunes famílies de caducifolis —aceràcies i hipocastanàcies— de distribució principalment boreal, que es troben discretament representades a les selves temperades d’Euràsia i l’Amèrica del Nord, generalment per espècies que també són presents a les boscanes decídues veïnes. El mateix passa amb la família de les juglandàcies, àmpliament difosa a les boscanes decídues, sobretot de l’Àsia oriental i de l’Amèrica del Nord, però que també es difon cap als tròpics asiàtics i americans. Als boscos mixtos de planifolis del baix Mississipí i del SE dels Estats Units són freqüents diferents espècies de cària (Carya), com la cària vera (C. glabra), la cària tomentosa (C. tomentosa) o la cària aquàtica (C. aquatica). A l’Himàlaia oriental, d’altra banda, s’hi fa Engelhardia [=Engelhardtia] spicata, de la qual s’aprofita l’escorça pels tanins que conté.
Les astèrides: freixes, catalpes i viburns
Les astèrides apleguen les famílies d’angiospermes dicotiledònies més evolucionades i, encara que en menor nombre que els representants d’altres grups d’angiospermes més arcaics, tampoc no falten a les selves temperades.
Les oleàcies, les boraginàcies i les verbenàcies
La família de les oleàcies, a la qual pertanyen oliveres i freixes, àmpliament distribuïda per les regions temperades i tropicals de tot el món, té també representants a les selves temperades. A les terres baixes humides, però no inundades, del SE dels Estats Units, del SE de Virgínia, fins a Louisiana i l’E de Texas, s’hi fa el freixe de Carolina (Fraxinus caroliniana), un arbret caducifoli que conviu a la seva àrea de distribució amb altres freixes americans d’àrea més extensa. Les troanes (Ligustrum japonicum i L. lucidum) són els representants més conspicus de la família a les laurisilves de l’Àsia oriental, mentre que als arxipèlags macaronèsics, les oleàcies estan representades pel gènere Picconia.
La família de les boraginàcies no té únicament representants herbacis, com sol passar a la conca mediterrània, centre mundial de la seva distribució, sinó que també en té de llenyosos i fins i tot de port arbori. Als estrats arboris inferiors de les selves de la regió del Paranà, per exemple, hi té un paper gens menystenible el “guajayvi” o “guayaibí” (Patagonula americana). La família de les verbenàcies, predominantment tropical i subtropical però amb alguns representants —gairebé tots herbacis— a les regions temperades, és present a les selves temperades de l’extrem austral d’Amèrica amb espècies com l“arrayán macho” o “espino blanco” (Rhaphithamnus spinosus) de les selves valdivianes. El “pūriri” (Vitex lucens), al seu torn, és un arbre comú als boscos amb predomini de coníferes de la meitat septentrional de l’Illa del Nord de Nova Zelanda.
Les bignoniàcies, les caprifoliàcies i les asteràcies
Les bignoniàcies són una família principalment tropical, centrada sobretot a l’Amèrica meridional, on té nombrosos representants arboris i arbustius, però també s’estén per les selves temperades americanes, asiàtiques i australianes. A les selves de la regió del Paranà, les bignoniàcies són particularment abundants, mentre que a les del SE dels Estats Units, sobretot al S d’Alabama i també a les regions adjacents de Florida, Geòrgia i Mississipí, hi creix la catalpa americana meridional o “catawba” (Catalpa bignonioides), extensament difosa com a arbre ornamental per totes les regions temperades. A les regions orientals de la Xina es donen altres espècies de catalpa.
Les caprifoliàcies són conegudes principalment com a lianes de les regions temperades i subtropicals, però també n’hi ha que són petits arbres. És el cas dels saücs (Sambucus) —el saüc canari (S. palmensis), per exemple, un rar endemisme limitat a comptades localitats de La Palma, La Gomera i Tenerife— i d’alguns viburns (Viburnum) —com el “follao” (V. rugosum) canari—. El viburn de Walter o viburn de fulla petita (V. obovatum) és un petit arbre perennifoli propi dels sòls estacionalment inundats del SE dels Estats Units, des de Carolina del Sud fins a Florida. L’“arrow-wood” (V. dentatum) també és propi de la mateixa àrea, en aquest cas dels substrats sorrencs no tan humits, però amb menys freqüència assoleix el port arbori.
També de la família de les asteràcies o compostes es troben representants arboris a les selves temperades. A Tasmània, per exemple, diferents asteràcies llenyoses dels gèneres Olearia i Brachyglottis, la majoria de port arborescent però algunes de port arbori, es fan més visibles a mesura que augmenta l’altitud; l’arbre del mesc (Olearia argophylla) arriba fins i tot a fer-se localment dominant en algunes àrees. En un altre continent, a les pinedes de “pehuén” (Araucaria araucana) de més altitud dels Andes meridionals, s’hi pot trobar el “palo santo” o “palo blanco” (Dasyphyllum diacanthoides).
Les plantes dels estrats inferiors del sotabosc i els epífits
A les selves temperades, per sota de l’estrat arbori inferior se sol trobar un estrat arbustiu més esponerós que el de les boscanes decídues, encara que no tan com el de les selves tropicals o, fins i tot, el de les mediterrànies. Els seus elements més característics solen ser arbrissons i arbustos de fulla perenne, bé que no falten tampoc, en les de latituds o altituds més extremes, els caducifolis. En condicions més càlides i humides també es presenten abundants falgueres arborescents i algunes palmes. Les lianes abunden arreu, però el sotabosc herbaci, en canvi, sol ser força pobre. No passa el mateix amb el brancatge, les enforcadures i la superfície dels mateixos arbres que, en el ric ecosistema multiestratificat de les selves temperades, esdevenen un dels hàbitats més interessants.
Els arbustos del sotabosc
La majoria dels arbustos del sotabosc de les selves temperades pertanyen a les mateixes famílies que els arbres. En alguns casos, fins i tot les mateixes espècies tenen formes de creixement arbòries o arbustives segons les circumstàncies del lloc en el qual creixen, o en funció de les pertorbacions que han sofert. Com els arbres, els arbustos presenten fulles planifòlies o aciculifòlies i, si bé els perennifolis són majoria, no falta algun caducifoli. Entre els arbustos planifolis, tal com passa entre els arbres, es poden reconèixer els laurifolis pròpiament dits, de fulla més gran i semblant a la dels llorers, i els mirtifolis, de fulla més petita i semblant a la de la murta.
Els casos més remarcables de sotabosc arbustiu planifoli de fulla perenne potser són els de les selves euxíniques i hircanianes, que conviuen amb cobricels dominats per espècies caducifòlies: boixos (Buxus colchica, B. hyrcana) a les àrees calcàries, i arços (Crataegus), galzerans (Ruscus), estafilees (Staphylea), nabius (Vaccinium) i altres arbustos a la resta. Els rododendres (Rhododendron ponticum, R. ungernii), presents a les selves euxíniques, estan absents de les hircanianes, bé que reapareixen en tota la seva esplendor a l’Himàlaia, on n’hi ha una diversitat d’espècies considerable que, a més, augmenta en direcció E.
A les laurisilves himalaianes, a més dels rodondendres, es troben també altres espècies d’ericàcies planifòlies (Lyonia, Gaultheria) i una colla d’espècies d’afinitats sinojaponeses de diferents gèneres de teàcies (Eurya, Camellia, Schima), simplocàcies (Symplocos), mirsinàcies (Embelia) i altres famílies. Les ericàcies tenen també un paper remarcable en el sotabosc arbustiu de les selves temperades de l’Amèrica del Nord, per exemple el “stagger-bush” (Lyonia mariana) o el “dwarf huckleberry” (Gaylussacia dumosa) que, al SE dels Estats Units formen un estrat arbustiu esclarissat sota algunes pinedes recorregudes regularment pel foc, o el “red huckleberry” (Vaccinium parvifolium), la “mock azalea” (Menziesia ferruginea), el salal (Gaultheria shallon) o el rododendre americà de fulla gran (Rhododendron macrophyllum), que es troben als densos sotaboscos de les selves plujoses de la costa NW.
Ericàcies i cletràcies de fulla acicular a l’hemisferi boreal i epacridàcies a l’austral donen una altra forma arbustiva característica, l’estereotip de la qual serien els brucs (Erica). Generalment ocupen els sòls més pobres o més drenats i de vegades formen bosquines sense cobert arbori (landes) en àrees que han estat molt pertorbades o que no reuneixen les condicions adients per al creixement de formes arbòries. D’arbustos d’aquest tipus, n’apareixen en moltes de les regions de selves temperades, en particular en àrees de condicions extremes —el cas de la cletra de marjal o “coast white-alder” (Clethra alnifolia) d’alguns marjals del SE dels Estats Units, o de la majoria d’epacridàcies de Nova Zelanda— o pertorbades per l’acció humana o per altres causes —el cas de les landes de bruc boal (Erica arborea) de les Canàries—.
Les palmeres i les falgueres arborescents
Les palmeres i les falgueres arborescents tenen en comú una semblança superficial, compartida amb les ciques, que es deu al ventall de fulles que corona els seus ‘troncs’ —que, en no tenir creixement secundari, no són veritables troncs—. En realitat, però, es tracta de dos grups de plantes molt allunyats. Mentre que les falgueres arborescents són pteridòfits i no tenen flors ni fruits, les palmeres són angiospermes monocotiledònies que solen posseir grans inflorescències i produeixen fruits generosos.
Encara que algunes arriben a superar els 15 m, la majoria de les palmeres de les selves temperades tenen port arbustiu, fins i tot per la seva ramificació simpodial. La seva presència és particularment important a les selves temperades de latituds subtropicals, com les de la regió del Paranà o les australianes, però també a les macaronèsiques o les del SE dels Estats Units. A les selves de la regió del Paranà, en boscos madurs, les palmeres solen acompanyar les lauràcies, encara que només esdevenen dominants quan les condicions són adverses per a aquestes; el “pindo” o “geriba”, també anomenat impròpiament “coco” (Syagrusromanzoffiana), és la palmera més característica, i es dóna freqüentment a les zones fredes, mentre que l’“ysyi” o “palmito” (Euterpe edulis) és una espècie localment acompanyant que, en alguns indrets, com per exemple a la selva missionera d’Iguaçú, pot formar extensos “palmitales”. A l’àrea macaronèsica només s’hi fa la palmera canària (Phoenix canariensis), endèmica de les illes Canàries i, de fet, pròpia dels vessants de sotavent de les illes, on no arriba la influència de les boires que possibiliten l’existència de les laurisilves. Al sotabosc dels boscos mixtos de planifolis del SE dels Estats Units s’hi fan palmes com el “saw palmetto” (Serenoa repens), el “blue-stem palmetto” (Sabal minor) o Rhapidophyllum hystrix, més aviat de port arbustiu que no arbori. Les palmeres de Nova Zelanda i illes adjacents (Rhopalostylis) són les més australs del planeta.
Les falgueres arborescents tenen un port semblant al de les palmeres, i són particularment abundants a les selves temperades australs, per exemple a la selva missionera, on es coneixen amb el nom collectiu de “chachís”; espècies com Alsophila procera, A. [=Cyathea] atrovirens, Dicksonia sellowiana i Cyathea [=Hemitelia] setosa, totes elles de la família de les ciateàcies, formen part important del sotabosc en els indrets més humits. També a Austràlia i a Nova Zelanda apareixen falgueres arborescents, concretament dels gèneres Dicksonia (D. antarctica a Tasmània o D. squarrosa a Nova Zelanda) i Cyathea.
Les lianes
També les plantes enfiladisses són molt comunes a les selves temperades. A les selves euxíniques, per exemple, n’hi ha una desena d’espècies, encara que no formen bardisses, ni tan sols a les vorades del bosc. En són especialment característiques l’heura còlquica (Hedera colchica) i la vinya de seda (Periploca graeca), l’arítjol còlquic (Smilax excelsa) i la vidalba (Clematis vitalba). A les laurisilves macaronèsiques les lianes són menys comunes, però hi predominen l’heura canària (Hedera helix canariensis), que és molt abundant, la “gibalbera” o “alicacán” (Semele androgyna) i l’arítjol canari o “zarzaparrilla sin espinas” (Smilax canariensis), Rubus bollei, que és un esbarzer endèmic, el gatmaimó canari (Tamus edulis) i el “corregüelon” o “corrigüela de los montes” (Convolvulus canariensis).
Entre les espècies de lianes més freqüents a la selva missionera es troben la dioscoreàcia Dioscorea bulbifera, la vitàcia Cissus gongylodes, diferents apocinàcies del gènere Forsteronia, i diverses espècies de bignoniàcies, euforbiàcies, cactàcies, dil·leniàcies, rubiàcies, sapindàcies, etc. A la part més septentrional de la selva valdiviana, les lianes, com els epífits, són també força abundants. Hi destaquen la vitàcia Cissus striata i la gesneriàcia Mitraria coccinea. Més al S, als boscos mixtos de “coihue” d’altituds mitjanes, hi ha la hidrangeàcia Hydrangea integerrima, que pot assolir 40 m de longitud, i l’esmilacàcia Luzuriaga radicans.
Les lianes i altres plantes enfiladisses estan ben representades als boscos planifolis mixtos meridionals del SE dels Estats Units. S’hi compten la vinya muscadina salvatge (Vitis rotundifolia), la vinya verge (Parthenocissus quinquefolia), la “cross-vine” (Bignonia capreolata), la “poison-ivy” (Rhus [=Toxicodendron] radicans) i el gessamí de Carolina (Gelsemium sempervirens).
Els epífits
Entre els organismes epífits més característics de les selves temperades figuren molts líquens, ja que les condicions càlides i humides del bioma són ideals perquè hi creixin. Als boscos més vells, sembla que pengin de cada branca i poden arribar a amagar el tronc. Els líquens, en particular, tenen un paper essencial a la vida dels boscos plujosos de coníferes; efectivament, en un ecosistema complex, on cada membre interactua amb molts d’altres, els líquens són els cooperatius per excel·lència, ja que ells mateixos estan constituïts per espècies de fongs i algues (o cianobacteris) que viuen en simbiosi. Les algues i els cianobacteris, gràcies a la seva capacitat per a portar a terme la fotosíntesi, proporcionen energia al seu soci, i alhora els fongs aporten nutrients i donen protecció a l’alga o el cianobacteri.
Hi ha líquens de totes formes, colors i mides. Molts són foliacis i formen crostes sobre els troncs dels arbres i les roques, mentre que altres són filamentosos o fruticulosos i pengen de les branques com un guarniment. La majoria creixen molt a poc a poc i, per la seva mateixa naturalesa, són molt abundants en boscos primaris madurs. Hi ha moltes espècies de líquens extremadament sensibles a la contaminació atmosfèrica, tant si es tracta de gasos com de pluja àcida, i per això desapareixen molt de pressa a les regions industrials.
Algunes espècies poden arribar a dominar gairebé totalment alguns microhàbitats de les selves temperades. Així, a les selves plujoses de coníferes de la costa nord-occidental de l’Amèrica del Nord, el liquen foliaci Lobaria oregana assoleix densitats de gairebé 125 000 kg/ha i, tot i que als boscos densos és pràcticament invisible de terra estant, cobreix les capçades dels arbres més alts, on pot arribar a amagar les acícules, i algunes vegades la seva biomassa es pot acostar a una cinquena part de la del fullatge del mateix arbre. Lobaria oregana té una estructura encara més complexa que la de la majoria de líquens; està constituït bàsicament per un fong i una alga verda però, a més, inclou un tercer organisme, un cianobacteri del gènere Nostoc, que té la capacitat de fixar nitrogen atmosfèric. Aquest liquen és tan eficaç que sovint presenta taxes de fixació de nitrogen de l’ordre de 5 a 10 kg/ha, tret que el converteix en una font essencial de nutrients per a l’ecosistema. Els líquens tenen un paper important en la cadena alimentària perquè són consumits per un ampli ventall d’invertebrats i vertebrats, fins i tot per grans herbívors. En l’àmbit del bioma són especialment importants en indrets on fa prou fred perquè a l’hivern hi nevi, com ara al NW de l’Amèrica del Nord i a Xile. En aquestes regions, la caiguda de líquens morts a conseqüència de ventades fortes proporciona una provisió alimentària essencial quan la neu ha cobert el sòl. S’ha calculat que, a la costa nord-occidental de l’Amèrica del Nord, la quantitat de fragments caiguts de líquens de les espècies més comunes, com ara diferents espècies d’Alectoria, pot superar en algunes vegades la producció de fulles.
Moltes falgueres petites i mitjanes es comporten com a epífits a les selves temperades i creixen entremig dels tous de molses i líquens de troncs i branques. Ocasionalment, aquest és el cas de diferents espècies d’Asplenium, Trichomanes, Polypodium, Davallia, Diplazium o Hymenophyllum de les laurisilves macaronèsiques, o de Polypodiumserratum o Dryopteris alexeenkoana a les selves euxíniques. Als boscos de pins i roures de la muntanya mexicana, les branques d’aquests darrers constitueixen un ambient particularment propici per al desenvolupament de plantes epífites, entre les quals també abunden les falgueres, però des del paral·lel 21°N cap al S, fan incursions en aquest nínxol nombroses plantes d’origen neotropical, sobretot orquídies i bromelàcies, i el mateix succeeix amb les selves temperades de l’Amèrica del Sud.
Les orquídies i les bromeliàcies figuren entre les famílies d’epífits més característiques, la primera pràcticament cosmopolita a totes les regions càlides i temperades del món, la segona restringida a l’àmbit neotropical, amb algunes penetracions N enllà, a través de l’istme mesoamericà. Així, entre els epífits que creixen entre els tous de molses i líquens que cobreixen les branques dels boscos d’araucària i les laurisilves de la regió del Paranà hi figuren orquídies (Campylocentrum, Oncidium) i bromeliàcies, com Aechmea recurvata i diferents clavellines d’aire (Tillandsia), i també la piperàcia Peperomia tetraphylla. L’anomenada molsa espanyola o “Spanish-moss” (Tillandsia usneoides) és un epífit comú als boscos planifolis mixtos meridionals del SE dels Estats Units i és omnipresent als aiguamolls poblats de taxodis, els quals pot arribar a cobrir gairebé totalment.
Les bromeliàcies presenten la particularitat de poder absorbir vapor d’aigua de l’atmosfera gràcies a les parets higroscòpiques de les seves cèl·lules i als tricomes morts que recobreixen completament aquestes plantes. Així, quan la humitat relativa augmenta, en començar la nit, sovint s’observa un màxim d’absorció. A més a més de les característiques anatòmiques i morfològiques per a poder fer front a la manca d’aigua, a moltes espècies d’epífits, particularment de les famílies de les bromeliàcies, les orquidàcies i totes les espècies epífites de les cactàcies, la via metabòlica de la fotosíntesi és l’anomenada CAM, i no la C3, cosa que els permet augmentar l’eficàcia en l’ús del diòxid de carboni i de l’aigua.
La vegetació de les selves temperades del Vell Món
Des dels arxipèlags atlàntics de la Macaronèsia fins al centre del Japó, amb importants discontinuïtats marcades per les extenses regions de la conca mediterrània, el Sàhara i l’Àsia occidental sotmeses a períodes de sequera, diverses àrees del Vell Món conserven testimonis de la flora arctoterciària en les seves laurisilves i boscos mixtos. Cal situar l’origen d’aquestes selves temperades en els ambients miopliocènics de la conca mediterrània, diferents jaciments fossilífers de la qual testimonien l’existència de selves tropicals o subtropicals de característiques florístiques semblants, però amb una diversitat més elevada que la de les selves temperades que actualment ocupen la Macaronèsia.
Les laurisilves i els boscos de coníferes macaronèsics
La gran dispersió territorial dels arxipèlags macaronèsics i la desigual distància entre ells i de cada un respecte als continents veïns expliquen les considerables diferències entre unes illes i altres. Així, mentre és considerable l’afinitat florística i faunística entre les Canàries i Madeira, que comparteixen unes quantes espècies de fanerògames arbòries, arbustives i herbàcies, un nombre elevat de criptògames i de fongs i bona part de la fauna, a les Açores abunden els endemismes vicaris o exclusius i la flora està més relacionada amb la de l’Europa atlàntica. Són molt rares les espècies endèmiques compartides per tots tres arxipèlags macaronèsics; pràcticament només es poden citar com a endemismes comuns el llorer Laurus azorica [=L. canariensis], amb varietats diferents en cada arxipèlag, i el ranuncle Ranunculus cortusifolius.
El component arbori és significatiu, encara que no arriben a la trentena les espècies arbòries autòctones. De la combinació d’aquestes espècies, en resulten uns estrats arboris de composició florística molt variable, amb alçades que, segons la localitat i l’exposició, poden variar entre els 5 i els 20 m i poden permetre o no l’existència d’una selva pluriestratificada. La dominància d’unes o altres espècies fa possible la diferenciació de les comunitats, que al seu torn estan condicionades pel clima: laurisilves típiques amb dominància de lauràcies als vessants exposats als alisis —entre els 600 i els 1 200 o 1 400 m—; brugueres (“fayal-brezal”) caracteritzatdes per la dominància de la “faya” (Myrica faya) i el bruc boal (Erica arborea) en vessants no tan ben orientats, en cotes altes o com a formacions secundàries en llocs on ha estat destruïda la laurisilva; pinedes de pi canari (Pinus canariensis) per damunt de la laurisilva; el “fayal-brezal” fins als 2 000 m aproximadament, etc.
És igualment notable la quantitat d’arbustos que componen la vegetació climàcica o la dels espais poc pertorbats, alguns d’ells pertanyents a gèneres endèmics, com la lamiàcia Bystropogon canariensis, la urticàcia Gesnouinia arborea, les escrofulariàcies del gènere Isoplexis, les campanulàcies del gènere Musschia, i les rosàcies dels gèneres Bencomia o Argyranthemum. En el component herbaci destaca la gran quantitat de pteridòfits, molts dels quals es poden comportar com a epífits, i la importància del seu recobriment i la seva biomassa.
Les selves euxíniques i hircanianes
Una climatologia excepcional que ha experimentat escassos canvis des del Terciari explica la pervivència del que resta d’aquestes selves, filles dels vents frescals i humits procedents respectivament de la mar Negra i de la Càspia. A diferència d’altres laurisilves, no és pas l’estrat arbori, integrat majoritàriament per caducifolis, sinó el complex del sotabosc i en particular el dens estrat arbustiu amb boix (Buxus colchica, B. hyrcana) i altres arbustos perennifolis el que caracteritza aquestes selves temperades. Es pot afirmar sense exagerar que als pendents de les gorges i les valls de la Còlquida i regions costaneres adjacents es concentren gairebé tots els representants més característics de l’antiga flora forestal mesòfila. Una cinquantena d’espècies d’arbres i arbustos i una vuitantena de plantes herbàcies, entre les quals moltes falgueres, apareixen en les més característiques d’aquestes selves, encara que la composició global de la flora és, naturalment, encara més rica.
Els cinc arbres fonamentals que configuren aquests boscos són el castanyer comú (Castanea sativa), el roure de Hartwiss (Quercus hartwissiana), el faig oriental (Fagus orientalis), el carpinus del Caucas (Carpinus caucasica) i el vern barbat (Alnus barbata). Les tres primeres espècies constitueixen els estrats arboris de les selves primàries, mentre que el carpinus i, especialment, el vern són espècies secundàries que apareixen quan s’ha produït alguna pertorbació. Totes les espècies esmentades es troben en composicions variades i sovint barrejades amb altres espècies arbòries, com ara el tell del Caucas (Tilia caucasica), el plàtan fals (Acer pseudoplatanus), l’erable (A. platanoides), els oms Ulmus eliptica, i U. scabra i, en alguns casos poc freqüents, el teix europeu (Taxus baccata) i altres gimnospermes.
Els elements més característics del sotabosc, com ja s’ha dit, són els arbrissons i arbustos de fulla perenne. A les regions calcàries, a l’estrat arbori inferior, abunden els poblaments de boix còlquic (Buxus colchica), encara que, en conjunt, aquesta espècie ha esdevingut escassa als boscos de la regió de la Còlquida a causa de l’explotació de què ha estat objecte. En moltes gorges d’aquesta zona, especialment a Abkhàzia, Geòrgia i Adjària, el sotabosc de les selves euxíniques es caracteritza per l’esponerós desenvolupament de petits arbres perennifolis, com el llorer cirer (Prunus laurocerasus) i el grèvol europeu (Ilex aquifolium), i d’arbustos com els rododendres (Rhododendron ponticum, R. ungernii). Són també molt freqüents les mates de galzerans (Ruscus), que formen els estrats perennifolis més baixos del sotabosc. Les selves euxíniques es caracteritzen també per la presència d’un cert nombre d’arbustos caducifolis, com el rododendre groc (Rhododendron flavum), les estafilees (Staphylea colchica, S. pinnata), la xeringuilla del Caucas (Philadelphus caucasicus), el nabiu caucàsic (Vaccinium arctostaphylos), el saüc europeu comú (Sambucus nigra) i molts d’altres.
Un cert nombre de lianes, especialment l’heura còlquica (Hedera colchica), la vinya de seda (Periploca graeca), l’arítjol còlquic (Smilax excelsa) i la vidalba (Clematis vitalba), són força característiques de les selves euxíniques, i les falgueres també hi són presents. D’entre totes les espècies de lianes esmentades, només Hedera colchica es troba en abundància, no solament enfilant-se pels troncs dels grans arbres, sinó també estenent-se pel brancam i formant un cobricel compacte; les seves rabasses arriben a assolir diàmetres d’uns 20 cm.
A la regió hircaniana, per sota de l’estatge de les fagedes, s’estén el de les selves hircanianes, molt semblant al de les euxíniques. L’arbre dominant a l’estrat arbori superior sol ser el roure de fulla de castanyer (Quercus castanaefolia), amb un estrat inferior dominat pel boix Buxus hyrcana o, localment, pel carpinus del Caucas (Carpinus caucasica) o el fals om hircanià (Zelkova hyrcana); també hi ha àrees, però, en les quals l’arbre dominant és l’hamamelidàcia endèmica Parrotia persica, la parròtia persa. Allà on el nivell freàtic s’acosta a la superfície apareixen vernedes amb diferents espècies de vern (Alnus barbata, A. subcordata). Altres espècies arbòries notables són Diospyros lotus, la falsa acàcia càspica (Gleditsia caspia) o l’acàcia taperera o arbre de seda (Albizia julibrissin), mentre que al sotabosc es troben espècies arbustives com el grèvol hircanià (Ilex hyrcana), el galzeran hircanià (Ruscus hyrcanus), el llorer alexandrí (Danae racemosa), o arços com Crataegus lagenaria i C. kyrtostila, i lianes com l’arítjol còlquic (Smilax excelsa), la vinya de seda (Periploca graeca) o l’heura Hedera pastuchovii.
Els boscos perennifolis himalaians i sinojaponesos
Els vessants meridionals de l’Himàlaia constitueixen una zona de transició entre els boscos secs de l’W i els humits de l’E, i també entre diferents formes fisiognòmiques arbòries, des dels arbres perennifolis tropicals i subtropicals d’altituds inferiors als 2 500-3 000 m fins a les coníferes i els caducifolis temperats d’altituds més elevades. Aquesta característica de la vegetació montana de l’Himàlaia contrasta tant amb la de les muntanyes tropicals, on els perennifolis prevalen monòtonament a qualsevol altitud, com amb la de les muntanyes temperades, en les quals des de les terres baixes fins al límit superior del bosc es van succeint boscos de tres fisiognomies diferents: perennifolis de fulla ampla, caducifolis i coníferes.
La regió nord-occidental de l’Himàlaia és més aviat eixuta i presenta una vegetació de tipus subdesèrtic, amb més afinitats amb la de l’Àsia central, pertanyent a la regió florística iranoturaniana, que amb la de la regió sinojaponesa o la dels tròpics humits de la regió indomalaia. La vegetació arbòria es troba limitada al cinturó nebulós de les altituds mitjanes i a les àrees humides, ja sigui per l’existència de precipitacions locals o pel fet d’estar prop de cursos d’aigua permanents. Els boscos més freqüents al cinturó nebulós de les àrees montanes són els de coníferes, amb pi blau indi (Pinus wallichiana), pi de tres fulles de l’Himàlaia (P. roxburghii), pícea de l’Himàlaia (Picea smithiana), avet de l’Himàlaia (Abies spectabilis) i cedre de l’Himàlaia (Cedrus deodara). Per sota seu apareixen boscos esclerofil·les amb “kharsu” (Quercus semecarpifolia) i altres alzines (Q. baloot, Q. leucotrichophora), més semblants a les mediterrànies que a les selves temperades humides més orientals. Als vessants obacs i al fons de les valls es desenvolupen boscos caducifolis amb castanyer d’Índia de l’Himàlaia (Aesculus indica).
En canvi, les selves humides temperades de l’E de l’Himàlaia s’assemblen molt als boscos nebulosos de les altes muntanyes equatorials del SE d’Àsia. El cobricel d’un i altre tipus de bosc està dominat per arbres de les famílies de les fagàcies (Castanopsis, Lithocarpus i Quercus) i de les lauràcies (Cinnamomum, Neolitsea, Litsea i Persea) —raó per la qual són sovint anomenats ‘boscos laurofagacis’—, i molts dels arbres acompanyants, com per exemple les hamamelidàcies Altingia excelsa i Exbucklandia populnea, la teàcia Schima wallichii o la juglandàcia Engelhardia [=Engelhardtia] spicata, són comuns a tots dos.
Al NE de la serralada himalaiana, cap a les províncies sud-occidentals i meridionals de la Xina, el nombre d’espècies acompanyants d’aquestes selves va decreixent i la seva composició florística s’empobreix mentre davalla en altitud fins a assolir els seus límits septentrionals, al curs baix del riu Iang-Tsé o a les costes centrals del Japó. L’estructura forestal, però, no canvia ni amb l’altitud ni amb la latitud. El tancat dosser arbori es troba a una alçada de 20 o 30 m i no hi ha estrat arbustiu o està molt poc desenvolupat —excepte al fons de les valls o a les vorades i clarianes, on la quantitat de radiació que travessa el dosser és més important—. Els arbres i arbustos típics de l’estrat arbustiu també són espècies perennifòlies pertanyents a les famílies de les teàcies, simplocàcies, mirsinàcies, rubiàcies i encara altres, i l’estrat herbaci està format per diverses falgueres perennes, com Dryopteris, Polystichum, Diplazium o Plagiogyria, i herbes higròfiles, com les urticàcies dels gèneres Elatostema i Pilea, les poàcies del gènere Oplismenus o diferents gingiberàcies.
Al seu límit superior, les selves perennifòlies dels vessants meridionals de l’Himàlaia deixen pas a boscos temperats de coníferes dominats per pícees (Picea), tsugues i hemlocks (Tsuga), avets (Abies) i pins (Pinus). Alguns grans exemplars d’aquests arbres, de 30 o 40 m d’alçada, sobresurten emergents per sobre el cobricel de la selva perennifòlia, mentre que en algunes de les regions més humides d’aquests vessants meridionals de la serralada els perennifolis del dosser es barregen amb espècies caducifòlies, com alguns aurons (Acer) i diferents araliàcies. La presència d’algunes espècies de caducifolis relictes, com el tetracèntron xinès (Tetracentron sinense) o Euptelea polyandra, únics representants de dues famílies monotípiques amb caràcters molt primitius, és particularment significativa i indica que algunes de les espècies de la rica flora terciària relicta del SW de la Xina arribaren a penetrar a la regió oriental de l’Himàlaia. En anar d’E a W, la presència de les espècies mèsiques es fa cada vegada menys evident, fet que s’exemplifica clarament en el cas dels rododendres (Rhododendron); les 84 espècies presents a Sikkim i Bhutan es redueixen a una trentena al Nepal oriental i a 15 al voltant de la vall de Dudh Kosi, al Nepal central; el nombre segueix decreixent fins a 11 a les rodalies de Katmandu i, més cap a l’W, a Kumaon i al Caixmir, tan sols creixen quatre espècies de rododendres (R. anthopogon, R. arboreum, R. campanulatum i R. lepidotum). En sentit oposat, els boscos de coníferes típics de l’W de l’Himàlaia es prolonguen, en una distribució disjunta, ben enllà de la regió humida oriental. Així, les pinedes de pi de tres fulles de l’Himàlaia (Pinus roxburghii) s’estenen fins al SW de la Xina, encara que a la regió oriental humida de la serralada ocupen només les valls seques o altres indrets de condicions localment més eixutes, com per exemple els solells rocallosos.
Estatges de vegetació a les muntanyes de l’E i el SE asiàtic
Idem a partir d’Ohsawa, 1990 i 1993
Perfil vertical dels estatges de vegetació de les muntanyes de l’E i el SE d’Àsia. Als ambients tropicals i subtropicals d’aquestes muntanyes es desenvolupen selves plujoses i boscos monsònics de dipterocarpàcies i de planifolis dominats per fagàcies perennifòlies. Als ambients tropicals, les fagàcies dominants solen ser espècies del gènere Castanopsis, mentre que als ambients subtropicals són substituïdes per Quercus i Lithocarpus, que sovint apareixen barrejades amb diverses lauràcies (Persea i Litsea, entre altres) i algunes teàcies (per exemple Schima) formant laurisilves. Els boscos de Castanopsis ocupen extensions més grans que no pas els de dipterocarpàcies, ja que s’estenen fins als 37° de latitud N a les costes del Japó. En canvi, les dipterocarpàcies assoleixen el seu límit més septentrional a Yunnan, al SW de la Xina, on avancen fins als 20°N. Per sobre dels 20-30°N, als ambients temperats típics, creixen laurisilves a les regions més càlides i boscanes decídues a les més fresques, mentre que als ambients temperats freds de les regions circumboreals i de l’alta muntanya s’hi fan boscos de coníferes perennifòlies. Les formacions caducifòlies estan constituïdes pels parents de fulla decídua de les espècies que es troben a les regions subtropicals, és a dir, per fagàcies, lauràcies i teàcies, que van sovint acompanyades d’arbres d’altres famílies endèmics d’aquestes muntanyes, com Tetracentron, Cercidiphyllum, Euptelea, i de les coníferes també endèmiques Cathaya, Pseudolarix i Cryptomeria. Als ambients temperats freds els arbres dominants són pícees (Picea), avets (Abies) i pins (Pinus).
Un tret característic de les selves temperades sinojaponeses és la gran extensió que hi han anat ocupant els boscos secundaris de bambú, en bona part d’origen antròpic. Hom ha plantat bambús per recollir-ne els rebrots tendres, que són comestibles, o bé per aprofitar-los en qualsevol de les seves múltiples aplicacions. A les regions de clima temperat i subtropical de la Xina han estat descrits dotze tipus de bosc de bambú, i al Japó, en canvi, solament se n’han trobat quatre. Les espècies de bambú, com Phyllostachys heterocycla i P. bambusoides, eviten els esllavissaments en estendre llurs rizomes pel sòl, i per això al Japó es planten principalment als vessants que poden amenaçar els habitatges que tenen al peu. Les dues espècies esmentades de bambú, que tenen una alçada de 10 a 15 m, són seriosos competidors de les selves perennifòlies humides, ja que poden envair els boscos joves estenent llurs rizomes des de l’exterior; tanmanteix, atès que es tracta de plantes monocàrpiques, mostren una persistència limitada, i després d’haver produït nombroses canyes al llarg d’unes quantes dècades, tots els seus rizomes moren simultàniament. Les del gènere Pleioblastus envaeixen ràpidament els terrenys abandonats; en algunes de les illes Ryūkyū (o Nansei-shotō) formen poblaments purs en què difícilment hi pot créixer cap arbre, i amb canyes que arriben a fer 5 m d’altura i que resulten impenetrables per la seva extrema densitat.
La vegetació de les selves temperades del continent americà
Tot i que, sobretot a l’Amèrica del Nord, no falten estudis locals o regionals sobre moltes de les àrees americanes del bioma, la seva fragmentació, les diferències en l’estat de conservació en què es troben les diferents zones i la diversitat de mètodes utilitzats en els estudis duts a terme dificulten una descripció unitària dels tipus de vegetació predominants.
Els boscos plujosos de coníferes vancouverians
L’exemple paradigmàtic de selva plujosa temperada amb predomini de coníferes és el que constitueixen les majestuoses masses forestals de les costes nord-occidentals de l’Amèrica del Nord. Aquesta és, a més, la part més ben coneguda del bioma, tant pel que fa a la seva reputació internacional com a l’esforç d’investigació que ha fomentat.
Els boscos d’avets, pícees i hemlock occidental
El bosc dels imponents avet de Douglas (Pseudotsuga menziesii) i hemlock occidental (Tsuga heterophylla), acompanyats, entre d’altres, de la tuia gegant (Thuja plicata) i l’avet roig americà (Abies amabilis), es distribueix en un interval d’altituds que va des del nivell de la mar fins a uns 1 000 m. Es tracta de grans masses de bosc madur pluriestratificat, dominat per arbres de fins a 90 m d’alt i 3 m de diàmetre. A l’estrat arbori inferior l’arbre dominant és el teix del Pacífic (Taxus brevifolia), mentre que els planifolis de fulla caduca i creixement ràpid, com el vern d’Oregon (Alnus rubra), l’erable de fulla gran (Acer macrophyllum) o el cirerer d’Oregon (Prunus emarginata), només són comuns en indrets que han sofert pertorbacions recents o bé a les riberes dels cursos d’aigua, on s’associen amb altres caducifolis més pròpiament riberencs, com el pollancre califòrnià (Populus trichocarpa) i el freixe d’Oregon (Fraxinus latifolia). A la part septentrional de les Muntanyes Rocalloses, l’arbre que domina als boscos madurs de les terres altes és el hemlock occidental, que és acompanyat a l’estrat arbori superior per la tuia gegant (en sòls humits), i per la varietat glauca de l’avet de Douglas (Pseudotsuga menziesii var. glauca, en indrets ben drenats i anteriorment afectats per alguna pertorbació).
Un segon complex propi d’aquestes selves plujoses és el que formen la pícea de Sitka (Picea sitchensis) i el hemlock occidental (Tsuga heterophylla). Aquest tàndem constitueix la vegetació dominant en una estreta franja costanera que s’estén des del N de Califòrnia fins al golf d’Alaska, on el clima és perhumit a causa del règim de precipitacions i el marge de temperatures és variable (pot ser entre fred i molt fred a l’extrem septentrional i entre temperat i subtemperat al meridional). A les illes d’Afognak, Kodiak i altres veïnes del golf d’Alaska, no hi creix cap altra conífera autòctona que la pícea de Sitka, que és l’arbre dominant a l’estrat arbori superior d’aquest complex, mentre que més al S, bé que rarament a altituds superiors als 750 m, el hemlock occidental és codominant i l’avet de Douglas (Pseudotsuga menziesii) i la tuia gegant (Thuja plicata) subdominants; el vern roig americà (Alnus rubra) apareix al sotabosc als indrets pertorbats. Al llarg de tota la costa d’Oregon i Washington, en indrets extremament humits o inundats o en el front de mar, també és comú el pi de Murray (Pinus contorta), i se sol trobar un dens sotabosc arbustiu, en el qual destaquen dues ericàcies, el “red huckleberry” (Vaccinium parvifolium) i la “mock azalea” (Menziesia ferruginea). A la península d’Olímpic, la pícea de Sitka i el hemlock occidental assoleixen alçàries de 60 m i un diàmetre de 2,30 a 3,30 m a 1,5 m de la base.
A la serralada de les Cascades del N d’Oregon i de Washington, el complex d’avet roig americà (Abies amabilis) i hemlock occidental (Tsuga heterophylla) caracteritza l’estatge montà entre els 1 000 i els 1 500 m d’altitud. Aquesta zona montana apareix també al Coastal Belt de la Colúmbia Britànica, als 48°N i entre els 750 i els 1 450 m d’altitud, i davalla fins al nivell de la mar en el seu límit septentrional. En tota l’àrea de distribució de l’avet roig americà, el clima és fred, regular i perhumit, i aquesta espècie apareix acompanyada pel hemlock occidental, l’avet pròcer o avet blau d’Oregon (Abies procera) i l’avet de Douglas (Pseudotsuga menziesii), etc.
Finalment, els boscos d’avet de Mertens (Tsuga mertensiana) formen l’estatge subalpí de la serralada de les Cascades, l’Insular Belt i les Coast Ranges, i s’estenen cap al N de les Muntanyes Rocalloses d’Idaho. A altituds menors, al N de l’illa de Vancouver, l’avet de Mertens es troba en torberes àcides i indrets humits, i a la badia de Prince William domina tot el paisatge. A la Colúmbia Britànica, l’arbre codominant a les torrenteres és el cedre groc o “Alaska yellow-cedar” (Chamaecyparis nootkatensis), i als vessants i carenes l’avet roig americà (Abies amabilis).
Els boscos de sequoia
Els boscos de sequoia costanera (Sequoia sempervirens) del S d’Oregon i del N i el centre de Califòrnia també poden assimilar-se a aquestes selves humides de coníferes, encara que el seu extrem meridional termeneja amb l’àrea mediterrània de Califòrnia. L’àrea de distribució d’aquests boscos és més aviat discontínua i forma una banda llarga i estreta de 740 km i amplada variable, relacionada amb la distribució del ‘cinturó de boira’ de la costa californiana septentrional i central, tret climàtic essencial per a la supervivència de la sequoia.
Els boscos de sequoia gegant (Sequoiadendron giganteum) es desenvolupen entre el nivell de la mar i els 915 m, però la majoria es troben en altituds compreses entre els 30 i els 760 m; són absents de la línia de costa perquè no toleren la salabror ni els vents oceànics. Acompanyant la sequoia solen aparèixer també el cedre de Port Orford (Chamaecyparis lawsoniana) i l’avet de Douglas (Pseudotsuga menziesii), així com la nou moscada californiana (Torreya californica), el pi de Jeffrey (Pinus jeffreyi), el pi de Lambert (P. lambertiana), el pi blanc americà (P. monticola) i el pi de Murray (P. contorta), entre d’altres.
Els boscos madurs de sequoia comprenen una rica barreja d’arbres de diferents dimensions i edats. La major part es troben en parcs estatals o nacionals i ocupen poc més de 80 000 ha en total. A mesura que augmenten l’altitud, l’aridesa i el pendent, la talla de les sequoies disminueix i apareixen altres espècies. Són arbres molt resistents, que es beneficien del fet que els incendis eliminen les espècies competidores, consumeixen els troncs caiguts, reciclen els minerals, obren clarianes i estimulen la caiguda de les llavors. Les inundacions tenen conseqüències semblants per a les sequoies, ja que els proporcionen les condicions ideals per a la germinació i el desenvolupament de les llavors i, a més, fertilitzen el sòl i s’enduen la virosta.
Els boscos mixtos de coníferes i roures
El bosc mixt d’avet de Douglas (Pseudotsuga menziesii) i planifolis de fulla perenne és el tipus de bosc més comú de la part septentrional de la Coast Range de Califòrnia i de les muntanyes Klamath, al N d’aquest estat. És un bosc de transició entre els boscos humits de la costa dominats per l’avet de Douglas i les mediterrànies californianes, i alguns autors el consideren un subtipus d’aquestes últimes. Però allà on l’avet de Douglas forma un estrat superior continu i alt (fins a 65 m), per damunt d’un altre de més baix (fins a 35 m) d’altres arbres, és més adient tractar aquest bosc com un tipus de bosc costaner de coníferes. En alguns indrets, l’avet de Douglas és l’únic arbre de l’estrat arbori superior, i el “tan oak” (Lithocarpus densiflora) forma un sotabosc espars sota seu. Més al N, cap a Oregon, la comunitat forestal amb avet de Douglas i planifolis de fulla perenne es troba confinada als vessants submèsics, sobre un substrat de diorita i a altituds d’uns 1 000 m; en aquests indrets forma comunitats de quatre estrats, amb l’avet de Douglas com a única espècie dominant a l’estrat arbori superior.
Per damunt dels 500 m d’altitud, sobre un substrat de serpentinita, als vessants de pendents moderats orientats al NE de les muntanyes Siskiyou, l’avet de Douglas (Pseudotsuga menziesii) és codominant en un bosc a mig camí entre el bosc mixt de coníferes i el bosc mixt amb planifolis de fulla perenne. Entre els arbres acompanyants hi ha el cedre d’encens californià (Calocedrus decurrens), el pi de Jeffrey (Pinus jeffreyi), el pi blanc americà (P. monticola), el pi de Lambert (P. lambertiana) i el pi d’Eldorado (P. attenuata), i el sotabosc és constituït per arbustos com el “huckleberry oak” (Quercus vaccinifolia), el “tan oak” (Lithocarpus densiflora), el “red huckle-berry” (Vaccinium parvifolium) i la gàrria de fulles de boix (Garrya buxifolia), entre d’altres. Als fons de les valls amb subtrats molt bàsics (peridotita, dunita i serpentinita) hi domina el xiprer fals de Lawson (Chamaecyparis lawsoniana), acompanyat pel pi blanc americà, el pi de Jeffrey i l’avet de Douglas.
Els boscos plujosos de coníferes temperats i subtropicals
A diferència de la costa nord-occidental de l’Amèrica del Nord, a l’Amèrica del Sud no s’hi fan boscos plujosos en què predominin absolutament les coníferes. Tanmateix, hi ha boscos en els quals coníferes com les araucàries (Araucaria araucana, A. angustifolia), el làrix andí o làrix americà (Fitzroya cupressoides) o el xiprer xilè (Libocedrus [=Austrocedrus] chilensis) són arbres emergents del cobricel i dominants en la vegetació.
Els boscos de làrix americà
El làrix americà (Fitzroya cupressoides) té una distribució discontínua i es troba disseminat entremig de les selves valdivianes de faigs australs (Nothofagus), per una regió dels Andes compresa entre els 39° 50’ S i els 43° 30’ S i per un sector de la Cordillera de la Costa comprès entre els 40°S i els 41° 10’ S. Acostuma a trobar-se en llocs deprimits i sobre sòls mal drenats, a altituds entre els 700 i els 1 000 m. Forma rodals gairebé purs o bé mixtos amb arbres com el “coihue” comú (Nothofagus dombeyi) i el “canelo” xilè (Drimys winteri), per damunt de les capçades dels quals destaca el làrix com a arbre emergent. Al sotabosc creixen, entre d’altres, el bambú Chusquea nigricans, la glossulariàcia Escallonia leucantha i els “michayes” (Berberis). En altres punts, el làrix americà es pot presentar associat amb el “coigüe” de Chiloé (Nothofagus nitida). Els boscos de làrix americà han estat molt explotats a causa de la bona qualitat de la fusta i del considerable volum dels arbres, que pot arribar a ser de fins a 3 000 m3/ha; els millors es troben als Andes, entre els 40° 30’ S i els 42°S, zona en la qual el làrix americà creix juntament amb el “mañío” de fulles punxants (Podocarpus nubigenus), sobre sòls prims i àcids (pH=3,7) derivats de cendres volcàniques. Els pocs rodals que queden presenten una regeneració escassa i es creu que estan en regressió.
Els boscos xilens d’araucàries
L’araucària xilena o “pehuén” (Araucaria araucana) forma boscos monoespecífics en sòls esquelètics dels vessants exposats al N i a altituds pròximes al límit altitudinal del bosc, i també ocupa petits claps associats a afloraments de roca i corrents d’aigua, als contraforts orientals dels Andes, cap a la Patagònia. En la resta de localitzacions sol formar boscos mixtos amb diferents espècies de faig austral (Nothofagus), ja sigui en situació de dominància o de codominància.
Els més importants d’aquests boscos mixtos de “pehuén” (Araucaria araucana) i faigs australs són els de “pehuén” i lenga (Nothofagus pumilio), en els quals aquest faig austral caducifoli domina l’estrat arbori inferior. Aquest bosc s’estén a altituds de 1 000 a 1 400 m, però, a Xile, no creix als vessants obacs, on s’hi fan boscos purs de lenga; als Andes argentins, en canvi, on la influència marítima és menor i la sequedat major, els boscos mixtos d’araucària xilena i lenga creixen precisament als obacs. Sovint s’hi desenvolupa un sotabosc molt espès de bambús del gènere Chusquea i també s’hi poden trobar arbustos com el “canelo enano” (Drimys winteri var. andina), la loganiàcia Desfontainia spinosa i diferents espècies de “michay” (Berberis).
Als Andes, per sobre els 900 m, es troben els boscos mixtos de “pehuén” (Araucaria araucana) i raulí (Nothofagus procera), un altre faig austral caducifoli, i de “pehuén” i “roble pellín” (N. obliqua), igualment caducifoli. Els boscos mixtos de “pehuén” amb “ñirre” (N. antarctica, un tercer faig austral caducifoli) al sotabosc són abundants a l’E dels Andes i al límit altitudinal del bosc. Aquest tipus de formació forestal és molt comuna als vessants orientals dels Andes on, probablement a conseqüència dels incendis, s’estén a altitud mitjana. Al S de la seva àrea de distribució, entre els 1 000 i els 1 300 m, l’araucària xilena s’associa amb el “coihue” comú (N. dombeyi), i formen un cobricel en què el “coihue” excedeix l’alçada de l’araucària en més de 30 m. Són boscos molt frondosos, amb un sotabosc ben desenvolupat, sobretot a les àrees meridionals, i amb una composició d’espècies similar a la dels boscos precedents. A la Cordillera de la Costa es desenvolupen a altituds més baixes, mentre que a la part argentina estan limitats als ambients més humits.
Els boscos brasilers d’araucàries
El “kuri’y” o “pino Paraná” (Araucaria angustifolia) és l’arbre emergent i dominant d’unes selves que tenen tres o quatre estrats arboris i estan formades per un gran nombre d’espècies de laurifolis i un nombre menor (1 de cada 4 o 5) d’espècies d’arbres caducifolis. En rodals densos s’han comptat fins a 65 exemplars de “kuri’y”/ha, però la mitjana supera en poc un terç d’aquesta quantitat. El diàmetre del tronc acostuma a ser d’entre 50 i 90 cm, però alguns arbres vells (el “kuri’y” pot assolir edats de 200 o 300 anys) arriben excepcionalment a fer 2 m. Aquests boscos estan interromputs pels “campos”, praderies sabanoides de gramínies, ocasionalment de caràcter edàfic però en la seva major part creades i afavorides per les tales i els incendis.
D’aquestes selves mixtes, amb el “kuri’y” (Araucaria angustifolia) com a arbre emergent, se’n diferencien quatre variants. La primera ocupa els terrenys al·luvials situats al llarg dels rius que neixen als vessants interiors de les serres costaneres, i les valls poc profundes del “planalto”; en els indrets més alts, el “kuri’y” es troba associat al “pinheirinho” (Podocarpus lambertii) i a la winteràcia Drimys brasiliensis. Una segona variant submontana, localitzada en els vessants per sota dels 500 m (exclosos els terrenys al·luvials dels fons de valls), ha quasi desaparegut a causa de l’explotació forestal. Una tercera variant, de caràcter montà, prospera a tot el “planalto” de la regió del Paranà, des dels 500 fins als 1 000 m d’altitud, als estats de Rio Grande do Sul, Paraná i Santa Catarina; aquesta variant presenta un sotabosc dens, amb arbres i arbustos com l’anacardiàcia Schinus spinosus, la winteràcia Drymis brasiliensis i la berberidàcia Berberis laurina, i falgueres arborescents com Dicksonia sellowiana, entre d’altres. Les branques dels arbres, d’altra banda, estan cobertes de molses, líquens i altres epífits, principalment orquídies i bromeliàcies, entre les quals diverses espècies de clavellina d’aire (Tillandsia). La quarta variant, altimontana, apareix a altituds superiors als 1 000 m; aquí, el “cedro” (Cedrela fissilis) comparteix l’estrat arbori amb l’araucària i també hi són presents els podocarps Podocarpus sellowii i P. lambertii.
Les pinedes de les planes nord-americanes sud-orientals
Les pinedes són un dels trets més característics de la vegetació del SE dels Estats Units, i ocupen aquest espai a causa de la pobresa i la textura sorrenca dels sòls i de la freqüència dels incendis. Sembla que el “southern pine” o pi de fulla llarga (Pinus palustris) era el més difós fins que la tala a què ha estat sotmès ha donat avantatge al pi “loblolly” (P. taeda). El pi d’Elliott (P. elliottii) i el pi cedre (P. glabra) hi tenen també un paper destacat.
En indrets molt secs (dunes fixades o turons sorrencs) poc o molt enlairats, amb sòls de sorra grossa i pocs nutrients, es fan pinedes esclarissades de pi de fulla llarga (Pinus palustris). Els arbres són esparsos, raquítics i malformats, i l’àrea basal és relativament petita. Aquesta pineda, l’estrat herbaci de la qual necessita cremar-se periòdicament per a mantenir-se, es desenvolupa sota un règim d’incendis freqüents però poc intensos. El pi de fulla llarga acostuma a ser l’únic arbre de l’estrat arbori, de vegades acompanyat pel “turkey oak” (Quercus laevis); l’estrat arbustiu, d’altra banda, més aviat baix i dominat principalment per ericàcies com el “stagger-bush” (Lyonia mariana) o el “dwarf huckleberry” (Gaylussacia dumosa), representa un 2% del recobriment. Als turons sorrencs de la regió del Piedmont dels Apalatxes, més frescal i menys castigada pels incendis, a l’estrat arbori inferior s’hi fan el roure de Maryland o “blackjack oak” (Quercus marilandica), el “sand post oak” (Q. margaretta) i el “bluejack oak” (Q. incana) i, secundàriament, el “tupelo” negre (Nyssa sylvatica) i el persimó (Diospyros virginiana), entre d’altres.
A les planes sorrenques baixes i deprimides els pins de fulla llarga o “southern pine” (Pinus palustris) assoleixen alçades considerables i apareixen sovint acompanyats pel pi d’Elliott (Pinus elliotti), el pi “loblolly” (P. taeda) i per unes quantes dicotiledònies que formen un estrat arbori obert. El sotabosc és dominat per un estrat arbustiu en el qual prenen importància cap al N els arbustos caducifolis i cap al S el “saw palmetto” (Serenoa repens), una palmera de port arbustiu; també hi ha un estrat herbaci en el qual abunden les poàcies. Als turons sorrencs més meridionals, principalment cap al N de Florida, s’hi fan pinedes mixtes amb un estrat arbori superior de pi de fulla llarga, pi d’Elliott i alzina de Virgínia (Quercus virginiana) i un estrat arbori inferior amb “turkey oak” (Q. laevis), “bluejack oak” (Q. incana), roure roig meridional (Q. falcata) i persimó (Diospyros virginiana). Un dels arbustos més comuns és la crisobalanàcia Chrysobalanus oblongifolius.
El “southern pine” (Pinus palustris), el pi d’Elliott (P. elliotti) i el pi “loblolly” (P. taeda) també són dominants en alguns tipus de boscos mixtos de les cotes altes de la plana costanera, però en indrets més ben drenats i més pobres en nutrients que els que ocupen els boscos mixtos de planifolis que es tractaran més endavant. Es tracta d’un estadi intermedi entre les pinedes i aquests boscos mixtos, amb una flora més pobra i una estructura més simple que les d’aquests darrers. L’estrat arbori superior comprèn també, a més d’aquells pins, dicotiledònies de fulla perenne com la magnòlia comuna (Magnolia grandiflora) i l’alzina de Darlington o alzina llorer comuna (Quercus hemisphaerica), i de fulla caduca, com el roure blanc americà (Q. alba), el “water oak” (Q. nigra), el “post oak” (Q. stellata), el tuliper de Virgínia (Liriodendron tulipifera) i el faig americà (Fagus grandifolia). També hi ha algun arbust caducifoli al sotabosc, com per exemple el sanguinyol florit (Cornus florida).
Els boscos mixtos de pins i roures mesoamericans
Queda encara molt per aclarir, i fins i tot per descobrir, sobre la taxonomia de les espècies arbòries de Mèxic i de l’Amèrica Central, però se sap que en aquesta regió hi ha més de 150 espècies diferents de roures o “encinos” (Quercus caducifolis i algun de perennifoli, però marcescent) i no menys de 80 de coníferes. No tots aquests “encinos” ni coníferes es comporten com a dominants, però la majoria sí que ho fan de manera regular o ocasional. Per això aquests boscos canvien de composició florística i de fisiognomia en funció de les condicions locals, en particular l’altitud i la latitud. La majoria de les coníferes, per exemple, tan sols arriben, cap al S, fins al N de Nicaragua, mentre que alguns “encinos” ressegueixen l’espinada del continent fins a Costa Rica i Panamà. A més, les coníferes es troben més aviat a altituds elevades, on les condicions els són més favorables; a Mèxic, fins a uns 2 000 m, els pins acostumen a aparèixer associats amb roures; més amunt, cap als 3 000 m, els rodals de pins esdevenen pinedes sense caducifolis, i són aquestes pinedes les que solen marcar el límit altitudinal del bosc, entre els 4 000 i els 4 200 m.
El més habitual és que en un indret donat convisquin dues o tres espècies arbòries (i sovint fins a vuit, deu o encara més), majoritàriament pins i roures, que de vegades estan barrejats amb altres coníferes, amb “madroños” (Arbutus i Arctostaphylos), amb el “capulín” (Prunus capuli), amb “ailes” (Alnus cordifolia i altres congèneres) i amb “tepozanes” (Buddleia americana i altres congèneres), la majoria pertanyents a gèneres originaris de l’espai holàrtic.
En aquests boscos, generalment, el cobricel superior deixa passar força llum als estrats inferiors, cosa que permet l’establiment al sotabosc de nombrosos arbustos i elements herbacis de famílies molt diverses, que són els que predominen quantitativament en qualsevol inventari florístic. Hi destaquen, sobretot, representants de la família de les compostes o asteràcies, així com labiades o lamiàcies, escrofulariàcies i solanàcies, moltes de les quals amb flors molt vistoses. A les pinedes sovint abunden les poàcies i, en condicions més ombrívoles i al peu de talussos, no hi falten diverses falgueres. També hi ha fongs en abundància, molts dels quals viuen en simbiosi amb les arrels dels arbres i produeixen atractius cossos fructífers en èpoques de pluges.
Al S de Mèxic, a les muntanyes més altes, els pins són substituïts gradualment per espècies de bosc nebulós. Els boscos de pins i roures també apareixen a les regions més elevades de Cuba i a la Hispaniola. En aquestes regions, ja subtropicals, es fa difícil delimitar biomes concrets. A l’estat d’Oaxaca, a Mèxic, hi ha selva tropical, selva temperada humida planoperennifòlia, bosc humit de coníferes i bosc nebulós en una mateixa àrea, que es caracteritza pel fet de presentar anomalies altitudinals i geogràfiques, com per exemple ombres pluviomètriques. En alguns casos, en una sola vall hi poden estar representats biomes diferents i encavalcats.
Els boscos inundables de les planes nord-americanes sud-orientals
Als aiguamolls inundables de les riberes del SE dels Estats Units, en sòls de pH baix, amb un elevat contingut en minerals i un estat d’inundació gairebé permanent, es donen poblaments de taxodi de Virgínia (Taxodium distichum) i de “tupelo” negre (Nyssa sylvatica). A les depressions mal drenades de tot el SE hi ha petites comunitats forestals dominades pel taxodi de marjal (Taxodium ascendens), que es diferencien ecològicament de les de taxodi de Virgínia pel fet que, al contrari que aquestes, apareixen en substrats pobres en nutrients, especialment en fòsfor, però de pH igualment àcid (entre 4,1 i 4,6). La clavellina d’aire localment anomenada “Spanish-moss” (Tillandsia usneoides), hi és omnipresent. Entre els arbres i arbustos acompanyants figuren el “dahoon” o cassina (Ilex cassine), el “tupelo” negre, el titi roig (Cyrilla racemiflora), la magnòlia d’aiguamoll (Magnolia virginiana), el “fetterbush” (Lyonia lucida), el baguer vermell o “redbay” (Persea borbonia), la cletra de marjal o “coast white-alder” (Clethra alnifolia) i diferents espècies de nabius (Vaccinium). Els rodals d’aquestes comunitats de marjal estan més ben desenvolupats al centre i al S de Florida, on es troben poblaments pràcticament purs de taxodis de marjal coberts de diverses espècies de clavellines d’aire (Tillandsia).
Damunt de substrats profunds de torba permanentment inundats i amb sorra a sota, en cursos d’aigua lents i al voltant de marjals permanents de la plana costanera atlàntica —des del N de Florida fins a Maine—, apareixen grans rodals densos i regulars de cedre blanc atlàntic (Chamaecyparis thyoides), sobretot a la costa de Carolina del Nord, Virgínia i S de Nova Jersey. En canvi, a la plana al·luvial del Mississipí aquests rodals ocupen tan sols 4 900 ha, i el cedre blanc atlàntic representa només el 50% de l’estrat arbori superior. En depressions mal drenades, com ara als marjals costaners i les depressions de rereduna amb sòls torbosos de les costes de Carolina del Sud i Carolina del Nord, apareixen tot un seguit de comunitats higròfiles que toleren un pH baix, poc oxigen i concentracions elevades d’àcids orgànics; els indrets permanentment inundats, en canvi, presenten una comunitat graminoide de poàcies i ciperàcies, que sovint forma tapissos flotants. Els marjals són dominats per poblaments arbustius amb exemplars esparsos de pi de marjal (Pinus serotina); aquestes formacions, anomenades “pocosin”, es fan en substrats de torba, sovint sense contacte amb el substrat mineral, i obtenen els nutrients de la pluja i de la virosta arrossegada pel vent.
Les laurisilves i els boscos mixtos austroamericans
A l’extrem austral del continent americà, fonamentalment al vessant pacífic i des dels 37° 45’ S cap al S, els faigs australs (Nothofagus) imposen la seva llei a les comunitats forestals, encara que localment poden fer-se dominants algunes coníferes que, cap al S, esquitxen el paisatge fins als 51°S de latitud. Els arbres planifolis més característics, i endèmics de la regió, són el “coihue” o “coigüe” comú (N. dombeyi), el “coigüe” de Chiloé (N. nitida) i el “coigüe” magallànic o “guindo” (N. betuloides). També són comunes altres espècies de planifolis, com l’“ulmo” (Eucryphia cordifolia), el “tineo” (Weinmannia trichosperma) i el “canelo” xilè (Drimys winteri), fins a sumar una cinquantena d’espècies arbòries de les quals el 95% són endèmiques de Xile i les regions adjacents de l’Argentina.
En correspondència, a grans trets, amb les regions climàtiques del S de Xile i de l’Argentina limítrofa, i substituint-se l’una a l’altra de N a S (sense límits precisos) en resposta al descens tèrmic, es reconeixen tres regions fitogeogràfiques: la selva valdiviana (entre els paral·lels 37° 45’ S i 43° 20’ S), els boscos plujosos nord-patagònics (entre els parallels 43° 20’ S i 47° 30’ S) i els boscos magallànics perennifolis (entre els paral·lels 47° 30’ S i 55°S). Moltes zones d’aquests boscos es mantenen molt inestables i estan subjectes a grans pertorbacions, sobretot a la regió de bosc plujós valdivià del vessant occidental dels Andes, on l’activitat volcànica i els terratrèmols causen sobtats moviments de terra i esllavissades que arranen vastes extensions i afavoreixen l’inici de noves successions. Després d’una erupció, les cendres volcàniques també destrueixen la vegetació existent en grans àrees, de manera que la freqüent activitat volcànica dels últims cinc segles sembla haver estat un factor determinant en la composició d’aquests boscos. Sobretot en àrees d’elevació mitjana, les espècies colonitzadores com el “coihue” comú (Nothofagus dombeyi) tenen tendència a formar rodals pràcticament monoespecífics, i a les clarianes que deixen les caigudes d’arbres s’implanten ràpidament espècies oportunistes, com el ”huahuán” o “tepa” (Laurelia philippiana).
Les selves valdivianes
La regió de la selva valdiviana presenta una gran diversitat d’arbres i una biomassa molt elevada. Les espècies laurifòlies més significatives són el “coihue” comú (Nothofagus dombeyi), l’“ulmo” (Eucryphia cordifolia) i el “tineo” (Weinmannia trichosperma), a les quals s’uneixen coníferes i una proporció variable d’espècies caducifòlies. A partir dels 900 o 1 000 m d’altitud, aproximadament, la laurisilva dóna pas als boscos decidus de lenga (Nothofagus pumilio) i “ñirre” (N. antarctica), que arriben fins al límit altitudinal del bosc; més amunt es desenvolupen prats altoandins. A altituds compreses entre els 1 000 i els 1 400 m també es troben els boscos de “pehuén” (Araucaria araucana) descrits anteriorment. La selva penetra també puntualment per les valls cap al vessant argentí de la serralada andina, com ara al N de Cerro Tronador, a Puerto Blest i al llac Frías, al parc nacional de Nahuel Huapi, però en aquests indrets és molt més pobra en espècies.
Perfil transversal de les muntanyes de Xile als 41° de latitud S, que mostra el continu de formacions vegetals que se succeeixen a l’extrem austral del continent americà, amb indicació de l’altitud i el volum de precipitacions. Les laurisilves d’aquesta regió estan dominades indiscutiblement pels faigs australs (Nothofagus), tant perennifolis com caducifolis. Així, l’anomenat “roble pellín” (N. obliqua) és un caducifoli que forma extensos boscos als vessants orientals de la Cordillera de la Costa, mentre que la lenga (N. pumilio), també caducifoli, domina a la serralada andina aproximadament entre els 1 250 i els 1 500 m, bé que al vessant occidental es barreja amb una espècie perennifòlia, el “guindo” o “coigüe” magallànic (N. betuloides).
Jordi Corbera, a partir de C. Donoso, 1993
A la depressió central xilena, en sòls ben drenats, es dóna el bosc de “roble pellín” (Nothofagus obliqua), espècie caducifòlia que forma un estrat emergent per damunt d’un cobricel dominat per les capçades de tres laurifolis, el llorer xilè o “laurel” (Laurelia sempervirens), el “lingue” (Persea lingue) i l’“olivillo” (Aextoxicon punctatum), que superen en nombre i en àrea basal el “roble pellín”. Altres boscos de la zona, de composició semblant, tenen l’“ulmo” (Eucryphia cordifolia) com a espècie dominant, i en aquest cas, a l’estrat arbori inferior hi destaquen, entre altres, l’“olivillo” i el “huahuán” (Laurelia philippiana), mentre que el “roble pellín” i les altres espècies de faigs australs en són absents.
En sòls poc drenats, entre Valdivia i Puerto Montt, es dóna el bosc de “coihue” comú (Nothofagus dombeyi) amb “canelo” xilè (Drimys winteri) i diverses mirtàcies. Creix sobre terrenys ondulats de sediments glacials on s’ha desenvolupat una capa d’òxids de ferro que impossibilita el drenatge (“ñadis”), o bé en indrets mal drenats a causa de la seva topografia. El “coihue” comú esdevé l’espècie arbòria dominant sobre un estrat format pel “canelo” xilè i altres mirtàcies que formen l’estrat arbustiu, i les falgueres i epífits hi són abundants. Com en el cas anterior, gran part del bosc ha estat artigat per fer-ne terres de cultiu i pastura. Sobre sòls encara més mal drenats, prop dels cursos d’aigua, i en una gran varietat de localitats es desenvolupen els “tepuales”, un bosc en el qual domina el “tepú” (Tepualia stipularis), una mirtàcia d’uns 8 m d’alçada i de tronc múltiple (tret que es considera una reminiscència dels manglars tropicals), de vegades acompanyat de la cupressàcia Pilgerodendron uvifera, el “ten”, i en altres ocasions del “canelo” xilè.
Als vessants occidentals de les muntanyes, entre els 400 i els 700 m, en sòls ben drenats derivats de cendres volcàniques, són típics els boscos mixtos dominats pel “coihue” comú (Nothofagus dombeyi). Aquest bosc és abundant als Andes, on les precipitacions, en aquestes altituds mitjanes, arriben als 4 000 mm, i és escàs a Chiloé; a la Cordillera de la Costa xilena no s’ha descrit. L’estrat arbori superior està format per “coihue” comú i “ulmo” (Eucryphia cordifolia) i a l’inferior hi ha “tepa” (Laurelia philippiana) i “mañío” de fulles curtes (Saxegothaea conspicua). A altituds superiors, fins als 950 m, sobre sòls profunds i rics en matèria orgànica, es desenvolupa un bosc similar a l’anterior però amb una composició una mica diferent; el “coihue” comú hi assoleix el seu màxim desenvolupament, amb alçades de fins a 50 m, diàmetres de més de 2 m i grans contraforts al peu del tronc. Als vessants orientals i occidentals de la Cordillera de la Costa, al S de Valdivia, i als vessants orientals de Chiloé, a altituds compreses entre els 600 i els 900 m, es fan boscos de “coigüe” de Chiloé (Nothofagus nitida), amb “mañío” de fulles curtes, “mañío” de fulles punxants (Podocarpus nubigenus), “tineo” (Weinmannia trichosperma) i “canelo” xilè (Drimys winteri). Es tracta de localitats on la humitat és molt alta i el drenatge dolent, amb sòls àcids (pH=4) i un contingut elevat de matèria orgànica. Als vessants dels Andes, per damunt dels 1 000 m, es forma un bosc de transició de “coihue” comú amb lenga (Nothofagus pumilio), espècie caducifòlia que, tal com s’ha indicat, més al N forma el límit del bosc. Aquí, en comptats punts més humits apareix, molt localitzat, el “mañío” de fulles punxants, i al sotabosc abunda el bambú Chusquea tenuiflora. Els sòls són prims i deriven de deposicions volcàniques recents. Prop del límit del bosc de lenga, al S del paral·lel 40° 48’ S, el “guindo” (Nothofagus betuloides) substitueix el “coihue” comú, i més al S encara és el “guindo” el que constitueix el límit del bosc.
Les laurisilves patagòniques i magallàniques
Les zones més meridionals del bioma estan menys explorades i per això els tipus de bosc estan definits amb menys precisió. A mesura que augmenta la latitud austral les temperatures baixen, decreix la diversitat i es perden alguns elements valdivians. El “guindo” (Nothofagus betuloides) desplaça el “coihue” comú (N. dombeyi), que tanmateix roman l’espècie dominant al cobricel fins als 45° 32’ S, acompanyats l’un i l’altre pel “tineo” o “tinel” (Weinmannia trichosperma), el “mañío” de fulles curtes (Saxegothaea conspicua) —que desapareix als 46° 41’ S—, el “mañío” de fulles punxants (Podocarpus nubigenus) i el “canelo” xilè o “boighe” (Drimys winteri).
Les tres espècies perennifòlies de faig austral, el “coigüe” de Chiloé (Nothofagus nitida), el “guindo” (N. betuloides) i el “coihue” comú (N. dombeyi), formen un bosc mixt que s’estén al llarg de la franja de costa fins als 48°S. També hi són presents el “mañío” de fulles punxants (Podocarpus nubigenus), el “canelo” xilè (Drimys winteri), el “maitén negro” (Maytenus magellanica) i la tiaca (Caldcluvia paniculata), entre altres. El bambú Chusquea quila és important al sotabosc fins gairebé els 47°S de latitud. A la zona costanera de sòls més humits predomina el “guindo”, i a l’interior, més sec, el “coihue” comú, que no passa dels 48°S. Al sotabosc també són importants les falgueres arborescents com Blechnum magellanicum i B. chilense, i les lianes i epífits.
Al S dels 47° 30’ S, en un clima temperat fred molt humit, sobre sòls podzòlics de transició a torbosos i, en general, sobre sòls en els quals predominen els processos hidromòrfics, apareix la selva plujosa magallànica dominada pel “guindo” (Nothofagus betuloides), que arriba fins als 55°S, la latitud més extrema que assoleix a l’hemisferi austral la vegetació forestal. Aquesta selva plujosa no conté lianes, ni plantes enfiladisses, ni bambús. A la costa, on les condicions són més suaus, es desenvolupa un bosc mixt dominat pel “guindo” com a arbre emergent a l’estrat arbori superior, sobre un estrat arbori inferior format per “canelo” xilè (Drimys winteri), “maitén negro” (Maytenus magellanica) i “tepú” (Tepualia stipularis), encara que aquestes espècies no sempre apareixen juntes. L’estrat arbustiu és dens però pobre. Allí on el drenatge és bo, la lenga (Nothofagus pumilio), que és un arbre caducifoli, forma rodals mixtos amb el “guindo”. El “mañío” de fulles punxants (Podocarpus nubigenus), resisteix els sòls més mal drenats i arriba fins al paral·lel 51°S, formant rodals marginals enmig del bosc magallànic.
Els boscos de “guindo” (Nothofagus betuloides), sovint acompanyat del “ñirre” (N. antarctica), arriben fins al límit altitudinal del bosc i fan el paper de boscos subalpins; aquí, els arbres tenen formes rabassudes i la regeneració és vegetativa, per rebrots que surten de la base del tronc. El límit altitudinal del bosc se situa als 400 m a la Terra del Foc, i als 600 m al N de l’àrea de distribució d’aquest bosc. Més amunt, les parts altes i planes de les muntanyes que es troben al S del paral·lel 45°S es desenvolupa la tundra magallànica, que es diferencia de la boreal per l’absència de permafrost. La vegetació, que es desenvolupa sobre sòls àcids torbosos, és oligotròfica i està formada per esfagnes, plantes pulviniformes, ciperàcies i poàcies. La tundra domina també la banda costanera externa d’illes a partir del paral·lel 47°30’S, zona en la qual els boscos —que persisteixen fins als 55°S— queden acantonats als indrets més arrecerats.
Les laurisilves de la regió del Paranà
Encara que la biodiversitat de les selves de la regió del Paranà i missionera és menor que la de les selves tropicals del mateix continent, com les amazòniques i les de la “mata” atlàntica, aquestes selves plujoses subtropicals semiperennifòlies tenen un particular interès ecològic, justament a causa del seu caràcter extratropical. S’hi troben línies filogenètiques d’origen amazònic (s’estima que gairebé un 50% dels gèneres de plantes —encara que només un 5% de les espècies— són compartits amb la selva amazònica) i adaptades a condicions ambientals menys uniformes i, per tant, sotmeses a un estrès moderat. Les selves missioneres més ben conservades són exuberants, molt espesses i ombrívoles, i els escau la qualificació de laurisilves pel fet que hi dominen els arbres de la família de les lauràcies i els laurifolis d’altres famílies, tot i que també s’hi troben arbres caducifolis en proporcions variables, en funció de les condicions edàfiques, topogràfiques i geogràfiques, i del règim de pertorbacions de cada punt. Les selves missioneres de l’Alto Paraná, a Paraguai, i la de Misiones, a l’Argentina, tenen una gran semblança en molts aspectes, i les diferències més grans s’observen entre zones selvàtiques amb diferents històries d’intervenció, independentment de la regió. En general es caracteritzen per presentar una organització vertical en dos (de vegades tres) estrats. L’estrat arbori superior pot arribar a assolir fins a 32 m d’alçada, mentre que el segon arriba fins als 18 m i és més dens i atapeït.
Les lauràcies i les palmeres predominen als boscos madurs, bé que aquestes darreres només esdevenen dominants en determinats estadis de degradació; els arbres de les famílies de les meliàcies i les sapotàcies, d’altra banda, són importants en els indrets de sòl argilós ben drenat. Les rutàcies dels gèneres Balfourodendron i Zanthoxylum [=Fagara] abunden en tots els boscos de sòl sorrenc, mentre que damunt dels terrenys mal drenats predominen les til·liàcies i les sapindàcies. Les famílies de lleguminoses són més importants durant els estadis inicials de la successió, i les mirtàcies abunden en els ecotons entre la selva i la vegetació sabanoide dels “campos”, en els rodals isolats de bosc, allà on la vegetació forestal envaeix zones desforestades i en sòls mal drenats. L’herba mate o “ka’a” (Ilex paraguariensis) és una espècie esciòfita i higròfita selectiva, característica del sotabosc d’aquestes laurisilves. Generalment creix associada amb la “imbuia” (Ocotea porosa), O. pulchella, el “laurel negro” o “aju’y hu” (Nectandra angustifolia), la “canela amarela” o “aju’y” (N. lanceolata), el “cedro” o “ygary” (Cedrela fissilis) i altres Ilex (I. brevicuspis, I. microdonta, I. theezans i I. dimosa).
Les laurisilves altes semiperennifòlies
En la seva forma més típica, les laurisilves de la regió del Paranà es fan en sòls argilosos, arenosoargilosos i, més rarament, arenosos. Les cesalpinàcies i mimosàcies presenten una alta diversitat d’espècies, entre les quals destaquen alguns arbres emergents del cobricel, com la “grapia” o “yvyra pere” (Apuleia leiocarpa) o el “timbó” o “timbaúva” (Enterolobium contortisiliquum), però el nombre d’exemplars de lauràcies que formen l’estrat intermedi més alt, de 20 m, és superior. Les espècies més característiques són lauràcies com Ocotea puberula i Nectandra angustifolia, i la palmera més característica és el “pindo” o “geriba” (Syagrus romanzoffiana), que té una gran plasticitat. L’arbust més característic és l’henecàrtia (Hennecartia omphalocarpa), de la família de les monimiàcies.
Sobre terres ben drenades, àcides i de color vermell fosc, a les zones més altes dels costers i els plans més elevats del “planalto”, la meliàcia Cedrela fissilis, anomenada a la regió “ygary”, “cedro” o “cedro misionero”, és l’espècie que domina en l’estrat superior, mentre que als estrats inferiors es troben la “canjerana” o “cedro ra” (Cabralea canjerana) i encara altres arbres de la mateixa família, les meliàcies, com Guarea kunthiana, l’“yrupe rupa”, i diferents espècies de Trichilia. Per aquesta raó aquestes selves han estat anomenades de vegades ‘selves de meliàcies’. Les dues primeres d’aquestes espècies es troben extensament distribuïdes a les regions neotropicals, però són singularment més abundants al SE del Brasil, l’E de Paraguai i el NE de l’Argentina. L’“aguai” o “aguaí dulce” (Chrysophyllum gonocarpum), una sapotàcia important de l’estrat intermedi d’aquestes selves, també té una distribució molt extensa.
Una altra variant és la selva de lauràcies amb “guatambú blanco” o “yvyra neti” (Balfourodendron riedelianum), una rutàcia que es fa a les riberes del Paranà, al Brasil, l’Argentina i el Paraguai, en sòls argilosos o arenosoargilosos però ben drenats. Abans de les explotacions selectives, aquesta fou possiblement la laurisilva més estesa i representativa de la regió, però actualment, encara que manté una elevada biodiversitat, hi són rars els arbres de més de 30 m. Aquest tipus de selva de “laurel” i “guatambú” (lauràcies i Balfourodendron) sembla que està més ben conservada a l’Argentina, on s’han enregistrat densitats de 442 arbres/ha, pertanyents a una seixantena d’espècies, amb una àrea basal de 18,5 m2 i un diàmetre mitjà de 23,1 cm.
Les laurisilves baixes amb mirtàcies
Entre les laurisilves altes i les sabanes, en sòls eixuts i arenosos i sòls hidromòrfics, a les zones en què la vegetació viu en condicions més extremes, les selves baixes amb mirtàcies formen una banda de transició. El seu tret distintiu és que l’alçada del cobricel només és d’uns 9 m. En la composició florística també hi ha espècies de la selva semiperennifòlia de lauràcies, però dominen les espècies de mirtàcies, anacardiàcies i mirsinàcies. La majoria dels arbres perden el fullam durant l’estació seca. Les selves de mirtàcies tenen un paper molt significatiu com a boscos invasors de la sabana, on formen rodals que faciliten l’establiment de les espècies de la laurisilva alta. Colonitzen també les zones més fluctuants pel que fa a les condicions d’humitat edàfica, per exemple les planes d’inundació dels cursos d’aigua.
Seguint les riberes dels rius al llarg de les planes d’inundació es fa una selva en galeria o de ribera d’amplada variable però que a les riberes pel Paranà, per exemple, pot arribar als 5 km. Les fluctuacions dels cursos fluvials originen meandres i dipositen materials formant barres, damunt les quals creix una laurisilva amb vegetació molt variada. Les espècies més higròfiles, com les “ingas” (Inga, família de les mimosàcies) o algunes espècies de mirtàcies, ocupen les zones més exposades. Algunes euforbiàcies com les del gènere Sapium o la “sangre de drago” o “uruku’ra” (Croton urucarana) i altres congèneres també són molt abundants i solen ocupar estrats inferiors.
Les laurisilves i els boscos mixtos de les planes nord-americanes sud-orientals
A les terres altes ben drenades però riques en nutrients de la Plana Costanera Atlàntica es fa el típic bosc planifoli mixt meridional. L’estrat arbori superior el solen constituir planifolis de fulla caduca: faig americà (Fagus grandifolia), liquidàmbar americà (Liquidambar styraciflua), cària vera (Carya glabra), diferents roures (Quercus), i sovint aurons (Acer) i tells (Tilia). Entre els planifolis de fulla perenne hi ha la magnòlia comuna (Magnolia grandiflora), l’alzina de Darlington o alzina llorer comuna (Quercus hemisphaerica), el grèvol americà comú (Ilex opaca), i localment l’alzina de Virgínia (Quercus virginiana) i, pel que fa a coníferes, el pi cedre (Pinus glabra), el pi “loblolly” (P. taeda) i, localment, el pi americà de fulla curta (P. echinata). A l’estrat arbori inferior es troben el carpinus de Carolina (Carpinus caroliniana), el sanguinyol florit (Cornus florida), la morera americana (Morus rubra) i l’òstria americana comuna (Ostrya virginiana), i a l’estrat arbustiu palmes com el “saw palmetto” (Serenoa repens), el “blue-stem palmetto” (Sabal minor) o Rhapidophyllum hystrix.
Als sòls humits de les parts més enlairades de les planes inundables es donen boscos mixtos amb una major representació de caducifolis. Els boscos de transició cap a boscos mixtos d’aiguamoll estan dominats pel roure castanyer d’aiguamoll (Quercus michauxii) i el roure d’escorça de cirerer (Q. pagodaefolia), acompanyats per “water oak” (Q. nigra), alzina de Virgínia (Q. virginiana) —a les àrees properes a la costa—, cària vera (Carya glabra), pi cedre (Pinus glabra) i pi “loblolly” (P. taeda). Els arbres de l’estrat arbori inferior són principalment el grèvol americà comú (Ilex opaca), la papaia nord-americana anomenada “pawpaw” (Asimina triloba) i el “spicebush” (Lindera benzoin). Al sotabosc de les regions meridionals i orientals s’hi fan palmeres arbustives (al S sobretot el “blue-stem palmetto” (Sabal minor) i a l’E el “saw palmetto”, (Serenoa repens), respectivament).
Als sòls estacionalment inundats adjacents a rius i corrents d’aigua es forma un continu de boscos interzonals, dominats per arbres de fulla caduca i amb planifolis de fulla perenne confinats usualment al sotabosc. Aquests boscos ocupen 6,1 milions d’ha a les terres al·luvials del Mississipí, sobretot al N dels 31°N, i s’hi han reconegut una considerable varietat de tipus, els principals dels quals, per ordre decreixent de superfície ocupada, es caracteritzen per tenir com a conjunts respectius d’espècies dominants el liquidàmbar americà (Liquidambar styraciflua), el roure de Nuttall (Quercus nuttallii) i el roure salze (Q. phellos), el lledoner dolç (Celtis laevigata), l’om blanc americà (Ulmus americana) i el freixe roig americà (Fraxinus pennsylvanica), el roure castanyer d’aiguamoll (Quercus michauxii), el roure d’escorça de cirerer (Q. pagodaefolia), l’“over-cup oak” (Q. lyrata) i la cària aquàtica (Carya aquatica) i tot el complex d’arbres planifolis de ribera (bedolls, plàtans, freixes, oms, erables, salzes i pollancres).
Als marjals de terra baixa, en sòls torbosos, es fan els anomenats “bayforests” (a Carolina del Nord, a Carolina del Sud i Geòrgia), “bayheads” (a Florida) o “baygalls” (a Texas), selves d’aiguamoll denses i altes en les quals dominen la magnòlia d’aiguamoll (Magnolia virginiana), el baguer vermell o “redbay” (Persea borbonia), la gordònia americana o “loblolly-bay” (Gordonia lasianthus) i el “tupelo” negre (Nyssa sylvatica var. biflora, a través del cobricel del qual emergeixen les capçades d’alguns pins esparsos). De fet, el terme anglès “bay” (equivalent al català ‘baguer’, el peu femení del llorer) s’aplica en aquesta regió a un gran nombre d’arbres i arbustos laurifolis de manera que el terme “bayforest” i els seus sinònims equivalen a laurisilva. Al centre de Florida el domini dels laurifolis és absolut, però la presència i la importància dels caducifolis augmenta en direcció N, fins a arribar a Carolina del Nord, i cap a ponent fins a l’E de Texas.
La vegetació de les selves temperades d’Austràlia i Nova Zelanda
Les selves humides d’Austràlia i Tasmània ocupen una proporció relativament petita del total del territori australià, principalment a causa de l’aridesa generalitzada d’aquell continent, el més eixut després de l’Antàrtida. Es desenvolupen sota diferents règims de pluviositat, però sempre amb precipitacions de com a mínim 1 500 mm, encara que es troben vegetacions afins o claps de selva humida al llarg de cursos d’aigua, en indrets amb pluviositats de l’ordre dels 500 mm anuals (ocasionalment 300 mm). A l’W de Tasmània (amb clima temperat i pluges d’hivern) i al cap York, a l’extrem septentrional d’Austràlia (amb clima tropical i màxim relatiu de pluges a l’estiu), les pluviositats més elevades s’acosten als 3 500 mm anuals. Però el règim de pluges varia notablement entre aquests dos extrems: al N es dóna un règim monsònic d’estiu, estació que arriba a concentrar el 90% de les pluges, i al SE d’Austràlia i a Tasmània un règim temperat, amb pluges i precipitacions de neu (a les cotes més altes) principalment hivernals.
A Nova Zelanda, ben al contrari, el clima està fortament influït per la peculiar situació geogràfica, enmig de l’oceà (a latituds compreses entre els 30° i els 50°S) i pel fet que aquest arxipèlag rep regularment vents de ponent carregats d’humitat; aquestes condicions fan que la totalitat dels boscos autòctons, un terç del territori del país, s’assimilin a selves plujoses temperades. L’arrenglerament de les muntanyes en un sentit N-S constitueix una barrera per als vents dominants de l’W que causen precipitacions, sovint fortes i prolongades, a les àrees occidentals i a les terres altes. En aquestes àrees, la precipitació anual pot sobrepassar els 10 000 mm/any, amb fortes nevades i una coberta de neu de llarga durada per sobre dels 1 000 m d’altitud. La part oriental del país, en canvi, sofreix els efectes d’ombra pluviomètrica de les muntanyes, que són particularment acusats en algunes conques interiors localitzades entre les serralades axials dels Alps Meridionals neozelandesos i les serres exteriors. A l’Otago central, al S de Nova Zelanda, les condicions arriben a ser semiàrides i les precipitacions atenyen tan sols els 300 mm anuals.
Les selves temperades denses australianes i tasmanianes
La vegetació de les selves plujoses australianes és molt diferent de la de la resta del continent. En general són pluriestratificades, el cobricel és dens i les seves afinitats florístiques amb els altres tipus de vegetació australians són relativament petites, no sols pel que fa a espècies, sinó fins i tot a gèneres i famílies; només un petit nombre d’aquestes (dil·leniàcies, fabàcies, mimosàcies, mirtàcies i rutàcies) és àmpliament compartit dins i fora de les selves plujoses. La seva distribució de N a S al llarg de la costa oriental del continent fa que pràcticament no hi hagi discontinuïtat entre les selves plujoses tropicals i les temperades. Al SE de Queensland i a les àrees adjacents de Nova Gal·les del Sud, on la pluviositat és relativament menor, es desenvolupen selves plujoses subtropicals en les quals les cunoniàcies prenen una rellevància creixent de N a S. Al llarg de les serres i planes costaneres d’aquesta regió, fins a uns 1 000 m d’altitud, predominen associacions, sovint molt riques florísticament, primer dominades per diferents esterculiàcies del gènere Argyrodendron i, més cap al S, per cunoniàcies com el “coachwood” (Ceratopetalum apetalum) i altres.
La baixa intensitat lumínica i l’acumulació de brossa donen sovint com a resultat una coberta poc densa arran de terra i una gran abundància de falgueres. Els estrats arbustius estan poc desenvolupats. Cap al S, diverses famílies i gèneres tropicals van desapareixent a mesura que assoleixen als seus límits meridionals i hi ha una pèrdua notable de diversitat. Allà on la pluviositat baixa fins a 1 100-1 400 mm, es poden donar les selves plujoses ‘seques’, normalment amb dos únics estrats arboris, el superior constituït pels arbres emergents del cobricel que es distribueixen de manera discontínua i sovint són araucàries, principalment el “hoop pine” (Araucaria cunninghamii), que dóna a aquestes ‘pinedes’ un aspecte semblant a les de “kuri’y” (A. angustifolia), de la regió del Paranà. Més al S es generalitzen les selves denses dominades per cunoniàcies, i a les àrees de clima més frescal i humit, les dominades per faigs australs.
Les selves plujoses de faig austral i afins
Al SE d’Austràlia i a Tasmània les selves temperades es caracteritzen per la dominància dels faigs australs (Nothofagus) o de conjunts d’arbres en els quals algun faig austral és rellevant. Se’n poden distingir dos tipus generals: la selva densa baixa i l’alta. La selva densa baixa és montana i la seva àrea de distribució queda restringida a les terres altes de Victòria i de Tasmània. Es caracteritza per la gran quantitat de molses i líquens que cobreixen els troncs dels arbres, la fusta morta i el terra, per la presència de nombrosos arbres petits (a vegades amb port arbustiu, ramificats des de la base) i per tenir un cobricel relativament baix, que rarament supera els 10 m. La selva densa alta es dóna en altituds inferiors i pot enfilar el cobricel a alçades de fins a 30 m. Generalment, la densitat de troncs, que tenen un diàmetre major i no se solen ramificar des de la base, és molt inferior.
Les selves plujoses temperades de Tasmània són especialment notables, tant pel que fa a les espècies i associacions característiques que s’hi troben com pel seu atractiu visual. L’espècie més notable entre les dominants és el “myrtle beech” o faig austral de fulla de murta (Nothofagus cunninghamii). A la costa occidental de Tasmània, a les muntanyes Hartz i a la regió del llac Saint Clair, n’apareixen poblaments gairebé purs, amb exemplars de gran port (poden assolir una alçada de 50 m), i d’elevada densitat, amb poques plantes vasculars als estrats inferiors a causa de l’atapeïment i la poca obertura del cobricel; s’hi pot fer, però, un sotabosc discontinu amb una important presència de la falguera arborescent Dicksonia antarctica. A la terra baixa tasmaniana el faig austral de fulla de murta s’associa sovint amb el sassafràs de Tasmània (Atherosperma moschatum), que sol formar un estrat arbori intermedi per sota del cobricel del faig austral, o que fins i tot pot esdevenir dominant en alguns emplaçaments o després de determinades pertorbacions. També s’hi associen el pi de Huon (Dacrydium franklinii) i l’om de Tasmània (Eucryphia lucida) i, als boscos subalpins, el “celery-top pine” (Phyllocladus aspleniifolius, que també es pot trobar a menor altitud) i Eucryphia millinganii.
Globalment, la flora que pot associar-se amb la selva densa alta de “myrtle beech” (Nothofagus cunninghamii) potser no és gaire nombrosa, però inclou espècies ben característiques, com el bruc enfiladís tasmanià (Prionotes cerinthoides), que no és un bruc, ni tan sols una ericàcia, sinó una epacridàcia endèmica de Tasmània. A les selves tasmanianes, a diferència del que passa a les de Victòria, no hi arriben espècies pròpies de les selves plujoses més càlides de més al N. No obstant això, alguns gèneres, i fins i tot espècies, són compartits amb Nova Zelanda, i altres amb l’extrem austral de l’Amèrica del Sud. A l’estat de Victòria, en canvi, no està gaire estès el “myrtle beech”, i sovint es troba confinat en barrancs frescals i humits enmig del bosc de freixe de muntanya de Tasmània (Eucalyptus regnans).
A les àrees de Tasmània i Victòria que no són prou frescals i humides perquè hi domini algun faig austral tendeixen a fer-se dominants algunes espècies de la família de les cunoniàcies: el “coachwood” (Ceratopetalum apetalum), Anodopetalum biglandulosum i Schizomeria ovata, representatives de l’antiga flora gondwaniana. Des dels 30°S cap al N, les selves plujoses esdevenen molt més riques en espècies i s’hi fan predominants les esterculiàcies, en particular les del gènere Argyrodendron, però també s’hi troben lauràcies, rutàcies, meliàcies i representants d’altres famílies pròpies d’àrees predominantment boreals. A les àrees septentrionals més seques, els boscos d’Argyrodendron són substituïts pels boscos monsònics. A Nova Gal·les del Sud i el SE de Queensland, el “negrohead beech” o faig austral de Moore (Nothofagus moorei) esdevé dominant només en emplaçaments rics en nutrients, i en emplaçaments menys fèrtils, és reemplaçat amb èxit per plantes de la selva plujosa tropical del N, excepte en altitud. Als límits més septentrionals del bosc de faig austral, als confins entre Nova Gal·les del Sud i Queensland, el faig no es regenera per llavor sinó per rebrot.
Les selves temperades de ribera
Al llarg dels cursos d’aigua, de vegades adjacents a tipus de vegetació força diferents, apareixen boscos de ribera que a Austràlia són qualificats de ‘selves plujoses en galeria’. Se’n donen dues formes particulars, una amb espècies característiques que voregen els cursos d’aigua dins de zones de selva plujosa, i una altra localitzada en franges boscades, normalment massa seques per a la selva plujosa. Els exemples de la primera són nombrosos; inclouen el pi de Huon (Dacrydium franklinii) al llarg dels rierols que travessen boscos de “myrtle beech” (Nothofagus cunninghamii), i franges de l’eleocarpàcia Elaeocarpus grandis al llarg dels rierols que travessen boscos d’esterculiàcies del gènere Argyrodendron.
La segona situació és molt característica pel contrast que implica. De vegades les espècies pròpies de les selves plujoses formen un sotabosc en què hi ha associades espècies de tipus de vegetació molt més secs. Com a exemple d’aquest segon tipus de bosc en galeria es poden esmentar les extensions de terra endins de boscos caducifolis de lianes, que apareixen al costat de rierols que amb prou feines poden mantenir eucaliptus i casuarines, típics d’àrees més àrides. Aquestes franges de bosc poden tenir cobricels força alts (de fins més de 20 m d’alçada) amb una considerable diversitat d’espècies, incloent-hi palmeres.
Els boscos plujosos d’eucaliptus
L’enorme diversitat del gènere eucaliptus (Eucalyptus), amb més de 600 espècies, explica que calgui referir-s’hi tant en parlar de les zones desèrtiques australianes com en tractar les selves temperades. Alguns eucaliptus, en efecte, són arbres immensos i exigents pel que fa a la humitat, capaços de vertebrar comunitats forestals de caràcter selvàtic, i prosperen amb particular puixança a la zona sud-occidental.
Els boscos densos d’eucaliptus del SW australià
En aquestes selves del SW no es troben arbres de selves plujoses d’afinitats malèsiques ni antàrtiques. El bosc de “karri” (Eucalyptus diversicolor), un arbre alt i de creixement ràpid que pot assolir els 85 m d’alçada, està associat amb les localitats amb precipitacions més elevades. Aquests boscos tenen dos estrats, el superior amb l’espècie o les espècies dominants i un estrat molt més baix d’arbres petits i arbustos alts. Les lianes són escasses i els epífits inexistents. De vegades es poden arribar a formar boscos purs de “karri”, però aquesta espècie sovint és codominant amb altres eucaliptus; el bosc de “karri”, a més, també és transicional cap a altres tipus de boscos i arbredes. Els estrats arbustius poden ser molt diversos florísticament i presentar una àmplia gamma de gèneres i famílies, molts dels quals es troben igualment a les bosquines mediterrànies de les regions limítrofes més àrides. Probablement, els incendis naturals han estat sempre un factor fonamental en la dinàmica d’aquests espais forestals; com que el “karri” no es regenera bé després de grans incendis, sovint es provoquen focs controlats per reduir la brossa sense afectar aquests arbres, sobretot als llocs on es talen els boscos per extreure’n la fusta.
En emplaçaments calcaris de precipitacions lleugerament inferiors, la selva humida és dominada de vegades pel “tuart” (Eucalyptus gomphocephala), un altre gran eucaliptus que pot arribar a assolir els 40 m d’alçada. Aquesta selva forma una estreta franja costanera, a vegades en poblaments purs de “tuart” i altres en poblaments mixtos amb altres espècies d’eucaliptus. El “jarrah” (E. marginata), el típic eucaliptus mediterrani de l’àrea sud-occidental d’Austràlia, i el “marri” (E. calophylla) formen una associació característica en àrees on la precipitació anual és d’entre 650 i 1 300 mm, sobretot en sòls laterítics. Aquest bosc és complex, tant en la seva composició florística com en la seva relació amb altres tipus de vegetació. Tots dos arbres dominants poden aparèixer també en bosquines o formar boscos esclarissats de 10 a 15 m d’alçada en punts costaners.
Els boscos alts i oberts d’eucaliptus del SE australià
Es tracta dels més humits dels boscos oberts. Actualment, a Austràlia s’anomena bosc obert aquell que presenta recobriments d’entre un 30 i un 70%. Els més humits són també, lògicament, els més alts, i això fins al punt que els boscos oberts alts de la regió sud-oriental d’Austràlia inclouen els arbres més alts del món, eucaliptus que arriben a mesurar més de 100 m, com és el cas del freixe de muntanya de Tasmània (Eucalyptus regnans). El sotabosc, molt per sota del cobricel superior, en alguns casos encara té una alçada considerable, i sovint és molt exuberant i ric en espècies. A més, pot contenir una gran quantitat de falgueres arborescents (com per exemple Dicksonia i Cyathea).
La pluviositat elevada i regular és un determinant primari d’aquest tipus de bosc i de la seva distribució. Les característiques del sòl constitueixen factors secundaris importants, en particular un bon drenatge i uns nivells moderats de fòsfor, ja que als sòls rics en nutrients i més humits sempre s’hi fa un nombre major d’espècies. Els règims de temperatura dels boscos oberts alts varien molt, des de frescos a temperats i càlids, ja que abasten una gran varietat d’emplaçaments, des de Queensland fins a Tasmània. Aquests boscos són freqüents al llarg de les serres i planes de primera línia de costa, on el clima els és particularment favorable. En llocs més humits, sobretot on els sòls són rics en nutrients, les falgueres arborescents dominen els fons dels barrancs. La principal característica ambiental és que el règim de pluges és constant i les precipitacions moderadament altes. A més, per efecte orogràfic, la gran alçada del bosc madur pot fer incrementar localment la precipitació de manera efectiva, i ajudar el mateix bosc a mantenir-se.
Tot i això, en aquests boscos es produeix un procés natural d’aclarida, ja que en arribar a edats d’entre 25 i 40 anys la majoria dels arbres veuen frenat precoçment el seu creixement i moren. Així, el nombre d’exemplars, quan el bosc assoleix una edat d’uns 150 anys, pot haver quedat reduït d’una densitat de quasi 2 000 troncs/ha a menys de 80. Claps de bosc d’edat uniforme i carbó vegetal enterrat suggereixen una llarga història d’incendis a les regions forestals del SE d’Austràlia. Fos quin fos el règim de foc prehistòric a les parts més humides del continent, és probable que constituís un factor primordial en l’evolució dels boscos oberts alts.
Les selves temperades neozelandeses
La major part de les selves temperades neozelandeses són boscos mixtos que combinen coníferes i planifolis, encara que una part, sobretot a l’Illa del Sud, siguin fagedes de faigs australs. Els arbres forestals principals són, doncs, les grans coníferes, sobretot el “kauri” (Agathis australis), una araucariàcia que pot arribar a assolir longevitats considerables (fins a 1 700 anys), i podocarpàcies com el “rimu” (Dacrydium cupressinum), també força longeu (fins a 1 200 anys), el “kahikatea” (Dacrycarpus dacrydioides) i diferents “tōtara” (Podocarpus), els faigs australs (Nothofagus) i tot un seguit de laurifolis que pertanyen generalment a gèneres representats per poques espècies. Seria el cas del “pōhutukawa” (Metrosideros excelsa) i els “rātā” (Metrosideros), que són mirtàcies, del “kāmahi” (Weinmannia racemosa) i el “tawhero” (Weinmannia), de la família de les cunoniàcies, del “pukatea” (Laurelia novae-zelandiae), una monimiàcia, etc. Les formes de creixement dels arbres de Nova Zelanda són molt variades i abasten des d’arbres alts de tronc dret i únic, que poden excedir els 60 m d’alçada, com el “kauri” i el “kahikatea”, fins a arbres de troncs múltiples i forma arrodonida. Al límit altitudinal del bosc, el faig austral muntanyenc neozelandès (Nothofagus solanderi var. cliffortioides) de vegades s’agarriga i s’ajeu. No hi ha pròpiament arbres caducifolis, només algun semicaducifoli, la majoria de muntanya, cap dels quals no forma part del nucli d’espècies dominants. De fet, es pot dir que molt pocs arbres són parcialment caducifolis. L’esclerofil·lisme és comú i sovint les fulles són densament tomentoses per la cara inferior.
Les selves temperades de coníferes i planifolis
Presenten normalment dos estrats arboris, un de superior format per les grans coníferes i l’inferior integrat per arbres planifolis alts, i també s’hi pot trobar un tercer estrat d’arbres més petits. Arran de terra hi són presents herbes altes i falgueres, i les criptògames solen dominar un estrat muscinal de menys de 10 cm d’alçada. Sovint abunden també els epífits. La considerable variabilitat d’aquestes selves es correlaciona amb els canvis en la fertilitat del sòl, el pendent, l’exposició i l’altitud.
En els sòls menys fèrtils de les regions septentrionals de l’Illa del Nord, l’arbre dominant a l’estrat superior sol ser el “kauri” (Agathis australis), si bé avui dia aquest arbre es troba principalment confinat als cims i als altiplans elevats. En altres selves de terra baixa de l’Illa del Nord dominen podocarpàcies com ara el “rimu” (Dacrydium cupressinum), el “kahikatea” (Dacrycarpus dacrydioides), el “mataī” (Prumnopitys taxifolia) i el “tōtara” (Podocarpus totara). També són elements importants en aquests boscos mirtàcies com el “northern rātā” (Metrosideros robusta), cunoniàcies com el “kāmahi” (Weinmannia racemosa), proteàcies com el “rewarewa” (Knightia excelsa) o lauràcies com el “tawa” (Beilschmiedia tawa). Les selves de podocarpàcies i planifolis més grans de Nova Zelanda es troben a la part occidental de l’Illa del Sud. La més comuna és la de “rimu” i “kāmahi”, que es transforma gradualment en altres tipus de selva com ara selves mixtes de podocarpàcies, selves de “kahikatea” a les maresmes i selves muntanyenques de “southern rātā” (Metrosideros umbellata).
També és a la façana occidental de l’Illa del Sud, on hi ha una àmplia zona on no creixen faigs australs, que aquestes selves presenten el seu òptim. S’hi fan moltes de les espècies dels boscos de les terres baixes, però són florísticament més pobres. Els arbres dominants solen ser el “southern rātā” (Metrosideros umbellata) i el “kāmahi” (Weinmannia racemosa); entre les podocarpàcies s’hi troben també el “tōtara” (Podocarpus totara) i el “tōtara” de muntanya (P. hallii) i, als estrats inferiors, apareixen cupressàcies com el “kaikawaka” (Libocedrus bidwillii), grossulariàcies com Quintinia acutifolia, winteràcies com el “horopito” (Pseudowintera colorata), araliàcies com els Pseudopanax i falgueres arborescents. Asteràcies llenyoses de petit port arbori o de port arborescent dels gèneres Olearia i Brachyglottis, així com podocarpàcies més petites, apareixen a mesura que augmenta l’altitud.
Les selves temperades costaneres
Un altre tipus de selves temperades es fa al llarg dels estuaris protegits i dels braços de mar i a les parts més baixes de les valls costaneres, on dominen les influències marítimes. A l’extrem septentrional de Nova Zelanda hi predomina sovint el “pōhutukawa” (Metrosideros excelsa) i a la part meridional s’hi poden fer prominents les asteràcies arbòries. Aquestes selves costaneres de les regions septentrionals són molt variades i poden tenir com a arbres dominants proporcions variables de “pōhutukawa”, “kohekohe” (Dysoxylum spectabile), “karaka” (Corynocarpus laevigata), “taraire” (Beilschmiedia tarairi) i “tawa” (B. tawa). El “puka” (Meryta sinclairii) es troba localment a l’illa de Taranga, i el “kānuka” (Kunzea ericoides) i el “rewarewa” (Knightia excelsa) es troben sovint en llocs que han estat recentment alterats.
Les selves temperades de faigs australs
Es fan a la major part de Nova Zelanda i la seva composició varia des de la presència exclusiva de faig a l’estrat del cobricel fins a les variants mixtes, amb contribucions considerables de podocarpàcies, de planifolis o d’unes i altres. El rang altitudinal abasta des del nivell de la mar fins al límit altitudinal del bosc. A l’E de l’Illa del Sud les fagedes de faig austral muntanyenc neozelandès o “mountain beech” (Nothofagus solanderi var. cliffortioides) ocupen el límit altitudinal del bosc de manera pràcticament ininterrompuda al llarg de moltes de les serralades axials i dels Alps Meridionals. Aquestes selves muntanyenques de faigs australs són notables pel fet que sovint aquesta espècie es troba pràcticament sense cap acompanyant o només amb la presència escadussera d’alguna altra espècie.
A cotes més baixes, el faig austral dominant és el faig austral de terra baixa o “hard beech” (Nothofagus truncata) a les regions més septentrionals, mentre que el faig austral roig o “red beech” (N. fusca), el més alt i de creixement més ràpid de Nova Zelanda, ocupa les cotes més baixes, gairebé a qualsevol latitud tret de les més extremes. En les regions septentrional i nord-occidental de l’Illa del Sud domina sovint el faig austral argentat o “silver beech” (N. menziesii), mentre que a la part oriental d’aquesta illa només s’hi fan fagedes de faig austral muntanyenc (N. solanderi var. cliffortioides). Més al S, a les regions septentrionals i orientals de Fiordland, a la façana sud-occidental de l’Illa del Sud, el faig austral argentat domina aclapadorament a les valls excessivament plujoses, mentre que en sòls porosos sobre materials parentals pobres en nutrients s’hi poden trobar formes nanes d’aquest faig determinades genèticament. A la resta de la regió coexisteixen, en hàbitats diferents, les tres espècies.
Les selves de les illes perifèriques
A les illes Kermadec, al límit septentrional (29° 16’) de la regió botànica neozelandesa, domina una mirtàcia que està estretament emparentada amb la “pōhutukawa” (Metrosideros excelsa) de Nova Zelanda: és la “pōhutukawa” de Kermadec (M. kermadecensis). Aquest arbre coexisteix amb una gran varietat d’altres planifolis, amb falgueres arborescents i amb una palmera endèmica, la “nikau” de Kermadec (Rhopalostylis baueri var. cheesemanii). A les illes Chatham, plenament oceàniques —ja que són situades a 800 km a l’E de Nova Zelanda—, el bosc endèmic de la sapindàcia “akeake” (Dodonaea angustifolia), caracteritzat per la presència al sotabosc de l’asteràcia Olearia chathamica, havia dominat inicialment les àrees costaneres, si bé actualment la seva extensió s’ha vist dràsticament reduïda. Els boscos de “karaka” (Corynocarpus laevigata) s’estenien antigament pels sòls fèrtils de terra baixa, a l’illa de Chatham (Rēkohua) i l’illa de Pitt (Rangiauria), les més grans de l’arxipèlag. En aquestes dues illes i a les South-East, d’altra banda, les zones amb una pluviositat baixa i sòls orgànics foren originàriament dominades per boscos de “tarahinau” (Dracophyllum arboreum).
Les illes situades al S de Nova Zelanda posseeixen una gran varietat de comunitats forestals. A les petites illes Snares, la pressió dels nombrosos efectius de les poblacions d’ocells marins n’han alterat força la composició; hi predominen les margarides arbòries dels gèneres Brachyglottis i Olearia, que exclouen pràcticament la presència de qualsevol altra planta llenyosa. Més al S, a les illes Auckland (50° 44’), al llarg de les costes i les valls arrecerades, l’arbre dominant és el “southern rātā” (Metrosideros umbellata), al qual acompanyen l’epacridàcia Dracophyllum longifolium var. cockayneanum, la rubiàcia Coprosma foetidissima, l’araliàcia Pseudopanax simplex i la mirsinàcia Myrsine divaricata. Atesa la incapacitat dels sòls profunds i humits de les torberes que cobreixen la major part de les illes per suportar el pes dels troncs, els arbres tenen una inclinació característica. Encara més al S, a l’illa de Campbell (52° 33’ S), els boscos estan poc desenvolupats, i la planta de més alçària és l’epacridàcia Dracophyllum longifolium, que assoleix els 5 m d’alçada i habita en valls costaneres estretes i arrecerades.