La fotosíntesi té lloc gràcies a l’existència de pigments específics en els cloroplasts (bacterioclorofil·la i clorofil·les a, b, c, d i e com a pigments principals, i carotens i ficobilines com a accessoris, capaços d’absorbir l’energia i de transmetrela a la clorofil·la). Les molècules de pigment amb capacitat de transformar l’energia lumínica en energia química constitueixen els centres de reacció. Moltes molècules de pigment només transporten l’energia absorbida fins al centre de reacció i constitueixen una pantalla.
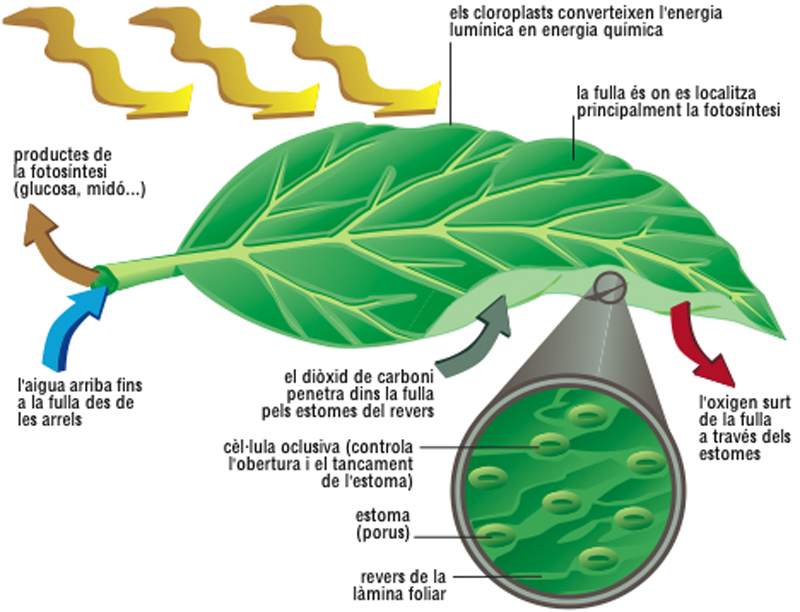
La fotosíntesi
El conjunt centre de reacció-pantalla és anomenat unitat fotosintètica, i en ell per cada molècula del centre de reacció n'hi ha unes 300 de pantalla. Cada unitat fotosintètica va associada a cadenes enzimàtiques que reben l’energia absorbida pel centre de reacció i la transformen en energia química; aquesta associació constitueix un sistema fotosintètic o fotosistema. L’equació general de la fotosíntesi en els vegetals superiors és
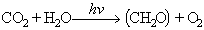
CO2 essent el diòxid de carboni, H2O l’aigua, (CH2O) la unitat dels glúcids i O2 l’oxigen. Als bacteris purpuris del sofre l’equació de la fotosíntesi és
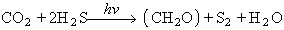
H2S essent l’àcid sulfhídric i S2 el sofre. Això significa que el O2 que es desprèn en els vegetals superiors prové del H2O i no del CO2, com hom havia pensat inicialment. Van Niel sintetitzà la fotosíntesi d’una manera global segons l’equació
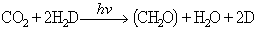
de manera que el CO2 és l’acceptor de H+, que es redueix a (CH2O), el H2D és el donador de H+, i D és la forma oxidada. Abans hom creia que la fotosíntesi comprenia dos processos: en el primer calia claror, mentre que el segon es devia donar a les fosques; però això no fou comprovat bioquímicament fins el 1937: Hill demostrà que en il·luminar cloroplasts aïllats d’espinacs es desprenia O2 en presència d’acceptors artificials d’electrons, i veié que no calia CO2 per al despreniment de O2. L’any 1950 Ochoa demostrà que l’acceptor natural d’electrons en la fotosíntesi és el NADP+ (nicotinamida-adenina-dinucleòtid-fosfat), segons l’equació
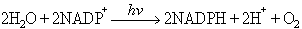
NADPH essent la forma reduïda del NADP+.
La utilització d’isòtops 18O incorporats al H2O i al CO2 fou una evidència més en el sentit de demostrar que el despreniment de O2 i la reducció del CO2 són independents.
Esquema de la fotosíntesi: salts d’energia (fletxes vermelles) gràcies als quanta de llum absorvits
El 1954 Arnon demostrà que si il·luminava cloroplasts aïllats se sintetitzava ATP (adenosina trifosfat) a partir d’ADP (adenosina difosfat) i PO4≡ (fosfat). Hom comprovà que aquesta reacció no depenia de la presència del CO2. El fet que la claror fos necessària per a sintetitzar ATP (molècula molt energètica) i NADPH (molècula amb un gran poder reductor) feu pensar que aquestes dues molècules eren les utilitzades en la fase fosca de la fotosíntesi.
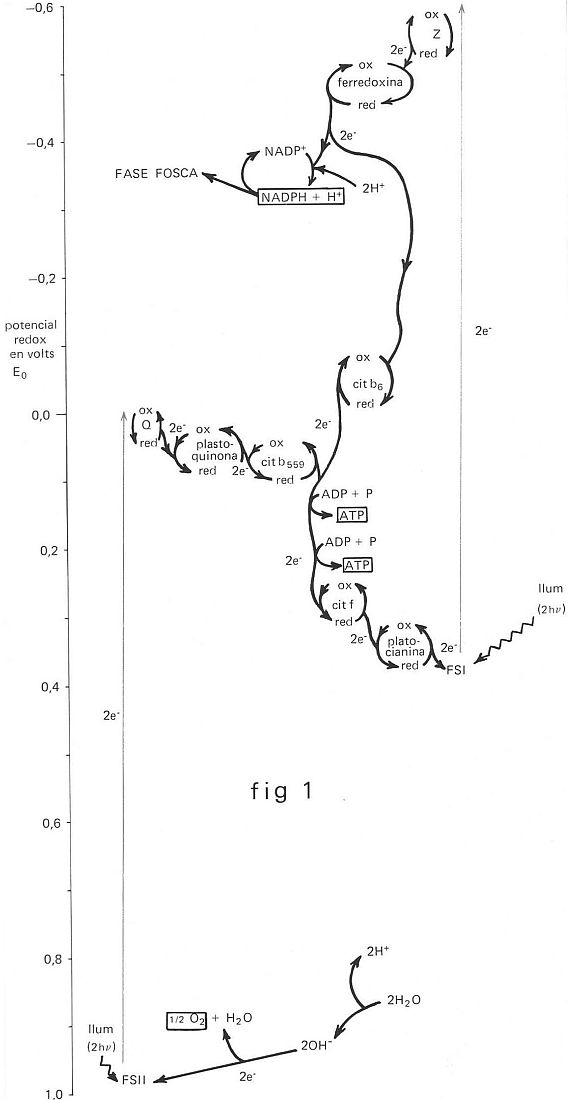
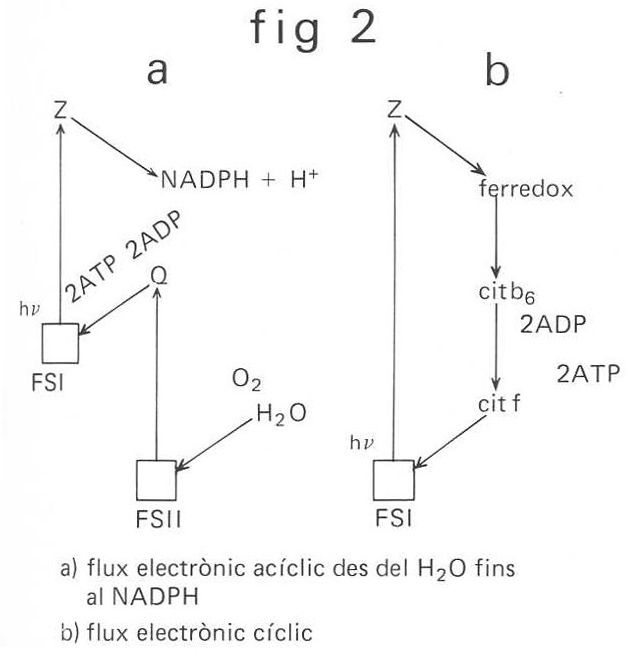
El flux electrònic induït per la llum en la reacció de Hill és acíclic, puix que els electrons passen del H2O, o d’un altre donador d’electrons, al NADP+, o a un altre acceptor d’electrons, a través de la clorofil·la. D’altra banda, la síntesi d’ATP, sense que s’acumuli cap substància reduïda, es deu a un flux cíclic d’electrons que comença amb l’excitació de la clorofil·la per la llum. L’existència de dos fluxos electrònics suggereix la idea de dues reaccions lumíniques en la fotosíntesi. Aquesta suposició es fonamenta en les diferències que hi ha entre la fotosíntesi dels vegetals superiors i la dels bacteris, i sobretot en l’existència de l'efecte Emerson, que consisteix en el fet que l’eficàcia quàntica, en il·luminar simultàniament amb llum de 690 nm i llum d’uns 650 nm, és superior a la suma de les eficàcies quàntiques obtingudes en il·luminar separadament amb aquestes longituds d’ona. Duysen postulà que hi ha una reacció lumínica en què no es desprèn O2; activada amb una excitació màxima a 700 nm i amb intervenció de la clorofil·la a i els carotens, la qual és atribuïda a l’anomenat fotosistema I (FS I). Afirmà també l’existència del fotosistema II (FS II), activat per longituds d’ona inferiors, amb despreniment de O2, el qual fotosistema posseeix associades la clorofil·la a i altres clorofil·les, ficobilines i xantofil·les.
Els vegetals superiors posseeixen tots dos fotosistemes, mentre que els bacteris només tenen el FS I. El conjunt d’aquests dos fotosistemes forma una unitat estructural al cloroplast, anomenada quantosoma. La llum excita la molècula de clorofil·la i aleshores els electrons poden transferir-se a una cadena de transportadors. L’esquema de transferència és el característic d’una reacció redox: els electrons agafats per la forma oxidada la converteixen en la forma reduïda de la mateixa molècula, que es reconverteix en la forma oxidada en cedir els electrons a una altra molècula oxidada d’un potencial redox més alt. En aquest procés, el dèficit electrònic del FS II és satisfet pel H2O, que forneix, amb la seva descomposició (fotòlisi), els electrons que proporciona l’hidroxil (OH-).
Esquema de la fotosíntesi: a) flux electrònic acíclic; b) flux electrònic cíclic
El dèficit electrònic del FS I és satisfet pels electrons que, excitats en el FS II, posen en funcionament la cadena redox (Q, plastoquinona, citocrom b 559, citocrom f, plastocianina) en l’anomenat flux acíclic, o bé els electrons que exciten el FS I posen en funcionament una altra cadena redox (Z, ferredoxina, citocrom b6, citocrom f, plastocianina), i es torna al FS I, en l’anomenat flux cíclic. En el flux acíclic es forma ATP i NADPH (el H+ —hidroxil— és proporcionat pel H2O en la fotòlisi), mentre que el flux cíclic proporciona només ATP.
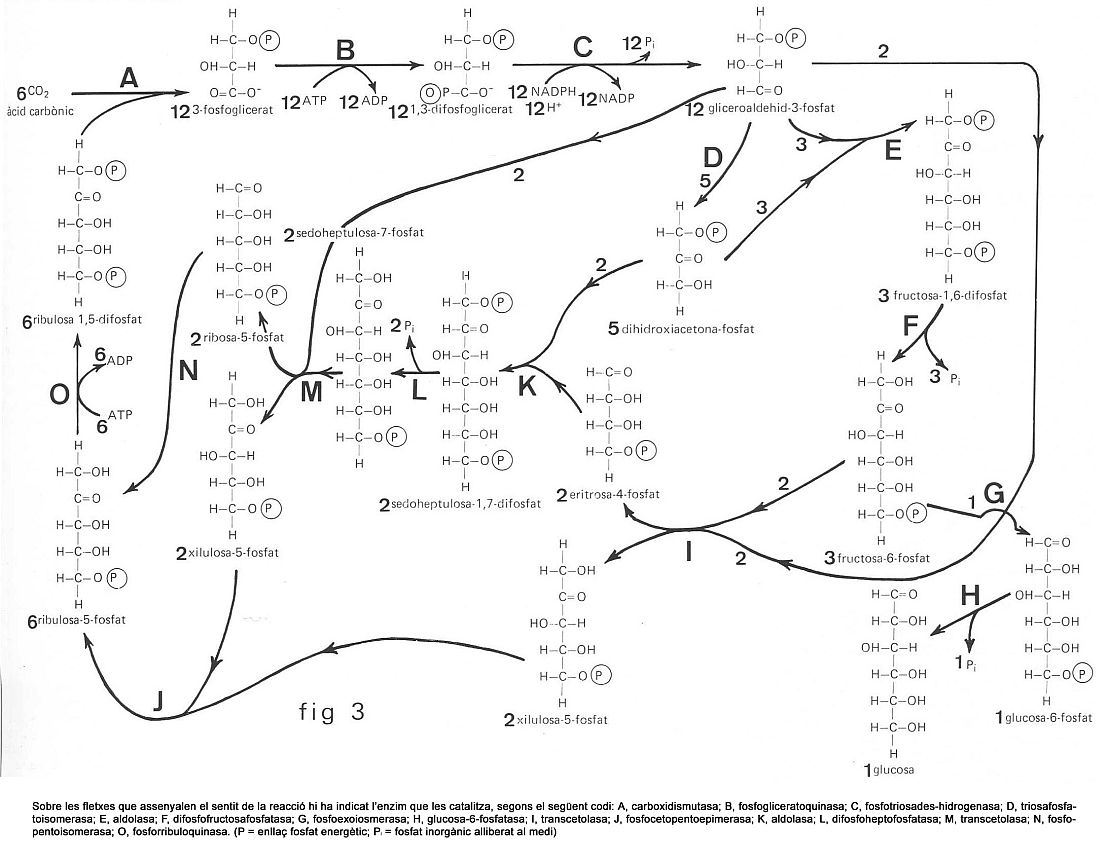
La fixació del CO2 i la posterior reducció a hidrats de carboni és un procés purament bioquímic i no necessita claror (reacció fosca). M.Calvin i els seus col·laboradors han esbrinat la complexitat d’aquest procés, en què al final hom obté hexosa, que passarà a midó o a uns altres productes de reserva. Cal fer notar que la ribulosa 1,5-difosfat, que incorpora el CO2 i forma dues molècules de 3-fosfoglicerat, es regenera completament; com a matèria nova hi ha només el CO2, que prové de l’atmosfera, puix que les molècules d’ATP i NADPH + H+ es formen en la fase lumínica. Com a producte final apareix la glucosa.
Per a establir el balanç i el rendiment de la fotosíntesi cal veure que per reduir sis molècules de CO2 i formar-ne una de glucosa (C6H12O6) són gastats divuit ATP i dotze NADPH + 12H+, o sigui, que la fixació i la reducció de CO2 a (CH2O) implica d’utilitzar tres ATP i dos NADPH + 2H+.
Esquema de la fotosíntesi: fixació del CO2 i reducció a glúcids
Si hom té en compte la hipòtesi dels dos fotosistemes acoblats, veu que calen quatre electrons en el FS I per a formar dos NADPH + H+ i quatre electrons més en el FS II per a formar una molècula de O2 i quatre ATP; tot plegat, calen vuit electrons, quantitat suficient, puix que hom obté un ATP més dels necessaris. En la síntesi de la glucosa (C6H12O6) a partir del CO2 i H2O, per a passar del CO2 al grup unitat dels glúcids (CH2O) calen tres quanta. En comparar tres quanta teòrics respecte als vuit quanta necessaris, el rendiment quàntic en la fotosíntesi és del 40%, aproximadament. En la natura el rendiment encara és més baix que el calculat per al procés molecular bàsic. Tant per als organismes aquàtics com per als terrestres, hom calcula que només és aprofitat el 2% de l’energia solar rebuda.