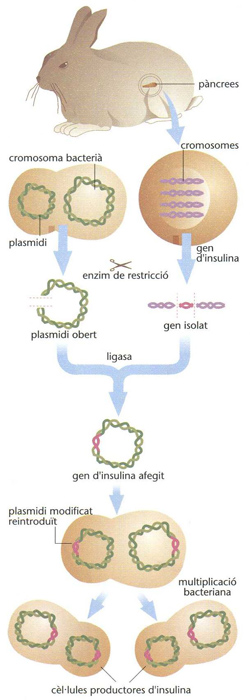
Pel fet que l’ADN és l’estructura responsable de la producció de proteïnes (transcripció, traducció) i, en general, de tota la producció cel·lular, l’enginyeria genètica es fonamenta en la introducció de nous patrons d’ADN dins el sistema genètic d’un organisme per tal de fer-li sintetitzar proteïnes que altrament no sintetitzaria. Per bé que a la pràctica de l’enginyeria genètica hom no ha exclòs els organismes superiors, és en els microorganismes unicel·lulars on més s’ha desenvolupat, i especialment en el bacteri Escherichia coli, pel fet d’ésser el més estudiat i conegut per la genètica bacteriana. E. coli presenta encara dos avantatges: el d’ésser procariota (és a dir, no té membrana nuclear) i el de contenir plasmidis (segments circulars d’ADN no integrats en el cromosoma bacterià, i que es dupliquen independentment del cromosoma). El procés emprat en aquests casos comença per l’extracció i purificació de l’ADN a implantar. Aquesta macromolècula és massa llarga per a ésser integrada sencera, i convé fragmentar-la bé per mitjans mecànics o per enzims de restricció (enzim de restricció), que tallen l’ADN per punts específics, i sovint deixen els fragments amb els extrems cohesius (és a dir, que es poden unir directament a altres segments, la qual cosa facilita la integració del fragment a l’ADN hoste). Com a vector del fragment, hom empra el factor F (o factor sexual), plasmidis (segments d’ADN de l’ordre d’1/100 de la talla del cromosoma) o bé virus bacteriòfags. Així, doncs, hom prepara el vector, que, en el cas del plasmidi, és extret, obert el seu cercle per un enzim de restricció, sense malmetre el sistema de duplicació i hom hi insereix el segment d’ADN aliè. Es tanca altre cop el cercle i és introduït novament al bacteri. Un cop dins el bacteri, la molècula d’ADN (hom l’anomena recombinant, híbrida o quimèrica si conté ADN provinent d’espècies diferents) es multiplica mercès al sistema de duplicació propi del vector. Hom pot acumular plasmidis en un bacteri pel fet que es dupliquen en presència de cloramfenicol, contràriament a l’ADN cromosòmic bacterià. En multiplicar-se l’organisme que conté els plasmidis, dóna lloc a un clon (hom anomena també aquest procés tècnica de clonació).
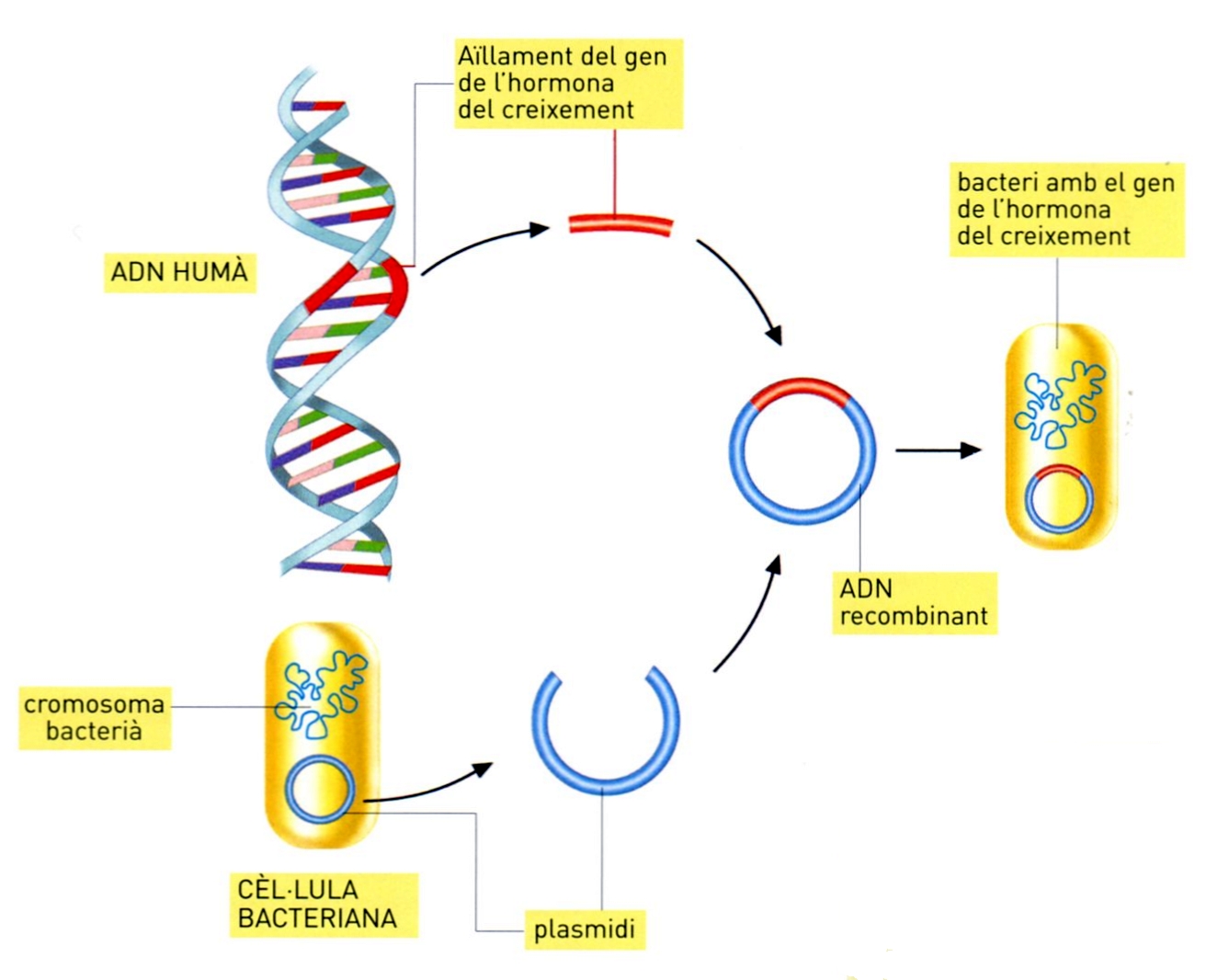
La producció de l’hormona del creixement
© fototeca.cat
Tot i que el fonament teòric semblava d’antuvi clar, pel fet que el codi genètic de l’ADN és universal, és a dir, el mateix per a tots els organismes vius, hom ha topat amb seriosos inconvenients pràctics, com ara l’expressió gènica, que necessita uns senyals d’iniciació (promotor) de la transcripció i la traducció adequats a l’organisme hoste, i que presenten diferències importants entre procariotes i eucariotes; alguns d’aquests problemes han estat enginyosament superats, per bé que en molts casos no s’ateny encara el nivell de producció proteica esperat. Amb tot i això, d’ençà del 1971, en què Paul Berg inicià aquestes pràctiques, una gran quantitat de genetistes moleculars d’arreu començaren d’emprendre treballs en aquest sentit, per tal com s’adonaren de les possibilitats revolucionàries que aquest camp oferia, i tot seguit es desenvoluparen diverses línies de treball tendents a perfeccionar i a diversificar els mètodes d’extracció i síntesi d’ADN, i de la seva integració i expressió genètica. La producció de proteïnes fou iniciada per l’aconseguiment d’una hormona de l’hipotàlem, la somatostatina humana (de 14 aminoàcids), i continuada cronològicament per la proinsulina de rata (82 aminoàcids), l’ovoalbúmina (381 aminoàcids), la dihidrofolat reductasa, l’hormona de creixement de rata, la insulina humana, l’antigen HBc del virus de l’hepatitis, l’hormona de creixement humana i l'interferó. En l’esforç d’integrar ADN en eucariotes, hom ha reeixit a incloure gens de conill en cèl·lules renals cultivades, i hom preveu que en el futur s’aconseguirà la introducció de gens isolats d’organismes superiors, en cèl·lules en cultiu del mateix organisme, i per mitjà de virus. Hom treballa també en la inclusió dels gens per a la fixació del nitrogen en ADN de bacteris associats a les arrels d’altres plantes de les lleguminoses, o bé directament dins els cromosomes de les plantes de conreu, amb la qual cosa deixarien d’ésser dependents dels adobs nitrogenats. Les aplicacions a la medicina inclouen l’obtenció d’hormones, d’enzims, de certs vaccins i d’interferons. La indústria ha promogut un ràpid increment de l’activitat en aquest camp: als EUA hi ha prop de 20 societats constituïdes per a la recerca i explotació dels productes de l’enginyeria genètica, i d’altres empreses al Japó, a la Gran Bretanya i a la República Federal d’Alemanya. Entre les recerques iniciades es compten les de la producció d’alcohol a baix preu amb finalitats de combustible (gasohol); de colorants, detergents, matèries plàstiques, cautxú sintètic, plaguicides, etc. Hom ha reeixit a modificar un bacteri del gènere Pseudomonas i dotar-lo de capacitat per a descompondre hidrocarburs (amb aplicació en la solució de marees negres). Amb tot, ja d’un bon començament, una part de l’opinió científica es manifestà repetidament hostil a aquestes activitats de recerca, i el 1974 s’aturaren per la pressió d’alguns científics temorencs de la perillositat que comportaven. El 1975 es reuniren a Asilomar (Califòrnia) 140 investigadors de 17 països i establiren una normativa de seguretat per tal de continuar la recerca. A partir d’aquell moment, les restriccions s’anaren atenuant, un cop demostrat que els perills no eren tan greus com alguns havien suposat.