
Els mol·luscs són animals de cos tou i musculós, sovint protegit per una conquilla calcària. Es tracta d’un gran llinatge separat molt aviat de la resta de metazous, el qual ha experimentat una diferenciació intensa quant a morfologia, fisiologia i ecologia, donant lloc a una enorme diversitat En aquesta fotografia veiem un exemplar d’Atlanta peroni. un cargol caçador que forma part del plàncton marí. L’adaptació al medi pelàgic ha comportat l’alleugiment de la conquilla, l’expansió del peu per a formar una aleta, i un increment notable en els òrgans sensorials.
Claude Carré
L’embrancament dels mol·luscs forma un grup immens, probablement amb més de 100 000 espècies vivents. N’hi ha en gairebé tots els ambients on la vida a la Terra és possible, i sovint assoleixen una importància ecològica cabdal. La varietat de formes, mides, tipus de vida i cicles vitals és sorprenent. El registre fòssil dels mol·luscs es remunta als orígens dels animals multicel·lulars, i afegeix dimensions insospitades a la diversitat actual. De fet, l’estudi dels mol·luscs, la malacologia, ens obliga a reconsiderar críticament nocions com ara l’escala evolutiva, la rigidesa dels plans estructurals, o la divisió, sigui amb la intenció que sigui, dels metazous en vertebrats i invertebrats. Des de la nostra perspectiva de «nomol·luscs», i potser acostumats a forçar la realitat quasi inabastable de la vida d’aquest planeta, encabint-la en esquemes classificatoris simplificats, no és fàcil de trobar una definició senzilla del que és un mol·lusc. Per tal d’evitar l’artificiosa discussió de tipus generalitzats o avantpassats hipotètics, és necessari examinar amb cert detall els diferents sistemes orgànics, i així poder inferir homologies, establir relacions filogenètiques, i demostrar la unitat bàsica del fílum.
Fora de l’àmbit teòric, la nostra civilització deu bastant als mol·luscs: l’existència de malacòlegs es deu en bona part al consum alimentari de musclos, ostres, cloïsses, cargols i calamars. Però hi ha molts altres motius d’interès per a l’estudi dels mol·luscs. La gravetat de diverses malalties, de gran dispersió i prevalença, transmeses per mol·luscs obliga a discernir la identitat i ecologia dels temuts vectors. La necessitat de mantenir ecosistemes funcionals en un medi global cada cop més humanitzat empeny a intentar descobrir les relacions de moltes espècies de mol·luscs amb llurs ambients. L’abundància de mol·luscs fòssils ens permet una lectura detallada dels estrats geològics.
Morfologia
Els sistemes esquelètics i els moviments
En el cos d’un mol·lusc podem trobar tres tipus de suport mecànic. La majoria d’espècies presenten algun tipus de conquilla, un exosquelet calcari i quitinós al qual l’animal està adherit mitjançant músculs potents. Aquesta musculatura s’estén per bona part del cos, formant un suport per a ella mateixa, que faculta així moviments complexos i altament coordinats. Finalment, hi ha cavitats internes que poden funcionar com un esquelet hidrostàtic. La diversitat morfològica del fílum és en bona part el resultat de combinacions d’aquests tres esquelets, cadascun amb una àmplia gamma de possibilitats de formes i disposicions.
La conquilla

La conquilla dels mol·luscs és un exosquelet calcari que protegeix els òrgans interns i contribueix a donar suport al cos. El creixement de la conquilla acompanya el de l’animal, de manera que és un veritable registre del desenvolupament individual. L’enrotllament característic de les conquilles respon a un disseny òptim entre les necessitats d’economitzar materials, maximitzar la resistència i aconseguir una forma global adequada al medi. Aquí veiem una conquilla de cargol bover tallada per a mostrar-ne l’estructura espiral.
Jacques Six / Firo-Foto
L’observació de qualsevol cargol o bivalve permet de descobrir una expansió laminar del cos que recobreix la superfície interna de la conquilla i embolcalla altres òrgans: aquesta estructura rep el nom de mantell. Es tracta d’un òrgan epitelial, sovint amb elements musculars i connectius, el marge del qual conté abundants cèl·lules secretores de carbonat càlcic. Aquesta substància cristal·litza a l’exterior del cos, en general en forma d’aragonita, però de vegades també com a calcita; en els solenogastres, caudofoveats i poliplacòfors, i també en alguns bivalves, la cristal·lització s’inicia intracel·lularment. El mantell també secreta una substància quitinosa de composició complexa, la conquiolina, la qual es diposita sobre la superfície externa de la capa calcària, i forma un estrat orgànic anomenat periòstrac; aquest segon nivell és essencial per a evitar la dissolució de la conquilla en ambients àcids. En general el mantell també produeix pigments de tota la gamma de colors, que generen l’existència en el periòstrac de tota mena de patrons cromàtics. El nombre, gruix i estructura cristal·lina de les capes de la conquilla varia considerablement. La deposició de material nou té lloc al marge ja format, de manera que la conquilla creix més o menys isomètricament, com un con o tub en expansió.
La conquilla d’un cargol terrestre típic consta d’una sola peça, però en molts altres gasteròpodes hi ha una segona estructura, l’opercle, que serveix de porta protectora. Els bivalves tenen, és clar, dues valves, però poden tenir una colla de peces accessòries de formes sorprenents. Hi ha una família de gasteròpodes opistobranquis l’opercle embrionari dels quals sofreix una hipertròfia tal, que l’adult es troba protegit per una conquilla efectivament bivalva. Els cefalòpodes amb conquilla calcària presenten aquesta estructura dividida internament en cambres plenes de gas. Els escafòpodes tenen una conquilla tubular, en forma d’ullal d’elefant, oberta pels dos extrems. Els poliplacòfors són recoberts per vuit plaques articulades i vorejades per un cinyell d’espines; tant les plaques com les espícules són calcàries i produïdes pel mantell. En els caudofoveats i els solenogastres només hi ha espícules.
La increïble varietat de formes i colors de les conquilles ha captivat de sempre l’home. Per al fabricant, i habitant alhora, d’una conquilla, tota l’estètica és un accident de la interacció de les limitacions de disseny que imposen les lleis físiques, de la tendència a l’economia de materials que suposin una despesa energètica, de pressions selectives com són la predació o l’estabilització en un determinat substrat, i de l’herència de models de desenvolupament. És possible de deduir molt del mode de vida d’un mol·lusc a partir de la forma i detalls estructurals del seu exosquelet; això és particularment important per a la comprensió del registre fòssil dels mol·luscs, riquíssim precisament gràcies a la composició química de llurs conquilles. Ara bé, hi ha molts mol·luscs que no tenen una conquilla prou gran per a servir de defensa, o fins i tot que no en tenen de cap mena. El cas de quasi tots els cefalòpodes vivents (els coleoides) és il·lustratiu: les sèpies i els calamars tenen conquilles internes, més o menys importants en termes funcionals, però en els pops la conquilla arriba a mancar completament.
La musculatura i els esquelets hidrostàtics
Els moviments dels tentacles dels cefalòpodes i l’expansió del cos d’un cargol fora de la conquilla són dos exemples de la importància d’una musculatura tan complexa en el seu funcionament: la tensió d’un grup de músculs es complementa amb la contracció d’uns altres per a modificar la forma del conjunt (els moviments de la llengua humana són el resultat del mateix principi). Dins els gasteròpodes, trobem una sèrie d’espècies amb la conquilla reduïda en grau divers, en les quals la musculatura assumeix les funcions de l’exosquelet, incomplet o absent: hi ha llimacs amb conquilles estranyament petites, d’altres amb vestigis interns, i fins i tot d’altres que no en tenen ni rastre, i en tots ells la forma del cos respon a la consistència i contraccions d’un peu molt extens.
Sovint l’acció dels músculs es fa efectiva mitjançant el canviament de la forma d’una cavitat plena de líquid. Aquest principi mecànic és present en els caudofoveats i alguns solenogastres, en l’espai entre el mantell i la faringe, ocupat per teixit mesequimatós; en els bivalves, el si hemocèlic principal contribueix decisivament als moviments del peu.
Els diferents tipus de suport per als músculs permeten el control dels moviments i, per tant, el desplaçament de l’animal, amb mecanismes molt variats. Només en els solenogastres la locomoció es realitza per mitjà dels cilis de la superfície ventral, encara que poden tenir un paper important en qualsevol mol·lusc de dimensions reduïdes.
Els caudofoveats i alguns solenogastres s’enterren en el sediment mitjançant moviments peristàltics de l’extrem anterior del cos: un eixamplament representa un ancoratge efectiu, mentre que una elongació facilita la penetració. Aquest desplaçament és conseqüència de la interacció de la musculatura longitudinal i l’esquelet hidrostàtic. Els bivalves que viuen enterrats en el sediment es regeixen pel mateix principi, encara que en aquest cas el sistema muscular i l’esquelet hidrostàtic són força més complexos.
Una de les característiques més remarcables de cargols i llimacs és el rastre lluent que deixen per allà on passen. Si s’observa el mateix animal desplaçant-se sobre un vidre, és fàcil de detectar ones rítmiques de contracció que recorren el peu; aquest moviment és possible gràcies a la complexa estructura de la massa muscular en contacte amb el substrat. L’experiment no s’acaba si es posa el vidre en posició vertical, o fins i tot si s’inverteix: això demostra que hi ha algun factor addicional. En efecte, el mucus que l’epidermis dels mol·luscs secreta copiosament té propietats mecàniques extraordinàries: en recollir-lo amb els dits hom pot adonar-se que és un líquid viscos, i així es comporta sota qualsevol zona del peu que estigui separada del substrat; però quan el mucus és comprimit entre el peu i el substrat (o entre els nostres dits), la secreció es converteix en un sòlid elàstic, que evita el lliscament i funciona com un adhesiu. Així doncs, cada ona locomotriu passa acompanyada de transformacions viscoelàstiques. Hi ha un límit a la tensió que el mucus pot suportar abans de deixatar-se, i això podria determinar màxims absoluts de mida i velocitat per als organismes que tenen aquest mètode de desplaçament.
El peu dels mol·luscs pot tenir formes molt diferents, de manera que no sempre s’efectua el moviment per la progressió d’ones de contracció. Els pops acostumen a caminar sobre el fons marí amb l’elegància de les seves vuit extremitats coordinades, i ajudats per les ventoses, que els proporcionen un ancoratge efectiu. Alguns bivalves i gasteròpodes salten usant el peu, musculós i llarg, com a perxa.
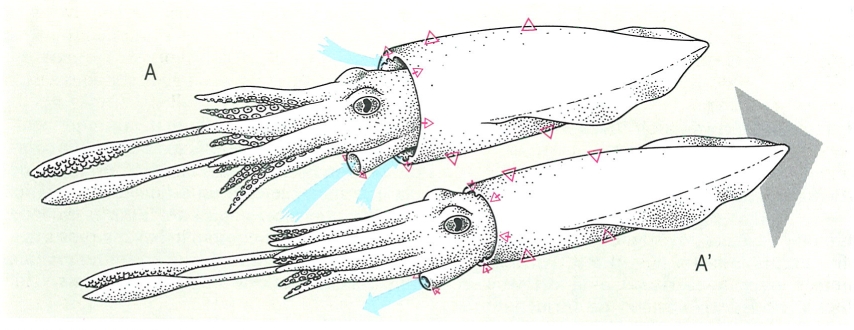
El sistema de propulsió dels cefalòpodes coleoides és un cas especial de desplaçament dins el grup dels mol·luscs. Comporta el bombament alternat, en sentits oposats, d’aigua a la cavitat pal·liat (A). Els cartílags sifonals ancoren el mantell durant la contracció, el celoma esmorteix els canvis de pressió damunt les vísceres, el sifó dirigeix el líquid efluent, i el cos sencer es desplaça en sentit oposat a aquest flux (A’)
Biopunt, original de l’autor
El mantell dels coleoides és una mena de bossa essencialment muscular, escassament unida a la resta del cos, que l’envolta quasi del tot. La contracció dels músculs circulars del mantell força l’aigua que hi havia a dins a sortir per la boca de la bossa, i l’animal es desplaça, per reacció, en sentit oposat; la sortida de l’aigua té lloc a través del sifó, que és alhora una mànega per a dirigir el flux i una vàlvula per a controlar-ne la pressió. L’acció de músculs antagonistes expandeix el mantell i permet l’entrada d’aigua per a recomençar el cicle. Els nautiloides, en tenir la superfície externa formada per una conquilla rígida, retreuen la massa cefàlica per a generar el flux a pressió. Els pectínids i els límids són bivalves capaços de nedar amb gràcia considerable a base d’expel·lir aigua a pressió pel sifó exhalant, el qual té la mateixa funció en aquests animals que en els coleoides.
Cal recordar els nombrosos mol·luscs nedadors, els quals fan servir expansions del cos com a aletes. N’hi ha amb habilitats més aviat escasses, com és el cas d’alguns opistobranquis, normalment bentònics, que baten les vores del peu com si fossin ales, però hi ha gasteròpodes planctònics que mostren dots sorprenents en llurs delicats moviments. Les sèpies i els calamars tenen un parell d’aletes laterals al mantell, els moviments ondulatoris de les quals permeten maniobres exactes en totes direccions.
Els cefalòpodes que tenen una conquilla amb cambres internes poden regular la quantitat de gas que aquestes contenen; això possibili ta migracions verticals importants amb una mínima despesa energètica directa. Entre els cargols d’aigua dolça, n’hi ha que poden contraure el pulmó, la qual cosa permet de modificar el volum de gas dins el cos, i generar moviments verticals a l’aigua.
La cavitat pal·lial

La respiració s’efectua en la majoria de mol·luscs a través d’una o dues brànquies de tipus característic, anomenades ctenidis. les quals s’allotgen a l’interior de la cavitat pal·lial. En casos excepcionals els ctenidis són visibles exteriorment, com és el cas d’aquest opistobranqui, en el qual l’únic ctenidi és força ramificat.
Josep M. Gili
El mantell (pallium) engloba part del cos en tots els mol·luscs, i la cavitat que així defineix conté els òrgans respiratoris. En els poliplacòfors i triblídides, la cavitat pal·lial és només l’estret espai entre el peu i el mantell que el circumda, essent elevat el nombre de brànquies. L’estructura d’aquestes brànquies és molt característica, amb aspecte de pinta amb dents eixamplades: el ctenidi és, doncs, una bateria de làmines la funció de les quals és incrementar la superfície disponible per a l’intercanvi de gasos; a l’interior de cada làmina flueix la sang, que va des d’un conducte aferent general i perpendicular a les làmines fins a un altre conducte, d’estructura semblant a la del primer, però on la sang circula en sentit oposat. La superfície del ctenidi és ocupada per cèl·lules secretores de mucus i d’altres proveïdes de cilis, el moviment dels quals genera els corrents necessaris per a la renovació del medi immediat. La sang i l’aigua flueixen en sentits oposats, per un sistema de contracorrent que fa més eficient la respiració.
Els gasteròpodes tenen la cavitat pal·lial orientada cap endavant, de manera que el cap i el peu s’hi poden retreure, almenys en les espècies amb una conquilla funcional. Algunes espècies marines tenen un parell de ctenidis, però quasi sempre presenten una sola branquia. En els opistobranquis el ctenidi acostuma a ésser força ramificat, i sobresurt a l’exterior com un plomall. Els cargols i llimacs terrestres, i també molts dels que viuen a les aigües dolces, no podrien respirar amb brànquies, perquè la manca de rigidesa i l’adhesió entre les làmines (per tensió superficial) en col·lapsaria la delicada arquitectura; aquests mol·luscs tenen la superfície de la cavitat pal·lial profusament irrigada i l’intercanvi gasós es realitza a través de l’epiteli, de manera que la cavitat funciona com un pulmó. Alguns cargols i solenogastres tenen brànquies que són expansions laminars de la paret del mantell i que, per tant, no es poden considerar homòlogues als ctenidis.
Els caudofoveats i solenogastres tenen la cavitat pal·lial limitada a l’extrem posterior; només els primers tenen un parell de ctenidis, encara que alguns solenogastres tenen dues brànquies d’estructura diferent. Els bivalves no presenten cefalització, i la cavitat pal·lial envolta quasi tot el cos; hi ha només dos ctenidis, però en general són de grans dimensions, perquè la producció de mucus i els corrents ciliars s’aprofiten per a recollir partícules alimentàries de l’aigua. Les brànquies dels septibranquis, un grup de bivalves de les grans profunditats, grosses, robustes i perforades, funcionen com una nansa per a atrapar els animals de què s’alimenten. Alguns cargols aquàtics també usen el sistema filtrador, de manera exclusiva o com a complement de llur alimentació. Els escafòpodes no tenen brànquies, però sí bandes ciliars que funcionen de manera semblant als ctenidis dels bivalves típics.
Els ctenidis dels cefalòpodes són grans i ramificats, i reben sang bombada des de cors accessoris situats a llur base. La ventilació de la cavitat pal·lial s’efectua mitjançant el mecanisme de propulsió ja descrit i, en els nautiloides, també amb el moviment de la part interna del sifó.
L’organització interna

La concentració d’òrgans sensorials a la part anterior del cos és característica dels mol·luscs que no menen una vida sedentària en l’estat adult. Aquesta cefalització és clarament visible en els mol·luscs caçadors o carronyaires. Moltes espècies de cargols marins, com ara Hinia incrassata, un buccínid, presenten un sifó, una extensió tubular del mantell a l’extrem anterior, la qual permet de discernir la direcció d’on provenen els estímuls químics.
Joan M. Benavent
La musculatura no incorporada a la massa del peu, el cap o el mantell forma tres sistemes bàsics: l’un just sota el mantell, l’altre al llarg dels marges laterals, i finalment una sèrie de parells de feixos dorso-ventrals. El primer és important només en solenogastres i caudofoveats, tot i que es troba també en els poliplacòfors; el segon sistema muscular, característic d’aquest darrer grup (encara que també es troba en alguns caudofoveats, els solenogastres i, modificat, en els triblídides), permet l’enrotllament del eos; el sistema de feixos dorsiventrals es limita a l’extrem anterior en els caudofoveats, és seriat però poc definit en els solenogastres, seriat i definit en els poliplacòfors i els triblídides, i limitat a pocs elements grossos i de funció fonamental en la resta de les classes.
El sistema alimentari de quasi tots els mol·luscs conté una ràdula, una cinta formada per fileres repetides de denticles quitinosos. La forma i disposició dels denticles es relaciona amb el tipus d’alimentació, atès que l’aliment és raspat i dut a la boca en cicles d’extensió i retracció del conjunt muscular de la faringe. El tracte alimentari dels solenogastres és simple i recte, amb constriccions que corresponen als músculs dorso-ventrals. Els caudofoveats tenen una ràdula peculiar i presenten un enorme cec digestiu com a única complicació. La resta de mol·luscs tenen com a mínim una mandíbula quitinosa i extensions labials a la boca, dos parells de glàndules digestives (anteriors i medials), un estómac més o menys complex, i un intestí llarg i enrotllat. L’estructura interna de l’estómac i la divisió d’aquest en cambres té relació amb el tipus d’alimentació: en els carnívors és molt més senzill que en els herbívors. El sistema digestiu és pràcticament obsolet en aquells mol·luscs que viuen com a paràsits interns en l’estat adult, i en els que contenen endosimbionts.

El grau de complexitat dels ulls en els mol·luscs és molt variable: poden ésser des de simples agrupaments de cèl·lules sensibles a la llum, fins a formar cambres tancades dotades d’una lent i una retina. la posició i el nombre d’ulls varia àmpliament: n’hi ha al marge del mantell d’alguns bivalves, en petits solcs de les valves dels poliplacòfors, en el sifó de diversos cargols marins, etc. els gasteròpodes i cefalòpodes tenen un parell d’ulls cefàlies, de complexitat molt variable. en els pulmonats terrestres, com aquest cargol bover, són a l’extrem d’uns tentacles llargs i retràctils: el segon parell de tentacles, proper a la boca, és de funció fonamentalment tàctil.
Jacques Six / Firo-Foto
El sistema nerviós dels mol·luscs és format bàsicament per quatre cordons longitudinals, amb les parts principals protegides per una túnica prima de teixit connectiu. Encara que la disposició de ganglis i nervis és molt variada, sempre hi ha un anell nerviós entorn de la part anterior del tub alimentari, del qual parteixen tres parells de nervis cerebrals, que innerven la regió perioral, l’àrea ventrofrontal, i la zona faríngia. La resta del cos és innervada pels cordons longitudinals. En molts cargols els cordons nerviosos posteriors a l’anell periesofàgic es creuen; en altres gasteròpodes aquest anell pot ésser més o menys desenvolupat, i arriba a ésser massiu en els pulmonats. El grau màxim de concentració nerviosa s’observa en els cefalòpodes, on es pot parlar d’un autèntic cervell, protegit per un crani cartilaginós. Els tentacles dels cefalòpodes reben innervació cerebral directament. La cavitat pal·lial de quasi tots els mol·luscs conté un òrgan quemoceptor anomenat osfradi, sempre innervat de manera semblant, encara que la seva forma i situació varia considerablement. En els caudofoveats hi ha un parell d’osfradis com a únics òrgans sensorials. Els solenogastres tenen un osfradi modificat, l’òrgan dorsoterminal, i l’òrgan atrial, associat a la boca. La resta de mol·luscs tenen un quemoceptor anomenat òrgan subradular, atès que sovint és associat a la ràdula; els coleoides tenen, a més, papil·les o fossetes olfactòries al cap. La majoria d’opistobranquis tenen un parell d’apèndixs cefàlies anomenats rinòfors. L’anell nerviós periesofàgic és connectat a un parell de petits sacs, els estatocists, que contenen concrecions calcàries i permeten el sentit de l’equilibri. Hi ha estatocists en tots els mol·luscs, excepte en els caudofoveats i els poliplacòfors. Gairebé tots els mol·luscs, excepte els caudofoveats i els solenogastres són sensibles a la llum. L’organització, nombre i disposició de fotoceptors varia molt d’un grup a l’altre, de manera que és improbable que en tots els casos es tracti d’estructures estrictament homòlogues.
L’excreció de metabòlits es realitza a través de la paret del tracte digestiu en els solenogastres i els caudofoveats, mentre que en la resta dels mol·luscs travessa la paret de l’aurícula vers el líquid pericàrdic; també té lloc per ultrafiltració en un òrgan format pel conducte del pericardi, amb plecs o diverticles laterals. Cal destacar que l’òrgan excretor dels mol·luscs, encara que anomenat ronyó, no apareix en cap altre fílum.
El sistema circulatori dels mol·luscs consta d’un cor sistèmic format per dues aurícules (excepte el de la majoria de gasteròpodes, que només en té una), i un ventricle, i els vasos i hemocels associats; aquests sins poden ésser poc extensos, com ara en els cargols terrestres, o molt amples, com s’observa en els bivalves excavadors. Els poliplacòfors, els triblídides i els nautiloides transporten la sang aferent vers el cor a través de sins particularment allargats. El cor sistèmic es troba embolicat pel pericardi, la funció del qual sembla que és la de generar la pressió negativa necessària per al reompliment de les aurícules. Hi ha dues excepcions a aquest model: d’una banda, el cor dels escafòpodes és molt reduït; de l’altra, el dels coleoides, que és un sistema tancat amb cors secundaris. És important de destacar que el ventricle d’aquests cors bronquials és probablement homòleg a l’aurícula d’altres mol·luscs, mentre que les aurícules dels coleoides són eixamplaments secundaris dels vasos veïns.
La reproducció i el desenvolupament

Els ous dels mol·luscs són de mida, forma i nombre molt variats. En el cas dels murícids, les femelles poden arribar a pondre més d’un centenar de càpsules ovígeres en forma de copa plena. Cada càpsula conté una munió d’ous, entre els quals n’hi ha que no són fèrtils i proveeixen d’aliment als embrions.
Xavier Safont / M. Alba Camprubí
La reproducció dels mol·luscs és exclusivament sexual; pot ésser en individus de sexes separats o bé en hermafrodites i, en aquest cas, simultanis o consecutius, amb capacitat d’autofecundació o sense. En general hi ha un parell de gònades, encara que poden ésser més o menys fusionades. La fertilització pot ésser externa o interna, sovint mitjançant espermatòfors(sacs acel·lulars plens de gàmetes masculins) d’origen divers; si és externa, el sistema és senzill: les gònades dels caudofoveats i solenogastres buiden llurs productes en el pericardi, mentre que els bivalves, els escafòpodes, els poliplacòfors i els triblídides obren els gonoductes directament a l’exterior. El sistema reproductor dels gasteròpodes és força complicat: en l’aparell masculí hi ha cambres i túnels destinats a emmagatzemar gàmetes, sovint altres cambres on es generen els espermatòfors, i en moltes espècies existeix un òrgan eversible i intromitent, que no sempre es desenvolupa de la mateixa manera; l’aparell femení inclou receptacles per a rebre els productes masculins, digerir part del material rebut, emmagatzemar espermatozoides, i formar una gran varietat de cobertes per als ous. Els coleoides mascles tenen un braç modificat, l’hectocòtil, destinat a transferir espermatòfors.
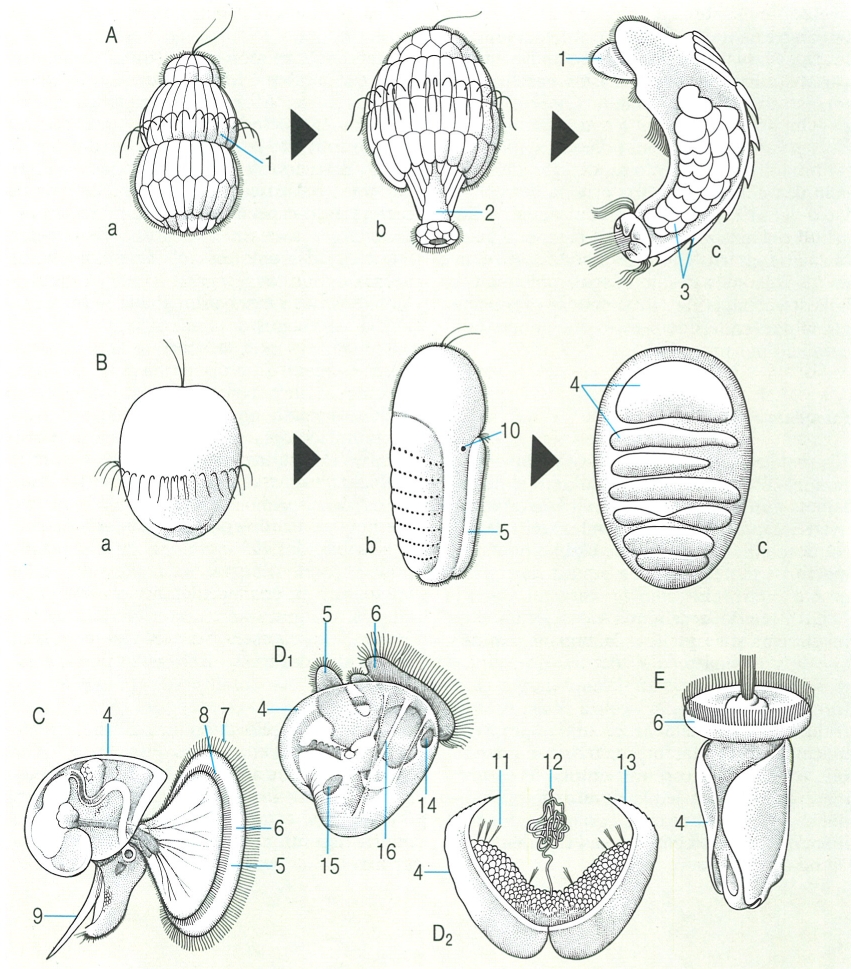
Formes larvals dels mol·luscs: A solenogastres, B poliplacòfors, C gasteròpodes, D bivalves, E escafòpodes. L’estadi de pericalimma (a) és absent del desenvolupament dels mol·luscs conquífers. La larva que se’n deriva per elongació del cos imaginal en solenogastres i poliplacòfors (b) acaba presentant una organització semblant en ambdós grups (c). Les larves de tipus vèliger (C) o pedivèliger (D1 i E) són característiques dels conquífers. Una excepció notable és la dels bivalves unionoïdeus, les larves gloquidi (D2) dels quals han de fixar-se a la superfície d’un peix o amfibi, on viuen com a paràsits fins a completar-hi la metamorfosi. Hom ha indicat en el dibuix: 1 calimma, 2 cos imaginal, 3 cossos calcaris, 4 conquilla, 5 peu, 6 vèlum, 7 prototroca, 8 metatroca, 9 opercle, 10 ocel, 11 cilis sensorials, 12 filament adhesiu, 13 ganxos, 14 múscul adductor anterior, 15 múscul adductor posterior, 16 múscul retractor del vel.
Biopunt, a partir de fonts diverses
El zigot dels mol·luscs sofreix divisions espirals, el blastòpor dóna lloc a l’estomodeu i l’anus apareix com un orifici nou. En els solenogastres es desenvolupa una larva del tipus pericalimma, caracteritzada per una ampla coberta episfèrica (la calimma), delimitada per una banda equatorial de cèl·lules ciliades (el trocus) i amb l’extrem apical ocupat per una placa i un feix de cilis. El pol ventral es diferencia en el cos imaginal, que creix en longitud, i la calimma és reabsorbida o abandonada. La larva dels poliplacòfors té ocels laterals i es desenvolupa de manera semblant. El zigot dels cefalòpodes conté una gran quantitat de vitel, i això fa que les divisions cel·lulars i el desenvolupament embrionari tinguin lloc exclusivament en un pol de l’ou fecundat. La majoria de gasteròpodes marins tenen una larva anomenada vèliger, en la qual el trocus de l’estadi de pericalimma s’hipertrofia en una làmina ciliada (el vèlum), que genera corrents d’aigua vers la boca; molts òrgans de l’estat adult ja són presents en aquesta larva, fins i tot els ulls. La vèliger té una conquilla hemisférica, que s’hi uneix per dos músculs retractors; un d’aquests músculs creix moltmés que l’altre, com a part (no causa) de la rotació de 180° que sofreix la massa visceral continguda en la conquilla: aquest procés rep el nòm de torsió, i és el responsable de la asimetria dels òrgans interns dels gasteròpodes. Els bivalves i escafòpodes tenen una larva anomenada pedivèliger, semblant a la dels gasteròpodes, però sense el fenomen de torsió. Els bivalves unioïdeus passen per estadis embrionaris molt peculiars, lligats a una fase parasitària.
Les larves poden estar proveïdes de material nutritiu (larves lecitotròfiques), o bé no tenir reserves alimentàries (larves planctotròfiques). En el primer cas, la fase larval és curta, mentre que en el segon poden passar mesos abans de la metamorfosi; això implica diferències en el grau d’isolament, i, per tant, ritmes d’especiació més intensos en el segon cas. El desenvolupament directe es dóna de manera que les fases larvals puguin ésser reduïdes en grau variable. Els cefalòpodes i els gasteròpodes terrestres i límnics tenen sempre el desenvolupament directe. En cap moment no s’observa la diferenciació d’una cavitat interna homologable al celoma d’altres fílums: el complex gonopericàrdic s’origina per acumulació del mesènquima resultant de la desintegració del mesoblastemma. En general aquesta cavitat interna és força reduïda, encara que en els coleoides s’estén considerablement; en aquests cefalòpodes té la funció d’esmorteir els sobtats canvis de pressió que genera la locomoció per propulsió.
Filogènia i sistemàtica
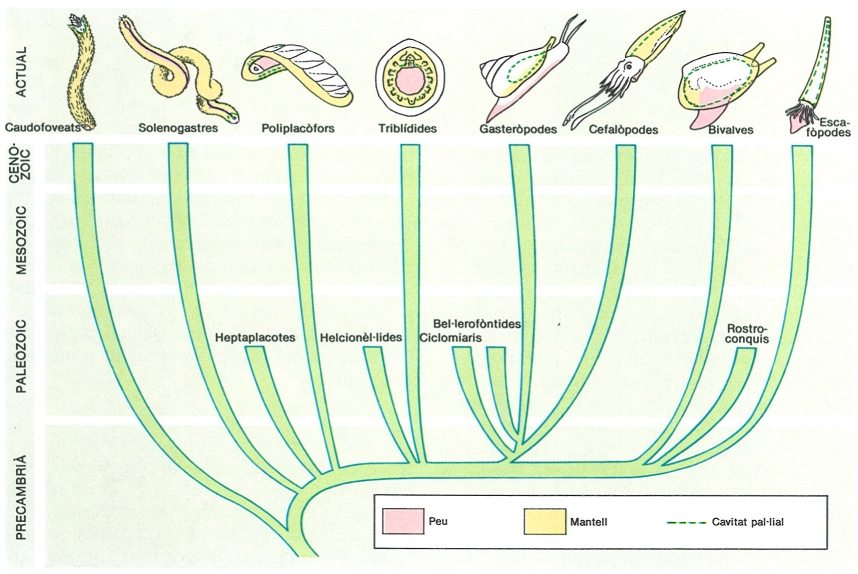
La filogènia dels grans grups de mol·luscs es pot inferir a partir de la distribució de molts caràcters homòlegs. Les característiques externes, però. són extremament variades. El peu varia molt de posició i de mida, reflectint el mode de locomoció. El mantell i la cavitat pal·lial reflecteixen el mode de respiració, la relació (en molts bivalves i gasteròpodes) entre la respiració i l’alimentació per filtració i la posició i la importància de la conquilla. La conquilla es forma al marge del mantell, de manera que creix amb la resta del cos. És present en la majoria de les classes, encara que la diversitat de formes i mides és impressionant i forneix molts exemples d’evolució convergent.
Biopunt, original de l’autor
Els mol·luscs són un llinatge de metazous clarament diferenciat, relacionat amb el dels celomats autèntics, però que n’ha divergit molt aviat. D’una banda, la desintegració del mesoblastemma s’observa en platihelmints, nemertins i mol·luscs, i, en aquest darrer grup es pot interpretar com un caràcter primitiu. D’altra banda la calimma larval és un caràcter derivat que mol·luscs, sipúnculs, equiürs i anèl·Iids comparteixen, encara que aquests tres grups són celomats i tenen larves trocòfores, amb metatroca, ocels apicals, protonefridis i cavitats secundàries. Així doncs, la formació d’una cavitat interna d’origen mesodèrmic (el celoma en sentit ampli) no és per si mateixa un indici fiable de proximitat filogenètica en els metazous, car ha aparegut de manera independent diverses vegades.
Els mol·luscs es divideixen en dos grans grups, els escutòpodes (Scutopoda), representats únicament pels caudofoveats (Caudofoveata), i els adenòpodes (Adenopoda), que inclouen tots els altres. Malgrat la semblança externa entre els caudofoveats i els solenogastres (Solenogastres), no és possible de reconciliar els plans estructurals d’aquestes dues classes: ambdues es distingeixen per una sèrie de caràcters molt especialitzats a un mode de vida concret (excavar en el fang els primers, predar hidroides colonials els segons), de manera que un avantpassat comú a tots els mol·luscs, tot i ésser prou sòlida la seva existència a principis del Fanerozoic, devia ésser un organisme molt més simple que qualsevol espècie vivent. El grup germà dels solenogastres és el dels testaris (Testaria), tots els mol·luscs amb conquilla; dins aquests darrers, els poliplacòfors, en els quals tota la morfologia és el resultat d’una adaptació curosa als ambients sobre substrats rocosos del litoral marí, són el primer llinatge divergent. Un altre llinatge de divergència primerenca és el dels triblídides (Tribíydiida), en els quals la multiplicació d’òrgans associats al sistema circulatori és una peculiaritat clarament exclusiva, malgrat que fou considerada com un indici de segmentació veritable en un possible arquetipus del fílum. Els gasteròpodes (Gastropoda) i els cefalòpodes (Cephalopoda) d’una banda, i els escafòpodes (Scaphopoda) i els bivalves (Bivalvia) de l’altra, comparteixen característiques bàsiques: en el primer grup hi ha diferenciada una massa cefàlica amb tentacles d’innervació cerebral, un parell d’ulls també cerebrals, el mantell limitat a la massa visceral, i el tracte digestiu amb boca i anus anteriors; en el segon grup el mantell cobreix quasi tot el cos, el peu és molt gran, en principi per a l’excavació en fons sorrencs, no hi ha un cap diferenciat, i el tracte digestiu té boca anterior i anus posterior.
Entre els mol·luscs paleozoics hi hagué una diversitat de plans estructurals bàsics més gran que en períodes posteriors; així, hom ha descrit fòssils que no encaixen en cap dels grups vivents, i que cal considerar com a representants d’altres classes extingides. Els heptaplacotes, els ciclomies, els helcionel·lis, els bel·lerofontis i els rostroconquis són tractats breument en l’apartat de sistemàtica de les classes actuals més properes.
En aquesta obra s’estudien totes les classes i ordres de mol·luscs, però només s’esmenten aquelles superfamílies i famílies presents als Països Catalans, o bé les que, tot i ésser exòtiques, són d’interès especial.