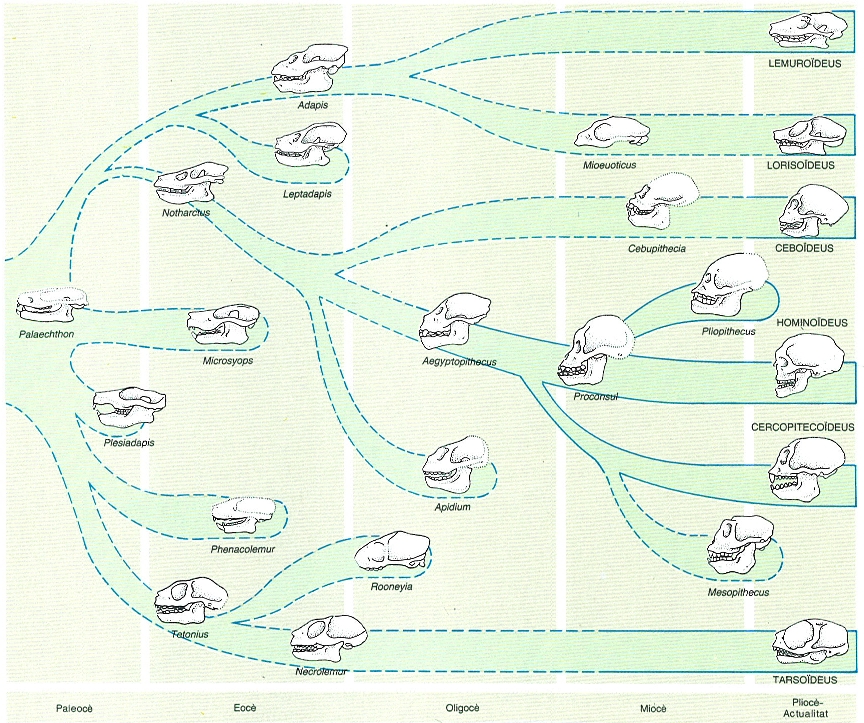
Arbre filogenètic dels primats. L’evolució dels primats és un cas típic de radiació adaptativa, amb una diversificació constant de grups i espècies. La categoria sistemàtica dels prosimis no inclou una única branca filogenètica, sinó dues ben separades, els tarsioïdeus per una banda i els lemuroïdeus i lorisoïdeus per l’altra. Dins dels avantpassats dels antropoides, hi ha una antiga diversificació entre platirrins i catarrins i una posterior entre cercopitecoïdeus i hominoideus. El gènere Aegyptopithecus, descobert a El Fayum (Egipte) i datat en uns 30 milions d’anys, presenta una semblança general amb els antropoides del nou continent. Clarament ja dins dels hominoideus, el Proconsul era primitiu i poc especialitzat, sense que s’assembli a cap dels primats superiors actuals en l’aspecte general.
Amadeu Blasco.
L’única espècie de primats present als Països Catalans —domesticismes a banda— és la pròpia espècie humana. La seva presència en aquest àmbit, d’altra banda, és molt antiga, amb restes fòssils que testimonien una ocupació mil·lenària. El sentiment antropocèntric que, ben comprensiblement, presideix la manera humana de mirar-se el món arriba a trobar xocant l’autoconsideració en el context d’obres com la present, però la coherència científica aconsella de tractar-hi l’home, tanmateix en termes d’antropologia biològica; altra cosa fóra que fos tractat, aleshores ben desencertadament, en termes etnogràfics, d’antropologia cultural o, molt pitjor encara, en termes històrics o socials. L’home com a primat —singularíssim, però primat al capdavall— és present en l’àmbit dels Països Catalans sota formes racials i amb característiques antropològicament tipificables, i mereix —exigeix—, doncs, un tractament concret i específic.
De fet, cal subratllar que fins a començament del segle XX —potser sota la influència del gran sotrac que representaren les descobertes evolucionistes del segle XIX, confirmades per les troballes paleoantropològiques— era corrent en qualsevol història natural dedicar un gran apartat a l’home i que és modernament que aquestes referències han desaparegut moltes vegades. Pot ser una reflexió interessant el fet d’intentar conèixer-ne les causes.
La locució «història natural de l’home» ha estat una denominació molt emprada en l’antropologia biològica, és a dir, en la disciplina que s’encarrega de l’estudi biològic de l’home o, més ben dit, de les poblacions humanes. La història natural de la nostra espècie té moltes connotacions, de les quals en podem ressaltar algunes: s’estudia una única espècie, per la qual cosa l’observació sol ser extraordinàriament minuciosa, i es dedica atenció a poblacions que habiten àrees petites; les tècniques d’estudi no són únicament les clàssiques, basades en la morfologia, sinó que hom n’ha integrat d’altres, sobretot provinents de la genètica, que permeten anàlisis a escala molecular que han resultat molt clarificadores; el coneixement detallat de la història de cada individu ha permès l’estudi de l’estructura i la dinàmica de les poblacions i ha donat dades empíriques als models i processos de diferenciació i evolutius; finalment, la comprensió biològica de l’home va estretament lligada al fenomen adaptatiu, al qual el sorgiment de la cultura ha donat el caràcter distintiu més important de l’home. Analitzar i destriar en el binomi biologia-cultura significa reconèixer les bases de l’adaptació, tant de l’home com a espècie com de les diferents poblacions humanes. Aquests aspectes, entre altres, han ajudat que moltes vegades no es consideri l’home dins dels estudis del món natural. Però, clarament, la biologia té quelcom a dir sobre l’home. Aspectes com l’evolució, la diversificació racial, la biodinàmica i diferenciació de les poblacions, la variació humana i molts altres entren de ple en la visió naturalística de l’home.
El marc evolutiu de l’espècie humana
L’home entre els primats: pòngids i homínids
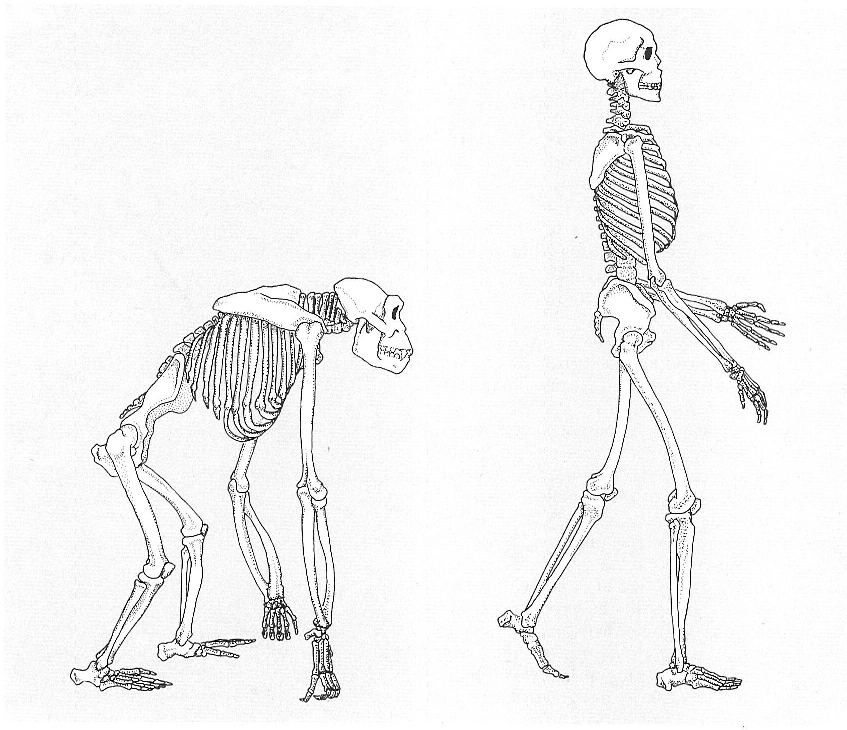
Esquema comparatiu entre l’esquelet d’un home i el d’un pòngid: tot i que el seu pla general és molt semblant, hi ha nombroses diferències relacionades amb dues adquisicions típiques dels homínids, la postura erecta i la cefalització. En l’esquelet postcranial podem observar que la columna vertebral presenta tres corbes en l’home i dues en els pòngids; la pelvis és més estreta i amb els ossos ilíacs estrets i verticals en els pòngids; en l’home els braços són més curts i les cames molt més llargues que en els pòngids, els quals tenen els ossos de les extremitats més robustos i arquejats. La mà és més curta en l’home, però el polze és més allargat; el peu té els dits més curts i el dit gros, que no és oposable, està molt desenvolupat i és paral·lel als altres quatre dits.
Ferran Rodríguez.
La semblança entre els pòngids i l’home s’ha mostrat molt gran en els diferents camps biològics en què s’ha portat a terme la comparació. Resulten colpidors els resultats moleculars, que ens mostren unes diferències realment minses: menys del 2 % a l’estructura de l’ADN nuclear entre el ximpanzé i l’home. Pel que fa a la morfologia, la comparació resulta il·lustrativa perquè ens permet entendre l’estratègia evolutiva humana, ens defineix els caràcters típicament humans i ens mostra en què cal fixar-nos en estudiar les restes fòssils del procés de l’hominització. Podríem dir que tota la diferència gira entorn d’un binomi que caracteritza la nostra espècie: la postura erecta i la cefalització. Pràcticament totes les diferències anatòmiques són interpretables a partir d’això.
La cefalització o increment de la capacitat cranial implica una globalització de la volta del crani, amb una forta alçada. No hi ha cresta sagital i la zona d’inserció de la musculatura mandibular és petita, a les línies temporals. La capacitat cranial és grossa, no únicament en termes absoluts (400-500 cm3 en els pòngids i 1300-1500 en l’home actual), sinó que ho és molt més respecte al pes corporal. Hi ha diferències importants en la dentició, relacionades amb la dieta, bàsicament herbívora en els pòngids i omnívora en l’home. En aquest, es produeix una disminució general de les peces dentàries, sobretot de les dents canines, que mai sobresurten de l’alçada de les altres peces. Per això, mai no es troben diastemes —separacions entre dents i ullals que permetren l’oclusió dentaria—, que són usuals en els pòngids, sobretot en els mascles.
La postura erecta implica moltes modificacions, que inclouen des de la posició del forat occipital al crani fins a l’estructura del peu. La conseqüència més important és l’alliberament de la mà de la funció locomotriu, la qual cosa possibilita el seu ús per a altres funcions. En el peu, en haver de portar a terme ell sol la locomoció, es produeixen canvis cap a una especialització, com la disminució de la mida dels dits externs i la pèrdua d’oposabilitat del dit gros. De fet, en tot l’esquelet postcranial podriem resseguir les diferències marcades per les adaptacions locomotrius: llargada, estructura i proporcions de les extremitats, columna vertebral, pelvis i altres.
L’estratègia evolutiva humana
L’evolució és marcada per un fet distintiu i únic en la història de la vida: l’adaptació al medi no es basa exclusivament en les modificacions de l’organisme, sinó també en les modificacions del propi medi. Hi ha hagut un continu deslligament de les condicions ambientals, una menor subjecció a les pressions selectives, gràcies a la capacitat de manipulació d’instruments, és a dir, gràcies als processos culturals.
L’estratègia evolutiva humana, que té com a punt bàsic el sorgiment de la cultura, pot analitzar-se des d’una perspectiva biològica dins del binomi d’especialització i generalització. L’home, considerat com a ésser biològic, és altament generalitzat en les seves estructures. La dentició, les extremitats anteriors o el cervell són exemples clars de la seva versatilitat funcional. Té, per tant, els avantatges dels éssers generalitzats. Però, en aquest cas, i com a procés innovador, la generalització representa la base orgànica per a l’adquisició d’especialitzacions fora del cos per mitjà de manipulació i fabricació d’instruments. Aquesta estratègia es beneficia dels avantatges dels dos grups, però amb un interès addicional: les especialitzacions fora del propi organisme són intercanviables i, a més, evolucionen molt ràpidament, ja que qualsevol procés cultural segueix els mecanismes lamarckians de transmissió. Aquest deslligament de l’entorn i la pròpia modificació de l’entorn per adaptar-se a l’espècie configuren, a grans trets, la gran innovació de l’estratègia evolutiva humana.
L’evidència fòssil de l’evolució humana
Les primeres restes fòssils que són clarament d’homínid, és a dir, les que pertanyen a una època en què ja estan separades les línies que porten als diferents pòngids actuals, es troben exclusivament a l’Àfrica. Deixant de banda restes més antigues molt fragmentàries, l’estadi més antic, ben diferenciat, que coneixem actualment és el d’Australopithecus afarensis, datat d’entre fa 3 i 3,7 milions d’anys. Tant per l’anàlisi morfològica de les restes d’Hadar (Etiòpia) com per les petjades deixades en caminar sobre cendres volcàniques a Laetoli (Tanzània), s’ha comprovat que tenia postura erecta i marxa bípeda. Les característiques cranials recorden, però, la seva proximitat evolutiva amb el ximpanzé.
A les terres africanes i fins fa uns dos milions d’anys, hi hagué una gran extensió i diversificació dels homínids, inclosos com a diferents espècies del gènere Australopithecus. És entorn d’aquesta època que, tant per les característiques morfològiques de les restes —capacitat craniana, dentició— com perquè aquestes es troben associades a una indústria lítica —primeres pedres tallades—, hom considera que ens trobem davant del primer home o, més ben dit, d’una espècie del nostre mateix gènere, Homo habilis.
Poc després, es produeix la sortida d’Àfrica i la conquesta del continent d’Euràsia. Les formes que visqueren entre fa 1,5 milions d’anys i 300 000 anys se les agrupa sota la denominació d’Homo erectus, tot i que els límits són poc precisos, a causa del canvi lent i gradual que s’observa en el procés evolutiu humà. Com sigui que la distribució en l’espai i en el temps d’aquesta espècie és tan gran, no és estrany que trobem una forta variabilitat morfològica en les restes i que el pas cap a formes posteriors —H. sapiens— es produeixi asincrònicament en l’àrea de distribució.
Les restes europees de menys de 300 000 anys són les primeres que hom assigna a la nostra espècie. L’evolució, però, no s’aturà, i en diferents regions geogràfiques sorgiren races diferenciades, entre les quals cal destacar l’home de Neandertal. Amb una morfologia ben característica, aquest home de Neandertal és una raça que visqué a Europa i l’Orient Mitjà des de fa 100 000 anys fins a 35 000, i és associat, a grans trets, a les cultures del Paleolític mitjà.
La sobtada desaparició de la morfologia neandertaliana, encara no prou ben explicada, va lligada a l’aparició de l’home modern o home del Paleolític superior, que, a Europa, coneixem com a raça o home de Cromanyó.
La raciació i el procés de poblament

Jaciment mesolític del Roc del Migdia. És situat prop de Vilanova de Sau (Osona) i s’hi trobaren les restes humanes d’un individu femení que pertany clarament a la raça mediterrànida. El jaciment forma part d’un conjunt d’abrics de reompliment que van cronològicament des del Paleolític superior fins al Neolític. Les datacions del Roc del Migdia corresponen al moment de la retirada dels gels de l’última glaciació i de la crisi de l’activitat caçadora de l’economia paleolítica. En el conjunt faunístic hi ha restes d’herbívors (cavall, cérvol, cabra, isard, toro salvatge), carnívors (llop, gat mesquer) i ocells. Hi ha també representació de la fauna malacològica marina.
Daniel Turbón.
Deixant a part els registres fòssils de l’existència de l’home al nostre país podem dir que tots els individus del Paleolític superior es consideren morfològicament moderns, en el sentit que llur tipologia no és gaire diferent de la que es troba en períodes posteriors. En cap d’ells, però, no es troben les característiques típiques de la raça que, majoritàriament, ocupa els Països Catalans des del Neolític fins a l’actualitat, la raça mediterrànida. La diferenciació d’aquesta raça es produeix in situ a la Mediterrània occidental i, per això, no té sentit plantejar-se problemes de grans migracions, ocupació, poblament o expansió de la raça mediterrànida.
La resta més antiga que presenta una morfologia típica mediterrànida és la de l’esquelet femení del Roc del Migdia, a Vilanova de Sau (Osona). Data de 9570 ± 220 anys aC, en un moment de trànsit entre el Paleolític i el Mesolític, coincident amb el final de la darrera glaciació. Les característiques mediterrànides hi apareixen clarament: estatura baixa i esquelet gràcil. En el crani, s’observa dolicocrània moderada, forma ovoide en norma superior, cara allargada —leptoprosòpia— i altres característiques que confirmen la seva assignació. Restes del jaciment proper del Cingle Vermell, també a Vilanova de Sau, que daten del 7810 ± 160 aC, no permeten una anàlisi tipològica per llur mal estat de conservació, però la dentició presenta una característica que cal destacar: l’elevada incidència de càries, poc usual al Mesolític.
La raça mediterrànida
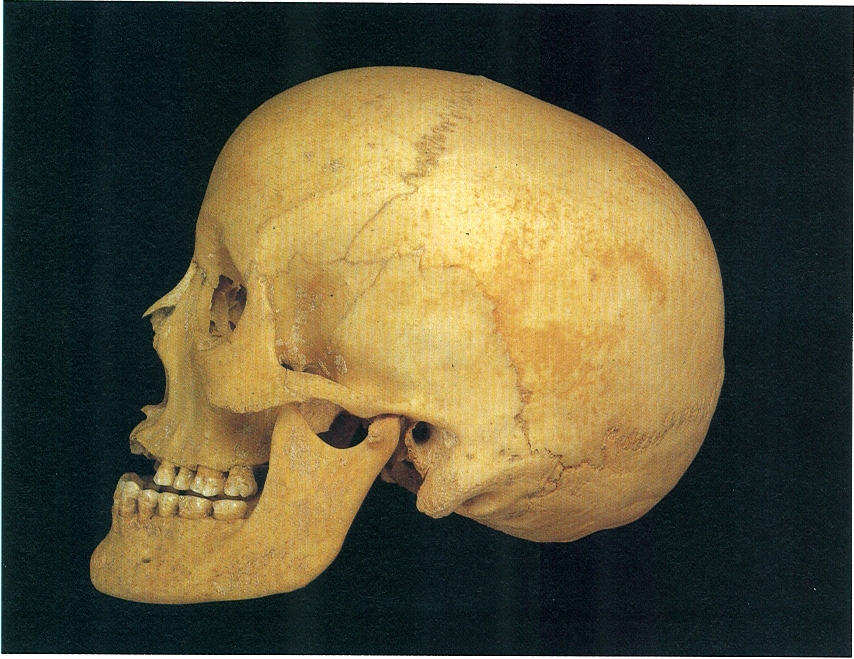
Crani d’home de la raça mediterrànida procedent de la necròpolis talaiòtica de Son Oms (Ciutat de Mallorca). El conjunt talaiòtic, actualment desaparegut per les obres de l’aeroport, data de l’inici del període talaiòtic i després d’un abandonament fou reocupat amb finalitats funeràries entre els segles IV i II aC. Des d’un punt de vista tipològic, la majoria d’individus pertanyen al tipus mediterrànid, caracteritzat per una gracilitat general de l’esquelet, crani allargat i no gaire alt, amb aplatament postobèlic (regió anterior a la confluència dels parietals i l’occipital) i cara i nas alts i estrets. És encara avui el tipus més freqüent a les nostres terres.
Jordi Vidal/Departament d’Antropologia.
Aquesta raça constitueix, actualment, un conjunt heterogeni, àmpliament estès a les ribes de la Mediterrània occidental i el N d’Àfrica. Es caracteritza per una pell opaca, més o menys fosca (és la pell més fosca dins la variació gradual que hi ha a Europa), cabell i ulls foscos, nas recte i prim, cara ovalada i allargada, cap dolicocèfal o mesocèfal i estatura força baixa. Dins de la raça mediterrànida, solen distingir-se dues subraces, però és més correcte considerar-les, senzillament, com a tipus diferents, ja que, en llur distribució geogràfica, no poden ser ben delimitades. Són, per un costat, el tipus mediterrànid gràcil (o subraça iberoinsular), de complexió prima, tronc llarg, extremitats curtes, estatura baixa i, pel que fa al cap, tendència a la mesocefàlia i volta craniana baixa, amb una clara depressió postobèlica; i el mediterrànid robust (o subraça atlantomediterrànida), d’estructura corporal més robusta i de més alçada.
La població humana des del Neolític
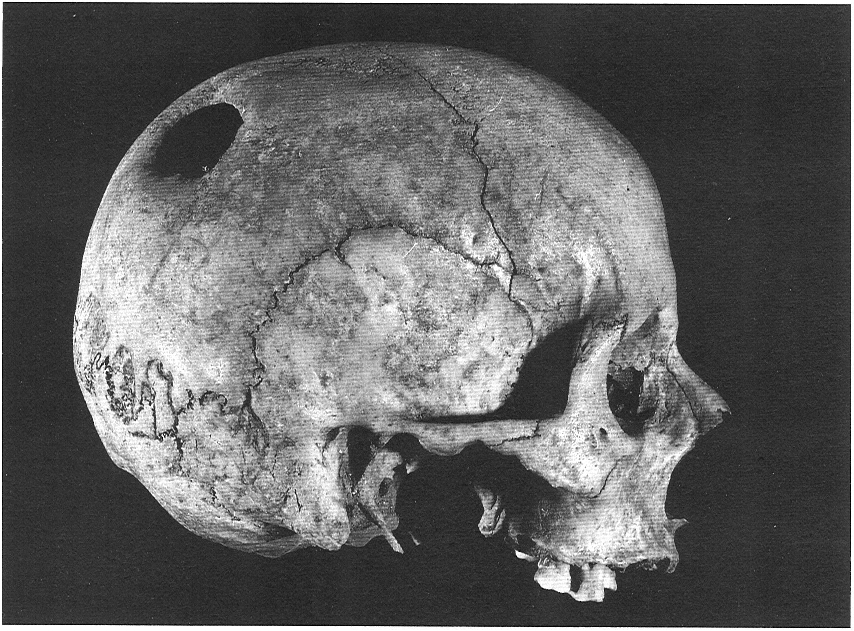
Crani d’home de tipologia alpina i trepanat, procedent del jaciment de la Torre d’en Cornet (Solsonès). Els primers individus de tipologia alpina que es troben a Catalunya s’inscriuen en els contactes migratoris amb l’Europa central: hi ha coincidència entre els representants més antics d’aquesta tipologia, l’inici de les cultures dels metalls i la tècnica de la trepanació. El crani és curt, ample i elevat, amb l’occipital corbat i la cara i el nas més amples i baixos que en els mediterrànids. L’estructura corporal és baixa i robusta. La trepanació, ben documentada des del segon mil·leni aC, es troba inicialment concentrada al Solsonès.
Oriol Clavell.
El Neolític representa un període cultural de la humanitat caracteritzat tradicionalment pel coneixement del poliment de la pedra, que té com a trets fonamentals els coneixements sobre agricultura i ramaderia que canviaren —o, més ben dit, revolucionaren— completament la societat. En viure menys subjecte a la caça i a la recol·lecció, l’home tendeix cap a una sedentarització, amb canvis en l’estructura social i en els hàbits alimentaris. Aquest fenomen arriba a les nostres terres vers l’any 5000 aC, tot i que s’han trobat indicis de domesticació referents al setè mil·leni aC. L’entrada al Neolític marca l’establiment definitiu de la tipologia mediterrànida com a substrat fonamental de la població, que persistirà, amb pocs canvis, fins als nostres dies. La constància morfològica durant més de 5000 anys és un punt interessant de reflexió pel que fa a la comprensió de la població dels Països Catalans. Tot i ser un lloc de pas per a moltes migracions, els canvis culturals s’han succeït amb diverses influències, però les variacions morfològiques a nivell poblacional —detectables en la població en conjunt— han estat poques.
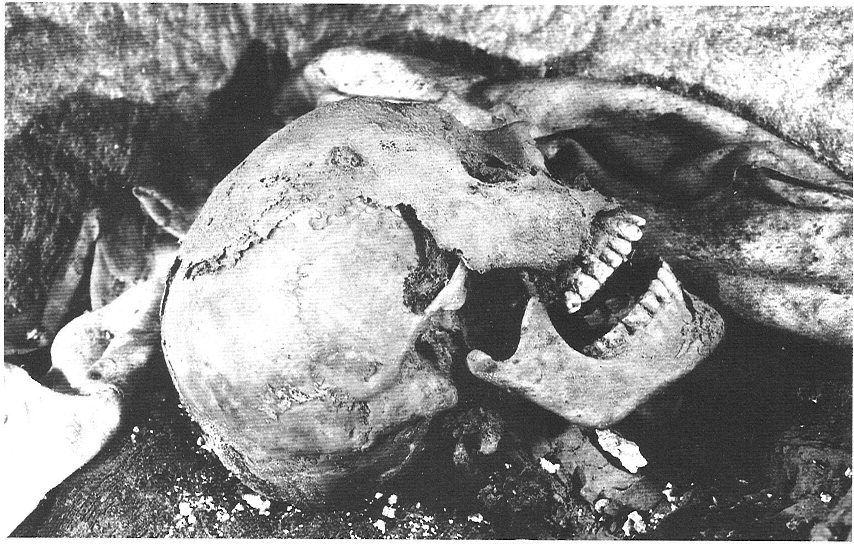
Restes atribuïdes a Ramon Berenguer II, "Cap d’Estopes", que es troben a la catedral de Girona. L’esquelet és complet i per sobre poden observar-se restes de teixits momificats de manera natural. L’esquelet correspon a un individu masculí d’una edat entre 25 i 35 anys, de gran robustesa i estatura (1,80 m). El crani és gros, de contorn superior ovoide, forma allargada i alt. La cara i el nas són molt alts i estrets. Aquestes característiques apunten cap a una tipologia nòrdica (que el color ros del cabell, inspirador del seu sobrenom, confirmaria), excepcional dins del context racial dels Països Catalans.
Jaume Bertranpetit.
Els homes neolítics es coneixen, als Països Catalans, per les troballes realitzades als sepulcres de fossa. A més de diversos jaciments on s’han trobat individus amb una clara tipologia mediterrànida, destaquen les restes de Sant Quirze de la Serra (Vallès Occidental), amb una morfologia que fa pensar en la persistència de característiques de l’home paleolític. No és fins al Neolític final i l’Edat del Bronze (segon mil·leni aC) que, pel nombre d’individus estudiats, podem fer una anàlisi antropològica més acurada, tant a nivell poblacional com tipològic, gràcies a les monografies sobre el bronze català i el Neoeneolític del País Valencià.
La semblança entre ambdues sèries és molt gran i mostren una composició racial molt propera a les poblacions posteriors, inclosa l’actual. Cal destacar un predomini més gran del tipus mediterrànid robust a llevant i, sobretot, la controvertida presència d’individus braquicèfals a les nostres terres. Aquests braquicèfals són de tipologia típicament alpina i serien, per tant, d’origen centreuropeu. La penetració al nostre país la van fer per la Cerdanya, seguint el curs del Segre, i s’instal·laren prop de Solsona; és molt probable que fossin els introductors de la metal·lúrgia del bronze i, entre ells, es troba estesa la pràctica de la trepanació. Aquesta influència alpina es fa sentir, molt atenuada, més al S (al barranc de Rifà, Baix Camp), fins al País Valencià, però dins d’un context mediterrani més ampli (Provença, Narbona-Rosselló, Principat, País Valencià) cal destacar la disminució de la influència alpina de N a S per dilució de l’influx de les poblacions centreuropees. Aquest element és minoritari, però la seva integració en el decurs de les generacions alterà, en part, les característiques de la població neolítica. La nova fisonomia, que continua essent predominantment mediterrànida, es manté invariable durant les importants novetats culturals posteriors.
Caldria fer esment d’un altre element forà, ben caracteritzat però menys nombrós, de tipologia braquicèfala plano-occipital, també detectat a partir de l’Edat del Bronze. Amb gran semblança amb les races dinàrica i armènida, la seva presència mostra una certa difusió racial en sentit E-W dins la Mediterrània.
Els costums incineradors —apareguts a finals de l’Edat del Bronze i vigents durant l’Edat del Ferro i l’època ibèrica, és a dir, durant el primer mil·leni aC— no permeten fer-se una idea de les característiques físiques dels habitants dels Països Catalans durant aquest mil·leni, en el qual els corrents migratoris procedents d’àrees del N són freqüents. Les restes òssies dels enterraments en coves o en urnes solament ofereixen una idea indicativa del nombre d’individus incinerats, i són escasses les precisions d’ordre demogràfic i pràcticament nul·les les morfològiques. La pràctica de la inhumació, recuperada a l’època romana —coexistent, però, amb la incineració—, contribueix al coneixement de les restes humanes. Les necròpolis de Tarragona i Empúries palesen la continuïtat morfològica.
Durant el mil·leni medieval, les esglésies fan de marc dels cementiris cristians i les necròpolis són, en general, petites. El manteniment de la tipologia mediterrànida i la pràctica absència de braquicèfals i d’altres formes fan pensar que el repoblament tenia un caràcter tancat, especialment a les àrees rurals, on la fortesa de l’esquelet postcranial és remarcable. Una necròpolis de caràcter especial és la de Montjuïc, a Barcelona, lloc d’enterrament de la comunitat jueva barcelonina dels segles XI al XIV, on aparegueren esquelets gràcils, amb base mediterrànida i algunes formes distintes que són interpretades com la prova d’un origen plural de la població. Els estudis fets fins ara indiquen que l’artrosi, les infeccions orals i els traumatismes són els problemes que afectaren amb més incidència la salut dels nostres avantpassats.
Les poblacions humanes prehistòriques balears
L’arribada de l’home a les Balears és, dins del context de l’evolució humana, recent. Per això, a diferència de les poblacions continentals, es pot parlar del poblament insular amb un significat precís: quan hi arriba l’home, de quin context racial prové i a quines influències posteriors és sotmès. L’inici del poblament és problemàtic i no és clar si és contemporani a totes les illes. Així, per exemple, si bé a Mallorca són ben documentats els temps antics, les dades sobre les Pitiüses són molt fraccionàries.
La presència humana més antiga s’ha detectat a la cova de Muleta, a Mallorca, amb una datació, per C14, de 3985 ± 109 aC, i a l’abric de son Matge, també a Mallorca, de 3800 ± 115 aC. Molt possiblement, l’establiment fos anterior, i es postulen dates entorn del 5000 aC. Una datació per C14 del gòral nan de les Balears (Myotragus balearicus) associat a restes humanes ha donat una datació de 5185 ± 80 aC. Els fets que no s’hagin trobat espècies animals domesticades ni ceràmica fins al 2700 aC, fan pensar que hi havien exclusivament arribades accidentals. Les primeres migracions que podríem anomenar «motivades» no es produeixen fins a l’inici del període pretalaiòtic, en dates més recents del 3000 aC. Raons geogràfiques i culturals apunten que les poblacions de la península Ibèrica serien l’origen dels primers habitants balears. No hi ha, en principi, raons per pensar en poblacions provinents de l’E de la Mediterrània.
Pel que fa a l’illa de Menorca, tot i que no hi ha evidències tan clares com a Mallorca, sembla que s’hi seguí el mateix procés i que el gòral nan també hi era extingit, per l’acció humana, a l’inici del període pretalaiòtic. La datació per C14 més antiga, fins als nostres dies, l’han proporcionada ossos d’animals trobats en el jaciment de Torralba d’en Salort, de 890 anys aC.
Els coneixements respecte de les illes Pitiüses han canviat radicalment arran de les excavacions de Can Sergent, a Eivissa, i Ca Na Costa, a Formentera, amb clares relacions amb el pretalaiòtic de les Balears. Fins fa poc, les restes més antigues es relacionaven amb el món púnic.
Han estat realitzats estudis antropològics sobre conjunts amplis d’individus dels períodes pretalaiòtic i talaiòtic mallorquins, sobre diverses necròpolis talaiòtiques mallorquines, i també s’han recopilat les dades referents a les restes menorquines. A grans trets, les poblacions prehistòriques de les Balears presenten molt poques diferències respecte a les poblacions que hem descrit de les zones peninsulars properes. L’element alpí hi és també present, així com, més tardanament, el dinàric. La composició racial, doncs, és totalment assimilable a la de la resta dels Països Catalans.
La població actual
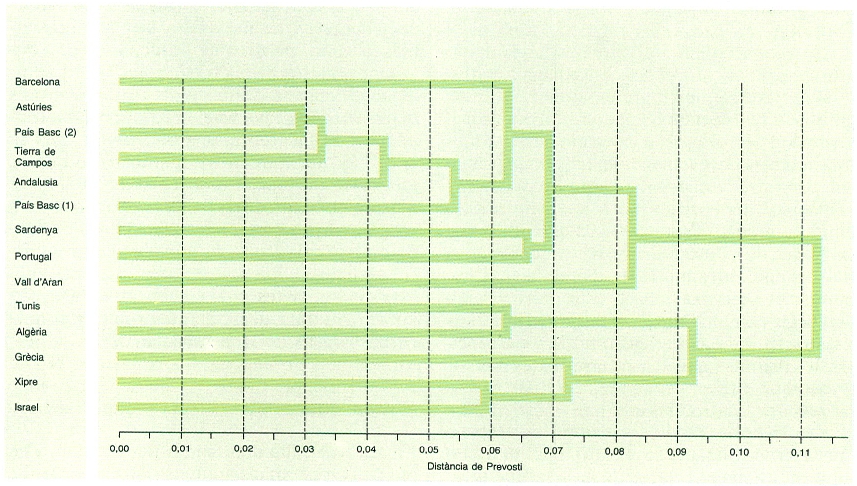
Relació de parentiu entre les poblacions de diferents zones a partir de dermatoglifs. El grau de semblança entre poblacions és en relació amb el seu parentiu genètic i és una eina interessant per a la tipificació biològica de les poblacions humanes i per a detectar influències entre elles. En aquest cas, s’ha emprat per definir en cada població la freqüència dels diferents tipus de figura que en cada un dels deu dits de la mà presenten els relleus dermopapil·lars o dermatoglifs.
Josep Manuel Melo, original de l’autor.
L’anàlisi antropològica d’individus vius aporta molta més informació que la consideració exclusiva de les restes esquelètiques, ja que tenim una gran quantitat d’informació aportada tant per la morfologia de les parts toves —teixits no perdurables— com per la variabilitat molecular. Dins del primer cas, queden compresos, entre altres, els estudis de somatologia (com la pigmentació de pell, cabells i ulls), de somatometria (mesures del cos), de somatoscòpia (observació de caràcters discontinus) i dels dermatoglifs (formacions epidèrmiques especials a dits, palmell i planta del peu). L’anàlisi molecular s’ha convertit, en els darrers anys, en una de les eines més potents per a entendre la biodinàmica de les poblacions humanes. Ja d’antic, s’han fet estudis emprant els diversos grups sanguinis: ABO, Rh, MN, P, Kell, Duffy i molts altres. Les tècniques electroforètiques han proporcionat, per altra banda, informació molt minuciosa sobre la composició genètica de les poblacions analitzant proteïnes plasmàtiques i enzims eritrocitaris.
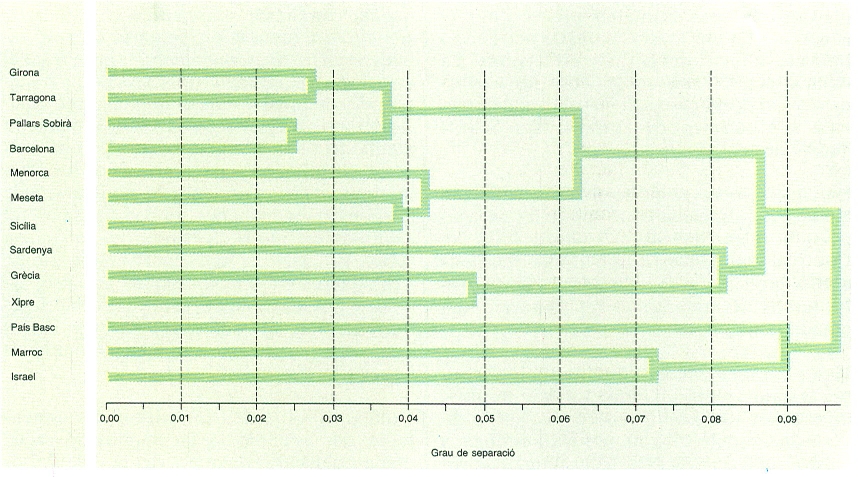
Relació de parentiu entre les poblacions de diferents zones a partir dels grups sanguinis. En aquest cas, hom compara les freqüències amb què es presenten els gens que codifiquen per diversos grups sanguinis —ABO, Rh (CDE), P, Kell i Duffy—, a partir de les quals s’obté una matriu de distàncies entre poblacions. La seva representació més senzilla i acurada és el dendrograma, interpretable en termes de semblança o parentiu entre les poblacions, que s’ha fet mitjançant l’algoritme matemàtic UPGMA.
Josep Manuel Melo, original de l’autor.
Quin significat tenen totes les dades esmentades per a entendre la composició racial d’una població, en aquest cas la dels Països Catalans? En parlar de composició racial, en realitat, no ens plantegem els problemes del poblament inicial, ja que el temps passat des de la primera presència d’homínids és prou gran perquè la diversificació produïda des de llavors sigui molt superior a la que podem trobar per àrees extenses. Entendre la composició racial no és altra cosa que veure les conseqüències de la dinàmica de les poblacions dins d’una àrea geogràfica extensa i sense uns límits concrets. I, per aconseguir-ho, es tracta d’establir i mesurar les relacions entre un conjunt ampli de poblacions, de manera que ens serveixi per a mesurar el parentiu que presenten, l’intercanvi genètic que han tingut entre elles. Tot i que qualsevol característica biològica que tingui una distribució geogràfica coneguda pot ser emprada amb aquest propòsit, és important que tingui una base genètica coneguda. I una limitació molt important són les dades disponibles (la informació sobre els País Valencià és, per exemple, pràcticament nul·la).
Les dades que permeten comparacions més extenses són les referents als dermatoglifs i als grups sanguinis. Les poblacions dels Països Catalans incloses són poques: Barcelona i Vall d’Aran pels dermatoglifs, i Girona, Tarragona, el Pallars Sobirà i Menorca pels grups sanguinis. Aplicant entre les poblacions la distància de Prevosti, s’han construït, mitjançant un algoritme matemàtic, uns dendrogrames que poden ser fàcilment interpretats en termes d’una semblança més o menys gran entre les poblacions comparades. Així com, dins de la península Ibèrica, els bascos representen un grup biològicament ben diferenciat, detectable sobretot en els grups sanguinis, hi ha una forta homogeneïtat entre les poblacions de la Mediterrània occidental, amb una discontinuïtat evident a l’estret de Gibraltar; en canvi, les relacions amb el N d’Àfrica, a les quals antigament s’havia donat gran relleu, no s’aprecien en aquesta anàlisi i, de fet, l’intercanvi d’individus deu haver estat escàs. Deixant de banda els bascos, de significat no ben comprès, la homogeneïtat biològica de les poblacions humanes de la península Ibèrica i, en general, de la Mediterrània occidental, es correspondria amb un antic i continu procés d’intercanvi d’individus que no ha permès l’establiment de discontinuïtats biològiques entre les diferents poblacions humanes. Aquesta conclusió està d’acord amb les dades osteològiques i rebutja les hipòtesis d’entrada de contingents importants provinents d’altres terres; les entrades exteriors, entenent-les com a integració d’individus en la població reproductora, han estat, doncs, poc importants. Raciològicament, la població dels Països Catalans se situa dins d’un àmbit molt més ampli de variació, encara que sempre immersa dins de les característiques típiques mediterrànides.
Recentment, s’ha fet un estudi hemotipològic extraordinàriament complet de la població de Menorca per tal de poder descriure i tipificar genèticament la població i, mitjançant comparacions, circumscriure-la dins l’àmbit de variació tant de les poblacions properes com d’un context més general de la variabilitat humana. S’han analitzat les variants bioquímiques en nombrosos polimorfismes: grups sanguinis (A1A2BO, CDE, MNSs, Lewis, Kell, Duffy i P), enzims eritrocitaris (ACP-1, EsD, ADA, 6-PGD i GLOI) i proteïnes sèriques (subtipus d’haptoglobines, de transferrines, del sistema Gc i de l’α1-antitripsina). La comparació dels resultats amb els d’altres sèries ha mostrat que la població actual de Menorca és plenament integrada dins de les poblacions del seu entorn geogràfic (Mediterrània nordoccidental) i ha fet palesa la màxima semblança amb poblacions catalanes, fet congruent amb les dades històriques. Tot i això, hi ha diferències importants en alguns dels marcadors entre Menorca i aquestes sèries catalanes: hi ha una més gran afinitat amb la població de Barcelona, caracteritzada per una forta influència d’altres zones de la península Ibèrica, que no amb altres que han rebut menys immigració, com Girona i Tarragona. Així, doncs, es mostra que la població actual de l’illa de Menorca presenta unes característiques diferencials pròpies que són atribuïbles tant al caràcter insular com a la peculiar història de la població.
Els canvis recents

Els trets característics de la raça mediterrànida poden observar-se en la gran majoria dels pobladors autòctons dels Països Catalans. Aquesta raça es troba actualment estesa pel sud i oest europeus, nord d’Àfrica i part d’Orient Mitjà, amb ramificacions fins a l’Índia. Es caracteritza per una pigmentació fosca, que augmenta per insolació, i cabells arrissats o ondulats foscos; dolicocefàlia o mesocefàlia, cara i nas alts i estrets i una conformació esquelètica feble, amb una estatura baixa.
Jaume Bertranpetit.
En les poblacions humanes occidentals, s’ha produït en els darrers anys —l’últim segle, de manera general— un seguit de canvis que han originat profundes transformacions en llur substrat biològic, detectables tant a nivell genètic com morfològic. Les causes primàries estarien en l’augment general de la mobilitat dels individus i en la millora alimentària, ambdós fenòmens associats als canvis socioeconòmics, però també s’han afegit altres factors a l’hora de produir aquestes modificacions. Així, les millores sanitàries, sobretot pel que representen en l’eliminació de les parasitosis, tenen incidència en la configuració morfològica de l’adult. Igualment són de gran transcendència els canvis demogràfics, amb implicacions genètiques importants, entre els quals cal constatar la disminució de la fecunditat i de la mortalitat infantil, amb gran incidència en les possibilitats d’actuació de la selecció natural; d’altra banda, la disminució de la mortalitat, sobretot a edats baixes, juntament amb la reducció de la natalitat, han produït canvis importants en l’estructura demogràfica, amb un increment constant i no aturat de la proporció de vells en la població.
A nivell genètic, es detecta una tendència a una uniformització entre les poblacions que, si bé no és perceptible a una escala geogràfica àmplia —entre continents o països allunyats— sí que és important dins d’una escala més reduïda, com en el cas de les poblacions pirinenques, en què la manera de viure de la societat tradicional havia permès un grau de diferenciació entre elles, basat, sobretot, en l’aïllament. L’augment de la mobilitat i les migracions de grans contingents humans ha produït, darrerament, uns canvis en la composició genètica de la població dels Països Catalans que, ara per ara, són difícils d’avaluar, però que, òbviament, apunten cap a una homogeneïtzació de les poblacions de la península Ibèrica i les illes Balears.
Els canvis socioeconòmics, base dels fenòmens migratoris esmentats, tenen repercussions morfològiques molt directes per les seves implicacions en els canvis alimentaris. És interessant ressaltar el fet que les dimensions corporals, que han estat un suport important de la taxonomia dels grups humans, són fortament afectades pels hàbits nutricionals i que el patró de creixement dels individus, les proporcions corporals i l’estatura final n’han resultat alterats.
Els canvis en el patró general de creixement han pogut ser analitzats gràcies als estudis escalonats en el temps referits a escolars barcelonins, de sexe masculí, que abasten des de 1944-45 fins a 1978-79. El ritme de creixement actual es caracteritza per un període puberal (entre els dotze i els catorze anys) de màxima crescuda, més prematur i intens que anys enrere. També ha augmentat el ritme en l’estirada infantil (entre els set i els nou anys), però aquest fenomen no és tant important.
A més de les variacions en ritme de creixement i en l’estatura final, hi ha importants canvis en les mesures parcials i en les proporcions. En el tronc, hi ha una reducció de l’amplada d’anques que, juntament amb un allargament considerable de les extremitats inferiors, donen un aspecte més longilini i esvelt. La llargada relativa de l’extremitat superior es redueix, sobretot en l’avantbraç; en la mà s’ha mantingut l’amplada i la llargada és una mica superior i en el peu han augmentat les dimensions, sobretot l’amplada. Els canvis en les dimensions cefàliques són poc importants: destaca una disminució de l’índex cefàlic perquè augmenta la llargada del crani i en disminueix l’amplada, però és manté dins la mesocefàlia; també s’han incrementat l’amplada frontal mínima i la bigoníaca (corresponent a l’amplada de la mandíbula). Tot i que es coneix, també, el patró general de creixement referent al sexe femení, no podem establir-ne la variació temporal perquè no es coneix en etapes anteriors.
L’estatura ha mostrat un canvi espectacular en aquest segle: les dades disponibles mostren que l’augment produït des de finals del segle XIX fins a 1983 és de gairebé 7 cm per al conjunt d’homes dels Països Catalans, la qual cosa representa un 4 % més d’estatura.
L’acceleració màxima en tot el segle es produeix a la dècada 1970-80 —coincideix, per tant, amb l’època de creixement dels nascuts en els moments de màxima expansió econòmica. Hi ha importants diferències geogràfiques relacionades amb el desenvolupament econòmic, però, en el decurs del temps, es van reduint.
L’edat de la primera menstruació (edat de menarquia) es troba en un important procés de canvi, semblant a l’observat en l’estatura. Les dades disponibles, referents únicament al Principat, mostren que entre les dones nascudes entre 1925-29 i les nascudes entre 1950-54 hi ha hagut una disminució de gairebé un any (de 13,71 anys a 12,75 de mitjana).
Tots aquests canvis són interessants d’analitzar, sobretot perquè ens mostren la forta interrelació entre la biologia i la cultura, o, més concretament, entre la morfologia i l’alimentació, fet que fa prendre amb precaució els estudis que pretenen ser exclusivament morfològics.
Les malalties genètiques humanes amb rerefons adaptatiu
Parlar de malalties dins del context de la història natural pot resultar sorprenent si considerem el procés patològic com quelcom independent dels individus i de llur bagatge genètic. Però hi ha malalties que no poden ser enteses només com a processos patològics, ja que presenten una base genètica i un rerafons adaptatiu que donen, a les poblacions on es presenten, una més gran eficàcia per a sobreviure en un medi concret. En aquests casos, es pot dir que els interessos de l’individu i els de la població no coincideixen: per a una millor adaptació global, hi ha alguns individus que són portadors de gens que poden desencadenar processos que els portin, fins i tot, a una mort prematura.
Entre els nombrosos casos descrits, n’hi ha dos que, per llur distribució geogràfica, difosa per la Mediterrània, i llur avantatge davant d’una malaltia tan important com ha estat la malària —anomenada també paludisme—, presenten un interès especial en les poblacions humanes dels Països Catalans: el favisme i la β-talassèmia.
El favisme
Des de finals del segle passat, es coneix el fet que alguns individus, en principi perfectament normals, poden patir una peculiaritat curiosa: pel fet de menjar faves o simplement de travessar un camp quan aquestes estan en flor, són víctimes d’una crisi hemolítica: hi ha una destrucció en massa de glòbuls vermells, amb fatiga, pal·lidesa i icterícia consegüents, que pot ser molt greu i ocasionar la mort. Les faves tendres i crues són les que provoquen els accidents més espectaculars, i molts dels accidents mortals es produeixen en infants de pit, ja que les substàncies que ocasionen l’hemòlisi passen a la llet materna. A Menorca, un refrany hi fa clara referència: «Qui de fetge va, que no passi pel favar».
Des de l’any 1956, se sap que la malaltia és associada a un deficiència de l’enzim glucosa 6-fosfat deshidrogenasa (G6PD). El gen per a la seva síntesi és situat en el cromosoma X, i és, doncs, un caràcter lligat al sexe. Abans d’aquest descobriment, i animats per les altes incidències descrites a poblacions mediterrànies (Grècia, Xipre, Itàlia i Sardenya, entre altres), Surinyach, Marcolongo, Alcobé i Llebaria estudiaren, l’any 1951, el focus de favisme de Menorca, que era particularment intens a la part més occidental de l’illa, entorn de Ciutadella. Reuniren 26 genealogies que incloïen un total de 51 fàvics; llur treball, però, no permet una anàlisi epidemiològica de la incidència, ja que no es féu un mostratge dins de la població. Alcobé posà en clar la coincidència familiar de la malaltia i féu una anàlisi genètica que s’adequava a les genealogies trobades. De fet, l’interès actual del treball és escàs pel desconeixement que hi havia dels fonaments bioquímic i genètic del fenomen.
La incidència de favisme s’estudia, actualment, a partir de la deficiència en G6PD: els homes poden ser normals o deficients —fàvics— i les dones, normals, portadores (heterozigotes) o deficients (homozigotes). Per això, l’anàlisi sol centrar-se en els homes. Els treballs realitzats en poblacions dels Països Catalans permeten constatar la presència de favisme tot i que el nombre d’individus és petit en gairebé totes les mostres. Les freqüències més elevades semblen trobar-se a les Illes; la idea inicial d’Alcobé de trobar unes incidències més altes en les zones més tancades i aïllades sembla, per tant, confirmarse. De tota manera, no s’arriba a les altes xifres trobades a altres illes de la Mediterrània.
A continuació, la taula reuneix dades sobre la incidència de la deficiència de l’enzim glucosa 6-fosfat deshidrogenasa (G6PD) en algunes poblacions dels Països CatalansDonat que és un caràcter que es transmet segons el model d’herència lligada al sexe, les dades es refereixen, tret del cas de Barcelona, al sexe masculí. La deficiència de l’enzim provoca la malaltia coneguda com a favisme. (Dades de l'autor.)
| POBLACIÓ | NOMBRE (HOMES) | DEFICIENTS | % | FONT | |
|---|---|---|---|---|---|
| Mallorca | 110 | 0 | Flatz i Düren | ||
| Sineu | 132 | 3 | 2,3 | Flatz i Düren | |
| Mallorca total | 991 | 5 | 0,5 | Miguel i altres | |
| donants | 222 | 0 | Miguel i altres | ||
| d’escoles | 241 | 1 | Miguel i altres | ||
| laboratori d’anàlisi | 528 | 4 | Miguel i altres | ||
| Menorca | 98 | 0 | Flatz i Düren | ||
| Ciutadella | 114 | 1 | 0,9 | Flatz i Düren | |
| Ciutadella | 640 | 5 | 0,8 | Miguel i altres | |
| València | 400 | 0 | Flatz i Düren | ||
| Sueca | 104 | 1 | 1,0 | Flatz i Düren | |
| Delta de l’Ebre | 96 | 0 | Flatz i Düren | ||
| Barcelona (homes i dones) | 400 | 0 | Guasch |
La β-talassèmia
Hi ha diverses anomalies que es caracteritzen per una disminució o absència genèticament determinada de síntesi d’alguna cadena d’hemoglobina. Són les talassèmies (de thalassos, mar, per llur associació amb individus de les ribes de la Mediterrània). Quan la cadena afectada és la β, es parla de ß-talassèmia, que és el cas d’interès en les poblacions mediterrànies, tot i que es troba en altres llocs. Els individus heterozigots tenen una anèmia lleugera, amb augment d’altres tipus d’hemoglobina, especialment A2, i no els cal tractament mèdic. Els homozigots, en canvi, tenen una anèmia greu i requereixen múltiples transfusions. A la infantesa, en absència de tractament, produeix la mort. Tot i que seria d’esperar una freqüència molt baixa d’una malaltia tan greu, es troba una incidència important en algunes poblacions i és un problema de salut pública a llocs com Xipre o Sardenya.
Les primeres dades referents als Països Catalans es troben en un estudi (1967) fet per Pellicer a Madrid, que comprèn un nombre molt baix d’individus de l’àmbit català: 39 del Principat i 30 de València, amb un únic cas detectat en aquest darrer lloc. El grup GEHBTA, de l’Asociación Española de Hematología y Hemoterapia, ha recollit dades molt completes sobre la incidència de les diverses talassèmies. Les dades corresponents a 1986 donen les següents freqüències de β-talassèmia, en tants per mil (‰): 0,0 a Lleida, 1,0 a Barcelona, 0,3 a Girona, 4,4 a València, 4,4 a Alacant, 3,2 a Mallorca i 50 a Menorca; les dades són d’àmbit provincial, excepte les de les Illes, i suposen una mostra superior a 1000 individus, excepte en el cas de Girona, en què el resultat es considera provisional. El valor de Menorca sembla que haurà d’ésser modificat per una xifra superior, entorn del 70 ‰. Cal destacar d’aquests resultats la gran incidència a Menorca, que supera llargament la dels altres llocs estudiats. Mallorca presenta un valor més baix, poc per sota del del País Valencià. A Catalunya, la freqüència és molt baixa, amb una incidència pràcticament nul·la a Lleida i Girona. És molt possible, de tota manera, que la incidència tingui una distribució molt més complexa, amb fortes oscil·lacions en considerar entitats poblacionals reduïdes que en llur història hagin configurat un patró adaptatiu peculiar.
El favisme, la β-talassèmia i la malària
La presència d’aquestes malalties genètiques ha d’explicar-se de manera que donin a les poblacions on són presents algun avantatge amb vista a la selecció natural i a l’adaptació, car si no desapareixerien. Tant en el cas del favisme com en el de la β-talassèmia, l’avantatge radica en el fet que confereixen als individus portadors (les dones heterozigotes pel favisme i els heterozigots per la talassèmia) una resistència enfront de la malària o paludisme. Actualment, coneixem força bé les bases bioquímiques d’aquesta protecció.
El panorama és complex: una malaltia endèmica de moltes parts del món i una protecció per mitjà d’un caràcter genètic que pot produir la mort. Però en el cas del favisme el model és, encara, més complicat: el consum de faves per individus normals, pel mateix mecanisme bioquímic que provoca hemòlisi en els fàvics, és un protector davant del paludisme.
Les preguntes que podrien sorgir són moltes, i una de les més interessants és la referent a l’origen d’aquesta complicada interrelació. Per intentar contestar-la, la paleopatologia ens ofereix una eina extraordinària: les anèmies produïdes per la talassèmia provoquen unes lesions detectables en les restes òssies. La manca de glòbuls vermells produeix, entre altres anomalies, un engruiximent de les parets cranials, coneguda com a hiperostosi osteoporòtica; en la part orbital del frontal, també poden formar-se una sèrie de trabècules, fet conegut com a "criba orbitalia". També l’anèmia consegüent a les crisis de favisme produïdes a la infantesa pot provocar lesions cranials que s’assemblen a l’hiperostosi porosa d’origen talassèmic.
Els primers casos descrits referents al conjunt de la Mediterrània es remunten, possiblement, a finals del Paleolític i es troben ben documentats al Mesolític, al Neolític, a l’Edat del Bronze i a la Grècia clàssica, amb clara relació amb hàbitats prop d’aiguamolls, llocs que afavoreixen l’expansió de la malària. En els Països Catalans, es troben casos típics ja des de finals del Neolític.
A la Cova del Frare (Matadepera, Vallès Occidental), hi ha restes d’un infant d’un o dos anys amb "criba orbitalia". Es tracta d’una cova d’hàbitat i enterrament del Neolític final, datat pel C14 en 2500 anys a.C. Al País Valencià, a la Coveta Emparetà (prop d’Alcoi), hi ha restes d’un infant de dos o tres anys amb una hiperostosi osteoporòtica que correspon al Neolític final. En el jaciment hi ha senyals de domesticació, tot i que hi ha moltes restes d’animals procedents de la caça. Podria correspondre, per tant, a una economia caçadora-recolectora amb ramaderia incipient. Aquest fet és interessant, ja que hi ha hipòtesis que relacionen l’expansió del mosquit Anopheles, vector del Plasmodium, productor de la malària, amb l’adopció de l’agricultura a les societats mesolítiques.
No s’han descrit casos amb aquestes anomalies en les sèries òssies estudiades de Mallorca, ni tampoc no hi ha cap cas en les restes prehistòriques de Menorca. Tot i això, és de suposar que aquestes anèmies hi eren presents i que podrien ser detectades en jaciments que subministressin restes òssies infantils.
Totes aquestes dades conflueixen a mostrar que les malalties genètiques que donen una resistència envers la malària han estat molt importants en la història biològica de les poblacions dels Països Catalans. Les diverses variants de resistència han tingut una importància diferent segons els llocs, sobretot en considerar poblacions reduïdes, i, en l’actualitat, per causa del trencament de l’aïllament, es van fent cada cop més difícils de detectar les estratègies poblacionals concretes amb vista a una adaptació que durant mil·lenis ha estat d’una importància cabdal per a la supervivència.