
La dominància del color blau en la visió espacial de la Terra és en gran manera deguda a l’abundància superficial d’aigua líquida i també gasosa. Això justifica el nom de «planeta blau» que se li aplica i explica que la biosfera, capa viva de la Terra, i igualment superficial, estigui tan vinculada a l’aigua.
Fotolia.com
L’observació directa de la Terra des de l’espai ha popularitzat la denominació de «planeta blau» per al nostre món. D’una banda, fa referència a la quantitat d’aigua que hi ha al nostre planeta: 1348 milions de km3 d’aigua líquida als oceans, almenys 27 milions de km3 d’aigua sòlida (glaç) i poc més de 10 000 km3 d’aigua a l’atmosfera i als rius. D’altra banda, hi ha una part considerable d’aigua, difícil de precisar, circulant a l’escorça sòlida de la Terra i en capes fondes. En termes de superfície, dels 510 milions de km2 que integren la Terra, 361,1 milions de km2 (el 70,8%) corresponen als oceans. La proporció pot ser diferent en qualsevol àrea arbitràriament delimitada (per exemple, els Països Catalans), però la proporció és significativa en l’àmbit de tota la Terra. Si més no, ens diu que l’extensió oceànica és molt gran i que, quant al funcionament de la Terra com a sistema ecològic (en l’àmbit de la biosfera), ha de ser important. Dit d’una altra manera, per a entendre el funcionament de la natura viva, objectiu central de l’ecologia, ens cal tenir una visió adient de la vida marina, ultra l’interès directe que pugui tenir, per exemple, si considerem les aigües litorals com a integrants d’un país i que contribueixen a la seva vida. Val la pena de recordar, també, que la designació de «planeta blau», esdevinguda gairebé un eslògan, recorda la unitat i l’aïllament de tota la Terra; aquesta unitat és essencial en la formulació de les ciències biològiques, com l’evolució i l’ecologia, i és, prospectivament, important si es considera que l’esdevenidor de l’home en el mateix planeta és un altre objectiu, potser més popular, de la ciència ecològica. De fet, l’ecologia té en compte l’aspecte extern, l’escenari, on opera la vida i el caràcter de sistema que tenen els conjunts d’individus, però pressuposa el coneixement intern, fisiològic, dels actors del drama. La major part de la resta de l’obra s’ha dedicat a la presentació dels organismes, amb una extensió major o menor en els aspectes ecològics. En aquesta part, volem centrar la nostra atenció en l’aspecte ecològic més sintètic, en allò que hom podria anomenar la macroecologia, per analogia amb la macroeconomia.
Val la pena de recordar unes quantes nocions molt necessàries i molt senzilles. La biomassa és la quantitat de matèria viva, i és convenient d’expressar-la d’una manera unificada, que pot ser en grams de carboni orgànic per metre quadrat; en ecosistemes terrestres els valors queden, sovint entre 1000 i 10 000 g C/m2. La producció (neta) és la quantitat de compostos orgànics de C sintetitzats anyalment pels productors primaris, descomptant-ne la respiració dels mateixos productors primaris; els valors mitjans són a la ratlla de 100 g C/m2·any en comunitats marines, i d’uns 300 g C/m2·any en els ecosistemes terrestres.
La producció sobrant dels vegetals o productors primaris nodreix, en forma d’energia dels enllaços químics i de tota mena de materials, els organismes anomenats heteròtrofs, que poden ser relativament petits i absorbir-la a través de tota llur superfície, que és molt extensa (bacteris i fongs, principalment), o bé relativament grossos i, llavors, obtenen l’aliment per ingestió (animals). El pas de materials i d’energia dels vegetals als heteròtrofs es considera, també, una producció, que hom qualifica de secundària, per a distingir-la de la producció primària, en l’àmbit dels vegetals. La producció secundària és, doncs, una fracció compresa dins de la producció primària bruta o total, que és un dels conceptes més interessants en ecologia. Podríem dir que mesura el cost de mantenir i de fer funcionar tota la biosfera. Per això, tota comparació entre la producció i la biomassa, siguin les totals, siguin les que es refereixen a un grup definit, és un indicador d’una propietat fonamental dels sistemes vius: llur taxa de renovació. La taxa de renovació, que es pot considerar, també, una productivitat —és a dir una mesura relativa de la producció amb relació a un factor important de producció com és, sens dubte, la biomassa— es pot simbolitzar, doncs, per P/B i té la dimensió d’una velocitat. El seu invers, B/P, correspon, per tant, a un temps, i ens dóna el temps necessari mitjà de la renovació del conjunt de la biomassa.
La biosfera té una reserva limitada d’elements biògens, com ara el carboni, que es considera el més característic, o el fòsfor, que és, relativament, el més escàs i, per tant, és el limitant efectiu en la producció de nova matèria viva. La producció biològica s’equilibra amb els processos de descomposició i de retorn d’aquests elements al medi. Els anomenats cicles ecològics o biogeoquímics consisteixen en aquest continu circular dels materials; llur velocitat, de la qual depèn la productivitat i, sovint també, la biomassa, no és sols regulada pels organismes, sinó també per l’es tructura funcional de l’entorn, tal com veurem. Ara bé, el camí que va dels productors primaris al retorn final dels elements al medi, en una forma que ja no es considera orgànica (generalment en forma de compostos oxidats relativament senzills, com ara CO2, etc.), s’efectua a través d’una considerable acumulació de materials que podem qualificar d’orgànics, però no vius. La biosfera, en efecte, porta darrere seu, com una ombra, una «necrosfera» de material mort, pendent de reciclatge. Es pot dir, gràficament, que moltes molècules sintetitzades en el cos d’organismes, tenen una «vida» molt llarga, que duren més temps mortes que no pas vives. Això ocorre en graus diferents i d’acord amb la natura i el significat d’aquestes molècules. Per exemple, són particularment persistents les molècules que són part d’estructures que convé que durin, el mecanisme generador de les quals ha estat sotmès a una evolució, per selecció natural, que ha premiat la capacitat de produir substàncies cada vegada menys fàcilment degradables. Qualitat que es demana en materials com és ara la fusta (cel·lulosa, lignina), els pèls i les plomes (ceratines), les substàncies de reserva que convé que estiguin protegides amb pany i clau (que només puguin ser metabolitzades pels enzims específics) enfront de la possible utilització per part d’altres organismes i, finalment, els missatgers químics de tota mena (feromones, antibiòtics), la funció dels quals demana una conservació de les estructures moleculars durant un temps considerable. A la llarga, molts dels materials més resistents es caracteritzen pel fet d’incorporar anells fenòlics a les molècules, amb una gran varietat en l’ocupació de les valències que queden lliures, és a dir, amb la possibilitat de tenir configuracions molt diverses amb molt poca diferència o nul·la en energia química entre les unes i les altres. Això és característic d’allò que, en general, hom anomenava i encara anomena substàncies húmiques, amb el benentès que «humus» es referia a la matèria orgànica del sòl — humus vol dir terra en llatí— quan ja havia perdut una estructura que permetia de reconèixer-ne l’origen. Tant per l’experiència com pel fet que es conserven certes característiques moleculars, es creu que una bona part de l’humus ve de la lignina, és a dir, de la fusta o les parts rígides dels vegetals; però a la mar hi ha una gran quantitat de matèria orgànica que, molecularment, no és gaire diferent de l’humus del sòl, sense que hi hagi fusta en el seu origen, sinó que és el resultat d’una sèrie de transformacions molt variades a partir de materials igualment diversos. Es pot dir, més aviat, que, tant a la terra com a la mar, es tracta de molècules orgàniques particularment resistents, a les quals es pot arribar per vies diverses, i que s’acumulen, simplement, perquè tenen menys probabilitats de ser destruïdes que altres tipus de molècules.
Es pot comprendre fàcilment que la cua de matèria orgànica morta que la vida deixa darrere seu és heterogènia, amb un enriquiment progressiu de les molècules de més durada, com les suara esmentades i d’altres, a mesura que passa el temps (moltes tenen milers d’anys, o fins milions, com en el cas del petroli). Fet i fet, hi ha més materials a la necrosfera, esperant ser descompostos, que a la biosfera, encara que és difícil de fer estimacions fiables. En els ecosistemes continentals hi pot haver entre 8 i 14 vegades més de matèria morta amb característiques orgàniques que matèria viva, i a la mar la proporció pot ser superior. Només cal pensar que els oceans contenen, per columna vertical d’un metre quadrat de secció, una concentració mitjana d’almenys 400 g de carboni orgànic no viu en molècules dissoltes, és a dir, sense comptar la matèria orgànica morta i encara en forma de partícules, que, ella sola, ja és superior a la biomassa. Es pot pensar que la quantitat i la qualitat de la necrosfera, amb relació a la biosfera, varia considerablement d’uns llocs a uns altres: a terra, per exemple, no acumulen parts mortes de la mateixa manera la tundra i la selva tropical humida.
Els ecosistemes terrestres i els ecosistemes oceànics

Model de l’energia externa i la regulació del flux de l’assimilació fonamental.
Carto-Tec, original de R. Margalef i M. Estrada.
Val la pena de continuar comentant les semblances i les diferències entre els sistemes terrestres i els oceànics. No s’ha d’oblidar que, venint de terra, nosaltres mateixos portem sovint una sèrie de prejudicis amb els quals volem interpretar el funcionament dels ecosistemes marins. A més, per a nosaltres, aquesta comparació no és inútil i ens fa copsar millor l’essència dels problemes ecològics més generals. Un tema constant de referència pot ser l’afirmació feta en els primers paràgrafs d’aquesta introducció: la superfície de la terra emergida és una tercera part de la superfície coberta per l’oceà, i la producció mitjana per unitat de superfície és tres vegades més gran a la terra que a l’oceà. És a dir, la producció anyal total de la biosfera, que s’estima diversament segons els autors però amb una notable convergència entorn dels 66 000 milions de tones de carboni per any, es distribueix, si fa no fa equitativament, entre els continents i l’oceà. L’oceà, doncs, per unitat de superfície, és crònicament molt menys productiu, i aquesta serà una constant sempre present en la nostra consideració de la vida marina.
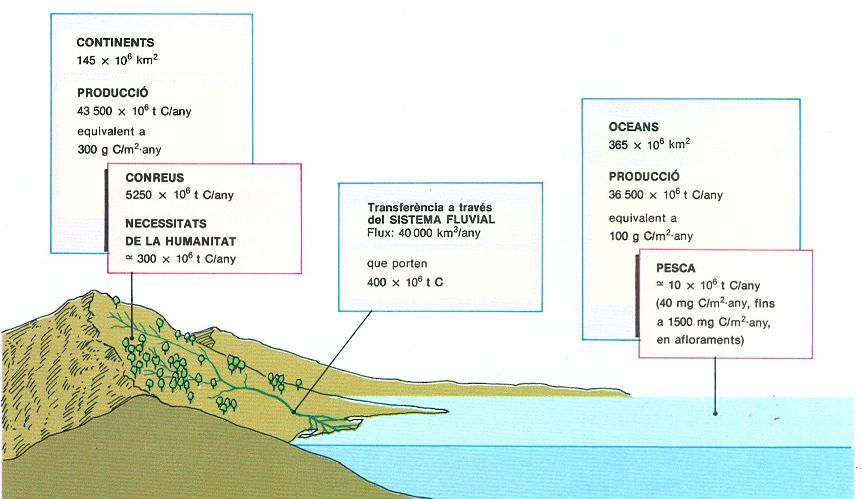
Valors globals de la producció primària en els continents i en els oceans, i transferència dels primers als segons que passa a través dels sistemes epicontinentals (aigües dolces) i maresmes.
Román Montull, original de l’autor
La producció primària és el resultat de l’adquisició de poder reductor per part de les cèl·lules vegetals, gràcies a l’energia de la llum solar. El poder reductor s’aplica a compostos químics de l’entorn i, així, per exemple, de manera principal, però no única, el CO2 passa a formar carboni orgànic, que és la forma en què, com hem vist, generalment s’expressa la producció.
L’aprofitament de la llum del Sol per a la producció orgànica es fa de manera diferent a terra i a l’oceà. A terra, les fulles s’interposen en la direcció dels raigs del Sol, i en part absorbeixen i en part reflecteixen la llum o la difonen en direccions diverses. Tanmateix, dins els boscos, àdhuc en els més densos, hi ha llum. L’alçada més gran de la vegetació terrestre, l’alçada màxima dels arbres, és entorn dels 110 m, però els boscos amb capçada contínua, en les millors condicions de creixement, rarament superen els 30 m d’alçada. La clorofil·la pot absorbir pràcticament tota la radiació efectiva del Sol en concentracions de mig gram de clorofil·la per metre quadrat, que és la concentració habitual en les fulles de les plantes. Així, amb un sol estrat de fulles (extensió de les fulles/extensió de la superfície del sòl dintre la qual cau la seva projecció = índex foliar = 1) n’hi hauria prou. Tanmateix, una gran part de la radiació és reflectida i, per un dels dispositius d’excés de seguretat necessaris a la natura, els índexs foliars de la vegetació terrestre acostumen a ser molt superiors a la unitat, generalment entre 4 i 10. Això fa que, fins i tot en les millors condicions, no tota la clorofil·la pugui funcionar simultàniament.
A l’oceà les condicions són molt diferents; els organismes productors més importants són microscòpics (la majoria fan entre 1/100 i 1/10 de mil·límetre) i no hi ha la distinció entre les fulles, que són els òrgans de producció, i les tiges i els troncs, que són els òrgans de suport i transport. Es pot dir que els principals productors marins són equivalents a les fulles, soltes i molt petites. Més endavant parlarem d’altres organismes vegetals marins, costaners, algues i fanerògames marines, que s’assemblen més a les plantes terrestres convencionals; fet i fet, llur contribució a la producció marina total deu representar entorn d’un 3% i, certament, no passa d’un 5%. La major contribució a la producció primària oceànica procedeix dels petits organismes en suspensió a l’aigua que, col·lectivament, formen l’anomenat fitoplàncton, una paraula formada d’arrels gregues, com gairebé tots els mots emprats en ciència: fito, que vol dir planta, fa referència a la condició vegetal, i plankton ve d’un verb que vol dir «derivar» o «errar a la ventura», com Ulisses, o com l’intel·lecte errant («plankton noon») en Parmènides. A diferència de l’aire, l’aigua absorbeix fortament la radiació solar, i la llum necessària per al manteniment de la vida vegetal desapareix ràpidament anant avall. Això és una propietat de l’aigua i no dels materials dissolts o de les partícules en suspensió que pugui contenir. No obstant això, si hi ha plàncton o altres partícules en suspensió, la llum és absorbida o s’extingeix molt més aviat, igual que passa amb les fulles en un bosc, de manera que l’ús de la llum és limitat. En tot cas, perquè la llum sigui utilitzada, cal que el fitoplàncton se situï a les capes superiors, on la mateixa absorció de l’aigua no l’hagi extingida. Això limita la producció marina a un gruix d’aigua superficial de 50 a 100 m només i, potser, és més que una coincidència el fet que es tracti d’una longitud semblant a l’alçada màxima dels arbres a la vegetació terrestre. Val a dir, doncs, que en tot el globus, uniformement, la utilització de la llum solar per a la vida es fa en un gruix que només és com 10−5 vegades el radi de la Terra, una pel·lícula relativament minsa si pensem que ha fet funcionar tota la vida i ha portat endavant l’evolució. La quantitat de clorofil·la que el fitoplàncton conté per unitat de superfície acostuma a ser molt inferior a la clorofil·la de la vegetació terrestre. Valors entre 0,01 i 0,1 g de clorofil·la/m2 són d’allò més freqüents. De tot això, en parlarem amb més detall més endavant.
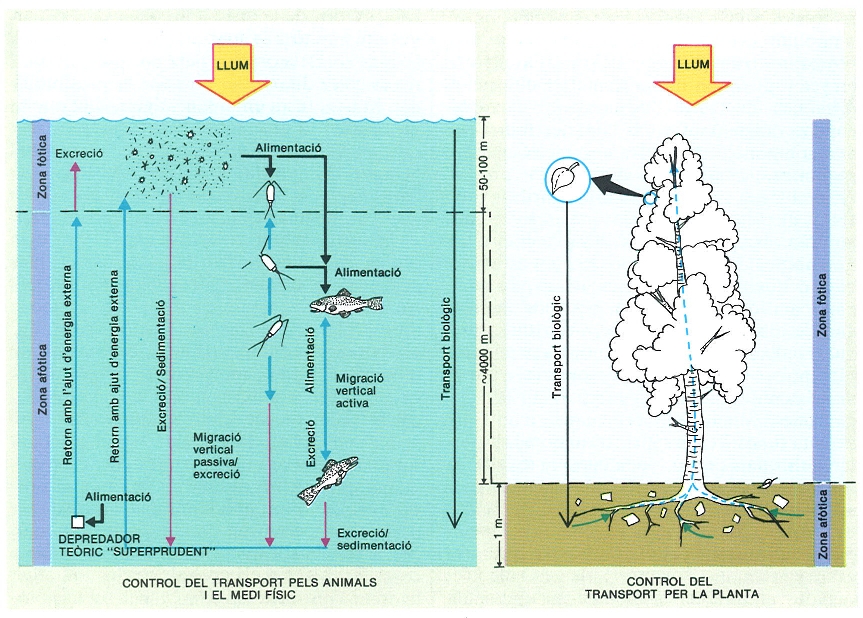
El funcionament del plàncton no és pas molt diferent del del bosc. Sempre es necessita energia externa per a fer pujar el nodriment químic a l’estrat il·luminat o zona fòtica i sota l’acció de la gravetat els elements esmentats tornen a nivells baixos. En la mar, hom podria imaginar l’existència d’un predador teòric «superprudent» que menjaria a la zona profunda i excretaria a la superfície; però aquesta activitat seria impossible de continuar sense tenir accés a l’energia externa.
Ricardo Génova, original de l’autor.
Potser, la diferència més important entre la vegetació terrestre i la vegetació planctònica és que la primera conté una gran part de materials relativament inactius, encara que igualment importants, a les tiges i els troncs, cosa que no té una analogia en el cas del plàncton. Com que el mecanisme de suport i de transport és fet de materials relativament resistents, té una taxa de renovació força més lenta que la part productora, que són les fulles, i uns i altres tenen una taxa de renovació més lenta que la del plàncton. Quant a la part vegetal, doncs, els ecosistemes marins tenen una taxa de renovació (P/B) molt més alta que els terrestres. Aquest fet, conegut de fa temps, havia fet néixer esperances exagerades en el futur dels conreus d’algues. Val la pena de parlar-ne una mica més, perquè té un valor didàctic amb relació a la comprensió de les diferències entre sistemes terrestres i sistemes marins.
Un organisme del fitoplàncton es pot dividir una vegada cada dia; això vol dir que, en un dia, una determinada massa vegetal produeix una biomassa nova igual a ella mateixa. És una velocitat de multiplicació insòlita en la vegetació terrestre. A més, les algues del plàncton són més riques en proteïnes que una planta que conté una gran part de tiges, arrels i troncs. Per això, s’ha estudiat la possibilitat del conreu d’algues planctòniques per a la producció de proteïna. En condicions naturals, però, la productivitat de les algues del plàncton és molt limitada, per causa de l’exhauriment del nodriment mineral en les aigües il·luminades i per la mateixa sedimentació de les algues, que les porta fora dels estrats on poden rebre llum. Si, no obstant això, volem aprofitar la capacitat de producció que sembla característica dels organismes unicel·lulars en un sistema artificial de cultiu, ens trobem immediatament amb un seguit de dificultats que es poden resoldre d’una manera consecutiva amb l’aplicació de diversos recursos tècnics: ens caldrà mantenir els organismes en un líquid il·luminat, agitat i amb un subministrament suficient dels diversos elements químics necessaris; caldrà també retirar una part del material produït, no solament perquè és el material que ens interessa, sinó per aconseguir que la resta mantingui unes condicions adients de concentració. Fet i fet, haurem construït un arbre tècnic o tecnològic, que és una rèplica del que ha fet la natura en la construcció evolutiva de l’arbre espontani. Una part molt important dels vegetals terrestres té, doncs, la missió de fer de suport, diguem-ne «tecnològic», d’un sistema de producció que és relativament petit, però equivalent al sistema total de producció del plàncton. Però, en el plàncton, les funcions de suport i transport queden confiades a l’aigua i als animals, i ens fan l’efecte que són poc organitzades. Aquest serà un punt molt important a considerar.
Les condicions de producció a la terra i a la mar
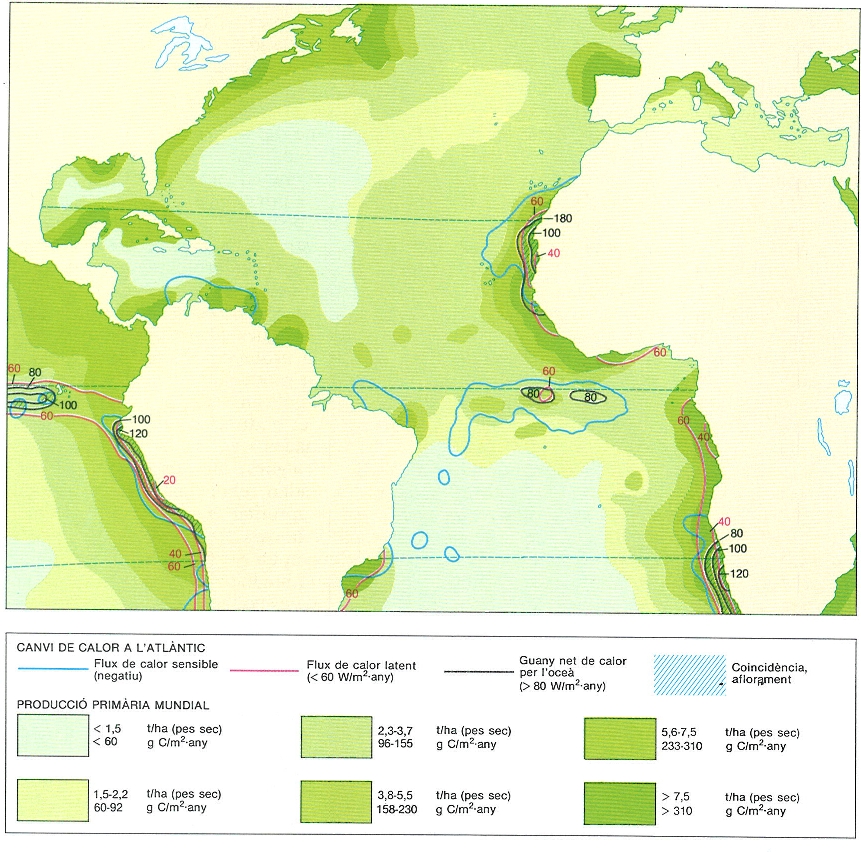
Mapa del canvi de calor de l’Atlàntic i dels valors de la producció primària mundial (a baix). En el mapa s’observa com se superposen les àrees on el guany net de calor per l’oceà excedeix els 80 W/m2 amb aquelles on el flux de calor sensible és negatiu (l’aigua s’escalfa a la superfície o en passar a la superfície) i amb aquelles on el flux de calor latent (calor invertida en evaporar aigua) és inferior a 60 W/m2. Les àrees d’aflorament tenen un canvi d’energia elevat que es manifesta en un escalfament considerable i una evaporació relativament baixa.
Carto-Tec, original de R. Margalef i M. Estrada.
Abans hem donat una estimació de la producció primària total de la Terra no més enllà de 70 000 milions de tones de carboni orgànic assimilat per any. Coneixent l’energia —química— que entra en els compostos formats, resulta que la producció primària de la biosfera és entorn de 100 o poc més bilions de watts (100 a 110 × 1012 W). Ara bé, l’energia del Sol que és interceptada per la Terra és de 178 × 1015 W, de manera que és clar que la biosfera, com a sistema viu, no empra més que una fracció molt petita, menys de l’1‰, de l’energia que arriba del Sol. La primera impressió sol ser de sorpresa davant de la poca avidesa de la vida, que només fa servir una fracció tan petita d’aquest bé de Déu que prové del Sol. Més endavant, potser, pensarem que la vida «s’interessa» més per l’ocupació de la Terra amb una estructura molt rica, i no empra més energia que la necessària. En tot cas, de l’energia no utilitzada a través de la producció primària, una part és reflectida directament vers l’espai, i l’altra actua diversament en l’enorme màquina tèrmica constituïda per atmosfera i hidrosfera, i aquesta part, que és la major, no és indiferent respecte al funcionament de la biosfera i al manteniment de la vida. De l’energia que passa efectivament a través de la producció primària, en podem dir energia endosomàtica o interna, ja que circula a la intimitat dels organismes. De l’altra energia, que és significativa en la biosfera, en podem dir energia exosomàtica o externa.
La vida depèn de les característiques globals del nostre planeta, i aquestes estan condicionades, entre moltes altres coses, pel flux d’energia solar: per la recepció d’energia del Sol, per la degradació i els bescanvis d’aquesta energia, principalment en les cobertes fluides que són constituïdes per atmosfera i hidrosfera, i per l’emissió d’energia vers l’espai, amb un espectre canviat amb relació al solar, en el sentit d’un allargament mitjà de la longitud d’ona —que correspon a una «pèrdua de qualitat» de l’energia—. Aleshores, la producció primària pot arribar a ser directament proporcional a aquesta energia externa i a la seva efectivitat.
El plàncton marí n’és un exemple excel·lent. Els organismes del plàncton tenen o arriben a tenir una densitat superior a la de l’aigua i, naturalment, acaben sedimentant-se. Els animals del plàncton sovint mengen més a prop de la superfície de la mar i excreten a una fondària més gran i, d’aquesta manera, contribueixen, ells també, a moure cap avall els elements químics que havien estat assimilats en els estrats més superficials. D’una manera general es pot dir que, per a un àtom definit, el fet de passar de formar part d’una substància dissolta en l’aigua a formar part d’una partícula sòlida i, per tant, d’un organisme, comporta una probabilitat definida d’anar a parar a un nivell més profund que aquell on fou assimilat, on és alliberat de nou. Els organismes del plàncton es podria dir que treballen contra la seva pròpia persistència, perquè transporten els elements necessaris per al seu nodriment (carboni, fòsfor, nitrogen, silici, etc.) de les capes il·luminades a les aigües profundes on no arriba la llum. La situació final, previsible, és que on hi ha llum no hi ha nodriment, i on el nodriment s’ha acumulat, no hi ha llum per a poder-lo mobilitzar. Aquesta tendència a una segregació dels reactants, que porta a una relativa immobilització de la vida i de la seva activitat, correspon, en certa manera, a la dita popular «Déu dóna faves a qui no té queixals». En el funcionament dels sistemes naturals, sembla també que les coses estan mal repartides i que tots els processos s’aturen tard o d’hora, perquè sempre hi falta alguna cosa.

La productivitat de la Mediterrània i dels subdeserts és aproximadament la mateixa. Això significa que aquesta bosquina litoral de les dunes de Guardamar (País Valencià) fixa per metre quadrat i any la mateixa quantitat de carboni que la mar immediata. La productivitat dels oceans és més elevada, com també ho serà la de la pineda incipient que hom ha plantat en aquestes dunes.
Jordi Vidal.
Tanmateix, hi ha un retorn de nutrients als estrats il·luminats, però limitat, i tan decisiu que d’ell depèn la producció primària. És en aquest sentit que dèiem que l’energia externa o exosomàtica és important, perquè sovint l’energia directament implicada en aquest retorn es pot mesurar. Potser avui tenim consciència de la importància d’aquesta energia externa per dues circumstàncies que comporta la nostra civilització; d’una banda, veiem que no solament emprem l’energia de la nostra dieta —les tres mil quilocalories al dia, o els 140 watts que utilitzem directament en el nostre metabolisme—, sinó també una energia externa a la cuina, la calefacció, la il·luminació, el transport i moltes altres necessitats; es tracta d’una energia externa que depèn del tipus de civilització, però que, per a nosaltres, que llegim llibres, viatgem i mirem la televisió, queda entre 10 i 100 vegades l’energia interna. D’altra banda, l’agricultura mecanitzada ens fa veure com el rendiment de les collites depèn d’una energia suplementària emprada en el regatge, el treball de la terra, l’addició de fertilitzants i l’eliminació de plagues, afegida a l’energia «natural» —però també externa— de la pluja.
En els ecosistemes continentals, l’energia externa més important és la implicada en el cicle de l’aigua, l’evaporació, i la pluja, que demana unes 500 calories per gram d’aigua. Sabem que les plantes depenen de l’aigua, però no pensem tan sovint que aquesta és mobilitzada i posada a disposició de les plantes per mitjà de la màquina de l’atmosfera i de la circulació de les aigües. Tant en un cas com en l’altre, tant en el plàncton com en la vegetació terrestre, l’energia exosomàtica més directament esmerçada a suportar la producció és gairebé de 10 a 100 vegades l’energia endosomàtica de la producció biològica, i l’energia exosomàtica que es relaciona amb la producció i el suport dels sistemes vius és tota la del Sol, o en la seva major part, que és, com ja hem dit abans, almenys 1000 vegades superior a l’energia de la producció primària.
Els ecosistemes terrestres són confinats per les capes superficials de la Terra. Totes les parts mortes de la vegetació i dels animals es reciclen en el gruix del sòl, que fa, a tot estirar, només un metre. Allí hi ha una vida molt intensa, i la circulació de l’aigua en el sòl i les arrels permeten el retorn, relativament ràpid, dels materials en el cicle de la vida. En els oceans, l’àmbit en el qual té lloc la descomposició del material i el retorn a la forma inorgànica, és relativament molt més vast: enfront del metre escàs del sòl, tenim els quatre mil metres i escaig de fondària mitjana dels oceans mundials. No és estrany que el retorn d’aquests materials posi un fre als cicles naturals i que la producció per unitat de superfície en l’oceà sigui solament un terç del que és a la superfície dels continents. Com ja s’ha dit, les vies de retorn són sota el control de la vida en els sistemes terrestres molt més que en els marins. A la mar l’aigua es pot barrejar verticalment, o bé ascendir en àrees limitades (àrees de surgència o d’aflorament) per causes «naturals», és a dir, pel treball produït per l’energia exosomàtica dels moviments de l’aigua. A terra, malgrat que l’energia de l’evapotranspiració també és externa, els organismes proporcionen una canalització que la fa més efectiva. Ja aprendrem a calcular, a la mar, el flux vertical necessari per a mantenir una producció.
D’una manera aproximada, podem posar un exemple que ens orientarà quant a magnituds. Una producció de 100 g de carboni per m2 i any necessita 2,5 g de fòsfor, i l’elevació en uns 400 m (una mica més d’un metre diari) d’una columna d’aigua que permetí una diferència de 6 mg de fòsfor per m3 entre l’aigua «rica» i l’aigua «exhausta». —400 × 6 mg = 2,4 g—, unes xifres adients per a una primera interpretació de la nostra Mediterrània. A terra podrem tenir una producció tres o més vegades més gran i el vehicle també ha de ser l’aigua. Si suposem que l’1% de l’extensió del camp o del bosc és representat per les tiges, i que 1’1% d’aquestes tiges és una secció activa en el transport (el xilema dels arbres), solament l’1 per 10 000 de la superfície terrestre és activa en l’ascensió de l’aigua procedent del sòl, aigua que és una solució diluïda dels elements essencials. Equival a un metre quadrat en una hectàrea. I la velocitat en els vasos de les plantes ha de ser, per tant, almenys deu mil vegades més gran que la velocitat ascensional en una aigua marina fertilitzada de manera equivalent, és a dir, no un metre per dia sinó decímetres per minut.
La màquina de produir biomassa

Les concentracions d’ocells oceànics, com aquesta colònia nidificadora de mascarells del Cap (Sula capensis) a Lambert’s Bay (regió sudafricana de clima mediterrani), són possibles gràcies a l’extensa zona marina que exploten com a àrea alimentària, zona marina que, d’altra banda, és molt fèrtil per l’existència de surgències d’aigües profundes. Si, com els ocells terrestres, aquests mascarells nidifiquessin en els seus territoris de caça (de pesca, en aquest cas), es trobarien dispersos en una àrea amplíssima, proporcional a la productivitat marina real.
Jaume Altadill.
Resulta alliçonador comparar el retorn dels elements a les capes il·luminades de l’oceà per moviments més o menys desordenats, amb el retorn d’aquests a les parts superiors de les plantes, d’una manera regular mitjançant un mecanisme de transport molt més ordenat, en forma de columnes paral·leles. Entre altres coses, ens permet de copsar diferències entre mecanismes diversos, uns que s’assemblen més a màquines fetes de parts rígides, com ara la vegetació, i altres que es podrien comparar, més aviat, amb els rellotges tous de Dalí. Sembla que una màquina feta de parts toves ha de ser molt poc efectiva en la transmissió a distància d’una energia i en la realització d’un treball. I, precisament, el medi oceànic té aquestes característiques. És molta l’energia que es degrada o s’esmerça en els moviments marins, sense realitzar un treball prou efectiu, que es manifesti, per exemple, en el sosteniment d’una elevada producció dels sistemes vius. Però no hem parlat dels rellotges tous de Dalí debades: cal recordar que, encara que tova, un fluid té una estructura que li dóna la qualitat de funcionar com una màquina i, a més, de funcionar amb discontinuïtats. Tots estem familiaritzats amb els fronts atmosfèrics que produeixen els canvis de temps, ben sobtats de vegades. I l’oceanografia ens mostra tota mena de mecanismes marins, tot fent-nos veure com les diferències de massa —densitat— en l’aigua continuen acoblades a sistemes de moviment i de transport, amb una gran riquesa de situacions que poden ser més o menys estacionàries. Des del punt de vista de la producció marina, els elements més importants d’aquesta estructura oceànica mundial són els sistemes d’aflorament a llevant dels més importants girs oceànics, a les costes del Perú, Namíbia, Califòrnia i el Sàhara, com també l’aflorament que segueix la línia equatorial. Tot plegat, això representa injectar a la zona fòtica o il·luminada dels oceans almenys la meitat del fòsfor que cal per a mantenir la producció de tots els oceans. Una part d’aquest fòsfor és reciclat diverses vegades en les capes superiors i hi ha, a més, altres aportacions de menor importància que complementen el total utilitzat.
Hi ha, doncs, llocs preferents de fertilització, que sovint tenen la forma d’estructures allargades, com els fronts marins allunyats de la costa, comparables als fronts atmosfèrics. Aquests fronts són, relativament, menys dinàmics, per la major densitat i la major capacitat calorífica que té l’aigua amb relació a l’aire. També són efectives estructures d’una altra mena, com ara les que resulten de la interferència de la inèrcia del flux marí i la topografia costanera. Sembla evident que, d’una manera o d’una altra, les màquines generen treball —en aquest cas un treball que dóna una producció biològica— en les discontinuïtats, en els límits entre llurs peces mòbils. En tindrem exemples excel·lents en un estudi més acurat de la producció marina a les nostres aigües. Ara, cal dir solament que aquest fet ha estat reconegut i distingit recentment amb la proposta d’anomenar ergoclines (superfícies on es produeix treball) aquelles interfases o discontinuïtats sobtades on, per raons diverses, es donen condicions apropiades per a la ràpida multiplicació dels organismes. Es tracta, simplement, de posar en contacte els diferents ingredients d’una reacció natural: llum, elements químics necessaris per a la vida i llavor, és a dir, alguns organismes; la productivitat es dispara sola. Evidentment, el treball es manifesta en una degradació de l’energia, per exemple de l’energia cinètica dels corrents, que es dissipa en forma de moviment turbulent. És, doncs, apropiat d’associar la turbulència amb una producció elevada, però podríem considerar la turbulència tant una propietat que permet una difusió del nodriment cap a les cèl·lules, com la manifestació final d’una degradació d’energia en un moviment que ha aproximat entre ells els ingredients necessaris per a una reacció. En resum, aquesta manera de veure les coses fa créixer l’interès que pugui tenir una anatomia funcional de l’oceà, és a dir, la dissecció de l’oceà en masses d’aigua i l’estudi dels seus moviments respectius; aquest és el veritable suport de la productivitat marina i conté l’explicació fonamental de les diferències locals.
En un sentit una mica diferent, però en el fons semblant, es pot dir el mateix de la superfície dels continents. També ací hi ha una estructura que concentra la producció en llocs preferits. Aquesta estructura és més rígida si és la que s’expressa en la topografia del sòl. És evident que el sistema fluvial canalitza aigua i recursos —nodriment dissolt en l’aigua— en àrees preferents, de tal manera que s’ha reconegut que la conca hidrogràfica és la unitat natural per a l’estudi de processos de producció a terra. D’altra banda, el pas dels fronts atmosfèrics, llur freqüència i efectivitat, condicionaments del clima local, defineixen també altres aspectes de la productivitat. Ja ha quedat clar que el concepte d’anatomia funcional de l’entorn s’aplica tant als oceans com a la superfície dels continents i, malgrat que les estructures significatives poden arribar a dimensions mínimes, les que són realment efectives, les úniques que manegen quantitats importants d’energia, són les de dimensions compreses entre 100 i 10 000 km. Són, per tant, estructures idònies per a ser estudiades sinòpticament des de l’espai, mitjançant tota la tecnologia moderna basada en la interpretació de senyals captats des dels satèl·lits artificials, senyals que són emesos des de la Terra en una sèrie de bandes de diverses longituds d’ona. Dins l’ecologia moderna hi ha, doncs, un espai científic molt interessant, en l’acoblament entre la captació i la interpretació de senyals des de l’espai, i el reconeixement del significat —que passa a través del concepte d’energia externa o exosomàtica— que té una anatomia funcional de l’entorn.
Els ecosistemes epicontinentals
En superposició a moltes d’aquestes estructures hi ha, òbviament, sistemes de transport sobre grans distàncies. Ja que hem parlat tant, de manera comparativa, de sistemes terrestres i sistems marins, cal fer una referència, encara que sigui breu, als sistemes epicontinentals o d’aigües dolces, que, a més de constituir un tipus propi i important d’ecosistema, estableixen un pont que uneix els sistemes terrestres amb els marins. De fet, la part més important d’aquest transport és a càrrec dels rius que, si bé contenen una fracció molt petita de l’aigua que hi ha a la superfície de la Terra —probablement no molt enllà d’una milionèsima del total—, tenen una importància incomparablement més gran, fins i tot per a la vida humana. De fet, els llacs s’han de considerar com a diverticles dels rius, interessants, això sí, però potser no gaire més importants que els processos de segregació i de sedimentació que s’hi realitzen. De l’aigua de pluja que cau a la superfície dels continents, dues terceres parts tornen a l’atmosfera per evaporació directa o per la transpiració de les plantes, i la resta, que és d’uns 40 000 km3 l’any per a tot el món, va a mar. Aquestes aigües pluvials porten tota mena de materials en solució, com també moltes partícules en suspensió. Tot això té una importància indubtable sobre la vida marina, principalment la costanera, i s’haurà de tenir en compte. No obstant això, pot ser principalment significatiu el fet que els rius porten a mar aproximadament un 1% de la producció primària continental (en forma de matèria orgànica morta) i almenys 300 milions de tones de carboni orgànic anyalment. Aquesta xifra certament fa pensar, si recordem que el producte total de la pesca mundial no va més enllà d’uns 10 milions de tones de carboni per any (uns 100 milions de tones, pes fresc). De fet, els oceans funcionen com a heterotròfics amb relació als continents: digereixen i respiren part de la matèria sintetitzada als continents. Per tant, des del punt de vista del bescanvi d’oxigen, i pel que fa a l’atmosfera, els oceans més aviat consumeixen oxigen que no pas en produeixen, almenys en els temps actuals.
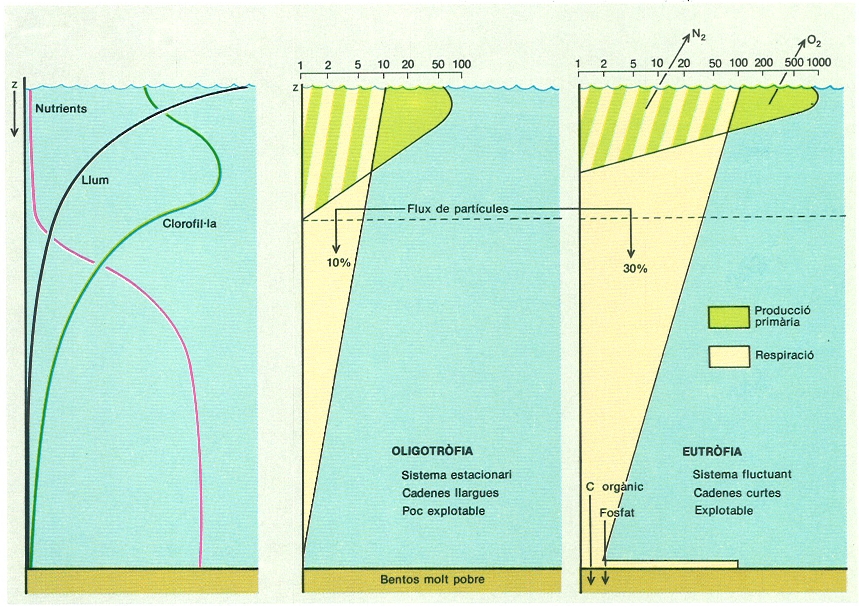
Model vertical de segregació dels elements de producció, llum, cèl·lules i nutrients, al cap d’un temps d’estabilització creixent (a l’esquerra), i distribució vertical dels valors de producció i respiració totals en escales horitzontals logarítmiques, mostrant les diferències entre sistemes eutròfics i oligotròfics. La mar en general, i la Mediterrània especialment, és oligotròfica i la pressió que rep dels continents no arriba a forçar-la. El gràfic de l’esquerra expressa també la tendència a fer mínima la relació entre el grau de coincidència de les distribucions dels factors de producció i la biomassa total mantinguda.
Ricardo Génova, original de l’autor.
Fent de pont entre els sistemes continentals i els sistemes marins, els rius, en general, treballen sota tensió —i amb presses—, i són molt efectius en la tasca de contribuir a l’atmosfera amb oxigen i nitrogen (desnitrificació), i als sediments amb carboni orgànic i amb fòsfor. Aquesta separació és característica dels sistemes que funcionen sota una càrrega més gran, que són els anomenats eutròfics. Ho són sovint les maresmes costaneres, però rarament es pot qualificar d’eutròfica una mar, si no és en casos comptats, com la Bàltica. Però, en algunes mars, aquestes tendències són mínimes, com s’esdevé en el cas de la Mediterrània, que es pot qualificar d’oligotròfica, que vol dir pobra o, encara més pròpiament, no carregada de fora amb una tensió excessiva.
Els ecosistemes pelàgics i els ecosistemes bentònics
El contrast entre els sistemes continentals, organitzats principalment entorn de la vegetació, i els sistemes oceànics és especialment marcat pel que fa als sistemes pelàgics, que són pròpiament els de les aigües lliures, i centrats en el plàncton. Però, certament, hi ha una vida costanera i de fons, subjecta o dependent del substrat sòlid de les costes i del fons, que, per això, semblaria més comparable amb els sistemes terrestres. De fet, hauríem de fer algunes distincions.
En primer lloc, la vida vegetal és limitada per la llum, però el manteniment de la vida vegetal demana un equilibri subtil entre la taxa de multiplicació —dependent de la llum i del nodriment— d’una banda, i les partides negatives, constituïdes per la respiració pròpia, el consum per part d’animals i la pèrdua per sedimentació que pot portar els organismes fora de l’àmbit de la llum. Es comprèn que organismes vegetals de fons que no poden sedimentar-se més avall, que tenen pocs enemics i un metabolisme baix, com les algues roges incrustants, puguin sobreviure a fondàries que pràcticament són el doble de les màximes fondàries en les quals viu el fitoplàncton: posem 250 i 120 m, respectivament. Hi ha, doncs, una vida vegetal de fons, o bentònica, representada per algues superiors, principalment rodofícies i feofícies, i per diverses fanerògames o plantes de flor, que també són anomenades «algues» per la gent. És clar que no sempre aquesta vegetació s’estén fins a les fondàries màximes esmentades abans, però pot ser important fins a uns 50 m i més. Es tracta d’organismes en els quals pot haver-hi trasllat de substàncies d’unes parts a les altres, però que, d’altra banda, pràcticament extreuen la major part del nodriment que necessiten de l’aigua.
En els sistemes marins, el sediment acostuma a ser pobre, i no estrictament comparable a un sòl terrestre, encara que sovint, especialment amb relació a maresmes i estuaris, els sediments emmagatzemen elements químics necessaris i intervenen en un reciclatge important, i no solament per mitjà de l’activitat dels organismes bentònics. De totes maneres, cal recordar que, a escala mundial, només el 7% de l’extensió marina té menys de 200 m de fondària, i que l’extensió colonitzable pels productors primaris del bentos és molt més migrada. Però una cosa és certa: aquesta vegetació té menys pèrdues que el fitoplàncton, i la relació P/B és més baixa. Encara que la vegetació litoral pugui arribar a ser impressionant com és el cas de les grans feofícies, en conjunt, la contribució a la producció marina és baixa, ja que segurament se situa entre el 3 i el 5% del total.
D’altra banda, per sota de l’estrat productiu dels oceans hi ha un espai grandiós que no és exempt de vida, però aquesta vida, ja sia en forma de bacteris, o d’animals, depèn estrictament del material produït o sintetitzat a les capes fòtiques o il·luminades. Una gran part dels habitants són pelàgics, és a dir, es troben perpètuament en suspensió. Però també hi ha organismes que reposen directament en el substrat o sediment profund, i que, com és natural, obtenen el seu nodriment del «mannà que cau del cel», és a dir, de l’excés de producció del sistema pelàgic. Podríem fer, també, un balanç del sistema litoral; sembla que, generalment, aquest importa més aliment del plàncton que no pas exporta matèria orgànica cap al plàncton. Fins i tot moltes espècies litorals que tenen larves pelàgiques podríem dir que envien els seus infants «a dida», a explotar el plàncton, si bé aquest bescanvi s’ha d’analitzar amb més cura i admet altres interpretacions complementàries. El que volem assenyalar ací és que, en la relació de bescanvi mutu entre els sistemes pelàgics i els bentònics, en general els bentònics hi guanyen.