
El llac de Banyoles en visió aèria, amb la seva forma característica.
Jordi Vidal
Els llacs es caracteritzen perquè mantenen una quantitat relativament gran d’aigua respecte de la que hi entra o en surt. La seva taxa de renovació és d’un any com a mínim, en mitjana. D’altra banda, el nivell de l’aigua no sol variar gaire, per la qual cosa s’hi desenvolupa un cinyell de vegetació, si les condicions climàtiques ho permeten. Aquesta característica els diferencia dels embassaments. Als Països Catalans, els llacs són relativament petits i emmagatzemen poca aigua. La seva forma, el volum i les seves característiques limnològiques estan relacionats amb el seu origen geològic.
L’origen geològic dels llacs o estanys és molt variable. Als Països Catalans podem assimilar-los a dos tipus principals: d’una banda, els estanys pirinencs, d’origen glacial, excavats en circs o bé formats per les morrenes terminals de les glaceres; i de l’altra, els llacs d’origen càrstic. Els estanys d’origen glacial són de formació recent (com a màxim tenen 10 000 anys) i es distribueixen pels Pirineus en diferents nuclis; són d’aigües fredes i oligotròfiques. Els llacs càrstics són d’origen més antic (uns quants milions d’anys) i se situen en zones on els guixos són abundants; tenen forma arrodonida, de dolina, i les seves aigües són força mineralitzades. Tractarem separadament, doncs, els dos tipus de llacs, ja que el seu funcionament i els organismes que hi viuen són molt diferents.
Els estanys pirinencs

Els llacs de muntanya constitueixen un món molt peculiar dins els ecosistemes limnètics dels Països Catalans, tant per la seva abundància (n’hi ha més d’un miler en tots els Pirineus) com per les característiques de les seves aigües, la flora i la fauna. La majoria són de dimensions mitjanes o petites, sense vegetació a les vores, com el de la fotografia, de Sotllo (Pallars Sobirà), i resten coberts d’una capa de neu i gel durant una gran part de l’any.
Jordi Catalan
Durant les glaciacions del Pleistocè, a més de la gran placa de gel del N d’Europa, que s’estenia d’Escandinàvia a Sibèria, les serralades altes situades més al S desenvoluparen el seu propi sistema glacial. Els estanys dels Pirineus són resultat de l’acció erosiva i remodeladora del gel en aquell període i constitueixen estrictament l’únic districte lacustre dels Països Catalans. S’estenen des de la Cerdanya fins a l’extrem W dels Pirineus, i la màxima densitat la trobem a la Vall d’Aran i al Pallars. A la part catalana dels Pirineus ja n’hi ha més de quatre-cents. La seva distribució és desigual, d’acord amb les condicions topoclimàtiques, però en general són més abundants al vessant N de les valls. Pel que fa a l’altitud, al vessant S se situen, de mitjana, uns 100 m més amunt que al N, cosa que reflecteix la diferència de nivells de les neus perpètues durant el període glacial.
Els estanys pirinencs i el seu entorn són de tots els nostres paratges els que més s’identifiquen amb el medi natural; no obstant això, la mà de l’home com a agent modificador no hi ha estat absent. Des d’antic l’activitat ramadera als Pirineus ha comportat l’eutrofització d’alguns estanys i basses utilitzats per a abeurar el bestiar. Actualment l’eutrofització també pot ser deguda a l’augment de l’excursionisme i del turisme en els estanys més accessibles.
De més abast encara ha estat la influència de l’aprofitament hidroelèctric. Molts llacs han estat modificats, simplement fent-hi una presa per augmentar-ne el volum o foradant-los per intercomunicar-los, i darrerament s’hi han fet galeries de circulació reversible. L’oscil·lació del nivell d’aigua i el transvasament comporten modificacions dinàmiques i estructurals de la biologia de l’estany: augmenta l’intercanvi entre l’aigua i el sediment i augmenta també la terbolesa deguda a material mineralògic en suspensió; es barregen poblacions d’organismes de diferents estanys; es trenca la congruència tèrmica de l’estacionalitat natural; disminueix el temps de residència global de l’aigua a l’estany i varia la renovació relativa de les capes segons el nivell d’on es pren l’aigua; etc. És probable que tot plegat porti cap a sistemes més eutròfics, amb una dinàmica molt fluctuant que segueix les pautes del règim d’explotació hidroelèctric. Tanmateix, manquen estudis que en valorin les modificacions reals. D’altra banda, l’efecte sobre les comunitats litorals és clar, ja que les oscil·lacions de nivell n’impedeixen el desenvolupament. Darrerament ha aparegut un nou possible agent alterador, la pluja àcida. Els estanys dels Pirineus hi són potencialment més sensibles que altres sistemes aquàtics del país, per la lenta resposta neutralitzadora d’àcids del substrat en què es troben.
Dinàmica, funcionalitat i tipologia
Origen i morfologia
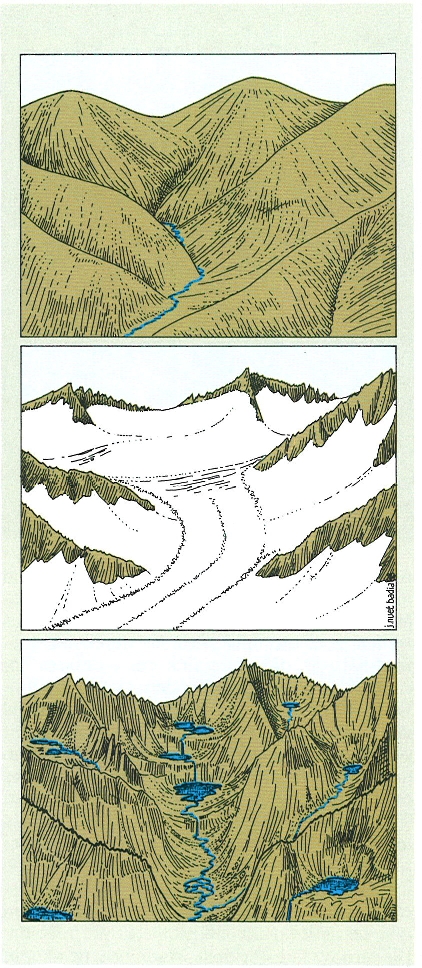
Formació dels estanys pirinencs per sobreexcavació de les glaceres del Pleistocè sobre un relleu fluvial previ. En els circs, zones d’acumulació de neu, es formen estanys arrodonits. En les valls per on baixen les glaceres els estanys són més allargats i poden esglaonar-se formant una sèrie en rosari. En les zones que reben més material sedimentan, acaben perdent les característiques de llac i s’hi desenvolupen mulleres.
Josep Nuet i Badia, a partir de fonts diverses
El principal element diferenciador entre els estanys pirinencs és la morfologia. La resposta biològica dels estanys enfront d’uns mateixos agents externs (vent, precipitació, radiació, etc.) varia segons la forma de l’estany, la fondària, la situació dins la conca, la grandària d’aquesta conca, el pendent de les parets, etc. Molts dels noms que tenen els estanys fan al·lusió a la seva forma (Llong, Redó, Tort) o, indirectament, a la fondària (Negre, Nere). Fent una primera aproximació, hom pot distingir entre estanys de circ, formats a les àrees d’acumulació nival, i estanys de vall, originats per les llengües glacials, sia per sobreexcavació o per obturació per una morena terminal o lateral. Als Pirineus, no hi trobem els grans llacs de vall marginals que hi ha als Alps; els més similars foren dues grans cubetes actualment reblertes de sediment, situades a Bono i Barruera (Alta Ribagorça).
La relació entre l’àrea de la conca i el volum del llac regula alguns aspectes importants de la dinàmica biològica dels estanys. D’una banda, l’entrada de nutrients i d’elements minerals de rebliment a l’estany depèn de l’àrea de drenatge. En aquest sentit, els llacs o estanys de vall, per exemple, que normalment drenen àrees més grans, es rebleixen molt més de pressa, són de menys fondària i tenen processos deltaics importants, alhora que són també més productius. El volum del llac regula la sortida, de manera que el temps de residència de l’aigua a l’estany disminueix amb el quocient «àrea de la conca / volum del llac». El temps de residència pot oscil·lar des d’uns quants dies, en els llacs de vall de menys volum, fins a cinc o més anys, en els llacs de circ de més volum. Lògicament, com més renovació de l’aigua hi ha, més influïble és la dinàmica del sistema per les característiques de l’entrada.
La relació entre l’àrea del llac i la fondària determina l’estabilitat de la columna d’aigua davant l’acció de barreja del vent. Els gradients de densitat als estanys dels Pirineus no són gaire importants (puix que els gradients de temperatura tampoc no ho són) i, a més, es donen a temperatures baixes. Per aquesta raó només es forma termoclina estacional, que divideix l’estany en un epilímnion i un hipolímnion diferenciats, en aquells llacs profunds i d’àrea relativament petita, i en se situa a una fondària considerable (10-20 m). En els estanys més soms, l’estratificació estiuenca hi és més irregular i imprecisa. En aquells en què es forma una estratificació en dues capes, l’epilímnion no ha d’entendre’s com a zona fòtica, ja que la transparència de l’aigua és tan elevada que l’hipolímnion està totalment o suficientment il·luminat perquè s’hi donin processos fotosintètics.
Un tercer element morfològic diferenciador és la relació entre el volum en un determinat nivell del llac i la superfície del sediment en contacte amb l’aigua. En molts estanys pirinencs on l’entrada al·lòctona de nodriment és força migrada, la reutilització dels nutrients és important per a la producció. L’intercanvi amb el sediment és, doncs, un factor de primera importància.
La mineralització, el cicle tèrmic i la producció

Variació estacional de la temperatura, la clorofil·la i la saturació d’oxigen a l’estany Redó (Vall d’Aran). S’hi distingeix un període d’estratificació a l’estiu, en el qual el màxim de clorofil·la (productors primaris) és a una fondària intermèdia i l’oxigen produït s’acumula sota la termoclina. Durant el desgel i la tardor l’estratificació es trenca i totes les línies s’uniformitzen en l’espai vertical. A l’hivern, la capa de gel i neu no permet el pas de la llum; hi ha un predomini dels processos respiratoris, que produeixen un esgotament progressiu de l’oxigen, més apreciable prop del sediment.
Carto-Tec, original de l’autor
El substrat rocós que conté els estanys és sobretot granític o metamòrfic; pràcticament no n’hi ha sobre substrat calcari. Aquest fet és important perquè determina un contingut baix en sals dissoltes, una de les característiques comunes de les aigües d’aquests llacs. La conductivitat varia entre 4 i 80 µS/cm, Però són molt comuns els valors situats entre 10 i 30. L’alcalinitat, en la majoria dels casos, és per sota de 0,5 meq/l, i pràcticament tota és deguda a l’ió bicarbonat. El pH es manté prop de la neutralitat (6,5-7,5), tot i que pot presentar oscil·lacions importants durant el cicle anual.
La situació dels estanys en conques petites, despoblades i de sòl poc desenvolupat, fa que l’entrada de sals nutritives —de fòsfor i de nitrogen sobretot— depengui bàsicament de la precipitació. Són llacs oligotròfics, limitats principalment pel fòsfor (menys de 0,1 µM). La relació entre el fòsfor i el nitrogen inorgànic a les seves aigües (1:50-5000) és lluny de la dels organismes (1:16-30). L’oligotròfia i la manca d’entrades importants de materials mineralògics de gra fi fan que l’aigua dels llacs sigui molt transparent, cosa que dóna un color blau fosc als de més fondària. Tret dels de més profunditat, la majoria reben prou llum a les parts fondes perquè s’hi realitzi fotosíntesi.
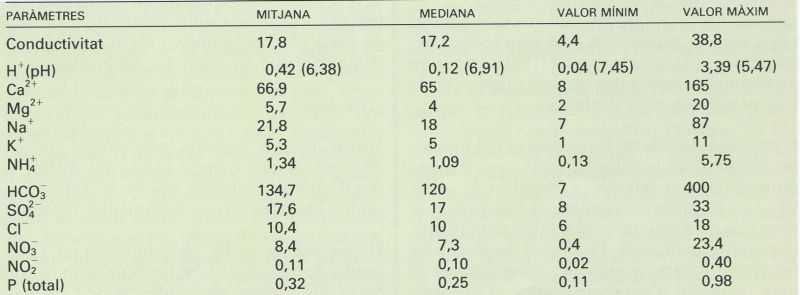
Paràmetres químics principals de l’aigua d’una setantena d’estanys pirinencs, des de la Cerdanya fins a la vall d’Aran (s’han exclòs els de particularitats locals molt acusades). Les concentracions es donen en µM i la conductivitat en µS/cm.
Dades de J. Catalan, E. Ballesteros, L. Camarero, O. Delgado, E. Gacia i A. Palau
Tèrmicament, els estanys segueixen la pauta del clima pirinenc. Durant bona part de l’any són glaçats: de quatre a vuit mesos, depenent de la seva altitud, la seva grandària i la seva orientació. En aquest període hi ha un gradient invers de temperatura sota el gel; la temperatura més alta, de 4°C com a màxim, es dóna prop del fons. Després del desglaç l’aigua s’escalfa ràpidament, tot i que rares vegades sobrepassa els 15°C a la superfície. La columna d’aigua tendeix a estratificar-se en gradients de densitat, a causa de la distribució desigual de la calor al fons. A partir de l’agost els estanys perden calor cap a l’atmosfera. La isotèrmía de la columna d’aigua de tardor dura més que la de la primavera, i es prolonga pràcticament fins que es glacen els estanys entre novembre i desembre.
Per les seves característiques oligotròfiques, de temperatura baixa i d’enfosquiment hivernal a causa de l’absorció i la reflexió de la llum en la coberta de neu, són llacs poc productius. La seva producció primària és probablement inferior a 40 g C/m2any, incloent-hi plàncton i bentos.
La coberta hivernal de gel i neu

La formació d’una capa de gel i neu als estanys pirinencs defineix unes característiques especials per a la vida durant els mesos d’hivern. D’altra banda, la manera com es forma i es fon el gel determina una gran part dels processos de producció del llac a la primavera. Aquesta coberta no és mai uniforme i depèn dels canvis climàtics que es van produint dia rere dia. La fotografia mostra la formació de blocs, a l’època de fusió, a l’estany Redó (Vall d’Aran).
Enric Ballesteros
La pèrdua de calor dels estanys pirinencs a la tardor és tan intensa que tots s’arriben a glaçar. La formació de gel, que és molt transparent i dur, comença per les vores del llac, però en unes quantes hores, especialment en una nit de calma, pot estendre’s per tota la superfície. L’aigua, en glaçar-se, expulsa sals i gasos que conté en dissolució; si el gel es forma molt de pressa, queden ben visibles les bombolles dels gasos atrapades dins el bloc de gel. La pèrdua de calor del llac continua i el gel va creixent en gruix. Amb les fortes nevades s’acumula neu sobre el gel, que, molt menys conductora i menys transparent, fa d’aïllant; així, els intercanvis d’energia entre l’estany i l’exterior es minimitzen, i l’engruiximent del gel s’atura. L’alternança de dies de tempesta amb dies de bon temps fa que la neu sofreixi ben aviat transformacions, que principalment es manifesten en un enriquiment d’aigua a les capes inferiors del tou de neu. D’altra banda, les fluctuacions tèrmiques i el pes de la neu fan que el gel s’esquerdi i que l’aigua del llac envaeixi el gruix de neu, fins a arribar a un equilibri hidrostàtic. A més, les capes de la coberta més riques en aigua, amb aspecte de granissat, si es glacen formen un gel de color blanc, molt menys transparent i menys dur que aquell format a partir de l’aigua del llac. Al llarg de l’hivern tots aquests processos es repeteixen i se superposen, de manera que la coberta es va transformant en un conjunt complex de capes de diferent gruix i de propietats molt distintes.
L’expulsió de sals en formar-se el gel implica un augment de la salinitat en la capa d’aigua immediatament inferior. L’expulsió és selectiva i no té gaire efecte sobre el fosfat, de manera que no s’estimula apreciablement la producció fitoplantònica. Durant el desglaç hi ha un efecte invers de dilució, estimulat per la gran quantitat d’aigua que entra al llac procedent de la fusió de la neu de la conca. L’aigua que entra se situa immediatament sota la superfície del gel, per la qual cosa poden formar-se capes de pH baix (entre 4 i 5), on el contingut de protons pot superar els bicarbonats; es parla llavors d’alcalinitat negativa.
La formació del gel i l’acumulació de la neu modifiquen la dinàmica biològica, ja que fan disminuir el pas de la llum i impedeixen l’acció del vent i l’intercanvi de gasos. Un mateix gruix de coberta té un efecte ben diferent sobre el pas de la llum segons que hi predomini la neu o el gel. Als Pirineus és freqüent trobar molt més gruix de neu que de gel, fet que contrasta amb la situació dels estanys que es glacen a latituds més altes. La manca de llum i d’intercanvi de gasos fa que els processos respiratoris provoquin una davallada de la quantitat d’oxigen de l’aigua sota el gel, que en alguns estanys soms i molt rics en vegetació podria provocar la anòxia en tota la massa d’aigua, amb el conseqüent efecte nociu sobre els organismes de l’estany, de les truites en particular; però encara no hi ha dades documentals que ho constatin als Pirineus.
Tipologia

Els estanys pirinencs més grans i profunds solen tenir l’aigua ben oxigenada durant tot l’any i formar una termoclima estival; alhora, és freqüent que la cubeta tingui les parets molt inclinades, la qual cosa dificulta la colonització de la zona litoral per helòfits. Quan l’home els modifica construint-hi una presa, com ha passat en el de Certescans, que veiem a la fotografia (juliol de 1987), la fluctuació del nivell d’aigua deguda a l’explotació hidroelèctrica contribueix també que no arrelin els helòfits; alhora és probable que la dinàmica natural de les comunitats planctòniques s’alterí per l’augment de la terbolesa i la variació de les pautes d’intercanvi de sals nutritives del fitoplàncton amb els sediments.
Jordi Catalan
Vistos aquests aspectes morfològics, podem distingir, d’una manera ideal, uns quants tipus d’estanys pirinencs, que en la realitat presenten totes les formes intermèdies possibles.
D’una banda, hi ha els estanys grans i profunds, de més de 15 hm3 i més de 30 m de fondària, amb temps de residència de l’aigua de 2 a 5 anys, dimíctics típics, amb la termoclina d’estiu ben definida entre 10 i 20 m. A l’estiu, els màxims de clorofil·la se situen per sota de la termoclina o més avall, i es diferencien, si més no fisiològicament, poblacions fitoplanctòniques epilimnètiques i hipolimnètiques. El litoral normalment és ocupat per esbaldregalls i no té cinyells de vegetació; les parets de la cubeta són molt inclinades i el sediment fi —això és biològicament important— se situa a fondàries superiors a la de la termoclina d’estiu. Per tant, els períodes de producció de l’estany són marcats per l’arribada de la barreja al nivell d’aquell sediment fi. Aquests estanys estan tot l’any ben oxigenats, tant a l’estiu com en el període d’hivern. Només en les parts més fondes hi ha un consum apreciable d’oxigen, però sense arribar probablement a la anòxia; per les seves característiques, són de producció molt baixa.

Els estanys pirinencs petits mostren sovint un desenvolupament important de la vegetació litoral, composta generalment de diverses espècies de càrexs, que formen un cinyell tot al seu voltant, i diverses fanerògames submergides. En aquests llacs la producció i la biomassa vegetal és més elevada. Aquest tipus d’estany sol ser més freqüent a alçades intermèdies i rep quantitats més grans de sediments i nutrients, que en condicionen força les característiques. Sovint estan voltats de pastures, de manera que la visita dels ramats incrementa notablement les entrades de nutrients i de matèria orgànica. La fotografia, de l’estany Pedó (Pallars) és un exemple extrem on, a causa de la poca fondària i de l’eutròfia, la vegetació de càrex típicament litoral ha ocupat la part central de l’estany.
Enric Ballesteros
Un segon tipus el representen aquells estanys encara dimíctics, però de mida i sobretot fondària més petita (15-20 m), que presenten un desenvolupament de la comunitat vegetal litoral i bentònica més important. Una quantitat notable de llum arriba sobre el fons, on es desenvolupa generalment una població important d’asprelles (Nitella), unes algues macroscòpiques. Les vores tenen un cinyell de càrexs —que sovint només envolta l’estany parcialment—, i diverses fanerògames aquàtiques hi viuen submergides. Els valors, més alts, de biomassa per unitat de volum fan que puguin presentar, durant les fases productives, augments notables de pH (se n’han mesurat valors superiors a 8,5), perquè les plantes consumeixen CO2 de l’aigua per a la fotosíntesi. Aquests estanys poden assolir la anòxia hivernal a les parts més fondes. Si la fondària dels estanys és encara més petita, poden convertir-se en polimíctics. El resultat és un comportament del fitoplàncton més homogeni en tota la columna, i el desenvolupament d’un poblament més adaptat a la radiació alta. Tots aquests llacs de menys volum es congelen i es desglacen més aviat que els grossos situats a una mateixa altitud i, per tant, l’estructura de la coberta de gel és diferent perquè presenta una proporció de gel dur més elevada.
Moltes vegades estanys similars als anteriors es troben en una vall, en lloc d’un circ, i llavors tenen algunes característiques diferents. Normalment són bastant més allargats, amb un procés deltaic a l’entrada d’aigua que els dóna una asimetria longitudinal. Si el rebliment de la cua de l’estany és de materials fins, pot formar-se una zona de mullera. Normalment són poc fons, són situats a menys altitud que els altres i drenen una àrea més gran. Tenen un temps de residència de l’aigua molt baix, i és improbable que arribin a la anòxia, perquè la circulació d’aigua es manté poc o molt durant pràcticament tot l’hivern. La fusió es produeix més aviat. A més del poblament, l’estany rep molts elements a través del rierol que el travessa; així és comú trobar en el seu plàncton una proporció més elevada de diatomees bentòniques. En algun cas, sota el gel s’ha trobat un gran nombre de zoòspores de Hydrurus, una crisofícia tal·losa que recobreix les pedres dels rius de muntanya.
A mesura que els estanys es van reblint de sediments, la producció neta anual supera cada cop més la respiració. El fred o fins i tot la congelació alenteixen la degradació i hi ha una acumulació de matèria orgànica. Els estanys evolucionen des de l’oligotròfia cap la distrófia, fins a convertir-se finalment en mulleres i en basses temporals. Aquests ambients terminals són els que presenten un conjunt d’espècies més diferenciat dels de la resta d’estanys.
Les comunitats límniques pirinenques

Esquema idealitzat d’un estany pirinenc (a dalt), amb una gran diversificació d’ambients i organismes, i principals organismes que formen part del plàncton (a baix). 1 pèl caní (Nardus stricta), 2 càrex inflat (Carex rostrata), 3 càrex fosc (Carex fusca), 4 esfagne (Sphagnum), 5 cotonera de fulla estreta (Eriophorum angustifolium), 6 isoet (Isoetes lacustris), 7 espargani de muntanya (Sparganium angustifolia), 8 Nitella, 9 Nostoc zetterstedtti, 10 estatoblast de Cristatella, 11 Spirogyra, 12 Cricotopus 13 truita (Salmo trutta fario), 14 Chironomus, 15 Tubifex tubifex, 16 sangonera, 17 esponges, 18 Hydra rubra, 19 Lymnaea, 20 Crucigenia, 21 Synura, 22 Uroglena, 23 Daphnia, 24 Kellicotia, 25 Cyclops, 26 Asplanchna, 27 Sphaerocystis, 28 Peridinium, 29 Melosira, 30 Mallomonas, 31 Chrysoikos, 32 Rhodomonas, 33 Pseudokephyrion, 34 Bitrichia, 35 Monosiga, 36 Cryptomonas, 37 Strombidium, 38 Ochromonas, 39 Dinobryon, 40 Polyarthra, 41 Eurycercus lamellatus, 42 Biapertura affinis, 43 Micrasterias, 44 Staurastrum, 45 Cosmarium, 46 Achnanthes, 47 Frustulia, 48 Ceratoneis arcus, 49 Stauroneis, 50 Eunotia, 51 Pinnularia.
Roman Montull i Maria Rieradevall, a partir de fonts diverses
El poblament dels estanys pirinencs és resultat d’una colonització recent, postglacial, de fa menys de 10 000 anys. Hi predominen les espècies de tipus cosmopolita o, si més no, de distribució àmplia a l’hemisferi nord i, en menor nombre, s’hi troben espècies de característiques bòreo-alpines. La diferenciació respecte d’altres indrets dels Països Catalans és determinada sobretot pels requeriments ecològics particulars. En general, els llacs presenten una diversitat específica baixa i només algunes espècies hi són quantitativament importants.
El fitoplàncton i la producció primària
En el fitoplàncton predominen les formes flagel·lades i petites: al voltant del 80% de les espècies presents fan menys de 20 µm. La diversitat d’espècies és generalment baixa i només algunes arriben a constituir poblacions importants al llarg de l’any. Les formes flagel·lades petites són principalment crisofícies d’organització molt senzilla (Chromulina, Ochromonas). Són també un element característic les crisofícies loricades (Dinobryon, Bitrichia, Chrysolykos), les formes més petites de les quals (Pseudokephyrion) poden ser l’element dominant, en nombre i en biomassa, en determinades èpoques de l’any. Aquests estanys de muntanya són ambients on és fàcil trobar crisofícies amb esquames silícies (Synura, Mallomonas). Les diatomees apareixen sobretot durant els períodes de barreja, en formes colonials, com Asterionella, Melosira i Tabellaria. Algunes espècies heliòfiles de Cyclotella poden mantenir-se a l’epilímnion durant el període estiuenc. Al plàncton de tots els estanys hi ha sempre un component variable d’espècies típicament bentòniques (Pinnularia, Navicula, Nitzschia, etc.).
Entre les formes no flagel·lades, tret del període d’aparició de les diatomees, dominen les clorofícies clorococals (Sphaerocystis, Crucigenia, Tetrastrum, Ankistrodesmus, Monoraphidium, Botryococcus). Les desmidiàcies apareixen esporàdicament i augmenten en aquells estanys més relacionats amb àrees de torberes. Les criptofícies, Rhodomonas sobretot i Cryptomonas, són un component habitual i, en alguna època de l’any, poden ser importants en biomassa. Els dinoflagel·lats, en canvi, rarament arriben a ser un element dominant. Les formes petites (Gymnodinium) apareixen durant tot l’any, però les formes més grosses amb teca (Peridinium) són preferentment d’estiu i Ceratium hi és rar. Els cianobacteris i els euglenòfits són molt infreqüents. Algunes espècies sideròfiles de Trachelomonas apareixen en aigües especialment riques en ferro.
La periodicitat anual del poblament fitoplanctònic no presenta una pauta clara general per a tots els estanys. Fins i tot en un mateix estany hi ha diferències entre un any i l’altre. En general, després de la fusió del gel, el creixement fitoplanctònic és de crisòfits, bé de crisofícies (Dinobryon) bé de diatomees. El poblament d’estiu és el més variat i variable d’un estany a l’altre: criptofícies, clorofícies i dinoflagel·lats, sobretot. A la tardor, en un altre màxim de productivitat, poden tornar a aparèixer diatomees, però sovint són les mateixes clorofícies d’estiu les que incrementen el poblament. Sota el gel la situació és poc coneguda, però sembla que el poblament de tardor, en condicions de poca llum i baixa temperatura persisteix força i va perdent biomassa lentament, mentre que la proporció d’espècies de flagel·lats incolors (Gymnodinium, Monas, Monosiga, Desmarella) augmenta. Si prop del fons s’arriba a condicions d’anòxia, llavors els canvis són més radicals i s’incrementa molt el nombre de bacteris.
D’acord amb l’oligotròfia dels estanys, la concentració de la clorofil·la no és mai gaire alta: d’1 a 3 µg/l a l’estiu i fins a valors de 4 a 7 µg/l durant la barreja. A l’estany Redó (Vall d’Aran), s’hi han trobat els màxims anuals de clorofil·la unes quantes setmanes després de glaçar-se i poc abans que el gruix de neu aturés l’entrada de llum. Molts dels llacs passen bona part de l’any amb valors inferiors a l’1 µg/l. Alguns estanys eutrofitzats poden arribar a més de 10 µg/l, especialment alguns de petites dimensions, freqüentats pel bestiar a l’estiu.
Encara queden per conèixer molts dels aspectes de producció dels estanys pirinencs. La producció primària neta del plàncton deu oscil·lar entre els 10 i 150 mg C/m2/dia. Hi ha fotoinhibició del fitoplàncton en els primers metres de la columna d’aigua, tot i que les poblacions de l’epilímnion són més eurifòtiques. L’òptim fotosintètic se situa a la fondària on arriba d’un 7 a un 12% de la il·luminació superficial. La fotoinhibició estimula la fracció de productes excretada, que arriba a ser d’igual magnitud o més alta que la producció retinguda com a biomassa. La renovació de la biomassa fitoplanctònica pot atènyer el 100% diari en els períodes més productius.
El zooplàncton
Les poblacions de ciliats prenen importància sota el gel. Juntament amb components comuns com Halteria, també s’han trobat formes més particulars com un Strombidium loricat o algun Dileptus, entre moltes altres formes.
Els rotífers constitueixen d’un 2% a un 10% de la biomassa zooplanctònica. Els més comuns són Asplanchna priodonta i Polyarthra vulgaris, espècies cosmopolites que a l’estiu es troben en molts estanys. Kellicottia longispina i Ascomorpha ovalis són espècies de caràcter bòreo-alpí, força esteses també, i la primera, de vegades, amb alts valors de biomassa. Menys freqüents són altres espècies de distribució nòrdica (Trichocerca longispina, Lecane ligona, Filinia cornuta). Keratella hiemalis, és una espècie estenoterma que rarament es troba a temperatures superiors a 6°C i que habitualment és hipolimnètica; com les tres anteriors, la seva presència a la península es limita als Pirineus. A la Cerdanya apareixen dos rotífers que no s’han trobat als Pirineus centrals, Ploesoma hudsoni i Conochilus unicornis, espècies de caràcter atlàntic que també viuen a l’W de la cadena cantàbrica.
En els crustacis planctònics dominen els elements cosmopolites, de poc interès biogeogràfic, probablement a causa de la colonització recent. Cyclops abyssorum apareix a la majoria d’estanys, igual que Daphnia longispina, encara que aquesta té pautes d’abundància més fluctuants, pròpies de la idiosincràcia dels cladòcers; ambdues espècies són d’expansió ràpida, s’han estès per molts dels embassaments de la península i presenten poblacions morfològicament bastant variables. Scapholeberis mucronata és un altre cladòcer força freqüent, que es distribueix per la regió holàrtica nord i presenta una adaptació morfològica de les valves que li permet explotar l’aliment de la pel·lícula superficial de l’aigua. Holopedium gibberum, un element clarament bòreo-alpí, és molt més infreqüent. Algunes espècies de cladòcers només s’han trobat, als Pirineus, als estanys de la Cerdanya i a Andorra (Daphnia obtusa, D. pulicaria, Polyphemus pediculus).
Els diaptòmids, en canvi, tenen un gran interès biogeogràfic. Als Pirineus falten les dues espècies més freqüents als Alps i a d’altres serralades europees. Hi apareixen dos diaptòmids grossos, Diaptomus cyaneus i D. castaneti, i dos de petits, Mixodiaptomus laciniatus i Eudiaptomus vulgaris. Les dues espècies grosses o les dues espècies petites no coexisteixen en un mateix estany, sinó que s’exclouen mútuament, mentre que en els estanys més grans i profunds pot arribar-se a trobar un diaptòmid gros i un de petit. Generalment les espècies petites ocupen els estanys d’altitud moderada, mentre que les més grosses viuen en els estanys de més altitud i també en aigües temporals. Les dues espècies de diaptòmids petits poden viure en un marge bastant ampli de condicions, i sembla que la seva distribució va lligada a la història de la seva dispersió i a una forta competència entre elles, amb avantatge per a l’espècie fundadora. Mixodiaptomus laciniatus és pròpia dels Pirineus occidentals; la localitat més oriental on fins ara s’ha trobat és l’estany de Toro, a la part W de la Vall d’Aran, encara que al vessant francès arriba més a l’E. És una espècie bòreo-alpina que s’estén per la franja atlàntica, molt abundant al NW de la península i a les muntanyes cantàbriques, i apareix també al Sistema Central, i a la Sierra Nevada i a l’Atles, que esdevenen els seus marges de distribució. Eudiaptomus vulgaris, que ocupa pràcticament tots els Pirineus catalans, és una espècie centreuropea que no s’ha estès ni cap al N ni cap a la Mediterrània; els Pirineus i alguns embassaments que s’hi relacionen en són el límit meridional: Diaptomus cyaneus, de distribució mediterrània occidental, és l’espècie més freqüent de diaptòmid al vessant S. Les formes d’Andorra i de la Cerdanya són diferents de les de la part central dels Pirineus. D. castaneti també apareix per tots els Pirineus catalans, però no entra cap als Pirineus aragonesos.
El cicle anual dels crustacis als estanys es coneix poc. En general, hom considera que tenen un desenvolupament lligat al règim tèrmic del llac i que passen l’hivern en estat alentit en forma d’adult o subadult (els ciclòpids) o en forma d’ous de resistència (Daphnia, i els diaptòmids). No obstant això, a ple hivern, tres mesos després d’haver-se format la coberta, s’han trobat densitats altes de Daphnia sota el gel i, durant tot un seguiment hivernal de l’estany Redó (Vall d’Aran), s’han trobat sempre alguns ciclòpids en estat adult o de copepodit. El període reproductiu té lloc durant la fusió o immediatament després. La biomassa màxima del zooplàncton es dóna al final de l’estiu, moment en què pot sobrepassar vint vegades la del fitoplàncton. Els adults, tant de Daphnia com de Cyclops, presenten moviments migratoris clars de la superfície al fons del llac, mentre que els nauplis d’aquests gèneres i els rotífers no en tenen. Als Pirineus no hi ha peixos planctòfags, de manera que Cyclops representa el nivell més alt de predació per al plàncton, tret que a l’estany no hi hagi larves del dípter Chaoborus, les quals cacen crustacis del plàncton i larves de quironòmids.
El bentos i els organismes de la zona litoral

El destí final de tots els llacs és quedar reblerts de sediments i de vegetació, procés el pas intermedi del qual és la formació de mulleres. En els bassiols i rierols de les mulleres, s’hi troba una comunitat biològica rica i diversa, força diferenciada de la dels estanys. La fotografia mostra l’estany dels Güells de Marimanya (Vall d’Aran), pràcticament tot ell convertit en una mullera.
Enric Ballesteros
Es podria fer una primera divisió dels estanys pirinencs entre aquells que tenen una vegetació macrofítica ben desenvolupada i aquells que tenen el litoral ocupat per cúmuls d’esbaldregalls.
En els estanys de més altitud i situats en conques amb poc sòl, la vegetació del litoral és constituïda exclusivament per diverses espècies de molses, que, a més, no són típicament lacustres. Sovint són formes que també es troben als torrents o als seus marges, i que colonitzen també les aigües estanyades (Blindia acuta, Anisothecium palustre, Cratoneuron commutatum varietat falcatum). Algunes, en l’ambient lenític, modifiquen la seva morfologia: allarguen les tiges, la disposició de les fulles es fa més laxa i els rizoides són més prims i febles. Poden ocupar des de la vora de la superfície fins a fondàries considerables, ja que se n’han trobat fins a 30 m (Drepanocladus exannulatus) i és possible que arribin a més profunditat. Durant l’hivern mantenen un aspecte verd, tot i que no els arriba la llum.
En els estanys de vegetació més rica poden distingir-se tot un seguit de components, que citarem a continuació, bé que, òbviament, no sempre es troben a tots els llacs. Començant per la vora de l’estany, la transició entre el prat alpí i les comunitats submergides la realitza un cinyell de ciperàcies i juncàcies (Carex nigra, C. echinata, C. lepidocarpa, Juncus filiformis, Eriophorum angustifolium, Scirpus caespitosus, Juncus alpinus, etc.) i algunes altres plantes característiques com la viola d’aigua (Viola palustris), la fetgera blanca (Parnassia palustris) i la pingüícula (Pinguicula vulgaris); tot i que aquesta comunitat només s’inunda temporalment, sempre manté una gran quantitat d’aigua, sovint afavorida per un substrat d’esfagnes i d’altres molses (Drepanocladus revolvens, Calliergonella cuspidata, etc.). Alguns estanys amb zones força reblides de sediment presenten una torbera d’esfagnes associada, en la qual es formen bassiols que mantenen una vida aquàtica independent. Cap a l’interior de l’estany, pot haver-hi un altre cinyell, en el qual domina, i molts cops com a únic component, Carex rostrata, sovint ja inundat, i de vegades hi apareix, barrejat, el trèvol d’aigua (Menyanthes trifoliata). Fins als dos primers metres de fondària, s’estén una comunitat ja típicament aquàtica, constituïda per isoets (Isoetes lacustris, I. echinospora); subulària (Subularia aquatica), una crucifera de distribució àrtico-pirinenca; ranuncles d’aigua (Ranunculus aquatilis), amb fulles de dos tipus, unes que suren i les altres que són submergides; i l’espargani de muntanya (Sparganium angustifolium), de llargues fulles acintades, que es troba molts cops superposat a aquestes plantes petites, però potser més freqüentment sol. Habitualment les clapes més importants d’espargani se situen prop de la zona d’entrada del rierol a l’estany; un conjunt dens d’aquesta planta pot modificar notablement l’ambient lumínic subaquàtic. La zona de fondària situada entre 1 i 4 m sovint és ocupada pels potamogètons (Potamogeton berchtoldii, P. alpinus), als quals es pot afegir el ranuncle Ranunculus trichophyllus, en la seva subspècie lutulentus, de fulles i tiges reptants, i exclusiva de l’alta muntanya alpina, i també Myriophyllum alterniflorum o Ranunculus aquatilis, entre d’altres. Les fanerògames rarament colonitzen les zones que es troben més avall dels 5 m. El fons normalment l’ocupa una alga cloròfita macroscòpica, Nitella, el recobriment i la biomassa de la qual pot arribar a ser molt important, més de cent vegades més gran que la del fitoplàncton; en canvi, la seva producció és inferior, de 30-80 mgC/m2dia; l’òptim el té cap als 10 o 12 m, encara que en alguns estanys viu a pocs metres de la superfície.
En els estanys pirinencs sorprèn el contrast entre la poca presència de cianobacteris en el plàncton i, en canvi, la seva abundància sobre les roques del litoral. L’espècie més característica en els primers metres és Nostoc zetterstedtii, que forma uns petits coixinets gelatinosos blavosos, tot i que n’hi poden haver moltes més (Schizothrix, Pleurocapsa, Hydrococcus, etc.); en alguns estanys també hi ha Stigonema, un cianobacteri d’aspecte muscinal, de 2-3 cm, fosc, que representa l’organització morfològica més complexa dintre del grup, amb ramificacions veritables. Les diatomees són l’altre element dominant del microfitobentos: algunes espècies colonitzen tots els substrats en totes aquelles fondàries on arriba la llum (Achnanthes, algunes Cymbella i Nitzschia), i hi ha una heterogeneïtat elevada en qualsevol direcció; la clorofil·la sobre les pedres és de 10-20 mg/m2. En els estanys amb mulleres i en les cubetes situades entre esfagnes hi ha un augment de les desmidiàcies (Cosmarium, Euastrum, Staurastrum, Micrasterias, etc.) i d’algunes diatomees (Eunotia, Frustulia, Pinnularia).
El microbentos epífit no difereix radicalment del que es troba sobre substrats inorgànics, encara que la proporció d’algunes espècies típicament epífites pot augmentar (Cocconeis). La distribució sobre la planta no és homogènia: algunes algues se situen preferentment en determinades zones d’una planta, i fins i tot una cara de la fulla pot ser colonitzada i l’altra no.
La fauna d’invertebrats que viu relacionada amb les parets d’un llac pirinenc arriba a l’entorn del centenar d’espècies tot i que molt poques hi tenen un paper quantitatiu important. La distribució batimètrica depèn del substrat, de les condicions tèrmiques i de l’oxigenació. Un 50% són larves d’insectes que no desenvolupen el seu cicle vital complet dins del llac.
Normalment hi ha més diversitat de crustacis bentònics que no pas planctònics, bé que han estat molt poc estudiats. Són comuns alguns cladòcers cosmopolites (Biapertura affinis, Chydorus sphaericus) o bòreo-alpins (Eurycercus lamellatus); Alona rustica hi viu associada a la vida de les torberes. Els harpacticoides dels Pirineus són poc coneguts; Canthocamptus staphylinus s’encista formant una membrana hialina entorn del cos doblegat dorsalment, i ho fa tant a l’estiu com a l’hivern, diverses vegades. A partir dels 5 m de fondària, a les parts amb sediment fi, hi viuen ostràcodes (Cypria, Candona). El creixement i la reproducció dels crustacis s’esdevenen durant el període sense gel. En el cas dels cladòcers, després del desgel hi ha l’eclosió dels ous durables. La màxima reproducció partenogenètica és a l’agost i al setembre, i al final del període hi ha una fase de reproducció sexual. Els ciclòpids tenen un desenvolupament en fases larvals: la primera, que és anomenada naupli i és molt diferent de l’adult, passa després de cinc mudes a una segona fase més semblant a l’adult, la de copepodit, que amb unes quantes mudes més donarà l’adult. En el cas dels ciclòpids, en el moment del desgel, els adults ja són en fase de reproducció, i les femelles, ovígeres o no, són molt més abundants que els mascles. De mitjan agost a mitjan setembre, la població té majoria de copepodits, mentre que al final de l’estiu el poblament és de mascles, de femelles no ovígeres i dels darrers estadis de copepodit.
Entre els altres invertebrats que completen tot el cicle vital a l’estany hi ha: mol·luscs (Lymnaea, Pisidium), sovint amb la closca molt feble a causa del baix contingut de calci de les aigües; esponges (Spongilla lacustris); briozous (Plumatella, Cristatella); celenteris (Hydra rubra); coleòpters (Agabus solieri, Donacia, que respira l’aire contingut en les tiges de les fanerògames submergides); planàries i oligoquets. Aquests últims tenen importància quantitativa, especialment a la part més fonda, i s’hi troben, entre d’altres, Tubifex tubifex, Limnodrilus hoffmeisteri, Lumbriculus variegatus (que viu en els fons entapissats de Nitella) i Peloscolex ferox (a les parts poc fondes); les sangoneres abunden en els estanys freqüentats per les vaques.
Les larves d’insecte són el component quantitatiu i qualitatiu més important del zoobentos, tot i que els tricòpters, els plecòpters i els efemeròpters hi tenen poca representació. En canvi, els quironòmids poden tenir prop d’una quarantena d’espècies en un sol llac, i pot distingir-s’hi un grup d’espècies de litoral i un grup de fons, a més d’espècies que es troben per tot arreu; generalment el fons l’ocupen espècies estenotermes fredes; però en aquells estanys en què al fons hi ha anòxia són substituïdes per espècies eurioiques, com ara Chironomus plumosus, i també C. commutatus, element típicament bòreo-alpí, que suporten les baixes tensions d’oxigen; es dóna la paradoxa que en llacs oligotròfics, com els pirinencs, apareixen espècies típiques d’ambients eutròfics. Els quironòmids presenten una sincronització molt marcada: en 10 dies tots els imagos d’una mateixa espècie emergeixen, els del litoral una mica abans que els del fons. Els quironòmids que tenen la larva del quart estadi petita (de menys de 8 mm) realitzen el cicle de creixement i d’emergència en un any; però les espècies més grans triguen dos anys; la larva del megalòpter Sialis lutaria n’està tres.
Els peixos i altres vertebrats
Als Pirineus hi ha dues truites, la comuna, autòctona (Salmo trutta fario), i la introduïda (Salmo gairdneri), força menys abundant als estanys. Ambdues volen aigües netes, ben oxigenades i fredes, tot i que l’espècie introduïda és més laxa en els requeriments, ja que ocupa els ambients més extrems, aquells que a l’estiu s’escalfen per sobre dels 15°C, o els de més altitud. L’època del zel i de la fresa de la truita comuna té lloc durant els mesos d’octubre i novembre i es realitza en zones de poca fondària, d’aigües quietes i amb fons d’arena o grava, on les truites excaven un petit pou. El nombre d’ous depèn del pes de la femella (uns 400 ous/kg). El desenvolupament embrionari es retarda durant tot l’hivern. Les truites de llac passen més temps estàtiques que les de riu, per la qual cosa el volum del cos augmenta en relació amb el del cap. La coloració varia força d’un estany a l’altre. La truita de rierol (Salvelinus fontinalis) ha estat introduïda en alguns estanys, encara que amb poc èxit; aquesta espècie és originària de l’Amèrica del Nord, mentre que el seu congènere europeu (S. alpinus) no es troba als Pirineus. El principal aliment de les truites dels estanys és constituït per les larves d’insecte, capturades en el moment d’emergència, i pels insectes aeris atrapats a la pel·lícula superficial. D’altra banda, la depredació tant d’exemplars de la mateixa espècie com d’ous és un hàbit comú en les truites. Un altre peix freqüent és el barb roig (Phoxinus phoxinus), que no és realment un barb i que només té rojos el ventre i els llavis del mascle durant l’época de zel. És de mida petita i es troba preferentment cap a la part oriental dels Pirineus, on conviu amb la truita en alguns estanys.
A part els peixos, hi ha pocs vertebrats en els estanys. Alguns amfibis (Rana temporaria, Bufo bufo) hi passen la vida larval, que dura uns 20 dies, i el tritó pirinenc (Euproctus asper), que és endèmic dels Pirineus, viu en els llacs o a prop, encara que és d’activitat més aviat nocturna.
Els llacs càrstics
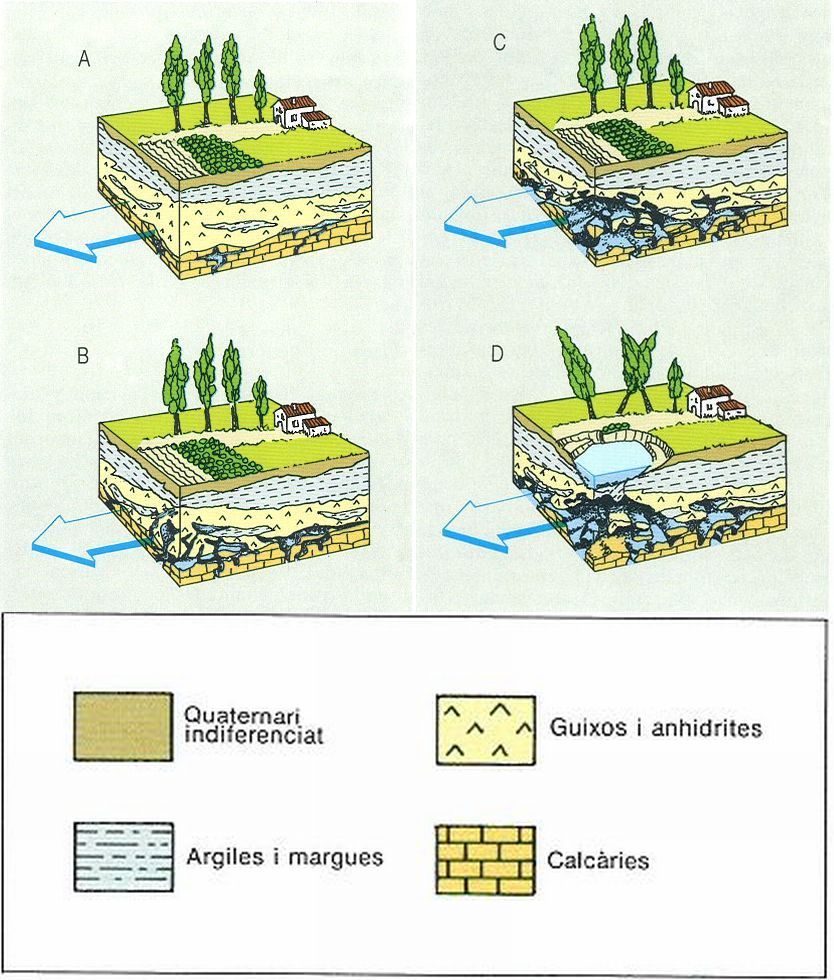
Esquema sintètic de la formació d’un estanyol càrstic. A Inici del procés de carstificació per flux d’aigua a pressió en l’aqüífer d’un terreny calcari. B La carstificació creixent dissol de baix a dalt els materials evaporítics. C Se sobrepassa l’equilibrí mecànic de les cavitats, però la compressió hidràulica les manté. D La descompressió temporal del sistema provoca un col·lapse gravitacional; una nova injecció hídrica generarà un estanyol.
Jesús Garcia Gil, original de D. Brusi, L. Palli i M. Sanz
Els llacs càrstics dels Països Catalans es troben situats al S dels massissos càrstics paral·lels a la serralada pirinenca i pràcticament alineats des de l’W amb els llacs d’Estanya, a Benavarri (Baix Ribagorça), fins a l’estany de Banyoles (Gironès), passant pels de Montcortès de Pallars (Pallars Sobirà) i de Basturs, a Isona (Pallars Jussà). Això és així perquè els massissos càrstics esmentats actuen com una zona de recàrrega de l’aqüífer que després alimentarà les masses d’aigua. També als flancs del Maestrat, al Sistema Ibèric, prop de Tírig (Alt Maestrat), hi ha petites dolines amb aigua o estanyols càrstics. Testimonis del que poden haver estat dolines, ara completament reblides, queden al Garraf.
Origen, morfometria i hidrologia
Els mapes batimètrics dels llacs càrstics en reflecteixen l’origen geològic. La dissolució de roques calcàries, sovint associades a guixos (encara més solubles), per l’aigua que percola, origina, per enfonsament posterior del terreny, cubetes semisfèriques o troncocòniques, que reben el nom de dolines. Per això, els llacs càrstics solen tenir la forma arrodonida d’una dolina o bé d’un camp de dolines («polje»), més o menys complex, inundat. A poc a poc, i per efecte de la dissolució provocada per l’aigua, les cavitats van augmentant de diàmetre fins que el sostre es precipita i la cubeta s’engrandeix. Una característica general de tots els llacs càrstics és, precisament, la seva alimentació mitjançant deus subterrànies. Normalment l’alimentació subterrània és comparativament més important que la superficial, al contrari del que passa a la resta de tipus de llacs.
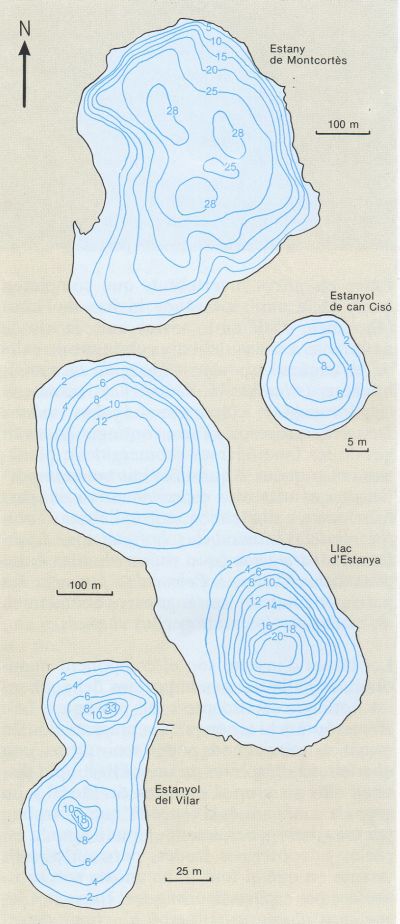
Mapa batimètric de diferents estanys i estanyols càrstics catalans, on les isolínies indiquen profunditats cada vegada més grans. La forma arrodonida és la més corrent, i en alguns casos hi ha una doble dolina, com en el llac d’Estanya i l’estanyol del Vilar, o formes més complexes, com la de l’estany de Banyoles.
Maria Rieradevall, a partir de fonts diverses
En el cas de l’estany de Banyoles, com a resultat del procés de dilució, el mapa del fons o mapa batimètric és força complex. És constituït per cinc cubetes o dolines principals, el centre de les quals correspon als punts de fondària màxima i alhora als punts d’entrada d’aigua, que es manifesten en la presència de sediment en suspensió. Gràcies a l’observació del mapa batimètric d’altres cubetes ja enfonsades (les anomenades cubeta 1 i cubeta 2), es pot deduir que per sota del forat de l’anomenada cubeta 3 hi ha una enorme cavitat de forma irregular, en lloc, com en altre temps s’havia pensat, d’una xemeneia de parets rectes fins als 132 m. Aquestes cavitats subterrànies són plenes d’aigua i de sediment en suspensió en l’aigua, que són els que afloren pels forats de les cubetes del llac i per molts estanyols del voltant. Al modelat actual de l’estany, hi contribuïren, hom creu, els efectes de dissolució i d’enrunament que ajuden a explicar certs fenòmens com el de l’octubre de 1904, que va fer recular les aigües dels recs de sortida de l’estany uns 50 m, amb la qual cosa es parà momentàniament el seu flux.

Els estanys càrstics són molt característics de les serres d’altures mitjanes revestides de calcàries mesozoiques, en les quals la dissolució del terreny (guixos i calcàries) per les aigües subterrànies propicia la formació de dolines. Són sistemes oligotròfics (el fòsfor precipita fàcilment amb el calci), amb poques entrades d’aigua superficial, però amb aportacions d’aigua subterrània de més o menys importància, tant pel fons com lateralment (per exemple, a l’estany de Montcortès). L’estany gran de Basturs (Isona), que veiem a la fotografia, rep aigua pel fons durant tot l’any.
Narcís Prat i Maria Rieradevall
En el cas de l’estany de Banyoles, els afluents no són rius sinó rieres habitualment poc cabaloses; però, en canvi, de l’estany surten cinc recs de cabal variable que històricament no s’han assecat mai. Els cabals de les rieres d’entrada i dels recs de sortida són molt diferents i variables; la riera de Morgat és la de més cabal que hi entra i el rec Major, el més gran que en surt. El balanç, doncs, és favorable a la sortida. Són remarcables les grans diferències entre les entrades i sortides totals, que oscil·len entre els 38 000 m3/dia en períodes secs (al final de l’estiu) i els 67 000 m3/dia en períodes plujosos (a la primavera). Per tant, aquestes diferències representen els valors dels cabals que corresponen a l’alimentació subterrània de l’estany de Banyoles. A aquestes dades caldria afegir els valors de l’evaporació i la diferència (positiva o negativa) entre les entrades i sortides dels aqüífers de flux difús.
L’origen de les aigües de l’estany de Banyoles ha estat estudiat des de principi de segle. Sembla demostrat que les aigües de Banyoles es dipositen en forma de pluja a la zona de l’Alta Garrotxa, a les conques hidrogràfiques de la riera d’Oix i dels rius Borró i Llierca i penetren a la terra a través de les fissures dels materials calcaris de la zona, formant un enorme aqüífer confinat o captiu. Les seves aigües circulen per dintre de la terra, sota el riu Fluvià i, en arribar a la zona de l’estany, emergeixen a la superfície; és a dir, passen de circular de forma quasi horitzontal a pujar verticalment. Aquest canvi de direcció té una explicació geològica: les calcàries fissurades per les quals circula l’aqüífer queden trencades sota l’estany per la falla d’Albanyà, fet que provoca que, a partir d’aquesta, l’aigua topi amb materials impermeables (gresos o marés i margues) que actuen com un dic. Així, la pressió de l’aigua de pluja que va entrant pel costat de recàrrega de l’aqüífer a la zona de Sadernes empeny, per vasos comunicants, l’aigua verticalment sota la zona lacustre de Banyoles. Aquest és l’origen de la surgència del Clot d’Espolla. Igual que a la superfície de la terra hi ha bulïidors, estanyols, deus, fonts, etc., en el fons de l’estany hi ha també enormes bullidors que es corresponen directament amb els punts dels quals es pot afirmar actualment que entra aigua a pressió pel fons. Aquesta entrada d’aigua provoca corrents ascensionals que transporten material del fons de l’estany: l’anomenat sediment en suspensió. Així, doncs, per aquesta observació indirecta coneixem alguns dels punts d’entrada d’aigua a l’estany. Això no exclou que n’hi hagi d’altres, ja sigui amb sediment en suspensió o sense, ara per ara desconeguts. En el decurs de l’ascensió a la zona lacustre de Banyoles, l’aigua va dissolent els materials solubles que travessa, principalment guix (CaSO4), en volums que s’estimen entre 50 i 130 m3/km2 any. La dissolució de carbonats es calcula entre 7000 i 20 000 m3/any.
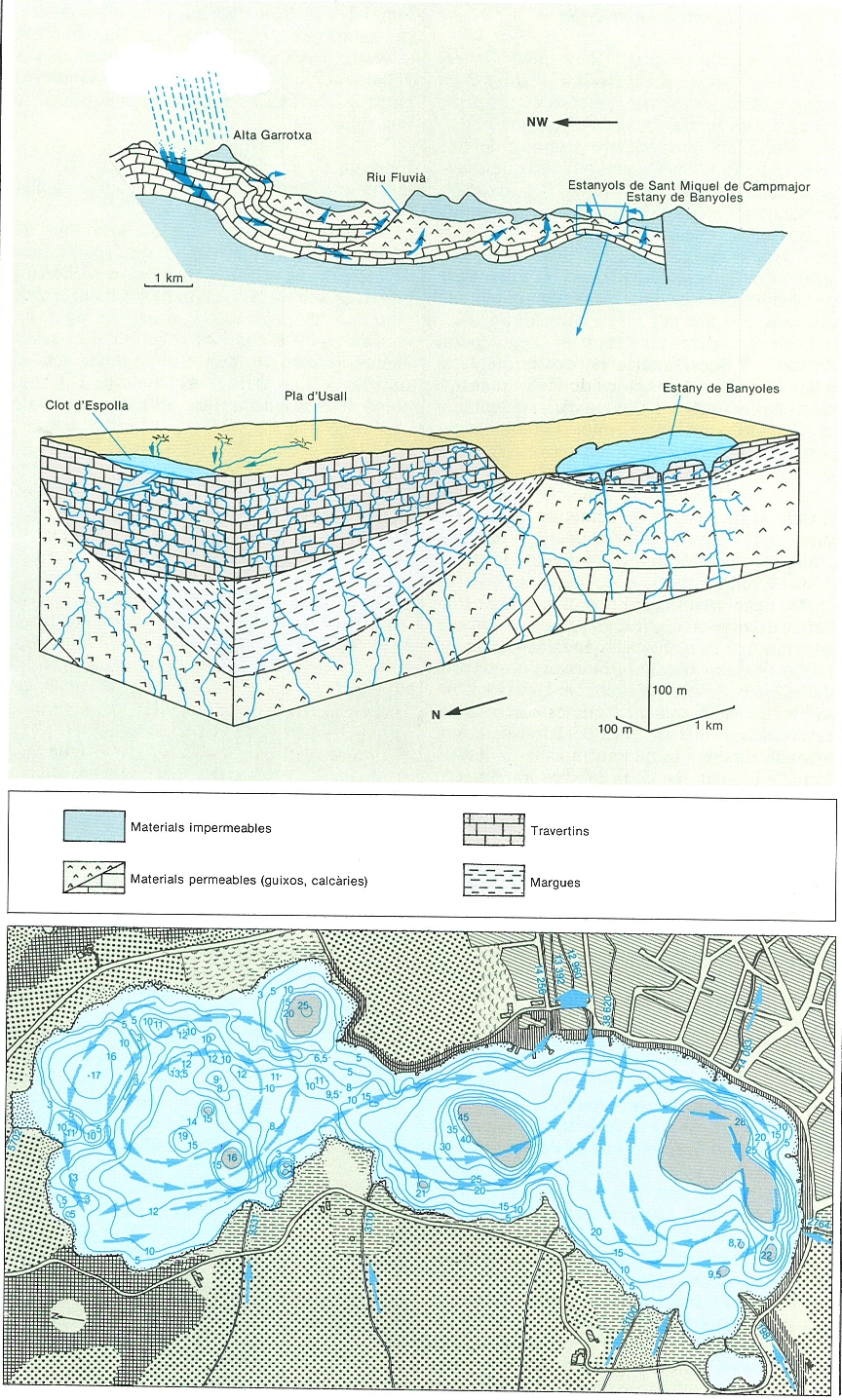
Origen i funcionament del sistema càrstic de Banyoles (a dalt). Les aigües de pluja que cauen a les serres de l’alta Garrotxa s’infiltren per les calcàries eocèniques i circulen a través d’elles fins que troben la falla que va enfonsar l’Empordà. Allà les aigües topen amb materials impermeables que actuen com un dic i provoquen la seva ascensió a la zona de Banyoles per vasos comunicants. Finalment, les aigües surten del llac cap al riu Terri i la conca del Ter. Quan plou molt i Taqüífer és ple, l’aigua pot ascendir a través d’escletxes fins a cotes més altes que les de l’estany; llavors s’omple l’estany temporani del Clot d’Espolla, situat a l’altiplà d’Usall, a uns 40 m damunt de l’estany de Banyoles. Una vegada és ple, del Clot d’Espolla surt un riu i les aigües retornen al Fluvià i no al Ter, formant un salt d’uns 60 m. A baix, mapa batimètric de l’estany de Banyoles, amb indicació de les vies d’entrada, sortida i circulació superficial de l’aigua, i de les àrees on hi ha fang en suspensió per l’existència d’una font d’aigua profunda. El volum d’aigua que entra i surt de l’estany pels afluents i efluents s’indica en m3/dia.
Maria Rieradevall, a partir de fonts diverses; mapa: Carto-Tec, original de R. Moreno i Amich i A. Garcia Bentou
Els estanyols que formen part del mateix sistema lacustre que l’estany de Banyoles, de fet, són surgències similars a les del fons de l’estany, però que han quedat abandonades per les aigües de l’estany major o bé s’han format de bell nou a partir d’ell. Els estanyols de la vall de Sant Miquel de Campmajor i l’estany d’Espolla, amb les diferents surgències, tenen el mateix origen càrstic i comparteixen amb l’estany l’alimentació hídrica subterrània. L’estanyol de la Cendra (d’uns 40 m de diàmetre) rep aquest nom per la presència de sediments del fons, de color cendrós, en suspensió (xalió o margues blaves disgregades), des del fons (20 m) fins a la superfície. Els estanyols Nou i de can Cisó (o estanyol Vermell) formen, juntament amb un important bullidor, el complex de la riera Castellana, una zona d’aiguamolls molt interessant des del punt de vista paisatgístic i ecològic; el Nou es va formar sobtadament la nit del 17 de novembre de 1978 en un camp de fruiters i pollancres, i actualment té una fondària màxima de 7 m i 40 m de diàmetre; a poca distància, hi ha el de can Cisó o Vermell situat també a la dreta de la riera Castellana. El seu nom és en relació amb el color que pren l’aigua, de vegades vermell porpra en època hivernal, que és causat per la presència de denses poblacions de bacteris fototròfics vermells en suspensió. L’estanyol del Vilar, el més gran i conegut de tots (de 170 m de longitud i 80 m d’amplada) és situat al S de l’estany de Banyoles i s’hi comunica per un canal artificial. És format per dos enfonsaments sensiblement circulars (dolines) i fàcilment visibles. Al fons també hi ha dos forats amb sediment en suspensió i amb fondàries de 33 i 18 m, per on entra l’aigua. Va sofrir una forta contaminació durant la dècada dels setenta per filtracions de pous morts o per abocaments directes de clavegueres, de la qual avui encara no s’ha recuperat: té l’aigua molt verda, amb una gran densitat d’algues en suspensió.
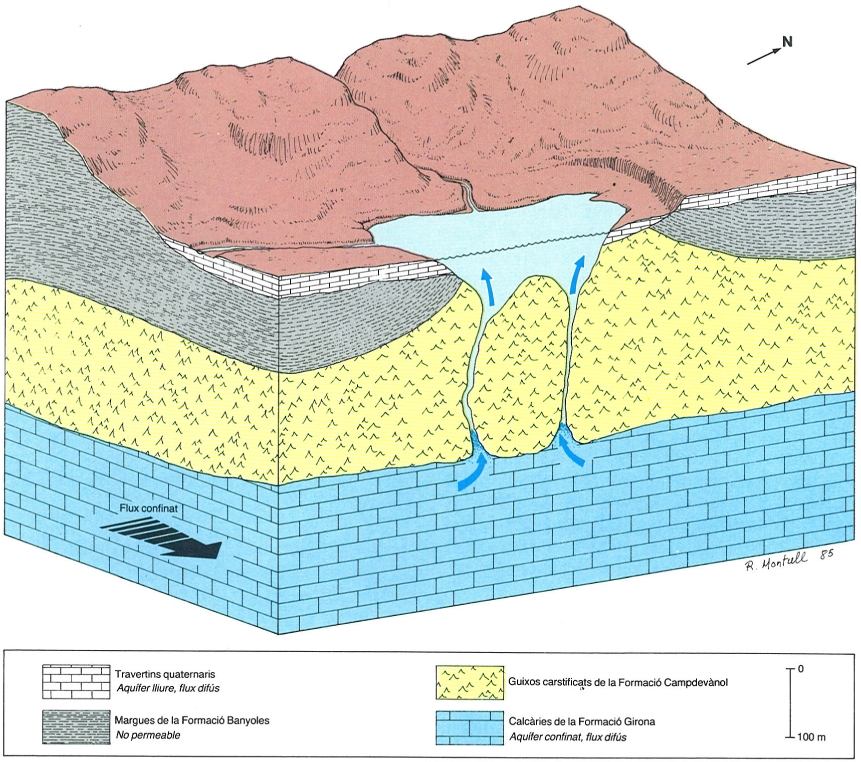
Aqüífers calcaris de flux difús en el sector de Banyoles: superposició d’aqüífers carbonatats amb flux difús, confinats i lliures en calcàries eocenes i travertins quaternaris.
Romà Montull, original de l’autor

Lluís Solé i Sabarís (1908-1985) fou el primer geòleg català que creà escola. Conreà tots els camps de la geologia, encara que s’especialitzà en els camps de la geomorfologia i la història del Quaternari. En els seus treballs inclou sovint blocs diagrames que il·lustren la relació entre el relleu i l’estructura geològica, com el que reproduïm, que mostra la geologia de la regió de Banyoles.
Jordi Vidal
L’estany de Montcortès, situat a la conca del Flamicell, i a 1065 m d’altitud, és gairebé circular, amb una fondària màxima d’uns 30 m; té una sola cubeta i és principalment d’alimentació subterrània. Tant aquest com els de Basturs funcionen com a limnocrens. Mentre el primer és feblement mesotròfic i meromíctic, els estanys de Basturs, que poden considerar-se un de sol amb dues cubetes separades, són d’aigües molt netes i d’una taxa de renovació molt alta. L’origen i la hidrologia dels estanys de Basturs (Pallars Jussà), el de Montcortès (Pallars Sobirà) i els tres d’Estanya, anomenats Gran de Baix, Gran de Dalt i Petit (Baixa Ribagorça), no es coneixen tan bé com els del sistema de Banyoles, que hem descrit àmpliament.
Paràmetres morfomètrics principals d’alguns estanys càrstics dels Països Catalans.
| LLACS CÀRSTICS | ÀREA SUPERFICIAL (m)2 |
PROFUNDITAT (m) |
VOLUM (m3) |
||
| MÀXIMA | MITJANA | ||||
| Estany de Banyoles | |||||
| Totalitat de l’estany | 1,1 × 106 | 40,0 | 15,75 | 1,7 × 107 | |
| Cubeta 3 | 7,5 × 104 | 30,0 | 10,92 | 8,2 × 105 | |
| Estanyols de Banyoles | |||||
| Vilar | 1,2 × 104 | 9,0 | 4,50 | 5,4 × 104 | |
| Cisó | 4,6 × 102 | 7,5 | 4,78 | 2,2 × 103 | |
| Nou | 8,4 × 102 | 7,0 | 2,26 | 1,9 × 103 | |
| Coromines | 2,9 × 103 | 5,0 | 2,90 | 8,4 × 103 | |
| Negre | 5,1 × 102 | 5,2 | 2,94 | 1,5 × 103 | |
| Estany de Montcortès | 9,3 × 104 | 30,0 | 20,43 | 1,9 × 106 | |
| Llac Gran d’Estanya | 2,1 × 103 | 20,0 | 6,60 | 1,4 × 103 | |
| Dades compilades pels autors |
Dinàmica i funcionalitat: mineralització, cicle anual i producció

Cicle de diferents paràmetres a la cubeta 3 de l’estany de Banyoles, on hi ha una meromixi característica. El dibuix mostra les variacions, segons la fondària i al llarg de dos anys (1977-78). de la temperatura, la conductivitat, l’oxigen i els sulfurs. La capa d’aigua es presenta dividida sempre en tres parts: la superior o mixolímnion va des de la superfície fins a uns 18 m de fondària (varía lleugerament segons els períodes de mescla i estratificació d’aquestes aigües); la intermèdia o monimolímnion, dels 18 als 25 m; i la més fonda, feta d’aigua amb sediment, a partir dels 25 m de fondària. Durant els estius, a més, la capa superior queda estratificada en un epilímnion superior i un hipolimnion, separats per una termoclina clarament marcada. Aquesta estratificació fa que l’aigua de les dues capes del fons mai no es barregi amb la de dalt ni tingui contacte amb l’atmosfera. Per això, el monimolímnion es caracteritza per la seva anòxia. L’aigua amb sediment en suspensió té una conductivitat més elevada, una temperatura constant tot l’any (de manera que a l’hivern dóna lloc a una inversió tèrmica) i un alt contingut de sulfurs resultat, fonamentalment, de l’activitat dels bacteris reductors de sulfats, que són abundants a l’aigua. La difusió de l’àcid sulfhidric cap als 14 o 15 m, on arriba una mica de llum, permet la presència de bacteris fotosintetitzadors del sofre, que formen una de les comunitats més interessants dels estanys i els estanyols de l’àrea de Banyoles.
Carto-Tec, amb dades de C Abellà
Per les característiques químiques generals de la majoria dels llacs càrstics dels Països Catalans hom pot fer la separació en dos grans grups respecte de la concentració de sulfats: el format pels llacs de Montcortès, de Basturs, de la Coromina i Negre, de baixa concentració de sulfats, i la resta, d’elevada concentració de sulfats, fins a màxims de més d’1 g/l al fons d’algunes cubetes de l’estany de Banyoles. La conductivitat reflecteix directament, en general, l’elevada concentració d’anions d’aquestes aigües. El pH quasi sempre és neutre o lleugerament alcalí (màxim de 8,5), encara que al fons pot ser lleugerament àcid a causa de la freqüent presència de l’àcid feble sulfhídric. Per tot això, l’aigua de l’estany de Banyoles, i dels llacs càrstics en general, pot ser considerada de baixa qualitat; pel contingut de sals, especialment sulfats, la concentració dels quals oscil·la entre 0,5 i 1,2 g/l, es troba en els límits de l’aigua potable. L’aigua és, per tant, sulfatada i bicarbonatada.
Tanmateix, en el transcurs de les estacions se succeeixen grans canvis en sentit vertical i horitzontal, que de fet separen masses d’aigua ben diferenciades. En els llacs càrstics, la columna d’aigua no solament pot estar estratificada (epilímnion, termoclina i hipolímnion) per l’efecte combinat de l’escalfament i la força del vent, com era el cas dels llacs dels Pirineus, sinó també per un gradient del contingut de sals dissoltes a l’aigua. Per tant, poden coexistir dues estratificacions, que delimiten dues masses d’aigua, una on es produeix una estratificació tèrmica en forma de cicle anual (mixolímnion), i una altra que resta gairebé permanentment capturada en el fons del llac (monimolímnion) per causa de la seva densitat més gran i del contingut més elevat de sals. La capa de separació entre la part superior d’aigua que es mescla (mixolímnion) i la que no es mescla (monimolímnion), rep el nom de quimioclina (gradient químic). A l’estany de Banyoles, a la cubeta 3 (però també a l’estany de Montcortès), l’aigua del fons hi és més densa (d=1,00274 g/cm3) que la superficial (d–l,00102 g/cm3). Aquesta estratificació sol ser permanent al llarg de l’any, fenomen conegut com a meromixi. Això afavoreix l’acumulació de substàncies químiques reduïdes procedents de l’activitat microbiològica al sediment, com CO2, NH+4, H2S, per processos de fermentació, putrefacció i reducció de sulfats, especialment en algunes cubetes i en aquells llacs amb una concentració més elevada de sulfats a l’aigua.
El monimolímnion i la quimioclina destaquen fàcilment si observem la variació de la temperatura, la conductivitat i la concentració d’oxigen i d’àcid sulfhídric al llarg de l’any. Així, la temperatura ens dóna clarament la idea del cicle anual, amb la formació de la termoclina, i alhora ens indica l’existència d’una massa d’aigua superior i una altra massa d’aigua inferior, amb diferències tèrmiques, en el cas que hi hagi quimioclina. D’altra banda, la distribució de l’oxigen al mixolímnion de la cubeta 3 de Banyoles segueix fidelment el procés d’estratificació i mescla, alhora que revela la presència del monimolímnion permanentment mancat d’oxigen. Els continguts màxims d’oxigen es troben freqüentment a la zona situada entre els 5 i els 10 m de fondària, indiquen el lloc de més densitat de fitoplàncton; els valors màxims solen detectar-se al final de l’hivern i al començament de la primavera. El procés d’estratificació pot provocar l’esgotament de l’oxigen a l’hipolímnion a la darreria de l’estiu (amb valors per sota els 4 mg/l a partir dels 9 m, a l’estiu de 1977, per exemple), a la qual cosa s’afegeix la anòxia que ja existia al monimolímnion del fons del llac. Com que el fons del llac sempre és anòxic i a l’aigua hi ha molts sulfats, aquests passen a la forma reduïda, el sulfhídric, que sempre hi té valors molt elevats. Per això, de vegades l’esgotament d’oxigen a l’hipolímnion és acompanyat de la difusió d’àcid sulfhídric des del monimolímnion, com va passar a la tardor del 1978, en què aquest compost reduït es començava a detectar a partir dels 12 m de fondària.
L’àcid sulfhídric és el compost característic del monimolímnion de la cubeta 3 de Banyoles, com ho era també a l’estany de Montcortès l’any 1976. Ateses les seves característiques d’un cert aïllament físic de la circulació general de l’estany, aquesta cubeta és meromíctica gairebé de forma permanent, a excepció d’algun episodi de mescla puntual (al final del 1978), seguida d’una nova i ràpida estratificació. Per això, els valors del sulfhídric són força constants al llarg del temps amb màxims relacionats amb l’augment de temperatura i, per tant, de l’activitat reductora de sulfats dels sediments durs del fons de la cubeta (no en el sediment aeròbic en suspensió) durant l’estiu i al principi de la tardor. En aquest moment, la presència de sulfhídric i de llum a la quimioclina permet el desenvolupament de poblacions denses de bacteris fototròfics.
Paràmetres químics principals de les aigües d’alguns estanys càrstics dels Països Catalans.| LLACS CÀRSTICS | pH | COND. (µS) |
ALC (meq/l) |
SO2-4 | PO3-4 | NO-3 | NO-2 | NH+4 | s2- | |
| (µg àt/l) | ||||||||||
| Estany de Banyoles | ||||||||||
| Cubeta 3 | 6,8-8,3 | 1200-2400 | 3,0-6,0 | 4,6-16,6 | 0-3,5 | 0,5-20,0 | 0,1-4 | 0-400 | 1,4 | |
| Cubeta 2 | 7,3-7,9 | 1200-1330 | 2,5-4,5 | 5,1-8,7 | 0,1-1,0 | 1,0-16,0 | — | — | 0 | |
| Estanyols de Banyoles | ||||||||||
| Vilar | 7,2-8,2 | 1100-2500 | 2,8-8,8 | 4,8-13,2 | 0-13,0 | 1,2-5,6 | 0-0,6 | — | 8,8 | |
| Cisó | 6,8-8,0 | 1900-2100 | 5,0-7,6 | 4,6-10,4 | 0-5,5 | 0-2,5 | 0-2,0 | 0-250 | 7,3 | |
| Nou | 7,0-8,5 | 1900-2400 | 5,5-13,7 | 3,0-13,3 | 0-80,0 | 0-0,8 | 0-0,4 | — | 1,4 | |
| Coromina | 7,8-9,0 | 800 | 4.1-5,4 | 0,3-0,7 | 0-23,1 | 0-0,6 | 0-0,6 | — | 2,3 | |
| Negre | 8,0-8,5 | 780 | 4,9-5,7 | 0,3-0,6 | 0-15,3 | 0,1-13,4 | 0-20,7 | — | 0,2 | |
| Estany de Montcortès | 6,5-8,5 | 500 | 2,5-4,0 | 0,7-1,9 | 0-0,1 | 0-5,0 | 0-0,1 | — | 0,5 | |
| Llac d’Estanya | 7,2-8,4 | 1950 | 2,0-3,5 | 6,6-8,4 | 0-1,0 | 0-8,5 | 0-0,9 | 1,1-90,5 | 1,1 | |
| Dades compilades pels autors |
Les comunitats
El fitoplàncton i la producció primària
Dos factors importants controlen el fitoplàncton dels llacs càrstics: la peculiar circulació de l’aigua, gairebé sense entrades superficials (però, en canvi, amb sortides) i l’elevat contingut de calci, que fa precipitar el fosfat. Això determina que a les aigües gairebé no hi hagi nutrients (principalment nitrogen i fòsfor), especialment a l’estiu, període d’estratificació tèrmica de les aigües. A la zona més il·luminada, el creixement de les algues és limitat, doncs, pels nutrients, i el plàncton presenta característiques oligotròfiques; és a dir, aigües amb molt pocs nutrients, clares i netes. Per altra banda, la influència de les surgències subterrànies i l’estratificació tèrmica que pot determinar una zona rica en nutrients prop de la termoclina pot estimular un creixement hipolimnètic de la biomassa planctònica. I en el cas que, a més, es produeixi una segona estratificació al fons, i que es diferenciï un monimolímnion anòxic que faciliti l’alliberació de nutrients del sediment, es pot establir just sobre seu, si hi arriba la llum, una comunitat planctònica formada per diverses poblacions que, tot distribuint-se en capes en el perfil vertical, presenten densitats extraordinàries.
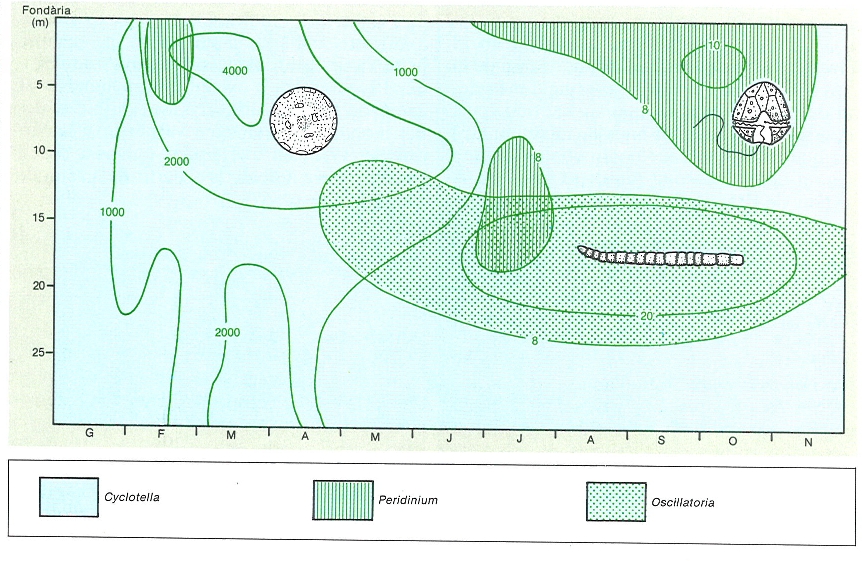
Distribució anual i vertical de la concentració màxima dels grups del fitoplàncton en l’estany de Montcortès, on la meromixi separa una capa anòxica a partir dels 15-20 m. Hi és característica l’abundància d’Oscillatoria, just als límits superiors de la capa anòxica; a la primavera, amb les aigües barrejades, hi domina especialment Cyclotella, i a l’estiu i la tardor, les dinoflagel·lades (Peridinium), que poden nedar i desplaçar-se en l’aigua encalmada i amb pocs nutrients, bé que al principi de l’estiu també es troben prop de la interfase entre les capes òxica i anòxica.
Maria Rieradevall, a partir de fonts diverses
Així, doncs, la part de dalt dels llacs es caracteritza per un plàncton d’algues petites o nanoplàncton, de baixa concentració i dominat fonamentalment per diatomees cèntriques del gènere Cyclotella. C. compta és la més important als llacs d’Estanya i de Montcortès, i també a Banyoles, bé que en aquest llac són fins i tot més abundants C. melosiroides i C. glomerata, presents gairebé tot l’any, amb màxims a l’hivern i a la primavera. El fitoplàncton de cada un d’aquests llacs és compost d’unes 80 espècies diferents, i algunes de les més freqüents coincideixen en un llac i altre i presenten cicles anuals d’abundància força regulars. De les dades existents sobre Banyoles, Montcortès i Estanya, hom pot resumir-ne la successió anual del fitoplàncton. A l’hivern, l’aigua no és estratificada, però té tendència a diferenciar-se lleugerament cap al fons. El plàncton té una densitat molt baixa, a la ratlla de 1000 cèl./ml (no més d’1 µg/l de clorofil·la a) i és constituït principalment per cèl·lules de Cyclotella acompanyades, en menys proporció, per Rhodomonas minuta i algunes crisofícies. Després de la barreja vertical de primavera es dóna un màxim de fitoplàncton, que pot arribar a ser d’uns 5000 a 10 000 cèl./ml (2 a 7 µg/l de clorofil·la a), constituït fonamentalment per Cyclotella; la diversitat és llavors força baixa (2,5 bits). Segons els anys i el llac, aquest màxim té com a espècies secundàries Cryptomonas, a més de Rhodomonas i crisofícies. A l’estiu, la densitat de població es manté alta: les Cyclotella hi són en abundància, bé que ara se situen en capes més profundes i són les principals responsables del màxim de producció al fons; ara bé, les espècies acompanyants ja són unes altres. Es poden fer importants les clorofícies que de vegades presenten pics de creixement molt alts a la superfície segons els anys, com és el cas de Planctonema lauterborni al llac d’Estanya. També són abundants els dinoflagel·lats, sobretot de Gymnodinium i de Peridinium, a tots els llacs, i a Banyoles, a més a més, Ceratium hirundinella presenta un creixement important. Al final de l’estiu la diversitat augmenta fins als valors màxims (3,5 bits) i la densitat tendeix a disminuir. La mescla vertical de la tardor provoca un nou màxim de Cyclotella que va acompanyat també de Rhodomonas, com a espècie subordinada; més avançada l’estació, es poden donar també màxims de clorofícies (Dictyosphaerium pulchellum, Chlorella vulgaris). El nombre de cèl./ml torna a ser molt alt i la diversitat es manté bastant alta, però va disminuint fins als mínims d’hivern (1,5 bits). Las comunitats fitoplanctòniques d’aquests llacs corresponen a les que hom ha descrit per a llacs d’Europa i de l’Amèrica del Nord oligotròfics d’aigües mineralitzades riques de calci i de pH alcalí.
Les comunitats estratificades de fondària presenten màxims importants, sobretot a l’estiu. L’establiment de la termoclina facilita l’estratificació més profunda, i el màxim primaveral de producció proporciona matèria orgànica que s’acumula a la quimioclina o, si més no, a una determinada fondària, on es pot originar algun gradient de densitat. El més general és que s’estableixi una microzonació molt marcada de diversos organismes fotosintetitzadors. Es freqüent, doncs, trobar a l’estiu les següents poblacions d’algues en el perfil vertical: diverses espècies de clorofícies, més aviat a la superfície; diverses espècies de Cyclotella, just sota la termoclina, que pot ser la població més important; diverses espècies de Peridinium, per sota d’aquella; algunes espècies de Cryptomonas, just sobre la inferfase (especialment C. phaseolus), on n’hi pot haver en grans quantitats; més avall, un gran creixement d’Oscillatoria rubescens, que en el cas dels estanys de Montcortès i d’Estanya pot donar màxims de clorofil·la de 20 µg/l; finalment, les comunitats bacterianes.
Per causa del dèficit de nutrients esmentat, la producció primària en aquests llacs és baixa a les capes del mixolímnion, on és molt inferior al límit superior que es dóna per als llacs oligotròfics. Per exemple, hom ha estimat valors de 85 g C/m2·any en un estudi fet al llarg de l’any 1970, en una cubeta no meromíctica de l’estany de Banyoles; aquest valor és similar al de la producció de la Mediterrània prop de les nostres costes, és a dir, una producció molt baixa. Recordem que la producció de la mar sempre és molt baixa comparada amb la dels sistemes d’aigua dolça, i la Mediterrània és, a més, una mar poc productiva. La producció prop del monimolímnion deguda a la proliferació de les espècies d’Oscillatoria o dels bacteris fotosintetitzadors encara és més petita. Sembla que es fan poblacions molt denses, però molt poc actives: l’assimilació de carboni hi és sorprenentment baixa, molt per sota del que faria pensar la concentració de clorofil·la. La causa és que aquestes poblacions són limitades per la llum, cosa que s’ha demostrat en diverses ocasions fent mesures de producció d’aquestes poblacions traslladades a medis d’intensitats lluminoses més grans. És a dir, quan una mostra de fondària (posada en una botella transparent) s’incuba prop de la superfície, la producció resulta unes quantes desenes de vegades més gran que quan s’incuba a la mateixa fondària on es trobava.
D’altra banda, els estanyols són molt més productius. A l’estanyol del Vilar, la producció primària en 1970-71 era de 277 g C/m2·any, tres vegades més gran que la de l’estany de Banyoles. Malgrat que l’aigua del Vilar és la mateixa que alimenta l’estany, amb el qual estava en comunicació, les poblacions fitoplanctòniques són molt diferents, perquè l’estanyol va ser aïllat de l’estany gran per a fer-hi un viver de peix i perquè hi ha pous morts prop seu. Les algues dominants aquell any ja no van ser les del gènere Cyclotella sinó de Chlorella i altres clorofícies, i la concentració de clorofil·la va arribar a 40 µg/l durant els màxims de densitat fitoplanctònica. Els estanyols, per causa de les seves dimensions reduïdes, són molt més sensibles a la contaminació.
El zooplàncton
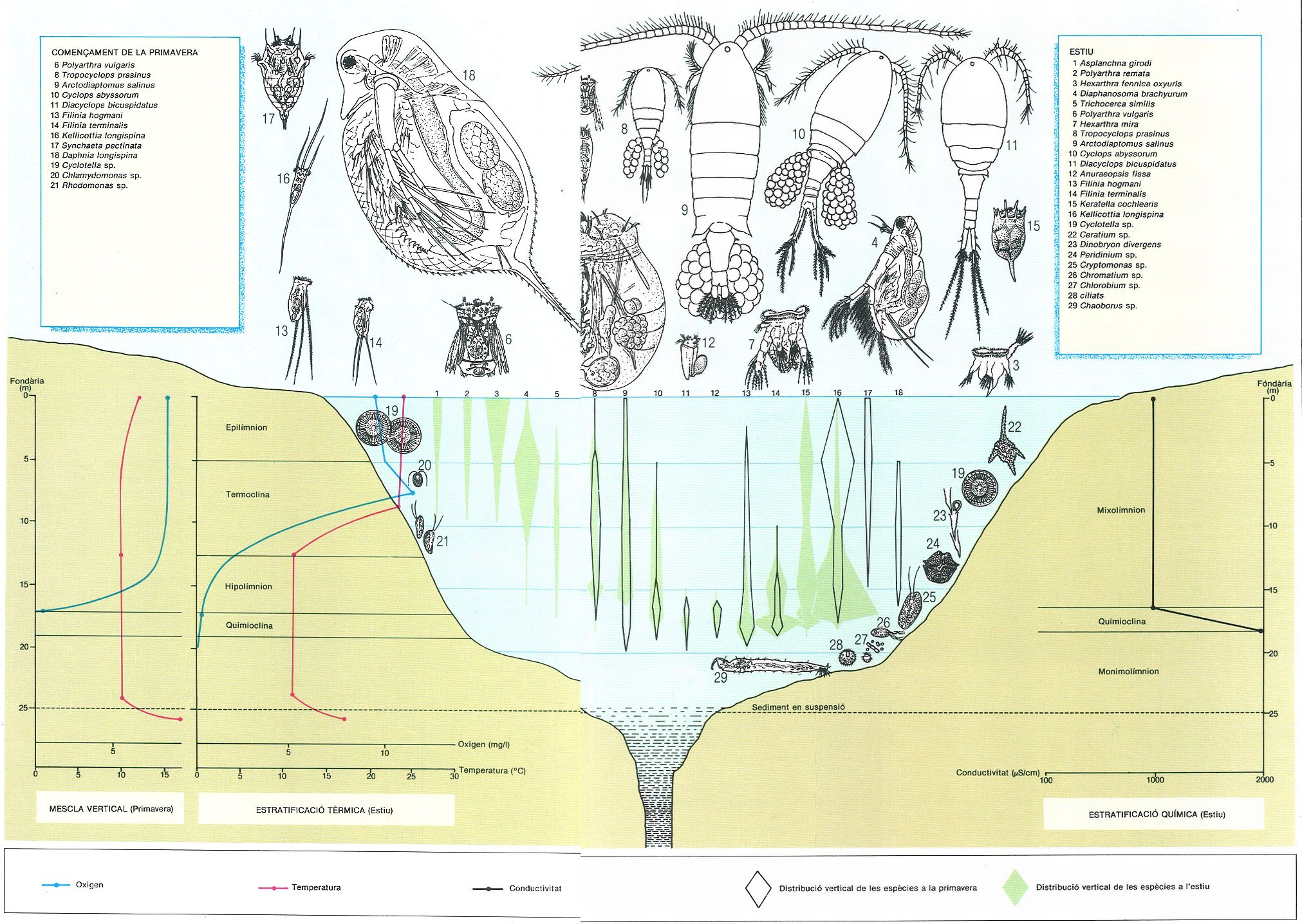
Distribució en el perfil vertical de les principals espècies del zooplàncton a l’estany de Banyoles, en un dia del principi de la primavera (mescla vertical) i un dia d’estiu (estratificació). Al període de mescla l’oxigen i la temperatura tenen valors homogenis en tot el perfil, fins que es produeix un augment de la temperatura en el sediment en suspensió o fins que s’arriba a l’extinció de l’oxigen a l’altura de la quimioclina, que és marcada també per un augment de la conductivitat. Les espècies del plàncton es distribueixen també força homogèniament tret d’algunes que se situen sempre al fons, prop de la quimioclina (s’indiquen també les espècies més importants del fitoplàncton en aquesta època). Al període d’estratificació s’observa una distribució diferencial marcada de les espècies planctòniques. Es forma una termoclina i la concentració d’oxigen presenta un màxim just a la part alta d’aquesta, que correspon al màxim de producció primària. Les espècies del zooplàncton es distribueixen en tres zones: unes presenten màxims marcats de fondària, prop de la quimoclina; d’altres ho fan a les capes superficials; i d’altres, prop de la termoclina. També s’han indicat les espècies més representatives del fitoplàncton, en les fondàries corresponents als seus màxims.
Román Montull
L’estructura de la comunitat zooplanctònica del mixolímnion dels llacs càrstics correspon a la que es troba normalment en llacs de característiques trònques i morfomètriques semblants. Es compon de crustacis i de rotífers, en una quantitat semblant uns i altres pel que fa al nombre d’individus, però amb una clara dominància de crustacis en biomassa (més del 75% del total del zooplàncton), ja que aquests són de dimensions molt més grans que els rotífers.
Entre els crustacis, les espècies majoritàries són un diaptòmid dominant filtrador, un ciclòpid petit filtrador relativament abundant (Tropocyclops prasinus, generalment), un ciclòpid depredador (espècies del gènere Cyclops) i dos cladòcers filtradors, un d’hivern i de primavera (Daphnia) i un altre d’estiu, força abundant (Diaphanosoma). Els crustacis, particularment els copèpodes, són bastant estables en el temps, quasi sempre són presents en el plàncton i són capaços de fer migracions verticals importants. Normalment només poden coexistir espècies de grups o famílies molt diferents. Dels llacs càrstics del nostre país, només dos, Banyoles i Basturs, reuneixen les condicions necessàries per als diaptòmids; mentre que en els altres llacs hi ha tan sols ciclòpids. Els diaptòmids són de distribució restringida i de gran interès biogeogràfic. El diaptòmid de Banyoles és Arctodiaptomus salinus, espècie d’aigües mineralitzades i de distribució circummediterrània, però que s’estén fins a l’Àsia central. Als estanys de Basturs, hi viu Eudiaptomus padamus, espècie europea que només s’ha trobat en alguns llacs dels Prealps.
Els rotífers varien extraordinàriament en el temps, i en els llacs en coexisteixen una vintena d’espècies, moltes de congenèriques, que s’exclouen en l’espai (perfil vertical de la columna d’aigua) i se succeeixen en el temps. Solen ocupar fondàries més delimitades que els crustacis i no fan grans desplaçaments.
La variació estacional del zooplàncton constitueix un exemple de les tendències de la successió. A la primavera, la mescla vertical fertilitza les aigües, es desenvolupen els productors primaris i s’afavoreix el creixement d’unes quantes espècies (la diversitat és baixa), que ocupen pràcticament tot el perfil vertical: espècies petites de rotífers filtradors de partícules fines i nauplis de copèpodes. A mesura que avança l’estiu les poblacions es diversifiquen i s’estratifiquen en el perfil vertical seguint l’estratificació físico-química. Els organismes s’acumulen a les discontinuïtats, és a dir, a la termoclina i a l’oxiclina. En el cas particular dels rotífers, es dóna una clara diferenciació de les comunitats en les diferents capes i una sorprenent estratificació d’espècies congenèriques: a l’epilímnion, Hexarthra fennica, Polyarthra remata i Asplanchna girodi; a la termoclina, Hexarthra mira, Polyarthra dolichoptera-vulgaris, Keratella quadrata i Trichocerca similis; a l’hipolímnion, Polyarthra longiremis, Keratella cochlearis i Kellicottia longispina. Totes les espècies de Polyarthra són d’hàbits més o menys raptors i s’alimenten de fitoplàncton més aviat gran. La resta de les espècies citades són filtradores, excepte Asplanchna girodi, que és depredadora. Si el llac o la cubeta té monimolímnion, llavors a l’oxiclina hi ha un desenvolupament de denses poblacions d’espècies del gènere Filinia (una o dues espècies) i Anuraeopsis, i en menys proporció, de Polyarthra. Els rotífers de l’oxiclina Filinia i Anuraeopsis són fonamentalment detritívors o filtradors de partícules fines (bacteris), mentre que Polyarthra captura Cryptomonas. A la zona de l’oxiclina hi ha també un important desenvolupament de les poblacions de diferents ciliats; alguns tenen algues en simbiosi; però, a més, s’alimenten també, uns de bacteris i els altres de Cryptomonas. Els crustacis ocupen una part més àmplia del perfil. El ciclòpid filtrador petit i els cladòcers tendeixen a ser epilimnètics, i els diaptòmids i els ciclòpids depredadors, mesolimnètics o hipolimnètics. Les larves naupli dels copèpodes creixen normalment a capes d’aigua més profundes que els altres estadis i també es concentren a l’oxiclina.
El bentos, els organismes del litoral i els peixos
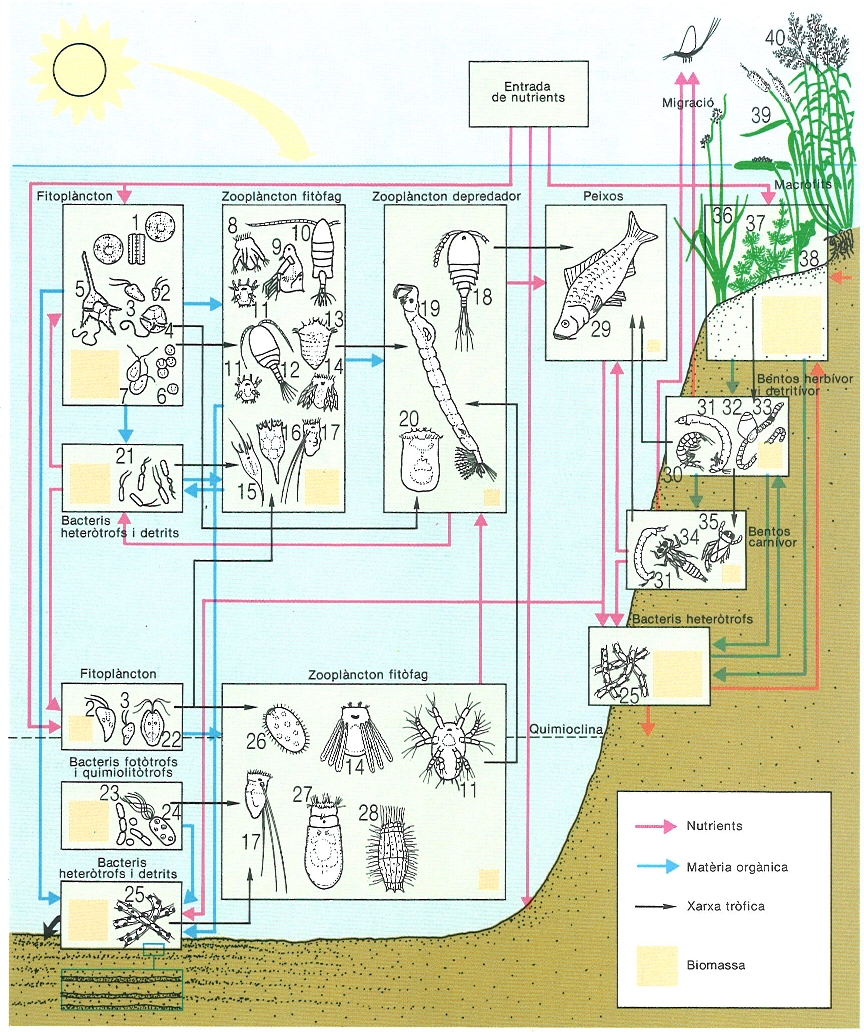
Flux tròfic de l’estany de Banyoles. Els productors primaris (sobretot diatomees petites) són filtrats pels crustacis planctònics; rotífers i ciliats filtren principalment partícules més fines, minúscules algues flagel·lades i detrits, i fan grans creixements prop de la quimioclina, on també mengen bacteris fotosintetitzadors; uns altres s’alimenten d’algues grans (Cryptomonas) i els depredadors capturen dinoflagel·lats i animalets diversos. Al litoral els productors són els macròfits, que mantenen directament la comunitat de macroinvertebrats i indirectament la de peixos. En el dibuix, s’han representat: 1 Cyclotella, 2 Rhodomonas, 3 Cryptomonas, 4 Peridinium, 5 Ceratium, 6 Chlorella, 7 Chlamydomonas, 8 Hexarthra, 9 Diaphanosoma, 10 Arctodiaptomus salinus, 11 naupli, 12 Cyclops abyssorum, 13 Synchaeta, 14 Polyartha, 15 Kellicottia, 16 Keratella, 17 Filinia, 18 Cyclops, 19 Chaoborus, 20 Asplanchna, 21 Pseudomonas, 22 Tetraselmis, 23 Chlorobium, 24 Chromatium, 25 Leptothrix, 26 Prorodon, 27 Anuraeopsis, 28 Coleps, 29 carpa (Ciprinus carpio), 30 gammàrid (Echinogammarus), 31 quironòmid, 32 gasteròpode, 33 oligoquet, 34 odonat, 35 hemípter coríxid, 36 jonca (Schoenoplectus), 37 Myriophyllum, 38 nimfea (Nymphaea), 39 boga (Typha), 40 canyís (Phragmites). Als llacs meromíctics els nutrients i la matèria orgànica que entren a la quimioclina ja no recirculen sinó que queden atrapats al monimolímnion, d’on més tard van incorporant-se al sediment, on van formant bandes de colors; en el litoral sí que recirculen i poden ser novament utilitzats a les capes superficials.
Maria Rieradevall, original dels autors
En els llacs càrstics es pot observar la zonació de vegetació litoral típica dels estanys europeus, amb una gran riquesa d’espècies, que es conserva particularment bé a l’estany de Banyoles. Aquesta vegetació consta normalment d’una faixa de canyissar amb bogues (Phragmites, Typha), mansegues (Cladium, Cyperus) i jonques (Scirpus), seguida d’una de prats molls amb grans càrexs (Carex), on es poden trobar plantes força interessants; a continuació, cap enfora del llac, en llocs no tan humits, s’hi fan les jonqueres (Juncus); la banda següent ja és el bosc de ribera, del qual no resta gran cosa. La vegetació pròpiament aquàtica consta de gespes submergides formades per les fanerògames Myriophyllum i Potamogeton a tota la vora i als llocs poc profunds. En alguns punts, aquestes gespes són constituïdes per caràcies, bé que aquestes algues, molt sensibles a la contaminació, han disminuït molt en pocs anys, especialment a l’estany de Banyoles (Chara, Nitellopsis). A l’estany de Montcortès, on aquesta zona litoral és pobra en nutrients, creix Utricularia, fanerògama carnívora que captura petits animals gràcies a uns utricles o ampolletes especials i n’obté els nutrients necessaris.
A la zona de macròfíts la fauna invertebrada associada és molt rica. A més d’amèbids, ciliats, rotífers, copèpodes i cladòcers, s’hi troben animals més conspicus, com ara cargols i cloïsses d’aigua dolça, planàries, esponges i crustacis superiors. A Banyoles són abundants el decàpode Atyaephyra desmarestii, que era capturat per fer-lo servir d’esquer de pesca, i l’amfípode Gammarus pungens. Ambdós són crustacis de distribució circummediterrània, com Arctodiaptomus salinus.
La fauna bentònica pròpiament dita és formada per quironòmids, crustacis, nematodes i oligoquets. A la zona sublitoral (5 m de fondària), la fauna és més diversa i abundant (fins a 25 000 individus/m2), mentre que al fons de les cubetes sense monimolímnion (20 m) la fauna queda reduïda a unes quantes espècies i en densitats menors (2500 individus/m2). Entre els quironòmids, les larves dels quals viuen enterrades als sediments, destaquen Procladius, Cladopelma, Microchironomus i Chironomus com a gèneres característics. Entre els crustacis cal destacar la presència de Gammarus (Echinogammarus) pungens, que pot arribar a viure a 20 m de fondària a les èpoques en què l’oxigen és abundant al fons. A l’interior, més profund, de les cubetes o dels llacs amb monimolímnion tan sols hi ha la larva del dípter Chaoborus flavicans, capaç de desplaçar-se àmpliament en sentit vertical i d’adaptar-se a tolerar ambients sense oxigen; sovint s’hi presenta en densitats extraordinàries.
No hi ha peixos característics d’aquests llacs, i els més abundants van ser-hi introduïts per l’home. A l’estany de Banyoles, la carpa (Cyprinus carpió), el gardí (Scardinius erytrophthalmus), la perca americana (Micropterus salmoides) i el llop (Esox lucius) són les espècies principals. Aquestes, entre altres, han estat introduïdes des de principi de segle, quan Darder va institucionalitzar la «Festa del peix». En canvi, les espècies autòctones de Banyoles, com la tenca (Tinca tinca), el barb (Barbus meridionalis) i la bagra (Leuciscus cephalus), pràcticament han desaparegut, i d’altres que havien estat citades també com a habitants del llac, com el sorelló (Gasterosteus aculeatus), la babosa (Blennius fluviatilis) i l’anguila (Anguilla anguilla), no en queda sinó el testimoni. A l’estany de Montcortès, la truita (Salmo trutta) és el peix més abundant, també per les repoblacions efectuades.
Les comunitats bacterianes

Els llacs d’Estanya (Baixa Ribagorça) constitueixen un sistema càrstic força interessant bé que no tan extens com el de Banyoles. El formen dos llacs principals (l’un format per dues dolines, i l’altre, per una), d’alimentació subterrània i mancats d’afluents de sortida funcionals. Com a l’estany de Banyoles, hi ha comunitats bacterianes a una certa fondària que aprofiten l’elevada concentració de sulfats de l’aigua i la anòxia estival. Les aigües tenen la blavor típica dels ambients càrstics, causada per l’alta concentració de carbonat càlcic i pels sediments que hi pot haver en suspensió.
Miquel Alonso
Les poblacions microbianes no han de ser considerades elements accessoris de la comunitat, sinó uns dels seus components essencials, tant per la seva aportació a la biomassa total com per la seva activitat. D’una banda, la biomassa microbiana equival al 10-50% de la biomassa del fitoplàncton. De l’altra, els microorganismes canalitzen directament o indirectament el 50% de la producció del fitoplàncton, la qual cosa els converteix en un element clau per al funcionament de l’ecosistema. Gràcies a l’extraordinària diversitat del seu metabolisme, els diferents grups de bacteris realitzen nombroses funcions que permeten el reciclatge dels elements químics, especialment del fòsfor, el nitrogen i el sofre.

Estany de can Cisó (Banyoles) tenyit de vermell per les grans poblacions de bacteris porpres que s’hi formen en l’època de mescla vertical de les aigües, quan hi ha àcid sufhídric a tota la columna. El desenvolupament de grans poblacions de bacteris fotosintetitzadors és característic dels nostres sistemes càrstics. Són microorganismes capaços d’emprar l’àcid sulfhídric produït en els sediments anòxics com a donador d’electrons i de fixar carboni per mitjà d’uns pigments particulars.
Ricard Guerrero
Finalment, hom ha pogut comprovar que les relacions entre els components de la comunitat microbiana d’un llac són molt més complexes del que fins ara hom pensava. Per exemple, hi ha protists fotòtrofs que mengen bacteris, bacteris que ataquen i devoren altres bacteris, i bacteris que realitzen producció primària en condicions molt diverses. A més, hom pot trobar una gran varietat d’associacions simbiòtiques, paràsites i comensals entre diferents grups de microorganismes.
Entre els microorganismes que es troben habitualment en els ambients lèntics cal destacar els bacteris heteròtrofs dels gèneres Achromobacter, Flavobacterium, Brevibacterium, Micrococcus, Bacillus, Pseudomonas, Nocardia, Streptomyces, Micromonospora, Cytophaga, Spirillum i Vibrio, que són implicats en el reciclatge de la matèria en aquests ambients. També és freqüent trobar-hi bacteris autòtrofs pertanyents a diferents grups taxonòmics.
Als llacs càrstics, uns dels grups de bacteris més interessants són els que es relacionen amb el cicle del sofre. La presència de l’oxiclina i els gradients d’oxidoreducció que es formen donen origen a una gran diversitat morfològica i funcional. Els bacteris fotòtrofs del sofre que es troben en aquestes masses d’aigua pertanyen principalment a dues famílies: les cromatiàcies (bacteris vermells del sofre) i les clorobiàcies (bacteris verds del sofre). Aquests microorganismes, amb un característic metabolisme fotòtrof i anaeròbic, utilitzen l’àcid sulfhídric en presència de llum, i el converteixen primer en sofre i després en sulfat. Els representants més freqüents de les cromatiàcies són Chromatium minus, Amoebobacter, Thiocystis violacea i Thiopedia rosea. Tots tenen en comú l’acumulació intracel·lular de sofre i la presència de bacterioclorofil·la a. El principal carotenoide d’aquestes espècies és l’okenona, excepte en Thiocystis violacea, que presenta rodopinal. Les clorobiàcies, molt més petites que les cromatiàcies, són immòbils i no acumulen sofre intracel·lular. Els estudis realitzats indiquen que Chromatium minus és l’espècie més àmpliament distribuïda i que domina al llarg de l’estiu en tots els llacs càrstics excepte en la Massona, i els estanys de la Coromina, Negre i Montcortès; Amoebobacter es troba a l’estany de can Cisó coexistint amb Chromatium minus, tot i que és durant la tardor i l’hivern que arriba a la seva màxima concentració i esdevé dominant; Thiopedia rosea i Thiocystis violacea s’han trobat tan sols a l’estany Negre, cap a la meitat de l’estiu; Chlorobium limicola és abundant durant l’estiu als estanys de la Coromina, Negre, de can Cisó, Montcortès i Estanya; C. phaeovibrioides (espècie marina) constitueix la població dominant a les llacunes costaneres de Cullera i Massona, mentre que C. phaeobacteroides es troba durant l’hivern en concentracions elevades a l’estany de can Cisó, a la cubeta 3 de l’estany de Banyoles, i als del Vilar i Montcortès.
El model de l’estanyol de can Sisó
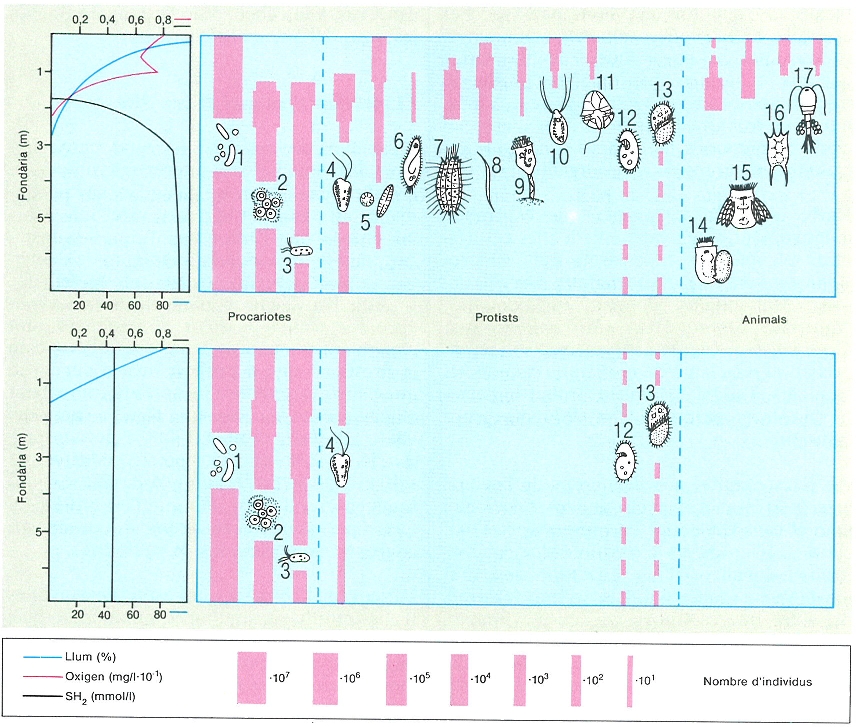
Comunitats característiques de l’estany de can Cisó i abundància i distribució dels organismes. A l’estiu (a dalt), amb l’estratificació tèrmica, la presència de la termoclina permet que l’oxigen quedi a la capa superior de l’aigua i que hi hagi una producció primària basada en diferents tipus d’algues, que permet una gran diversificació de la microbiota: hi ha epècies de les capes superiors, altres de la intermèdia i altres especialitzades a viure en les aigües anòxiques i carregades de sulfurs del fons. A l’hivern (a baix), quan l’aigua es barreja i tota la columna es carrega de sulfurs, només poden mantenir-s’hi algunes espècies, i especialment bacteris fototròfics, que poden arribar a tenyir l’aigua de color vermell si arriben a la superfície. S’ha representat en el dibuix: 1, bacteris heteròtrofs. 2 Amoebacter, 3 Chromatium, 4 Cryptomonas phaseolus, 5 diatomees, 6 Paramecium, 7 Coleps hirtus, 8 Hyaloraphidium, 9 Vorticella, 10 Carteria, 11 Peridinium, 12 Plagiopyla, 13 Metopus, 14 Anuraeopsis fissa, 15 Polyarthra, 16 Keratella, 17 Cyclops.
Maria Rieradevall, original de C. Pedrós-Alió
L’estanyol de can Cisó ha estat particularment estudiat pel seu interès microbiològic. És situat uns 300 m a l’W de l’estany de Banyoles, al costat de la riera Castellana, i forma una cubeta d’enfonsament quasi semisfèrica, de 8 m de fondària i uns 25 m de diàmetre; la seva superfície és tan sols de 487 m2. L’aigua d’aquesta superfície experimenta canvis espectaculars de color. A l’estanyol, tant el pendent pronunciat de la seva cubeta, com la protecció que li proporciona la vegetació que l’envolta, eviten la barreja i la consegüent oxigenació de la columna d’aigua a causa del vent. Tot això, combinat amb l’elevada producció de sulfhídric per bacteris reductors de sulfat del tipus de Desulfovibrio, que són presents en el sediment, fan que l’estanyol sigui quasi permanentment anòxic i que mostri concentracions elevades de sulfhídric.
Durant unes setmanes, a l’estiu, l’estanyol es troba estratificat, amb un epilímnion aeròbic i un hipolímnion anaeròbic. En aquestes condicions, hom hi pot trobar rotífers, especialment Anuraeopsis fissa, que es concentra al metalímnion amb una abundància que arriba fins a 104 individus/ml, mentre que és pràcticament absent a la resta de l’estanyol. En algunes èpoques de l’any hom hi pot trobar també, bé que en baixes concentracions, el copèpode Thermocyclops, i, ocasionalment, els cladòcers Scapholeberis i Daphnia; i de tant en tant, alguna larva del gènere Chaoborus. Però la major part de la biomassa és constituïda per microorganismes. A l’epilímnion sols hi ha alguns dinoflagel·lats i diatomees; i a l’hipolímnion, totalment anaeròbic i amb concentracions de sulfhídric elevades (entre 1 i 2 mM), bacteris anaeròbics.
El metalímnion (que correspon a una oxiclina) resulta la zona més interessant. Els canvis sobtats en tots els paràmetres físico-químics fan d’aquesta zona un lloc amb multitud d’hàbitats que poden ser ocupats per microorganismes de metabolismes diferents. Aquí coexisteixen, en una franja molt estreta, l’oxigen, la llum i el sulfhídric. A la part superior del metalímnion hi ha poblacions de dos protists: Cryptomonas phaseolus (fins a 105 cèl./ml) i Coleps hirtus (fins a 104 cèl./l). Sembla que el primer és principalment fotòtrof, encara que hom no ha exclòs que pugui tenir un metabolisme mixt o que, fins i tot, mengi bacteris. El segon és un ciliat que conté cèl·lules de Chlorella endosimbiòtiques, cosa que fa sospitar que té un metabolisme parcialment fotòtrof. Les concentracions d’aquests dos microorganismes són elevadíssimes i l’aigua d’aquesta zona és de color marró, a causa dels pigments de Cryptomonas phaseolus. Per sota d’aquesta capa hom troba poblacions de dos bacteris vermells del sofre, Chromatium minus i Amoebobacter, amb 106 cèl./ml cadascun. Com a conseqüència, l’aigua d’aquestes fondàries és de color vermell. Barrejats amb aquests bacteris també hi ha bacteris incolors del sofre. A alguns centímetres per sota pot haver-hi poblacions de bacteris verds del sofre, normalment constituïdes per diferents espècies del gènere Chlorobium, algunes de color verd, com ara C. limicola, i d’altres de color marró, per exemple, C. phaeobacteroides.
Un dels aspectes més interessants d’aquesta comunitat és la variació de la posició dels microorganismes a la columna d’aigua durant un cicle dia-nit. Durant el dia, els microorganismes mòbils es desplacen uns 40 cm cap a la superfície, cercant la llum. Cap al vespre, els organismes tornen a nedar vers la part inferior, per passar la nit en aigües més fredes i anaeròbiques. Mentre hi ha llum, els organismes fotòtrofs realitzen la fotosíntesi fixant CO2. Els organismes eucariòtics Cryptomonas phaseolus, i potser també Coleps hirtus, produeixen oxigen i acumulen reserves de midó. Els bacteris fotòtrofs acumulen glicogen i sofre i oxiden sulfhídric. Així, doncs, durant les hores de llum, tot contribueix a oxigenar el metalímnion. Durant la nit, Cryptomonas phaseolus descendeix i roman en una zona completament anaeròbica. Les reserves de midó van sent consumides i, cap a la matinada, les cèl·lules més grans es divideixen.
Sembla que aquesta comunitat, formada per Anuraeopsis fissa, Cryptomonas phaseolus, Coleps hirtus i bacteris fotòtrofs, és molt estable, i ha estat trobada en d’altres llacs de característiques similars, com ara l’estanyol del Vilar, i el llac d’Estanya.
Com que les poblacions fotòtrofes es fan ombra mútuament, les que es poden col·locar més amunt tenen un clar avantatge. En general, la presència d’algues filtra prou llum per a dificultar el creixement dels bacteris fotòtrofs. A l’estany de can Cisó sembla que hi ha una correlació inversa entre la quantitat de Cryptomonas phaseolus i de clorobiàcies. A més, els bacteris vermells també determinen l’espècie de Chlorobium que podrà desenvolupar-se per sota: per causa del diferents pigments accesoris de què disposa, C. phaeobacteroides predomina quan abunden les cromatiàcies, mentre que C. limicola és més abundant quan els bacteris vermells s’hi troben en baixes concentracions.
Un altre aspecte molt interessant d’aquesta comunitat és que hom hi ha descobert uns bacteris anaeròbics depredadors d’altres bacteris. Els dos gèneres identificats, Daptobacter i Vampirococcus, ataquen i devoren específicament bacteris fotòtrofs, i, per consegüent, són consumidors primaris, igual que el zooplàncton i els animals herbívors. Tanmateix, el paper que aquests bacteris depredadors anaeròbies poden tenir en l’ecologia del llac és encara desconegut.
Durant la resta de l’any, tot l’estanyol està barrejat, el sulfhídric arriba fins a la superfície i l’aigua esdevé totalment anaeròbica. En aquestes condicions, els organismes eucariòtics desapareixen totalment, amb l’excepció d’alguns exemplars de Cryptomonas phaseolus. En canvi, els bacteris fotòtrofs ocupen la totalitat de la columna d’aigua.
Per consegüent, es pot concloure que l’estratificació vertical de les poblacions de bacteris fotòtrofs anaeròbies del sofre compleix les regles més generals de la teoria ecològica. De fet, l’estany de can Cisó recorda un ecosistema de l’eó Arqueà (des de fa 3800 a 2500 milions d’anys), quan l’atmosfera de la Terra no tenia oxigen i els microorganismes procariòtics n’eren els únics habitants. En aquella època, els bacteris fotòtrofs anaeròbies eren els principals productors primaris i el cicle del sofre es tancava amb els bacteris reductors de sulfat, exactament com succeeix a d’estany de can Cisó durant gran part de l’any. Així, doncs, aquest estanyol particular constitueix un model excel·lent per esbrinar com eren els ecosistemes aquàtics fa uns 3000 milions d’anys. L’estanyol de can Cisó és una finestra oberta al passat del nostre planeta; només cal que hi mirem amb cura i entusiasme, amb imaginació i rigor.
El model de l’estany de can Cisó es repeteix de forma parcial o total en tots els llacs càrstics quan hi ha conjunció de llum, manca d’oxigen i força sulfhídric. Qualitativament, i sobretot quantitativament, és prou important per a fer un paper central en l’ecologia dels llacs càrstics.
Les llacunes temporànies i les basses

Una de les basses o estanyols de Bell-lloc del Pla (Plana Alta), d’origen càrstic. En aquesta comarca i a la del Maestrat n’hi ha diverses, disperses, i no són més que petites dolines de poca fondària, plenes d’aigua.
Joaquim Garcia Flor
Dins d’aquesta mateixa part de l’obra, hom ha parlat dels llacs o dels estanys com a entitats ben definides. Però entre aquests sistemes i allò que anomenem llacunes i basses hi ha tot un gradient de variació, principalment afectat per la relació entre la superfície de la cubeta i el volum d’aigua que allotja, que fa que aquests conceptes siguin difícils de delimitar. A més d’aquesta dificultat de precisar els límits entre ambdós tipus de sistemes hi ha, encara, un altre factor d’imprecisió: la manca d’equivalència entre els noms que reben aquests sistemes en els diferents països. Allò que en els països deficitaris d’aigua considerem una llacuna és considerat com una bassa en països amb abundància de llacs, i a l’inrevés; només cal recordar el cas de l’estany de Banyoles, entre nosaltres tingut per un llac (el segon de la península Ibèrica pel que fa a les dimensions) i comparar-lo amb les grans masses d’aigua continentals de l’Amèrica del Sud, que allà hom anomena «lagunas».

Les basses, els tolls i les llacunes temporànies són sistemes limnètics extraordinàriament diversos, que es poden diferenciar dels llacs més per la seva feble fondària, que impedeix la formació d’una termoclina estable a l’estiu, que no pas per la seva extensió o volum. Als Països Catalans són petites (les llacunes de la depressió de l’Ebre, antigament tan extenses, han desaparegut fa temps), però són interessants per la seva elevada productivitat biològica, que permet que concentrin una gran biomassa. L’estany de Vilaüt, a l’Empordà, d’interès biològic reconegut, pot ser-ne un bon exemple.
Jordi Vidal
És cert que, en principi, allò que defineix una llacuna i la separa d’un llac és que no participa dels principis limnològics fonamentals d’aquests: no té estratificació vertical a l’estiu, el fons de la cubeta pot ser ocupat per vegetació en tota la seva extensió, etc. Però, d’una manera general, el concepte queda definit pel grau d’interacció entre el medi aquàtic i el medi terrestre que l’envolta, i una bona mesura és la relació entre la superfície inundada i el volum d’aigua: els valors grans indiquen una interacció més important i el sistema s’acosta més al model de bassa; els valors petits, per contra, s’apropen més al model de llac.
L’interès científic que tenen les petites basses d’aigua i els tolls d’inundació temporal no es correspon, en general, amb els esforços dedicats al seu estudi. La causa principal d’aquesta poca atenció segurament és la manca de models d’estudis fets als països pioners en limnología (Europa i Estats Units, d’on ens arriba la major part de la literatura científica), països en els quals és enormement més gran la quantitat de sistemes aquàtics d’envergadura i on solen menysprear aquests sistemes que per a nosaltres constitueixen el tipus més freqüent, autòcton i de valor regional. En contra d’allò que hom pogués pensar, els sistemes aquàtics petits i sovint afectats de variacions marcades al llarg de l’any contenen organismes d’un interès extraordinari. Com que són sistemes d’una gran diversitat, en els quals les variacions ambientals són molt acusades i les possibilitats de riscs catastròfics molt altes, les biocenosis que s’hi allotgen són fortament especialitzades i pràcticament exclusives d’aquests ambients, de tal manera que actualment tenen per a nosaltres un gran interès ecològic i biogeogràfic. Des del punt de vista històric, es comporten com a refugis de fauna relicta i d’organismes molt antics, formes que han tingut una gran importància en l’evolució de molts altres organismes. Hi ha, encara, altres valors igualment destacables, amb relació a les basses i els tolls d’aigua: a més de la seva importància en el regatge i en la ramaderia, hom els pot atribuir també una rellevància paisatgística notable, ja que són una nota de color enmig dels paisatges unicroms propis de les zones àrides i, alhora, constitueixen llocs de repòs per als ocells de pas.
Estructura, dinàmica i funcionalitat
Esquema idealitzat del cicle anual d’una bassa temporània, en el qual s’han representat les estratègies més freqüents per a resistir el període de dessecació. Algunes espècies (5) s’encisten en un estat de desenvolupament avançat, de manera que en la nova fase d’inundació aviat hi ha adults; d’altres (6, 7, 8) deixen ous durables protegits per a resistir la dessecació, o (9, 10, 11) utilitzen la fase humida per als primers estadis de desenvolupament i abandonen la bassa en forma d’adults, quan s’asseca. Les algues unicel·lulars i la vegetació aquàtica es desenvolupen gradualment, i deixen les espores o les llavors enterrades al fang; més tard apareixen les comunitats terrestres de substitució. Insectes aquàtics, ocells i comunitats terrestres tenen un paper important en la transferència d’energia entre aquests i altres sistemes. 1 Vegetació nitròfila, 2 insectes terrestres, 3 ocells, 4 ocells aquàtics, 5 ciclòpides, 6 anostracis, 7 diaptòmides, 8 cladòcers, 9 notonèctids, 10 odonats, 11 coleòpters, 12 algues unicel·lulars, 13 ranuncles aquàtics, 14 caròfits, 15 ocells limícoles.
Román Montull, original de l’autor
La bassa, encara que és de petites dimensions, té unes característiques pròpies i un funcionament ben determinat: configura un ecosistema. En termes generals, l’organització d’una llacuna o una bassa és similar a la d’un llac, però en petit; hom fa servir la mateixa terminologia per a definir-hi elements equivalents.
La colonització vegetal d’una bassa s’endreça fent cercles de diferent fondària, i també d’acord amb tres estratègies biològiques, que defineixen els tres tipus biològics principals: els helòfits (canyes, bogues, etc.), que necessiten tenir les rels inundades, però tenen l’aparell vegetatiu aeri; els amfífits, que tenen les fulles inferiors submergides i les superiors aèries o flotants, que porten aire cap a les rels (com els ranuncles o els lliris d’aigua); i els limnòfits, que viuen submergits, completament independents del medi aeri (fanerògames, molses i algues carofícies). Hi ha, encara, altres estratègies biològiques que són d’una gran importància en la xarxa alimentària d’aquests sistemes: la del perífiton i la del fitoplàncton.
Pel que fa a la colonització animal, els grups més representats en aquests sistemes són invertebrats, principalment larves d’insectes (coleòpters, efemeròpters i dípters, sobretot), insectes adults (hemípters, coleòpters, etc.), crustacis, rotífers, àcars, nematodes, turbellaris, mol·luscs, oligoquets i hirudinis; entre aquests, alguns tanquen tot el seu cicle vital dins de l’aigua (com els crustacis), mentre que d’altres utilitzen la bassa només en una fase del seu desenvolupament. Entre els vertebrats, tenen una incidència igualment important en el sistema els amfibis i els rèptils, que poden viure-hi durant una etapa més o menys llarga del seu desenvolupament, com els ocells, que només s’hi estan de pas, però hi fan un paper de transportadors important. Es poden distingir bé els animals bentònics, però per contra, no es pot parlar en aquests sistemes d’un autèntic zooplàncton, perquè l’escassa fondària d’aigua fa que els seus organismes es relacionin sempre d’una manera o d’una altra amb el substrat del fons. Els microorganismes neustònics, associats a la pel∙lícula superficial de l’aigua, són abundants en aquests sistemes, i inclouen especialment algues (clorofícies i cianofícies, sobretot), pol·len i tricomes de plantes superiors, fongs i amebes.
El cicle de matèria i energia és igual al dels llacs tot i que el flux de matèria i enegia és més obert: hi ha una entrada important d’elements al·lòctons detrítics i de llum, i una sortida en forma d’explotació per part dels depredadors i d’emergència dels individus adults voladors a partir de formes larvals aqüàtiques. El metabolisme d’aquests petits sistemes aquàtics és molt elevat a causa del grau alt d’il·luminació que reben totes les parts de la cubeta, cosa que permet una activitat biològica intensa. El retorn dels nutrients del sediment a l’aigua, en lloc de fer-se, com en els llacs, per barreja vertical a determinats moments de l’any, el fan els vegetals per bombament actiu del sediment a la superfície. En les aigües més somes, el bentos sol tenir avantatges sobre el plàncton en la competència pels nutrients i, malgrat que són sistemes molt eutròfics, les aigües tendeixen a ser relativament netes.
En aquests sistemes, la substitució d’unes espècies per unes altres al llarg del temps és molt patent, i es pot diferenciar clarament entre la successió anual i la successió a llarg termini, amb relació a la maduresa creixent del sistema.
Criteris per a una tipificació
Hi ha molts tipus diferents de basses, segons les característiques ambientals, de manera que hom en pot fer una classificació. La limnologia tradicional sol fer-la considerant separadament el medi físic i el medi biològic, però actualment hom tendeix a pensar que una classificació físico-ambiental i una altra de biològica fetes per separat coincideixen en gran part en els seus resultats.
Des del punt de vista ambiental, els paràmetres que són més importants en aquesta classificació són el règim d’inundació de la cubeta i les característiques de l’aigua.
El règim d’inundació de la cubeta

A les depressions naturals de terrenys argilosos poden formar-se basses que acumulen aigua de pluja. Si la cubeta és prou profunda o si hi ha entrades d’aigua subterrània la bassa pot ser permanent. Moltes vegades, hom n’aprofundeix la cubeta artificialment, com el cas de la fotografia, d’una bassa de El Campillo (Baix Cinca), i això permet aprofitar-la com a abeurador de bestiar, fins i tot a l’estiu.
Román Montull
L’existència d’una bassa comporta la d’un substrat impermeable. Des del punt de vista hidrològic, les basses poden ser llocs de descàrrega d’aqüífers importants; també poden dependre d’aqüífers de rius i d’aqüífers superficials de poca potència, que depenen exclusivament de la pluviositat local. Les basses poden mantenir l’aigua més o menys temps segons la pluviositat i la fondària de la cubeta, factors que, alhora, es relacionen amb d’altres, més difícils de quantificar, que pertanyen al camp de la hidrogeologia. Les fluctuacions de nivell poden associar-se a la dinàmica dels aqüífers, i, també, a les basses de pluja la persistència de l’aigua pot posar-se simplement en funció de la pluviositat local i de la profunditat de la cubeta. En termes generals, el balanç d’aigua es pot resumir en una equació en la qual la suma de les entrades superficials, les entrades subterrànies i la precipitació local ha de ser igual que la suma de l’evaporació i la pèrdua per drenatge. Als Països Catalans, encara que la pluviositat regional és molt variable, es pot considerar que la fondària «de seguretat» a partir de la qual les aigües són permanents és d’1,5 a 2 m.
Entre les diferents possibilitats, hom distingeix les estrictament permanents, que mai no s’assequen; les temporals, que tenen un cert risc d’assecament; les periòdiques, que tenen un període de sequera regular, que pot ser d’un any (tenen aigua a l’època plujosa de cada any) o de més d’un any; i les efímeres, o d’assecament imprevisible, com, per exemple, les lligades a sistemes càrstics, que s’inunden i s’assequen sobtadament, sense que hi hagi un procés d’assecament regular.
Com és natural, alteracions tan importants per als organismes aquàtics, com poden ser les que afecten l’aigua, han de tenir implicacions molt importants sobre comunitats senceres d’organismes. En les aigües temporals, les adaptacions dels organismes es poden resumir de la manera següent. En primer lloc, la capacitat de sobreviure al període sec es fonamenta en tres mecanismes: abandonar el medi quan les condicions es fan adverses (cosa que fan, sobretot, els insectes odonats, coleòpters i heteròpters), produir formes latents que es conservin en el sediment, i produir ous durables (per exemple, els crustacis i les carofícies). En segon lloc, la capacitat de suportar grans variacions ambientals, ja que la reducció del volum d’aigua comporta canvis de la composició química i de la capacitat de tamponament tèrmic; aquests canvis poden ser afrontats pels organismes que presenten eurihalinitat o euritèrmia. En tercer lloc, l’adaptació dels cicles vitals a períodes humits de durada imprevisible: la velocitat dels processos vitals s’incrementa amb vistes a aconseguir aviat la maduresa sexual, tot disminuint la possibilitat de morir sense descendència (estratègia molt notable en els eufillòpodes i en els caròfits, que ha permès que aquests organismes tan antics no hagin estat, encara, desplaçats per d’altres espècies més evolucionades), i es produeixen formes de resistència. La vida a la bassa es converteix, així, en una cursa per la supervivència, i, cada període humit, circumstàncies diferents, mostra condicions probablement irrepetibles.
A les aigües permanents, les adaptacions dels organismes responen a factors d’altres tipus, com ara la temperatura, la salinitat o l’estat tròfic, i a les relacions amb d’altres espècies. La persistència de les aigües permet la colonització continuada, de manera que les biocenosis es van complicant al llarg del temps fins a arribar a situacions estables, en les quals la probabilitat de canvis estructurals o d’introducció de noves espècies és reduïda al màxim. Aquests sistemes afavoreixen l’existència de nínxols ecològics nombrosos, ocupats per una gran diversitat de gèneres de vida, que allarguen les xarxes tròfiques.
La terbolesa

Poblament, en un estany de jardi eutrofitzat i sotmès a dessecacions periòdiques, d’algues dels gèneres Scenedesmus i Botryococcus, principalment responsable de la coloració intensament verda (Olesa de Bonesvalls, Garraf).
MC
La terbolesa depèn de la concentració de fitoplàncton a l’aigua, o bé del material inorgànic en suspensió. La seva intensitat té una repercussió important en l’ecosistema, ja que determina les possibilitats del creixement de la vegetació arrelada al fons. Les aigües molt eutròfiques, amb molt fitoplàncton, generalment són sistemes sotmesos a tensió, que delaten una situació anòmala (generalment es tracta de basses contaminades). Quan la terbolesa es deu a matèria inorgànica en suspensió, pot ser més o menys persistent: és temporal quan coincideix amb les etapes inicials d’ompliment de la cubeta i s’acaba amb l’assentament dels sediments, i és permanent en terrenys argilosos, capaços de cedir a l’aigua partícules fines que poden romandre resuspeses formant una suspensió col·loïdal.
En el cas de les aigües tèrboles, les llacunes es mantenen en fase heterotròfica permanentment, ja que el bentos no és colonitzat per plantes i hi predominen els animals detritívors i els carnívors. L’energia entra al sistema en forma de detrits orgànics, que poden ser assimilats com a matèria orgànica dissolta per bacteris i fongs o bé directament pels detritívors, i és exportada pels insectes, pels ocells i pels mamífers.
La mineralització i els nutrients
Entre les característiques químiques de l’aigua, les més importants són la riquesa de nutrients i la mineralització total; en ambdós casos, el grau de variació és molt ampli.
Els nutrients provenen de la pluja, del rentatge dels sòls (principalment el nitrogen, i el fòsfor quan hi ha camps de conreu pròxims tractats amb adobs químics) i de la pol·lució. En les basses que es troben en fases madures de colonització els nutrients no solen trobarse dissolts a l’aigua en concentracions gaire elevades.
La mineralització total es deu fonamentalment als ions principals (carbonat, bicarbonat, clorur, sulfat, calci, magnesi, sodi i potassi), i la seva quantitat absoluta i relativa depèn del substrat i del règim hídric de la bassa. En substrats poc solubles (granits, esquistos, quarsites), la mineralització acostuma a ser baixa (menys de 500 µS/cm de conductivitat elèctrica de l’aigua), mentre que en substrats més solubles és més elevada: en calcàries, la conductivitat de l’aigua pot ser de 2000 a 4000 µS, i en evaporites, fins a 30 000 µS. D’altra banda, l’enriquiment de sals també és influït indirectament pel clima: en llocs secs resulta afavorit l’endorreisme (procés pel qual les aigües que donen lloc a les llacunes no tenen drenatge horitzontal i la seva via de sortida del sistema és només per evaporació). Les aigües sotmeses a aquest règim tendeixen a concentrar sals i poden arribar a atènyer salinitats molt superiors a la de la mar. Aquest mecanisme es complica encara més en sistemes lacustres lligats a situacions de descàrrega d’aqüífers, ja que, ací, la natura química de l’aigua es troba molt alterada per processos d’envelliment i, a més, integra les característiques de tots els substrats per on han passat les aigües subterrànies, que, en el cas de paquets sedimentaris terciaris, poden ser complexes, ja que de vegades inclouen fins i tot nivells salins. Les relacions de proporcionalitat entre els diferents ions mostren una gran variabilitat perquè, en general, les aigües de les basses s’allunyen dels equilibris habituals, tant de les aigües continentals de volum gran (les dolces), com de les aigües marines (les més salades). Les aigües dolces acostumen a ser més riques en bicarbonats, perquè el carboni pot ser incorporat a partir de l’atmosfera; no obstant això, aquest anió també pot ser dominant en aigües més mineralitzades, quan les concentracions de calci són més elevades (en zones calcàries, típicament) o quan ho són les de sodi (en sistemes esteparis endorreics o lligats a descàrregues d’aqüífers regionals). En sistemes pròxims a la costa, les aigües dolces també poden ser enriquides per clorurs, que els arriben per la pluja i el vent. En aigües més mineralitzades poden dominar els clorurs o els sulfats, segons la natura del substrat, i, en cas que es trobin sotmeses a evaporació, segons l’estadi de sequera en què aquestes es trobin, ja que pel fet que el producte de solubilitat del sulfat és més baix que el de clorur, a mesura que les aigües es concentren, el clorur pot incrementar-se proporcionalment.
Amb relació a la salinitat, l’adaptació dels organismes pressuposa l’existència de mecanismes fisiològics variats destinats a regular les relacions osmòtiques entre el medi intern i l’ambient. Hom pot distingir tres tipus d’adequació. La primera es basa en el manteniment del medi intern hipertònic en aigua dolça, però amb la impossibilitat de regular-lo en aigües hiperosmòtiques, per la qual cosa aquests organismes només poden viure a les aigües dolces. En segon lloc, hi ha organismes que són isosmòtics en aigües que tenen una salinitat aproximadament igual que la del mar i no toleren desviacions respecte d’aquesta; aquests organismes poden viure en aigües salabroses del litoral, però es van enrarint a mesura que ens allunyem de la costa. El tercer tipus d’adequació a la salinitat caracteritza els organismes que regulen eficientment (osmoreguladors) i poden mantenir el medi intern en aigües amb diferents salinitats, és a dir, els organismes eurihalins; en aquests organismes, la capacitat d’homeòstasi ha estat adquirida versemblantment com una adequació a la vida en les aigües continentals, molt més variables i diversificades que les de la mar, de manera que els colonitzadors de les aigües atalasohalines salades presenten una afinitat filogenètica més gran amb les espècies de les aigües dolces que no pas amb les d’aigües marines o litorals. La salinitat intervé, almenys en alguns grups d’organismes, en la fenogènesi d’una manera semblant a com ho fa la temperatura, fins i tot accelerant-ne el desenvolupament.
La influència de les diferents proporcions aniòniques no sembla important en les aigües dolces, ja que com més petita és la mineralització total, les aigües mostren una concentració relativa de carbonats i bicarbonats més gran. En les aigües molt mineralitzades, per contra, les concentracions iòniques tenen més importància; a mesura que la relació Cl-/ SO2-4 augmenta, s’arriba a una situació més semblant a la de la mar i apareixen més espècies que són emparentades amb les de la costa.
Les basses permanents

Les basses i els tolls permanents afavoreixen la presència d’una comunitat rica i diversa, pròxima a la que es pot trobar a les vores d’un llac. Els macròfits (Potamogeton, a la fotografia) poden desenvolupar-s’hi amb abundància i constituir la base d’una comunitat variada d’animals. És en aquests ambients on els ocells migradors poden trobar recursos per a recuperar forces o, si l’ambient és prou gran i complex, fins i tot criar-hi.
Ramon Margalef i Mir
Són les que atenyen més volum i, en general, superen els 2 m de fondària. En realitat, aquest tipus de bassa és escàs al nostre país, i generalment, les que hi ha, són d’origen artificial. A les zones de poca altitud, la majoria són d’origen freàtic, com els ullals de Tortosa, per exemple, o fluvials, amb una marcada tendència a la distròfia, amb sediments torbosos, o amb relació als sistemes càrstics, com les basses de la zona de Banyoles. A les zones altes, hi són representades per les llacunes d’origen glacial (basses i estanys dels Pirineus).
La transparència de les aigües en aquests sistemes permet el desenvolupament d’una vegetació abundant, la qual forma diferents zones distribuïdes segons la profunditat. A les llacunes més profundes es desenvolupa una vegetadó de fons, resistent a la baixa intensitat de llum. Les comunitats d’helòfits són variades; la jonca (Schoenoplectus lacustris) hi és un colonitzador habitual, com també espècies pròpies de llocs inundats: el lliri groc (Iris pseudacorus) i la menta d’aigua (Mentha aquatica), entre altres. Les comunitats de limnòfits poden arribar a ser-hi molt denses, amb Myriophyllum, Ceratophyllum, utriculària (Utricularia vulgaris), diverses espècies de potamogètons (Potamogeton coloratus, P. pectinatus, etc.), i algues carofícies (Nitella translucens. N. flexilis, Chara globularis, C. vulgaris, C. hispida). Entre els amfífits són corrents Polygonum amphibium, el lliri d’aigua blanc (Nymphaea alba) i diversos ranuncles (Ranunculus) i potamogètons (Potamogeton natans, P. gramineus). El plàncton animal en aquests sistemes és empobrit, ja que generalment hi ha peixos que, si bé no solen ser planctívors, ho són en la fase d’ alevins. El litoral es troba densament poblat per rotífers bentònics (Plathyas quadricornis, Trichotria pocillum, Euchlanis dilatata, Lecane) i larves d’insectes (Cloeon dipterum, Plea, ditíscids, driòpsids), àcars i mol·luscs. En el plàncton és característica la presència de Daphnia longispina i Tropocyclops prasinus en aigües de la plana, i Cyclops abyssorum en les de muntanya, sempre formant part d’àmplies comunitats.
Les llacunes i basses temporànies

L’existència d’un període d’assecament és un fenomen comú en les basses somes, que pateixen una evaporació elevada i que no reben altra entrada d’aigua que la de pluja. Després de la dessecació, el sediment acumulat s’esquartera reté entre les escletxes restes d’organismes, petits animalons vius i ous i formes de resistència diversos que esperen el nou període l’inundació. El fang és, doncs, la llavor per a l’any vinent.
Miquel Alonso
Les aigües temporànies al nostre país formen petites cubetes, generalment de menys d’un metre de fondària. Les d’aigües menys mineralitzades es formen a les zones de muntanya a l’estiu (Pirineus) o sobre materials sorrencs o amb graves, en llocs més baixos, a l’hivern i a la primavera (per exemple, els estanys de Tordera i can Nadal). Sobre substrats miocens (Baix Urgell, Casp), a la plana, i sobre calcàries (Prepirineus, Banyoles) es poden trobar cubetes amb aigües més mineralitzades i tèrboles.
La colonització vegetal de les basses temporànies sol ser protagonitzada per algues carofícies, entre els limnòfits, i per ranuncles entre els amfífits, aquests molt representatius ja que en l’època de florida (primavera) donen una imatge molt característica d’aquestes basses, en cobrir-les de vegades completament de flors blanques. A la zona central més profunda, si es manté una mica d’aigua durant tot l’any, hi pot créixer Zannichellia palustris, Groenlandia densa o Potamogeton pectinatus (en aigües lleugerament mineralitzades), mentre que a les zones que s’assequen cada any, hi viuen espècies de cicle curt, com carofícies (Nitella opaca, Tolypella glomerata), Callitriche, Elatine, Mentha, etc.). A les de muntanya, hom pot trobar-hi l’associació d’isoet (Isoetes lacustris) i subulària (Subularia aquatica), amb espargani de muntanya (Sparganium affine borderei). Al litoral, els helòfits solen ser representats per joncs de poca alçada (Juncus), castanyoles (Cyperus) i eleocaris (Eleocharis palustris). Entre les comunitats animals, en els abeuradors de bestiar abunden les sangoneres (hirudinis) i els insectes heteròpters i odonats, els gasteròpodes i els amfibis (granotes i tritons).
A les més efímeres és característic el desenvolupament de grans poblacions de crustacis eufil·lòpodes, com ara anostracis, tortuguetes, etc. Els primers omplen els tolls que es formen a les zones de muntanya (com els del pla de Beret) a l’estiu, i els segons viuen als arrossars. Les basses que es formen als costats dels rius durant les avingudes primaverals també han de ser considerades ací, ja que són de formació efímera. Un altre cas és el de les basses que es formen en relació amb els rius quan el riu porta poca aigua, en època de sequera; aquestes basses constitueixen sistemes que es recolonitzen cada any, ja que quan el riu torna a anar ple, s’enduu avall les espècies colonitzadores, i tenen la particularitat de funcionar com a basses precisament en l’època que les basses temporànies són seques.
Als terrenys càrstics, és típica la formació de basses que s’inunden ràpidament i s’assequen també imprevisiblement. El cas del conegut Clot d’Espolla (a la zona de Banyoles) n’és un exemple ben vàlid i comparable, pel seu funcionament i per la seva biota, al sistema dels arrossars. En terrenys àcids, amb sorres o gresos, les basses que es formen són colonitzades per comunitats d’organismes comparables a les de les basses de muntanya.
Les basses temporànies o semitemporànies que es formen sobre terrenys argilosos de zones estepàries presenten una fisiognomia característica: absència gairebé absoluta de macròfits (o únicament, una estora monospecífica de carofícies al fons i alguns ranuncles al litoral) i desenvolupament d’una comunitat de crustacis típica, on dominen els eufil·lòpodes, cladòcers i copèpodes. Basses d’aquest tipus es troben disperses per tot el país, allà on el substrat en permet l’existència (a l’Urgell, als Ports, a la Terra Alta, etc.). Generalment tenen poques espècies helofítiques, però de vegades hi pot créixer l’eleocaris (Eleocharis palustris). En substrats argilosos acostumen a ser molt tèrboles, cosa que impossibilita la presència de limnòfits al fons de la cubeta. Com hem dit més amunt, aquest tipus de basses pot contenir aigües salades, com les abans conegudes basses d’Ivars d’Urgell, que foren assecades artificialment, i les d’Alcanyís.
Les basses naturals que hom aprofita com a abeuradors de bestiar solen ser aprofundides artificialment a la seva zona central per tal de prolongar el període de permanència de l’aigua. Aquestes basses solen tenir més nutrients i ser lleugerament mineralitzades i tenen la particularitat que poden allotjar alhora comunitats pròpies d’aigües temporals, a les vores, i comunitats típiques d’aigües permanents, al centre.
Els arrossars, d’altra banda, són un sistema particular, també d’inundació temporània, sotmesa a les exigències del conreu i, per tant, el seu cicle es pot considerar aleatori pel que fa a les estratègies adaptatives de les espècies. Solen fer-s’hi grans pradells de carofícies (Chara vulgaris i C. braunii especialment) i fanerògames (Zannichellia palustris, molt comuna), que són explotats per les tortuguetes (notostracis), que arriben a convertir-s’hi en flagell.