Consideracions generals
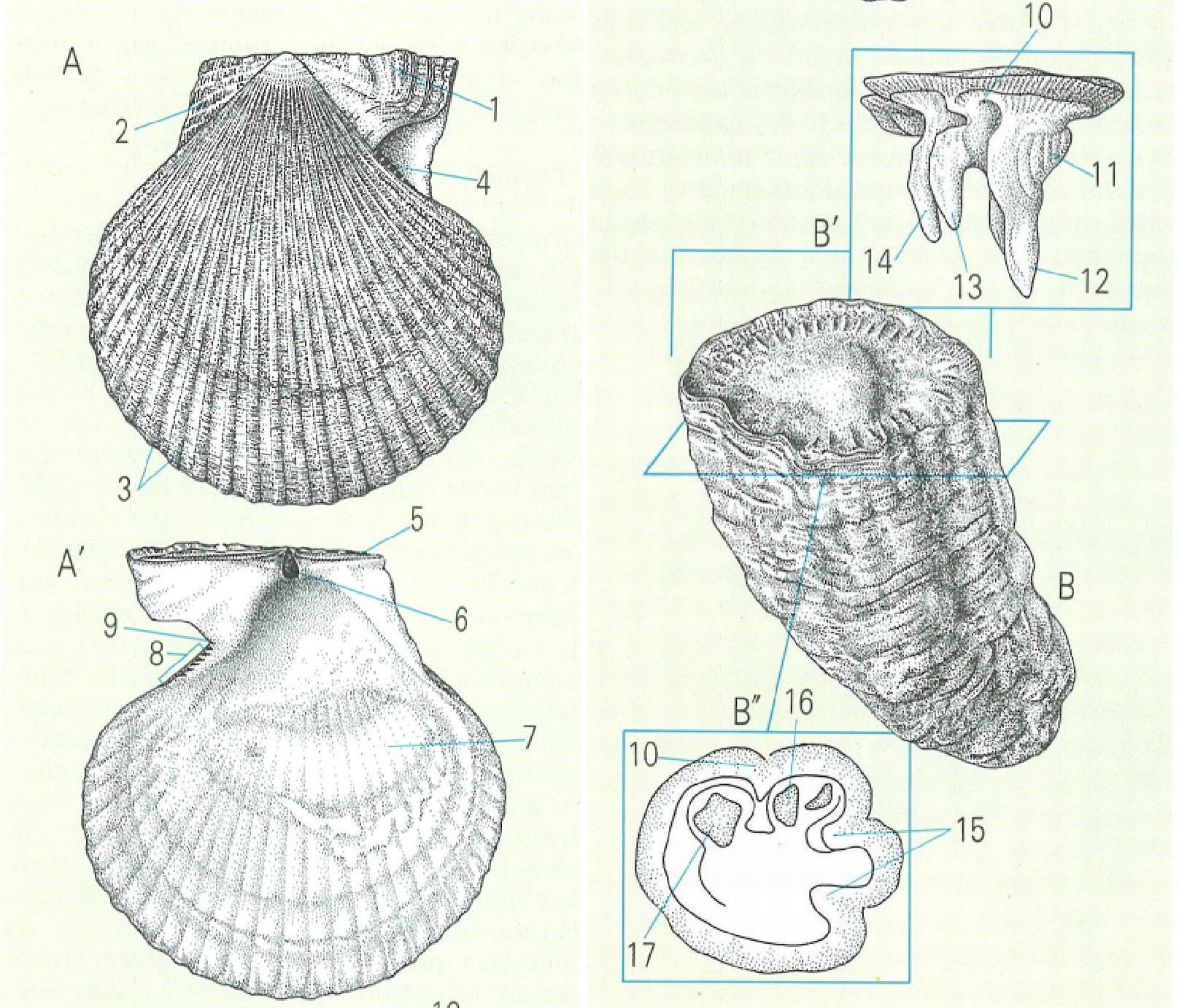
La morfologia general dels bivalves o lamel·libranquis pot ser molt modificada respecte del model de vida lliure proveït de dues valves simètriques, que en el dibuix és representat per un pectínid (A, A’ cares exterior i interior d’una valva dreta). Potser el cas més complex de modificació és el dels rudistes, en què una de les valves, en forma de con (B), és la que es fixa al substrat, mentre que l’altra pren forma d’opercle (B’; B" secció transversal de la valva inferior) Hom ha indicat en el dibuix: 1 orelleta anterior, 2 orelleta posterior, 3 còstules o costelles, 4 dents pectinidials, 5 vora auricular, 6 resilífer, 7 impressió del múscul, 8 ctenòlium, 9 sinus bissal, 10 aresta lligamentària, 11 apòfisi per a la fixació del múscul adductor anterior, 12 dent anterior, 13 dent posterior, 14 apòfisi per a la fixació del múscul posterior, 15 pilars on recolzen els sifons, 16 fosseta que allotja la dent posterior, 17 fosseta que allotja la dent anterior.
Lluís Centelles, a partir de fonts diverses
Els bivalves són mol·luscs aquàtics de simetria bilateral, amb una conquilla de carbonat càlcic formada per dues valves que l’existència d’una xarnera en posició dorsal permet obrir i tancar; aquest moviment es fa gràcies a la presència d’un lligament elàstic i d’un sistema de músculs. Han rebut també el nom de lamel·libranquis (Lamellibranchia) i de pelecípodes (Pelecypoda), termes que fan referència a les característiques de les parts toves del cos, com la forma de les brànquies i la del peu, respectivament. Els òrgans vitals de l’organisme són envoltats d’un mantell format per dos lòbuls que corresponen a cadascuna de les dues valves. Es tracta d’un grup àmpliament diversificat, que s’ha adaptat, deixant de banda les condicions marines, a les aigües de baixa salinitat i, fins i tot, a les aigües dolces. Tanmateix, dins el medi marí és on hi ha un nombre més gran d’espècies, i on han ocupat nínxols i substrats diferents, i fins i tot hàbits alimentaris diferents. Geogràficament s’han estès per totes les latituds d’arreu del món, bé que, en determinats períodes dels temps geològics, alguns grups hagin tingut una distribució més aviat limitada.
Pel que fa al registre fòssil, són formes que apareixen des del Cambrià, tot i que al llarg de la seva història, alguns grups com els rudistes, d’àmplia significació en el Mesozoic, s’han extingit completament. Paleontològicament són importants tant en el camp de la biostratigrafia com en el de la paleoecologia.
La conquilla
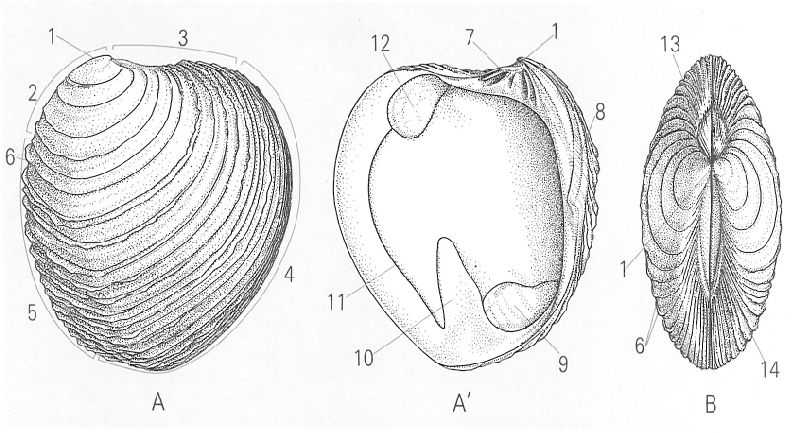
Característiques morfològiques generals de la conquilla dels bivalves: A, A’ valva dreta en visió externa i interna, respectivament; B visió dorsal. Hom ha indicat en el dibuix: 1 umbe, 2 marge dorsal, 3 marge anterior, 4 marge ventral, 5 marge posterior, 6 lamel·les, 7 xarnera, 8 lligament, 9 impressió del múscul adductor posterior, 10 sinus pal·leal, 11 línia pal·leal, 12 impressió del múscul adductor anterior, 13 lúnula, 14 escut.
Lluís Centelles, a partir de fonts diverses
La part inicial de la conquilla larval (protoconquilla o, més correntment, prodisoconquilla) moltes vegades es conserva, bé que només en traces, en estat fòssil; a partir d’ella la conquilla creix per deposició de carbonat càlcic a la vora lliure fins a constituir la forma adulta (teleconquilla). Les dues valves en contacte formen una comissura que, en general, és continguda en un pla que coincideix amb el pla sagital de l’organisme. Les valves s’orienten de tal manera que en relació amb el cos de l’animal es designen com a valva dreta i valva esquerra. En alguns casos, el contacte entre les dues valves no és complet i queden petites obertures per a la sortida dels diferents òrgans (el peu, els sifons, el bissus). En relació amb la simetria bilateral, la conquilla pot presentar fortes modificacions, de forma (com en alguns pectínids), de dimensions o d’ornamentació (cas dels hipurits i d’alguns ostrèids).
El marge de la closca on se situa la xarnera es considera el marge dorsal, i l’oposat el marge ventral o també pal·leal; el situat al costat de la boca és el marge anterior, i el contrari el posterior. D’aquesta manera, les valves queden dividides en quatre marges, els límits entre els quals, però, no sempre són ben delimitats. Quan les dues valves són iguals, és a dir, quan la simetria bilateral és perfecta, la closca és equivalva, i és inequivalva en el cas contrari; termes semblants s’utilitzen per a definir la igualtat o la divergència respecte dels marges anterior i posterior (valva equilateral i inequilateral, respectivament). La part inicial de la prodisoconquilla acostuma a tenir forma de punxa o de bec, i representa l’umbó (l’àrea que l’envolta forma la regió umbonal). La seva posició respecte de l’eix umbo-pal·leal permet establir tres tipus diferents d’umbons: ortògirs, si es troben enfrontats; opistògirs, si són corbats o inclinats posteriorment; o prosògirs si ho són anteriorment. En algunes formes d’hipurits, els umbons presenten un enrotllament espiral cap enfora del pla sagital: són els umbons qualificats d’espirògirs, com els del gènere Diceras, entre altres. Entre els ostrèids, als gèneres Gryphaea i Exogyra, la secció de l’umbó dibuixa una espiral d’una o més voltes que, en general, poden correspondre als tipus prosògirs o als opistògirs. A la regió dorsal, a cada costat de l’umbó, és freqüent la presència de dues àrees petites que presenten una ornamentació lleugerament diferent de la que es troba a la resta de la conquilla; aquestes àrees formen la lúnula, de mida més petita i situada al costat anterior, i l’escut, que en general és més allargat i es troba al costat posterior. Si els dos umbons són separats, hi ha una plataforma, que pot ser ampla, i que forma l’àrea cardinal, a la qual es fixa el lligament; això es pot apreciar bé en les conquilles d’Arca o de Glycymeris, per exemple.
Per a determinar quin és el marge anterior i quin és el posterior d’una valva solta (és a dir, si la valva és dreta o esquerra), hom utilitza diferents criteris: si l’umbó és inclinat cap a un dels costats, llavors el marge més curt correspon a l’anterior (hi ha excepcions en bivalves amb umbons opistògirs); la presència d’una carena única umbo-pal·leal sempre indica que aquell és el marge posterior; si hi ha plaques lligamentàries, sempre són al marge posterior; el sinus pal·leal sempre és al marge posterior; les impressions musculars úniques normalment es troben desplaçades vers el marge posterior; si una de les impressions musculars és molt més desenvolupada que l’altra, correspon al múscul posterior. En el grup dels ostrèids, però, cal emprar altres criteris.
La xarnera
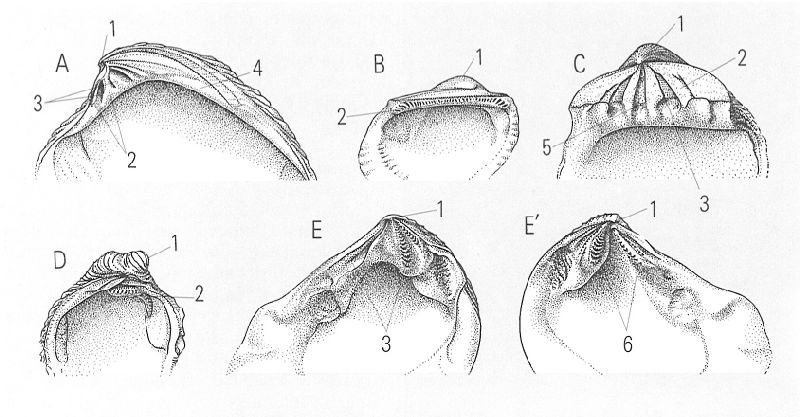
Principals tipus de xarneres en els bivalves: A heterodonta, amb dents cardinals sota l’umbe; B taxodonta, amb nombroses dents petites sota l’umbe que, en avançar cap als extrems, es van fent més grosses i s’inclinen cap a l’interior de la conquilla; C isodonta, amb dents situades simètricament a cada costat del resilífer; D paquidonta, amb una sola dent molt gruixuda sota l’umbe; E i E’ esquizodonta, amb dents bífides. 1 umbó, 2 dent, 3 fosseta, 4 dent lateral, 5 resilífer, 6 dent bífida.
Lluís Centelles, a partir de fonts diverses
La xarnera ocupa una posició dorsal per sota dels umbons. Una vista interna de les valves permet observar la morfologia i la disposició dels elements que la componen. Les estructures que sobresurten de la plataforma cardinal són les dents, mentre que les que hi són enfonsades representen les fossetes. Les estructures d’una valva encaixen perfectament amb les de l’altra. Això permet mantenir les valves juntes durant els moviments d’obertura i de tancament. Per la posició i la forma de les dents, hom distingeix les que se situen sota l’umbó (dents cardinals) de les situades en el marge anterior i posterior de la xarnera (dents laterals). La disposició, el nombre i la forma dels dos elements estructurals, dents i fossetes, donen lloc als diferents tipus de xarnera que tenen una gran significació en taxonomia i en la determinació de les espècies. La xarnera actinodonta es considera de tipus primitius, i té les dents allargades, que divergeixen a partir de l’umbó. La xarnera taxodonta, és llarga i estreta, amb dents curtes i nombroses; es considera que apareix secundàriament en els arcacis, i caracteritza Nucula, Arca, Glycymeris, etc. La xarnera esquizodonta té una dent bífida o partida, com a Trigonia. L’heterodonta té les dents diferenciades en cardinals i laterals, com en els venèrids. La paquidonta, típica dels rudistes, té les dents molt gruixudes. La isodonta té dues dents iguals a cada valva, simètriques, a cada costat d’una fosseta triangular, que correspon a la inserció del lligament, com a Spongylus. La disodonta té les dents molt petites i properes a l’umbe, com en el musclo (Mytilus). I la criptodonta té les dents poc manifestes; hom la considera característica de formes antigues, però també s’ha trobat en algunes formes recents.
Hom ha intentat, des de temps antic, d’establir sistemes que permetin expressar les característiques de la xarnera, i el que té més aplicació és el que pensaren Bernard i Munier-Chalmas. Es tracta d’una notació on figuren el tipus de dents i la seva ontogenia a partir de les làmines primitives. En aquesta notació s’indica, a dalt i amb nombres senars el que correspon a la valva dreta, i a baix i amb nombres parells el que correspon a la valva esquerra. Les xifres àrabs es refereixen a les dents cardinals i les romanes a les laterals. Les lletres corresponen a la posició de la dent: a i b per a les dents cardinals anteriors i posteriors, respectivament; i A i P per a les laterals, anteriors o posteriors, respectivament. En aquest tipus de notació, la correspondència entre dents i fossetes entre les dues valves també hi queda representada.
Cal assenyalar, encara, un altre fenomen que dóna més complexitat a la xarnera, i és l’anomenada transposició que presenta la d’algunes espècies, i que és especialment significativa en el cas dels càmids. En aquesta família la transposició va lligada a la fixació de la valva al substrat. Pot fixar-se tant la valva dreta com l’esquerra, però la que es fixa sempre presenta dues dents, mentre que la lliure en té una. Hom parla de disposició «normal» quan la valva fixa és la dreta, i de disposició «inversa» quan la fixa és l’esquerra.
El lligament

Vista dorsal (a dalt) i lateral (a baix) de Corbicula laletana, on es veuen les plaques (nimfes) sobre les quals es fixa el lligament. Característic del Maastrichtià del Prepirineu de Catalunya, aquest exemplar prové del barranc de la Posa (Pallars Jussà). Té un diàmetre antero-posterior de 28 mm.
Jordi Vidal / Col·lecció Jaume de Portal.
El lligament és una estructura elàstica, feta de material proteïnic (conquiolina) produït per l’epiteli del mantell. Té una estructura complexa, en capes que formen làmines, i serveix per a mantenir les dues valves juntes; és a dir, actua en sentit contrari als músculs. És per això que els exemplars morts que hom troba a la platja sempre tenen les valves obertes. El lligament se situa a la part dorsal de la conquilla i pot ocupar una posició externa o interna.
El lligament extern se situa al costat posterior (lligament opistodètic), però pot fer-ho a l’anterior (prosodètic); quan la disposició és a cada costat de l’umbe és amfidètic. La fixació del lligament es fa sobre una àrea més o menys gran i de morfologia variada (l’àrea lligamentosa). Algunes formes presenten plaques especials per a la fixació del lligament, com Corbicula laietana, força freqüent al Maastrichtià del barranc de la Posa, a Isona (Pallars Jussà). En el cas del lligament amfidètic, l’àrea lligamentosa és triangular i hi poden quedar marcades les posicions que ha anat ocupant el lligament durant el creixement; això es pot observar bé en Arca i Glycymeris.
En el cas del lligament intern, ocupa una cavitat més o menys triangular i fonda, que constitueix el resilífer. Presenten lligament intern, entre altres, els pectínids i el gènere Spondylus. Un resilífer ben particular és el que hom troba a Mactra i Lutraria (entre altres), ja que tenen el lligament fixat damunt d’una estructura sortint, i en aquest cas rep el nom de condròfor. També és especial el resilífer dels ostrèids. Hom utilitza, per a descriure el tipus de lligament, els termes alivincular, quan la capa lamel·lar es troba tant a la part anterior com a la posterior de la capa fibrosa; paravincular, quan és allargat longitudinalment i se situa posteriorment a l’umbe; multivincular, quan correspon a una successió d’elements de tipus alivincular (com a Isognomon); i duplivincular, quan la capa lamel·lar es repeteix en bandes, que són les que deixen els solcs característics en les conquilles d’Arca i d’altres gèneres.
Les impressions musculars

Les empremtes dels músculs adductors poden mantenir-se després del procés de fossilització de les conquilles. Noteu-les a la cara interna de la valva dreta d’aquest exemplar d’Arctica islándica (× 1,3), una espècie que actualment viu a l’Atlàntic Nord i que fou relativament abundant en les aigües costaneres de Catalunya durant el Würm.
Jordi Vidal / Col·lecció Jordi Martinell.
El cos dels bivalves es fixa a la part interna de les valves per diferents tipus de músculs. Sobre les diverses àrees de fixació queden ben marcades les impressions musculars. Sens dubte, les impressions que ocupen més extensión són les dels músculs adductors, l’acció dels quals permet mantenir tancades les dues valves i, en aquest sentit, actuen contràriament a l’acció del lligament. N’hi ha un parell a cada valva, l’un prop del marge anterior i l’altre en el posterior; la seva forma i dimensions són útils per a la determinació de les espècies. Si les impressions musculars s’han conservat bé, hom pot observar-hi dues àrees de mides diferents, que corresponen a la impressió de les fibres llises i les fibres estriades; això sovint es veu perfectament en alguns pectínids. Els bivalves que presenten dues impressions musculars s’anomenen dimiaris, i si són de dimensions semblants o iguals, isomiaris o homomiaris. En moltes formes hi ha una reducció de les dimensions del múscul anterior que, fins i tot, pot arribar a desaparèixer; en aquest cas, parlem d’anisomiaris. Dins d’aquest últim grup, si la diferència entre els dos músculs és gran, reben el nom d’heteromiaris, i si només queda el múscul posterior, de monomiaris; en aquest cas, acostuma a ser més gran i la impressió és més aviat central, com en els ostrèids.
Els lòbuls del mantell es fixen a la conquilla per mitjà d’un sistema de fibres musculars que també deixen una impressió, la qual segueix, és clar, un traçat més o menys paral·lel al del marge pal·leal i s’estén entre les impressions dels músculs adductors: és la impressió pal·leal o línia pal·leal, que pot ser contínua (integripalleal) o bé pot dibuixar una inflexió cap a l’interior de la closca (sinus pal·leal); el sinus es troba desplaçat cap al marge posterior, i és un dels caràcters morfològics emprats per a determinar si una valva és dreta o esquerra. Les característiques morfològiques i, especialment, la fondària del sinus, tenen relació amb la paleoecologia dels bivalves, ja que donen una mesura indirecta de la fondària a la qual es poden enterrar alguns elements de la infauna.
L’ornamentació

Dos tipus d’ornamentació característics en els bivalves. A dalt, conquilla amb les línies de creixement ben marcades de Panopea glycymeris, una espècie de la infauna profunda que sovint es troba fossilitzada com a motlle intern i que es coneix des del Miocè fins a l’actualitat; la conquilla, allargada en sentit antero-posterior i que fa uns 160 mm de diàmetre màxim, mostra una obertura entre les dues valves, a la part posterior. A baix, conquilla amb espines sobre les còstules radials de Cardium hians, especialment ressaltades vers el marge posterior on les còstules també esdevenen més anguloses. Aquesta espècie, de 69 mm de diàmetre antero-posterior, es coneix des del Miocè. Totes dues fotografies procedeixen del Pliocè del Baix Llobregat.
Jordi Vidal / MGB.
Com en altres grups d’invertebrats, la superfície de les valves presenta elements morfològics que, en conjunt, determinen la seva ornamentació. En general, l’ornamentació està relacionada amb els hàbits de l’animal i és un element morfològic funcional. Les línies de creixement, visibles a la valva, són determinades per engruiximents que representen interrupcions del ritme de secreció de carbonat càlcic. Però la veritable ornamentació és una escultura que es projecta al damunt de la superfície de la conquilla que respon a una disposició més o menys concèntrica, seguint un recorregut no del tot paral·lel al marge lliure. D’altra banda, també hi ha una escultura en disposició radial, que s’estén des de l’umbe fins al marge pal·leal; les costelles, separades per espais deprimits, que presenten molts pectínids en són un bon exemple. Hom utiltza una terminologia especial per a descriure l’ornamentació de les valves, segons la intensitat amb què es presenten aquests elements. No són poques les formes de bivalves que mostren una superposició dels dos sistemes d’ornamentació, concèntrica i radial, i prenen així una ornamentació reticulada; en aquests casos, és freqüent que en els punts d’intersecció hi apareguin tubercles, pústules, espines, etc. Una ornamentació particular és la divaricada, que pren el nom del gènere Divaricella, formada per parells d’elements divergents; hom creu que aquesta ornamentació facilita l’enterrament de la conquilla, tot i que és de forma circular.
Sistemàtica i evolució

Durant el Würm foren freqüents en les aigües costaneres espècies de mars fredes, com Chlamys islandica (× 0,7), que actualment viu a l’Atlàntic Nord. Noteu a la fotografia (valva dreta) la gran densitat de còstules, totes iguals, que caracteritzen l’espècie.
Jordi Vidal / Col·lecció Jordi Martinell.
Els bivalves constitueixen una classe de mol·luscs, molt abundant i diversificada. Les estimacions més moderades suggereixen un nombre d’espècies actuals pròxim a les 8000; si es consideren també les formes fòssils, hom en comptabilitza uns 9000 gèneres. La llarga història evolutiva dels bivalves des del Cambrià fins a l’actualitat permetria de seguir una extensa filogènia, però l’existència de convergències, paral·lelismes i adaptacions, entre altres factors, dificulten aquesta tasca. La comparació de les diferents obres de malacologia actual i fòssil manifesta la diversitat de criteris emprats per a seleccionar els caràcters sobre els quals hom estableix la classificació, i això es tradueix en una gran disparitat pel que fa a la fixació de les categories taxonòmiques i al nombre d’ordres, superfamílies i famílies.
La sistemàtica que hom sol emprar per als bivalves actuals pren com a caràcters bàsics els de les parts toves de l’organisme, com les bfànquies, l’estómac, etc., mentre que en els estudis de fòssils, hom ha donat més importància a la xarnera. Hom ha fet intents de buscar una classificació més objectiva, a base de comparar la informació en forma de taules i establir uns cladogrames que reflecteixen les possibles relacions filogenètiques. En aquesta línia cal citar els intents de Boss (1978) per a la superfamília dels padoracis, i la classificació de tots els bivalves de Purchon (1978), que relaciona caràcters de les parts toves amb caràcters de la conquilla. El diagrama obtingut en aquest darrer cas s’expressa en percentatges de similitud, i les agrupacions que s’obtenen coincideixen força amb les sis subclasses que tenen més acceptació actual, d’acord amb el «Treatise on Invertebrate Paleontology», de Moore (1969), i que seguirem ací: paleotaxodonts (Paleotaxodonta), criptodonts (Cryptodonta), pteriomorfs (Pteriomorphia), paleoheterodonts (Palaeoheterodonta), heterodonts (Heterodonta) i anomalodesmats (Anomalodesmata); hi ha, encara, qui pensa que caldria subdividir la classe en només dues subclasses: la dels protobranquis (Protobranchia) i la dels lamel·libranquis filtradors. Altres classificacions poden trobar-se a Pojeta (1971) i a Clarke (1985).
L’origen dels bivalves és encara una qüestió molt debatuda, especialment en dos punts: quin fou el primer bivalve, i quin és el grup que ha donat origen als primers bivalves.
A la part alta del Cambrià inferior de Nova York, als Estats Units, es va trobar (1881) un conjunt de formes molt petites que Barrande identificà com a un bivalve (Fordilla troyensis) que tenia dues valves amb impressions musculars, línia pal·leal i altres impressions que podrien correspondre al múscul retractor del peu; altres exemplars, procedents del Cambrià inferior de Sibèria, mostraven, a més de la línia pal·leal, una dent cardinal i una fosseta a cadascuna de les dues valves. No obstant això, hi ha altres opinions que contradiuen aquesta, i que consideren que F. troyensis no seria un bivalve. Pel que fa a la paleoecologia d’aquesta espècie, podria tractar-se d’un suspensívor infaunal, d’acord amb la compressió lateral de la conquilla; o bé podria viure sobre el fons marí. Posteriorment (1980) s’ha suggerit que l’hàbitat primitiu dels bivalves fou un substrat dur, i que la colonització del substrat no es devia donar fins després de l’Ordovicià. D’altra banda, al Cambrià inferior de Portugal es trobaren altres formes que podrien interpretar-se com a bivalves, entre elles Ctenodonta, que tenia una xarnera de tipus taxodont, i Modiolopsis (de la subclasse dels paleoheterodonts). A Murero (Saragossa) es va descriure (1962) un bivalve datat del Cambrià mitjà (segons la fauna de trilòbits que l’acompanyava), amb el nom de Lamellodonta simplex, que hom considerà com el bivalve més antic fins que una revisió posterior (1978) d’aquest material l’ha considerat com un braquiòpode inarticulat del gènere Tramatobolus (ordre dels obolèl·lids). Així, doncs, hom considera que els bivalves més antics trobats fins ara, Babinka, Redonia, Cycloconcha i Ctenodonta, corresponen al Tremadocià (Ordovicià inferior).
Pel que fa a l’origen dels bivalves, s’han assenyalat diverses vies. Una d’aquestes vies podria ser a partir dels rostrocònquids, i una altra dels monoplacòfors. La transició de formes univalves a la condició de mol·luscs bivalves devia fer-se a través de diferents gèneres, entre ells Heraultipegma, considerat per alguns com un rostrocònquid del Cambrià inferior. Al Cambrià inferior d’Austràlia s’han trobat un nombre de formes de dimensions petites (1,2 mm de llargada), que presenten una xarnera formada per una o dues dents a cada valva. Un conjunt de cinc o sis impressions musculars, de forma oval o arrodonida, s’estén entre els dos músculs adductors que són de mesures diferents. Aquesta forma, descrita com a Pojetia runnegari, s’interpreta com el bivalve més antic i correspondria a un paleotaxodont que s’enterrava en el fang carbonatad És considerat un «detritívor inhalant». El gènere Babinka, que apareix a dues àrees d’Europa (a Bohèmia i a la Montanha Negra), és una forma amb xarnera formada per dues dents i una sèrie d’impressions musculars, caràcter que suggereix una possible vinculació amb els monoplacòfors. La presència de nuculoides (Nuculoidea) en el Tremadocià de França i l’Argentina (Paleoneilo i Ctenodonta) indica que els bivalves ja es troben ben desenvolupats i que presenten unes característiques encara primitives.
A l’Ordovicià ja es troben representades la majoria de les subclasses de bivalves, i hi ha la primera gran diversificació del grup. Durant el Silurià, la fauna de bivalves resta molt semblant, tot i que cal assenyalar els primers representants dels pectinacis i el començament de la branca que desemboca en els rudistes, formes que tenen molta significació en el Cretaci, principalment l’europeu. Les primeres formes d’aigua dolça apareixen en el Devonià i tenen característiques molt semblants a les formes actuals; els pectínids ja hi són presents, amb els gèneres Aviculopecten i Posidonia. És durant el Carbonífer que els bivalves no marins adquireixen una gran importància per la seva abundor en determinats indrets i per l’interès que tenen en el camp de la biostratigrafia, ja que permeten establir correlacions en les conques carboníferes; en són gèneres significatius Anthracosia, Carbonicola i Naiadites, entre altres. Les faunes de tipus no marí continuen durant el Permià, i la fauna marina no presenta grans diferències, en general, respecte de la del Carbonífer.
El pas al Triàsic comportà l’aparició de diverses formes i l’extinció d’altres dins d’aquest període. Algunes formes de pectinacis són significatives del Triàsic, com per exemple, Daonella. Apareixen, també, per primera vegada els ostrèids i els trigònids, i tenen un gran desenvolupament els representants dels miofòrids. Durant el Juràssic, és especialment important l’aspecte d’algunes formes de pectínids, i l’abundància i la diversificació dels ostrèids (Liostrea, Exogyra i Lopha). Entre els hipurits, destaca l’aparició del gènere Diceras en el Juràssic superior (Oxfordià). L’últim representant dels modiomorfacis, el gènere Hippopodium presenta una dispersió discontínua. En el Cretaci, sens dubte, el fet més important és el desenvolupament dels rudistes, grup que presenta una morfologia molt diferent de la de la resta dels bivalves, i que desapareix completament al final del Mesozoic; és un grup molt ben representat a tot Europa i als Països Catalans, que ha tingut una forta rellevància en la biostratigrafia del Cretaci. Al mateix període, presenten una gran expansió els inoceràmids, que ja van fer l’aparició en el Juràssic, i també els trigònids, que encara són molt abundants. Entre els pectínids d’aquesta època cal destacar el gènere Neithea per la gran asimetria de les seves dues valves. També cal assenyalar l’aparició dels càmids, que tenen una manera de fixar-se al substrat semblant a la dels rudistes. Tota la resta de les subclasses de bivalves ja tenen una àmplia representació durant el Cretaci.
En el Terciari, els bivalves ja tenen un aspecte modern. Alguns grups tenen una àmplia expansió: els ostrèids, que cobreixen un espectre d’adaptació a diverses condicions i despleguen una gran plascititat morfològica, en ocupar medis diversos; i els mateixos pectínids, que són abundantíssims en el Miocè i el Pliocè i presenten una gran significació biostratigràfica. La immensa majoria dels bivalves terciaris tenen representants actuals.
El grup que ha experimentat més reducció en el nombre de formes deu ser la família dels trigònids, de la qual actualment només viu un gènere, a la part oriental d’Austràlia.
Durant el Quaternari, el fet més important per les àrees de latituds mitjanes respon a les oscil·lacions que presenten les faunes marines al ritme de les variacions climàtiques. Així, és freqüent veure en dipòsits de les nostres costes, com ara a l’Hospitalet de l’Infant (Baix Camp) o en terres alacantines, faunes qualificades de càlides, que ara viuen en regions tropicals o en indrets molt puntuals de condicions especials, i faunes fredes, com les que s’han trobat a la plataforma del davant de les costes del Principat, amb exemplars d’Arctica islandica, Chlamys islandica, etc., que avui no baixen per sota de les costes de l’Atlàntic del N d’Europa.
Des del punt de vista de l’alimentació, també hi ha hagut una evolució que ha permès abandonar el model detritívor, primitiu. Per canvis estructurals en els òrgans de la cavitat del mantell, els palps labials, els ectenidis i l’estómac, es desenvolupà un nucli d’organismes suspensívors que es nodrien de partícules orgàniques en suspensió. És a partir d’aquest moment que comença la possibilitat de colonitzar altres hàbitats, d’acord amb el desenvolupament dels tipus de brànquies (fil·lobranquis, pseudolamel·libranquis i eulamel·libranquis). L’estadi septibranquial que presenten algunes formes carnívores correspon a una transició. Els suspensívors, que són els més nombrosos han pogut explotar amb èxit totes les possibilitats dels hàbitats aquàtics, des de la zona litoral, amb totes les seves variacions quant al medi (tipus de substrat, nivells d’energia, salinitat, etc.), fins a la zona sublitoral i a les grans fondàries on avui viuen alguns protobranquiats.
Els paleotaxodonts

El paleotaxodont Nuculana pella es caracteritza pel rostre posterior truncat i per la presència de carenes umbopal·leals, que s’aprecien a l’exemplar superior de la fotografia. La xarnera és taxodonta i lleugerament corba. Espècie infaunal de sediments fins, és freqüent en el Pliocè del Baix Llobregat. La llargada de la conquilla és de 13 mm.
Jordi Vidal / Col·lecció Jaume de Porta.
La subclasse dels paleotaxodonts comprèn formes amb una xarnera taxodonta, equivalves i isomiàries. Representen el prototipus dels protobranquiats, que s’alimenten de detrits. Tenen representants tant a l’epifauna com a la infauna, i són considerats com a formes primitives. Pojetia runnegari, del Cambrià d’Austràlia, pertany a aquest grup. S’estenen en el temps des del Cambrià fins a l’actualitat. Comprenen un sol ordre, el dels nuculoides (Nuculoida), trobats al nostre país en terrenys del Triàsic, concretament del Ladinià i de l’Anissià, de Camposines i Centelles (Osona), on s’ha trobat Palaeonucula goldfussi; també hi ha citacions de Paleoneilo en el Ladinià d’Espejeras (en terres alacantines). Espècies de Nucula, Nuculana i Leda són freqüents al Terciari del Principat, com al Miocè del Penedès i al Pliocè de Girona, i també al Terciari superior de les Balears.
Els criptodonts

Conquilla fòssil de Cardiola interrupta, de 2 cm de diàmetre, característica del Silurià i freqüent a diverses localitats de Catalunya. (La de la fotografia prové de Riu Major.) Noteu l’ornamentació reticulada i molt marcada per la confluència de còstules o costelles radials i lamel·les horitzontals. Pertany al gran grup de bivalves criptodonts.
Jordi Vidal / MGB.
Constitueixen una agrupació de formes encara poc conegudes, amb conquilla de paret fina, equivalves, dimiaris i amb lligament extern de tipus amfidètic opistodètic. Hom en considera dos ordres: el dels solemioides (Solemyoida), que comprèn les úniques espècies de la subclasse encara vivents (per exemple, el gènere Solemya), bé que es coneixen des del Devonià; i l’ordre dels precardioides (Precardioida), que reuneix formes exclusivament fòssils, encara mal conegudes (algunes, com el gènere Eopteria del Cambrià, són considerades per molts autors com a crustacis). Del Silurià, és coneguda la presència de Cardiola interrupta a Santa Creu d’Olorda (Baix Llobregat), a Camprodon (Ripollès) i a Sant Sebastia de Buseu (Pallars Sobirà), entre altres.
Els pteriomorfs
La subclasse dels pteriomorfs consitueix un grup molt diversificat, per al qual es fa difícil d’establir unes característiques comunes i generals. Són anisomiaris, i en algunes formes especialitzades el múscul anterior és reduït i arriba a perdre’s, com en els pectínids i els ostrèids, entre altres. El lligament és duplivincular en molts casos, però també n’hi ha que el tenen alivincular i parivincular. En general són representants de l’epifauna, bé que també inclouen bivalves perforadors de substrats durs. Les dades paleontològiques indiquen que es tracta d’un grup monofilètic, que s’estén des del Cambrià inferior fins a l’actualitat. Comprèn tres ordres: els arcoides (Arcoida), els mitiloides (Mytiloida) i els pterioides (Pterioida).
Els arcoides

Entre els pteriomorfs hi ha grups d’origen molt antic i alguns d’una àmplia distribució en els temps geològics. Els àrcids, família a la qual pertany Anadara diluvii, que veiem a la fotografia, són especialment abundants en els sediments fins miocens. La conquilla, de talla mitjana (40 × 26 mm), és típica per la seva ornamentació radial de còstules nombroses i separades per solcs més estrets; i també ho és per l’àrea lligamentosa, estreta, amb solcs que assenyalen les posicions que ocupava el lligament en l’exemplar viu (bé que no es pot veure a la fotografia).
Jordi Vidal / MGB.
Aquest ordre inclou, entre altres, les famílies dels àrcids (Arcidae), els noètids (Noetidae) i els glicimèrids (Glycymeridae). Als Països Catalans n’hi ha representants des de l’Anissià de Centelles (Osona), amb Parallelodon, i posteriorment, en el Cretaci inferior s’ha trobat Arca cymodoce a Morella (Ports), al Castell de Cabres (Baix Maestrat) i a Tortosa (Montsià). Són especialment nombrosos els àrcids del Terciari: Barbatia barbata, Scapharca i d’altres, al Miocè del Penedès; Anadara diluvii i A. pectinata, del Pliocè de l’Empordà i el Baix Llobregat, i també a Alacant. Els glicimèrids tenen un marcat caràcter conservador, amb poca variació morfològica de la conquilla, i s’han adaptat a medis inestables.
Les primeres formes reconegudes al nostre país daten del Cretaci, amb Glycymeris buchi de Carbonils (Alt Empordà). En el Terciari són coneguts de l’Eocè de la vall d’Àger (Noguera) i encara molt més freqüents en el Miocè i el Pliocè; s’han reconegut també a diferents indrets del Miocè de la Depressió Prelitoral Catalana.
Els mitiloides
Són formes fortament inequilàteres, que secundàriament poden viure soterrades, com en el cas d’Atrina, que ha estat descrita en el Pliocè del cementiri de Siurana d’Empordà (Alt Empordà) i en el Pliocè de la vall del Llobregat. Modiolus és conegut del Ladinià de València, en el Liàsic d’Alfara de Carles (Baix Ebre), com M. moralejai i altres espècies es coneixen del Terciari a moltes localitats. Entre les formes perforadores, les citacions històriques de diverses espècies de Litophaga (=Lithodomus) del Cretaci inferior de Garraf no han estat confirmades; en canvi, són freqüents a tot el Terciari i el Quaternari. Els mitiloides comprenen també el gènere Fordilla, del Cambrià.
Els pteroides
Són, dins la subclasse dels pteriomorfs, el grup més important, tant per la quantitat de formes com per la seva diversificació. Moltes espècies tenen importància biostratigràfica; Bakevellia costata i les dels gèneres Gervillia, Hornesia, Cassianella, Leptochondria i Posidonia, en el Triàsic del Països Catalans.

Els pterioides de la família dels inoceràmids són molt ben representats en el Cretaci i hi tenen valor biostratigràfic. L’exemplar de la fotografia, d’Inoceramus (Platyceramus) cycloideasahsenensis, prové del Cretaci superior de Montesquiu (Pallars Jussà). Noteu la conquilla allargada (19 mm) en sentit umbopal·leal i amb l’ornamentació concèntrica ben marcada i irregular.
Jordi Vidal / UAB.
La família dels inoceràmids (Inoceramidae) formen un grup extingit i ben representat en el Cretaci, on tenen una forta incidència biostratigràfica. Als Països Catalans s’han descrit espècies d’Inoceramus, atribuïdes abans a altres gèneres, totes del Cretaci superior.
Dels pectinacis (Pectinacea) són importants en el Triàsic, concretament en el Ladinià, Posidonia i Daonella lommeli. La família dels pectínids (Pectinidae) és la més ben representada, en nombre d’espècies i de localitats. Els pectínids tenen unes característiques morfològiques peculiars: la conquilla és de contorn circular o oval, i té unes expansions laterals a cada costat de l’umbe que formen les orelletes; en general, el marge anterior presenta un solc pel qual surt el bissus i on es pot observar un conjunt de petites dents (les dents pectinidials) que serveixen per a conduir els filaments del bissus. Tot el conjunt rep el nom d’ectenòlium. La línia de la xarnera és recta i el lligament es fixa a diverses fossetes, de les quals la que es troba sota l’umbe s’anomena fosseta del condròfor. A cada costat del resilífer hi ha unes làmines anomenades crures cardinals. De vegades hi ha, a més, unes petites protuberàncies a la zona on les orelletes s’ajunten a la valva que reben el nom de crures auriculars. L’única impressió muscular és gran i ocupa una posició central o subcentral. La superfície de la conquilla, en general, mostra una ornamentació a base de costelles de característiques variades que tenen significació taxonòmica per a separar les espècies. Les formes sedimentàries jeuen sobre la valva dreta, que és bombada, mentre que l’esquerra ho és poc o gens.

Dos pectinacis del nostre registre fòssil. A dalt, Daonella lommeli, força freqüent en el Triàsic dels Països Catalans. L’exemplar és d’Alfara de Carles (Baix Ebre) i té un diàmetre de 38 mm. Noteu el marge cardinal rectilini i l’ornamentació, a base de grups de costelletes molt fines separades clarament. A baix, valva dreta de Neithea (47 mm), de Morella, subtriangular i molt convexa, amb 4 o 6 costelles, separades per espais amples, amb costelles molt més fines.
Jordi Vidal / MGB.
Les primeres troballes de pectínids corresponen al Triàsic dels Catalànids, amb el gènere Chlamys. Weila lacazei apareix a diversos punts del Liàsic tarragoní. Al Cretaci, el nombre d’espècies trobades és més alt; de l’Aptià; s’ha trobat Chlamys echaetes a Morella (Ports) i a punta de l’Àliga (Baix Ebre), i Entolium orbiculare a Alcalà de Xivert (Baix Maestrat). Del Santonià, Camptonectes virgatus, s’ha trobat a les collades de Basturs (Pallars Jussà), Chlamys espaillaci al Montsec i a Fígols, i Mimachlamys cretosa, també al Montsec. Del Campanià-Maastrichtià, Merklinia catalaunica, a Fígols (Alt Urgell), a Berga (Berguedà) i a Salàs del Pallars (Pallars Jussà). Durant l’Eocè, els pectínids són representats per algunes formes, però no hi ha determinacions segures. Al Miocè i al Pliocè hi ha nombrosíssims exemplars en moltes localitats del Penedès i del Tarragonès, i també, en el cas del Pliocè, a l’Empordà. La llista d’espècies trobades és quantiosa, de manera que només n’indicarem les més importants; del Miocè: Chlamys (Macrochlamys) latissima nodosiformis, C. (Aequipecten) seniensis, C. (A.) jackloveciana, C. (Macrochlamys) tournali, Pecten (Hopenheimopecten) joslingi i P. (Flabellipecten) costisulcatus; del Pliocè: Amussium cristatum, Chlamys (Macrochlamys) latissima, C. (Aequipecten) seniensis, C. (Manupecten pesfelis, C. (Chlamys) multistriata, Palliolum (Lissochlamys) excisum i Pecten (Pecten) benedictus.

Exemplar del pectínid Chlamys latissima, de closca gruixuda i grossa (fa 18 cm), procedent del Miocè mitjà d’Altafulla (Tarragonès), però abundant en general al Miocè i al Pliocè en ambients de mars somes. És característic perquè té les còstules principals molt desenvolupades i poc nombroses.
Jordi Vidal / Col·lecció Jaume de Portal.
Del Quaternari fred (Würm) s’han extret dels fons marins, davant de les costes de Begur (Baix Empordà) diversos exemplars de Chlamys (Chlamys) islandica, i també se n’han trobat a l’alçada de Vilanova i la Geltrú (Garraf); aquestes formes també es coneixien al cap de Creus. Del grup de Neithea, a més d’una citació dubtosa atribuïda al Juràssic tarragoní, concretament del Liàsic de mas Ramé, s’han trobat en el Cretaci dels Pirineus N. regularis, al Santonià de Santa Engràcia (Pallars Jussà); i, al Montsec, N. striatocostata al Santonià-Maastrichtià del Flamicell-Pallaresa, Erinyà (Pallars Jussà) i Fígols (Berguedà), N. syriaca a l’Aptià de Costa de Reinals (Montsià) i Morella (Ports), Mas de Barberans (Montsià), a la punta de l’Àliga (Baix Ebre), a Marmellà (Tarragonès) i al Garraf.
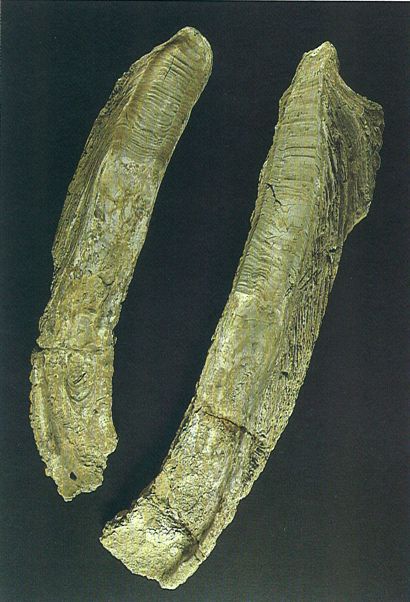
L’ostrèid Crassostrea gryphoides, de conquilla gruixuda i enorme (alguns exemplars poden atènyer els 60 cm de llargada), pot presentar una certa variabilitat en la forma general segons les condicions del medi. Abundant en els medis marins d’aigües somes, i en general a tot el Miocè català, pot formar bancs d’una gran extensió. A l’exemplar de la fotografia, procedent del Miocè mitjà del Penedès, s’observen les bandes de creixement en l’àrea lligamentosa de totes dues valves.
Jordi Vidal / Col·lecció Carles Martín.
Els ostrèids (Ostreidae), una família del subordre dels ostreïnis (Ostreina), no tenen xarnera quan són adults, de manera que el lligament és l’única estructura que permet l’obertura i el tancament de les valves. L’àrea lligamentària o taló consta d’una part central (resilífer) i de dues àrees laterals (els «bourrelets»); en l’àrea es poden observar unes bandes que corresponen als dipòsits estacionals del creixement i que s’han emprat per a determinar l’edat aproximada dels exemplars. A la vora de l’àrea lligamentària, algunes formes presenten unes estructures anomenades comes («chomata»), que a la valva dreta formen petits tubercles (anacomes) i a l’esquerra, els seus homòlegs, prenen forma de fossetes (catacomes); aquestes estructures es poden estendre fins al marge pal·leal. Sota l’àrea lligamentària hi pot haver una cavitat umbonal, com a Saccostrea. La impressió muscular, que és única, és de forma molt variable i té importància en la sistemàtica. L’estructura de la conquilla és complexa, i destaquen els elements vesiculars típics de la subfamília dels neopicnodontins (Neopycnodontinae). La larva dels ostrèids es fixa damunt d’un substrat dur, i pot quedar-s’hi fixa fins que mor l’adult. Si el substrat és una partícula petita, quan la conquilla creix hom la considera pràcticament lliure, però normalment sol quedar ben fixada; això produeix deformacions i casos de xenomorfisme. Per a la identificació de les dues valves hom utilitza criteris específics: la valva esquerra sempre és la fixa (o té el senyal deixat per la fixació), i és més convexa i, en general, més gruixuda, a part que és la que porta la cavitat umbonal i les catacomes, quan n’hi ha. Algunes formes toleren les variacions de salinitat i s’adapten als medis estuarins. Poden formar masses importants, com ara esculls, d’extensió i forma variables. La formació de plecs o plicatura en el marge de la comissura de les valves s’ha interpretat com una adaptació de defensa. Pel que fa a les seves dimensions, algunes formes poden arribar fins als 600 mm d’altura. Els primers ostrèids trobats als Països Catalans corresponen a l’espècie Enantiostreon difforme, de l’Anisià-Ladinià dels Catalànids i del País Valencià; a l’Aptià tarragoní i castellonès s’ha trobat Aetostreon couloni, A. latissimum, Gryphaeostraea canaliculata i Ilymatogyra pellicoi. Del Coniacià de les collades de Basturs i del Campanià-Maastrichtià de Tremp (Pallars Jussà), Pycnodonta vesicularis. En el Terciari, a l’Eocè del Principat són relativament freqüents Ostrea uncifefera i Cubitostrea multicostata, i aquesta darrera forma bancs importants a diferents localitats, especialment a la vall d’Àger (Noguera). En el Terciari superior, especialment en el Miocè, hi ha les espècies de dimensions més grans. Als Països Catalans són abundants i es troben reunides un gran nombre d’espècies a cada localitat; destaquen Ostraea lamellosa, O. digitalina i Crassostrea gryphoides.
Els paleoheterodonts

Els paleoheterodonts del grup dels trigonioides tenen una forma trigonal molt típica, bé que els caràcters que defineixen el grup afecten altres característiques, com el tipus de xarnera esquizodonta i l’umbe opistògir, és a dir, inclinat cap a la part posterior entre d’altres. És característica la presència d’una carena marginal prominent, amb un costat ornamentat diferencialment. Les fotografies mostren una vista posterior (a dalt) i lateral (a baix) d’una espècie de Trigonia de 75 mm, procedent del Cretaci inferior de Rodonyà (Alt Camp).
Jordi Vidal / MGB.
Els paleoheterodonts són bivalves de conquilles iguals (equivalves), amb la xarnera formada per poques dents divergents en l’umbe. El lligament és amfidètic o bé opistodètic. Fins fa poc (1978) hom considerava que aquest grup contenia els bivalves més antics. L’atribució de Lamellodonta simplex als braquiòpodes fa que la dispersió de la classe hagi quedat força més restringida i també obliga a eliminar l’antiga família dels lamel·lodòntids (Lamellodontidae).
L’ordre dels modiomorfoides (Modiomorphoida) conté, però, dos dels gèneres més antics: Redonia i Actinodonta, tots dos de l’Ordovicià mitjà de França i de Bohèmia.
L’ordre dels unionoides (Unionoida) comprèn bivalves no marins del Devonià al Permià, amb gèneres importants, com Carbonicola i Anthracosia, del Carbonífer d’Astúries. Dins del mateix ordre hi ha altres formes d’aigua dolça, que presenten una capa nacrada molt característica i que s’han trobat al Ladinià dels Catalànids i del País Valencià; n’és un exemple Unionites munsteri. Del Maastrichtià dels Prepirineus s’han trobat algunes formes relacionades amb fàcies garumnianes. És un grup molt complex, en el qual les condicions ecològiques tenen una forta influència sobre la morfologia de la conquilla. Destaca la presència d’Unio garumnica a Vallcebre (Berguedà).
Els representants de l’ordre dels trigonioides (Trigonioida) tenen la conquilla equivalva, triangular i amb el costat posterior truncat. La xarnera de la valva esquerra presenta una dent mitjana, forta, i la de la valva dreta, dues. Les impressions musculars se situen prop de la xarnera. De la família dels miofòrids (Myophoriidae), coneixem algunes formes de l’Anisià-Ladinià als Països Catalans: Myophoria vulgaris, Costataria goldfussi i Neoschizodus laevigatus; al Cretaci esdevenen més importants. De l’Aptià de Morella (Ports) provenen espècies atribuïdes al gènere Trigonia. De Masbonés (Tarragonès) i altres localitats tarragonines s’han citat Pterotrigonia caudata i Linotrigonia (Oistotrigonia) ornata.
Els heterodonts

Valva esquerra de l’heterodont Tellina planata, un tel·línid procedent del Miocè de Montjuïc (Barcelonès), bé que l’espècie encara és actual. La closca, de dimensions mitjanes (6,5 cm), és típica per la carena umboposterior i per la fina ornamentació a base de cordons, més marcada en el rostre posterior.
Jordi Vidal / MGB.
Les característiques comunes, vàlides per a la majoria dels representants d’aquesta subclasse són les següents: la xarnera heterodonta, amb una clara separació entre dents cardinals i dents laterals; el lligament opistodètic, la lúnula i l’escut ben diferenciats; i l’estructura de la conquilla lamel·lar creuada, complexa o prismàtica. Constitueixen aquest grup els ordres dels veneroides (Veneroida), els mioides (Myoida) i els hipuritoides (Hippuritoida), dels quals només els dos primers tenen representants actuals.
Els veneroides
Els veneroides, de tots tres, són els més nombrosos i diversificats. S’han adaptat a una gran varietat d’hàbitats, i n’hi ha de detritívors, com els de la família dels tel·línids (Tellinidae), i de suspensívors, que són la majoria. Hi ha formes tan epifaunals com infaunals i l’ordre inclou una de les formes més antigues, el gènere Babinka, de l’Ordovicià mitjà. Als Països Catalans se’n coneixen del Triàsic i el Juràssic, i han esdevingut cada vegada més nombrosos. Hi ha una llarga llista d’espècies citades al nostre país. Un grup que mereix referència a part són els dreissènids (Dreissenidae) i els limnocàrdids (Limnocardidae) del Pliocè del Baix Llobregat, que indiquen unes condicions d’aigües salabroses.
Els mioides
L’ordre dels mioides reuneix formes bàsicament excavadores, de closca fina, que tenen les valves equivalves o no, però sempre són molt inequilàteres. La xarnera té una sola dent a cada valva. Inclou la majoria de les formes perforadores, que pertanyen a les famílies dels gastroquènids (Gastrochaenidae), hiatèl·lids (Hiatellidae) i folàdids (Pholadidae). Són especialment abundants en el Terciari, i en general es troben associades a medis de molta energia. La presència de forats produïts per aquestes formes en els penya-segats s’ha emprat com a mètode per a assenyalar les variacions del nivell del mar durant el Quaternari. Els representants d’aquest ordre s’estenen des del Carbonífer fins a l’actualitat. Indiquem la presència de Jouannetia, un folàdid (Pholadidae) en els esculls miocens del Papiol (Baix Llobregat) i de can Sala (Penedès).
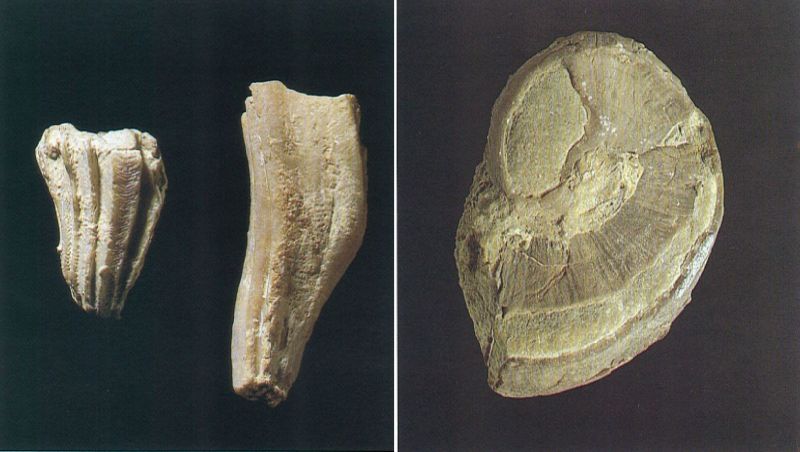
Els hipuritoides o rudistes

Rudista característic del Maastrichtià dels Pirineus catalans, Hippurites castroi, tot sencer, amb les dues valves juntes. La inferior, molt més gran que la superior, és la que es fixava al substrat; noteu-hi les còstules irregulars i les línies de creixement, ben marcades. A la valva superior, que fa com una mena d’opercle, s’hi poden veure els òsculs, situats marginalment. L’exemplar fotografiat, d’una altura de 82 mm, procedeix del barranc de la Posa (Pallars Jussà).
Jordi Vidal / Col·lecció Jaume de Porta.
Els hipuritoides inclouen les superfamílies dels megalodontacis (Megalodontaceae) i dels hipuritoidacis (Hippuritoidacea), que r.eben en general el nom de rudistes. Formen un conjunt d’organismes completament extingits a la fi del Cretaci, que presenten una condició inequivalva molt accentuada. La conquilla, de parets espesses, té una xarnera de tipus heterodont, amb poques dents, amorfes i gruixudes; en realitat, es tracta d’heterodonts aberrants, que vivien fixos al substrat per una de les valves. Els primers representants del grup són equivalves, excepte Megalodon, i desapareixen en el Cretaci inferior; la presència d’aquest gènere en el Triàsic dels Països Catalans és dubtosa. Els rudistes de la família dels hipuritacis (Hippuritacea) tenen les valves molt diferents. La valva fixa en general és la més gran i és enrotllada; la lliure o superior és plana, però també pot ser enrotllada i aleshores esdevé la més gran. El caràcter aberrant que presenten els membres d’aquest grup fa que sigui difícil d’aplicar el concepte de valva dreta i valva esquerra com a la resta dels bivalves. Alguns autors antics han interpretat les valves dels rudistes d’una manera paral·lela a allò que passa al gènere actual Chama, és a dir, aplicant el procés conegut com a inversió de la xarnera. Així, hi hauria unes formes dites normals i unes altres dites inverses. D’altres autors rebutgen aquesta proposta i, per tal d’evitar confusions, és aconsellable parlar només de valva fixa i valva lliure. La xarnera consta de dents (dues a la valva lliura i una a la fixa) i de fossetes. La dent de la valva fixa s’allotja en una fosseta que queda entre les dues dents de la valva lliure. El lligament podria estar situat tant a la part externa com a la interna de la conquilla, i devia fixar-se damunt d’una fosseta o d’una aresta en el primer cas, o en una estructura en forma de solc en el segon. El sistema muscular devia correspondre a dos músculs adductors fixats a la paret de la conquilla. La superfície d’inserció d’aquests músculs és molt variada i fins i tot canvia d’una valva a l’altra. A grans trets, podem indicar tres tipus de superfícies: plaques miofòriques, disposades perpendicularment al pla d’unió de les dues valves; apòfisis amb una forta projecció; i una estructura semblant a un esperó o un contrafort. Alguns representants presenten altres estructures que en alguns casos són típics de les famílies. Les cavitats accessòries són estructures que estan connectades amb les dents i les fossetes, especialment en els caprotínids (Caprotinidae), però també en la valva fixa dels caprínids (Caprinidae), dels hipurítids (Hippuritidae) i dels radiolítids (Radiolitidae), en els quals s’allotgen a la regió cardinal. Les estructures sifonals són formades per bandes, solcs i pilars, i estan associades en els hipurítids. Han estat interpretades de maneres diferents: com a estructures per a la fixació de músculs, zones sifonals i també com a suports d’òrgans sensorials. La presència de porus i òsculs a la valva lliure, situats per sobre dels pilars, devia relacionar-se amb la generació de corrents d’aigua. La paret de la closca presenta en els radiolítids una estructura cel·lular prismàtica, força típica, i en els caprínids hi ha canals marginals complexos.
Dins del grup dels rudistes o hipuntoides podem trobar una certa diversitat de morfologies. A dalt veiem un exemplar de 20 mm d’alçària de Birradiolites angulossissimus, de forma molt cònica, amb costelles longitudinals força desenvolupades i amb bandes que corren en el mateix sentit i que es diferencien de les costelles i dels solcs; i, al costat, un exemplar d’Agriopleura, d’uns 30 mm d’alçària, amb lleugeres estríes transversals i amb una zona plana entre dues bandes. A baix, valva inferior de Toucasia, amb un enrotllament en espiral bastant plana (la valva superior és de tipus opercular, també espiral·lada, però força més petita); és una forma freqüent en el Cretaci inferior.
Josep M. Moraleja / MGSB i Jordi Vidal / Col·lecció Jaume de Portal.
En el Juràssic superior (Oxfordià i Kimmeridgià) apareixen per primera vegada Diceras i Heterodiceras. Als Països Catalans són moltes les espècies identificades i també les localitats on apareixen són nombroses. A l’Aptià del massís de Garraf s’ha reconegut Toucasia, Pseudotoucasia santanderina, Matheronia, Horiopleura i d’altres. En el Maestrat, Matheronia oligostoja apareix en el Berriasià, i Toucasia i Matheronia ho fan en el Barremià, mentre que a l’Aptià hi ha Pachytraga i Offneria. Als Prepirineus, són nombroses les localitats que han fornit mostres de les principals famílies de rudistes: de requiènids (Requieniidae), com Apricardia; de monoplèurids (Monopleuridae), com Monopleura; de caprínids, com Mitrocaprina i Plagioptychus; d’hipurítids, com Hippurites, que hi és present amb diversos subgèneres; de radiolítids, com Radiolites, Sphaerulites, Rosellia, Sauvagesia, Agriopleura i Lapeirouseia, entre altres. S’han descrit moltes d’aquestes formes al nostre país, i també s’ha fet una biozonació del Cretaci superior dels Prepirineus.
Els anomalodesmats

Entre els anomalodesmats hi ha espècies carnívores, com Cuspidaria cuspidata, que veiem a la fotografia, característica per l’allargament del rostre de la part posterior de la conquilla. L’exemplar fou trobat al Pliocè del cementiri de Siurana d’Empordà (Alt Empordà), però l’espècie es troba també, encara que no tan abundant, en el Miocè.
Jordi Vidal / Col·lecció Jordi Martinell.
Són bivalves de conquilla equivalva que tenen la xarnera amb el marge molt engruixit (o bé enrotllat) i sense dents o només amb una de sola i, encara, poc marcada. Alguns gèneres són septibranquis. Entre ells hi ha els únics bivalves que són carnívors. La família més ben representada és la dels foladòmids (Pholadomiidae). Bé que el grup en conjunt apareix en el Paleozoic, els primers representants que trobem als Països Catalans són triàsics (de l’Anisià i el Ladinià), com ara Pleuromya elongata. Són abundants en el Juràssic i el Cretaci i s’hi han trobat diverses espècies, encara, però, de determinació insegura. Cuspidaria, un representant de la família dels cuspidàrids (Cuspidariidae), és conegut del Pliocè del Principat.
Paleoecologia dels bivalves

Visió lateral (a dalt) i posterior (a baix) d’una conquilla de Glycymeris de mida mitjana (11,6 cm), procedent del Miocè de Montjuïc. En general, els glicimèrids són formes globoses i de contorn subcircular, a les quals el lligament es fixa en una àrea subtriangular situada sota l’umbe.
Jordi Vidal / MGB.
En els bivalves, la morfologia de la conquilla i l’hàbitat estan molt relacionats, com passa en altres grups d’invertebrats i per això els estudis de morfologia funcional tenen un gran interès per a la paleoecologia; l’adaptació a diversos tipus de vida i a les condicions del substrat ha produït una gran diversificació en aquest grup. Pel que fa a l’hàbitat, hom els agrupa en tres grans categories: l’epifauna, la semiinfauna i la infauna.
L’epifauna és formada pel grup que ha aconseguit més facilitat d’adaptació, i cadascuna de les seves adaptacions s’ha traduït en una morfologia. Dins dels mol·luscs epifaunals, podem distingir entre les formes que viuen fixes al substrat per un bissus, les que s’hi cimenten i les formes lliures. Les que són fixades per un bissus tenen un ampli ventall de possibilitats pel que fa a l’elecció de substrat: viuen tant sobre substrats orgànics (algues, branques de coralls, etc.) com inorgànics. En general tenen la conquilla inequivalva i poc convexa, ben adaptada als corrents. El gènere Pteria és un representant molt típic d’aquest tipus. D’altres formes viuen en petites depressions de les roques, com fa Arca, o en les fissures, com alguns Chlamys. En les formes que viuen fixes per cimentació, aquesta cimentació té lloc per mitjà de dipòsits calcaris. La fixació pot ser temporal o permanent, com en el grup anterior, i moltes vegades estan exposades a condicions d’alta energia. La valva fixa presenta una deformació en relació a les irregularitats del substrat, i per tant, les conquilles són fortament inequivalves, inequilàteres i moltes vegades de parets gruixudes. En les formes lliures, la conquilla presenta un contorn més aviat arrodonit i una xarnera curta. El perfil circular permet una àmplia extensió dels òrgans dels sentits per fora del marge del mantell; algunes espècies, com les de Glycymeris, poden rodolar per damunt del substrat gràcies a la seva convexitat, i fins i tot poden soterrar-se lleugerament. Entre els pectínids hi ha una gran diversitat; alguns tenen les valves fortament asimètriques: reposen sobre la valva dreta, que és convexa i quasi sempre costellada, mentre que l’esquerra és molt plana, i es desplacen obrint i tancant les valves amb moviments enèrgics; d’altres són equivalves, de conquilla llisa i fina, i molt poc convexes, com Amussium i Propeamussium, que són nedadors actius.
La semiinfauna és representada per les formes que es desplacen poc o molt. Viuen amb una part de la conquilla soterrada, però és el costat posterior el que queda fora del substrat. En general són de conquilla allargada i de paret fina, i Pinna és el tipus més representatiu.
El grup infaunal comprèn les formes que passen la major part de la seva vida soterrades en un substrat. Per la manera d’enterrar-se, hom separa les formes adaptades a substrats durs de les que ho fan en substrats tous. Les conquilles llises o amb una ornamentació a base de solcs divergents o divaricada s’enterren més ràpidament; les espines, en canvi, actuen com un element d’estabilització. Els bivalves que foraden substrats durs poden instal·lar-se al damunt de roques, de branques de coralls, d’altres organismes i, fins i tot, de fusta. S’enterren per una combinació de desgast mecànic i de dissolució química del substrat. En general tenen la conquilla allargada; Lithophaga, Pholas, Botula i Hiatella són exemples actuals i fòssils. També hi ha formes de conquilla globosa, com Jouannetia, del qual coneixem restes fòssils en els esculls del Miocè del Penedès. Quan resta solament el forat on va ser l’animal, l’obtenció de motlles permet reconèixer algunes espècies ja que, fins a un cert punt, el forat reprodueix la forma del bivalve.