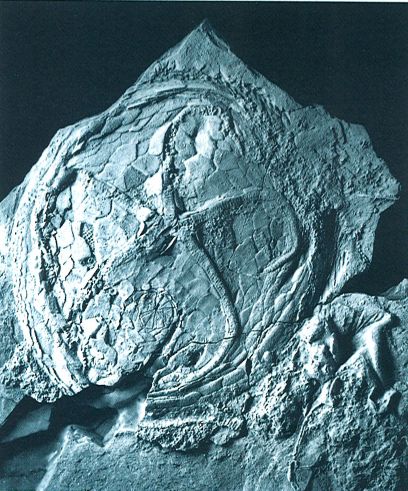
Els edrioasteroïdeus formen l’única classe totalment fòssil dels eleuterozous. Suspensívors de vida sèssil, tenien els solcs ambulacrals protegits per plaques, un periprocte format i una estructura bucal definida. L’exemplar de Krama devonica que veiem a la fotografia, del Devonià de Luanco, Astúries, és l’únic trobat fins ara a la península Ibèrica. El disc fa uns 19 cm de diàmetre.
Andrew Smith (Lethaia 20/1 Oslo) / Dept. Geol., Oviedo.
Els eleuterozous són equinoderms de teca globular, discoïdal o estel·lada, amb la superfície dorsal igual o més petita que la ventral, i gairebé mai prolongats en una tija. En aquests equinoderms, els ambulacres formen part integrant de la teca, i mai no es prolonguen lliurement.
De les cinc classes que inclouen els eleuterozous (Asteroidea, Ophiuroidea, Holothuroidea, Echinoidea i Edrioasteroidea), només els equinoïdeus tenen una importància real per als paleontòlegs, i només una, la dels edrioasteroïdeus és totalment fòssil, bé que no s’ha trobat representada, fins ara, als Països Catalans. Els edrioasteroïdeus són eleuterozous amb els solcs ambulacrals protegits per sèries de plaques recobridores, amb una estructura bucal fixa formada per les plaques ambulacrals inferiors més adorals, i amb un periprocte que s’obre ventralment en l’interambulacre posterior. Pel que sembla, vivien sempre fixos al substrat per mitjà de la seva superfície dorsal. El grup es compon d’un petit nombre de formes. Entre les més antigues, de vida lliure, es troben probablement els avantpassats d’altres grups d’eleuterozous. Eren suspensívors d’ambients marins soms, amb la superfície oral orientada cap amunt, en direcció oposada al fons marí. S’han trobat en materials del Cambrià inferior al Carbonífer superior. Dels quatre ordres que hom en reconeix, només el dels isoròfides (Isorophida), que visqueren de l’Ordovicià mitjà fins al Carbonífer superior, han estat reconeguts a la península Ibèrica: un exemplar de Krama devonica, descobert en l’Emsià superior (Devonià) dels voltants de Luanco (Astúries).
Els asteroïdeus
Els asteroïdeus són eleuterozous encara actuals, però apareguts a l’Ordovicià inferior, que tenen forma estel·lar, amb les superfícies dorsal i ventral aproximadament iguals i ambulacres biseriats; els braços presenten una àmplia cavitat corporal ocupada per les ramificacions dels òrgans principals, sense que hi hagi cap diferenciació important amb el disc central. Els canals hidràulics radials són a l’exterior de les plaques ambulacrals. L’anus s’obre dorsalment i l’estructura bucal no és fixa.
La diversitat dels asteroïdeus és molt gran i, bé que en els grups postpaleozoics hom ha establert una ordenació sistemàtica ben definida, no podem dir el mateix dels més antics, paleozoics. Les troballes d’exemplars complets són escasses, com sol passar en gairebé tots els altres grups d’equinoderms (tret dels equinoïdeus), i ara com ara només s’han arribat a reconèixer Lophidiaster faurai, de l’ordre dels paxil·lòsides (Paxillosida), trobats a l’Aptià d’Albinyana (Baix Penedès) i altres exemplars de l’ordre dels valvàtides (Valvatida), dels gèneres Metopaster, Crateraster i Pycinaster, apareguts en el Campanià superior-Maastrichtià de la conca de Tremp, Calliderma atagensis de l’Eocè de Gurb de la Plana (Osona) i del Malbalç (Bages) i Teichaster poritoides, de l’Eocè superior de Castellterçol (Vallès Oriental). Recentment s’han produït noves troballes d’asteroïdeus (com també d’ofiuroïdeus), que ben aviat ampliaran els coneixements actuals sobre les formes fòssils d’aquestes classes d’equinoderms.
Els ofiuroïdeus

La marcada diferenciació entre el disc central i els braços dels ofiuroïdeus és ben patent en aquesta vista dorsal d’Aspidura montserratensis (diàmetre del disc de menys d’1 mm), un petit ofiúride recentment descobert en el Muschelkalk superior dels voltants de Montserrat. Al contrari del que passa en els asteroïdeus, l’interior dels braços dels ofiuroïdeus no és recorregut per extensions del sistema digestiu.
Josep M. Moraleja / Col·lecció Daniel Gutiérrez i Assumpció Rodríguez.
Apareguts igualment a l’Ordovicià inferior, els ofiuroïdeus tenen forma estel·lada; els braços no són recorreguts interiorment per extensions del sistema digestiu i externament són ben diferenciats del disc central del cos. No tenen obertura anal i els canals radials són interns. El seu aparell mandibular és format per ossets ambulacrals modificats, que se situen prop de la boca.
Dels quatre ordres que hom reconeix entre els ofiuroïdeus, únicament s’ha trobat als Països Catalans un ofiúride (Ophiurida), Geocoma magna, que aparegué en el Toarcià (Liàsic) d’Alfara de Carles (Baix Ebre), i altres exemplars d’ofiuroïdeus, indeterminats fins ara, del Toarcià de Camarasa (Mont-roig, a la Noguera). En canvi, en sediments maastrichtians del port d’Olazagutia (Navarra), de fàcies i edats semblants a les dels nostres Prepirineus, s’han trobat Amphiura senonensis, Ophiura hagenowi i O. serrata, per bé que el fet d’haver-ne trobat només plaques soltes dels braços fa que no es pugui donar la seva determinació genèrica per absolutament certa. Hom creu que una recerca més acurada d’aquestes restes, que sovint són de dimensions petites, ha de donar resultats positius.
Els holoturoïdeus

Els holoturoïdeus fòssils no conserven complet el seu esquelet fora d’alguns casos molt excepcionals, com ara el de l’exemplar de la fotografia d’Oneirophantites tarraconensis, d’uns 15 cm de longitud, que procedeix del Muschelkalk superior de Mont-ral i que pertany a l’ordre dels elasipòdides. Noteu el cos simètric bilateralment, amb peus ambulacrals dorsals transformats en processos sensitius allargats.
Josep M. Moraleja / MGSB.
Els holoturoïdeus són equinoderms encara actuals (aparegueren al Silurià), de superfície dorsal fortament reduïda amb relació a la ventral. L’anus s’obre en la superfície dorsal i la boca és envoltada, en la majoria dels casos, per llargs peus ambulacrals tentaculars. Els ossets ambulacrals més adorals s’han modificat formant un anell intern calcari (corona perifaríngia). L’esquelet de la paret corporal és generalment reduït a unes espícules microscòpiques, anomenades esclerits; el canal hidràulic radial és completament intern. Actualment constitueixen un grup força divers i ben representat, adaptat a la nutrició suspensivora o detritívora. Viuen a totes les fondàries, per bé que són més abundants en aigües càlides i somes. Sembla que cal cercar els seus antecessors entre els ofiocistioïdeus (Ophiocistioidea), un grup molt peculiar del Paleozoic inferior, que tenia un esquelet a base de plaques i proveït d’aparell mandibular del tipus equinoide. La classificació paleontològica dels holoturoïdeus es fonamenta en un doble sistema: d’una banda, hom aplica una nomenclatura binomial, sense implicació jeràrquica subjecta al Codi Internacional de Nomenclatura Zoològica (paratàxons) per a les esclerites aïllades; de l’altra, s’empra un sistema linneà, binomial, per als exemplars complets.
Bé que la majoria dels equinoderms són ben coneguts gràcies a la preservació relativament fàcil dels seus exosquelets calcaris, els holoturoïdeus són pobrament representats en el registre fòssil. Fins fa ben poc (1986) se’n coneixien només tres espècies fòssils. Però, en canvi, fa aproximadament un segle que els micropaleontòlegs reconeixen i estudien els esclerits procedents de la disgregació d’aquests organismes en els sediments marins; s’han estudiat en el Carbonífer, el Triàsic, el Juràssic i el Miocè, si bé se’ls coneix des del Silurià i potser ja existien a l’Ordovicià. Als Països Catalans hom ha reconegut dues espècies d’holoturoïdeus, pel que fa a exemplars complets. Ambdues han aparegut en les calcàries del Muschelkalk de Mont-ral (Alt Camp), i pertanyen a la subclasse dels aspidoquirotacis (Aspidochirotacea), que reuneix els holoturoïdeus que tenen tentacles acabats en un disc aproximadament circular: Bathysynactites viai, de l’ordre dels aspidoquiròtides (Aspidochirotida), que tenen un cos simètric bilateralment i els peus ambulacrals dorsals transformats en papil·les; i Oneirophantites tarraconensis, de l’ordre dels elasipòdides (Elasipodida), que tenen el cos bilateralment simètric, amb els peus ambulacrals dorsals transformats en processos sensitius allargats.
Els equinoïdeus
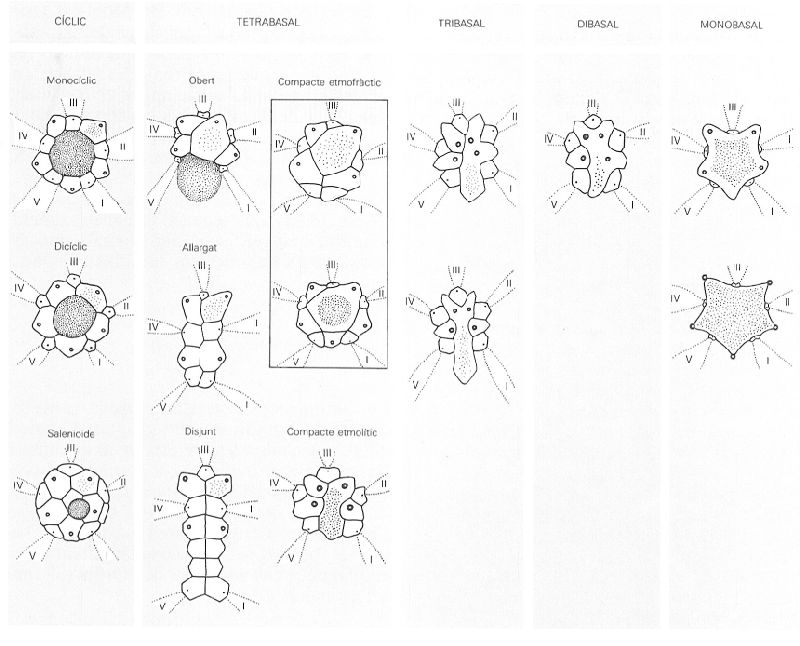
Tipus de sistemes apicals dels equinoïdeus. Entre els cíclics (5 plaques genitals), les plaques oculars poden estar en contacte amb el periprocte (monocíclic, com Diadema) o no (dicíclic, com Pedina); el tipus salenioide (Salenia) presenta plaques complementàries i el periprocte no centrat. Els sistemes tetrabasals (4 plaques genitals) presenten el periprocte desplaçat del sistema apical, que pot estar en contacte amb aquest (obert, com a Plesiechlnus) o pot haver emigrat totalment. En els compactes etmofràctics (Conulus, a dalt, i Coenholectypus, a baix), el madreporit no arriba a separar les plaques oculars I i V, mentre que en l’etmolític (Spatangus) si que es produeix la separació d’aquestes, tot i que el sistema apical manté una certa cohesió. En el sistema allargat (Echínocorys), la disposició de les plaques és allargada però contínua, mentre que en el disjunt (Dysaster) entre les plaques oculars I i V (bívium) i les plaques II, III i IV (trívium), s’hi intercalen altres plaques coronals no pertanyents al sistema apical. En els sistemes tribasals, el madreporit reuneix dues plaques genitals (Brissopsis) i en els dibasals (Brissopsis), tres. Els sistemes monobasals presenten les plaques genitals (4 en Echinolampas i 5 en Clypeaster) fusionades en una de sola, que forma el madreporit.
Amadeu Blasco, a partir de fonts diverses.
Els equinoïdeus són eleuterozous proveïts d’un esquelet format per plaques calcàries interconnectades; les corresponents als ambulacres són perforades per a permetre el pas dels peus ambulacrals. Tenen certes modificacions dels ossets ambulacrals més adorals: s’han convertit en un aparell mastegador, bé que el poden perdre secundàriament. Els equinoïdeus han viscut des de l’Ordovicià fins a l’actualitat.
Segons la hipòtesi més recent, fonamentada en el reconeixement d’estructures homòlogues entre els equinoderms i en la identificació dels elements morfològics que resultaren evolutivament innovadors per mitjà d’una metodologia cladística per a definir grups monofilètics, hom creu que els equinoïdeus són una de les classes d’equinoderms apareguda més recentment: únicament s’avançaren a l’aparició dels holoturoïdeus. No hi ha consens pel que fa al grup origen dels equinoïdeus, ja que hi ha diverses teories; per exemple, hom ha proposat com a grup ancestral dels equinoïdeus els helicoplacoïdeus, els edrioasteroïdeus, els asteroïdeus, els ofiuroïdeus, els holoturoïdeus, els cistoïdeus diploporits i, fins i tot, els blastoïdeus. De fet, els equinoïdeus representen un grup ben característic, però amb similituds òbvies amb les altres classes d’equinoderms. L’estudi de la distribució corporal i l’estructuració de les plaques, a més dels apèndixs, de l’aparell mandibular i del sistema vascular hidràulic (posició interna del canal hidràulic radial), i també de l’evidència bioquímica i l’estudi de les variacions ontogenètiques del desenvolupament larval, han fornit explicacions filogenètiques sobre l’origen dels equinoïdeus. D’aquest procés investigador es desprèn que els equinoïdeus i els ofiuroïdeus presenten un nombre d’innovacions evolutives importants, que mostren una separació relativament recent (Ordovicià inferior) dels dos grups.

Els equinoïdeus apareixen en el registre fòssil, sencers, i com a plaques aïllades o pues. La fotografia mostra la diversitat de les pues dels cidaroïdeus. D’esquerra a dreta, pues, de 24 a 37 mm, de: Balanocidaris gibberula, del Coniacià de Prats de Carreu (Pallars Jussà); Porocidaris schmideli, del Lutecià de la Pobla de Claramunt (Anoia); Cyathocidaris cyathifera, del Coniacià-Santonià d’Herbasavina (Pallars Jussà); Stereocidaris ramondi, del Maastrichtià inferior de Salàs de Pallars; i Balanocidaris pleracantha del Campanià de la Pobla de Segur (Pallars Jussà).
Jordi Vidal / MGB-UAB.
El registre fòssil proporciona una evidència que hi ha hagut una transformació gradual del conjunt dels asteroïdeus fins als ofiuroïdeus actuals. Els resultats de les anàlisis cladístiques porten a pensar que els equinoïdeus evolucionaren a partir d’un grup d’ofiuroïdeus primitius, després que aquests se separessin dels asteroïdeus, però abans que evolucionessin cap a una distribució de plaques ambulacrals oposades diametralment. Aquests ofiuroïdeus ancestrals segurament devien ser flexibles, amb una distribució de plaques ambulacrals alternes, el canal hidràulic radial intern, l’aparell mandibular ben desenvolupat, plaques laterals reduïdes en el disc i una sola placa madrepòrica en posició marginal. Tot això fa pensar en un representant dels subordres dels protrurins (Protrurina), parofiurins (Parophiurina) o lisofiurins (Lysophiurina) com a possible avantpassats dels equinoïdeus.
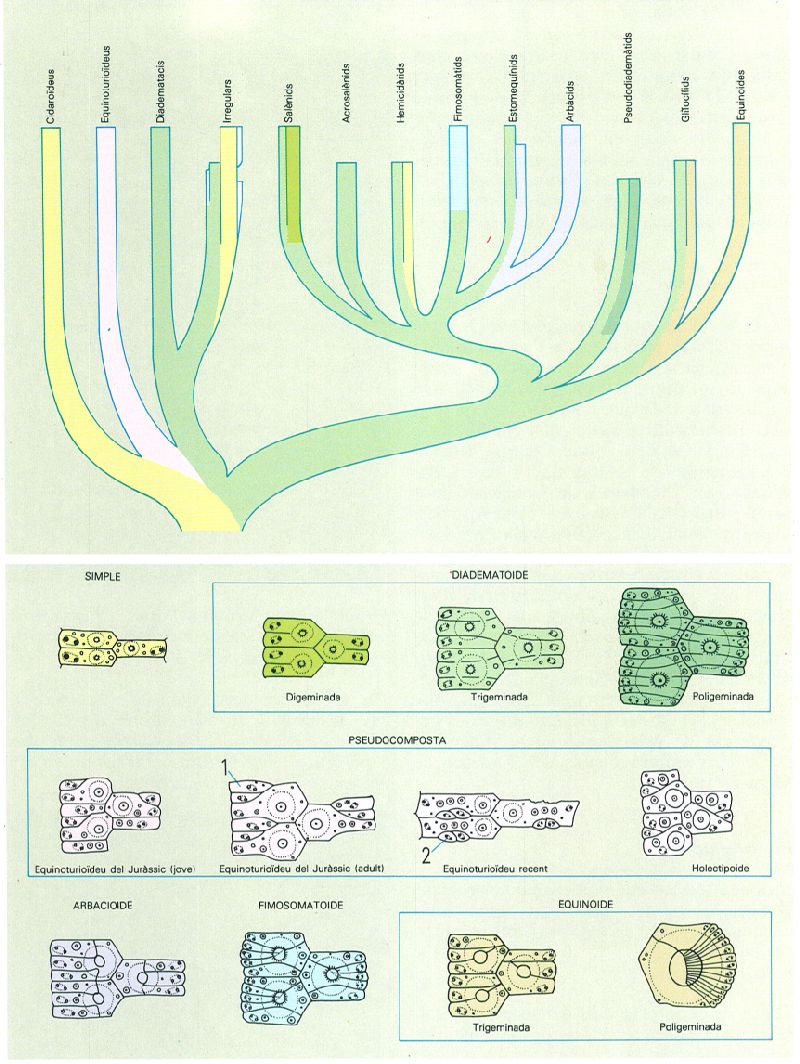

La disposició que adopten les plaques ambulacrals dels equinoïdeus postpaleozoics va des de les plaques simples dels cidaroïdeus, fins a les poligeminades, com en els equinoides. La part superior del dibuix il·lustra la distribució dels diferents tipus de plaques ambulacrals en la sistemàtica del grup i el seu possible desenvolupament evolutiu. Hom ha indicat en el dibuix: 1 hemiplaca, 2 placa ocluida.
Amadeu Blasco, original d’A. Smith.
El sistema de classificació més estès per als equinoïdeus, el del «Treatise on Invertebrate Paleontology», de Moore, és insatisfactori en certa mesura, però ha tingut una gran difusió entre els paleontòlegs, tant afeccionats com professionals. Entre els criteris utilitzats en aquest sistema de classificació hi ha el nombre de fileres de plaques ambulacrals i interambulacrals: els euequinoïdeus moderns tenen dues fileres de plaques per columna, mentre que els primitius periscoequinoïdeus en tenien un nombre variable. També es fa servir per a separar superordres i ordres la posició que ocupa el periprocte respecte del sistema apical, que pot ser inclòs (parlem de periprocte endocíclic) o bé extern (exocíclic); la disposició de les plaques del sistema apical, la presència o absència de llanterna i la morfologia de la dentició són altres caràcters utilitzats. La darrera opció sistemàtica presentada per Smith, que s’ajusta força més a una classificació filogenètica, contempla la divisió de la classe en tres subclasses: els periscoequinoïdeus (Periscoechinoidea), els cidaroïdeus (Cidaroidea) i els euequinoïdeus (Euechinoidea). Els periscoequinoïdeus no han estat trobats als Països Catalans. Inclouen tots els equinoïdeus paleozoics excepte els cidaroïdeus. Aparegueren a l’Ordovicià i s’extingiren al Permià. Reuneixen dos ordres: els equinocistitoides (Echinocystitoida), que tenien més de dues columnes en cada interambulacre i la teca flexible; i els palequinoides (Palaechinoida), d’esquelet rígid, sense anell perignàtic i amb interambulacres d’una o més de dues columnes.
Els cidaroïdeus

La persistència dels trets essencials dels cidaroïdeus al llarg dels temps geològics es manifesta en aguest exemplar de Leiocidaris montserratensis del Bartonià d’Avinyó (Bages), de 41 mm d’altura, amb ambulacres lleugerament sinuosos formats per plagues simples, i amb una forta tuberculació de les plaques interambulacrals. De fet, les diferències amb altres gèneres i espècies del mateix grup gairebé sempre són molt petites, i moltes vegades afecten la morfologia de les pues.
Jordi Vidal / MGB.
Els cidaroïdeus, que aparegueren al Devonià superior i han arribat fins a l’actualitat, són d’ambulacres bicolumnars i sistema apical amb un sol porus a cada placa genital. De l’ordre dels cidaroides (Cidaroida) podem citar Anaulocidaris faurai, del Muschelkalk superior de Colldejou (Baix Camp); Stereocidaris dallonii, de l’Albià de Pallerol (Alta Ribagorça); S. corbarica del Coniacià-Santonià d’Erinyà (Pallars Jussà); S. falgarsensis del Maastrichtià de Falgars (Berguedà); Rhabdocidaris salvae del Neocomià de Cocentaina (Comtat); Cyathocidaris cyathifera del Coniacià superior de la vall de Carreu (Pallars Jussà); Balanocidaris darderi de l’Aptià de Cocentaina; B. gibberula del Santonià dels Prats de Carreu (Pallars Jussà); Porocidaris schmideli del Lutecià-Biarritzià d’Amer (Selva); Prionocidaris sismondai del Langià de Vilobí del Penedès; Diplocidaris hispanicus de l’Oxfordià de Montanejos (Alt Millars); i Leiocidaris montserratensis del Biarritzià de Montserrat.
Els euequinoïdeus
Inclouen tots els equinoïdeus no cidaroïdeus postpaleozoics. Els dos grans grups dels euequinoïdeus són els regulars i els irregulars.
Els regulars

Els equinoïdeus regulars es diferencien dels irregulars pel fet que tenen el periprocte formant part del sistema apical. El pedinoide de la fotografia (Leiopedina tallavignesi), de 54 mm d’altura, és un dels equinoïdeus més comuns als jaciments catalans d’edat lutecianapriaboniana (l’exemplar prové de Manresa, al Bages). Noteu el contorn subpentagonal i les plagues compostes de tres elements.
Jordi Vidal / MGB.
Tenen el periprocte dins del sistema apical. Hom els classifica segons criteris fonamentats principalment en la presència o absència de perforació i crenulació en els tubercles de la corona, i pel tipus de llanterna. No tenim citacions al nostre país dels ordres dels equinoturioides (Echinothurioida), dels diadematoides (Diadematoida) ni dels micropigoides (Micropygoida). En canvi, els pedinoides (Pedinoida) sí que són presents al registre fòssil dels Països Catalans. Tenen els ambulacres formats per plaques simples o compostes, de tipus diadematoide. S’han trobat Diademopsis al Liàsic de Vandellòs, Hemipedina batalleri, també del Liàsic d’Alfara de Carles (Baix Ebre), Micropedina olisiponensis, del Cenomanià de Morella (Ports), i Leiopedina tallavignesi, tan freqüent en jaciments biarritzians de Catalunya.
De la cohort dels equinacis (Echinacea), caracteritzada per una corona rígida amb periprocte endocíclic, es distingeixen dos superordres: el dels estirodonts (Stirodonta) i els camarodonts (Camarodonta).
Els equinacis estirodonts

Tres equinacis estirodonts vistos per la cara apical. A dalt, el salenioide Salenia petalifera, del Campanià de Vallcebre (Berguedà), en el qual s’aprecia el seu sistema apical característic: l’anus desplaçat cap al costat dret o posterior i les plaques complementàries; aquest grup, els representants del qual són de petita talla (ací, 23 mm) i presenten forts tubercles, s’associa a fàcies d’esculls coral·lins o de rudistes. Al mig, el fimosomatoide Lambertechinus arnaudi, de 37 mm de diàmetre, procedent del Santonià de Clot d’Olsi (Montsec), on vivia arrecerat entre esculls de coralls i rudistes; el sistema apical dels eriçons d’aquest grup és caduc i destaca la semblança entre els tubercles ambulacrals i els interambulacrals. A baix, l’arbàcid Goniopygus, freqüent en materials cretacis (el de la fotografia prové del Santonià de la Llobera, al Montsec), del qual hom creu, en relació amb la densa i forta tuberculació de la corona i amb els tipus de sediments on es troba, que era propi d’ambients litorals; té un sistema apical dicíclic, que caracteritza aquest gènere.
Jordi Vidal / MGB i UAB.
Els estirodonts reben el nom del tipus de dentició de la llanterna que presenten. Els dos ordres que en diferenciem són presents als Països Catalans: els salenioides (Salenioida) i els fimosomatoides (Phymosomatoida).
Els salenioides són molt característics perquè tenen el sistema apical proveït d’una o més grans plaques supraanals poligonals, fortament connectades a l’anell oculogenital, que en conjunt recorda un calze, i fa que el periprocte quedi desplaçat posteriorment o posterodextralment. Alguns representants de l’ordre trobats al nostre país són: Acrosalenia faurai del Toarcià de Tortosa (Baix Ebre), Salenia prestensis, de l’Aptià de Canyelles (Garraf), Morella (Ports) o Montiberri (Alta Ribagorça), S. cenomanensis de Marmellar (Baix Penedès), S. personata, esmentada sense adscripció estratigràfica a Menorca, i Hyposalenia archiaci, de l’Aptià de la Casa Alta, a Castellet (Garraf).
Els fimosomatoides solen presentar plaques simples tot al llarg dels ambulacres o, més correntment, compostes del tipus diadematoide, trigeminades o polípores. En són exemples Phymosoma loryi de l’Aptià de Canyelles de Garraf, P. subnudum del Santonià del Clot d’Olsi (Montsec d’Ares), P. hexaporum del Maastrichtià de Salàs de Pallars, Leptechinus mestreae del Bajocià de Tivenys (Baix Ebre), Micropsis hispanica del Santonià de Berga, Porosoma faurai del Santonià del Montsec de Rúbies, P. cribrum del Bartonià de Castellolí i de Collbàs (Anoia); Rachiosoma rubiesensis, del Santonià del Montsec de Rúbies; Tylechinus vidali, del Coniacià-Santonià de la Nou (Noguera); Codechinus rotundus, de l’Aptià de Fredes (Baix Maestrat) i del Mas de Barberans (Montsià), i Trochalosoma vidali, del Santonià de Fígols (Berguedà).
Completen el subordre dels estirodonts les famílies dels hemicidàrids (Hemicidaridae) i dels arbàcids (Arbaciidae). De la primera coneixem Hemicidaris vilanovae, de l’Aptià d’Ador (Safor), Heterodiadema libycum del Cenomanià de cap Llentrisca (Eivissa), Acrocidaris nobilis, de l’Oxfordià de Montanejos (Alt Millars), Polydiadema gurgitis de l’Aptià de Puigcerver (Baix Camp), Tetragramma brogniarti, de l’Aptià d’Aiguamúrcia (Alt Camp), i T. variolare, del Cenomanià d’Abella de la Conca (Pallars Jussà). Dels arbàcids s’ha trobat, entre altres, Codiopsis alpina, de l’Aptià de la Morisca (Garraf), C. douvillei del Santonià de Clot d’Olsi (Montsec d’Ares), C. fontei del Maastrichtià de Carbonils (Alt Empordà), Coelopleurus coronalis, tan comú en el Lutecià català, Cottaldia royoi, de l’Aptià de la Mola de Miró (Ports), Goniopygus (Goniopygus) marticensis, també del Clot d’Olsi, i G. (G.) royanus, del Campanià de la Solana d’En Mauri (Berga) i del Maastrichtià de Fígols Vell (Berguedà).
Els equinacis camarodonts

Els ortòpsides, com aquest Orthopsis miliaris del Maastrichtià inferior de la serra dels Homes Morts (Pallars Jussà), són coneguts als Països Catalans en materials cretacis que van de l’Aptià al Maastrichtià. El sistema apical dicíclic, el perfil força aplanat de l’esquelet i la tuberculació regular i homogènia són elements propis, encara que no definitius, d’aquest ordre d’equinacis camarodonts de posició sistemàtica encara incerta. L’exemplar fotografiat ateny 34 mm de diàmetre.
Jordi Vidal / UAB.
En el superordre dels camarodonts, distingim quatre ordres: els temnopleuroides (Temnopleuroida), els equinoides (Echinoida), els plesiocidaroïdeus (Plesiocidaroida), de posició sistemàtica incerta i no trobats al nostre país, i els ortòpsides (Orthopsida). Els temnopleuroides es caracteritzen per tenir una corona normalment esculpida amb crestes (epistroma) i depressions suturals, almenys en els estats immaturs. Triplacidia vandenheki, del Priabonià de la Tossa de Montbui, Arbacina catenata de l’Helvecià d’Elx (Baix Vinalopó), Schizechinus duciei del Messinià de serra Colmenar (Baix Vinalopó), Prionechinus faurai de l’Helvecià de Montjuïc (Barcelonès), i Tripneustes gahardensis del Langià de Vilobí del Penedès són exemples d’aquest ordre. Els equinoides tenen la corona no esculpida i els solcs branquials soms. S’han pogut estudiar Psammechinus dubius, del Langià de Vilobí del Penedès i de la serra Colmenar, P. formosus també de Vilobí, i Isechinus delphinus del Burdigalià dels cingles de Bertí (Vallès Oriental i Osona). Dels ortòpsides, coneixem Orthopsis royoi de l’Aptià de Vallibona (Ports), i O. miliaris del Campanià de Vallcebre (Berguedà) i Maastrichtià de la serra dels Homes Morts (Salàs de Pallars).
Els irregulars

Entre els eriçons irregulars, de simetria bilateral, la tuberculació fina o inexistent és un fet generalitzat en el grup dels holectipoides, del qual veiem un exemplar de Coenholectypus similis, de 20 mm de diàmetre, procedent de l’Aptià de Morella (Ports). Els holectipoides, que vivien en ambients inframareals i d’escull, són especialment ben representats al llarg del Cretaci i del Paleogen.
Jordi Vidal / UAB.
Els equinoïdeus irregulars tenen el periprocte fora del sistema apical. Els grans grups s’estableixen segons la presència o absència de llanterna, tot i que aquesta sempre és fortament modificada respecte de la que presenten els regulars. Hom en considera dos superordres: els eognatostomats (Eognathostomata) i els microstomats (Microstomata). Tenen llanterna els eognatostomats i els microstomats de la sèrie dels neognatostomats.
Els eognatostomats
Dins d’aquest superordre, no coneixem al nostre país fòssils de pigasteroides (Pygasteroida) però sí dels holectipoides (Holectypoida). Els representants d’aquest grup són d’hemisfèrics a globulars i ovoides, amb un sistema apical monobasal o bé amb quatre o cinc plaques genitals, mentre que el periprocte s’obre ambitalment o adoralment. Força comuns en els jaciments mesozoics i cenozoics són, per exemple, Coenholectypus macropygus de cala Llentrisca i illa Espart (Eivissa), i C. similis de Morella (Ports), ambdós de l’Aptià; C. turonensis del Santonià del Montsec d’Ares, C. proximus del Maastrichtià de Salàs del Pallars; Discoides decoratus, de l’Aptià de Marmellar (Baix Penedès); Dixonia infera, del Cenomanià de Sopeira (Alta Ribagorça); Conulus haugi, del Campanià de Torallola (Pallars Jussà); C. gigas, del Maastrichtià de la serra dels Homes Morts; Globator ovulum del Coniacià-Santonià de la serra de Sant Corneli; G. darderi del Campanià de Bocairent (Vall d’Albaida), G. petrocoriensis del Campanià de Torallola, Conoclypus lucentinus, del Lutenià de la Pobla de Roda (Alta Ribagorça) i Callosa d’En Sarrià (Marina Baixa), i Echinogalerus belgicus, del Maastrichtià de Sapeira (Pallars Jussà).
Els microstomats neognatostomats

L’aspecte petaloide dels ambulacres i la forma allargada o en cor de l’esquelet són d’altres característiques que distingeixen fàcilment els eriçons irregulars. Aquestes, en concret, solen correspondre a eriçons d’hàbits excavadors o infaunals. La fotografia mostra el cassiduloide Echinolampas ovalis, de 59 mm de llargada, procedent del Lutecià d’Igualada (Anoia). Els cassiduloides, sempre ovalats, amb ambulacres petaloides i proveïts d’un sistema apical monobasal, són considerats com el grup del qual arrenquen els clipeasteroides.
Jordi Vidal / MGB.
Tant la sèrie dels neognatostomats com la dels atelostomats són ben representades en els diferents ordres que contenen al nostre país. Els neognatostomats inclouen els ordres dels cassiduloides (Cassiduloida), els oligopigoides (Oligopygoida), dels quals no tenim cap representant, els clipeasteroides (Clypeasteroida) i els neolampadoides (Neolampadoida), tampoc representats al nostre país.
Els cassiduloides tenen ambulacres petaloides a la cara adapical, a més de fíl·lodes i «bourrelets», i en estat adult no tenen mandíbules ni solcs branquials. Podem esmentar Pyrgorhynchus royoi de l’Aptià de Cinctorres, P. dallonii del Santonià de Montesquiu (Pallars Jussà), Echinolampas archiaci del Bartonià de Monistrol (Bages), E. barcinensis de Vilobí, i E. scutiformis de Menorca, ambdós del Langià; E. hemisphaerica del Tortonià de la serra de les Talaies (Vall de Cofrents); E. hoffmanni, del Pliocè de Santa Pola (Baix Vinalopó); Plesiolampas lorioli, del Lutecià de Montfalcó (Baixa Ribagorça), Areny (Alta Ribagorça) i Alacant; Parapygus nanclasi, del Santonià d’Aramunt (Pallars Jussà); Faujasia faujasi del Campanià de Berga; Fauraster priscus, del Maastrichtià d’Orrit (Alta Ribagorça); Nucleopygus, del Turonià-Santonià de la serra de Sant Corneli; Rhynchopygus, de la serra dels Homes Morts; Clypeolampas, del Santonià de Sant Corneli; i Studeria ficheuri, del Langià de les Cases de Montpaó (Baix Penedès).

Els clipeasteroides, representats encara actualment en mars càlides, són de contorn pentagonal o arrodonit i tenen ambulacres petaloides i suports interns. A la fotografia veiem, a dalt, la cara apical d’un Clypeaster indeterminat procedent de la conca del Vallès-Penedès, i a baix, el motlle intern, també en vista apical, de C. barcinensis del Tortonià de Montjuic (Barcelonès), on s’aprecien clarament els pilars i contraforts (en negatiu) i el sistema intern de canals ambulacrals; el color es deu a òxids de ferro presents en la matriu. Mesuren, respectivament, 150 i 125 mm de longitud antero-posterior.
Jordi Vidal / MGB.
Els clipeasteroides tenen una corona ovoidal aplatada, els ambulacres petaloides, les plaques genitals fusionades, els peus ambulacrals primaris de funció respiratòria i restringits als pètals, mentre que els accessoris, nombrosos, s’estenen fora dels pètals i en algunes formes arriben als interambulacres. El peristoma sol ser petit i l’esquelet normalment presenta suports interns. Podem esmentar, entre molts altres, Clypeaster doma, de Castellvell del Camp (Baix Camp); C. hardouini, de Sant Sadurní d’Anoia; i C. scutellatus de Clariana (Garraf), tots tres del Langià; C. galerus, de l’Helvecià de Muro (Mallorca), C. scillae, de Roda de Berà (Tarragonès) i de la serra Grossa (Costera), C. altus de la serra de Sancho (Alacantí), C. barcinensis de Montjuïc (Barcelonès), i C. lamberti de la serra d’Orxell (Baix Segura), tots del Tortonià. Hi ha, a més, Echinocyamus pusillus del Pliocè de Sant Miquel de Salines (Baix Segura), Scutella tarraconensis del Langià de Roda de Berà, S. bofilli de Castellbisbal (Vallès Occidental) i de Sant Pau d’Ordal (Alt Penedès), i S. lusitanica de Sant Sadurní d’Anoia, totes dues de l’Helvecià; i S. almerai de la Bisbal del Penedès i S. faurai de Montjuïc, totes dues del Tortonià. Esmentarem, encara, Amphiope montezemoloi de l’Helvecià de Cerdanyola del Vallès i de Montferri (Alt Camp), i A. bioculata del Tortonià del port de Maó (Menorca).
Els microstomats atelostomats

Eriçó microstomat (holasteroide), vist per la cara apical, on s’aprecia la forma típicament allargada del sistema apical. De vida semiinfaunal, la tuberculació és molt fina i gairebé imperceptible. Noteu, a l’exemplar de la fotografia (Echinocorys tenuituberculata, de 80 mm de longitud, procedent del Maastrichtià de Sapeira, a l’Alta Ribagorça), la disposició de les plaques simples en la corona.
Jordi Vidal / UAB.
Els irregulars de la sèrie dels atelostomats presenten plaques ambulacrals simples, i en l’estat d’adult no tenen llanterna ni anell perignàtic ni solcs branquials. Hom en distingeix tres ordres: els disasteroides (Disasteroida), que no tenim representats en el nostre registre fòssil, els holasteroides (Holasteroida) i els espatangoides (Spatangoida), que hi són profusament representats.
Els holasteroides tenen el sistema apical típicament allargat o disjunt, sense la genital 5. Presenten un plastró dèbilment diferenciat. Els pètals aparellats no són mai impresos i el sistema apical i el peristoma sovint es troben en posicions oposades. Poden mostrar diferents tipus de fascíoles. En són exemples: Holaster aptiensis de l’Aptià de Castellet (Garraf); H. bonansensis, de l’Albià de Bonansa (Alta Ribagorça); H. subglobosus i H. trecensis del Cenomanià de Sopeira (Alta Ribagorça); Cardiaster hermitei del Santonià de Santa Fe (Alt Urgell); C. integer del Santonià i C. heberti del Campanià de la serra de Sant Corneli; Echinocorys ovata i E. vulgaris del Coniacià-Santonià dels Prats de Carreu (Pallars Jussà); E. conoidea i E. heberti del Campanià de Sant Corneli i Sensui (Pallars Jussà); E. darderi del Maastrichtià de la serra de Bèrnia (Marina); Hemipneustes cerdanyolae del Campanià de Sant Julià de Cerdanyola (Berguedà), i H. pyrenaicus, del Maastrichtià de Sapeira i Orcau (Pallars Jussà) i Fígols Vell (Berguedà); Offaster dallonii del Maastrichtià de Sapeira, Pseudoffaster caucasicus del Maastrichtià de Sobrecastell (Alta Ribagorça); Stegaster novoi del Campanià-Maastrichtià del coll de Maigmó (Alacantí); Coraster vilanovae del Maastrichtià de Callosa d’En Sarrià i Orxeta (Marina Baixa); i Physaster vasseuri, del Cenomanià de Sopeira (Alta Ribagorça).

Tres exemples d’espatangoides. A dalt, un toxastèride típic, Heteraster oblongus, de 40 mm de longitud, procedent de Morella (Ports), desproveït gairebé del tot d’ornamentació; probablement vivia semienterrat en els fons margosos i sorrencs aptians, nivells que caracteritza i en els quals sol aparèixer juntament amb Toxaster collegnoi. Al mig, un dels micrasterinis més comuns de l’Eocè, Eupatagus, identificable per la seva morfologia, els seus ambulacres semipetaloides i per l’ornamentació, poc densa però acusada a la superfície apical; l’exemplar de la fotografia, de 74 mm de longitud, és de l’espècie E. faurai i prové del Lutecià de Navarcles (Bages). A baix, l’hemiasterí Proraster atavus, del Coniacià superior de la collada del Gassó (Pallars Jussà), de 21 mm de llargada; la morfologia general de l’esquelet correspon a la típica del grup: contorn suboval, amb un solc frontal més o menys acusat i amb ambulacres parells petaloides voltats d’una fascíola peripètala necessària per a la funció respiratòria, atesa la seva manera de viure enterrat.
Jordi Vidal / UAB, MGB.
Els equinacis de l’ordre dels espatangoides (grup d’equinoïdeus irregulars apareguts al final del Cretaci inferior) experimentaren una radiació evolutiva important que donà lloc a l’aparició de bona part dels grups actuals. Hom creu que tenien pues dorsals espatulades que els ajudaven a mantenir-se als seus habitacles, ja que eren excavadors i vivien en sediments fangosos. Els exemplars tenen un sistema apical compacte etmofracte o etmolític, no oposat al peristoma, amb quatre o menys gonoporus. La cara oral de l’interambulacre posterior normalment es diferencia en un plastró amb un parell de plaques esternals per sota del labre terminal. Les àrees ambulacrals de la cara adapical prenen totalment o parcialment forma petaloide, mentre que el peristoma sol ser excèntricament anterior i labiat, tot i que certes formes el presenten rodó o pentagonal. Molts espatangoides (excepte els de la família dels toxastèrids) presenten fascíoles peripètala, marginal, subanal, lateroanal o interna, o una combinació d’aquestes. Componen aquest ordre, a més de la família dels toxastèrids (Toxasteridae), els subordres dels micrasterinis (Micrasterina), els hemiasterinis (Hemiasterina) i un altre desconegut encara i inexistent al nostre país, representat per la família dels asterotomàtides (Asterotomatidae).
Entre els toxastèrids, que són formes que representen l’arrel dels espatangoides, coneixem Toxaster collegnoi de l’Aptià de Garraf; Aphelaster integer del Hauterivià d’Artà (Mallorca); Heteraster oblongus de l’Aptià de Morella i d’Herbés (Ports); Epiaster prior, de l’Aptià de Jafra (Garraf); E. crassisimus, de l’Aptià de cala Molins (Eivissa); E. dallonii, del Cenomanià de Sopeira (Alta Ribagorça); E. trigonalis, del Cenomanià de Sabinar (Serrans) i E. laxoporus, del Coniacià-Santonià d’Abella de la Conca (Pallars Jussà).
Del subordre dels micrasterins, caracteritzats per una fascíola subanal, absent en certs membres de la família dels lovènids (Loveniidae), que pot combinar-se amb una fascíola peripètala o interna, esmentarem Micraster brevis del Coniacià-Santonià dels Prats de Carreu i del congost d’Erinyà (Pallars Jussà); M. matheroni, del Santonià de Sant Corneli (Pallars Jussà); M. aturicus, del Campanià de Montesquiu; M. gourdoni, del Campanià de Sant Corneli i Vilacarle (Baixa Ribagorça); Brissopneustes vilanovae, del Maastrichtià de Callosa d’En Sarrià; Isopneustes heberti, del Maastrichtià de la serra Mariola; Brissus unicolor, del Messinià de la serra Colmenar i del pantà del riu Vinalopó; Brissopsis lusitanicus, del Langià dels Monjos (Alt Penedès) i de Sant Vicenç de Berà; B. lyrifera del Pliocè del Masrampinyo (Montcada i Reixac, al Vallès Occidental); B. papiolensis, del Pliocè del Papiol (Baix Llobregat); Eupatagus cranium del Lutecià de Navarcles (Bages) i de tantes altres localitats; E. cossmanni, del Lutecià-Biarritzià de Vilada (Berguedà) i de Roda de Ter (Osona); E. confractus del Bartonià de Collbàs (Anoia); Gualtieria almerai, del Lutecià de Sant Julià de Vilatorta (Osona); Schizobrissus, del Messinià de la serra del Colmenar i de Santa Pola (Baix Vinalopó); Trachypatagus tuberculatus, del Miocè menorquí; i Echinocardium cordatum, del Pliocè de Sant Onofre (Tortosa).
Dels hemiasterins, que presenten una fascíola peripètala combinada en certes famílies amb una altra de lateroanal o de marginal, s’han trobat, entre altres, Hemiaster aragonensis, del Cenomanià de Sopeira, H. punctatus del Coniacià-Santonià de la vall de Carreu, Hemiaster del grup prunella del Maastrichtià de Sapeira (Alt Ribagorça) i Orcau (Pallars Jussà), Ditremaster nux del Lutecià de Sant Climent d’Amer (Selva), de Collbàs i de Montbui (Anoia), Opissaster almerai del Languià de Castellet (Garraf), Palhemiaster ibericus de l’Aptià del S de la Foradada (Eivissa), Ornithaster evaristei del Maastrichtià de Callosa d’En Sarrià i de l’Alfàs (Marina Baixa), Pericosmus latus del Miocè menorquí, Schizaster montserratensis del Lutecià de la Calsina, S. spado del Lutecià-Biarritzià de Vilada, S. studeri del Priabonià d’Olot (Garrotxa), S. lovisatoi del Languià de Calafell (Baix Penedès), i dels Monjos (Alt Penedès), S. saheliensis del Tortonià de Montjuïc (Barcelonès), S. desori de l’Helvecià de Sant Sadurní d’Anoia, Agassizia pachecoi del Tortonià de Montjuïc, Linthia heberti del Lutecià d’Areny de Noguera (Alta Ribagorça), i Proraster atavus del Coniacià-Santonià de Corçà (Noguera).
Història evolutiva dels equinoïdeus
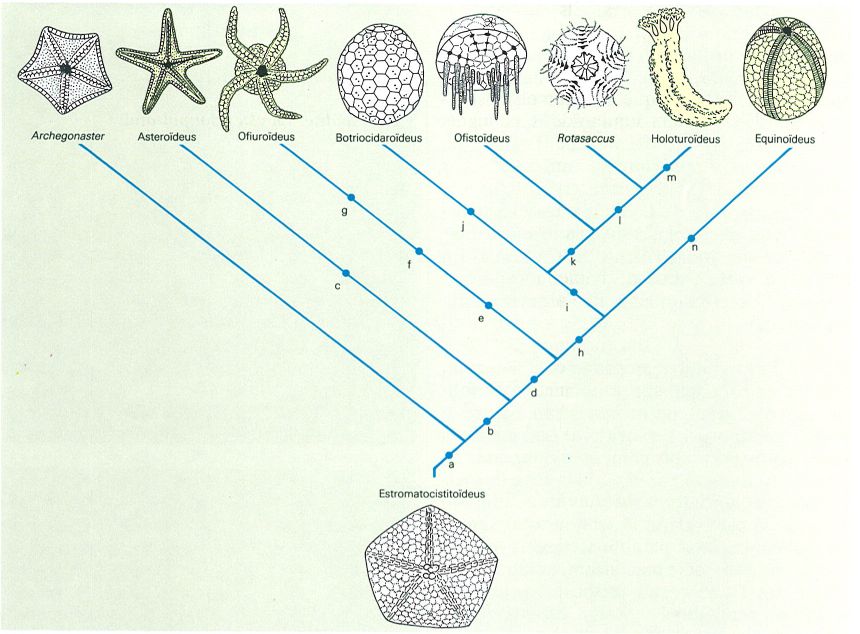
Cladograma que relaciona els diferents eleuterozous. Les lletres indiquen les innovacions esdevingudes dins de cada línia evolutiva: a aparició del madreporit i de les plaques modificades en adambulacrals i laterals; b adopció d’un perfil estel·lat (creixement radial); c madreporit aboral i larva bipinnària; d pèrdua de flexibilitat dels ambulacres i aparició de la primera mandíbula i de la larva de tipus plúteus; e ambulacres en posició oposada a l’odontòfor; f fusió dels ambulacres; g plaques dels braços dorsals i ventrals i escuts oral i radial; h periprocte que s’obre a la cara aboral, creixement meridional plaques laterals reduïdes a pues ambulacrals; i peus ambulacrals amb plaques i interambulacres reduïts a una columna o inexistents; j madreporit aboral i reducció dels elements mandibulars; k pèrdua de les pues i creixement dels peus ambulacrals orals; l paret corporal d’espícules circulars; m larva de tipus vitel·lària i llanterna reduïda a un anell calcari; n canal hidràulic radial intern i madreporit aboral. S’han representat en blanc els grups enterament fòssils i en groc els que encara són presents actualment.
Amadeu Blasco, original d’A. Smith, modificat.
La història evolutiva dels equinoïdeus pot ser interpretada com una sèrie d’innovacions adaptatives que portaren a l’adopció de nous sistemes de vida o que permeteren la colonització de nous hàbitats. Cada innovació adaptativa s’inicià amb l’evolució d’alguns trets morfològics nous d’importància i portà a un període de radiació i diversificació morfològica força ràpids a mesura que el nou nínxol era ocupat. Aquesta història evolutiva permet d’explicar com els equinoïdeus han evolucionat per a explotar la varietat d’hàbitats diferents que ocupen en l’actualitat.
Equinoïdeus paleozoics
La diversitat que presenten els equinoïdeus del Paleozoic és molt baixa si hom la compara amb la dels postpaleozoics. Els equinoïdeus devien estar limitats a ocupar hàbitats de poca energia en «offshore», i probablement mai no arribaren a adaptar-se a viure en la gran varietat d’hàbitats en què ho feren els postpaleozoics. Els primers equinoïdeus coneguts procedeixen de l’Ordovicià superior. Mentre que tots els equinoderms cambrians eren suspensívors, a l’Ordovicià aparegueren les primeres espècies menjadores de sediment, entre les quals hi havia els equinoïdeus proveïts d’un aparell mandibular mòbil, ben adaptat a tallar, i que probablement podien utilitzar una varietat més gran d’aliments, com ara algues o sediment lligat per organismes. L’evolució de la llanterna continuà, ja que en el Silurià inferior era emprada com a aparell mossegador. Aquesta nova tècnica alimentària permeté als equinoïdeus l’ús de fonts alimentàries no accessibles als altres grans grups d’equinoderms.
Durant el Silurià i el Devonià, el nombre d’espècies va créixer lentament mentre s’esdevenia una diversificació morfològica gradual, que establí les principals línies evolutives del Paleozoic superior. El Carbonífer inferior va veure el nombre d’espècies d’equinoïdeus màxim dins del Paleozoic, ja que proliferaren les espècies dels grups presents; els quatre principals grups presentaven sistemes de vida i hàbitats diferents. Així, els proterocidaròids (Proterocidaroidae), amb corones aplatades i amples ambulacres a la superfície oral, eren bàsicament detritívors, mentre que els arqueocidàrids (Archaeocidaridae), amb grans pues defensives, eren omnívors oportunistes que vivien en fons marins oberts. Els lepidèstids (Lepidesthidae), amb nombrosos peus ambulacrals, probablement mantenien una cobertora per damunt del seu esquelet com a camuflament; també, com en el cas dels arqueocidàrids, desenvoluparen estratègies defensives que semblen indicar un especial problema de predació sobre els equinoïdeus que vivien en zones obertes. No és el cas dels palequínids (Palaechinidae), que, associats a fàcies d’interescull i amb corones tessel·lades gruixudes, s’arreceraven entre antics talussos d’escull o en els seus forats i fissures.
El Carbonífer superior veié una gran davallada dels equinoïdeus, que arribà a l’extinció d’algunes línies evolutives (els palequínids). Aquesta tendència continuà durant el Permià, del qual tan sols coneixem sis espècies. Els equinoïdeus més comuns en aquells moments podrien agrupar-se en dos tipus generals: el dels grans i molt especialitzats protocidàrids, que eren detritívors, i el dels petits i oportunistes omnívors Miocidaris i el minúscul equinocistitoïdeu Xenechinus. A la fi del Paleozoic, en unes condicions més rigoroses i imprevisibles, es produí probablement una reducció global de les fonts alimentàries primàries, com demostra l’alta especialització que arribaren a desenvolupar els protocidàrids, que en iniciarse el Triàsic ja haurien desaparegut.
Equinoïdeus regulars postpaleozoics
Només Miocidaris sobrevisqué a la crisi del trànsit Permià-Triàsic i salvà de l’extinció els equinoïdeus. Si bé les condicions del Triàsic inferior i mitjà encara eren força rigoroses, en iniciar-se el Triàsic superior aquestes es feren favorables per a una nova expansió dels equinoïdeus, que ja no eren representats només pels miocidàrids (Miocidaridae), sinó també pels primers cidaroïdeus i acroequínids i pels plesiocidàrids. Més endavant devien aparèixer els primers equinacis, de manera que cap al final del Triàsic, les línies principals dels equinoïdeus regulars postpaleozoics ja eren establertes.
Tant els cidaroïdeus com els acroequínids desenvoluparen una estructura més forta de la llanterna i les mandíbules, amb una musculatura més eficient lligada per primera vegada a un anell perignàtic i una corona suturada que els donava la rigidesa necessària per a suportar aquesta musculatura. Aquests canvis demostren una adaptació a un nou règim alimentari que implicava una millora de l’acció rosegadora i d’arrencament de l’aliment. La manca de competitivitat afavorí el gran nombre d’innovacions morfològiques que, al seu torn, permeté la ràpida diversificació dels equinoïdeus. Aquesta tendència continuà durant el Juràssic, que aparegueren la majoria dels ordres tant dels equinoïdeus regulars com dels irregulars. Els cidaroïdeus canviaren poc des de la seva aparició, com tampoc no varià gaire el seu nínxol ecològic. En canvi, els acroequínids experimentaren una evolució adaptativa considerable que els permeté colonitzar hàbitats no explotats prèviament. I això fou per raó de l’existència d’una llanterna molt més lleugera i maniobrable que no la dels cidaroïdeus, i del desenvolupament de les plaques compostes que els permeté posseir pues ambulacrals més grosses i, per tant, una defensa global més perfeccionada.
Al principi del Juràssic, els pedinoides (Pedinoida) i els primers equinacis eren molt similars, si bé la mandíbula carenada dels darrers, mecànicament més forta que no la dels primers, els permeté millorar la seva acció sobre els organismes incrustants, i s’expandiren i es diversificaren com a brostejadors, mentre que els pedinoides conservaven una dieta menys especialitzada. Els pseudodiademàtids (Pseudodiadematidae), els primers equinacis coneguts, originaren diferents grups durant el Juràssic. Els hemicidàrids (Hemicidaridae), per exemple, són convergents amb els cidàrids i, probablement, ocuparen un hàbitat similar. Els acrosalènids (Acrosaleniidae), amb un periprocte excèntric, experimentaven un nou tipus de dieta. Els fimosomàtides (Phymosomatidae) eren coberts de pues curtes que els protegien dins les fissures de les roques on vivien. Els estomequínids (Stomechinidae) tenien uns forts peus ambulacrals orals i pogueren colonitzar zones rocoses somes que no havien estat encara mai ocupades pels equinoïdeus. Els arbàcids (Arbaciidae), lligats als anteriors, encara eren més ben adaptats a medis molt soms d’aigües càlides, gràcies al desenvolupament de peus ambulacrals respiratoris especialitzats.
La diversificació dels equinacis va continuar en un grau més petit durant el Cretaci. Els temnoplèurids (Temnopleuridae), amb l’adopció d’una llanterna camarodonta, competiren amb èxit amb els grups estirodonts preexistents i començaren a desplaçar-ne alguns grups cap al final del Cretaci, que els pseudodiademàtids, els hemicidàrids i els acrosalènids ja s’havien extingit. Els camarodonts esdevingueren llavors els equinoïdeus regulars dominants durant el Cenozoic i s’anaren diversificant amb l’aparició dels equinoides. Aquests camarodonts s’adaptaren als fons soms rocosos en franca competència amb els estomequínids i els arbàcids. Actualment, aquells són gairebé a punt d’extingir-se, però els arbàcids aconsegueixen perdurar gràcies probablement al fet que han aconseguit adaptar-se a aigües més calentes i somes més bé que la resta dels equinoïdeus.
Equinoïdeus irregulars
Hom creu que els equinoïdeus irregulars evolucionaren a partir d’equinoïdeus regulars oportunistes, com Eodiadema, que s’adaptaren a viure i a alimentar-se en sediments no consolidats en iniciar-se el Juràssic inferior, i que es produí una diversificació progressiva molt ràpida, acompanyada d’una diversificació morfològica a favor de la poca competència i pressió selectiva. El canvi d’estil de vida dels regulars als irregulars aparegué com un fet gradual. Els primers irregulars, com ara els pigasteroides i els holectipoides, només podien excavar en sediments relativament grollers i vivien semiinfaunalment. Probablement els calia tornar a la superfície per a cercar aliments, que encara col·lectaven per mitjà de la llanterna. Però cap a la fi del Liàsic inferior, ja existien galeropigoides que utilitzaven peus ambulacrals xucladors per a recollir l’aliment i que es movien en una direcció, cabussant-se en el sediment; una concentració més gran de les pues els permeté progressar en sediments de gra més fi. En un altre període de diversificació morfològica s’establiren les línies evolutives dels cassiduloides i els disasteroides.
Disasteroides i cassiduloides, probablement competidors mutus en els seus inicis, s’especialitzaren ràpidament per a diferents nínxols ecològics: mentre els cassiduloides desenvoluparen «bourrelets» i fíl·lodes, cosa que els feia aptes per a empassar grans quantitats de partícules de la mida dels grans de sorra, i per tant els permetia colonitzar zones amb sediments de contingut orgànic baix, els disasteroides desenvoluparen peus ambulacrals peniciliats per a la recol·lecció per adhesió mucosa de material de gra fi, de manera que eren més selectius en la seva alimentació i admetien un ventall més ampli de dimensions de partícules en la seva dieta.
L’evolució dels peus ambulacrals peniciliats portà, en el trànsit del Juràssic al Cretaci, a l’aparició d’espatangoides i holasteroides. Molt més ben adaptats a excavar, els espatangoides probablement podien viure infaunalment en sediments molt fins i fins i tot fangosos. Posseïen pues dorsals espatulades per a mantenir els seus habitacles i fascíoles per a crear corrents hidràulics que els servien per a millorar l’evacuació dels materials fecals i per a l’alimentació (transport de nutrients des de la interfase aigua-sediment per mitjà d’una adaptació de l’ambulacre anterior). Els holasteroides, en canvi, vivien epifaunalment o excavaven en sediments més permeables, cosa que feia innecessari el manteniment d’un conducte respiratori fins a la superfície dels sediments.
Després d’una davallada de la diversitat que s’esdevingué cap al final del Cretaci i durant el Paleocè, els espatangoides sofriren una radiació important que desembocà, durant l’Eocè, en l’aparició dels grups actuals, adaptats a gairebé tots els tipus de sediment. Els holasteroides, per contra, no superaren el trànsit entre el Mesozoic i el Cenozoic, i actualment els pourtalèsids, semiinfaunals i adaptats a ambients molt profunds, poden considerar-se un grup amb èxit. També els clipeasteroides aparegueren probablement a partir dels cassiduloides, durant el Paleocè i es diversificaren durant el Cenozoic. El fet de ser coberts d’un gran nombre de peus ambulacrals xucladors molt prims els permeté manipular sediments més fins que els dels seus avantpassats, amb la possibilitat de colonitzar nous hàbitats. Molts d’aquests aconseguien d’aportar partícules detrítiques a la boca a base de crear corrents ciliars per al transport d’aigua oxigenada, i alhora tamisaven el material fi del sediment que els cobria per mitjà de les pues adorals, com un sedàs. Això els portà a un aplanament del seu esquelet, a la ramificació i a la posterior expansió del seu sistema de solcs alimentaris i a l’augment del perímetre de la corona mitjançant el desenvolupament de diversos forats i lúnules.