Sistemes aquàtics canviants, fauna i flora adaptables
La coincidència del període eixut amb el càlid determina la principal característica dels sistemes aquàtics mediterranis: la irregularitat dels cabals fluvials i els canvis del nivell d’inundació en llacs i llacunes naturals. El règim hídric mediterrani té un cabal màxim de tardor associat al període principal de pluges, tant a l’hemisferi nord com a l’hemisferi sud, excepte a Austràlia, on es produeix a l’hivern. Però a les zones més humides n’hi pot haver dos, un a la primavera i l’altre a la tardor i passa el mateix a les conques mediterrànies que neixen a l’alta muntanya (Pirineus, Sierra Nevada d’Amèrica del Nord i Andes), on l’increment primaveral de cabal és conseqüència directa del desgel produït a les capçaleres.
Les conques amb un règim hídric estrictament mediterrani presenten una sèrie de característiques generals comunes. Són curtes (a excepció de la conca parcialment mediterrània del riu Murray, a Austràlia), tenen canvis de cabal molt pronunciats i mostren una tendència clara a la temporalitat: a l’època càlida el llit queda totalment o parcialment sec, o bé les aigües són intermitents. El grau de temporalitat arriba a la màxima expressió a les rambles de la Península Ibèrica i als “oueds” africans, que queden eixuts la major part de l’any. Els acusats períodes de sequedat solen anar acompanyats d’avingudes o riuades estacionals, que arriben a cert grau de catastrofisme, durant l’època de màxima precipitació, tardor o hivern segons el continent. Tanmateix, poden produir-se crescudes en altres èpoques de l’any, especialment durant el període càlid.
En aquest tipus de sistemes fluvials es diferencien dues zones. La zona superior o ritron té un fort pendent, substrat de còdols i flux turbulent, i correspon als trams de capçalera de les conques que drenen sistemes muntanyosos de diversa altitud. La resta de la conca o potamon, amb poc o cap pendent, substrat de grava i arena i flux laminar, comprèn la part del riu que circula per la plana fins a desembocar a la mar o en cubetes internes. La importància relativa d’un o altre tram depèn de l’estructura geomorfològica de la conca, i en l’àrea bioclimàtica mediterrània hi ha tota mena de situacions intermèdies.
Solen ser els trams de potamon, amb la seva plana al·luvial associada, els que acusen i registren de manera més dràstica els efectes de les avingudes i les riuades. Fins i tot en els rius que disposen d’àmplies conques de recepció a muntanya, i que tenen el cabal d’aigua assegurat durant l’estació seca, el règim estacional és marcat, i fa que a les parts baixes es produeixin inundacions periòdiques, que fertilitzen o arrasen, segons la magnitud de l’avinguda, les terres de conreu adjacents. A les planes al·luvials complexes com la del Murray, a Austràlia, es formen els “billabongs”, llacunes laterals al riu que, ja siguin meandres abandonats, llacunes d’inundació per efecte de les crescudes, o llacunes associades a la capa freàtica superficial, etc., reben en l’època de pluja l’embat de l’aigua, d’una força i un cabal imprevisibles.
La composició natural de l’aigua depèn de la seva mineralització, la qual al seu torn depèn de la composició de l’aigua de pluja (diferent a cada àrea geogràfica), del transport de sals per l’aire, de l’aflorament d’aigües subterrànies i de la dissolució de les roques que constitueixen el substrat de la conca de drenatge. La quantitat de sals és més gran si el substrat és calcari o evaporític. Les conques mediterrànies australianes tenen els valors de salinitat més elevats (de 500 a 3-000 mg de sals per litre d’aigua), exceptuant la capçalera del Murray, que és més dolça que la resta de rius mediterranis d’Austràlia. Als rius de la província australiana de Victòria, molt curts, els valors de salinitat depenen de la composició de l’aigua de pluja, que és idèntica a la marina, mentre que a gran part de la conca del Murray la salinitat és aportada pels rius salats que venen de la zona del “mallee”. A les restants àrees mediterrànies la salinitat mitjana de l’aigua és de 120 mg/l, clarament inferior a l’australiana. El grau de salinització que es manté a les conques lacustres tancades depèn força de la taxa de renovació de l’aigua. Per això, allà on l’evolució geològica del paisatge i les condicions climàtiques determinen la formació de grans extensions de terres desproveïdes d’un sistema de drenatge, les aigües d’escorrentia no troben el camí per desguassar a la mar i s’hi acumulen formant estanys i tolls atalassohalins, sovint molt soms, que no han estat mai connectats amb la mar; si l’evaporació no supera la precipitació, la inundació es manté tot l’any, però és freqüent que aquestes àrees endorreiques estiguin sotmeses a un cicle d’inundació i assecament anual, o de més d’un any (per exemple, la llacuna de Gallocanta, a la Península Ibèrica).
En àrees de relleu càrstic de la conca mediterrània la dissolució de la roca calcària estableix sistemes de cavitats ampliades i excavades contínuament per l’acció de l’aigua, que conseqüentment es carrega de sals. En aquests sistemes es poden formar estanys per enfonsament de dolines, en les quals hi ha una renovació constant de l’aigua i no s’arriba a valors elevats de salinitat. Als llacs càrstics poden haver-hi moviments tectònics, que en alguns casos provoquen l’enfonsament periòdic del fons, com és el cas dels llacs més grans i profunds de la conca mediterrània, per exemple la mar Morta, la qual, a causa tant del balanç evaporació-precipitació com de la composició del substrat, té una salinitat molt alta. Entorn de la conca mediterrània les fonts càrstiques tenen una gran importància, i són en molts casos l’origen de rius ressurgents, que sovint surten a la superfície amb una pressió artesiana considerable. Els seus cabals són, en més o menys grau, independents de les pluges, i en canvi més dependents de la potència i l’extensió del carst al qual estan associats (per exemple, la capçalera del Jordà o la del Llobregat). Aquestes aigües són interessants perquè constitueixen el refugi dels darrers supervivents de la biota originada a l’antiga mar de Tetis.
Si bé la quantitat d’aigua i la seva distribució temporal són, en bona part, conseqüència del clima mediterrani, també depenen dels sistemes terrestres als quals estan acoblats: tota l’aigua que reben els rius, excepte la que els arriba directament de la pluja, prové dels sistemes terrestres de la conca hidrogràfica per on han circulat anteriorment. La distribució i l’extensió de les masses forestals, la seva composició florística, els ritmes d’entrada de matèria orgànica procedent de la conca i l’existència i la importància de la vegetació de ribera tenen implicacions importants en la composició de les aigües, en la seva dinàmica i en la biologia dels seus ecosistemes. Els boscos mediterranis d’Eucalyptus condicionen el color i el grau d’acidificació de les aigües continentals australianes, fet que separa les característiques limnològiques d’Austràlia dels altres continents; per exemple, determinen la coloració terrosa fosca típica dels compostos fenòlics procedents de la descomposició de les fulles, l’escorça i els fruits, o l’acidesa més gran de les aigües (amb un pH de 5,3 a 6,5). A les àrees mediterrànies restants dominen les aigües bàsiques (valors de pH superiors a 7,8) i clares.
D’altra banda, la vegetació de la conca de drenatge incideix, no només en l’autotròfia i heterotròfia dels sistemes fluvials, sinó també en les aportacions de matèria orgànica procedents dels sistemes terrestres. En general, els rius que circulen entre masses forestades solen ser fonamentalment heterotròfics, és a dir, depenen de l’entrada de matèria orgànica procedents dels boscos, no tan sols per la importància de l’aportació en ella mateixa, sinó per l’efecte de para-sol que constitueix el cobriment total o parcial del llit per la vegetació terrestre. Si el riu és ample, l’efecte de para-sol desapareix i la producció de les algues i els macròfits aquàtics pot equilibrar o superar la importància de les entrades de matèria orgànica d’origen terrestre. En aquest cas el riu es comporta com a un sistema autotròfic. Als sistemes mediterranis, les zones de “ritron” són heterotròfiques i les de “potamon” autotròfiques, en trobar-se les àrees forestades, originàriament, als trams de muntanya. Constitueixen l’excepció els rius mediterranis xilens que, en néixer a altituds superiors als 2 500 m, no tenen capçaleres forestades. Una altra excepció és la conca australiana del Murray, amb àrees forestades a la part central de la plana al·luvial. En els sistemes mediterranis es pot parlar, fins i tot, d’exportació de la producció primària o vegetal fluvial cap als sistemes terrestres. De vegades les masses de vegetació aquàtica queden en sec i són aprofitades per diversos organismes durant el període de sequera en rius, llacunes, “billabongs” temporals, etc. En altres ocasions les masses vegetals aquàtiques són dispersades per l’avinguda i dipositades fora del sistema aquàtic.
Exceptuant el cas d’Austràlia, a les àrees mediterrànies durant la tardor hi ha una entrada màxima de matèria orgànica procedent dels boscos caducifolis, coincidint amb la circulació dels cabals màxims. Aquests permeten un transport i una redistribució ràpida de la matèria introduïda al sistema, com també la descomposició del seu component dominant (les fulles, que representen més del 60% del material) en períodes curts (2 o 3 setmanes en sistemes de la Península Ibèrica). Així s’impedeix el desenvolupament i l’assentament en aquests sistemes d’organismes estrictament trituradors. A les àrees mediterrànies australianes, tanmateix, la màxima entrada de materials procedents dels boscos d’eucaliptus es produeix a l’estiu (moment en què el cabal dels rius és menor) o fins i tot quan els cursos d’aigua temporals estan secs. Ambdues situacions produeixen, a causa dels diferents processos que pateixen, un retard en la disponibilitat de la matèria orgànica per al consum dels organismes trituradors.
La biota de les aigües continentals mediterrànies comparteix unes mateixes estratègies adaptatives, ja que ha convergit a viure en condicions semblants en retalls de continent separats d’arreu del món. Però, igual que passa amb la flora i la fauna terrestres, no comparteix les mateixes espècies, i moltes vegades ni tan sols els mateixos grups. Grans grups d’organismes que són cabdals a l’hemisferi nord, com ara els salmònids o les perles de les famílies dels pèrlids i els perlòdids, són substituïts per altres famílies a l’hemisferi sud o a l’àrea austral, per exemple.
Efecte de l’activitat humana
La intervenció humana sobre els sistemes aquàtics naturals és molt antiga i està àmpliament estesa a l’àrea mediterrània. Moltes de les antigues civilitzacions van sorgir al voltant de les valls àrides i semiàrides mediterrànies, aptes per a l’activitat agrícola. La necessitat de construir rescloses per regar i canalitzacions per distribuir l’aigua, tant de consum urbà com agrícola, complementades amb aqüeductes a l’època romana, com també la recollida i la canalització de les aigües residuals a les poblacions importants, i fins i tot les rescloses i els aprofitaments d’ús industrial, han originat una cultura de l’ús de l’aigua que tendeix al seu màxim reaprofitament i al desenvolupament d’una enginyeria hidràulica important. Les conseqüències actuals són una àmplia xarxa d’embassaments construïda als sistemes fluvials mediterranis, tant per emmagatzemar aigua com per preveure les avingudes mitjançant la regulació dels cabals. Entre les obres de prevenció d’avingudes, les encaminades a convertir els rius en canals rectes amb marges desforestats són especialment destructives, ja que originen una erosió més gran, i potencien els efectes destructius aigües avall d’aquests trams. El gran assentament urbà de les àrees mediterrànies, conseqüència de la benignitat del clima, amb grans nuclis de població a tots els continents, com també la transformació en regadiu de grans extensions de boscos i matollars arbustius, han transformat profundament les regions mediterrànies, de manera que es fa difícil reconèixer-hi el paisatge originari. Hi ha hagut grans processos erosius i, fins i tot, de desertització (especialment a la conca mediterrània), juntament amb abocaments de residus contaminants d’origen urbà, agrícola i industrial. El resultat global és un grau elevat de transformació de gran part dels sistemes aquàtics continentals mediterranis: rius estancats i contaminats, llits alterats, llacs i embassaments eutròfics, salinització dels cursos d’aigua i dels mantells freàtics superficials a causa de la irrigació, etc.
La introducció d’espècies de peixos forànies als ambients aquàtics mediterranis, amb finalitats esportives o comercials, facilitada per la semblança climàtica entre les diferents porcions mediterrànies del món, és una de les causes de l’empobriment d’espècies dels rius mediterranis. La carpa (Cyprinus carpio), procedent del Pròxim Orient, es va estendre per tota la conca mediterrània i actualment s’ha introduït a Califòrnia, Xile i Austràlia. Altres espècies que comparteixen aquesta distribució artificial són la truita comuna (Salmo trutta) i la gambúsia (Gambusia holbrooki). El grau d’alteració de les comunitats de peixos és molt variable segons el país i la conca. Per exemple, a la Península Ibèrica la proporció d’espècies forànies és del 36%, però a Catalunya arriba al 47%.
Els ambients aquàtics mediterranis
Viure en perill en rius, torrents i rambles
El règim hídric dels rius mediterranis, amb èpoques de sequera i períodes de cabal màxim associats a avingudes i fins i tot a riuades, determina una gran diversitat de condicions a les quals s’han d’adaptar les espècies que hi viuen. Als rius i trams permanents, la fauna d’insectes és dominada bàsicament per espècies que tenen més d’una generació a l’any. Molts tricòpters (per exemple els petits hidroptílids brostejadors) arriben a tenir quatre generacions, bé que, a causa de diferents toleràncies tèrmiques, algunes espècies tenen un únic cicle vital, centrat a l’hivern o bé a l’estiu (baètids d’aigües fredes, o efemerèl·lids d’aigües càlides). Hi ha dues comunitats d’insectes aquàtics ben definides, que s’alternen al llarg de l’any. L’una apareix amb les inundacions tardorals o hivernals, segons les diferents àrees geogràfiques, i hi roman fins a mitjan o final de primavera, quan dominen les espècies reòfiles dins les comunitats del zoobentos. L’altra és estival i es caracteritza perquè la major part de les seves espècies tenen els cicles vitals sincronitzats de tal manera que a l’inici del període d’inundació es troben en fase aèria fora del sistema aquàtic, o bé en fase de latència (ous durables). En alguns grups, com en molts coleòpters i heteròpters, els adults —els únics presents als cursos d’aigua a l’inici del període d’inundació— poden abandonar els sistemes fluvials i cercar refugi en altres tipus de masses d’aigua, o bé introduir-se als sediments de la llera, com ho fan, per exemple, els coleòpters de la família dels èlmids o les efímeres durant els primers estadis del desenvolupament. Als rius mediterranis el nou cicle hídric comença amb la inundació del sistema fluvial i l’elevació del nivell de l’aigua dels rius, i desencadena l’aparició de la comunitat hivernal d’insectes. Als rius de la Península Ibèrica calen de 20 a 25 dies per trobar la major part d’espècies que formaran la comunitat hivernal.
També les comunitats de peixos es caracteritzen per la seva capacitat d’adaptar-se a la variabilitat ambiental d’aquests rius, i per la seva permanència després de les avingudes, riuades i secades. Als rius curts i temporals la composició faunística és molt similar tot al llarg de la costa, excepte a la desembocadura i, a vegades, a la capçalera. A les conques més estructurades i permanents el nombre d’espècies és més gran, i s’observa un cert model de distribució, determinada per factors físics (velocitat del corrent, temperatura de l’aigua i ordre del riu), biològics i biogeogràfics. Durant el període de sequera les espècies es concentren a les basses, on romanen fins que la inundació del llit els permet expandir-se de nou. A l’hemisferi nord el model faunístic és similar a les dues regions amb clima mediterrani. A la conca mediterrània, excepte a les zones de muntanya, les espècies dominants són bàsicament ciprínids (80%), (Barbus, Leuciscus, Chondrostoma, Rutilus, Anaecypris, Iberocypris, Pseudophoxinus, etc.), juntament amb altres grups, com els cobítids. Als rius de la regió mediterrània de Califòrnia també dominen els ciprínids (30%), (Gila, Rhinichthys, Lavinia, etc), i altres grups, com els còtids (28%) (Cottus). A l’hemisferi sud, a Xile, entre les conques del Limari i el Bío-Bío, per raons històriques però també ecològiques, el nombre d’espècies és molt reduït, i entre les típicament d’aigua dolça destaquen els pigídids (Pygidium), els caràcids (Cheirodon), els aterínids (Basilichthys) i els tricomictèrids (Trichomycterus). A Austràlia abunden els galàxids (Galaxias), presents també a Sud-àfrica i a l’extrem d’Amèrica del Sud, els percíctids (Maccullochella, Macquaria) i els kúlids (Nannoperca, Nannatherina). La distribució dels galàxids s’explica, sobretot, per la deriva continental.
Vinculats a aquests ambients hi ha alguns vertebrats que no són exclusivament aquàtics. Per exemple les granotes Rana perezi i R. esculenta, a la conca mediterrània, R. aurora i R. catesbeiana, a Califòrnia, i Heleioporus eyrei, a Austràlia. Entre els rèptils hi ha serps d’aigua, com les dels gèneres Natrix, a la Mediterrània, Diadophis, a Califòrnia, i Austrelaps a Austràlia. També hi ha tortugues d’aigua, com Mauremys caspica i Emys orbicularis, a la conca mediterrània. Certs mamífers exploten també aquests sistemes aquàtics, com per exemple la llúdria (Lutra lutra).
Els rius temporals tenen un model simplificat respecte dels permanents. Els que queden secs durant part de l’any (4 o 5 mesos) són colonitzats per comunitats similars o equivalents a les hivernals dels rius permanents, amb algunes espècies diferents. Si el grau de temporalitat augmenta, dominen les espècies cosmopolites de cicle ràpid, com els dípters (quironòmids, simúlids, ceratopogònids). Si la temporalitat és molt extrema (queden secs deu mesos l’any o més), com a les basses romanents després de les crescudes pròpies de les rambles i als “oueds” de les zones àrides (SE de la Península Ibèrica, N d’Àfrica, Sud-àfrica i la zona de transició entre Califòrnia, Arizona i Mèxic), la comunitat que s’estableix correspon a sistemes lenítics temporals (microcrustacis, mosquits, heteròpters, etc.). De manera semblant, el nombre d’espècies de peixos va disminuint entre les conques moderadament temporals i les rambles, en les quals, com a molt, subsisteixen peixos a les basses que romanen a la desembocadura (ciprinodòntids i Gasterosteus). Així, en general, a les regions mediterrànies la riquesa d’espècies disminueix a mesura que augmenta la durada del període en què el riu és sec. Tanmateix, no hi ha grans diferències entre els rius permanents i els moderadament temporals, que mantenen sistemes de tolls més o menys romanents durant tot el període estival.
Resistir el corrent a les zones de muntanya
Sobre el substrat de roques, blocs i còdols es desenvolupa una variada comunitat de microalgues del “ritron” dominada fonamentalment per diatomees i molses capaces de resistir una dessecació parcial. La matèria orgànica que prové dels boscos es descompon ràpidament i és transportada en forma de matèria particulada fina en suspensió o queda retinguda a les àrees de sedimentació dels tolls que s’alternen amb els salts d’aigua.
En aquestes àrees dominen els filtradors, com els simúlids i els tricòpters (de les famílies dels hidropsíquids, helicopsíquids i leptocèrids), que poden interceptar aquestes petites partícules mitjançant la fabricació de xarxes o mitjançant la utilització d’apèndixs amb pintes de sedes (simúlids i leptocèrids). També hi abunden els fitodetritòfags, un grup que alterna el consum de microalgues amb el de partícules retingudes o dipositades als sediments; en són un exemple les efímeres de les famílies dels caènids i els efemerèl·lids, els coleòpters helòdids i la majoria de quironòmids (Orthocladinae).
Altres organismes característics d’aquests trams són els brostejadors especialment adaptats als corrents, els efemeròpters de cos aplanat (heptagènids a l’hemisferi nord i a Sud-àfrica, i leptoflèbids a Austràlia i a Xile), els mol·luscos (Ancylastrum) i algunes espècies de petits coleòpters de la família dels èlmids, tricòpters glossosomàtids i els del gènere Baetis, capaços de suportar corrents intensos. Entre els depredadors dominen els invertebrats caminadors com els grans plecòpters (Marthamea i Eoperla, a la conca mediterrània, diferents gèneres de gripopterígids i eustènids a l’Àmèrica del Sud i Austràlia), o els tricòpters riacofílids. Els peixos d’aquesta zona són els salmònids de l’hemisferi nord: la truita comuna (Salmo trutta), a la conca mediterrània, i la truita arc iris (Salmo gairdneri), Salmo clarki i Cottus, a Califòrnia.
Avingudes i fluència tranquil·la a les planes inferiors
En les zones de “potamon” el riu té poc pendent i la llera és formada totalment o parcialment per substrat tou. L’aigua circula sense turbulència, amb flux laminar que permet el desenvolupament d’herbassars submergits de macròfits (Potamogeton, Myriophyllum), o, a les àrees amb un substrat una mica més groller, de grans masses d’algues filamentoses (Cladophora), que constitueixen el suport d’algues microscòpiques (diatomees) i el refugi de gran part de la microfauna mòbil (cladòcers, copèpodes, ostràcodes, àcars) que s’amaga dels predadors, com ara els odonats (agriònids, hemiflèbids), els heteròpters (notonècnids, gèrrids) i els peixos. En aquestes zones la matèria particulada fina tendeix a dipositar-se, de manera que permet el predomini dels detritòfags, principalment efímeres (caènids, Tricorythodes) i quironòmids (Chironomini). Els filtradors com les efímeres (efemèrids, polimirtàcids) que excaven túnels als substrats tous abunden molt en aquestes àrees. Quan aquests trams tenen àrees forestades properes al riu, com a Austràlia, s’hi troben ben representats, també, els plecòpters trituradors (nemúrids, notonemúrids) i, fins i tot, espècies que participen en la descomposició de restes de troncs i branques (austropèrlids). Aquestes zones del riu constitueixen el paradís dels ciprínids (Barbus, Rutilus, Chondrostoma i Leuciscus, a la Península Ibèrica), que disposen de mecanismes d’alimentació especialitzats, com una mandíbula protràctil i dents faríngies; la majoria s’alimenten de petits insectes aquàtics, crustacis, cucs i detrits en proporcions variables.
Davant d’aquesta estructura de les comunitats, aparentment complexa i molt estructurada, les avingudes i les riuades introdueixen moments de cabals màxims, mobilitzen els substrats i arrenquen part de la vegetació dels herbassars submergits. Els efectes de les riuades sobre la fauna d’aquests sistemes només es coneixen parcialment, però sembla que estan associats a la predicibilitat dels diferents esdeveniments, de manera que les avingudes, esperades anualment, no hi destrueixen l’estructura de les comunitats. Tanmateix, calen entre tres mesos i un any perquè es recuperin les comunitats d’insectes després d’una riuada, i encara més temps perquè es reestructurin les comunitats de peixos.
Tolls romanents, refugi en època seca
La sequera estival pròpia d’aquest clima marca el grau de permanència de gran part dels cursos fluvials mediterranis, fins al punt que la majoria tenen trams temporalment intermitents. En aquests trams, fonamentalment associats a zones baixes de rius curts, el riu es transforma en un rosari de basses més o menys romanents. Aquí s’hi refugien els organismes que no tenen un altre tipus d’estratègia per resistir el període sec, com per exemple els peixos (que poden arribar a concentracions de 25 individus per metre quadrat), o bé espècies amb un cicle estival que acaba en aquests tolls romanents. Les basses són visitades esporàdicament per organismes voladors (coleòpters adults, heteròpters) provinents de sistemes lenítics, basses artificials, llacunes de plana al·luvial (“billabongs”), llacs i embassaments.
Llacunes de plana al·luvial o “billabongs”
Els “billabongs” australians són basses d’aigües més o menys permanents associades a un curs fluvial, que hi entren en connexió durant la crescuda de l’època de pluges. Són el paradís de la vegetació aquàtica i permeten especialment el creixement de plantes flotants (Riccia, Lemna), algunes de les quals (Azolla) s’afavoreixen quan hi ha esgotament dels nutrients dissolts. Les comunitats animals són molt variades i diferents de les del curs principal que circula al seu costat (280 espècies presents als “billabongs” i només 112 al riu). Quan el cabal del riu torna a disminuir, part de les espècies del “billabong” ja han colonitzat el curs principal i desplacen les espècies originàries, com ho ha fet, per exemple, el bivalve Velesunio ambiguus, que ha desplaçat Alathyria jacksoni. A Austràlia, els “billabongs” s’han transformat en un paradís també per a les espècies introduïdes, que poden constituir plagues, com l’hepàtica Riccia o el jacint d’aigua (Eichornia). Les espècies forànies de peixos (Salmo, Gambusia, i altres) poden arribar a eliminar les autòctones, sigui a causa de la competència o de la depredació.
Sal i calor variables en les llacunes somes interiors
Les llacunes salabroses tenen una dinàmica anual particular, ja que al cicle hidrològic d’alternança d’un període d’escassetat d’aigua i un període de reompliment per l’abundància de pluges, se superposa un cicle molt acusat de concentració i dilució de sals: en les èpoques d’estiatge, la forta evaporació fa que la concentració de sals en l’aigua augmenti, fins al punt d’atènyer valors de salinitat que poden ser dues i tres vegades més alts que la salinitat mitjana marina (34 a 35‰). L’alta insolació sobre la cubeta inundada encara té un altre efecte important per als organismes: el de la temperatura, perquè l’aigua, que hi fa una capa soma damunt el sediment, s’escalfa progressivament, sovint fins als 40°C o més. Les condicions per a la vida hi són, òbviament, molt especials.
A l’estiu, és fàcil que aquestes llacunes s’arribin a assecar totalment, de manera que la cubeta que allotjava l’aigua queda convertida en una extensa plana coberta d’una crosta de sals. Aquest tipus de llacunes atalassohalines d’inundació temporal són abundants en l’àrea mediterrània, mentre que a les terres australianes són més abundants les d’inundació permanent.
La salinitat de les aigües atalassohalines pot ser deguda al predomini d’uns tipus d’ions sobre uns altres (carbonats, clorurs i sulfats de sodi, potassi, calci i magnesi, i combinacions), predomini que mostra una distribució de tipus regional: els llacs salats australians solen tenir un predomini de clorurs, mentre que als de la cubeta mediterrània (per exemple, a la conca del Jordà) solen predominar les aigües carbonatades i fortament alcalines. Els valors de pH solen ser-hi alts: poden oscil·lar entre 7,5 i 11. A l’estiu, el contingut salí de l’aigua queda enregistrat en el sediment, on precipiten diferents sals segons les proporcions iòniques que aquella contenia (aragonit, guix, dolomita, mirabilita, epsomita, halita, etc.). Els materials evaporítics es dissolen periòdicament i es tornen a dipositar, la qual cosa fa que es diferenciï una zonació concèntrica de minerals menys solubles a més solubles, des del litoral cap al centre de la cubeta. El sediment sec i recobert d’una crosta dura de sals guarda tota la sembra de la llavor que ha de reconstruir les comunitats.
La vida en les llacunes salabroses mediterrànies obliga a estratègies que permetin superar els pronunciats canvis de salinitat i temperatura que es produeixen al llarg del cicle anual. En general, les comunitats són fetes de poques espècies, però en canvi pot desenvolupar-s’hi una successió al llarg de l’any d’organismes cada vegada més ben adaptats a la forta salinitat i l’alta temperatura. És característica la formació de tapissos algals i microbians (bacteris heteròtrofs i autòtrofs) recobrint el sediment, sobretot en les aigües somes del litoral, com també els extensos pradells de carofícies que cobreixen enterament la cubeta, especialment de Lamprothamnium papulosum i Chara galioides, sovint acompanyats de Ruppia, un gènere de fanerògames capaç igualment de suportar salinitats superiors a la del mar. Quan s’acosta el període de secada, les carofícies produeixen oòspores abundants, i deixen una sembra en el sediment capaç de recolonitzar la cubeta en la pròxima inundació, mentre que Ruppia deixa òrgans de resistència en les arrels que queden enterrades al sediment. Del plàncton destaquen les espècies eurihalines (Nannochloris, Dunaliella salina en el fitoplàncton, i entre el zooplàncton, el ciliat Fabrea salina, rotífers com Brachionus plicatilis i els gèneres Hexarthra, Asplanchna, Keratella, diversos cladòcers i copèpodes, però també espècies que, contràriament a aquelles, són capaces de suportar àmplies variacions de salinitat, moltes vegades cosmopolites (Brachionus plicatilis, Cyclops viridis). Un cas extrem de salinització és el de la mar Morta, que té una salinitat de 280 ‰ (contra els 34-35 ‰ de l’aigua marina!) i una fondària de 320 m, d’aigües fortament alcalines i amb continguts de calci, magnesi, potassi i brom molt elevats. Malgrat el nom, és l’hàbitat predilecte de Dunaliella salina i de bacteris, aerobis a les capes superiors i anaerobis al fons, i halòfils. Gràcies a la fondària i als alts valors de salinitat, que afecten la densitat de l’aigua, en aquest llac no es forma una termoclina anual com passa generalment en els llacs temperats, sinó que se succeeixen cicles més llargs, i l’època d’estratificació pot durar diversos anys.
Les aigües salades relacionades amb estuaris i aiguamolls litorals són explotades per la gambúsia (Gambusia affinis), espècie que ha estat introduïda en tots els continents; en els llacs australians, aquesta espècie representa un greu perill per a les espècies endèmiques d’hàbitat equivalent, principalment el galàxid Galaxias maculatus i Atherinosoma microstoma. Aquestes dues espècies són capaces de suportar alts valors de salinitat i de temperatura, com també ho és l’isòpode endèmic Haloniscus searlei, i la larva del quironòmid Tanytarsus barbitarsis. Els anostracis són un element típic dels sistemes atalassohalins costaners: Artemia salina, cosmopolita, es troba a tots els continents excepte a Austràlia, on és substituïda per Parartemia.
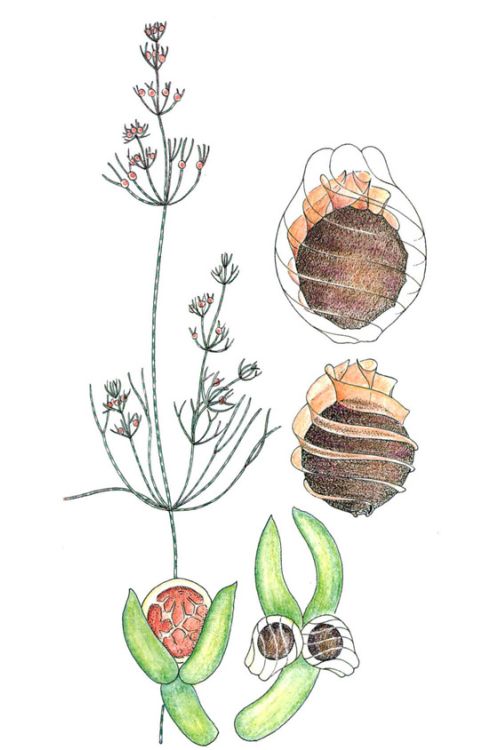
L’aparell vegetatiu de Nitella opaca , organitzat en nusos i entrenusos, pot arribar a fer 1 m de llargada, quan les aigües són fondes i permanents, mentre que fa petites gespes arrapades al llot en les basses d’inundació temporal. Es tracta d’una espècie d’àmplia distribució i de desenvolupament vernal, que completa el cicle en pocs mesos i deixa una sembrada d’oòspores resistents capaç de reconstruir la població en pocs dies quan arriben les pluges primaverals. La seva sensibilitat a les concentracions altes de fòsfor en l’aigua, com les provocades pels detergents, fa que els caròfits siguin indicadors de l’estat de salut dels sistemes aquàtics.
Montserrat Comelles / Miquel Alonso / ECSA
Les basses temporals de mineralització moderada i variable al llarg de l’any tenen més riquesa d’espècies. És fàcil que s’hi desenvolupi una zonació de vegetació, en àrees concèntriques, al voltant de la cubeta. El cinturó exterior d’helòfits (Phragmites, Juncus) és seguit cap a l’interior per plantes semiaquàtiques, que arrelen al fang però fan fulles flotants: Ranunculus pot fer-hi una florida abundant, que adorna la superfície de l’aigua amb un cercle de flors blanques. Als primers estadis d’inundació solen formar-s’hi pradells de carofícies pioneres (del gènere Tolypella en aigües força mineralitzades, Nitella en aigües més pures), que més tard són substituïts per altres espècies de caròfits (Chara) o plantes superiors enterament aquàtiques, com ara (Zannichellia palustris, Potamogeton pectinatus), ambdues capaces de suportar valors de salinitat força elevats.
Aigües quietes i fondes: els estanys càrstics
Els estanys càrstics són, en el món mediterrani, els únics sistemes limnètics naturals on l’aigua té un temps de residència suficient per a arribar a assolir una organització ecològica pròpia. Les aigües són força mineralitzades, i contenen quantitats elevades de carbonats i bicarbonats a causa de la naturalesa soluble dels materials del substrat. L’abundància de minerals finament particulats en suspensió en limita força la transparència.
El fitoplàncton dels llacs càrstics mediterranis mostra una successió anual acusada, ja que el règim alternat d’estratificació i barreja de les aigües fa que dues vegades l’any es creï una situació nova, que requereix la colonització d’espècies pioneres de ràpida reproducció. Es compon de diatomees (Cyclotella), crisofícies (Dinobryon), criptofícies (Rhodomonas, Cryptomonas) i dinoflagel·lades (Ceratium hirundinella, Peridinium) i algunes clorofícies (Ankistrodesmus). La producció primària, però, sol ser-hi baixa, per dèficit de nutrients, ja que l’abundància de calci fa que el fòsfor lliure sigui escàs. El zooplàncton que s’hi estableix sol ser dominat pels copèpodes per sobre dels rotífers i els cladòcers, tots de cicle de vida més llarg, sovint anual, i amb una alimentació diferent en les diferents etapes vitals. El bentos profund és pobre, ja que els sediments del fons no són un sistema estable a causa de l’entrada subterrània d’aigua, que els manté en suspensió. La representació bentònica és més rica al litoral, on viuen quironòmids, nematodes i bivalves (Unio a Europa; Velesunio a Austràlia). També al litoral pot fer-s’hi una comunitat macrofítica ben desenvolupada, especialment rica en caròfits (Chara, Tolypella, Nitella), però també fanerògames arrelades (Myriophyllum, Potamogeton). Són característics de les aigües alcalines dels sistemes càrstics els crustacis isòpodes (Gammarus a l’hemisferi N; Hyalella a l’hemisferi S i el freatoicideu Metaphreatoicus a Austràlia, en hàbitats meridionals equivalents). També destaquen els crustacis nedadors de desenvolupament larval planctònic (Atyaephyra, Dugastella), que d’adults viuen a la zona litoral.
Aigües ni quietes ni corrents dels embassaments
Els embassaments construïts pels humans han introduït en la zona mediterrània un hàbitat aquàtic que no li correspon de manera natural: les grans cubetes inundades permanentment d’aigües vingudes de muntanya, les quals són, a més, sotmeses a un règim hidrològic capriciós, que respon a les necessitats d’aigua de les poblacions humanes aigües avall. Tot i això, malgrat aquesta dinàmica hidrològica, els embassaments solen comportar-se com a llacs temperats quan la descàrrega d’aigües es fa per les capes superiors, ja que res no impedeix que en les parts més profundes la columna d’aigua continuï comportant-se com un llac natural, i s’hi desenvolupi una termoclina anual; a les vores, en canvi, sotmeses a canvis de nivell no climàtics, s’hi impedeix el desenvolupament normal dels sistemes litorals. L’efecte de la mà humana és molt més important a l’escala de la biologia dels sediments, que regeix en definitiva la biologia de tota la cubeta, quan hi ha un buidament periòdic dels fangs del fons. Aquesta acció no sols desorganitza completament l’activitat biològica i química de l’embassament, sinó també la del riu aigües avall, ja que difícilment pot assimilar la descàrrega de fangs fortament reduïts que s’acumulen per sota de la termoclina.
El temps de renovació de l’aigua en l’embassament té molta influència en la composició de les comunitats aquàtiques: com més de pressa es renova l’aigua, el sistema s’assembla més a un riu; com més a poc a poc, més s’assembla a un llac.
En les parts profundes, més properes a la presa, els embassaments es comporten com a sistemes lacustres quant al seu cicle tèrmic. A la cubeta mediterrània, els embassaments s’estratifiquen durant l’estiu. És a dir: les capes superiors d’aigua (epilimni) s’escalfen gràcies a l’intercanvi de calor amb l’atmosfera, mentre que les inferiors (hipolimni) es mantenen fredes; ambdues capes queden separades per una banda de canvi brusc de temperatura, la termoclina, tot i que la profunditat a la qual es forma és molt influïda per l’ús que els humans fan de l’embassament. La quantitat d’oxigen dissolt a l’aigua varia al llarg de l’any, principalment amb relació als processos biològics (respiració i fotosíntesi) i a l’estratificació estival: quan l’aigua està barrejada, hi ha una bona oxigenació en totes les fondàries; en canvi, en condicions d’estratificació, la capa superior manté una bona concentració d’oxigen perquè es manté en contacte amb l’aire i conté la major part dels productors primaris (algues del fitoplàncton), mentre que la inferior rep majoritàriament els efectes de la respiració dels organismes i oxidació de la matèria orgànica, i per això pot arribar a ser deficitària d’oxigen; s’arriba fàcilment a l’esgotament d’oxigen quan les aigües són eutròfiques. El grau d’eutròfia dels embassaments, conseqüència de la seva riquesa de nutrients, fa que s’hi desenvolupin unes o altres comunitats planctòniques, tant vegetals com animals. En el fitoplàncton, partint de les comunitats bàsiques de les aigües amb pocs nutrients, l’eutròfia fa que s’enriqueixi d’altres espècies, i quan arriba a valors alts, l’alteració de les proporcions normals de fòsfor i nitrogen fa que s’estableixi un predomini dels cianobacteris. En el zooplàncton, l’eutròfia porta associada una dominància de cladòcers i rotífers, de cicle de vida curt (sovint d’uns dies) per sobre dels copèpodes; ambdós grups mostren fenòmens pronunciats de ciclomorfosi amb relació a la temperatura estival de l’aigua. Els rotífers dominen completament en condicions de forta eutròfia. En el fons de la cubeta, amb relació als sediments, s’hi fan comunitats bentòniques habitants del fang, on fan galeries o tubs, o bé es mouen per la superfície dels sediments. Aquí és on viuen organismes capaços de resistir concentracions d’oxigen ben baixes; els tubifícids (Tubifex) i les larves de quironòmids (Chironomus) són els més abundants; els primers viuen d’explotar els detrits vinguts del litoral, mentre que els segons s’alimenten sobretot del fitoplàncton que cau al fons contínuament. Larves de caobòrids (Chaoborus) i alguns mol·luscs semblen estar-hi també adaptats. A les zones somes del litoral, molt fluctuants de nivell, s’hi fan comunitats d’organismes semblants a les de les basses d’aigua dolça, tots de cicle curt, poc exigents en el menjar i amb una producció nombrosa de descendents. La mateixa variació del nivell de l’aigua impedeix normalment que creixi un cinyell litoral d’helòfits; i quan l’estabilitat ho permet, en els rabeigs tranquils, s’hi fan comunitats de fanerògames aquàtiques (Potamogeton, Myriophyllum). Els peixos que hi viuen són d’espècies pròpies de sistemes fluvials i majoritàriament d’espècies introduïdes.
La vegetació de ribera

Vegetació de ribera a la Tet, al seu pas per Vilafranca de Conflent.
MC
Els terrenys que voregen els rius mediterranis són llocs privilegiats quant a l’abundància d’aigua, ja que, per més sec que sigui el paisatge, a la vora del curs d’aigua la secada s’amoroseix. Es formen en el paisatge bandes de vegetació, les anomenades formacions en galeria, que ressegueixen els cursos d’aigua, tot donant lloc a paisatges azonals, o sigui, més determinats per les condicions ecològiques locals que no pas per les climàtiques generals. La composició específica i l’aspecte fisiognòmic d’aquests paisatges de ribera són fortament condicionats per la fondària, l’abundor i la constància del mantell freàtic, però també per la magnitud i la recurrència de les crescudes, ja que tots dos factors determinen les característiques dels vegetals que hi viuen. Sota règim mediterrani trobem des de rius permanents fins a cursos d’aigua que només en porten durant períodes plujosos; els primers presenten un règim més regular que no els segons; les crescudes fortes obliguen els vegetals de ribera a fer front alternativament a la manca d’aigua i a la seva força destructiva.
Els boscos dels cursos permanents
Els rius mediterranis més cabalosos porten associada una franja de vegetació forestal dominada per caducifolis hivernals propis d’altres biomes, que desplacen els perennifolis en condicions d’absència de sequedat estival. Aquestes comunitats són boscos purs o mixtos, com ara les vernedes, dominades pel vern (Alnus glutinosa), les alberedes dominades per l’àlber (Populus alba), les omedes dominades per l’om (Ulmus minor), les salzedes, dominades per les diverses espècies de salze (Salix) i les platanedes, per plàtans (Platanus orientalis), o per les nogueres (Juglans regia) de la conca mediterrània.
En la regió californiana també trobem plàtans (Platanus racemosa) i algunes restes de les antigament abundants i esponeroses rouredes de ribera, formades pel roure anomenat “valley oak” (Quercus lobata), actualment poc abundants a causa de les tales fetes per aprofitar la fusta d’aquests roures, de la roturació dels fons de vall per implantar-hi hortes, de la davallada de la capa freàtica per l’explotació intesa dels aqüífers i de la urbanització de les parts baixes del territori. A la regió sud-africana, mancada d’arbres, trobem comunitats arbustives dominades per caducifolis. A la zona mediterrània xilena, hi ha també bosquetons de caducifolis que ressegueixen els cursos d’aigua, en els quals dominen el “sauce chileno” (Salix humboldtiana) i el “culén” (Psoralea glandulosa), acompanyats d’arbrets perennifolis com el laurifoli típic dels boscos subantàrtics anomenat “canelo” (Drimys winteri), el “chequén” (Luma chequen) o el “patagua” (Crinodendron patagua). En el cas australià trobem boscos de ribera perennifolis dominats per espècies d’eucaliptus com Eucalyptus camaldulensis, que a Austràlia Meridional sol estar acompanyat d’E. largiflorens, mentre que a la regió sud-occidental comparteix l’hàbitat amb el “yarri” (E. patens) i E. rudis.
La majoria d’aquests arbres i arbustos tenen capacitat de rebrotar de soca, cosa que representa una resposta adaptativa a la possible destrucció de les parts aèries per crescudes violentes. En zones prou nitrificades o riques de clorurs, ja sigui a causa de la natura dels sòls o de la proximitat a la mar en el cas de rius de baix cabal, els caducifolis deixen pas a arbrets perennifolis halòfits, com els tamarius (Tamarix gallica i T. africana) de la conca mediterrània.
Els bosquetons dels cursos intermitents
En les regions mediterrànies més àrides, dominades per caducifolis estivals o per comunitats arbustives perennifòlies molt eixarreïdes, la vegetació de ribera també s’empobreix, per tal com ha de suportar una relativa secada durant l’estiatge. Trobem en aquests casos formacions arbustives de ribera dominades per arbustos perennifolis o per arbres de mida petita.
A la conca mediterrània, al mig de la llera, als llocs on en profunditat sempre queda una mica d’aigua romanent, solen prosperar-hi salzes arbustius, com ara la sarga (Salix elaeagnos). A les vores, o en plena rambla i sota condicions d’intermitència més severa, predominen els baladres (Nerium oleander) i fins els alocs (Vitex agnus-castus) o les murtes (Myrtus communis). El caràcter perennifoli d’aquestes espècies ja denuncia l’escassa disponibilitat hídrica.
La nitrofília ripària
Quan els cursos d’aigua s’assequen periòdicament, o quan només porten aigua durant les èpoques plujoses, la mateixa llera del riu és colonitzada per vegetació zonal pròpia dels vessants. De fet, aquestes lleres són sistemes terrestres periòdicament devastats per riuades violentes i curtes que gairebé no donen temps que s’hi instal·li una comunitat aquàtica ben desenvolupada. Aquesta vegetació és constituïda per espècies pioneres, de creixement ràpid i cicle biològic curt, que deixen abundant descendència entre riuada i riuada. La majoria són plantes herbàcies com Aster squamatum, o joncs diversos, però hi trobem també alguns arbustos com l’olivarda (Inula viscosa), caducifoli hivernal de florida tardoral típic de les rambles de la conca mediterrània.
Totes aquestes espècies tenen una marcada tolerància davant dels compostos nitrogenats que dipositen les riuades, i fins i tot en demostren apetència. Per això, aquests ambients són el focus de dispersió de moltes espècies nitròfiles, com la mateixa olivarda (Inula viscosa), que després colonitzen àrees discretament humides i nitrificades per acció humana, com ara les vores dels camins. És així com ruderal, nitròfil i ripari s’agermanen.