Els elements de la vida
Els components químics vitals
La vida es basa en estructures materials i és tenaç a l’hora d’augmentar-ne la complexitat. Els sistemes vius han aconseguit una millora notable en guanyar organització en canvi de les despeses d’energia irrecuperable. Sembla que la gran riquesa d’estructura làbil, pròpia de la vida, no hauria d’anar lligada necessàriament a una naturalesa material molt precisa. Però, de fet, els elements químics possibles i a l’abast marcaren el camí. Certs tipus d’àtoms entraren més fàcilment que altres en un procés de complicació material. Com que no coneixem la vida fora de la Terra no sabem (i potser no ho sabrem mai) si s’hauria pogut prendre un altre punt de partida utilitzant materials més o menys diferents.
Sis elements (carboni, hidrogen, nitrogen, oxigen, fòsfor i sofre) representen més del 99% del pes de les cèl·lules vives. Els àtoms predominants són de nombre atòmic i pes atòmic baix.
El substrat material dels fenòmens vitals conté molta aigua i carboni. En estat líquid, l’aigua és un suport excel·lent. La seva estructura molecular és extraordinària ja que és feta de molècules polars, que li donen propietats importants, entre altres la de convertir-la en un dissolvent molt efectiu. A més, l’aigua forma grups de molècules elementals aproximades, de manera que, i contra el que se’n podia esperar, és un líquid de densitat relativament alta. La molècula d’aigua fa poc més de 2 àngstroms (1 àngstrom (Å) = 0,1 nm), mida que ens pot servir per a tenir una idea del grau de miniaturització en la constitució de la vida.
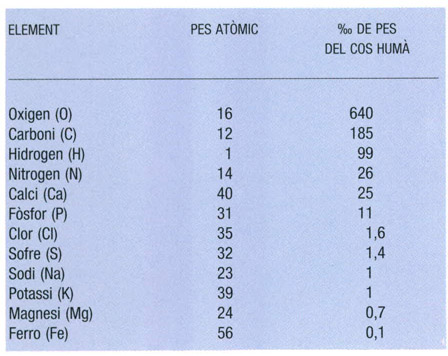
Principals elements químics components del cos humà. La llista, a la qual caldria afegir quantitats menors d’alumini, arsènic, bor, brom, coure, fluor, iode, manganès, silici i zinc, expressa en tants per mil de pes la composició elemental del cos humà. L’exemple s’ha triat com a representatiu de la matèria viva; i els pesos atòmics són els dels isòtops predominants de cada element.
Dades elaborades per l’autor
Entorn de les 3/4 parts de la matèria viva poden ésser aigua. A la resta és aplicable la proporció de carboni, nitrogen i fòsfor aproximada proposada pel biòleg nord-americà Alfred C. Redfield (1890-1983), referida inicialment a la composició mitjana del plàncton marí, i que en àtoms és de 106C:16N:1P. Recordant els respectius pesos atòmics (12, 14, 31) dels elements comparats, les proporcions calculades en pes s’apropen a 41:2,6:1. Aquesta relació és solament orientativa, però bona de recordar i útil quan es tracta de veure si la composició de l’entorn és favorable a la vida o per on pot fallar el subministrament. Quan hi ha desviacions es tracta generalment d’una quantitat “excessiva” de carboni, com se sol trobar en la vegetació terrestre, en la qual la fusta té molt de carboni i molt menys de nitrogen i de fòsfor.
El carboni
El carboni total de la biosfera podria situar-se entre mig quilo i un quilo per metre quadrat, sense comptar l’humus o la matèria orgànica morta (necrosfera), però incloent-hi materials que no és fàcil discernir si són vius o no. Es com la reserva de carboni atmosfèric en forma de CO2 que també és entorn d’l kg m-2. (Recordem que el resultat de multiplicar la pressió atmosfèrica [104 bars] per la concentració de CO2 [en volum] pel factor per passar de volum a pes de carboni [igual a 10 197 × 0,00035 × 12/44] és igual a 0,97 kg).
Els àtoms de carboni s’uneixen formant esquelets moleculars molt variats, generalment en forma de cadenes o anells. Recentment s’han descobert en el laboratori altres variants inèdites que afegeixen més possibilitats d’estructura: com a pilotes de futbol, o bé com a tubs de llargada indefinida. La distància entre els centres de cada àtom consecutiu és de l’ordre de 0,1 nm. Entorn d’una desena part del pes total i aproximadament el 40% del pes de la matèria seca dels organismes és carboni. Les cadenes o anell fets d’àtoms de carboni, estables i persistents, tenen valències lliures que s’articulen amb altres àtoms i grups d’àtoms amb funcions molt diverses. Els plàstics, d’ús tan generalitzat, són basats en les propietats de les molècules que tenen un esquelet de carboni. La vida encara n’ha tret més partit que la tecnologia humana: aigua i carboni abundants, i una temperatura que fa que la major part de l’aigua de la biosfera sigui líquida, sota un clima adient de radiació electromagnètica, constituïren la millor oferta de la Terra com a possible escenari de la vida.
L’aigua
Unes 3/4 parts de la massa (biomassa) de tots els éssers vius, per terme mitjà, són formades per aigua. Aquesta quantitat comprèn l’aigua com a tal, ja sigui líquida, ja sigui l’aigua semisòlida que s’adhereix fortament a les molècules de la matèria viva, més l’aigua que equival a l’oxigen i l’hidrogen que formen part de les molècules orgàniques.
El nitrogen
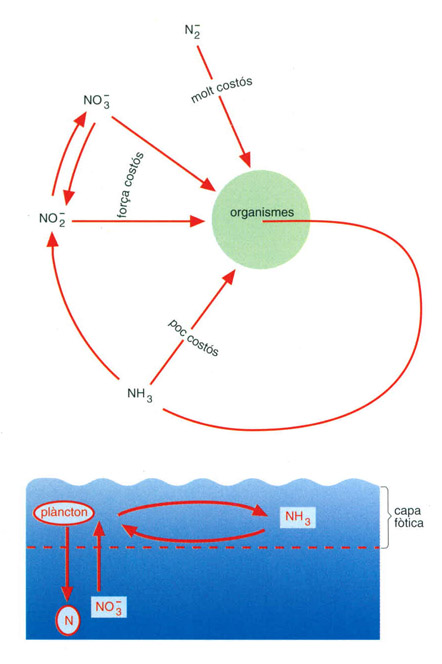
L’entrada de nitrogen en els organismes té un cost energètic diferent segons la forma en què es troba. Quan es troba en forma amoniacal (NH3) resulta menys costós que si es troba en forma de nitrit (NO2-) o nitrat (NO3-3-). La forma energèticament més cara és la de nitrogen gasós atmosfèric (N2). El reciclatge del nitrogen (per exemple, el que té lloc a les capes superficials dels oceans) es fa, en bona part, en forma amoniacal.
Editrònica, a partir de dades elaborades per l’autor
Altres elements químics s’han fet acceptables per a la vida i hi contribueixen amb les propietats peculiars dels seus àtoms. El nitrogen és notable per la varietat d’estats d’oxidació-reducció (nitrat i àcid nítric, nitrit, amoníac i l’element nitrogen en forma de gas o dinitrogen). El nitrogen és abundant a la Terra i és més aviat volàtil. Actualment, unes 4/5 parts de l’atmosfera són de nitrogen gasós. Dels dos elements que generalment limiten el creixement dels organismes, el nitrogen es recicla principalment entre la biosfera i l’atmosfera, mentre que el fòsfor es recicla més significativament entre la biosfera i la part sòlida de la Terra. Aquests dos elements es troben, tradicionalment, en totes les barreges de fertilitzants, i s’hi acostuma a afegir, també, potassi.
El nitrogen, juntament amb el carboni i els components de l’aigua, configura les molècules dels anomenats aminoàcids, per raó de la duplicitat de les seves funcions, que facilita la seva unió en cadenes. Alguns aminoàcids també contenen sofre. La vida utilitza amb una profusió especial no gaire més d’un parell de dotzenes de formes d’aminoàcids que, individualment, són relativament estables. Aquests aminoàcids tenen facilitat d’acoblar-se linealment els uns amb els altres per mitjà d’unions relativament més mòbils. De la mateixa manera que un nombre limitat de lletres, el mateix nombre que el d’aminoàcids corrents, ens serveixen per a confegir una infinitat de paraules, es pot construir un nombre pràcticament infinit de proteïnes a base dels aminoàcids comuns. L’analogia va més enllà, perquè les proteïnes tenen altres semblances amb les paraules; són com les paraules de la vida, les portadores d’una informació pràcticament il·limitada, sobre la base d’un abecedari molt efectiu. Les proteïnes fan serveis molt diversos, i excel·leixen com a catalitzadors, és a dir, enzims. La gran varietat de conformacions tridimensionals que permeten, especialment les de gran pes molecular (que consten de centenars d’aminoàcids), els dóna capacitat d’organitzar entorns particulars, i de situar de manera oportuna grups funcionals diversos, la qual cosa les predisposa per a funcionar com a enzims. També hi ha moltes proteïnes que fan una funció estructural i totes segueixen el pla de construcció modular que retrobem a escales tan diferents dins de la naturalesa.
El fòsfor
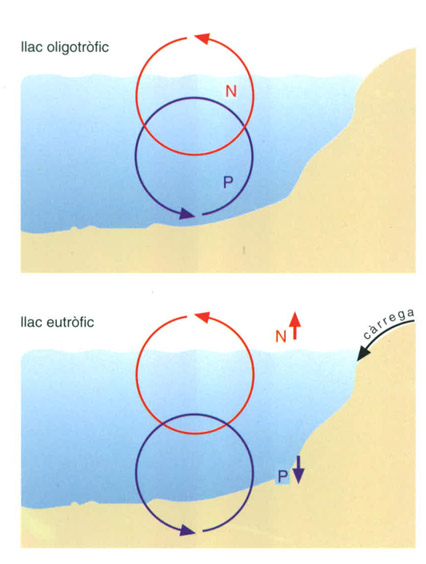
Els cicles del fòsfor i del nitrogen es comporten de manera diferent en un llac oligotròfic i en un d’eutròfic. En un d’oligotròfic, mentre que en el cicle del nitrogen predomina el pas de l’aire a l’aigua, en el del fòsfor predomina el pas del sediment cap a l’aigua. Si el llac rep una càrrega de nutrients i esdevé eutròfic, el cicledel nitrogen es decanta cap amunt, amb expulsió neta de nitrogen, mentre que el del fòsfor ho fa cap avall, amb pèrdua neta de fòsfor, que va al sediment.
Editrònica, a partir de dades elaborades per l’autor
El fòsfor és un altre element indispensable per a la vida. En els organismes es troba en la seva forma oxidada, com a àcid fosfòric i fosfats, i té funcions importants: pot ésser el component essencial de molècules molt efectives en l’emmagatzematge d’energia en enllaços relativament làbils, les quals actuen com a acumuladores i transportadores d’energia. Les més conegudes són els fosfats d’adenosina (AMP, ADP, ATP, monofosfat, difosfat i trifosfat d’adenosina respectivament; el segon i el tercer són importants acumuladors intracel·lulars). El fosfat associat als sucres i a les bases nitrogenades és el component principal dels nucleòtids, molècules que duen informació essencial per a la vida.
Mentre que el nitrogen és fugisser, ja que té més afinitat per l’atmosfera i els seus compostos són solubles en l’aigua, el fòsfor forma fàcilment compostos molt insolubles. Entre ells hi ha les combinacions amb calci (apatita) que encara són més insolubles quan s’hi afegeix fluor. Recordem l’esmalt dentari, en el qual els efectes de la mancança de fluor augmenta la possibilitat de solució per l’acció metabòlica combinada dels microorganismes de la placa dentària. El fòsfor es mou preferentment entre el medi líquid i el medi sòlid, en sediments i sòls. El fòsfor s’ha fet absolutament necessari per a la vida i podria ésser l’element decisivament limitant.
Els elements minoritaris
Altres elements químics s’han anat reclutant des de l’inici de la vida o bé al llarg de la seva evolució. El sofre és un element essencial i important, que és present en diversos aminoàcids. Altres elements freqüents (sodi, potassi, clor, calci) són part dels mecanismes elementals d’equilibri àcid-base i de sistemes de transport. Altres, com el silici i el calci, juntament amb el magnesi i l’estronci, componen esquelets rígids, entre altres funcions. Metalls com el ferro, que canvien fàcilment de valència química, tenen planer el canvi d’electrons. És com si diversos elements químics disponibles s’haguessin situat en els mecanismes de la vida: l’executant que troba el seu lloc a l’orquestra.
Aquests i altres elements poden aparèixer accidentalment, de manera erràtica, sense acomplir una funció precisa o necessària. O bé poden ésser francament nocius, si s’escau que troben facilitats per concentrar-se en alguns entorns des dels quals passen als organismes, com succeeix actualment amb molts productes industrials. Aquesta “contaminació” no és, però, cosa nova ni d’ara. Hom gosaria veure l’aspecte químic de l’evolució com una aventura bastant oportunista. Un procés ininterromput d’assaig i error en apropiar-se de certs elements químics i afegir-los a unes funcions que sempre han anat orientades a enriquir una capacitat informàtica.
L’arquitectura de les molècules vivents
Les complexes molècules vivents participen d’alguns trets comuns en la seva estructura i comportament. Mostren, per exemple, una contumaç tendència a mantenir la seva estructura i respecten determinades proporcions entre la complexitat estructural i la seva dimensió.
Molècules i persistència
Algunes de les molècules orgàniques formades per la vida i que la vida maneja resulten molt persistents. Sense anar més lluny, és el cas dels àcids desoxiribonucleics (DNA): tothom ha sentit parlar del DNA fòssil. És un aspecte d’una qüestió més general i molt important. La biomassa es refereix a la matèria dels organismes vivents. Hom pot parlar de necromassa per referir-se a materials sintetitzats per la vida, però ja sense vida i, en aquest estat, poden durar molt de temps.
Hi ha moltes molècules que passen més temps “mortes” que “vives”. En són un exemple els components de la fusta. En la fusta hi ha la cel·lulosa, que és feta de molècules d’un sucre unides de tal manera que formen una estructura semicristal·lina, molt difícil de descompondre per l’acció dels enzims comuns en l’àmbit on es troben organismes vius. La fusta també conté lignina, les molècules anulars de la qual també són químicament molt resistents i contenen nitrogen i altres elements. La fusta és un bonic exemple de la transició entre material desproveït de vida activa i material vivent, transició que hom pot observar entorn de la regió del càmbium, zona del tronc on la fusta creix cap a dintre i l’escorça cap a fora. En el petroli persisteixen cadenes d’hidrocarburs que foren produïdes per organismes ara fa desenes o centenars de milions d’anys, i continuen conservant en els seus enllaços interatòmics energia del Sol d’aquell temps; l’energia que actualment degradem en els múltiples usos que la nostra civilització fa del petroli.
Dimensió i estructura
Les dimensions fixes dels àtoms i les molècules defineixen l’escala de construcció d’organismes: totes les funcions cel·lulars necessiten unes dimensions força definides per a manifestar-se. El nucli cel·lular té una mida mínima fixada per la quantitat de DNA i pels sistemes accessoris necessaris per al seu maneig. Es reconeixen limitacions anàlogues en l’estructura de la membrana cellular (en la qual intervenen notablement lípids, i que conté punts especials de pas o de transport), en les fibres musculars, etc.
Els virus no tenen el mecanisme necessari per a fer les rèpliques, però conserven el material replicable: les seves dimensions són d’una fracció de micròmetre (0,05-0,2 µm) i són les estructures vives més petites. Sempre hi ha virus lliures en tots els ecosistemes i es troben en concentracions molt elevades, com proven anàlisis sistemàtiques recents fetes en aigües dolces i marines: els nostres coneixements són més limitats pel que fa als sòls.
Fora de cèl·lules excepcionaiment llargues, com les neurones i algunes altres, les dimensions cellulars es troben compreses, en general, entre 5 i 50 µm i el límit superior de la mida dels organismes és també reduït, per altres raons que se’ns faran evidents; l’animal més gros conegut, del passat i del present, és el rorqual blau (Balaenoptera musculus), que pesa fins a 150 t. El grau de quantificació de la vida és més important del que hom suposa per entendre el funcionament de la biosfera. Defineix almenys la forma d’acoblament entre els fluxos materials i la dinàmica de les poblacions. Les transferències de material entre parts dels ecosistemes es poden comparar, més que a un flux continu, a aixetes que degoten; això contribueix a fer que els sistemes parcials entre els quals s’estableix el bescanvi (organismes o conjunts d’organismes) estiguin fora d’equilibri.
Els camins dels nutrients: circuit tancat, circuit obert
Els dipòsits i els fluxos
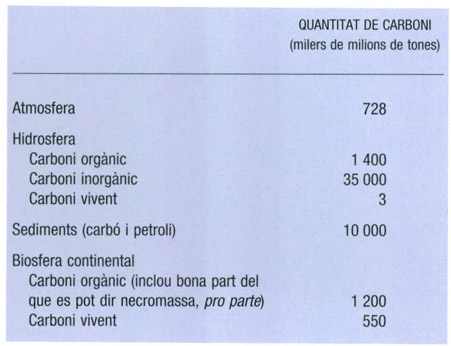
La distribució aproximada del carboni, expressada en milers de milions de tones (= petagrams), en els diferents compartiments en què es pot subdividir la perifèria de la Terra, és la que queda expressada en aquest quadre.
Dades elaborades per l’autor
En qualsevol apreciació material de la biosfera podem distingir entre dipòsits i fluxos. Fa temps que s’intenta una avaluació global o planetària d’uns i altres, que per diverses raons mai no pot ésser gaire exacta. Ara potser hi ha més urgència a causa de les preocupacions nascudes del més que probable increment de la concentració de CO2 a l’atmosfera; però cal dir que han augmentat més les especulacions que no els estudis acurats. L’aigua i els seus components són importants, però com a factors limitants vénen en segon terme. Hom pot fer diverses aproximacions de la quantitat de carboni en diversos compartiments de la perifèria de la Terra. Les quantitats d’altres elements importants amb relació a la vida (nitrogen, fòsfor) són encara menys precisables.
L’organització de la biosfera se simbolitza amb la construcció d’un model de capses cada una de les quals representa un reservori o contenidor important i definible com les capes superiors de la mar, el sòl, etc. Els fluxos entre uns reservoris i altres es representen amb sagetes i s’intenten quantificar i expressar per unitat de temps. Amb aquestes dades i si fos acceptable un estat estacionari, es podria assignar un temps mitjà de permanència al material de cada recipient. En realitat el seu contingut fluctua i es produeixen acceleracions i desacceleracions en els fluxos entre els contenidors. Encara que diversos elements puguin migrar de manera paral·lela d’uns reservoris a uns altres, no hem d’esperar que ho facin a la mateixa velocitat. El fòsfor del nucli de la cèl·lula és més persistent que no pas el fòsfor relacionat amb el sistema ATP → ADP, molt més làbil. La taxa de renovació de bona part dels elements és més alta en el fetge que no en el cervell, o en les fulles més que no pas en la fusta. Igual de grans poden ésser les diferències respectives entre parts d’ecosistemes.
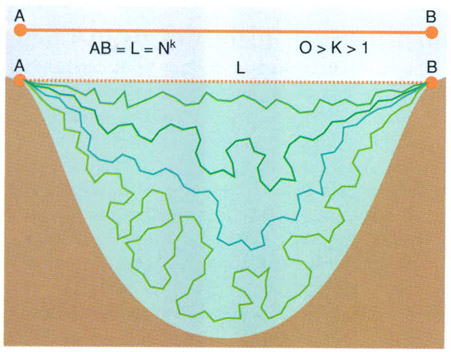
Entre dos punts extrems d’un espai limitat, per exemple d’un llac, hi ha tants trajectes com valors pot prendre k a l’expressió L = Nk. A la pràctica, però, la majoria de les vegades el valor de k és pròxim a 1/2 i varia segons que la gota d’aigua o l’organisme del fitoplàncton que considerem circuli prop de la superfície (valor més pròxim a 1, camí més directe) o més profunditat (valor més acostat a 0, camí més sinuós).
Biopunt, a partir de dades elaborades per l’autor
Combinar velocitats i trajectòries porta una dificultat que s’entén fàcilment si hom tracta d’imaginar com es mou l’aigua en un llac o en un embassament. Algunes molècules d’aigua van gairebé en línia recta i triguen molt poc temps a anar des de l’afluent fins a l’emissari; altres, ben al contrari, s’entretenen molt descrivint una llarga trajectòria sinuosa i irregular que tal vegada les porta a travessar nivells més profunds del llac. El model més senzill es fa relacionant la distància (L) entre dues posicions extremes, mesurada en unitats de “passos”, i el nombre de “passos” necessaris (N) per a recórrer-la. L’expressió serà
L = Nk, on 0
Es pot suposar que la distribució del flux d’un riu que travessa un embassament dóna una distribució de valors de k que correspon a un espectre bastant ampli representatiu d’una diferenciació de les velocitats, just com la que es troba dintre els organismes i en el teixit dels ecosistemes.
Si usem “passos” progressivament més curts apareixen complicacions d’un altre caire, que ens porten al concepte de quantitats fractals i a totes les paradoxes esmentades en llibres de divulgació quan tracten de la llargada de les fronteres entre països i dels conflictes en què es poden trobar els agrimensors. Problemes molt semblants es presenten a l’ecòleg, i no sols al que estudia fluxos, sinó també al que fa mapes o analitza els límits entre ecosistemes.
Cal tenir presents aquestes i altres compilacions a l’hora d’interpretar les mesures de producció, de jutjar la composició d’un sistema complicat o de fer i de llegir els gràfics i diagrames que il·lustren la major part dels llibres d’ecologia. I cal tenir-los presents perquè petits canvis de velocitat poden conduir ràpidament a distribucions força diferents, i la dificultat de mesurar aquella velocitat de flux, com també la variabilitat intrínseca dels mateixos fluxos, fa que hom hagi de ser escèptic davant d’un bon nombre de les prediccions que s’arriben a proposar en l’àmbit de l’ecologia global.
La recirculació del carboni a escala global pot ésser exemplar. Les plantes usen CO2 que treuen directament de l’atmosfera, la qual en conté en una proporció una mica superior al 0,3‰ del volum, o bé del que hi ha dissolt en l’aigua en forma de gas o d’àcid carbònic dissociat (CO2 + H2O → CO3H2 → H+ + CO3H- → H+ + CO3=). Part d’aquest carboni assimilat per les plantes és respirat per elles mateixes, i una altra fracció és menjada i respirada pets heteròtrofs (Corgànic + O2 → CO2). Els heteròtrofs depenen dels autòtrofs (plantes) i es reparteixen en dues categories: els que utilitzen materials orgànics en solució, o osmòtrofs, com els bacteris, els fongs i molts paràsits (plantes o animals) i els que ingereixen materials sòlids i els digereixen, o fagòtrofs. Amb relació a tots els elements importants convé reconèixer les limitacions que van lligades als cicles globals i als cicles inclosos dintre d’altres cicles.
El cicle del carboni
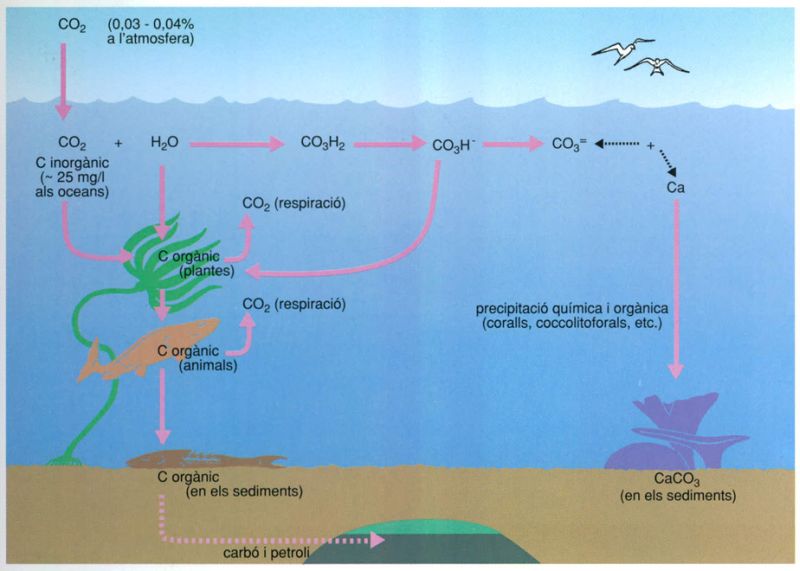
Biopunt, a partir de dades elaborades per l’autor
La circulació del carboni a escala global és un bon exemple de la influència mútua entre els cicles globals dels diferents elements (en aquest cas el calci) presents al medi i de la intervenció dels diferents tipus d’organismes en cadascun. Una part del carboni present a l’atmosfera en forma de diòxid de carboni pot combinar-se amb el calci i precipitar en estructures orgàniques persistents, com són els baixos de corall, els exosquelets de les coccolitoforals, etc., o perdre’s en els sediments. Una altra part, després d’haver passat com a carboni orgànic a través de les cadenes tròfiques, també pot acabar al sediment però en forma de carboni orgànic de la necromassa.
La circulació i el reciclatge
En oceanografia és habitual distingir entre una producció primària “nova” i una producció “reciclada”, amb referència a les capes superiors de la mar, és a dir, al gruix d’aigua suficientment il·luminada (el primer centenar de metres). Allí és on se sintetitza matèria viva nova, que en part és respirada o menjada per animals, i que torna a l’aigua en forma de CO2 i dels corresponents compostos oxidats (o reduïts, pensem en els productes amoniacals) dels altres nutrients essencials. El fòsfor circula sempre com a fosfat i les fosfatases solen ésser enzims ubics. Aquest material pot ésser assimilat novament per les algues, de manera que la disponibilitat de llum determina un cicle gairebé tancat en les capes superficials, talment com es pot aconseguir en un cultiu dintre d’un flascó de laboratori, o en una gota d’aigua mantinguda entre portaobjectes i cobreobjectes degudament tancats i disposats per a l’observació microscòpica. Fa uns anys es posaren de moda les “ecosferes”, sistemes amb aigua, nutrients i alguns organismes confinats en un recipient esfèric de vidre. Aquestes “ecosferes” anaven mantenint una vida més o menys intensa segons la il·luminació i la temperatura a les quals estaven sotmeses.
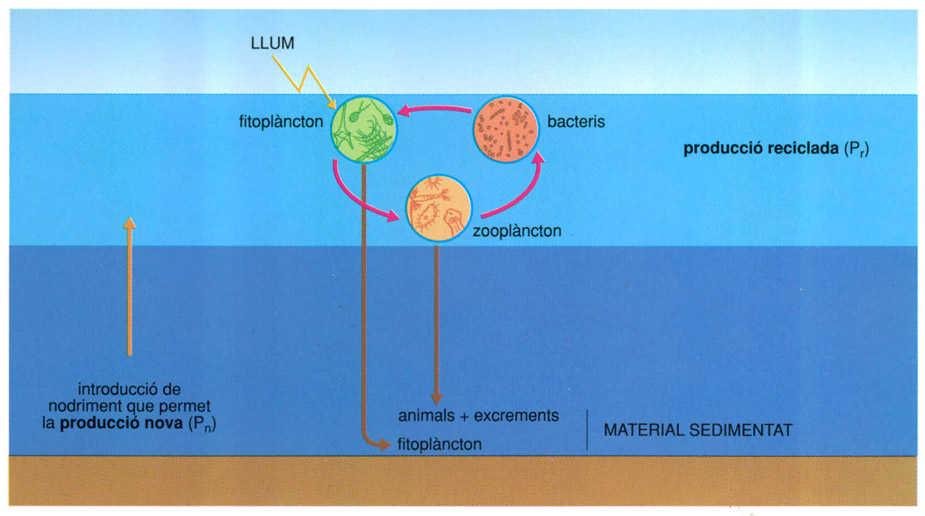
En mars i oceans, la producció primària “nova” (síntesi de matèria viva denovo) i la producció “reciclada” es distingeixen en el primer centenar de metres d’aigua. La relació entre la producció “nova” i la “reciclada’’ oscil·la d’ordinari entre un 10 i un 50%.
Biopunt, a partir de dades elaborades per l’autor
A la mar, una part del material se sedimenta, ja sigui directament o bé per efecte dels animals que mengen a prop de la superfície i després es mouen cap a nivells profunds, on deixen els productes de rebuig. De fet, el reciclatge a les capes superiors, on els materials es fan renovables, normalment comprèn entre el 60 i el 90% del total. Per mantenir unes condicions de producció estables la resta ha de venir de fora del compartiment format per la capa superior de la mar, i normalment hi arriba amb l’ascensió d’aigües més profundes. Mantenir un nivell uniforme de vida representa que l’entrada de nutrients sigui equivalent a la part de la producció primària que se sedimenta. Hi ha com una baga accessòria que s’estén i circula de dalt a baix. Precisament el retard que aquesta circulació secundària adquireix en un oceà profund és una causa de la menor productivitat general dels oceans amb relació a la superfície dels continents.
Els productors primaris, les plantes, per un costat assimilen CO2 (a la llum) i per l’altre produeixen CO2 en la respiració (a tothora). Si el CO2 és un producte limitant, l’evolució ha portat a fer recircular el que resulta de la respiració, és a dir, a prendre mesures per reassimilar-lo, cosa que implica mantenir-lo temporalment dintre la planta, en un altre cicle que comporta certs àcids no volàtils. Hi ha moltes plantes terrestres que han aconseguit aquest reciclatge intern, principalment plantes pròpies de temperatures altes i sovint pobladores de clarianes de boscos tropicals. Són plantes que creixen ràpidament i respiren molt, com el blat de moro, la canya dolça, els bambús, etc. Sembla que evolutivament es tracta d’una adquisició recent, ja que s’ha aconseguit per vies diverses convergents en llur funció, i podria haver evolucionat acompanyant, adaptativament, un descens de la concentració atmosfèrica de CO2 en temps geològicament recents. En les plantes aquàtiques la necessitat indicada no s’ha deixat sentir tant, i encara menys en les que viuen en aigües d’alta reserva alcalina, aigües que poden retenir molt carboni a l’abast en forma de bicarbonat.
Les estratègies dels ecosistemes
La naturalesa és força flexible i acomodatícia. Si en una suspensió d’algues, en un plàncton, escassegen els elements necessaris per a la multiplicació cel·lular (fòsfor, nitrogen), però hi ha carboni i llum, sovint les algues continuen la síntesi. Aquesta síntesi va acompanyada, però, de la producció de mucílags que poden anar embolcallant les cèl·lules, i que representa una mesura de precaució contra la dessecació en les algues costaneres. Una ralentització relativa comparable es dóna en boscos tropicals humits situats en planúries: la major part dels nutrients essencials s’han incorporat dintre dels organismes vius i aquests limiten el seu creixement a la reposició de parts malmeses. Al mateix temps, evolutivament, han aconseguit reduir al mínim el cicle extern de fòsfor i nitrogen. En aquestes condicions, qualsevol vertebrat té un capital en l’esquelet; potser per aquesta raó no hi ha gaires vertebrats, i gairebé cap cargol. Solament amb la proximitat de muntanyes i rius i amb l’accés de nous materials es manifesta una taxa més gran de creixement i renovació.
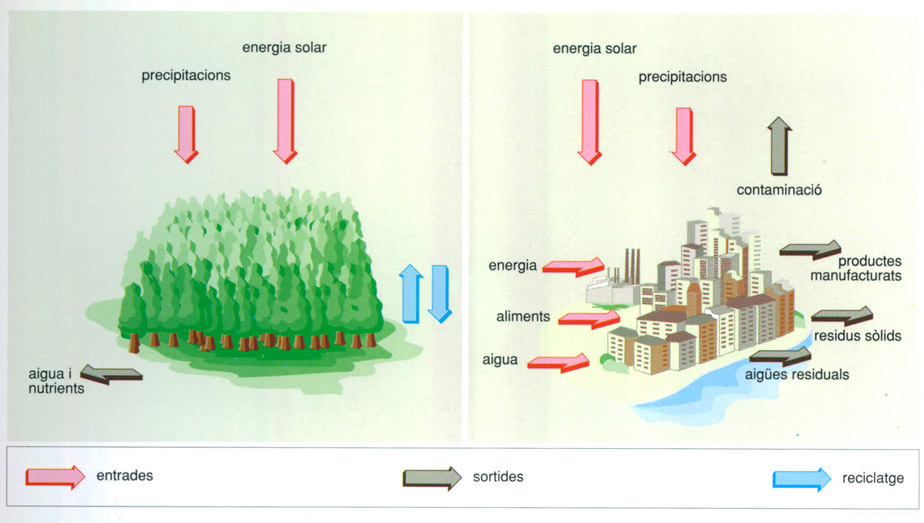
La selva i la ciutat són exemples extrems del paper del transport horitzontal en els ecosistemes. La selva exemplifica els ecosistemes on el flux horitzontal de nutrients es minimitza i pràcticament és perfectament divisible en columnes verticals funcionalment idèntiques. La ciutat, per contra, exemplifica els ecosistemes que no poden subsistir sense actius bescanvis laterals de materials o recursos essencials i organitzats internament de manera que el transport horitzontal és tant o més important que el vertical.
Editrònica, a partir de dades elaborades per l’autor
La interpretació de la polaritat dels ecosistemes porta a imaginar-los com sistemes integrats per columnes o per fibres verticals. Precisament el transport lateral és un criteri important d’anàlisi. Un reciclatge complet segons la vertical local caracteritza un sistema més tancat: a aquest model s’aproximaria l’Amazònia, per exemple, però també molts altres ecosistemes on les fronteres sobre la superfície del sòl no són llocs de flux net horitzontal de nutrients. Aquests sistemes són eminentment divisibles. Hi ha, per contrast, ecosistemes que canvien materials essencials amb altres ecosistemes situats lateralment, com la plana depèn de la muntanya, o com els camps de conreu i les ciutats apareixen connectats horitzontalment.
El significat especial de la dimensió vertical es visualitza a través d’un experiment molt senzill. Posem dos aquaris de condicions i de poblament semblants. En un es divideix el contingut en columnes adjacents, per mitjà d’envans col·locats verticalment, o per trossos de tubs de material plàstic oberts pels dos extrems, amb l’eix en sentit vertical. En l’altre aquari l’envà o els envans es disposen horitzontalment. En el primer, les diverses parts continuen essent molt semblants a com eren abans i no necessiten transport horitzontal per a subsistir en la seva condició primera, llevat de si hi havia algun peix molt gros que hagués quedat confinat en un dels espais. El segon experiment ha interferit amb l’organització vertical fonamental dels ecosistemes i ha creat condicions noves: a dalt hi ha contacte amb l’atmosfera i a baix no, i a més a baix hi ha una part de sediment amb materials procedents d’una etapa anterior de l’activitat de l’ecosistema.
Els sòls han tingut aquesta funció, i l’agricultura ha escapçat ecosistemes antics, substituint-ne la part superior i conservant la part inferior en qualitat de sòl agrícola. Idees semblants ajuden a interpretar formes de conreu, de fertilització, etc. Fonamentalment, els humans tendeixen a “obrir” tots els sistemes; per això una tasca de la conservació ha d’ésser restringir les funcions de transport extraordinàries que la humanitat els ha imposat.
Les estratègies dels organismes
Unes poblacions animals es renoven més de pressa que les altres. Unes tenen èxit perquè es reprodueixen ràpidament malgrat que tenen una gran pèrdua d’individus. Altres espècies estan més ben defensades i els individus viuen més temps i deixen menys descendents. Tot això també té a veure amb l’alimentació, i és a la base de curioses relacions biològiques.
Els sucs de les plantes són molt diluïts, amb excepció dels sucres. Insectes molt sedentaris explotadors dels vegetals, com per exemple els pugons i les cigales, claven una mena de bec en els vasos de la saba, d’on treuen aliment, però sovint els sobren els sucres i els excreten de manera gairebé directa mitjançant una mena de filtre que tenen en una llaçada annexa al tub digestiu; aquesta llaçada treu el sucre o manà de molts hemípters petits o l’escuma de les cigales d’escuma, tots ells animals notòriament poc actius i amb més necessitats de creixement que no pas d’energia per a moure’s. D’altra banda, tenim les formigues, insectes de vida més activa, la qual els comporta grans despeses d’energia lligades al moviment, que compensen aprofitant el sucre que als pugons precisament els sobra. I pel que fa als colibrís és clar que necessiten proteïna, no poden viure només de xarop; però pel fet d’ésser bestioles d’una gran activitat usen una elevada quantitat de sucre, que obtenen amb el nèctar de les flors.
La interacció entre els factors de producció
Hom sempre recomana als estudiants que fan els primers experiments de química d’agitar ben bé el tub d’assaig. Aquesta agitació és necessària per a fer avançar la reacció; altrament, poden quedar substàncies sense reaccionar perquè estan separades, per exemple, a nivells distints dins del vas reactor si el sistema es va estructurant, o bé si es tracta d’un material col·loïdal que es compartimenta. Tota la biosfera té una estructura molt rica i pot mantenir fàcilment separats components que, altrament, podrien reaccionar. El món, tota la bellesa del món, ve del fet que l’espai és un gran aïllant que manté separats reactius potencials. Així, hom podria relacionar la producció real amb el resultat de multiplicar l’energia externa pel grau de superposició en la distribució dels diferents factors de producció.
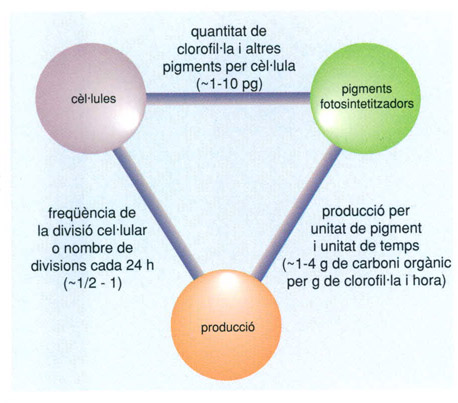
Normalment, en un programa d’estudi del fitoplàncton, les característiques que hom acostuma a estudiar són el nombre de cèl·lules, el contingut de pigments (valorables químicament) i la productivitat (capacitat de produir O2 o de fixar 14C). Però les relacions entre aquestes magnituds, relacions que es poden considerar totes com a característiques intensives, són igual d’importants o més. A l’esquema s’indiquen entre parèntesis els valors més freqüents d’aquestes relacions.
Biopunt, a partir d’un original de Ramon Margalef
És clar que els factors de producció poden ésser l’aigua, la llum, els elements nutritius i les algues microscòpiques del plàncton. Si tot això es troba junt es produeix el miracle de l’augment de la vida, i continua produint-se fins que algun element necessari comença a fallar per exhauriment. És com un procés de construcció o muntatge que s’interromp quan deixa de subministrar-se alguna de les parts necessàries, sigui quina sigui.
Sovint aquests elements essencials no es perden del tot, però queden segrestats durant un temps en algun reservori o espai particular. Per exemple, el confinament del fosfat en aigües profundes i la incertesa que es presentin condicions favorables per al seu retorn a la llum és la causa principal que la producció dels sistemes aquàtics es vegi deprimida, especialment en les regions centrals i subtropicals dels grans oceans. Això es podria expressar escrivint l’equació anterior d’una manera que permeti considerar una baga externa (de fet sovint tota una jerarquia de bagues externes): biomassa persistent en el lloc + exportació (sedimentació en sistemes aquàtics) = entrades de l’exterior + (energia externa × covariància en la distribució dels diferents factors de producció).
Els mecanismes de transport
La distinció entre recursos renovables i recursos no renovables conté una referència a la qualitat del reciclatge i de l’espai on es fa. És clar que en el nostre univers tot és potencialment reciclable, però si la nostra civilització importa des d’un lloc materials procedents de llocs diferents i, després de combinar-los i d’usar-los d’alguna manera, no retorna cadascun d’aquests materials al lloc d’origen, aixeca una barrera en l’espai i en fa més lent el retorn. Aquest és un dels riscos més importants de la contaminació, considerada com una malaltia del transport. Aquesta malaltia consisteix en el refús a pagar el preu del retorn, i a confiar-ho als mecanismes naturals de transport, els quals en aquest cas poden resultar excessivament lents per la pressa que porta la nostra civilització. Es comprèn que un ecosistema es pugui reactivar per subministrament del component deficitari, procedent de l’exterior. Això és el fonament de l’ús de fertilitzants o adobs. Aquest subministrament es pot considerar com un subsidi extern que sovint compromet una futura persistència del sistema.
Hom ha comparat les vies de transport natural a un model de passeig a l’atzar. Aquest model ofereix moltes possibilitats, com la d’entendre la diferència que hi ha entre la velocitat de l’aigua damunt del llit d’un riu i la del corrent subalvi que circula o percola en els sediments del mateix riu. És extraordinària la diversitat de mecanismes de transport natural que operen, i desperta interès i admiració la manera com es combinen i s’integren, o s’influeixen mútuament.
La Mediterrània es fertilitza per la modesta barreja vertical que es produeix en trencar-se la termoclina a la tardor, i per la contribució dels rius i del moviment ascensional que els mateixos rius creen amb l’aiguabarreig d’aigua dolça i marina; a això cal afegir-hi episodis d’aflorament costaner, que es produeixen sota l’impuls de vents favorables, i a més, s’ha de tenir en compte encara l’ascensió d’aigües profundes en llocs apropiats al llarg de fronts hidrogràfics més o menys persistents, sense oblidar la interacció de corrents marins amb illes, illots i fins a sobtades diferències de fondària. I cada espai marí conté una combinació diferent de mecanismes semblants, que defineixen la producció mitjana total. Així, els diversos mecanismes s’integren de manera diferent en la Mediterrània clàssica i en la Mediterrània americana o mar de les Antilles; en aquesta darrera, la situació és tal que dóna una producció total o mitjana una mica més elevada. D’altra banda cal dir que a vegades aquestes mars secundàries fan de més bon integrar que no els oceans principals, senzillament perquè la posició de les diverses estructures hidrogràfiques significatives en la producció resulta geogràficament més fixa, i més bona d’estudiar, en una mar relativament petita que no en una de més extensa.
Els ambients terrestres i els ambients aquàtics
L’evolució de la vida es pot veure com un arbre genealògic molt ramificat, en les branques del qual es manifesten, parcialment i en diverses direccions, les potencialitats de la vida en general. Hi ha unes certes regularitats o principis generals en el camí que va des del senzill al complex, a mesura que es van superant les proves a la supervivència posades per l’entorn. Aquest entorn planteja exigències ben definides, derivades bàsicament de les condicions ofertes per la Terra, amb relació a les quals es defineixen les principals direccions d’adaptació de la vida i de la seva evolució. Les condicions del món físic operen en primer lloc sobre els productors primaris, els vegetals, els quals, malgrat recolzar-se en uns mecanismes bioquímics comuns per a la captació de l’energia solar, adopten formes i estructures força diverses, segons si es tracta d’utilitzar el medi aeri (plantes terrestres o continentals) o el medi aquàtic. El fet que l’aigua sigui dolça o salada és secundari. Com que els animals depenen dels vegetals (o bé els uns dels altres), també mostren característiques d’adaptació que poden ésser divergents, però potser no en el mateix grau que en les plantes.
Els boscos i el plàncton
Una bona aproximació al dilema inicial consisteix a comparar el bosc amb el plàncton. El plàncton vegetal o fitoplàncton, que viu en suspensió en les aigües marines i en les aigües dolces o epicontinentals, és format per organismes unicel·lulars i, per tant, molt petits, d’entre 1 i 100 µm. Si les condicions són prou favorables, aquests organismes es multipliquen molt de pressa per divisió cel·lular i arriben a duplicar la seva massa en un temps que sol variar entre mig dia i una setmana. Sovint les cèl·lules que resulten de les divisions ràpides queden enllaçades les unes amb les altres fent cadenes, ventalls o agregats d’altres formes.
La rapidesa de multiplicació dels organismes unicel·lulars del plàncton ha fet pensar com serien d’útils per a fer cultius i obtenir proteïnes o materials de possible ús per a l’home. Duplicar la biomassa present en un dia no és possible en la vegetació terrestre; aquesta demana mesos per a produir una collita acceptable. Biològicament, la comparació és fal·laç, ja que en realitat la massa més gran de les plantes terrestres, o de les “plantes superiors”, consisteix en un dispositiu de suport i de transport fet de material especialment resistent i amb prou feines viu, i fins i tot en bona part no vivent; és el que passa amb la fusta, la qual forma una fracció molt considerable de la vegetació terrestre. I el mateix s’aplica als espermatòfits que han retornat al medi aquàtic, és a dir, al que se suposa que és el seu medi ancestral.
El que es pot veure com una planta individual pertany, com tots els éssers vius, a una espècie. La determinació de les seves característiques finals depèn, és cert, de l’estructura general de la tija, que és la que condiciona la tendència a un determinat estil de ramificació. Però no hi ha dues plantes de la mateixa espècie que tinguin una forma idèntica. Posseir una part persistent afavoreix molt les plantes perennes, plantes que solen tenir l’activitat vegetativa limitada a una estació favorable, perquè mitjançant el manteniment d’una estructura, que és alhora un banc gènic, mantenen ocupat un lloc i hi perduren, a punt per a treure fulles i flors (de vegades, primer les flors que les fulles) tan bon punt arriba l’estació favorable.
El significat exacte de les diferències d’estructura entre el bosc i el plàncton té molt a veure amb la naturalesa física de l’entorn i amb els mecanismes en virtut dels quals s’aconsegueix el reciclatge dels materials. El fitoplàncton viu suspès en l’aigua: alguns dels seus organismes, com ara les diatomees i moltes crisofícies i desmidiàcies, són passius i generalment tenen una densitat lleugerament superior a la de l’aigua, de manera que tenen una tendència natural a sedimentar-se, i ho fan a una velocitat generalment compresa entre decímetres i metres per dia; altres organismes del plàncton, com ara les dinofícies, les euglenals, moltes crisofícies, les coccolitoforals i diversos grups de flagel·lades, poden nedar, i si ho fan en la vertical i cap amunt, aconsegueixen de mantenir-se a un mateix nivell; i, finalment, altres organismes són tan petits (1-2,5 µm) que viuen esclaus de l’aigua per les forces de la viscositat i amb prou feines s’escapen d’aquesta servitud.
El fitoplàncton es multiplica en les capes illuminades de l’estrat superficial (en general, no més de 100 m d’aigua, i sovint molt menys), on consumeix els elements més escassos i necessaris per a l’edificació de les estructures vives. Generalment, el fòsfor i el nitrogen són els elements decisoris. Si el moviment net de les algues es fa cap al fons, els materials esmentats circulen formant part del seu cos cap a nivells més baixos. Els animalets del plàncton també hi contribueixen, ja que tenen, en general, el costum de menjar prop de la superfície durant la nit i de moure’s durant el dia cap a més fondària, on defequen. És corrent que els excrements d’aquests animalets del plàncton siguin compactes, i aleshores se sedimenten amb més facilitat. El resultat final és que la solubilització en l’aigua de compostos dels elements químics més limitants (el fòsfor i el nitrogen) es fa a un nivell de fondària que es troba bastant per sota del nivell on foren assimilats. A poc a poc, el reciclatge es fa lent, i el nivell mitjà, entorn del qual els elements limitants han retornat a l’aigua i es distribueixen, va esdevenint més profund.
El retorn dels elements que fan possible la continuació de la vida s’ha de produir per l’ascensió de les aigües profundes, la qual es fa de manera desordenada, sigui per barreja sota l’acció del vent, sigui (en les regions d’aflorament) com una manifestació local de moviments circulatoris més amplis, originats en la interacció entre l’atmosfera (principalment, vents) i l’aigua. L’aportació d’elements procedents dels continents que es fa principalment a través dels rius té una importància secundària i es limita a la proximitat de les costes. Els moviments marins més importants responen amb processos locals i irregulars de barreja quan interfereixen amb illes o esculls. Els moviments ascensionals són sempre desordenats i poc efectius.
En la part continental de la biosfera la situació és ben diferent. És clar que les plantes utilitzen els nutrients del sòl i que tenen els mateixos problemes d’esgotament d’aquells nutrients que són escassos i limitadors de tot el procés: nutrients nous han de procedir de l’erosió de materials sòlids que es trobin en nivells més alts. El reciclatge es fa sempre a través de l’aigua, de manera semblant al que passa en ecosistemes aquàtics. Però sobre terra es tracta de l’aigua que té el seu origen en la pluja, la qual, després de recollir elements minerals necessaris en el sòl, on forma una solució molt diluïda, puja per l’interior de les tiges i els troncs de les plantes (dels macròfits, amb translocació estesa almenys sobre alguns centímetres, caràcter que diferencia aquestes plantes dels micròfits del plàncton) sota l’acció de l’evapotranspiració a les fulles. El mecanisme de circulació de l’aigua no depèn directament del sistema de la fotosíntesi, sinó que depèn de l’energia exosomàtica de l’atmosfera: de la pluja i l’evaporació. La saba circula per vasos que s’estenen verticalment; aquesta forma d’organització controla la circulació del nodriment més eficaçment i més previsiblement que no pot fer-ho el plàncton, ja que aquest ha de confiar en un mecanisme extern turbulent i sovint imprevisible, rarament òptim.
La dimensió vertical del reciclatge en els continents es limita als primers decímetres de les capes actives del sòl i a l’alçada de les plantes. En canvi, en els oceans poden interessar fondàries molt més grans, cosa que en fa un mecanisme menys efectiu i que introdueix un fort retard. De fet, en les aigües profundes dels grans oceans es troben concentracions de fòsfor de fins a 50-75 mg m-3 mai observades prop de la superfície, si no és en aigües contaminades de rius i en llacs eutròfics. Això, naturalment, fa que sigui molt més difícil de mantenir una producció marina alta, ja que ocorre només en una petita extensió privilegiada. Fet i fet, les dificultats de l’ascensió del nodriment o el llarg camí que aquest ha de recórrer en les tenebres abissals poden ésser la causa principal que la producció primària dels oceans per unitat de superfície sigui solament la tercera part de la producció primària mitjana de la superfície dels continents.
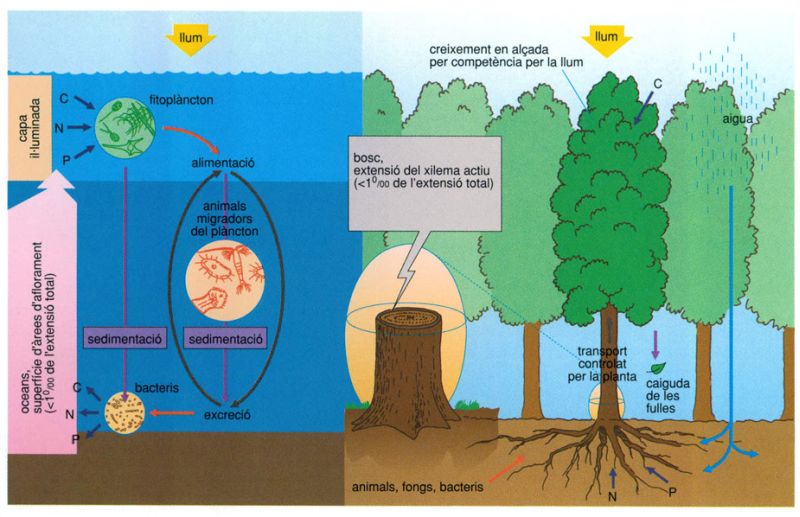
Malgrat les diferències d’estructura i de dimensió entre el plàncton i el bosc, resulta interessant analitzar les analogies que presenten i els seus límits, gairebé sempre relacionats amb la naturalesa física de l’entorn i amb els mecanismes mitjançant els quals es produeix el cicle dels materials. El dibuix en posa de manifest algunes: la ja assenyalada organització segons blocs verticals, que en el cas dels oceans es pot estendre en columnes de prop de centenars de metres, mentre que en el del bosc ho fa com a molt en columnes de desenes de metres; el predomini de la producció en els nivells superiors, relacionat amb l’accés a la llum, font d’energia indispensable per als productors, i el de la descomposició en els inferiors, tant si es tracta dels sediments dels fons oceànics com dels sòls; la lentitud del reciclatge, més acusada encara als oceans que als ecosistemes terrestres; remarquem també la coincidència en la proporció de la superfície de les zones d’aflorament respecte a la superfície total dels oceans i la de les seccions del xilema actiu, pel qual es transporten cap a les fulles els materials sense elaborar que la planta extreu del sòl, respecte a les seccions dels troncs sencers (menys d’un 1‰ en un cas i en un altre); i encara d’altres.
Biopunt, a partir de dades elaborades per l’autor
Les analogies i les diferències entre la vida vegetal terrestre i l’aquàtica es poden veure expressades de maneres molt diverses. La consideració d’una alga unicel·lular planctònica i d’un arbre que pot tenir una dimensió lineal de fins a 107 vegades més gran, fa pensar en altres diferències importants. La producció marina és mantinguda en el nivell actual perquè una fracció de l’extensió total dels oceans, potser una mica més de l’1‰, conté aigua que es mou cap amunt a velocitats d’1 a 10 m dia-1. Si la concentració de fòsfor de l’aigua que puja fos de 25 mg m-3 aquesta ascensió mantinguda durant un mes a l’any representa una aportació de fòsfor que permetria una producció primària de 100 a 500 g C m-2 any-1, la qual és comparable a la mitjana mundial observada (uns 100 g C m-2 any-1). Les estimacions de la producció marina es basen en els resultats d’incubacions en mostres confinades en ampolles, on la fixació de carboni es mesura mitjançant àtoms marcats (s’empra l’isòtop 14C del carboni), o bé mitjançant el bescanvi d’oxigen. En els continents, com és natural, la “collita” periòdica del material vegetal proporciona una estima més directa de la producció orgànica.
Per a introduir uns càlculs que permetin una certa analogia amb referència als arbres caldria conèixer la secció de les tiges de tota la vegetació continental sobre el pla horitzontal, i també la fracció d’aquesta vegetació ocupada per vasos llenyosos (xilema), que poden conduir cap amunt l’aigua i els materials que porta en solució. Els tècnics forestals empren una mesura, anomenada secció basal, la qual és la secció de tots els troncs a l’alçada convinguda d’l,3 m. Aquesta mesura s’expressa en metres quadrats per hectàrea (m2 ha-1), que equival a deu mil·lèsimes de la superfície del terreny, i els valors corrents se situen entre 20 i 50. El tant per cent que els conductes de la saba ascendent ocupen en un tronc no és una funció senzilla i constant de la secció, ja que es relaciona amb l’altura. De tota manera, per als troncs dels arbres europeus de diàmetre moderat és entre 1 i 10%. Multiplicant les dues xifres anteriors, tenim com a resultat entre 0,05 i 0,2 per mil de la superfície del sòl, valor que cal comparar amb l’1‰ que preníem per als oceans. Sembla inexplicable el poc interès que hi ha hagut a cercar informació útil o necessària per a fer aquesta aproximació. Ja que l’extensió de les fulles és almenys quatre vegades l’extensió del sòl sobre el qual es projecta el fullam, l’índex més interessant podria ser: extensió de les fulles / secció dels vasos del xilema actiu.
No hi ha hagut, però, gaire afecció a calcular-lo sobre observacions adients, i les poques xifres de què hom disposa donen valors situats entre 3 000 i 20 000 pel que fa als arbres; les “herbes” han estat quasi totalment negligides en un oblit imperdonable.
Les plantes i l’aigua
Per la manera com es presenta la qüestió de l’aigua en molts llibres de botànica, sembla que la vida de les plantes depèn quasi exclusivament de com poden controlar i limitar la pèrdua d’aigua.
Si fos així, el remei seria senzill: només caldria no tenir estomes i cobrir totes les fulles i fins i tot l’escorça amb algun dels vernissos impermeables d’alta qualitat que la naturalesa sap fabricar. Però no es tracta d’això: cal que l’aigua del sòl, que porta el nodriment mineral necessari, entri per les arrels i surti per les fulles, i que el flux sigui tan actiu com es pugui. Evidentment, una planta que habitualment deixa circular molta aigua i creix de pressa ha de poder controlar aquesta circulació per mitjà dels estomes i escapar-se del risc de marcir-se en algun moment. Els estomes també faciliten l’entrada de CO2 gasós i, com és natural, han de mantenir una certa relació amb l’evapo-transpiració, de manera que els estomes i llur control ajusten les dues funcions. Els estomes, petites “boquetes” d’una fracció de mil·límetre, es troben en nombre de diversos centenars per mil·límetre quadrat, especialment en la cara inferior de les fulles.
Tot funciona gràcies a l’evaporació que es produeix en els teixits immediats als estomes, l’anomenada evapo-transpiració, per la qual sol passar una tercera part de la pluja caiguda localment. Repetim que, com l’aigua oceànica elevada en les regions d’aflorament, aquesta circulació de l’aigua no és compensada per energia de la fotosíntesi sinó per l’energia externa del clima. L’evaporació que es produeix a les fulles aspira l’aigua del sòl i la fa pujar per dins la planta. La conducció es pot forçar per altres mitjans: per exemple, a la primavera, quan algunes plantes “ploren” en superfícies que hom ha tallat és perquè la saba puja abans de desenvolupar succió per evaporació, també per preparar les noves fulles; aleshores no és una saba molt diluïda, ja que hi ha materials de reserva (sucres, com els de l’erable de sucre [Acer saccharum]) que li donen una alta pressió osmòtica, capaç d’atreure fortament l’aigua de fora.
No coneixem amb seguretat la història evolutiva de la vegetació terrestre. Probablement les primeres plantes terrestres derivaren d’algues verdes del tipus de Coleochaete i altres de semblants que hom pot observar amb comoditat amb una lupa i que es veuen com minúsculs discos verds sobre les parets dels aquaris. A una escala més gran tenim les molses i una munió de criptògames. L’evolució ha portat cap a un augment de la mida i cap a la persistència dels sistemes de transport. D’aquesta manera, la planta, amb el banc de gens corresponent, sobreviu les temporades desfavorables perdent una part de l’aparell vegetatiu o de les fulles, i quan retorna una situació adient, pot aprofitar-se immediatament de les bones condicions. Aquesta manera de procedir sembla condemnar les plantes superiors més grans, els faneròfits, a una vida llarga i relativament pausada. Els botànics han reconegut sempre la superioritat que dóna a les plantes el fet de disposar d’una estructura superior persistent i el més desenvolupada possible.
Aquesta creença es reflecteix en la classificació ecològica de C. Raunkjaer (i en altres classificacions semblants fetes per altres autors), la qual consisteix, de fet, a proposar una terminologia grega (això sí, una mica més extensa o completa) per als conceptes vulgars d’herba, arbust i arbre. Hi ha dues formes molt importants de fer-se la competència: l’una és atrapar la llum abans que aquesta arribi a les altres plantes a base de créixer en alçada, i l’altra és agafar l’aigua abans que altres puguin utilitzar-la a base de construir un sistema radical prou extens o profund, segons demani la distribució de l’aigua en el terreny. Els arbres poden predominar fàcilment sobre l’herba: li intercepten la llum per dalt i estenen les arrels per baix. Sovint s’hi afegeixen d’altres formes de competència, sigui per substàncies específiques, sigui per l’acumulació de virosta. Això fa que hom pugui preveure al llarg de la successió ecològica no sols la dominància final de les formes arbòries, si la disponibilitat d’aigua ho permet, sinó que la manifestació del procés de successió pel que fa a la disminució del quocient producció/biomassa va essencialment lligada a l’acumulació de fusta, és a dir, de material poc actiu però amb una immensa capacitat per organitzar l’espai i assegurar-ne el domini.
La tendència dels arbres a allargar la vida determina una successió molt pausada de les generacions i, per tant, condemna les plantes superiors a una evolució relativament lenta. Hi ha fustes de diversa qualitat: una cosa és la fusta blanca i lleugera dels pollancres i els eucaliptus, i una altra cosa són les fustes “precioses”, molt denses, de gra fi i particularment resistents (caoba, banús), les quals han evolucionat sota l’amenaça constant d’un ambient humit i ple de fongs, en països tropicals. És clar que aquestes fustes amb prou feines poden ésser considerades com a recursos renovables; no ho són certament a l’escala de la nostra civilització.
Hi ha tot un seguit d’algues marines (Macrocystis i d’altres) que potser també mereixerien la qualificació de macròfits, ja que mostren transducció de diverses substàncies, almenys sobre distàncies decimètriques. És una evolució paral·lela a la de les plantes llenyoses terrestres, però mai no atenyen els mateixos nivells de persistència. Els macròfits originats a terra han envaït la mar, com en el cas de les posidònies (Posidonia), mal anomenades algues, o l’herba de la tortuga (Thalassia). Hi ha pocs animals de nissaga marina que les puguin digerir, de manera que els organismes que més les consumeixen són invasors secundaris de la mar, descendents d’avantpassats terrestres, com algunes tortugues marines i els sirenis (manatí i dugongs).