Les característiques estructurals
Les condicions ambientals que determinen l’organització i el funcionament de les selves temperades es caracteritzen per la benignitat del clima al llarg de tot l’any. L’abundància de precipitacions i la suavitat de les temperatures són condicions generals a totes les zones del bioma. Tanmateix, atesa la dispersió geogràfica d’aquest, hi ha diferències marcades en el funcionament de les diferents selves temperades i subtropicals pel que fa a la velocitat a què tenen lloc els processos de síntesi i descomposició de matèria orgànica, la utilització dels nutrients i el cicle de l’aigua. Les diferències són grans, en particular entre els dos pols més extrems de la gamma de les selves temperades: les laurisilves pròpiament dites i els boscos plujosos de coníferes.
Potser perquè la seva localització geogràfica és marginal i perquè l’extensió que ocupen és relativament petita en comparació amb altres biomes, les selves temperades han estat menys estudiades i les dades disponibles sobre producció i dissipació de la matèria orgànica, flux d’energia i circulació d’aigua i nutrients són més escasses. En molts aspectes, el funcionament de les laurisilves subtropicals és semblant al de les selves tropicals plujoses, però a les laurisilves de les zones temperades i temperades fredes situades a les costes occidentals dels continents les temperatures són més baixes i els processos fisiològics més lents. Al sòl de les regions on el clima fred es deu a una altitud o latitud més elevades, s’acumula una gran quantitat de matèria orgànica i el reciclatge de nutrients és lent.
Longevitat de les coníferes
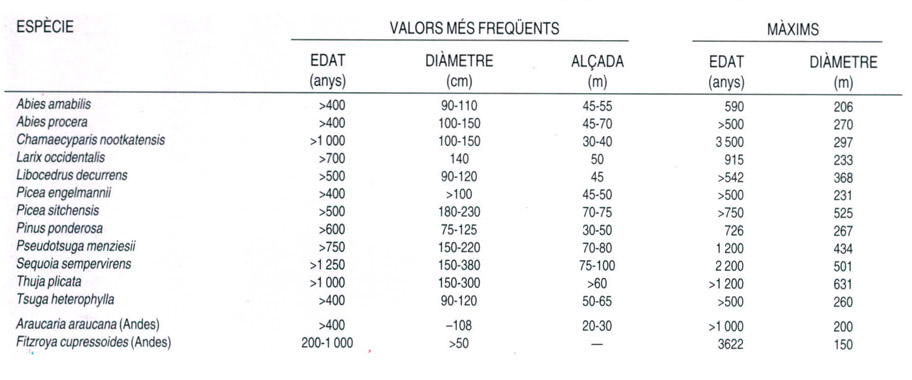
A partir de dades proporcionades per l’autor
Les coníferes que formen els boscos del NW de l’Amèrica del Nord, així com algunes de l’Amèrica del Sud, són de les més grans i longeves del planeta. Gairebé totes les espècies més freqüents en aquests boscos d’ambdós subcontinents poden viure com a mínim 500 anys, i són diverses les que superen fàcilment els 1 000 anys d’edat; fins i tot n’hi ha alguna d’especialment longeva, com el làrix americà (Fitzroya cupressoides) de les selves temperades de l’Amèrica del Sud del qual s’han trobat exemplars de més de 3 600 anys, tot i que els exemplars més vells d’aquesta conífera no solen superar el miler d’anys. Pel que fa a les dimensions, els exemplars més vells d’aquestes espècies poden créixer fins a alçades espectaculars, i el diàmetre dels seus troncs sol ser també considerable. El rècord se l’emporten la sequoia costanera (Sequoia sempervirens) i l’avet de Douglas (Pseudotsuga menziesii), amb alçades normalment superiors als 70 m i diàmetres de més de 150 cm, que en aquest cas superen clarament el longeu làrix americà.
L’estructura vertical
Sota unes condicions climàtiques benignes, la vegetació de les selves temperades es caracteritza per la seva exuberància, la seva complexitat estructural i l’organització del dosser en estrats, la variada composició de formes vivents i l’abundància de lianes i epífits. Els arbres emergents de més alçada poden superar fàcilment els 40 m i, en algunes laurisilves verges del Japó i de Xile, arriben a tenir diàmetres superiors a 1 m.
Les araucàries de les selves sud-americanes i australianes, per exemple el “pehuén” i el “kuri’y” (Araucaria araucana, A. angustifolia) o el “hoop pine” (A. cunninghamii), s’assemblen molt i tenen una arquitectura similar, amb un tronc cilíndric i recte que pot assolir gairebé els 50 m (encara que habitualment tenen de 35 a 40 m); la seva escorça és gruixuda i està dividida en plaques poligonals, i la capçada té una forma característica, amb verticils de 3 a 9 branques inserits a distàncies regulars al llarg del tronc. A la costa nord-occidental de l’Amèrica del Nord, és característica la presència d’araucàries enormes, de diàmetres que poden assolir i fins superar els 2 m i alçades de més de 100 m; evidentment es tracta de valors màxims, però les dimensions mitjanes d’aquests arbres també són considerablement més llargues que les dels arbres d’altres boscos, fins i tot si les comparem amb les dels arbres propis de les selves tropicals. Això es deu al fet que les espècies de coníferes de les selves temperades són molt longeves, normalment superen els 500 anys d’edat i poden arribar fins als 3 500 o més, com passa a Xile, on alguns exemplars de làrix americà o “alerce” (Fitzroya cupressoides) superen els 3 600 anys d’edat. Les coníferes estudiades a Austràlia i Nova Zelanda assoleixen edats màximes que superen els 1 000 anys, i els seus diàmetres també són semblants als dels arbres dels boscos de coníferes americans, llevat de la sequoia. El mateix es pot dir d’alguns grans eucaliptus (Eucalyptus).
Els arbres emergents no acostumen a formar un estrat continu, i per sota de les seves capçades es disposen un gran nombre d’arbres de mida mitjana, i sobretot petits, que omplen els buits i ocupen tot l’espai. Vista des de dalt, la superfície del cobricel de les selves temperades és rugosa i la seva alçada per sobre del nivell del terra força variable. Així, per exemple, en la majoria dels boscos de pins i “encinos” de Mèxic i de l’Amèrica Central, els arbres solen fer de 6 a 15 m, però no en falten, per l’extrem més sec, alguns que només assoleixen 2 o 3 m i, per l’extrem més humit, els que arriben als 40 o 50 m. Als arbres emergents se’ls acostuma a anomenar edificadors del bosc, i la complexa estructura vertical que resulta de la combinació de diversos estrats arboris crea una gran diversitat d’hàbitats on tenen cabuda nombroses espècies de plantes epífites, lianes, hemiparàsites i també una gran varietat d’espècies animals. Els herbívors disposen d’un ampli ventall de recursos alimentaris en la diversitat existent de fulles, flors i fruits, i els predadors també tenen on triar. Per sota del vol de les capçades, amb prou feines hi arriba a penetrar la llum i l’ambient és molt ombrívol durant tot l’any. En aquestes condicions, les espècies que poden mantenir un índex de fotosíntesi suficient per a compensar les despeses de manteniment són molt poques, i l’estrat de plantes herbàcies està escassament representat, llevat de les espècies que tenen per hàbitat les clarianes que es formen com a conseqüència de la mort d’algun arbre o grup d’arbres; en alguns casos, plantes herbàcies perennes gegants, com els bambús, poden formar un estrat molt dens i impenetrable de més de 3 m d’alçada.
La competència per la llum i l’índex foliar
A les laurisilves subtropicals i temperades la competència per la llum és molt intensa i constitueix un procés clau en l’organització de l’estructura vertical de la vegetació. És el comportament dels arbres de les diferents espècies envers la llum el que al capdavall determina la seva posició al dosser i la seva contribució estructural a l’ecosistema; així, els arbres de les espècies més heliòfiles ocupen els estrats superiors, en particular l’estrat dels arbres emergents, que, de vegades, són caducifolis. Per sota dels emergents, els arbres del cobricel són més tolerants a l’ombra. En un rodal ben desenvolupat d’una laurisilva, com en qualsevol bosc, la capçada dels arbres creix contínuament fins a assolir la màxima productivitat possible en funció de les condicions ambientals. En resposta als canvis, la disposició ramificada i apinyada de les capçades va essent modificada pel creixement i la mort de les parts aèries. La superposició en diferents estrats de les capçades distribueix les fulles en l’espai amb la màxima efectivitat i, en proporcionar més gruix al cobricel, determina que l’àrea foliar assoleixi valors elevats.
L’àrea total —en m2— de les fulles que es projecten verticalment sobre 1 m2 de terra constitueix l’índex foliar (LAI, per l’expressió anglesa “Leaf Area Index”). Atès que en la majoria d’espècies únicament hi ha estomes en una de les cares de la fulla, per a determinar l’índex foliar només es té en compte la superfície d’una cara. Aquest índex és una de les característiques més importants dels ecosistemes, ja que és a través de la superfície de les fulles que es produeix l’intercanvi de gasos en els processos de fotosíntesi i de transpiració i, per tant, aquest valor condiciona la producció primària. L’índex foliar sol ser molt elevat a les selves temperades, com es correspon amb condicions d’absència de dèficit hídric. Per a les laurisilves, els valors més comuns d’índex foliar oscil·len entre 5 i 9 m2/m2, i en algunes de madures s’arriba fins a 12 i 13 m2/m2, dades comparables a les de les selves tropicals plujoses, on de mitjana els valors estimats d’índex foliar estan compresos entre 8 i 12 m2/m2. Quan les condicions ambientals són més adverses, però, com s’esdevé en boscos magallànics de “guindo” (Nothofagus betuloides), a l’extrem austral del continent americà, a uns 55°S, l’índex foliar és molt més baix, i oscil·la entre 2,5 i 4,5.
Als boscos plujosos de coníferes aquest índex també és molt alt. Als boscos costaners, on predomina el hemlock occidental (Tsuga heterophylla), l’índex foliar varia entre 10 i 20 m2/m2, als boscos costaners de sequoia (Sequoia sempervirens) s’han mesurat valors pròxims a 22 m2/m2 i als boscos situats cap a l’interior i als vessants occidentals de la serralada de les Cascades (Cascade Range), on predominen l’avet de Douglas (Pseudotsuga menziesii), l’avet roig americà (Abies amabilis) i el hemlock occidental, l’índex oscil·la entre 6 i 10 m2/m2. Els valors de superfície foliar més grans, estimats als boscos de coníferes de la costa NW de l’Amèrica del Nord, s’han mesurat en condicions òptimes de temperatura i disponibilitat d’aigua, i són els que es prenen com a referència, és a dir, representen un màxim per sota del qual les condicions per al creixement són menys favorables. Al N de l’àrea de distribució d’aquests boscos (cap als 60°N), el principal factor limitador del creixement pot ser la baixa temperatura del sòl, que restringeix la taxa d’absorció d’aigua, mentre que a les zones meridionals i orientals, es considera que el condicionant més estressant és la manca d’aigua disponible durant l’estació de creixement, fins i tot a altituds elevades.
A l’interior de les selves temperades l’ambient és molt ombrívol durant tot l’any, ja que el tant per cent de radiació que arriba a terra és molt baix; en algunes laurisilves japoneses joves, per exemple, tan sols entre el 0,57% i l’1,7% de la radiació solar que incideix en el cobricel penetra fins a terra; la resta és interceptada per les fulles dels diferents estrats a mesura que va penetrant a través de les capçades. Quan la radiació solar travessa cada un dels estrats foliars experimenta una forta atenuació exponencial i, amb cada reducció, les condicions ambientals canvien per a les plantes i els animals dels successius estrats.
La relació entre índex foliar i àrea conductora
La suma de totes les seccions transversals dels troncs, mesurades a 1,30 m de la base, constitueix l’àrea basal total (AB). En 16 boscos estudiats a la laurisilva de Misiones, l’àrea basal oscil·la entre 39 i 15,8 m2/ha, i la mitjana és de 24,15 m2/ha. En general, aquesta àrea és més gran a les laurisilves pròpiament temperades que a les subtropicals. Cal tenir presents, com a dades comparatives, que per a les selves tropicals plujoses els valors mitjans d’àrea basal acostumen a estar compresos entre 23,1 i 35,3 m2/ha, i que valors d’entre 30 i 40 m2/ha són força habituals. A les selves temperades de coníferes l’àrea basal també és espectacular; a Nova Zelanda, els boscos mixtos i madurs de “kauri” (Agathis australis) tenen àrees basals de fins a 177 m2/ha, mentre que les àrees basals pròpies de les selves temperades mixtes de podocarps (Podocarpus) i planifolis prenen valors de 63 a 69 m2/ha (xifres referides només, en un i altre cas, a les espècies de coníferes). Als boscos costaners madurs de sequoia, l’àrea basal se situa entorn dels 343 m2/ha. Certament, aquests boscos creixen en indrets propers a la costa, sobre sòls rics en nutrients, formats sobre substrats al·luvials; aquí, les condicions de temperatura i humitat són especialment favorables, però als altres boscos de la costa NW de l’Amèrica del Nord, encara que no assoleixin valors tan alts, encara ho són molt si es comparen amb els que s’enregistren a les boscanes decídues o a les selves tropicals plujoses.
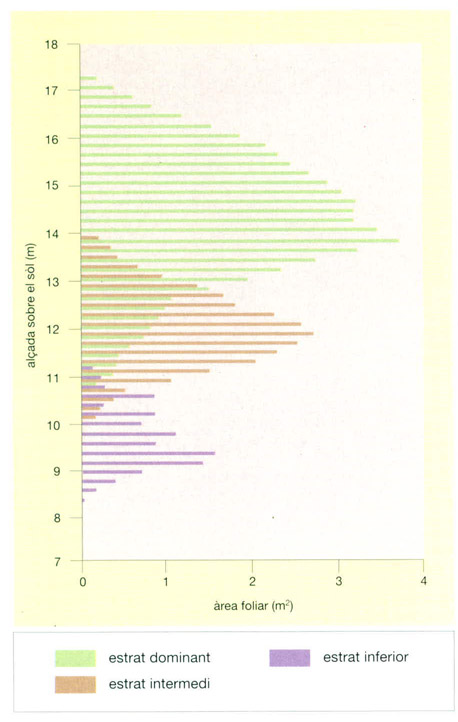
Distribució vertical de l’àrea foliar en una laurisilva canària d’una trentena d’anys. El dosser superior força tancat de les laurisilves i de la major part de selves temperades ja madures dificulta el pas de la llum del sol fins a l’estrat intermedi, de manera que les plantes que formen aquest estrat presenten una àrea foliar total més reduïda. El mateix succeeix als estrats inferiors del bosc, ja que el percentatge de radiació que reben és encara menor. La superposició de colors del gràfic mostra com l’àrea foliar de cada estrat és màxima a les respectives alçades mitjanes, i va disminuint cap als valors extrems, on les fulles de diferents estrats se superposen. En el cas concret d’aquesta laurisilva canària es calculà que el nombre total de fulles/ha arribava als 5 milions. Les plantes del gènere Myrica eren les que més fulles aportaven al bosc, gairebé 2,5 milions, seguides de les espècies de Laurus, Persea, Erica i Ilex. Tanmateix, el fet que una planta tingui un gran nombre de fulles no vol pas dir que tingui també una elevada àrea foliar, ja que moltes tenen fulles llargues i primes amb una superfície molt petita, com és el cas de les espècies del gènere Erica.
Jordi Corbera, a partir de D. Morales i altres, 1996
A més del seu caràcter estructural, les dades sobre l’àrea basal són interessants per la relació funcional que hi ha entre aquesta superfície i l’índex foliar. Tota l’aigua que transpiren les fulles a través dels estomes ascendeix al llarg del tronc canalitzada per vasos o traqueides. L’àrea basal, com l’índex foliar, sol ser força més elevada, ben bé el doble o el triple, en els boscos plujosos de coníferes de les costes del NW de l’Amèrica del Nord que en les laurisilves. Normalment, l’aigua transpirada per 1 m2 de superfície foliar és transportada per uns pocs cm2 (entre 1,5 i 15) de la secció del tronc. En una mostra estudiada de laurisilva canària, la relació entre l’índex foliar i l’àrea basal és de 0,23 m2/cm2; a les laurisilves en general, aquesta relació no és gaire diferent si es considera un índex foliar mitjà de 8 m2/m2 i una àrea basal de 30 cm2/m2, i encara és més baixa si es descompta l’escorça i es considera que no tota l’àrea basal és àrea conductora, perquè, a mesura que els arbres creixen, part de l’àrea basal deixa de tenir funcions de transport i tan sols la corona circular més externa és útil per a la circulació d’aigua i de nutrients.
A aquesta corona circular exterior formada per elements conductors i cèl·lules acompanyants vives se l’anomena ‘àrea basal de l’albeca’ (ABA). Es disposa de poques dades sobre l’àrea basal de l’albeca, en part per les dificultats que hi ha per a diferenciar-la. La relació entre índex foliar i àrea basal de l’albeca als boscos de “guindo” (Nothofagus betuloides) del S de l’Argentina és de 0,32 m2/cm2 a les zones més protegides, situades a les parts baixes de les muntanyes, però tan sols de 0,09 m2/cm2 a les zones subalpines de bosc, on les condicions són més dures. A Nova Zelanda s’han trobat valors semblants per a altres espècies de faig austral (Nothofagus), cosa que significa que l’àrea basal de l’albeca és més gran com més adverses són les condicions. Tot i això, per al “golden chinquapin” (Castanopsis [=Chrysolepis] chrysophylla), una fagàcia caducifòlia dels confins meridionals de les selves plujoses de la costa NW de l’Amèrica del Nord, el quocient és de 0,46 m2/cm2 en condicions benignes, i arriba a valors de 0,65 m2/cm2 en boscos monsònics de tec (Tectona grandis).
És apropiat fer la distinció entre àrea basal i àrea basal de l’albeca, per les implicacions que això té en el flux d’aigua que circula pel tronc i en la seva velocitat. En un bosc estudiat a la laurisilva canària, la transpiració anual és de 636 mm, l’índex foliar de 7,8 m2/m2, i l’àrea basal de 33,7 m2/ha. Si es considera que tota l’àrea basal és àrea basal de l’albeca, la velocitat amb què l’aigua remuntaria el tronc seria de 2,15 cm/h; si es considera que l’àrea basal de l’albeca és el 20% de l’àrea basal, però, la velocitat del flux seria de 10,77 m/h. Això significa que la velocitat del flux de l’aigua a través del tronc és 2 314,14 o 11 572,84 vegades superior, respectivament, segons el supòsit, a la velocitat d’evaporació.
La raresa o absència de caducifolis
A l’hemisferi meridional, les selves temperades, fins i tot les que es desenvolupen al continent sud-americà a latituds elevades (55˚S), es caracteritzen per l’absència gairebé absoluta de caducifolis. En efecte, estan formades per angiospermes perennifòlies i coníferes termòfiles. El color sempre verd d’aquests boscos contrasta amb els tons grocs, marrons, ataronjats i vermellosos dels extensos boscos caducifolis de l’hemisferi nord. Justament, el caràcter decidu o perennifoli dels arbres d’un i altre hemisferi reflecteix les diferències fonamentals dels ambients en què els arbres evolucionaren, situats a banda i banda del cinturó de boscos tropicals.
En general, les espècies d’angiospermes perennifòlies de les selves de l’hemisferi sud són poc resistents al fred, ja que als escenaris on evolucionaren no els féu falta cap adaptació en aquest sentit. La distribució asimètrica dels continents i oceans en un i altre hemisferi queda reflectida també en la distribució de la vegetació. A l’hemisferi meridional no hi ha grans extensions continentals com a l’hemisferi septentrional, on hi ha terres molt allunyades de la influència de la mar i el clima continental fred afavoreix el desenvolupament de coníferes i caducifolis. A l’Amèrica del Sud, les espècies caducifòlies de faigs australs (Nothofagus), com per exemple la lenga (N. pumilio) o el “ñirre” (N. antarctica), es troben restringides a les àrees més fredes, als vessants orientals dels Andes i a les parts altes de les muntanyes.
A Nova Zelanda, només el 4% de les espècies són caducifòlies. En aquest arxipèlag, les variacions més àmplies de temperatura es produeixen a l’interior i a la part oriental, i a l’W i al S és on la biodiversitat de la laurisilva és més gran. La distribució local de moltes espècies es correspon amb la seva resistència a les temperatures baixes, i així, per exemple, el faig austral negre (Nothofagus solanderi) no creix als fons de les valls, ja que les temperatures hi solen ser més baixes que a les parts altes de les muntanyes, on aquesta espècie constitueix el límit altitudinal del bosc.
En moltes localitats d’Austràlia, les variacions de temperatura són més grans que a Nova Zelanda, i les espècies d’eucaliptus (Eucalyptus) de les selves temperades són més resistents al fred que els faigs australs (Nothofagus) neozelandesos. Tanmateix, la majoria de coníferes australianes mostren una limitada resistència a les baixes temperatures —comparable a la que presenten les coníferes menys resistents del Japó—, com és el cas d’algunes espècies subtropicals de podocarps (Podocarpus) que ja pateixen danys a temperatures de –13˚C. En conseqüència, totes aquestes espècies estan confinades a les àrees de clima més benigne.
Al llarg de les costes centrals i meridionals de Xile, les temperatures no varien gaire al llarg de l’any i, per això, en aquesta zona dominen els arbres i els arbustos perennifolis. Les fulles de les proteàcies d’aquestes regions resisteixen temperatures de fins a –7 i –9˚C, i fins i tot les gemmes més baixes del “canelo” xilè (Drimys winteri), una winteràcia arbustiva, resisteixen temperatures semblants; els faigs australs (Nothofagus) perennifolis, que dominen als boscos de latituds més meridionals, són fins i tot més resistents al fred que el “coihue” comú (N. dombeyi), de la selva valdiviana, a Xile.
Uns estudis duts a terme per diversos autors posen de manifest que les espècies perennifòlies que resisteixen temperatures baixes (de fins –30˚C) no han evolucionat a les zones de clima càlid. Però hi ha una excepció, els rododendres (Rhododendron), representats per unes 600 espècies que viuen sota règims climàtics molt variats. D’altra banda, no és fàcil explicar per què les espècies resistents a les baixes temperatures que creixen perfectament als parcs i jardins de les zones temperades no formen part de la vegetació d’aquests climes més càlids. És probable que la competència amb altres espècies determini en gran manera la seva àrea de distribució.
Resistència a les glaçades
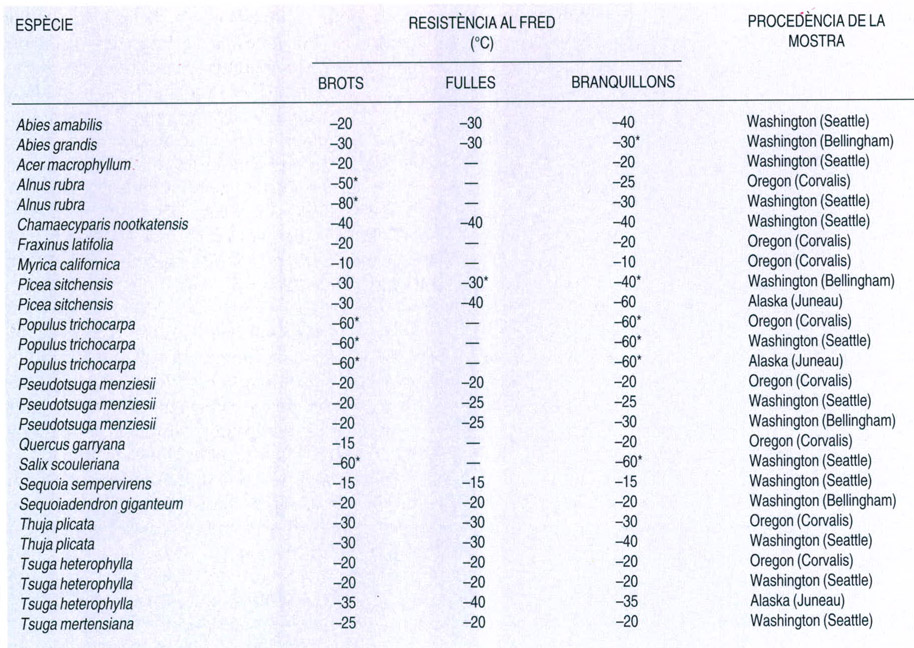
A partir d’A. Sakai & W. Larcher, 1987
Resistència a les glaçades de diferents espècies arbòries de la costa pacífica de l’Amèrica del Nord (els asteriscos indiquen valors de temperatura als quals els diversos teixits vegetals encara no resulten danyats). La majoria de coníferes i d’angiospermes d’aquesta regió pateixen danys en alguns dels seus teixits a temperatures inferiors als –20 o als –30°C. Només els salzes (Salix) i els pollancres (Populus) en són una excepció, ja que resisteixen bé fins als 60°C sota zero. En general, les espècies que habiten latituds més nòrdiques, les d’Alaska, resisteixen temperatures més baixes, i fins i tot dins d’una mateixa espècie són més resistents les poblacions més septentrionals. És el que passa, per exemple, en el cas de la pícea de Sitka (Picea sitchensis) i del hemlock occidental (Tsuga heterophylla). En algunes espècies, els diferents tipus de teixits vegetals mostren una tolerància idèntica, o semblant, a les glaçades, com és el cas de les sequoies (Sequoia sempervirens i Sequoiadendron giganteum) i del cedre groc o “Alaska yellow-cedar” (Chamaecyparis nootkatensis). Altres arbres, en canvi, tenen un determinat tipus de teixit especialment resistent, com ara els brots dels verns (Alnus) i dels branquillons de la pícea de Sitka.
Les coníferes han seguit el mateix patró que les angiospermes. Molt resistents a les baixes temperatures (fins a –70˚C), les coníferes han evolucionat a la zona boreal de l’hemisferi nord. Les de les selves temperades, però, tenen un grau de resistència menor, i no toleren temperatures inferiors a –30˚C. Algunes espècies de podocarps (Podocarpus) que viuen als cims de les muntanyes només aguanten temperatures de –23˚C, molt menys fredes que les que toleren les pinàcies de la taigà. Però les resistents coníferes de l’hemisferi nord no tenen èxit als climes de les selves temperades, i, a més, tampoc no poden superar les barreres dels tròpics.
A l’Amèrica del Nord, les coníferes i les angiospermes perennifòlies que no suporten temperatures inferiors a –15˚C estan confinades a les zones temperades sud-orientals i als boscos de la costa nord-occidental del Pacífic. És interessant remarcar que tant les coníferes, entre les quals el pi teia o pi d’Elliott (Pinus elliottii), el pi “loblolly” (P. taeda) o els taxodis (Taxodium), com les angiospermes perennifòlies, per exemple les magnòlies (Magnolia) i les alzines o “live oaks” (Quercus perennifolis) de les selves mississipianes, són força menys resistents al fred que les espècies decídues, molt abundants en aquesta zona. Les coníferes i les espècies planifòlies dels boscos vancouverians, que s’estenen des d’Alaska fins al N de Califòrnia, mostren en canvi una major resistència (aguanten temperatures properes als –30˚C). D’aquestes, la menys tolerants és la sequoia costanera (Sequoia sempervirens), de Califòrnia, les fulles, brots i gemmes de la qual no suporten temperatures inferiors als –20˚C. Les coníferes de la zona septentrional de la regió i de les terres interiors són més resistents que les que habiten prop de la costa, on les temperatures hivernals són molt més suaus.
De fet, és una característica força generalitzada que les angiospermes decídues tolerin millor les baixes temperatures que les perennifòlies. Un dels casos més interessants d’adaptació a grans canvis espacials i temporals és l’evolució del caràcter decidu de moltes espècies vegetals. Aquestes espècies aparegueren als boscos plujosos temperats de les latituds mitjanes de l’hemisferi septentrional durant el Cretaci, en resposta a condicions d’estrès hídric moderat durant els mesos hivernals, secs però sense glaçades. A mesura que la temperatura mitjana de la Terra anà baixant, les espècies caducifòlies s’anaren estenent cap a latituds superiors, on el clima era més fred i els canvis en el fotoperíode al llarg de l’any més marcats. Així, el caràcter caducifoli pot considerar-se com una preadaptació a les condicions ambientals dels nous hàbitats. Adquirit en zones càlides, serví per a colonitzar latituds superiors. Les espècies decídues d’hivern, que perden les fulles quan l’absorció de nutrients és més difícil i les temperatures són més baixes, tindrien avantatge durant aquesta estació, i per això, amb el refredament general del clima durant el Quaternari, la pèrdua del fullatge es convertí en una adaptació a les baixes temperatures hivernals.
El caràcter caducifoli o perennifoli és una propietat de les fulles considerades en el seu conjunt, mentre que la longevitat de les fulles és una característica individual. Normalment s’assumeix que les espècies perennifòlies tenen fulles que viuen més que les de les espècies caducifòlies, però no sempre és així. Un arbre perennifoli pot canviar les fulles més d’una vegada a l’any; en aquest cas, encara que l’arbre continuï sent perennifoli, la longevitat de les fulles no supera l’any. Quan les condicions ambientals permeten un període de creixement llarg, com sol ser el cas de les selves subtropicals i temperades, la majoria de les espècies arbòries són perennifòlies, independentment de la longevitat de les seves fulles. A les laurisilves, com a resposta a les variacions estacionals del clima, les espècies caducifòlies que acostumen a ocupar els estrats superiors del dosser perden les fulles anualment.
Als ambients on la disponibilitat de nutrients és baixa, el fet que les fulles presentin una longevitat major es tradueix també en un increment de l’eficiència en l’ús dels recursos. I, quan el balanç és positiu, encara que les taxes de fotosíntesi siguin baixes, la vida de les fulles acostuma a ser més llarga. A la selva tropical també s’observa que els arbres dels estrats inferiors, on la llum és un factor limitador, retenen les fulles durant més temps, i és lògic pensar que les seves taxes de fotosíntesi siguin més baixes. Així, el caràcter perennifoli o caducifoli, igual que les dimensions foliars, està relacionat amb el cost de la fabricació de noves fulles i amb els guanys que se’n puguin obtenir, atès que cal considerar les fulles com les fàbriques de recursos dels productors primaris. Es considera, doncs, que el caràcter caducifoli està associat amb una certa estacionalitat del clima. I les espècies perennifòlies, en efecte, són predominants als tròpics humits i decreixen en importància a mesura que augmenta la latitud, alhora que es produeix un augment de la presència d’espècies caducifòlies. En latituds elevades i temperatures molt baixes, com passa als boscos boreals, predominen les coníferes.
Dinàmica foliar
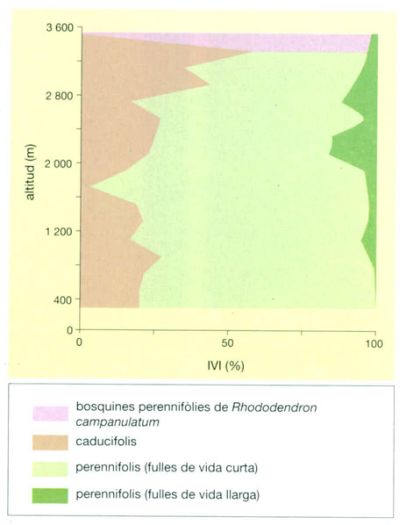
Idem, a partir de S.P. Singh i altres, 1994
En diversos boscos del vessant meridional de l’Himàlaia central es dugué a terme el 1994 un estudi molt complet sobre dinàmica foliar. Els arbres d’aquests boscos es dividiren en tres tipus: caducifolis, perennifolis amb fulles de vida curta (d’un any o poc més) i perennifolis de fulles de vida llarga (de diversos anys). També s’estudiaren les bosquines del rododendre Rhododendron campanulatum —perennifolis de fulles de vida curta— situats per sobre els boscos. Al llarg d’un gradient altitudinal (aproximadament dels 400 als 3 600 m) es quantificà la dinàmica foliar dels arbres mitjançant l’índex del valor d’importància o IVI (Importance Value Index), que suma la densitat relativa més la importància relativa més la freqüència relativa del fullatge de les diverses espècies. L’estudi mostrà que als boscos de les altituds més baixes dominaven les espècies perennifòlies de fulles de vida curta, mentre que a partir dels 1 800 m eren cada vegada més abundants els caducifolis i els perennifolis de fulles de vida llarga. No es disposa de dades d’altres laurisilves, però a la Terra del Foc i al llarg d’un gradient altitudinal de boscos de “guindo” (Nothofagus betuloides) s’ha observat que a les zones baixes de les muntanyes les fulles són retingudes fins a tres o quatre anys a les branques, mentre que a les zones altes s’hi mantenen fins a set anys.
L’estructura horitzontal
Una selva temperada madura, quan se n’observa una gran superfície, consisteix en molts petits rodals que es troben en diferents estadis de desenvolupament, amb arbres de totes mides i, probablement, també de totes les edats. Aquesta estructura en mosaic crea una distribució horitzontal d’ambients diversos que es combina amb una estructura vertical en la qual se succeeixen diferents estrats. Així, sovint s’hi poden observar espècies pioneres en espais oberts i clarianes, mentre que les espècies dominants creixen a les parts més velles. En un bosc madur, aquesta estructura en què coexisteixen múltiples estadis provoca també una gran variància horitzontal de la mida i edat dels arbres i de la seva biomassa.

Àrea basal i densitat de diversos gèneres arboris i arbustius en una laurisilva canària segons el diàmetre del tronc a l’alçada del pit (en cm). Els valors foren mesurats al cap de 30 anys d’haver-se talat el bosc i després de deixar que es regenerés al seu aire per rebrot. Els estrats arbori i arbustiu d’aquesta laurisilva eren formats per llorers (Laurus, 38%), brucs (Erica, 27%), Myrica (17%), Persea (14%) i grèvols (Ilex, 4%). Tanmateix, la major part de l’àrea basal corresponia a Myrica (38%), seguit de Laurus (26%), Erica (20%), Persea (14%) i Ilex (2%). El diàmetre dels arbres variava entre 4 i 46 cm, però els que tenien un diàmetre de 14 cm eren els més nombrosos, i els de 16 cm de diàmetre eren els que tenien l’àrea basal més gran. Tal com mostra el gràfic, tant el diàmetre com l’àrea basal mostraven una distribució marcadament asimètrica.
Jordi Corbera, a partir de D. Morales i altres, 1996
A les laurisilves verges i que no s’han vist afectades per pertorbacions naturals recents, els gràfics de distribució dels diàmetres dels troncs, mesurats a 1,30 m de la base, acostumen a prendre la forma de J invertida. És a dir, que els arbres grans són poc abundants, mentre que els de mida mitjana, i especialment els petits, són molt nombrosos. Això explica que, a les laurisilves, on la contribució dels arbres petits, que són els més abundants, és baixa, alguns paràmetres estructurals com la biomassa o l’àrea basal total tinguin valors poc elevats. L’estructura per edats és pràcticament impossible d’establir per a les laurisilves subtropicals, ja que el període de creixement es prolonga al llarg de tot l’any i els anells anuals de creixement no es diferencien amb claretat en el tronc; a les laurisilves de la zona temperada, en canvi, les discontinuïtats en el creixement, derivades d’una estacionalitat climàtica més marcada, acostumen a quedar reflectides en els anells anuals de creixement de moltes espècies, que a les coníferes i els faigs australs són clars.
Encara que amb menys claredat que les distribucions per edat, les distribucions per mides també reflecteixen la història de les pertorbacions, naturals o antròpiques, que ha sofert un bosc. Quan s’han realitzat aclarides selectives i s’han talat els arbres d’unes dimensions determinades, la disminució de la freqüència d’arbres de les classes que han estat objecte d’explotació provoca irregularitats en les distribucions de diàmetres; així, en aquests boscos es produeix un augment espectacular dels arbres més petits, que ocupen l’espai deixat pels arbres talats. En altres casos, quan els boscos són destruïts completament, la distribució de freqüències que s’observa al cap d’unes quantes dècades presenta una forma gairebé acampanada, asimètrica i esbiaixada cap a l’esquerra; aquest tipus de distribució reflecteix l’evolució simultània de tots els arbres de les diferents espècies instal·lades després de la pertorbació. Les variacions en aquest tipus de distribució són comunes, per exemple, a la laurisilva canària, allà on s’han practicat tales arreu i la regeneració de les espècies principals es produeix per rebrot; es tracta de boscos joves, en els quals no sembla haver-se arribat encara a una estructuració clara del cobricel en estrats diferenciats. En aquests casos, a causa de l’abundància d’arbres petits, les densitats acostumen a ser molt altes, i sovint superiors a 1 500 arbres/ha. En algunes laurisilves japoneses s’arriba als 4 000 i fins i tot als 6 300 arbres/ha.
El paper ecològic dels arbres morts
Per paradoxal que pugui semblar, els arbres morts donen vida al bosc: són aliment i refugi per a molts animals, constitueixen el substrat on les plantes poden créixer o recolzar-se i, en últim terme, com es veurà més endavant, són un component important de la necromassa i una font d’humus que enriqueix el sòl forestal. Una selva temperada madura es caracteritza, en definitiva, per la presència d’un nombre considerable de grans troncs morts ajaguts per terra, amarats d’aigua i recoberts de briòfits, falgueres i també fongs, així com d’un nombre igualment considerable de troncs morts en peu. En caure, aquests troncs obren clarianes, i en aquestes es desenvolupen millor els estrats inferiors del sotabosc; a més, afegeixen heterogeneïtat espacial i desencadenen l’establiment de l’organització de l’estructura en mosaic característica de les selves temperades. En absència de pertorbacions intenses, la selva es pot mantenir indefinidament en aquest estat estacionari dinàmic.
Un arbre, com qualsevol ésser viu, pot morir en molts moments del seu cicle biològic i per múltiples raons. La majoria moren quan tot just són plançons sortits de la llavor, molts quan només són arbrissons i víctimes d’insectes defoliadors, de fongs patògens o de les condicions ombrívoles del sotabosc. La taxa de mortalitat anual disminueix quan els arbres arriben a la maduresa i, en moltes espècies, els adults poden viure centenars i fins i tot milers d’anys abans de morir inevitablement i afegir-se al volum de fusta morta de l’ecosistema forestal. Encara que les investigacions sobre la importància de la fusta morta, pel que fa al seu paper en l’ecosistema, són encara a les beceroles, sembla probable que els microhàbitats més importants siguin els creats per la fusta morta procedent d’arbres grans i vells.
Els arbres adults dels boscos naturals poden morir de moltes maneres diferents. Alguns moren com a conseqüència de l’acció directa d’insectes i fongs patògens o altres plagues. Els rigors del clima, com ara les sequeres o les gelades, també causen la mort d’arbres en molts ecosistemes, però concretament al bioma temperat i humit de les selves temperades, el clima acostuma a ser un factor de mortalitat poc important, tret que es donin circumstàncies excepcionals o quan es tracta d’indrets de molta altitud. Una part dels arbres moren o queden fatalment malmesos a causa del foc o d’un llamp. Hi ha vegades que fins i tot un arbre sa pot caure a causa d’una tempesta violenta; en aquestes ocasions, es dóna el cas que alguns exemplars sobreviuen però queden tocats, sovint inclinats, mentre que altres viuen encara el temps just per a produir una darrera tongada de llavors i després moren. La mort també pot esdevenir-se lentament, de manera que un arbre viu pot tenir ‘fusta morta’ en alguna branca o a la part superior del tronc, constituint un microhàbitat durant molts anys, mentre de terra estant sembla que estigui del tot sa.
Les causes de mort acostumen a variar segons els boscos. En un bosc de hemlock occidental (Tsuga heterophylla) i pícea de Sitka (Picea sitchensis) de la costa d’Oregon, es trobà que el vent era la causa de mort del 83% dels arbres adults, mentre que en un bosc de hemlock occidental i avet de Douglas (Pseudotsuga menziesii) de la serralada de les Cascades no arribava a causar-ne ni el 50%. Hi ha vegades que en un sol arbre actuen diferents factors. Els arbres afeblits pels insectes defoliadors, per exemple, són més sensibles a l’acció dels insectes barrinadors, que al seu torn deixen l’interior dels arbres desprotegit i a l’abast dels atacs fúngics.
Les pertorbacions i l’estructura en mosaic de la vegetació
En absència de pertorbacions majors, que no siguin la mort i la caiguda d’arbres aïllats o de grups d’arbres, a la major part del bioma el bosc arribaria a la seva edat màxima i es regeneraria a través de la mort dels arbres vells i la seva substitució per arbres joves. Però ni tan sols les laurisilves subtropicals estan exemptes de patir pertorbacions de gran intensitat. Els incendis, els huracans i els ciclons són freqüents a moltes selves subtropicals (els huracans principalment a les regions més humides i els incendis a les més eixutes) i, en algunes zones de la laurisilva xilena i neozelandesa, els terratrèmols i l’activitat volcànica arriben a sovintejar tant que els boscos no poden assolir fases gaire avançades de la successió.
La incidència de les pertorbacions intenses rejoveneix àrees poc o molt extenses, de manera que, a més gran escala, el seu efecte es tradueix en l’aparició d’un mosaic de selves en diferents estadis de la successió, que presenta diferències fisiognòmiques i florístiques entre els diferents claps que l’integren. A les selves valdivianes, per exemple, els faigs australs (Nothofagus), que no toleren l’ombra, no es regeneren sota el dens cobricel de la selva madura, que amb prou feines deixa passar la llum i més aviat afavoreix la regeneració dels laurifolis. Tanmateix, com confirmen els rodals en què aquests arbres formen boscos coetanis, les espècies de faig austral de la selva valdiviana, concretament el “roble pellín” (N. obliqua), la lenga (N. pumilio) i el “ñirre” (N. antarctica), es regeneren bé als espais denudats per grans catàstrofes.
Els incendis, en canvi, semblen afavorir més les coníferes. Així, la distribució del “pehuén” (Araucaria araucana) coincideix amb la situació de molts volcans recentment actius. Les erupcions volcàniques dipositen gran quantitat d’escòria sobre els boscos i provoquen incendis, però, no obstant això, els exemplars grans de “pehuén” sobreviuen i actuen com a colonitzadors inicials gràcies a la seva bona adaptació al foc. Aquesta adaptació es tradueix en una sèrie de característiques específiques: una escorça de fins a 20 cm de gruix que protegeix el tronc dels focs de superfície; la capacitat —a diferència d’altres coníferes— de formar gemmes a les axil·les de les fulles; la de produir brots epicòrmics després del foc o d’haver-se produït altres ferides o, en fi, la presència de densos verticils de fulles que protegeixen les gemmes terminals i permeten que aquestes puguin continuar creixent després del foc. Un cop acabat l’incendi, es pot formar un bosc mixt amb “coihue” comú (Nothofagus dombeyi). Una dinàmica similar a la que es produeix després d’un incendi és la que es dóna quan hi ha grans lliscaments. Així, els grans moviments de terra sovint deixen pas a boscos semienterrats, arbres trencats i, també, alguns individus supervivents. Aquests arbres actuen de planters i produeixen nombroses plàntules i arbres joves que s’estableixen després de la pertorbació. Tots aquests mecanismes de resistència al foc i a diferents tipus de pertorbacions també existeixen en altres espècies d’araucariàcies, per exemple el “pino Paraná” o “kuri’y” (Araucaria angustifolia), que tolera malament l’ombra però s’estableix amb èxit als “campos”, i s’hi regenera bé després d’haver-se produït pertorbacions regulars com poden ser els incendis, als quals acostuma a sobreviure.
El metabolisme forestal
Les selves temperades, com tots els boscos, es caracteritzen per la gran acumulació de biomassa que concentren en les seves parts llenyoses, que constitueixen més del 90% de l’esquelet de l’estructura tridimensional del bosc. Aquesta arquitectura sosté una quantitat tampoc negligible de biomassa foliar, que, gràcies a la seva activitat fotosintetitzadora, és la que transforma l’energia solar en energia química, i alhora constitueix l’hàbitat de molts tipus d’animals i plantes.
L’acumulació de biomassa
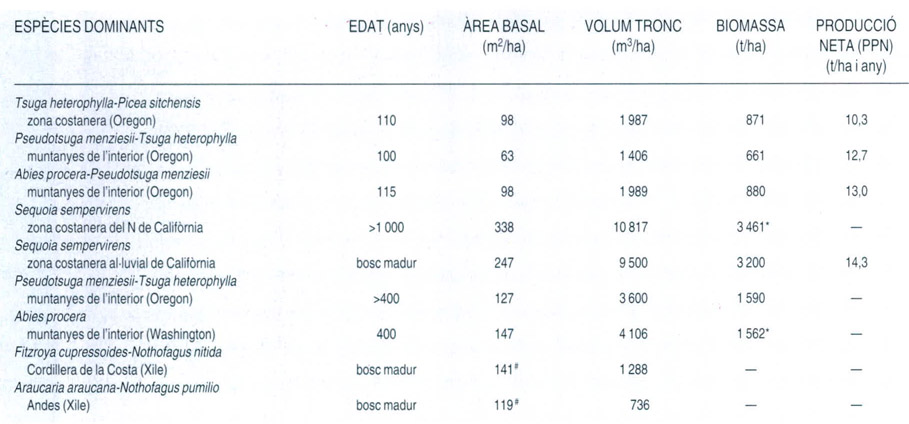
Biomassa i producció primària neta (PPN) d’alguns boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord (les dades que es refereixen solament a les coníferes van marcades amb #, mentre que * indica valors de biomassa calculats només per als troncs). El valor de PPN és de la màxima importància per al funcionament de l’ecosistema, ja que determina tota l’estructura de les xarxes tròfiques. La PPN es tradueix en l’increment de biomassa i en la renovació de les estructures dels vegetals verds (tronc, escorça, branques, fulles, arrels, radícules, flors, fruits, etc.). Els animals herbívors que s’alimenten d’aquestes estructures consumeixen directament una petita part de la PPN, mentre que una altra part, molt més important, va a parar al compartiment detrític del sòl en forma de material vegetal mort, que serà processat per organismes diversos. Així, per a cada interval de temps, la PPN és igual a l’increment de biomassa acumulada (∆B) més el consum dels herbívors (C) més la matèria orgànica que va a parar al compartiment detrític (D). Per tant, PPN = ∆B + C + D.
A partir de dades proporcionades per l’autor
La quantitat de biomassa de les selves temperades varia molt de les unes a les altres. En general, l’alçària del bosc i la biomassa acostumen a augmentar seguint un gradient latitudinal de temperatura, des de la zona subàrtica fins a la zona tropical. Tanmateix, en contra d’aquesta tendència general, als boscos plujosos de coníferes, que tenen un estrat arbori superior de 60 o 80 m, l’acumulació de biomassa és molt gran. Als boscos madurs de sequoia, comptant només la biomassa dels troncs, ja s’assoleixen valors de 3 461 t/ha en pes sec, i sumanthi la de les fulles, les branques i les arrels és de 4 525 t/ha. Aquests són boscos que creixen en òptimes condicions de clima i sòl, però en altres de no tan afavorits de la costa NW de l’Amèrica del Nord s’assoleixen també valors molt alts, si es comparen amb els dels boscos temperats o les selves plujoses tropicals. Seguint el gradient de vegetació de la costa cap a l’interior es produeix una disminució de la biomassa, de l’àrea foliar i d’altres característiques estructurals i funcionals de l’ecosistema.
Un dels estudis més detallats realitzats en aquest gradient W-E, entre els paral·lels 44 i 45˚N, comprèn des dels boscos dominats pel hemlock occidental (Tsuga heterophylla) de la zona costanera, fins als de la zona oriental on, en condicions xeròfiles, hi predominen els boscos de pi groc o pi ponderosa (Pinus ponderosa). Els resultats mostren que la biomassa dels boscos costaners dominats pel hemlock occidental (comptant només l’acumulada als troncs) està compresa entre les 865 i les 1 492 t/ha; aquests són valors molt alts, força superiors als valors de biomassa que s’han quantificat per als boscos boreals (422 t/ha), les laurisilves (575 t/ha), les selves tropicals (420 t/ha), i també als valors de biomassa estimats als boscos plujosos temperats freds de tsuga comuna (Tsuga sieboldii) del Japó, que són de 600 t/ha. En canvi, seguint el gradient assenyalat, es pot observar que els boscos de la muntanya mitjana plujosa de l’interior tenen biomasses compreses entre les 449 i les 528 t/ha, mentre que els boscos subalpins de hemlock californià o avet de Mertens (T. mertensiana) i els de pi groc o ponderosa tenen biomasses més baixes, de 278 i 136 t/ha, respectivament. També és important, d’altra banda, la biomassa acumulada a les fulles. Als boscos costaners on predomina el hemlock occidental, la biomassa estimada en el mateix estudi és de 23 a 32 t/ha i l’àrea foliar varia entre 10 i 20 m2/m2; als boscos situats a l’interior i als vessants occidentals de les serralades costaneres, en canvi, els valors de biomassa foliar estan compresos entre les 12 i les 18 t/ha.
Fonamentalment, el que determina l’elevada quantitat de biomassa d’aquests boscos és el creixement continuat en diàmetre i alçada, durant moltíssims anys, de les espècies arbòries dominants. A aquestes característiques cal afegir-hi la seva capacitat per acumular i mantenir una biomassa foliar molt alta i, com a conseqüència, índexs foliars també força elevats. En general, els boscos d’espècies planifòlies assoleixen els valors màxims de biomassa i àrea foliar en pocs anys, mentre que els boscos de coníferes triguen unes quantes dècades. S’ha calculat, per exemple, que els boscos d’avet de Douglas (Pseudotsuga menziesii) atenyen un valor màxim de biomassa i àrea foliar entre els 40 i els 60 anys i, si les condicions són desfavorables, encara més tard.
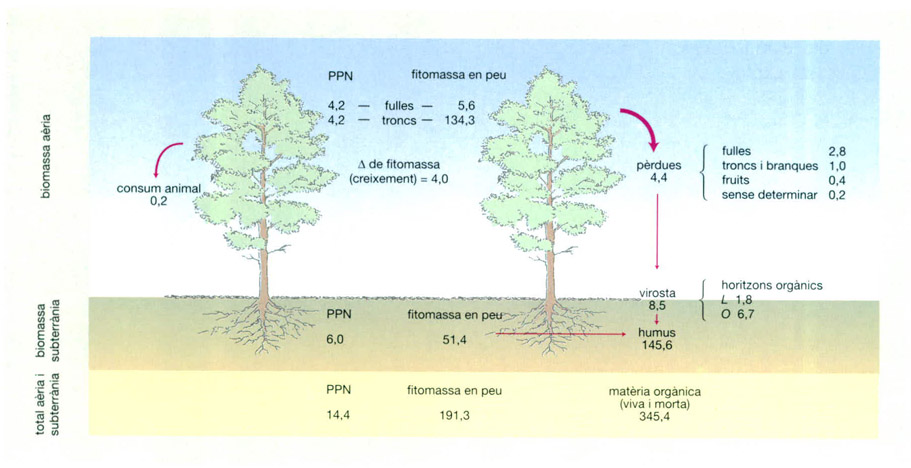
Biomassa i flux de matèria en una laurisilva semicaducifòlia de la conca hidrogràfica de Coweeta (Carolina del Nord, EUA) (els valors de biomassa es donen en t/ha i els de producció en t/ha i any). Uns experiments duts a terme en aquest bosc demostraren que cada any les aportacions de material detrític igualaven la quantitat de matèria orgànica morta que es descomponia, de manera que s’arribava a un estat estacionari. El temps de renovació de la matèria es calculà en 1,9 anys, és a dir, els nutrients trigaven gairebé dos anys a completar el cicle. Alguns, com el magnesi i el potassi, tenien temps de renovació inferiors (1,4 i 1,5 anys respectivament) mentre que els d’altres eren superiors (2,2 anys el nitrogen, 2,6 el calci i 2,9 el fòsfor). D’altra banda, l’acumulació de nutrients en la fitomassa era molt elevada, com se suposa que succeeix també en altres laurisilves. Així, les quantitats de fòsfor, potassi i calci acumulats en els diferents compartiments vegetals eren superiors a les quantitats que es poden trobar d’aquests nutrients al sòl. Tan sols el nitrogen i el magnesi es presentaven al sòl en quantitats superiors. La capa de virosta concentrava entre l’1,2 i el 7,6% del total dels nutrients, i, tret del nitrogen, es calculà que la resta podien ser reposats per l’erosió de la roca mare. La concentració diferencial de cada nutrient als diversos compartiments vegetals, les dimensions d’aquests compartiments i les taxes de renovació de cada nutrient eren aspectes determinants en la dinàmica del bosc.
Jordi Corbera, a partir de Monk & Day, 1988
A diferència dels de les selves plujoses de coníferes, els valors de biomassa aèria de les laurisilves subtropicals són baixos. La biomassa forestal aèria, tal com sol succeir a les regions humides, constitueix aquí més del 80% de la biomassa total, que es concentra als troncs i les branques. En aquestes laurisilves subtropicals, amb un estrat arbori d’uns 25 m d’alçària, els valors més alts estimats de biomassa no sobrepassen les 400 o 450 t/ha, i normalment són inferiors. A dues mostres de la selva de la regió del Paranà, al departament paraguaià de Misiones, no explotades durant les últimes dècades, s’obtingueren estimacions de biomassa de 251,6 i 211,2 t/ha respectivament, i en una d’explotada recentment de la mateixa regió el valor estimat fou de 210 t/ha. A la conca de Coweeta, a l’extrem meridional dels Apalatxes, les estimacions de biomassa aèria per a diferents comunitats forestals estudiades oscil·la entre les 126 i les 420 t/ha. A les laurisilves de les zones temperades i a les himalaianes s’assoleixen valors semblants als de la selva tropical plujosa (que són d’entre 300 i 650 t/ha), bé que sense arribar mai als valors de biomassa que s’acumulen als boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord. Però els valors de biomassa més elevats de totes les estimacions fetes en laurisilves s’assoleixen als boscos de “moru” (Quercus floribunda) de Kumaun, als vessants meridionals de l’Himàlaia, amb 782 t/ha, quantitat superior a les 710 t/ha dels boscos monsònics de sal (Shorea robusta) de la mateixa zona, o a les 744 t/ha estimades als boscos de tec (Tectona grandis) del N de l’Índia, fisiognòmicament semblants a les selves tropicals plujoses però sotmesos a un règim estacional de pluges força extrem. Pel que fa a la biomassa foliar, la variabilitat és menor entre els diferents tipus forestals. Al Japó, per exemple, la biomassa foliar mitjana d’una laurisilva s’estima en 8,6 ± 2,5 t/ha.
La relació entre biomassa aèria i subterrània (T/R) varia a les selves temperades entre 3 i 7, però a les laurisilves el marge d’oscil·lació d’aquest quocient es manté entre 4 i 6. A Minamata, al SW del Japó, en el marc del Programa Biològic Internacional (International Biological Programme, IBP), es realitzà un estudi sobre biomassa i productivitat de les laurisilves japoneses (l’únic estudi de l’IBP referit a una laurisilva). Es calculà que, en els rodals estudiats del bosc de Minamata, la biomassa total i la relació T/R eren respectivament de 400 t/ha i d’entre 3,5 i 4,4. La biomassa de les arrels (exceptuada la rabassa) era pràcticament la mateixa que la de les branques; les arrels fines (les d’un diàmetre inferior a 2 mm) suposen una biomassa de 0,8 a 0,9 t/ha, fet que implica que menys d’1 t/ha d’arrels fines subministra tota l’aigua i tots els nutrients necessaris a les 400 t/ha de biomassa forestal aèria existents.
Estructura del bosc de Minamata

Jordi Corbera, a partir de Nagano & Kira, 1978
Estructura del bosc de Minamata (Japó) representada segons la distribució vertical, en estrats de 2 m d’alçada, de l’índex foliar i de la biomassa de la fusta. L’índex foliar per hectàrea i la biomassa per hectàrea d’aquests estrats indiquen la distribució de la biomassa al bosc. És manifest que l’índex foliar és màxim al cobricel i als estrats més baixos del sotabosc, mentre que la biomassa s’acumula preferentment als estrats més baixos, que són els que han crescut més; a la base de les branques del cobricel el pes d’aquestes supera el dels troncs. Una altra manera de quantificar la biomassa consisteix a fer-ho en termes de densitat mitjana de biomassa a l’espai forestal, és a dir, fent el quocient entre la biomassa aèria per unitat de superfície i l’alçària mitjana del bosc. Les laurisilves madures, tant temperades com subtropicals, tenen una densitat mitjana de biomassa d’1,6 o 1,7 kg/m3, valor que en les primeres etapes de la successió es redueix a 0,5-1,0 kg/m3. Els valors màxims, però, poden ser molt més alts; en una bosquina costanera d’“ubamegashi” (Quercus phylliraeoides) del S del Japó, per exemple, s’han enregistrat valors de prop de 2,5 kg/m3, semblants als calculats en boscos plujosos de coníferes, i en un bosc de sequoies (Sequoia) de Califòrnia, amb un estrat arbori de 88 m d’alt, es calculà que la densitat mitjana de biomassa arribava als 4 kg/m3.
La producció primària
La biomassa (B) conté energia en forma d’enllaços químics, i també es pot expressar en calories o en funció del seu contingut en carboni. No obstant això, la biomassa no és en si mateixa una bona mesura del flux d’energia, i per això cal expressar-la amb relació al temps. D’aquesta manera sí que s’aconsegueix una bona mesura del flux energètic, és a dir, la producció (P), que es pot expressar en les mateixes unitats que la biomassa, però per unitat de temps; per als ecosistemes forestals s’utilitza l’any, i la producció s’expressa en t/ha i any. Cal distingir entre la producció primària bruta (PPB) i la producció primària neta (PPN), ambdues expressades en unitats de pes sec, carboni, o calories per unitat de superfície i temps. La producció primària neta és allò que queda de la producció primària bruta un cop se n’han descomptat les despeses respiratòries durant la fotosíntesi, les de manteniment de les estructures existents i de la síntesi de teixits nous; per tant, es pot considerar que la producció primària neta és la velocitat a la qual els productors primaris acumulen matèria orgànica.
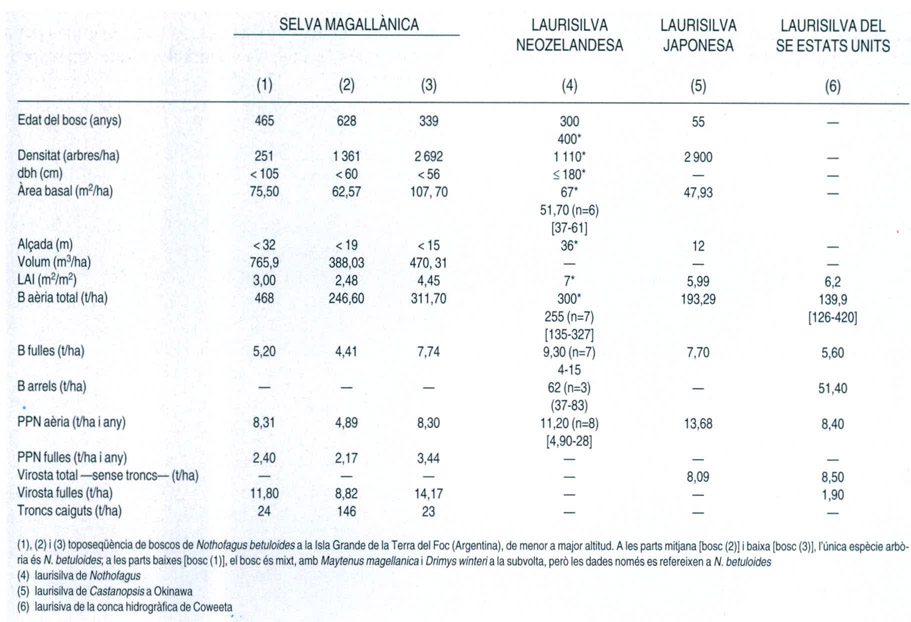
L’extrema variabilitat de les selves temperades, així com l’escassetat de dades quantitatives disponibles, queden ben paleses en aquesta taula que compara diferents paràmetres referents a estructura, biomassa i producció d’algunes d’aquestes selves. Les xifres entre claudàtors indiquen l’interval de variació dels valors de les mostres, mentre que els valors entre parèntesis corresponen a la mida de la mostra. Com ja s’ha dit, cal tenir en compte que la procedència de les dades és diversa i en dificulta la comparació.
A partir de dades de E. Gutierrez (selva magallànica), P. Wardle et al. (laurisilva neozelandesa), D. L. DeAngelis et al. (laurisilva japonesa) i C. D. Monk et al. (laurisilva del SE dels Estats Units)
La proporció de PPB que es destina a la respiració varia en funció de la temperatura, de manera que als ecosistemes tropicals, on el metabolisme és més accelerat, és més alta que a les laurisilves subtropicals, i en aquestes més alta que a les laurisilves de les zones temperades i als boscos plujosos de coníferes. Hi ha poques estimacions disponibles de PPB, però per poder comparar i situar-se poden ser útils les dades obtingudes per a diferents comunitats tropicals i temperades. Per a una mateixa taxa d’acumulació de biomassa (expressada com la relació B/PPN aèria), als ecosistemes de les zones tropicals la taxa de respiració dels productors primaris, expressada en percentatge de producció primària bruta, és un 15% superior que la de les zones temperades. A les laurisilves subtropicals japoneses, per exemple, la respiració autotròfica total estimada només arriba a representar un 64,3% de la producció primària bruta, un valor molt inferior al 94% estimat per als boscos plujosos de coníferes de la regió nord occidental de l’Amèrica del Nord i al 71% d’algunes selves tropicals. El fet que la respiració total de les laurisilves sigui menor que la dels boscos plujosos de coníferes es deu, fonamentalment, al fet que l’acumulació de biomassa també hi és menor, i això es tradueix en un cost total de manteniment també inferior, encara que les taxes de respiració siguin més elevades, tal com ja s’ha dit.
La taxa de respiració de les parts llenyoses acostuma a ser proporcional a la seva superfície, i la relació entre la superfície de les parts llenyoses aèries i l’índex foliar pot ser una manera de mesurar l’eficiència productiva d’un arbre aïllat o de tot un rodal. Aquesta relació sol ser més alta en el cas dels arbres petits i els boscos joves perquè no posseeixen tants elements llenyosos. La relació observada en els arbres de la laurisilva de Minamata oscil·la entre 2 i 10 per als arbres petits i decreix per als arbres més grossos; els que constitueixen el dosser del bosc acostumen a deixar de créixer quan la relació ateny un valor determinat, un valor llindar proper a 1,5. Es calcula que a la laurisilva de Minamata la relació és d’entre 2 i 3, pràcticament la mateixa mesurada en boscos caducifolis nord-americans.
L’equilibri entre fotosíntesi i respiració determina l’acumulació de matèria orgànica a l’arbre, cosa que depèn de la temperatura, de l’edat de l’arbre i de les seves dimensions. Els arbres de les laurisilves no arriben a tenir les dimensions ni la longevitat pròpies de les coníferes del boscos plujosos temperats en què aquestes dominen. En algunes laurisilves del Japó, les edats dels arbres més vells varien entre els 206 i els 360 anys, però la biomassa és molt inferior a la dels boscos plujosos de coníferes; només alguns boscos de la laurisilva valdiviana, de Nova Zelanda o de Tasmània, que tenen grans faigs australs emergents de gairebé 500 anys i algunes espècies de coníferes, podrien assolir valors elevats de biomassa i per tant de respiració, però no és fàcil que arribin a igualar els valors dels boscos plujosos de coníferes. A l’Himàlaia s’observa un gradient altitudinal en funció del qual la biomassa de les laurisilves d’altituds inferiors és menor que la de les selves d’altituds mitjanes, i el màxim de biomassa s’assoleix entre els 2 400 i els 2 600 m. Per contra, la producció té una tendència totalment diferent i es manté aproximadament constant a diferents altituds.
Pel que fa a la producció primària neta, els valors estimats per a les laurisilves varien entre 10 i 22 t/ha i any, encara que altres autors donen valors de 15 a 25 t/ha i any. A les selves subtropicals, la producció primària neta acostuma a ser una mica superior que a les temperades, i disminueix força quan les temperatures baixen en funció de la latitud o l’altitud, de la mateixa manera que la diversitat d’espècies i la complexitat estructural es redueixen. Així, a les selves perennifòlies magallàniques de “guindo” o “coigüe” magallànic (Nothofagus betuloides), que són les més extremes pel que fa a latitud i a les condicions climàtiques, la PPN varia entre 5 i 9 t/ha i any, encara que en aquest cas només s’inclouen en l’estimació els arbres d’aquesta espècie. Als boscos de l’Himàlaia, en canvi, la PPN i la biomassa se situen a l’extrem superior del rang indicat. De tota manera, falten estimacions de la selva valdiviana, on, igual que a l’Himàlaia, els arbres atenyen grans dimensions. A l’Himàlaia, fins als 2 600 m i malgrat un gradient tèrmic de 10˚C, la producció primària neta està compresa entre les 15 i les 20 t/ha i any. Aquest alt nivell de producció reflecteix probablement la coincidència de l’estació de pluges i de l’època de creixement. Al mateix temps s’observa que les selves situades a les altituds inferiors, on les temperatures són més elevades, tenen una biomassa menor que les selves de les zones muntanyoses mitjanes, on la taxa de respiració és més baixa.
Als boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord, la PPN, a diferència de la biomassa, no és gaire diferent de la d’altres boscos temperats. Als boscos joves, sobre sòls fèrtils i sota una influència marítima important, és on la producció és més alta; assoleixen una producció primària neta mitjana de 15 a 25 t/ha i any. Els valors més alts (36 t/ha i any de pes sec) corresponen a boscos joves de hemlock occidental (Tsuga heterophylla) i de sequoia costanera (Sequoia sempervirens). Es tracta de valors del mateix ordre que els màxims estimats per a les boscanes decídues i les laurisilves, que són respectivament de 24,1 i 28,0 t/ha i any, o que la producció de les plantacions de sugi o cedre roig japonès (Cryptomeria japonica) que és de 29,1 t/ha i any i és una de les plantacions forestals més productives. Als boscos madurs la producció primària neta és inferior, i els càlculs duts a terme per als diferents tipus de boscos plujosos de coníferes la situen entre 4,2 i 15,0 t/ha i any. Els valors més alts de producció sempre s’assoleixen a les zones costaneres i sobre sòls fèrtils, i els més baixos cap a l’interior i a altituds elevades. En general, els boscos plujosos de coníferes tenen una producció de 15 t/ha i any; es tracta d’una producció primària neta inferior a la de les selves plujoses tropicals (23 t/ha i any) i semblant a la de les boscanes mixtes (13 t/ha i any), però superior a la dels boscos boreals de coníferes (7 t/ha i any).
Les diferències degudes a la despesa total per respiració són molt més grans que les diferències en la producció primària neta. Ja que la producció primària neta és la velocitat d’emmagatzematge de la matèria orgànica o la quantitat d’energia fixada per unitat de temps un cop descomptada la respiració, les grans acumulacions de biomassa dels boscos plujosos de coníferes es deuen, tal com ja s’havia comentat, al creixement continuat durant centenars i fins i tot milers d’anys més que no pas a una producció primària neta molt elevada. El quocient PPN/PPB, que sol ser baix als ecosistemes forestals (perquè tenen una gran quantitat de biomassa acumulada a les estructures llenyoses), ho és particularment en aquests boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord, ja que el manteniment d’aquestes estructures els requereix grans quantitats d’energia.
El quocient producció/biomassa (PPN/B) representa la taxa de renovació de la biomassa, és a dir, la velocitat a la qual es renova. Als boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord, considerant les arrels, els troncs, les branques i les fulles d’arbres de diferents boscos estudiats, aquest quocient varia entre 0,014 i 0,087 anys–1. Es tracta d’una taxa molt baixa si la comparem amb el valor de 0,04 anys–1, que és la mitjana que s’obté per a la majoria dels boscos, incloent-hi les selves tropicals plujoses. Aquest quocient també es pot interpretar com a indicador de l’eficiència de la biomassa, o com la biomassa produïda per unitat de biomassa i, en aquest sentit, els boscos plujosos de coníferes són els que tenen una eficiència menor. L’invers d’aquest quocient (B/PPN) dóna el temps de renovació, i és obvi que valors baixos del quocient PPN/B corresponen a temps de renovació alts. Així doncs, es pot dir que la biomassa dels boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord és la que triga més temps a renovar-se.
La disponibilitat d’aigua
Les condicions climàtiques sota les quals es donen les selves temperades exclouen cap limitació dràstica de les disponibilitats d’aigua. Però els mitjans pels quals accedeixen a aquesta aigua els vegetals i, en particular, els arbres i arbustos dominants ja són una altra cosa. Així i tot, a la regió dels boscos plujosos de coníferes del NW de l’Amèrica del Nord es manifesta un període d’eixut relatiu, de durada i intensitat variables segons els anys i la zona. Sol començar amb l’estació de creixement i dura més cap a l’interior del continent, de manera que la fotosíntesi pot estar força limitada durant l’estiu, quan les condicions d’humitat són desfavorables. Aquesta deficiència hídrica durant l’estació de creixement provoca el tancament dels estomes per reduir les pèrdues d’aigua, però d’aquesta manera també s’impedeix l’absorció de diòxid de carboni i l’activitat fotosintètica es pot reduir fins a nivells molt baixos. El tancament dels estomes es produeix quan ja no queda aigua disponible al sòl i quan la demanda evaporativa és molt alta (dèficit de vapor d’aigua a l’atmosfera). Als llocs més eixuts, els efectes sobre l’activitat fotosintetitzadora són més acusats, fins al punt que les coníferes d’aquests indrets realitzen fins al 70% de la fotosíntesi fora de l’estació típica de creixement.
Les coníferes presenten adaptacions estructurals que els permeten superar amb èxit hipotètiques condicions d’estrès hídric i que les posen en avantatge enfront d’altres espècies. D’una banda, a les acícules l’intercanvi de calor està sotmès a menys impediments que a les fulles planes i més grans, i per això la temperatura de les acícules sempre és més pròxima a la temperatura ambient que la temperatura de les fulles de les angiospermes; com a conseqüència, en les coníferes, els nivells de respiració i transpiració són relativament inferiors. D’altra banda, l’efecte de la sequera estival es veu amortit en certa manera per la gran quantitat d’aigua que aquestes coníferes poden acumular a l’albeca. En algunes localitats, l’aigua de l’albeca es recarrega de seguida amb les tempestes d’estiu, i, en altres, després de l’estació de creixement. Una diferència crítica entre les coníferes i les angiospermes és que l’albeca de les primeres es recarrega fàcilment mentre que en les angiospermes aquesta recàrrega és més difícil, i en les d’anell porós és impossible. Això es deu al trencament de la columna d’aigua d’aquestes espècies i a la subsegüent cavitació als vasos (s’omplen d’aire). En les coníferes, les traqueides queden afectades d’una manera individual i són fàcilment recarregables, ja que els porus areolats que les comuniquen entre elles es tanquen en resposta al gradient de pressió que s’hagi generat.
La relació funcional que s’estableix durant l’estació de creixement entre l’àrea de l’albeca i l’àrea foliar es manté força estable, independentment de la mida de l’arbre, segons les condicions de creixement. Als ambients més estressants i exposats, l’àrea conductora que els arbres necessiten sol ser més gran que la dels arbres de les localitats protegides o costaneres. Per exemple, als boscos subalpins d’avet de Mertens (Tsuga mertensiana), la relació entre l’àrea foliar en m2 i l’àrea conductora en cm2 és de 0,16 m2/cm2, mentre que als boscos de hemlock occidental (T. heterophylla), on la influència marítima és més gran, aquesta relació és de 0,45 m2/cm2. En els boscos de la varietat menziesii de l’avet de Douglas (Pseudotsuga menziesii var. menziesii), la més típica, s’obtenen dades similars: 0,54 m2/cm2, en comparació als 0,25 m2/cm2 de la varietat glauca (P. menziesii var. glauca). Els boscos de la primera varietat creixen prop de la costa, mentre que els de la segona creixen més cap a l’interior. Als boscos de pi groc o ponderosa (Pinus ponderosa), que creixen en condicions més xeròfiles, la relació és de 0,25 m2/cm2.
La mida i el cicle de les fulles presenta diferències entre les laurisilves temperades, on l’estacionalitat és menys marcada, i les laurisilves subtropicals, en les quals el grau d’estacionalitat és major (especialment allà on el règim de plujes és monsònic). A les laurisilves tropicals sovint abunden les espècies caducifòlies, o si més no de fulles senescents, encara que això pot ser degut a la insuficiència de nutrients més que no pas a la manca d’aigua. La mida de les fulles és una altra característica molt important perquè determina la velocitat a què tenen lloc molts processos; tant als ambients molt freds com als ambients molt calorosos i amb manca d’aigua la regulació tèrmica de les fulles grans és difícil, i per tant les fulles tendeixen a ser petites. Si la mida foliar és adequada s’afavoreix la regulació de la temperatura, i les condicions són quasi òptimes per a la fotosíntesi. Així, en ambients calorosos on l’aigua és abundant, la mida de les fulles és més gran. A les laurisilves subtropicals, on les temperatures són elevades i l’aigua abundant, hi ha més espècies arbòries de fulles grans que a les laurisilves temperades. Les fulles dels “guindos” (Nothofagus betuloides) de la Terra del Foc, per exemple, només tenen uns 2 cm2 de superfície; en aquest cas és el fred la principal causa limitadora de la mida, perquè l’aigua és abundant durant tot l’any.
De vegades, com passa a les laurisilves de les illes Canàries, les boires persistents fan minvar la demanda evaporativa de l’aire a l’estiu, quan, en general, les precipitacions són escasses i es produeixen períodes de sequera. Si no fos per les boires, la laurisilva canària probablement desapareixeria, ja que, quan les precipitacions estivals són molt escasses, les plantes serien incapaces de resistir, fins i tot en condicions moderadament àrides. La regulació estomàtica d’aquestes espècies és poc eficient i, per tant, sense boires perdrien grans quantitats d’aigua per transpiració; la seva tolerància a la sequera és molt baixa i el seu sistema de transport d’aigua i nutrients també és bastant ineficient, de manera que, fins i tot en condicions d’aigua abundant i disponible al sòl, el seu sistema conductor no podria proveir suficientment les fulles d’aigua i nutrients.
La disponibilitat de nutrients
A les selves temperades, la disponibilitat de nutrients al sòl és en general força baixa i el reciclatge de la matèria orgànica és lent. A escala global, des de l’equador fins a la zona temperada, les espècies perennifòlies dominen en els sòls pobres sempre que la radiació o la temperatura no siguin factors limitadors. L’avantatge principal del caràcter perennifoli és la capacitat de les espècies d’allargar l’estació de creixement en aquells indrets on la disponibilitat de nutrients és baixa; si algun altre factor del medi fa que el període de creixement sigui més curt, però, aleshores les espècies perennifòlies es veuen desplaçades per les caducifòlies, de creixement més ràpid i amb un cicle de nutrients més accelerat. A escala local, l’aparició de boscos mixtos d’espècies caducifòlies i perennifòlies no es pot atribuir al clima i prou, sinó que les condicions edàfiques també hi tenen un paper molt important. Així, s’ha observat que les espècies caducifòlies eviten els sòls pobres en nutrients, els quals són ocupats per les espècies perennifòlies, que hi tindrien un avantatge competitiu pel fet que el seu cicle intern de nutrients és més conservador. A la regió de la costa NW de l’Amèrica del Nord, la major part de la descomposició de la matèria orgànica, i per tant l’alliberament de nutrients, té lloc durant l’hivern, i s’ha observat que fins al 90% de la pèrdua de pes es produeix en aquesta època de l’any. Segons els resultats obtinguts en diferents tipus de boscos i en altituds diferents, durant l’estiu, als mesos de juliol i agost la descomposició és gairebé nul·la.
Al fet que al sòl la concentració de nutrients sigui baixa hi contribueixen diferents factors: la gran biomassa dels arbres, que reté i immobilitza una quantitat elevada de nutrients en el si de les seves estructures; una relació carboni/nitrogen (C/N) força alta (aproximadament 40 o més) en la matèria orgànica, com és característic de les coníferes, fet que significa que la concentració de N és baixa, i per això ho és també l’activitat dels descomponedors; i, per últim, els grans incendis catastròfics, poc freqüents però de gran intensitat, que destrueixen el bosc i propicien la consegüent pèrdua de nutrients (per exemple, als boscos de la península d’Olímpic i en altres zones plujoses de la regió). A les zones més seques, els focs superficials eren freqüents abans de la política de supressió d’incendis, i resultaven beneficiosos perquè cremen el material detrític i acceleren el reciclatge dels nutrients.
El nitrogen és un dels nutrients més afectats pels incendis catastròfics; això no obstant, hi ha molts organismes capaços de fixar-lo i compensar així les pèrdues produïdes pel foc. Moltes espècies pioneres, entre elles el vern roig americà (Alnus rubra) i la ramnàcia Ceanothus velutinus, arriben a fixar entre 50 i 300 kg de nitrogen/ha i any, i això sense comptar el nitrogen produït pels líquens foliacis que viuen a les capçades de les coníferes, la majoria dels quals són endèmics de la regió; la fixació de nitrogen per part d’aquests líquens arriba a ser de fins a 5 kg/ha i any. Així mateix, els grans troncs que després del foc queden a mig cremar proporcionen una reserva addicional de nutrients i serveixen de substrat per a molts bacteris fixadors.
D’altra banda, les coníferes disposen de diversos mecanismes per fer front a l’escassetat de nutrients i, en general, tenen uns requeriments baixos. Així, en retenir les fulles durant molts anys (de 9 a 31 als boscos de la costa NW de l’Amèrica del Nord), redueixen les seves necessitats i, tot i que la biomassa d’aquestes fulles és elevada, la de les fulles noves només representa un 20%. La baixa concentració de nutrients a les fulles indica que aquests es requereixen en quantitats petites, fet que provoca una descomposició més lenta. En un bosc d’avet de Douglas (Pseudotsuga menziesii) de 450 anys, la concentració de nitrogen a les fulles és del 0,8% del pes sec, menys de la meitat que en les d’un planifoli, i el mateix passa amb altres nutrients, com el calci, del qual una conífera només en necessita la meitat que un planifoli.
A més, encara que no hi ha dades molt concloents, sembla que les coníferes són més eficients en l’absorció del nitrogen i el fòsfor; això sense comptar que absorbeixen i emmagatzemen aquests elements a l’hivern, estació de latència obligada per als caducifolis. Aquests darrers també estan en desavantatge durant part de l’estació de creixement, ja que en aquesta època l’índex de descomposició de la matèria orgànica del sòl és molt baixa. L’eficiència de les coníferes en l’ús dels nutrients, gran part dels quals són subministrats per retranslocació des de les fulles velles i senescents, també és força alta. El nitrogen que utilitzen els boscos madurs de coníferes del Pacífic prové gairebé en la seva totalitat del reciclatge intern, que interfereix en el procés de mineralització. En un bosc d’avet de Douglas (Pseudotsuga menziesii) de 100 anys d’edat, per exemple, la meitat del nitrogen és subministrat per retranslocació, i això fa que la demanda total d’aquest nutrient disminueixi en uns 50 kg/ha i any (als boscos més joves, en uns 30 kg/ha i any). Al llarg de la successió, doncs, la immobilització de nutrients a la biomasa augmenta, mentre que la qualitat de la matèria orgànica i la taxa de descomposició disminueixen. Aquest comportament no és exclusiu dels boscos d’avet de Douglas, sinó que per als boscos d’altres espècies s’han descrit dinàmiques similars. Les espècies decídues també retransloquen els nutrients abans que caiguin les fulles, però els seus requeriments són molt més alts. Als boscos caducifolis de les àrees orientals dels Estats Units, per exemple, el requeriment total anual de nitrogen és de 70 kg/ha, menys d’una tercera part del qual és subministrat per retranslocació.
Al mateix temps, grans quantitats de nutrients de les fulles són reabsorbits abans que aquestes caiguin al sòl. D’aquesta manera, les plantes interrompen el procés de descomposició del material vegetal; la biomassa foliar actua també com una reserva de nutrients i la reabsorció es constitueix en un mecanisme de renovació. La retranslocació anual és de 56,5 kg de nitrogen/ha, 13,5 kg de potassi/ha, 3,4 kg de magnesi/ha i 3,1 kg de fòsfor/ha. Si es comparen aquests valors amb les quantitats d’aquests nutrients que les mateixes plantes prenen del sòl, la retranslocació representa respectivament el 54% dels requeriments de nitrogen, el 26% dels de fòsfor, el 25% dels de magnesi i el 21% dels de potassi. D’altra banda, la quantitat de nutrients que queda atrapada amb l’increment anual de biomassa es pot calcular fent la diferència entre l’absorció de minerals i el retorn d’aquests al sòl, principalment en forma de fullaraca. Aproximadament entre el 6 i el 9% del nitrogen, el magnesi, el potassi i el calci, i el 25% del fòsfor s’incorporen anualment a la fracció de la producció primària bruta que contribueix a l’increment de biomassa, sobretot en forma de fusta, de la laurisilva. Aquestes dades il·lustren prou bé el fet que les plantes utilitzen grans quantitats de nutrients per a les fulles, que tenen una taxa de renovació alta, mentre que en fan servir concentracions petites per a la producció de fusta, la taxa de renovació de la qual és molt baixa.
La quantitat de nutrients minerals present en l’increment de biomassa comporta una reducció de la mateixa magnitud en les reserves de nutrients del sòl. Si aquest increment fos constant, el calci duraria uns 138 anys, el potassi uns 95, el magnesi uns 517, el fòsfor uns 12,5 i el nitrogen uns 1 031, mentre que el retorn de nutrients a través dels detritus vegetals i la lixiviació permetrien un increment constant de biomassa durant vuit anys. Cal destacar l’important paper de les espècies perennifòlies en el reciclatge de nutrients; normalment, en les seves fulles el contingut de nutrients és inferior al de les espècies caducifòlies. A la laurisilva de la conca experimental de Coweeta, per exemple, la biomassa foliar de les espècies perennifòlies representa el 35% de la total, i per tant suposaria el 35% del carboni. En canvi, les fulles contenen només el 21% de fòsfor, el 20% de magnesi, el 17% de nitrogen i un 14% de potassi.
La matèria orgànica viva a disposició dels herbívors
Als boscos en general, i també a les selves temperades, la proporció de la producció primària neta (PPN) que és consumida pels herbívors (tot i que cal incloure entre aquests, a més dels consumidors de fulles, fruits i altres parts dels vegetals, els paràsits i els animals que s’alimenten d’exsudats dels productors primaris) és molt petita. Les estimacions que s’han fet d’aquesta proporció per a diversos boscos donen valors d’aproximadament el 10%, tot i que aquest percentatge és més gran als boscos tropicals. Aquestes dades contrasten amb les d’ecosistemes com les praderies, on el consum per part dels herbívors és molt elevat i en molts casos supera el 50%.
Als boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord, el consum és molt baix a causa de la baixa qualitat de l’aliment i pel fet que aquest, a més, és poc mengívol; també s’ha de tenir en compte que les fulles, que són l’aliment habitual dels herbívors, es troben a força alçada i són físicament inabastables per a la majoria. Els insectes són els que en consumeixen més i, llevat dels períodes de plagues, el consum només representa entre l’1,5 i el 2,5% de la producció primària neta. Tanmateix, les poblacions d’aquests insectes augmenten periòdicament de densitat fins a esdevenir flagells de proporcions epidèmiques. Per exemple, les erugues del tortrícid Choristoneura occidentalis, que consumeixen fulles de pícees (Picea), i les del limàntrid Orgyia pseudotsugata, que s’alimenten de fulles d’avet de Douglas (Pseudotsuga menziesii), entre moltes altres espècies, poden defoliar grans extensions de bosc en un sol estiu.
En molts boscos, aquestes transferències massives de matèria i energia en la xarxa alimentària que té per base els insectes herbívors signifiquen, en primer lloc, una gran reducció de la producció primària neta durant uns quants anys i, en segon lloc, una disminució de la biomassa acumulada. D’altra banda, la gran abundància d’insectes implica que hi ha una font inusual d’aliment disponible per a moltes espècies d’ocells i altres animals, que l’any següent o els anys successius incrementaran les seves poblacions; al seu torn, els depredadors d’aquestes espècies tindran més aliment disponible i així fins a arribar als depredadors que ocupen les posicions més altes de la xarxa alimentària. De manera que aquestes pulsacions d’energia que van des dels productors primaris fins als consumidors es reflecteixen en la magnitud i l’expansió temporals dels fluxos d’energia i biomassa, que recorren com una ona la xarxa tròfica al llarg d’uns quants anys. Efectes similars s’observen en la xarxa tròfica basada en els detritívors quan reben grans quantitats de femta i organismes morts, principalment insectes.
La defoliació massiva mata molts arbres i altres pateixen una reducció dràstica del creixement, però, això no obstant, en determinades circumstàncies, els boscos d’algunes espècies com l’avet de Douglas es recuperen l’any següent, després que gran part de les seves fulles hagin estat menjades per erugues d’Orgyia pseudotsugata però el creixement, en canvi, no s’hagi vist gaire afectat. Això pot ser degut, per exemple, al fet que el defoliador actuï durant anys de sequera, circumstància en què la reducció de l’àrea foliar pot fins i tot resultar beneficiosa, ja que disminueix la superfície de transpiració; les fulles que queden poden augmentar la seva eficiència fotosintètica i compensar les pèrdues produïdes per la disminució del nombre de fulles. Als boscos destinats a la producció de fusta, generalment es tracta d’evitar l’herbivorisme mitjançant insecticides o altres mecanismes, però a llarg termini, sembla que un cert nivell estable d’insectes defoliadors és beneficiós perquè estimula la descomposició de la matèria orgànica i la circulació dels nutrients, i augmenta la productivitat. També s’ha suggerit que als boscos envellits les defoliacions massives per part d’insectes són beneficioses perquè estimulen la substitució subsegüent d’aquests boscos per boscos joves, molt més productius i menys susceptibles d’ésser defoliats.
Similar al consum estimat per als boscos plujosos de coníferes de les costes del NW de l’Amèrica del Nord resulta la fracció (tan sols un 2,38%) que consumeixen els herbívors a la localitat WS18 de la conca experimental de Coweeta, la subconca més ben estudiada d’aquest espai. A banda d’aquestes dades, no es coneixen estimacions basades en mesures per a cap altra laurisilva, encara que, pel que fa a les selves en general, s’acostumen a donar valors de consum per part dels herbívors propers a un 10%. La resta de la producció primària neta d’aquesta laurisilva de Coweeta bé s’acumula parcialment en forma de biomassa (un 45,2%), bé passa al compartiment detrític (un 52,4%). Es tracta d’una selva temperada relativament jove, d’uns 70 anys, i per això l’acumulació de biomassa encara constitueix una fracció important de la producció primària neta.
La matèria orgànica morta a disposició dels detritívors i descomponedors
Una part molt important de la producció primària neta (PPN) de les selves temperades va a parar al sòl, on és processada per diferents animals, fongs i bacteris fins que finalment és descomposta i s’alliberen els elements que constitueixen la matèria orgànica. Aquesta matèria orgànica morta té un paper molt important en els ecosistemes per dues raons: d’una banda, és una font d’energia per a tots els organismes heterotròfics, i de l’altra constitueix una reserva de nutrients que s’alliberen gradualment a mesura que és consumida pels organismes detritívors. Els nutrients, entre altres el fòsfor i el nitrogen, són finalment reutilitzats pels productors primaris i així es tanca el cicle de la matèria en l’ecosistema. Als boscos madurs, la producció primària neta que passa a formar part de la necromassa pot arribar a ser del 80%. Aquesta transferència de matèria orgànica viva al compartiment detrític constitueix la ruta principal de la matèria i l’energia als boscos, i és també el fonament d’una xarxa tròfica de gran importància per a l’ecosistema —més important que la que es basa en els herbívors—.
Les taxes de mortalitat i els seus patrons estacionals varien no tan sols segons el tipus de bosc, sinó també segons l’espècie i la part de la planta. Hi ha, però, una tendència general a acceptar que la taxa de mortalitat anual dels elements de dimensions reduïdes, com ara fulles i branquillons, és més alta a les zones de clima més càlid. Els patrons de caiguda de la virosta fina són força regulars en l’espai, però els de virosta gruixuda són irregulars tant en el temps com en l’espai. Es calculà que la taxa mitjana anual de caiguda de virosta en un rodal de 60 anys d’edat de la laurisilva de Minamata era de 8,4 t/ha i any (incloent-hi la taxa de caiguda de virosta gruixuda de més de 1,5 cm de diàmetre, que és de 2,4 t/ha i any); en aquest rodal, concretament, la virosta gruixuda constitueix el 28% de la virosta total, però la proporció de virosta gruixuda en un bosc madur acostuma a ser més alta, del 30 al 40%, de manera que la taxa de caiguda de virosta total d’una laurisilva pot ser de 8 o 10 t/ha i any, aproximadament la meitat de la taxa de les selves plujoses tropicals de les terres baixes de la Malàisia occidental.
La relació entre la taxa de caiguda de la fulla i la biomassa foliar expressa el tempsde residència, és a dir, el temps que tarda a renovar-se tot el fullam. A la laurisilva de Minamata, per exemple, aquest temps és d’1,9 anys, encara que les espècies dominants de l’estrat arbori, com ara el castanyer shii o “shii” (Castanopsis cuspidata) i algunes espècies d’alzines del subgènere Cyclobalanopsis, acostumen a tenir un temps de residència més curt que les espècies del sotabosc. El temps de renovació del “shii”, l’espècie dominant de l’estrat arbori, és tan sols d’1,6 anys, mentre que el del “sakaki” o arbre sagrat del Shintō (Cleyera japonica), una espècie de sotabosc, és de 6 a 7 anys. Els branquillons també es renoven ràpidament, igual que les fulles. Es calcula que en aquest bosc el temps mitjà de renovació dels branquillons de 2 mm de diàmetre, calculat mitjançant el mateix procediment que el de la fulla, és de dos anys. Les parts llenyoses més gruixudes presenten un temps de renovació superior, que correspon al seu diàmetre elevat a una potència de 0,9.
La quantitat de fullaraca produïda depèn de la biomassa de fulles que es renova cada any. Als boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord, per exemple, llevat dels anys de defoliacions massives per part dels herbívors, cada any es renova de mitjana fins al 20% de la biomassa foliar. La producció de fullaraca és menor com més gran és el nombre d’anys durant els quals les fulles han estat retingudes a les branques. Per exemple, als boscos d’avet roig americà (Abies amabilis) de la Colúmbia Britànica (Canadà), a la zona costanera, les fulles tenen un temps de permanència a les branques de fins a 9 anys, fins a 19 a la muntanya mitjana i fins a 26 als boscos de les zones de major latitud, a uns 1 700 m. En tots aquests boscos, la biomassa de fulles és similar, però la caiguda de la fullaraca varia de manera considerable: és menor als boscos on les fulles són retingudes més anys.
Hi ha una gran variabilitat en l’aportació de material detrític en les diferents selves temperades. En els boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord les aportacions de material detrític fi són de les més baixes (tan sols són inferiors a la taigà), mentre que a les laurisilves són de les més elevades (superades només per les selves plujoses tropicals). La virosta caiguda a les laurisilves de l’Himàlaia central, d’altra banda, és semblant a la d’altres laurisilves, però a la part oriental de la serralada els valors estimats s’assemblen més als de les selves tropicals. La virosta caiguda als boscos de pi de tres fulles de l’Himàlaia (Pinus roxburghii) oscil·la entre 4,4 i 7,6 t/ha i any però, tot i que es tracta de valors relativament alts, encara ho són més als boscos de pi “loblolly” (P. taeda) de Carolina del Sud, al SE dels Estats Units. La fracció de virosta caiguda que correspon a les fulles és del 54 i el 82% respectivament.
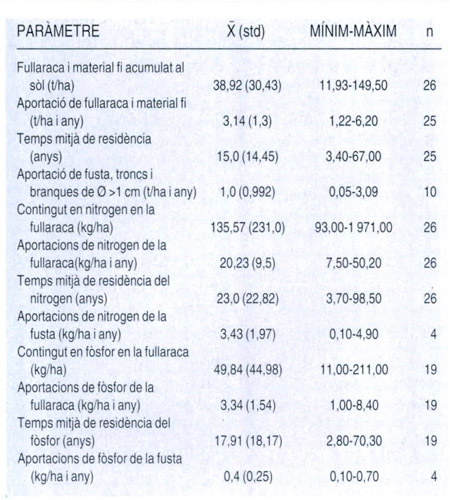
Característiques del material detrític vegetal de diferents boscos de coníferes del NE de l’Amèrica del Nord situats entre els 44° i els 47° de latitud N (n indica el nombre de boscos estudiats). Les dades s’expressen com el valor de la mitjana aritmètica i de la mitjana estadística (més representatiu que el primer perquè és més proper al valor que apareix amb més freqüència a la mostra). La quantitat de matèria vegetal morta que s’acumula al sòl de les selves temperades per unitat de temps és molt gran i varia en funció del tipus de bosc, l’estadi de desenvolupament en què es troba i el clima. Així, per exemple, el volum de fullaraca és superior als boscos dominats per espècies de fulla de vida curta que als boscos on les espècies dominants tenen fulles de vida més llarga. D’altra banda, a les selves temperades de les regions subtropicals l’acumulació de fullaraca sol ser menor que als boscos de les zones temperades, bé que amb altres tipus de materials detrítics pot succeir el contrari.
A partir de dades proporcionades per l’autor
A la laurisilva de la conca experimental de Coweeta la producció de material detrític és de 4,4 t/ha i any; a aquesta quantitat, les fulles hi contribueixen amb un 64%, fet que equival a dir que la producció de fullaraca és de 2,81 t/ha i any. Són valors bastant baixos en comparació amb la laurisilva canària o la japonesa o amb els valors generals estimats per a les laurisilves subtropicals o de la zona temperada. En aquesta laurisilva de la conca experimental de Coweeta també es pot apreciar que la diferència entre la biomassa de fulles (5,6 t/ha i any) i la producció de fulles (4,2 t/ha i any), i la diferència entre la producció de fulles i la de fullaraca (2,8 t/ha i any), és de 1,4 t, en tots dos casos, la qual cosa reflecteix la importància de les espècies perennifòlies.
Pel que fa a l’acumulació de matèria orgànica morta als sòls en forma de virosta fina i humus, s’ha constatat que segueix un gradient oposat i que sol ser més gran a les zones de clima més càlid, encara que, en fer disminuir l’activitat dels descomponedors, les baixes temperatures i els baixos continguts en nutrients de les coníferes de les regions més fredes també fan augmentar l’acumulació. Per això, als boscos plujosos de coníferes de la costa NW de l’Amèrica del Nord els nivells d’acumulació de virosta i humus a terra són dels més alts (38,9 t/ha), només superats pels de la taigà (44,7 t/ha); a les selves tropicals plujoses la mitjana publicada indica una quantitat de 22,5 t/ha, semblant a la de les laurisilves subtropicals i la dels boscos temperats càlids de coníferes (20 t/ha); els boscos temperats caducifolis presenten una acumulació de 32,2 t/ha i els boscos mediterranis d’11,4 t/ha. A la laurisilva de la conca experimental de Coweeta l’acumulació arriba a 8,5 t/ha, i com que l’entrada d’aquest material a la conca és de 4,4 t/ha i any, la durada de la capa de virosta sobre la superfície del sòl és en aquest cas d’uns dos anys, cosa que equival a una taxa de descomposició del 52% anual o del 0,1452% diari. Pel que fa a l’acumulació de virosta gruixuda, aquesta acostuma a dependre més de la maduresa del bosc que no de factors climàtics, i així, en boscos madurs, el volum de virosta gruixuda equival a un 10% de la biomassa aèria, independentment del tipus de bosc.
La quantitat total de necromassa forestal és encara més gran quan es considera la fracció que n’hi ha en forma d’humus. En general, es considera que l’acumulació d’humus en una laurisilva és aproximadament equivalent a un terç de la matèria orgànica total de l’ecosistema, proporció que correspon a una sisena part en un bosc plujós tropical de terra baixa i a la meitat en un bosc de coníferes subalpí. A la mateixa localitat WS18 de la conca experimental de Coweeta s’ha calculat que hi ha unes 145,6 t/ha de matèria orgànica en forma d’humus, enfront de les 191,3 t/ha de fitomassa viva (aèria i subterrània). El total de matèria orgànica (viva més necromassa) de tot l’ecosistema és de 345,4 t/ha, quantitat que equival aproximadament a 144 t de C/ha. Als boscos de “guindo” (Nothofagus betuloides) de la Terra del Foc, la quantitat total de necromassa al sòl, en les seves diferents formes, iguala gairebé la fitomassa aèria.
La descomposició i el reciclatge
La taxa de descomposició, i per tant la taxa d’alliberament de nutrients, depèn de les característiques de la mateixa matèria orgànica que es descomposa, dels factors ambientals (com temperatura i humitat), i dels organismes del sòl que puguin viure en un indret determinat. La pèrdua de pes sec al llarg del temps no és la mateixa si es tracta de virosta fina que si es tracta de virosta gruixuda: la taxa de pèrdua d’una unitat de pes sec de virosta fina és pràcticament constant al llarg del procés de descomposició, i això significa que el pes sec disminueix exponencialment. Aquesta constant s’anomena coeficient de pèrdua de pes sec. En el cas de la virosta gruixuda cal més temps per a preparar el substrat llenyós, especialment a l’estadi inicial; així doncs, la taxa de pèrdua és baixa a l’estadi inicial i s’accelera gradualment en els estadis posteriors, de manera que al llarg del temps forma una corba sigmoïdal inversa i es pot ajustar a una equació logística.
La temperatura accelera la taxa de descomposició, però els efectes dels canvis estacionals i diaris en les condicions ambientals són variables; els estacionals indueixen modificacions en la fauna i en la població de descomponedors, i, en les regions que enregistren pluges estivals, l’efecte es multiplica amb els canvis estacionals de precipitació. El contingut d’aigua també accelera la taxa de descomposició, però en cas d’excés d’humitat la descomposició de la virosta gruixuda i de la matèria orgànica del sòl s’aturen per causa d’una ventilació deficient. En una àrea humida, la taxa mitjana anual de descomposició de virosta nova al sòl del bosc es pot estimar aproximadament a partir de la temperatura mitjana anual en ˚C, tot i que la taxa diària pot fluctuar àmpliament en funció de canvis puntuals de temperatura, contingut d’aigua i altres factors.
Les taxes de descomposició i els temps mitjans de residència se solen calcular a partir de les aportacions degudes a la producció in situ i del material detrític acumulat. En la majoria dels casos, les entrades des de l’exterior de l’ecosistema i les exportacions no intervenen en els càlculs de les taxes de descomposició. El temps mitjà de residència està definit com el quocient entre la quantitat de material acumulat dividit per les aportacions, i l’invers d’aquest quocient es pren com una taxa de descomposició (k), o velocitat a la qual el material detrític desapareix; aquesta és una taxa finita i és diferent de la taxa calculada ajustant les dades de pèrdua de pes del material detrític (o nutrients), que normalment segueix una pauta exponencial; les taxes obtingudes d’aquesta manera són taxes instantànies i només són similars a les taxes finites quan són molt petites.
Els temps mitjans de residència del material detrític són llargs en comparació dels d’altres biomes de latituds més baixes. Així, la descomposició és més ràpida a les laurisilves subtropicals: per exemple, en una laurisilva xinesa (26° 10" N i 117° 20" E) on abunda el castanyer fals Castanopsis kawakamii s’ha estimat una velocitat de descomposició de 0,364%/dia (comparable a les calculades en boscos tropicals de l’Índia). La descomposició és més lenta a les laurisilves amb pi de tres fulles de l’Himàlaia (Pinus roxburghii), on la taxa de descomposició estimada és del 0,19%/dia. Tanmateix, la descomposició del material de P. roxburghii és més ràpida, 0,165%/dia, que la de les pinedes amb pi “loblolly” (P. taeda) de Carolina del Sud, al SE dels Estats Units, la taxa de descomposició estimada dels quals és de 0,120%/dia. Aquesta taxa és encara més baixa per als boscos de faigs australs de les selves temperades i temperades fredes de l’extrem austral del continent americà. Als boscos de “coihue” comú (Nothofagus dombeyi) del vessant oriental dels Andes, que domina la Patagònia, molt més fred que l’occidental, la taxa de descomposició és de 0,056%/ dia, proporció molt semblant a la dels boscos de “guindo” (N. betuloides) de la Terra del Foc. En aquests boscos, les baixes temperatures i la petita oscil·lació tèrmica anual no afavoreixen l’activitat dels organismes descomponedors, i es calcula que els troncs dels grans arbres caiguts poden romandre fins a 500 anys sense descompondre’s.
La major part de les dades sobre el material detrític es refereixen a les aportacions aèries; això no obstant, quan s’ha considerat l’aportació deguda a les arrels s’ha vist que en determinades situacions era superior a la de la fullaraca. Això és el que passa als boscos subalpins o desenvolupats sobre sòls pobres. Segons s’ha calculat, en dos boscos determinats d’avet de Douglas (Pseudotsuga menziesii), un desenvolupat sobre sòls pobres i força secs i l’altre sobre sòls fèrtils, la producció primària neta aèria és de 7,3 i 13,7 t/ha i any, respectivament, però la producció de radícules (
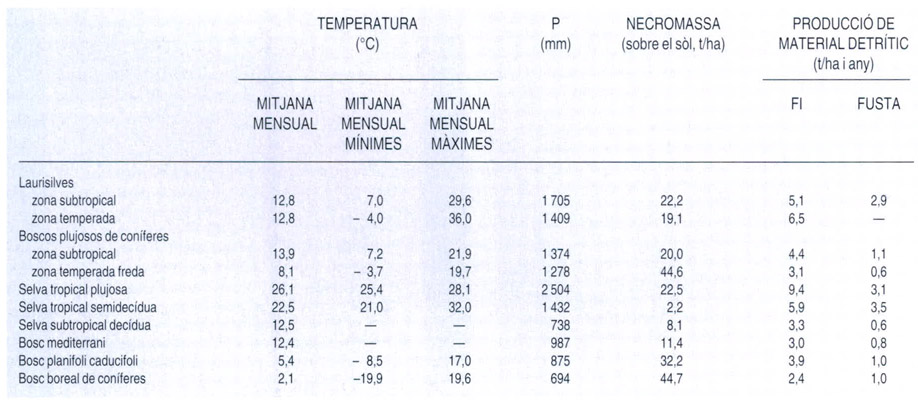
El volum de necromassa i la producció anual de material detrític varia als diferents tipus de selves temperades en funció de la temperatura i de les precipitacions, tal com succeeix també en altres biomes. Per exemple, els boscos plujosos de coníferes de la zona temperada freda presenten taxes d’acumulació de material detrític molt baixes, però les temperatures també són baixes i això retarda l’acció dels organismes descomponedors, de manera que el volum de necromassa que s’acumula al sòl és molt elevat. La temperatura i la pluviositat, però, no són els únics factors que determinen l’acumulació de material detrític i la quantitat de necromassa que hi ha al sòl d’un bosc, de manera que sovint es fa difícil observar una relació directa entre aquests factors.
A partir de dades proporcionades per l’autor
La xarxa tròfica assentada sobre el material detrític és la ruta més important del flux d’energia. Fongs i bacteris hi porten a terme gairebé tota la transformació de la necromassa. A les laurisilves, la mineralització l’efectuen principalment els fongs, després que l’activitat fragmentadora dels animals del sòl (principalment àcars i isòpodes en el cas de les laurisilves) els hagin preparat un substrat adient. En estadis posteriors, els cucs de terra també tenen un paper important en la humificació. Als boscos de coníferes, com que el material detrític és més àcid, els fongs són més abundants que els bacteris, i la descomposició també és més lenta. La biomassa dels descomponedors és petita amb relació a la quantitat d’energia que passa a través d’aquest nivell tròfic, ja que la seva mida també és reduïda i la seva vida curta. Com les arrels, en general han estat poc estudiats i no es diposa de gaires dades concretes publicades que facin referència a selves temperades. Els cucs de terra són poc abundants als boscos de coníferes, i en molts d’aquests boscos ni tan sols hi són presents a causa de la seva intolerància al tipus de material detrític que s’hi troba. Les termites són molt abundants a les selves subtropicals obertes i la seva activitat, perforant la fusta, fa que els troncs que han caigut desapareguin amb relativa rapidesa.
Els fongs i els bacteris constitueixen la principal font d’aliment per a moltes espècies de protozous i altres organismes de petites dimensions, i molts petits mamífers també consumeixen fongs de manera directa. A les selves patagòniques de faigs australs (Nothofagus) els fongs són molt abundants i canalitzen una gran part del flux d’energia. A la selva valdiviana, moltes de les espècies de petits mamífers més abundants, tant en nombre com en biomassa, també consumeixen fongs en quantitats considerables; algunes d’aquestes espècies són omnívores, i altres també inclouen en la seva dieta les parts verdes dels vegetals. Aquest fet posa de manifest que la xarxa tròfica basada en els descomponedors i la dels herbívors presenten interrelacions importants. En una selva temperada de Nova Zelanda, per exemple, la biomassa dels animals arriba a 504 kg de pes fresc/ha (sense comptar els invertebrats que viuen a les parts aèries de la vegetació), i a aquesta quantitat els cucs de terra hi contribueixen amb 333 kg, els artròpodes de la virosta amb 145 kg, els mamífers amb 25 kg i els ocells amb 0,6 kg. En selves de faigs australs, també de Nova Zelanda, la biomassa dels ocells és de 0,12 kg/ha. Com a dada comparativa, la biomassa dels cucs en un prat productiu pot oscil·lar entre 1 163 i 3 048 kg/ha.
Els canvis recurrents en el temps
Les variacions climàtiques estacionals, bé que moderades, són prou acusades al domini de les selves temperades perquè el cicle anual segueixi un model identificable. Paral·lelament a aquest cicle anual, a un ritme molt més lent, cada selva temperada descriu també un cicle vital marcat pel creixement i la mort dels arbres, la substitució d’unes espècies per unes altres en el paper de dominants i, també, per l’establiment repetitiu de determinades combinacions d’espècies al cobricel i al sotabosc en cada estadi de maduració de la selva en el cicle vital de la successió.
El cicle estacional
Les selves temperades de latituds mitjanes tindrien, aparentment, el mateix aspecte durant tot l’any, si no fos per la presència, a la primavera, d’algunes espècies de flors molt vistoses i pels caducifolis hivernals. Atès el clima benigne, els vegetals poden realitzar certa activitat fotosintetitzadora durant tot l’any, excepte a les àrees muntanyenques més enlairades. Tanmateix, l’activitat del càmbium i, per tant, el creixement en gruix, solen presentar un ritme anual molt marcat i, en moltes de les espècies arbòries, es formen anells anuals de creixement força ben diferenciats.
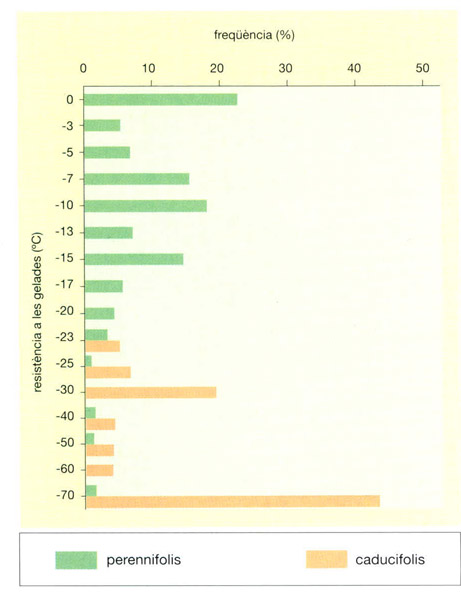
Resistència a les gelades dels branquillons dels perennifolis comparada amb la resistència dels branquillons d’espècies caducifòlies, segons un estudi dut a terme als boscos del Japó en el qual s’avaluaren 73 espècies de cada tipus. S’observa clarament com els branquillons de les espècies caducifòlies resisteixen temperatures molt més baixes que els branquillons de les perennifòlies, cosa que està completament d’acord amb la transició cap al caràcter caducifoli que s’observa en les formacions vegetals a mesura que es va cap a regions cada cop més fredes.
Idem, a partir d’A. Sakai & W. Larcher, 1987
L’hivern comporta baixes temperatures i una certa interrupció del creixement vegetal. Sota la capa de neu, els pinyons de “pehuén” (Araucaria araucana) i els d’altres coníferes romanen a l’espera de germinar a la primavera. Molts animals hivernen durant mesos, i els invertebrats sobreviuen enfonsant-se profundament sota terra, amagant-se a la fusta podrida o bé passant l’hivern en forma de pupa. Una part dels ocells migra a àrees més benignes, i els ocells i mamífers que es mantenen actius arrepleguen tot l’aliment que troben, inclosos els líquens i les molses que les tempestes hivernals han arrencat dels arbres.
A la primavera, així que la temperatura augmenta, la vida agafa embranzida. En el moment en què la neu es fon a les regions més fredes, un bé de Déu de tapissos de flors sembla aparèixer en un no res, sobretot a les clarianes obertes per la caiguda de grans arbres. Les coníferes treuen brots primaverals d’acícules tendres, mentre que els arbres planifolis floreixen i treuen fulla nova. Les diferents espècies de faigs australs, típics de les selves temperades de l’hemisferi sud, mostren els nous brots, en els quals es formen també, bé que no són gaire conspícues, les flors femenines i masculines, i el mateix fan altres planifolis. Els ocells migradors tornen i es disposen a aparellar-se i nidificar.
Tot i ser un bioma humit, a l’estiu hi acostuma a haver poca precipitació i, de vegades, autèntiques sequeres. Al S de Xile, a Nova Zelanda o a la costa NW de l’Amèrica del Nord, l’estació de creixement s’allarga durant tot l’estiu, encara que la sequera també afecta algunes àrees. No obstant això, l’estiu és un període de gran activitat, perquè els animals alimenten i crien els petits, i les plantes viuen un període de creixement ràpid i floreixen i fructifiquen. Coincidint amb el final de la brotada de les fulles de l’any, entre el final de la primavera i el començament de l’estiu, se sol produir la caiguda de la majoria de les fulles velles.
La tardor comporta una pèrdua important de fulles a tots els rodals, tret dels que són rodals purs de coníferes (on la caiguda d’acícules té lloc al llarg de tot l’any però la majoria es mantenen uns quants anys a l’arbre). Als boscos del S de Xile, amb faigs australs (Nothofagus) caducifolis, la tardor és especialment bonica, amb vessants plens d’arbres de fulla daurada esquitxats d’alguna conífera ocasional que sobresurt per damunt de l’estrat arbori principal. Els fruits dels arbres, un cop assolida la seva maduresa, cauen a terra i escampen les llavors que haurien de germinar la primavera següent. La producció abundant de fruits carnosos i secs, per altra banda, atrau molts éssers vius, tant si és amb l’objectiu d’acumular energia per a una llarga migració, com si es tracta d’emmagatzemar reserves de greix de cara a hibernar durant l’estació freda.
L’hivern és generalment el període més sec; és el moment en què les espècies caducifòlies perden totes o part de les seves fulles (entre el 75 i el 100%) i algunes floreixen cap al final de l’estació. En algun cas, però, com a Mèxic o als vessants meridionals de l’Himàlaia, els condicionaments climàtics són uns altres i l’hivern comporta condicions més humides —bé que la neu només és present a les muntanyes més altes—.
A les selves temperades situades a latituds més baixes, com les de la regió del Paranà, d’entre tots els factors climàtics, el que més limita el creixement i l’activitat dels vegetals són les baixes temperatures hivernals, bé que la majoria d’espècies no presenten períodes de latència llargs. L’activitat del càmbium de les espècies arbòries no té un ritme estacional gaire clar i les discontinuïtats que es manifesten en el creixement radial poden ser degudes a ritmes interns de fructificació i de repartició diferencial dels recursos, més que no pas a causes climàtiques. La formació d’anells anuals de creixement en el “kuri’y” (Araucaria angustifolia), tanmateix, és força regular, possiblement perquè les oscil·lacions de la temperatura són més acusades a les planes altes del S del Brasil que a les terres baixes on es desenvolupa la laurisilva d’aquesta regió.
Fructificacions de les principals espècies d’arbres de la selva valdiviana. Tant a les selves subtropicals com a les temperades, la floració i la fructificació es donen al llarg de tot l’any, bé que a causa d’una certa estacionalitat del clima, hi ha temporades en què aquestes activitats assoleixen el seu màxim. Per exemple, a la selva valdiviana la majoria d’arbres produeixen fruits a l’estiu i a la tardor. Durant un període de quatre anys (1980-83) s’observà que les espècies que produeixen més fruits (alternant anys de més i menys producció) són el “gevuín” (Gevuina avellana) i l’“olivillo” (Aextoxicon punctatum). El “roble pellín” (Nothofagus obliqua), un faig austral caducifoli, el “mañío” de fulles llargues (Podocarpus salignus), una conífera, i el ”tepa” o “laurela” (Laurelia philippiana), una monimiàcia laurifòlia, tenen màxims d’abundància de fructificació cada tres anys. Altres espècies, com el “canelo” xilè (Drimys winteri), l’“arrayán” (Lumaapiculata), l’“ulmo” (Eucryphia cordifolia) i l’“avellanillo” (Lomatia dentata) tenen una producció més baixa i no presenten cicles plurianuals definits.
IDEM, a partir de R. Murúa i L.A. González, 1985
Els arbres i arbustos normalment floreixen i fructifiquen una vegada l’any, al principi de l’estació humida, però algunes espècies ho fan dues vegades l’any i altres diverses vegades, segons un comportament arítmic independent de les condicions climàtiques de cada estació. La majoria d’espècies floreixen a la primavera o poc abans de l’estiu; algunes ho fan durant l’estiu o a l’hivern, però molt poques a la tardor. La maduració dels fruits és màxima a l’estiu, bé que al llarg de tot l’any hi ha espècies amb fruit. Sembla que hi ha certa relació entre l’època de maduració dels fruits i el mecanisme de dispersió de les llavors, de manera que quasi tots els que maduren a l’hivern produeixen llavors anemocores, bé que també els animals són agents molt importants en la disseminació de les llavors. Seria el cas del cridaner gaig blau americà o “gralha azul” (Cyanocorax caeruleus), disseminador principal de les llavors de “kuri’y” (Araucaria angustifolia); aquest còrvid té el costum d’emmagatzemar pinyons sota els troncs i les pedres i fins i tot en deixa caure en ple vol; d’aquesta manera, l’araucària té l’oportunitat de colonitzar nous territoris.
Els processos de successió
Les selves primàries madures són ecosistemes amb una funció ecològica singular. Protegides naturalment de les fluctuacions climàtiques extremes, tant de la calor i el fred com de les sequeres, la neu i el vent, la vida que s’hi desenvolupa acostuma a ser més estable, amb espècies animals i vegetals que en són exclusives i dependents, que no les poblacions, molt variables, dels estadis inicials de la successió. Assolir aquest estadi no és cosa de poc temps; s’estima que en els boscos plujosos de la costa NW de l’Amèrica del Nord, possiblement els més ben estudiats del bioma i alguns dels més ben estudiats del món, el cicle vital complet d’una selva madura s’allarga no menys de 400 anys: entre els 400 i els 500 anys s’assoliria l’acumulació màxima de biomassa i es mantindria al mateix nivell, reciclant la majoria dels nutrients, fins als 900.
En teoria, el procés comença quan alguna mena de pertorbació forta (un incendi espontani, per exemple) elimina l’antic recobriment forestal. Al llarg dels vint anys següents hi creix poca cosa tret d’herbes i matolls, tot i que també s’hi poden establir alguns plançons pioners de coníferes. A partir d’aquí, té lloc una colonització, que es tradueix en una progressiva dominància, per part de coníferes que no toleren l’ombra, com ara l’avet de Douglas (Pseudotsuga menziesii), que ve seguida d’un període de creixement ràpid que s’allarga fins que aquests arbres arriben a la maduresa, uns vuitanta anys més tard. Entre els 80 i els 200 anys hi ha una fase durant la qual les coníferes atenyen la seva taxa màxima de creixement anual abans de començar a desenvolupar les característiques del bosc madur. Entre els 200 i els 750 anys, en absència de cap pertorbació forta, la successió continua, i algunes espècies tolerants a l’ombra com el hemlock occidental (Tsuga heterophylla) i la tuia gegant (Thuja plicata) poden anar prenent el lloc als avets de Douglas a mesura que aquests moren i cauen.
En un interval donat, la biomassa total acumulada en les estructures dels productors primaris pot augmentar o disminuir. Els increments de biomassa són més grans durant els primers estadis de la successió, mentre que solen ser negatius després que una pertorbació, per exemple una tempesta de vent o de neu, tombi algun arbre i la seva biomassa es converteixi en necromassa. En un bosc d’avet de Douglas (Pseudotsuga menziesii) de la costa NW de l’Amèrica del Nord, la biomassa acumulada disminuí en 4,5 t/ha i any de mitjana, quantitat que tan sols representa el 0,52% de la biomassa total dels productors primaris, que és de 870,4 t/ha. Si es fa referència a la respiració, que és una mica superior al 97%, és probable que aquest bosc hagi assolit o estigui pròxim a assolir la seva biomassa màxima; en una situació com aquesta, les pèrdues de la biomassa acumulada podrien representar una fluctuació entorn de la mitjana d’aquest màxim, o bé un episodi més de la davallada que experimenten molts boscos envellits.
A banda dels primers anys de successió posteriors a una pertorbació forta, els boscos primaris es caracteritzen pel fet de presentar tot un ventall d’espècies i mides d’arbres. Aquesta varietat es manifesta de diferents maneres. Tant l’aparició de clarianes obertes per la mort natural dels arbres, com la successió constant d’espècies esciòfiles que té lloc sota els rodals inicials d’avet de Douglas (Pseudotsuga menziesii) (que provoca canvis a petita escala), com també les pertorbacions locals degudes a petits incendis i a caigudes d’arbres per causa del vent (que són factors igualment inductors de canvis), permeten que a l’estrat arbori d’un bosc madur o primari ja existent tingui lloc una nova successió reduïda. Així, un bosc primari presenta sovint plançons d’arbres i peus madurs de molts segles d’edat, que formen un mosaic irregular propi de cada bosc i cada moment concret. La presència de fusta morta caiguda i en peu, i de virosta llenyosa gruixuda, té una importància crucial per a moltes espècies, com ara els óssos, diferents ocells, insectes i plantes inferiors, sobretot líquens i fongs.
Altres regions del bioma es caracteritzen per un model successional en el qual són molt freqüents les pertorbacions de certa importància. Terratrèmols, erupcions volcàniques, esllavissaments, glaceres, allaus, vents violents, incendis i tales arreu indiscriminades influeixen en l’estructura, la dinàmica i la distribució de comunitats forestals com les de la selva valdiviana, on els grans faigs australs, les araucariàcies o les podocarpàcies, que sovint apareixen com a arbres emergents, no es regeneren sota el dens dosser de laurifolis dominat per vells arbres, sinó que només ho fan als espais denudats per grans catàstrofes o pertorbacions d’escala més modesta. La freqüència de les pertorbacions impedeix l’exclusió d’aquests arbres per les espècies esciòfiles, que acaben esdevenint dominants en algunes situacions particularment estables.