Els organismes del zooplàncton
Dins les cadenes tròfiques marines el zooplàncton constitueix l’anella que canalitza l’energia produïda pels organismes autòtrofs cap als consumidors secundaris. D’una manera molt simplificada, es pot afirmar que el zooplàncton s’encarrega de posar a disposició dels grans carnívors pelàgics l’energia generada pel fitoplàncton. Tanmateix, el paper del zooplàncton als ecosistemes marins és força més complex, fruit tant de la diversitat d’organismes i de formes que el componen com de la diversitat d’estratègies que segueixen les diferents espècies i grups. El zooplàncton és constituït per tots aquells organismes heteròtrofs —encara que recentment s’ha observat que alguns tenen també capacitat autòtrofa, gràcies a la simbiosi— que viuen en suspensió a les masses d’aigua i que depenen de la dinàmica d’aquestes, ja que els manca una capacitat de desplaçament suficient.
Entre les espècies del zooplàncton n’hi ha de carnívors que, per tant, són consumidors secundaris, però hom convé a reservar aquesta denominació per als depredadors marins més grossos (grans crustacis, peixos, etc.).
El meroplàncton i l’holoplàncton
Encara que la majoria dels organismes del zooplàncton són de petites dimensions, n’hi ha de mides que van des d’una vintena de micròmetres fins a 2 metres. Entre els més petits es troben els flagel·lats, i entre els més grans les meduses i els sifonòfors. Deixant una mica de banda les classificacions formals per mides i grups zoològics és important diferenciar en el zooplàncton dos tipus d’organismes: els que passen tota la vida al plàncton (holoplàncton) i els que en alguna fase del seu cicle vital colonitzen, en forma d’adults o de larves, el medi pelàgic (meroplàncton). Aquest és el cas de moltes fases larvals d’organismes bentònics sèssils o molt poc mòbils que utilitzen el medi pelàgic per a la dispersió. Les larves de peixos i d’un gran nombre d’espècies de crustacis decàpodes bentònics, per exemple, es desenvolupen enterament al plàncton.
Complexitat dels cicles biològics
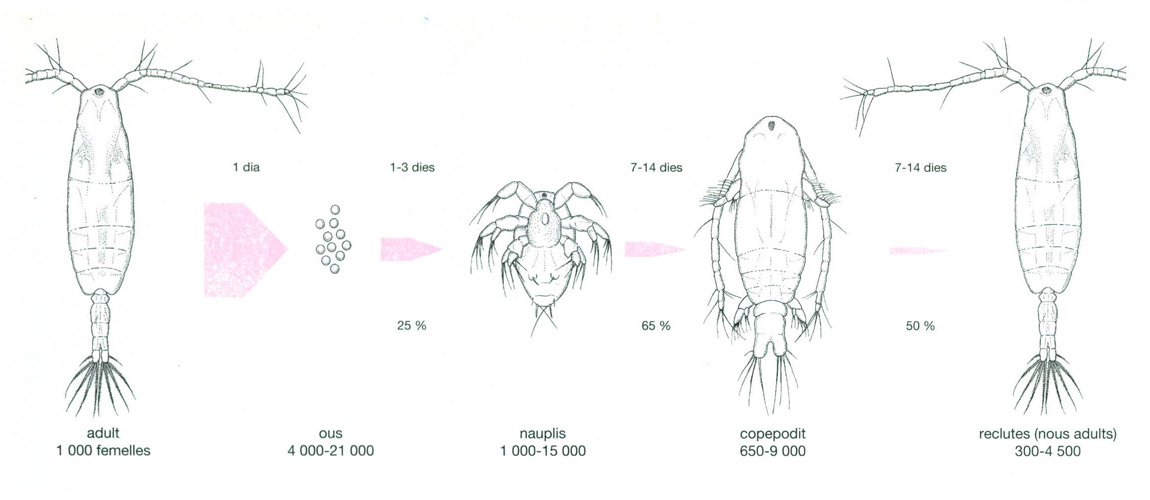
Diferents estadis del cicle biològic del copèpode marí Acartia clausi.
Jordi Corbera, a partir de fonts diverses
El cicle biològic de molts dels organismes del zooplàncton és força complex. Un exemple és el del copèpode Acartia clausi a les costes atlàntiques del continent americà. En el dibuix es poden veure els diferents estadis del cicle, amb el temps que duren els percentatges de supervivència i el nombre d’individus que van sobrevivint en cada estadi, a partir dels inicials nascuts de la posta. El cicle complet pot durar fins a 14 dies, però el de la població completa és més llarg, ja que les femelles adultes tenen una esperança de vida d’uns cinc dies i, mentre romanen en el plàncton, poden donar diverses postes d’ous segons la quantitat d’aliment disponible. En l’espècie A. tonsa, amb una concentració d’aliment d’uns 100 µg de carboni per litre, el 46% de l’energia s’inverteix en el creixement, el 33% en la reproducció i el 21% restant es perd per excreció. Si l’aliment s’incrementa quatre vegades, la ingestió ho fa 3,5 vegades i l’energia invertida en la reproducció es dobla. En canvi, en incrementar nou vegades l’aliment disponible, ho fa molt poc l’assimilació i gairebé no ho fa l’esforç reproductor.
Durant la seva estada al medi planctònic, els organismes del meroplàncton passen per diferents etapes de transformació morfològica fins a arribar a la fase de juvenil, que ja té una forma molt semblant a l’adult. El temps de residència de les fases larvals al plàncton varia molt segons les espècies, però per a totes representa el moment més crític del cicle vital. A més de les larves dels crustacis decàpodes, ja mencionades, altres grups d’animals bentònics contribueixen de manera semblant a la composició de les comunitats zooplanctòniques. S’han de destacar les larves d’equinoderms i les fases de medusa de molts cnidaris bentònics (pòlips), com també les de molts peixos. Aquests estadis larvals tenen un temps de residència al plàncton que oscil·la de setmanes fins a mesos i, durant curts períodes de temps, poden representar un percentatge elevat de la biomassa zooplanctònica dels sistemes costaners. Molts altres grups d’animals bentònics, com les esponges, els poliquets, els briozous, etc., produeixen també un gran nombre de larves planctòniques durant el període de reproducció sexual. Però, a diferència dels grups esmentats abans, les larves d’aquests amb prou feines romanen al plàncton enllà d’unes hores o uns dies i efectuen la primera metamorfosi així que han trobat un substrat on establir-se. La mortalitat d’aquestes larves és enorme; es beneficien del medi planctònic per a la dispersió, però paguen aquest avantatge amb la feixuga càrrega de constituir una font estacional d’aliment molt important per a altres organismes del zooplàncton costaner, que en devoren una gran part.
Si bé el meroplàncton pot ser un component important de les comunitats zooplanctòniques, la major part de la biomassa d’aquestes comunitats és constituïda per l’holoplàncton. Els copèpodes són de bon tros el grup dominant: constitueixen entre el 50 i el 90% de tots els individus d’aquestes comunitats; el seu cicle biològic es desenvolupa enterament en el plàncton. Els percentatges més alts de copèpodes s’han trobat a les plataformes continentals de la meitat nord de l’oceà Atlàntic, mentre que els més baixos s’han observat a les àrees tropicals de l’oceà Índic. Pel que fa a la biomassa, en valors de carboni orgànic, els copèpodes representen, a l’Atlàntic nord, una mica més del 50% a les zones oceàniques i prop d’un 75% a les àrees costaneres. Aquesta diferència de biomassa s’explica, en gran part, per la presència de grans poblacions oceàniques d’eufausiacis, que arriben a representar més del 30% de la biomassa zooplanctònica. La tendència observada a l’Atlàntic nord es repeteix en altres oceans, bé que a les àrees tropicals del Pacífic i de l’Índic la biomassa deguda als eufausiacis és més gran, i a més a més altres grups, com els amfípodes, els quetògnats, els cnidaris i els mol·luscs pteròpodes, representen cadascun prop del 5% de la biomassa total. Cal assenyalar que aquestes xifres generals subestimen la biomassa del zooplàncton gelatinós, format per meduses, sifonòfors, ctenòfors, mol·luscs pteròpodes, sàlpids i taliacis que, a causa de les seves grans dimensions i el fet de formar agrupacions denses o colònies llargues, no es recol·lecten eficaçment amb les xarxes de mostreig habituals per al zooplàncton. Recentment s’ha pogut avaluar en algunes àrees tant la seva biomassa com la seva activitat. A més a més de ser molt abundants, actuen com a carnívors voraços —especialment les grans meduses— que competeixen amb les larves de peixos com a depredadors del zooplàncton petit.
L’estructura tròfica del zooplàncton està marcada clarament per la dominància dels macroherbívors (filtradors i brostejadors) i els omnívors (en realitat herbívors capaços d’alimentar-se també de petites partícules inertes i de diminuts organismes). A les àrees temperades i polars representen més del 70% de la biomassa zooplanctònica entre els uns i els altres; en mars tropicals destaca l’abundància de depredadors (carnívors i raptors), que representen prop del 40% de la biomassa total. Aquestes diferències tròfiques estan relacionades amb la menor biomassa i producció del fitoplàncton de les mars tropicals, a causa de la pobresa en nutrients. A més a més, prop dels esculls de corall, els herbívors representen menys del 10% de la biomassa total, a causa de l’escassetat de fitoplàncton, ja que els esculls actuen com a trampes de nutrients i desenvolupen una rica flora endobiont i simbiont.
Les variacions estacionals
Els valors esmentats fan referència a una mitjana anual. Però, tal com passa als ecosistemes terrestres, els ecosistemes marins presenten variacions estacionals, més marcades com més allunyats es troben de l’equador. Aquesta variabilitat temporal es manifesta en una oscil·lació del nombre d’espècies i d’individus, causada, sobretot, per canvis d’activitat relacionats amb la modificació de les condicions hidrogràfiques de la columna d’aigua.
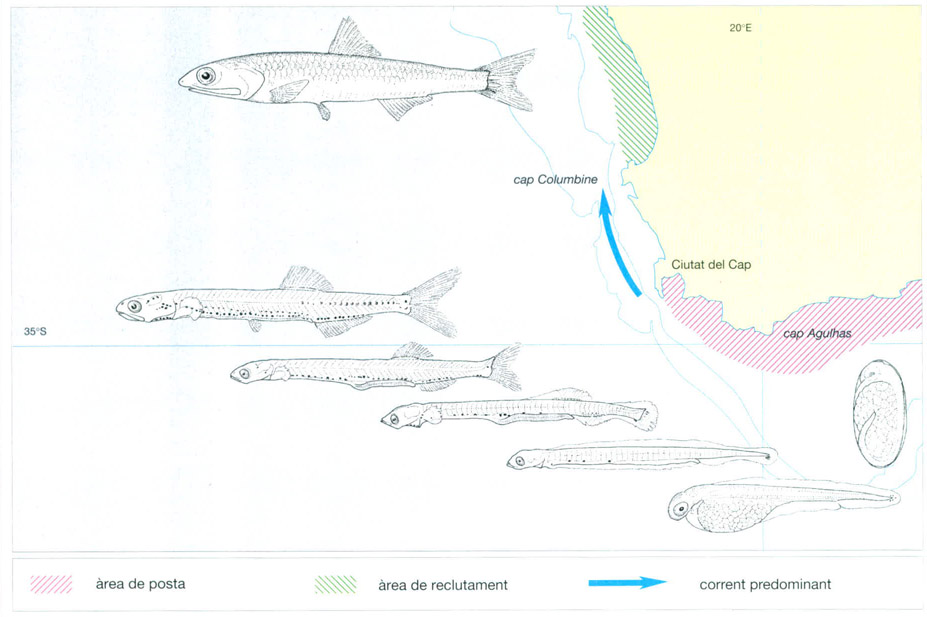
Els processos de dispersió i distribució de molts dels organismes del zooplàncton estan íntimament lligats a la dinàmica de les masses d’aigua. Un exemple d’aquest fenomen és el procés de dispersió que segueixen les larves de l’anxova Engraulis capensis, pròpia de l’Atlàntic sud-oriental. Els adults fan la posta a la zona del cap Agulhas, on la columna d’aigua està barrejada i té poca productivitat. Els ous eclosionen pocs dies després i surten unes larves lecitotròfiques que ja es poden alimentar per elles mateixes una setmana més tard. Després d’un mes fan gairebé 20 mm, i ja són juvenils quan fan de 60 a 70 mm de grandària; aleshores és quan es recluten. Les larves són arrossegades per un fort corrent de direcció N, que les transporta més de 240 km, per a deixar-les en una zona d’aflorament altament productiva, per damunt del cap Columbine, fenomen que els assegura una alimentació abundant. Els adults tornaran a la zona d’Agulhas a la primavera o l’estiu australs següents per fer la posta.
Jordi Corbera, a partir de fonts diverses
A les mars temperades el zooplàncton presenta dos moments de màxima abundància: un a la primavera i l’altre al final de l’estiu. En canvi, a les mars fredes es dóna un sol màxim d’abundància, durant l’estiu, mentre que a les àrees tropicals les concentracions de zooplàncton són constants al llarg de tot l’any. Aquests màxims d’abundància responen a un model hidrogràfic d’acord amb el qual a la primavera, quan augmenta el nombre d’hores d’insolació, s’accelera l’activitat fotosintètica del fitoplàncton, que, a més a més, aprofita els nutrients preservats de l’hivern anterior.
L’augment del fitoplàncton desencadena una resposta immediata per part del zooplàncton herbívor, constituït, majoritàriament, per copèpodes, que consumeixen entre el 20 i el 30% de la producció primària diària. El brostejament continuat dels copèpodes dóna lloc a un màxim de biomassa a la primavera, representada per individus de mida més gran que els de la resta de l’any. Els individus poden créixer ràpidament i arribar a la maduresa sexual en pocs dies. La seva esperança de vida és d’uns 20 dies i durant els últims 10 no paren de produir ous. Les femelles d’algunes espècies ponen uns quants centenars d’ous cada dia, bé que la taxa de producció decreix des de l’inici de la primavera, a causa de l’esgotament dels nutrients i, per tant, del fitoplàncton. El procés productiu generat pel zooplàncton herbívor és molt ràpid, de manera que en pocs dies la biomassa zooplanctònica augmenta fins més de 10 ordres de magnitud per damunt de la concentració hivernal. L’èxit dels copèpodes rau, en gran part, en les seves constants metabòliques, que els permeten créixer molt ràpidament. El fet que inverteixin més del 33% de l’energia que assimilen en el creixement i la reproducció, a més de tenir un coeficient d’assimilació de l’aliment capturat de vora el 60%, fa que siguin molt eficaços quan les condicions de concentració d’aliment ho permeten.
No s’ha de descartar que en mars càlides, durant la primavera, a més a més de copèpodes herbívors hi hagi concentracions elevades de salpes, capaces de consumir més del 40% de la producció primària diària, especialment quan s’ajunten en eixams densos. Aquests eixams poden omplir les xarxes de pesca, que poden recol·lectar diverses tones d’aquests organismes en pocs minuts d’arrossegament. En formar colònies de molts individus, les salpes poden competir eficaçment amb els copèpodes per l’aliment comú. Aquests últims solen presentar unes concentracions mitjanes de 5 a 10 individus per litre, mentre que les salpes poden superar els 20 individus.
Sincronitzat amb el màxim de zooplàncton herbívor, se succeeix un increment del carnívor, constituït per una gran diversitat de grups, des dels mateixos copèpodes fins a les grans meduses i les larves dels peixos. L’activitat del zooplàncton carnívor no pot impedir que els herbívors explotin amb gran eficàcia el fitoplàncton, ni que aquest esgoti els nutrients. Paral·lelament a l’augment d’activitat del zooplàncton es produeix un escalfament de les aigües superficials que origina una estratificació, més marcada a mesura que avança l’estiu. Tots dos fenòmens, el canvi de condicions hidrogràfiques i d’activitat dels organismes, porten a una disminució de l’abundància de zooplàncton. A la tardor, els vents que bufen arran de la superfície ajuden a trencar la termoclina estival, i els nutrients tornen a quedar a disposició del fitoplàncton, en resuspendre’s tota la matèria orgànica generada durant l’estiu i que en gran part estava retinguda sota les capes superficials per la termoclina. Es produeix, doncs, un segon màxim de concentració de zooplàncton, que en aquesta ocasió és format per espècies de copèpodes herbívors més petites —menys biomassa que a la primavera—, molt actives i eficaces, de manera que donen un màxim de producció. Posteriorment, quan amb l’arribada de l’hivern disminueix la temperatura i els dies es fan més curts, l’activitat del fitoplàncton es redueix considerablement, cosa que es correspon amb una disminució de la biomassa zooplanctònica.
A les mars càlides es generen fenòmens a macroescala de variació estacional de la diversitat i la biomassa zooplanctòniques, semblants al descrit per a les mars temperades, bé que depenen de factors diferents a les variacions tèrmiques, molt suaus a les àrees tropicals. Una diferència important és que, mentre a les mars temperades les variacions estacionals del zooplàncton succeeixen en qualsevol banda, als mars tropicals són locals i tenen lloc més freqüentment prop de la costa. Un exemple són els esdeveniments meteorològics a gran escala, com els canvis estacionals en el règim dels monsons a l’oceà Índic. Durant quasi la meitat de l’any els vents dominants són persistents i bufen de mar cap a terra, i durant l’altra meitat de l’any bufen en sentit contrari. En aquest últim cas es produeix un desplaçament de la massa d’aigua superficial cap a la mar oberta, desplaçament que es compensa amb un aflorament d’aigua profunda. Aquesta aigua és més freda i rica en nutrients, cosa que donarà lloc a una època de gran abundància de zooplàncton. Durant aquest període la comunitat està dominada per espècies que desenvolupen el cicle biològic en àrees costaneres amb un fort component meroplanctònic. En canvi, durant el període de vents procedents de mar oberta la comunitat zooplanctònica és dominada per espècies oceàniques, i la productivitat és menor, ja que la capa profunda queda retinguda sota la capa fòtica. Un exemple d’aquest tipus de variacions estacionals és la composició específica de les poblacions d’hidromeduses de les costes orientals de Papua Nova Guinea. Durant el període de vents costaners, la major part de les espècies tenen una fase pòlip bentònica que dóna lloc a un gran nombre de petites meduses planctòniques. Aquestes meduses romanen unes setmanes al plàncton abans que les seves larves, originades per reproducció sexual, tornin al fons. En canvi, durant el període de vents oceànics la comunitat és dominada per meduses de mida més gran, el cicle vital de les quals té lloc totalment al plàncton, on romanen diversos mesos.
Els genuïns monsons, molt més intensos, generen diferències estacionals importants a gran escala del zooplàncton de les mars càlides. Els monsons es corresponen amb les èpoques de pluges torrencials, quan els rius produeixen descàrregues de gran envergadura sobre la plataforma continental. L’elevada concentració de nutrients transportats per les aigües continentals dispara un cicle de producció biològic que afavoreix el desenvolupament de denses poblacions de zooplàncton herbívor. A causa de la persistència dels monsons, aquestes aportacions continentals són contínues, de manera que la successió a l’ecosistema planctònic permet la proliferació d’un gran nombre d’espècies carnívores i altres consumidors secundaris, bé que no s’arriba al nivell d’estructura i complexitat observat a les mars temperades durant la primavera.
Els sistemes de sustentació
El zooplàncton comprèn animals que viuen en contínua suspensió en l’aigua, però amb una certa capacitat de moviment, encara que limitada. L’activitat i la producció marines més grans es donen als primers 200 m de profunditat. Per això, la principal estratègia seguida pels organismes del zooplàncton consisteix a procurar no sedimentar. Per a aconseguir-ho han desenvolupat modificacions morfològiques o bé presenten algun tipus d’activitat que ofereixi resistència a la caiguda. Una de les adaptacions més generalitzades és disposar d’un seguit d’apèndixs o expansions corporals, de manera que la seva major superfície ofereix una major resistència a la sedimentació a través de la columna d’aigua. Així redueixen la necessitat de moure’s contínuament i, alhora, el desgast metabòlic que representaria una activitat locomotriu continuada. Hi ha nombrosos exemples d’aquesta estratègia, com les expansions de les larves dels equinoderms, els apèndixs i les prolongacions de les larves dels crustacis decàpodes, els parapodis dels mol·luscs o els apèndixs plomosos de molts copèpodes.
Una estratègia alternativa es basa en la inclusió dins del cos de fluids de baixa densitat, glòbuls d’oli o, fins i tot, petites cambres amb gas. Un exemple són els ous de peix, que contenen fluids diluïts per a garantir la flotabilitat, ja que en ser esfèrics i no tenir capacitat de locomoció caurien cap al fons ràpidament. Els sifonòfors presenten unes esferes plenes de gas al zooide superior de la colònia, que els permet mantenir-se permanentment en suspensió sense gaire esforç. Molts eufausiacis i copèpodes tenen inclusions olioses als teixits adiposos que, juntament amb la capacitat de desplaçament, els ajuden a flotar. Els organismes gelatinosos del zooplàncton (meduses, sifonòfors, ctenòfors, mol·luscs pteròpodes, sàlpids i taliacis) presenten unes propietats relacionades amb el canvi de l’equilibri iònic als teixits que els permeten regular la flotabilitat. Alguns sifonòfors i altres organismes, com el dinoflagel·lat Noctiluca, contenen cavitats amb solucions isosmòtiques de clorur d’amoni que poden regular per millorar la flotabilitat.
S’observa una gran diferència entre les espècies que han adaptat els seus cossos amb mecanismes de flotabilitat quasi neutra, que gairebé no els cal fer cap esforç per mantenir-se a un nivell, amb les que presenten alguna estructura pesada que les obliga a nedar contínuament per no enfonsar-se. Així, per exemple, els quetògnats del gènere Sagitta només han de nedar de tant en tant per mantenir-se a la profunditat desitjada, mentre que els mol·luscs del gènere Cavolinia, amb una closca pesada, han de nedar vigorosament mitjançant els seus apèndixs aliformes per no enfonsar-se. S’ha comprovat, d’una manera molt general, que els organismes que viuen en aigües tropicals (menys denses i viscoses que les temperades i les polars) són de mida més petita i tenen un nombre més gran d’apèndixs i de projeccions que les espècies d’altres mars.
La distribució del zooplàncton
La distribució del zooplàncton depèn principalment dels moviments de les masses d’aigua a nivell global. Tot i que els oceans estan interconnectats, el complex conjunt de corrents oceànics i estructures hidrogràfiques a macroescala i a mesoescala fan que en molt pocs casos les espècies del zooplàncton tinguin una distribució cosmopolita. Les espècies que presenten una distribució mundial han hagut de seguir tot un llarg procés de dispersió i adaptació des del moment en què es van originar, superant obstacles hidrogràfics, com les àrees de convergència i divergència. També han hagut de superar corrents a gran escala, com per exemple el de Cromwell (de 300 km d’amplada), que circula cap a l’E per sota de l’equador a una velocitat de tres nusos.
El zooplàncton es troba immers i en suspensió a les diferents masses d’aigua, a les quals adapta la seva fisiologia segons les característiques hidrogràfiques. Una massa d’aigua es caracteritza pels valors de temperatura i salinitat i, per tant, de densitat. En adaptar-se les diferents espècies a una determinada massa d’aigua s’origina una pauta de distribució molt heterogènia, com a conseqüència de la gran varietat d’aigües diferents que es poden trobar en un mateix ecosistema. Aquesta adequació de les comunitats zooplanctòniques a masses d’aigua determinades respon a una visió clàssica, contraposada a la visió més actual que atribueix a la hidrodinàmica un paper preponderant a l’hora d’explicar la distribució del zooplàncton a macroescala i a mesoescala. Es pot afirmar que la dinàmica de l’aigua origina mecanismes mitjançant els quals es crea l’heterogeneïtat en la distribució dels organismes planctònics.
Però no són només els factors ambientals els que influeixen en la distribució dels organismes zooplanctònics; s’han de considerar, també, els factors biològics resultants de l’activitat, la morfologia i la fisiologia dels mateixos organismes. La distribució heterogènia del zooplàncton s’expressa tant en l’espai com en el temps. L’opinió generalitzada, bé que no acceptada per tots, és que la variabilitat espacial és deguda a factors físics, mentre que les variacions temporals responen a factors biològics. Aquesta controvèrsia ha estimulat moltes publicacions, tot i que, en realitat, tots dos tipus de factors s’interrelacionen a cada àrea concreta, on la preponderància d’un dels dos depèn de l’escala a què té lloc el fenomen que s’observa.
La distribució a escala oceànica
A gran escala, són el règim de corrents oceànics i els remolins ciclònics els que delimiten les regions biogeogràfiques de les diferents faunes planctòniques. A les àrees de trobada d’aquest tipus de corrents es desenvolupen els anomenats corrents o zones de frontera, que actuen com a límits físics de la distribució de les espècies. A l’hemisferi sud, per exemple, la confluència entre el corrent de Benguela i el d’Agulhas (que prové de l’oceà Índic), a la costa occidental de l’Àfrica meridional, origina una zona de divergència que actua com a frontera entre les faunes del sud de l’Índic i les de l’Atlàntic. Zones de convergència-divergència a escala oceànica, com el front polar, també a l’hemisferi sud, actuen com a fronteres naturals tant per a la penetració d’espècies a l’oceà Antàrtic com per a la sortida de les espècies pròpies d’aquest.
La distribució a escales intermèdies
L’heterogeneïtat espacial i temporal a mesoescala del zooplàncton (aproximadament entre 100 i 10 km, i entre tres mesos i quinze dies) és regida per un seguit d’estructures hidrogràfiques, més o menys persistents, que produeixen fenòmens de dispersió i concentració. Com ja s’ha dit, el zooplàncton no presenta una distribució uniforme, sinó que tendeix a concentrar-se en certes àrees, perquè en aquestes tenen lloc fenòmens hidrogràfics a l’escala indicada. De fet, sembla evolutivament positiu que les diferents espècies hagin adaptat la seva biologia a l’existència d’una variabilitat hidrogràfica més o menys permanent, ja que d’aquesta manera aconsegueixen viure juntes en un mateix sistema marí, sense dispersar-se excessivament. L’organització que proporciona el marc espacial té una importància vital per al manteniment de les cadenes tròfiques marines, on la inversió en la recerca d’aliment no pot ser superior al guany que proporciona el fet de trobar-lo.
Els processos advectius
Uns dels fenòmens hidrogràfics a mesoescala més coneguts són els processos advectius. Els corrents que periòdicament desplacen masses d’aigua sobre la plataforma continental són responsables dels gradients de zooplàncton del llarg de les zones costaneres. Per exemple, les intrusions contínues del corrent de Califòrnia, ric de nutrients, són responsables de les variacions interanuals en la biomassa zooplanctònica d’aquelles aigües, i generen canvis en l’estructura de les comunitats de zooplàncton, en incrementar la quantitat d’herbívors associats amb l’extrem sud del corrent. La proliferació d’aquest zooplàncton herbívor desencadenarà un augment de les poblacions de carnívors de les costes de Califòrnia. D’aquesta manera, les variacions interanuals en les intrusions del corrent determinen la riquesa de preses per a les larves de peixos, amb la consegüent repercussió en l’augment de les poblacions anuals d’espècies comercials de peixos.
En altres casos l’impacte dels processos advectius sobre les comunitats zooplanctòniques vernacles pot no ser tan positiu. Per exemple, al nord del sistema de Benguela, coincidint amb una disminució de l’aflorament, es produeix una entrada d’aigües càlides amb el corrent d’Angola. Aquestes aigües transporten un elevat nombre de meduses i sifonòfors que, a causa de la seva voracitat com a depredadors, poden exercir un efecte negatiu sobre la resta del zooplàncton de l’àrea. De fet es considera que la concentració de copèpodes al nord del corrent de Benguela (tres vegades més petita que l’observada al sud durant la relaxació de l’aflorament en aigües superficials) es deu a l’elevada depredació de l’anomenat zooplàncton gelatinós aportat amb les intrusions d’aigua d’Angola. En altres àrees, com a les costes d’Oregon i a les costes catalanes, s’observa que el desplaçament d’un corrent paral·lel a la costa tendeix a delimitar l’espai de les comunitats zooplanctòniques de la plataforma. Així, la zona situada entre la línia de costa i el corrent es caracteritza per un zooplàncton que rarament s’allunya de la costa, mentre que a l’altra banda del corrent es desenvolupa una comunitat dominada per espècies holoplanctòniques. Entre totes dues, associada al corrent, es troba una comunitat típica de plataforma, que actua de transició entre la comunitat costanera i l’oceànica. Un relaxament del corrent donarà lloc a una penetració d’espècies meroplanctòniques prop de la costa, amb el consegüent augment de la competència específica o la dispersió i possible pèrdua de les espècies costaneres.
Un altre tipus de processos advectius molt importants per al zooplàncton són les intrusions d’aigües continentals procedents de rius grans, per exemple del Mississipí. Aquestes plomes d’aigua van associades amb processos productius, a causa de l’elevada concentració de nutrients, màxima a l’extrem de la ploma. Allà s’observa una gran aglomeració de zooplàncton que serveix d’aliment a les larves dels peixos, les poblacions dels quals augmentaran segons la intensitat de les aportacions del riu.
Les ones internes
Les ones internes també són una font de variabilitat de les comunitats zooplanctòniques. En relació amb el relleu del fons, el pas dels corrents sobre la plataforma continental genera un seguit d’ones internes a les quals s’associa un fenomen de concentració de nutrients i un increment de la producció primària. Al mateix temps actuen com a ones de transport que afavoreixen la concentració local del zooplàncton. La seva importància es fa palesa, per exemple, en el desplaçament d’algunes poblacions de larves de crustacis decàpodes, que es traslladen al llarg de la costa per a l’estrat íntimament associat a les ones internes, aprofitant la concentració d’aliment per sobreviure i dur a terme la metamorfosi.
Els fronts
Els fronts hidrogràfics actuen com a zones de convergència, creant una discontinuïtat important en la distribució horitzontal de les masses d’aigua. Van associats amb una elevada producció planctònica, que dóna un increment de la producció i la biomassa de zooplàncton. Així, els fronts actuen tant com a zones d’acumulació com de retenció i transport de zooplàncton. Al llarg del front es donen processos de pujada i baixada de les masses d’aigua, que retenen els organismes prop de la superfície i, gràcies als processos de mescla, eviten la sedimentació dels nutrients. En fronts localitzats a la mar Lígur o a la mar del Nord s’ha observat una concentració de larves de copèpodes molt superior a la de les aigües circumdants. En un front del canal de la Mànega les concentracions poden ser fins a 75 vegades superiors a les “normals” d’aigües costaneres properes al front. Això proporciona un hàbitat idoni per a la proliferació d’un ric zooplàncton carnívor, com les larves de peixos.
Mentre els fronts oceànics actuen com a zones de retenció, els de plataforma i de talús actuen, a més a més, com a barreres a la dispersió. En un front localitzat al final de la plataforma continental, a la Mediterrània occidental, s’ha observat una gran aglomeració de larves de peix, els adults de les quals es troben en aigües costaneres. Les larves d’aquestes espècies podrien dispersar-se cap a mar oberta, però els fronts de plataforma i de talús eviten la seva pèrdua i faciliten el seu posterior assentament. A més a més, associades al front es troben un gran nombre de larves d’espècies mesopelàgiques, fet que incrementa la competència interespecífica per l’aliment, possiblement resolta gràcies a l’elevada taxa de producció planctònica associada al front. Igual que en altres fronts de plataforma i de talús, a la cara oceànica hi circula un corrent, el qual és el causant de la formació del front on topen les aigües de plataforma amb les de talús. D’aquesta manera, l’efecte de barrera hidrogràfica limita la dispersió del zooplàncton de plataforma.
Els fronts de marea i els d’estuari també van associats amb valors de productivitat i biomassa planctòniques més alts que a les aigües circumdants. Per exemple, al George Bank s’ha constatat l’acció com a barrera d’un front de marees que evita la dispersió de les espècies planctòniques cap a fora. A més a més, a la part externa d’aquest front hi ha associada una elevada biomassa zooplanctònica, que s’introdueix cap a l’interior del banc mitjançant un corrent de fons compensatori. Aquest corrent associat al front té una gran importància perquè subministra l’aliment necessari per a mantenir les poblacions d’ictioplàncton, d’importància vital per a mantenir els estocs de les espècies que s’exploten a l’àrea. En alguns estuaris de la costa oriental del Canadà es va comprovar la sincronització entre l’existència d’un front estuàric i el desenvolupament de les poblacions de larves de peix. El front s’anava desplaçant cap a l’exterior de l’estuari, i associat a les zones de mescla es trobava un gran nombre de larves. Cap a l’interior de l’estuari, les larves eren molt petites i s’alimentaven de nauplis de copèpodes. A mesura que el front s’acostava a la desembocadura de l’estuari, les larves eren de mida més gran, i passaven a alimentar-se de copèpodes petits i posteriorment d’adults. L’adaptació alimentària de la larva amb l’increment de grandària permetia que els recursos no s’esgotessin abans de finalitzar el desenvolupament.
Els vents
A les costes occidentals dels continents es produeix un règim de vents favorable a un aflorament d’aigües profundes. Aquestes aigües que afloren són riques de nutrients, i disparen un procés de producció tan gran que situa aquestes zones d’aflorament entre les més productives de tots els oceans. El zooplàncton respon produint denses poblacions d’espècies oportunistes, majoritàriament herbívores, amb la qual cosa resulten unes comunitats poc diversificades, dominades per copèpodes calanoides. El sistema pot sustentar una elevada biomassa zooplanctònica, que és la base de l’aliment dels consumidors secundaris (peixos i cefalòpodes).
El desplaçament de les masses d’aigua superficials cap a mar oberta origina el corrent divergent d’Ekman, que és utilitzat per moltes larves de peix per allunyar-se de la costa. Però aquest desplaçament pot ser d’efectes negatius, en allunyar les larves del centre de producció. La solució utilitzada per evitar aquest transport forçat ha estat el desenvolupament de migracions verticals, que situen els individus per sota la capa d’aigua que es desplaça quan bufen els vents de terra cap a mar oberta. A les costes de Namíbia i el Perú s’ha observat que quan l’aflorament es relaxa, les comunitats zooplanctòniques esdevenen més complexes, en incrementar el nombre d’espècies i disminuir el nombre d’individus de les espècies dominants. Al mateix temps, les espècies més oportunistes de copèpodes són substituïdes per espècies amb un cicle de vida més llarg i amb una capacitat de producció d’ous i de creixement menors. Les densitats de copèpodes a l’aflorament de Benguela varien entre més de 4 000 individus per metre cúbic, quan l’aflorament és suau, fins a més de 12 000 quan la intensitat és màxima.
Els corrents
Els corrents de gran intensitat sovint produeixen una sèrie de filaments que desplacen masses d’aigua de grans dimensions cap a fora del cos central del corrent. Aquestes masses d’aigua formen uns remolins que engloben una comunitat zooplanctònica similar a la del corrent que l’ha originat. Els remolins queden immersos dintre d’altres masses d’aigua amb unes comunitats zooplanctòniques molt diferents. La circulació contínua dels remolins permet que la comunitat zooplanctònica del seu interior es mantingui prop de la superfície i pugui evolucionar. Al cap d’un temps, organismes procedents de les masses d’aigua circumdants penetren al seu interior i canvien l’estructura de la comunitat, de manera que esdevé més barrejada.
S’han utilitzat remolins originats al corrent del Golf per estudiar l’evolució de comunitats aïllades de zooplàncton, ja que la persistència del remolí pot ser de diversos mesos. Estructures semblants —com els anells o cercles d’aigua càlida associats al corrent de Kuro-Shivo o al de l’E d’Austràlia— actuen com a minisistemes que mantenen aïllades durant un cert temps comunitats de zooplàncton molt específiques. Aquestes estructures hidrogràfiques mantenen aïllades força espècies, que es desenvolupen sense quasi cap interferència exterior. S’ha observat que els remolins de les aigües de Hawaii engloben un gran nombre de larves de peix, els adults de les quals es troben als esculls de corall. Aquestes larves fugen de l’hàbitat dels progenitors, ja que els peixos adults són els seus principals depredadors. Duen a terme el seu creixement a l’interior dels remolins fins a arribar a l’estadi de juvenil, i tornen cap a l’hàbitat de l’adult, on s’aplegaran. Aquests mecanismes de retenció es consideren molt importants per a garantir la supervivència de les larves, ja que, confinades dins el remolí, poden accedir amb més facilitat als recursos alimentaris, més concentrats que en l’exterior, on els organismes es troben més dispersos.
Hi ha altres processos a mesoescala que també contribueixen a augmentar l’heterogeneïtat espacial i temporal del zooplàncton. Fenòmens com els corrents residuals associats a remolins o a fronts, la formació de capes discretes d’acumulació de plàncton o la circulació als canyons submarins introdueixen variabilitat en la distribució a mesoescala del zooplàncton. Als canyons submarins del George Bank, per exemple, es troben concentracions elevades d’eufausiacis (més de 1 000 individus per metre quadrat). Aquestes concentracions ajuden a explicar l’elevada producció de l’àrea, ja que la biomassa zooplanctònica de l’interior del banc i la seva producció no expliquen com es cobreixen les necessitats de les poblacions de peixos. La troballa de les denses concentracions de crustacis, associats als corrents de fons dels canyons submarins del banc, permet tancar el cicle productiu.
La distribució a petita escala
Tot i que a mesoescala i a macroescala la variabilitat en la distribució, l’abundància i la producció del zooplàncton és regida principalment per processos hidrodinàmics, a petita escala sembla que els factors biològics adquireixen una importància més gran. Tanmateix, hi ha un seguit de fenòmens físics relacionats amb la formació de cèl·lules de convecció o de Langmuir que poden influir en la distribució a petita escala del plàncton (entre 10 i 100 m).
Els factors que influeixen en la distribució
La pèrdua de calor de l’aigua superficial produeix un augment de densitat d’aquesta aigua, que durant la nit s’enfonsa i és substituïda per una aigua més calenta dels fons. Així es generen les cèl·lules de convecció, en un costat de les quals es produeix un procés d’enfonsament i en l’altre, d’emergència. Els organismes amb flotabilitat positiva podran situar-se a les zones de convergència, aprofitant la caiguda d’aliment des de la superfície. Els organismes amb flotabilitat negativa tendiran a trobar-se a les zones de divergència, mentre que els que tenen flotabilitat neutra podran concentrar-se a la base de la cèl·lula o a la seva superfície. Aquest suport físic de la distribució del plàncton afavoreix la tendència a l’agregació per ocupar els espais idonis segons les espècies. L’heterogeneïtat aconseguida és molt variada, ja que una cèl·lula de convecció pot fer des de pocs metres de diàmetre fins a quasi 200. Tanmateix, factors relacionats amb el comportament social de les espècies, la demografia, els cicles de vida i les relacions intraspecífiques i interspecífiques tenen un paper primordial a l’hora de buscar les causes de la distribució a petita escala.
El nivell d’heterogeneïtat (variació en el nombre d’espècies o d’individus) que es pot trobar en més d’un quilòmetre en sentit horitzontal equival al que s’observa en una desena de metres, o pocs més, en sentit vertical. La major part dels organismes del zooplàncton tendeixen a concentrar-se a la capa d’aigua compresa en els primers 200 m (a la capa fòtica o just per sota seu). Per exemple, a la plataforma continental atlàntica d’Amèrica del Nord la biomassa mitjana anual de zooplàncton als primers 50 m és de 199 cm3 per 1 000 m3 d’aigua; entre 50 i 100 m és de 94 cm3, de 35 cm3 entre 100 i 200 m, i a partir dels 500 m és inferior a 20 cm3 per 1 000 m3 d’aigua. Aquestes diferències dels valors de biomassa zooplanctònica són una mica més suaus en altres àrees, com a la mar del Sargassos o al corrent del Golf, però els valors de biomassa dels primers 100 m són sempre el doble, o més, que els detectats a partir dels 200 m, i deu vegades més elevats que els trobats a més de 500 m de profunditat. Paral·lelament a aquesta abundància més gran de zooplàncton a les capes superficials, s’ha observat una pauta de desplaçament dels organismes en sentit vertical que configura un ambient molt actiu davant la resta de la massa d’aigua oceànica.
Els eixams d’organismes
La tendència a viure agregats, formant taques o núvols monospecífics, és una estratègia àmpliament seguida pels organismes del zooplàncton. Diferències o gradients de salinitat i temperatura, gradients d’intensitat lluminosa, distribució dels recursos alimentaris, presència de depredadors i comportament social són causes que generen agregació. La mida de la taca o eixam depèn de cada espècie, i la grandària dels individus d’una determinada espècie varia segons els factors externs i durant el desenvolupament. Per exemple, en els copèpodes els eixams de nauplis generalment són més grans que els eixams d’adults, potser perquè hi ha un nombre més elevat d’individus per eixam. Al mateix temps, un eixam d’adults pot ampliar-se o reduir-se quan troba una acumulació d’aliment o detecta la presència d’un depredador. Així, els eixams funcionen com la manxa d’un acordió, i presenten unes dimensions fluctuants.
La mida de la zona per on es distribueix una espècie pot fluctuar durant el dia, al llarg del desenvolupament o segons les àrees on es trobi. Una espècie pot presentar fases d’agregació i fases de dispersió. Aquesta fluctuació en la distribució fa que l’avaluació de la densitat de població mitjançant els mètodes convencionals (xarxes o bombes de succió) sigui molt difícil. La densitat al centre d’un eixam de copèpodes és de 100 a 1 000 vegades més gran que la densitat mitjana de la població global de l’àrea. A més a més, el paper com a presa o depredador en les relacions tròfiques de la comunitat canvia molt si els copèpodes es troben en fase d’agregació o bé de dispersió.
Els avantatges o el valor adaptatiu de la formació d’eixams són encara avui molt discutits. Considerem, per exemple, un eixam de larves de sardina. Els individus del centre de l’eixam estan molt ben protegits dels depredadors; en canvi, estan molt allunyats de l’aliment, que es troba a la perifèria de l’eixam. Una alternativa consisteix a romandre junts durant el dia per evitar els depredadors i dispersar-se durant la nit, quan es troba aliment. En general i a escala de sistema, la formació d’agregats facilita la distribució i el reciclatge de l’energia disponible. Els individus solitaris s’han de desplaçar contínuament per capturar les seves preses i, al mateix temps, és molt difícil que siguin capturats, de manera que hi ha moltes probabilitats que es perdin per a la cadena tròfica (almenys en els primers 200 m de profunditat). Els productes d’excreció del zooplàncton són vitals com a nutrients per al fitoplàncton. Els productes excretats pels eixams són més fàcils de captar per les algues, ja que es troben molt menys dispersos que els produïts pels individus errants.
Les característiques d’un agregat o eixam són molt diferents entre les diferents espècies. Si comparem, per exemple, dues espècies de copèpodes com ara Calanus finmarchicus i C. tonsus, podrem observar pautes diferents. La primera espècie forma eixams que duren entre 12 hores i alguns dies, després es desfan i al cap d’uns dies es tornen a formar. Els eixams de C. finmarchicus tenen entre 1 i 3 m de diàmetre, i una densitat de més d’un milió d’individus per metre cúbic. La segona espècie forma eixams de 100 m de diàmetre a 1 km, amb una densitat de 10 000 individus per metre cúbic. La distància mitjana entre individus d’un mateix eixam és molt variada. En copèpodes pot ser inferior a 1 cm, mentre que en eufausiacis és de 3 a 5 cm i en meduses grans de fins a 100 cm. Estudiant els eixams de copèpodes que viuen als esculls coral·lins s’ha observat que un mateix grup pot formar eixams de característiques diferents. Per un costat es formen denses agrupacions monospecífiques d’un metre cúbic de volum amb densitats de més de mig milió d’individus per metre cúbic, situades just sobre la superfície de les colònies de coralls. Hi ha eixams constituïts per individus de la mateixa mida però d’espècies diferents, sense una relació social entre ells. Altres eixams mostren un comportament social, com molts eixams de peixos.
Els eixams d’una mateixa espècie presenten una composició demogràfica diferenciada. Per exemple, en agregats de meduses com Ropilema sculentum (amb una ombrel·la de 20 a 40 cm de diàmetre) els individus de la mateixa mida s’agreguen, segons la seva capacitat de natació, formant un conjunt d’eixams dins d’un eixam més gran. Tots els diferents eixams es desplacen plegats, a una velocitat d’uns 200 m/h, per trobar aliment. En moltes espècies, l’origen d’un eixam és reproductiu. Tots els individus d’un eixam s’han originat al mateix temps per una població d’adults. Per exemple, els adults d’un eixam de sardines efectuen la posta tots junts, de manera que els ous formen un sol agregat que queda a mercè dels corrents. A mesura que va creixent, l’eixam perd una part de la seva població, especialment els individus de la perifèria, mentre que els restants continuen junts fins a l’estat de juvenils o fins i tot d’adults.
Els desplaçaments verticals del zooplàncton
Els desplaçaments verticals de les espècies zooplanctòniques responen a diverses causes, bé que el factor que sembla desencadenar la migració és la llum. Les migracions verticals també poden tenir un component ontogènic, ja que els eixams de les formes larvals, els juvenils o els adults d’una mateixa espècie poden seguir pautes de migració diferents. En els quetògnats s’ha observat que mentre els adults es desplacen al llarg de la columna d’aigua, els juvenils formen eixams situats a profunditats molt discretes, prop del nivell de màxima abundància de les seves preses, que són eixams de copèpodes de mida petita.
Els moviments migratoris
La majoria d’espècies pugen cap a la superfície durant la nit o el vespre, i davallen en fer-se de dia. Com s’ha comprovat per algunes espècies de copèpodes i altres crustacis, canvis molt suaus en la intensitat lumínica que incideix a la superfície de la mar provoquen una reacció de desplaçament més o menys llarga, depenent de la freqüència de l’estímul. Altres tipus d’obstacles que poden situar-se entre la llum i l’organisme receptor produint ombra també estimulen l’individu a desplaçar-se en sentit vertical. Tot i que la major part de les migracions tenen lloc a les capes superiors de la columna d’aigua, també són habituals en organismes que viuen per sota dels 500 m de profunditat. La diferència consisteix en el fet que a la capa fòtica els ritmes d’activitat són molt curts, perquè responen al cicle dia-nit, mentre que a profunditats més grans els ritmes de migració són més lents, d’uns quants dies.
De tota manera, la velocitat de desplaçament vertical i la intensitat de la migració depenen del grup i de les condicions hidrogràfiques de cada àrea geogràfica. Els copèpodes de mida mitjana, per exemple, poden desplaçar-se entre 30 i 60 m en una hora, mentre que els eufausiacis poden ascendir entre 100 i 400 m en una hora. Altres espècies més litorals, com algunes larves dels glans de mar del gènere Balanus, pugen tan sols uns 15 m en una hora, bé que per a aquests organismes és fonamental que les larves no s’allunyin massa de l’hàbitat dels adults, i si migressin a grans distàncies serien arrossegades pels corrents. La velocitat de descens, aprofitant la gravetat, pot ser bastant més elevada, amb el consegüent avantatge d’escapar-se dels depredadors. A més a més, en moltes espècies les migracions poden ser inverses, o sigui, durant la nit són descendents, allunyant-se també del camp d’acció dels depredadors.
Les migracions alimentàries
Les migracions representen un cert cost metabòlic, que es compensa, però, amb els beneficis que comporta. El sentit evolutiu de la migració es basa en l’envergadura d’aquests beneficis. Per una banda, el fet de desplaçar-se al llarg de la columna d’aigua està relacionat amb la recerca d’aliment. Moltes poblacions de copèpodes se situen per sota el nivell màxim de clorofil·la, i durant la nit pugen a alimentar-se, mentre que d’altres resten a la superfície on pasturen sobre el fitoplàncton. La migració dels copèpodes comporta un desplaçament sincrònic dels seus depredadors, que persegueixen els eixams que pugen o baixen. S’ha de tenir en compte que molts depredadors són caçadors visuals, com les larves de peix, de manera que les preses durant el dia migren cap a capes menys il·luminades, i així és més difícil que els depredadors les localitzin. Molts investigadors consideren la migració com una estratègia per escapar-se dels depredadors. Una altra explicació, aquesta metabòlica, resideix en el fet que a les aigües superficials, més càlides, els copèpodes produeixen un nombre més gran d’ous i, a més a més, tenen taxes de creixement més elevades. En canvi, en aigües més profundes i fredes es frena el metabolisme, i el creixement de la població és menor.
De tota manera, el punt en què estan d’acord tots els investigadors és que la migració és regida per un conjunt de factors biològics íntimament relacionats. Un model representatiu de com s’explica la migració podria ser el següent. Un eixam d’una determinada espècie de copèpode, que durant el dia se situa a uns 100 m de profunditat, quan s’amaga el Sol comença a desplaçar-se cap a la superfície. L’eixam es desplaça regularment, de manera que els individus de l’extrem apical arribaran més amunt. Quan arriben a la capa de màxima densitat de fitoplàncton, pasturen contínuament i els individus que ja estan tips comencen a baixar, mentre pugen els restants membres del grup. A l’alba, la població baixa ràpidament, ja que en tenir els estómacs plens d’algues es fan més visibles als depredadors. Mentre romanen a la superfície, s’alimenten i excreten contínuament i, en accelerar el seu metabolisme, també poden produir un nombre més gran d’ous. De fet, la posta es realitza majoritàriament durant la nit, com s’ha observat, per exemple, en el copèpode Acartia pacifica a la mar del Japó; durant el dia la producció és d’uns 30 ous per metre cúbic i hora, mentre que a la nit és de 150. Un cop han arribat a capes més fredes relaxen els moviments i, en disminuir l’activitat metabòlica, passen el temps digerint l’aliment capturat durant la nit.
Les migracions defensives
Molts organismes zooplanctònics són transparents, per exemple els quetògnats i les meduses. Durant les hores de llum aquests organismes són quasi invisibles per als seus depredadors visuals i poden restar a la superfície. Però per alimentar-se han d’allunyar-se de les zones il·luminades de la superfície, ja que les preses els fan visibles, i, per tant, baixen a capes menys il·luminades. En aquestes capes la temperatura és menor, de manera que poden digerir les preses més lentament i esperar la nit o el dia següent —segons la velocitat de digestió— per tornar a pujar a caçar.
Hi ha moltes espècies que en determinades circumstàncies no migren. Si allà on es troba el depredador hi ha una concentració suficient de preses disponibles, aquest no té necessitat de migrar, i s’estalvia els costos que representa. Si la zona on es troba el nucli d’un eixam coincideix amb una concentració suficient d’aliment disponible, i no es detecten depredadors propers, l’eixam opta per no desplaçar-se.
Els obstacles migratoris
A part les circumstàncies tròfiques, també hi ha factors físics que alteren la migració de moltes espècies. L’existència d’una termoclina ben formada pot actuar com a barrera a la migració. Un exemple de l’acció de la termoclina és la pauta de distribució vertical observada en diferents grups de zooplàncton del nord de l’ecosistema de Benguela. En aquesta zona les intrusions d’aigües càlides d’Angola donen lloc a una forta termoclina, d’uns 6°C, situada a uns 50 m de profunditat. Moltes espècies de meduses, quetògnats i amfípodes es concentren just per sobre la termoclina. Les elevades concentracions de copèpodes a la superfície durant tot el cicle diari i l’elevat cost que representa creuar la discontinuïtat tèrmica de la termoclina són les causes que les espècies no es moguin, o bé que realitzin desplaçaments curts entre la termoclina i la superfície.
L’estratègia migratòria
Dins la comunitat planctònica, un dels guanys de la migració està relacionat amb l’excreció dels copèpodes. Aquests, en desprendre els seus excrements des de les capes superficials en forma de paquets fecals, contribueixen a la remineralització dels nutrients. D’aquesta manera, els productes de defecació poden ser descompostos gràcies a l’acció dels bacteris, prop de la capa fòtica durant el seu trànsit descendent. Cal assenyalar la importància de l’activitat descomponedora dels bacteris en les cadenes tròfiques marines, ja que remineralitzen els productes d’excreció a les capes fòtiques, i amb la seva acció generen un increment dels nutrients per al fitoplàncton. Un altre grup important són els flagel·lats, que a més a més de presentar una alimentació de tipus heteròtrof, com la majoria de protozous, també posseeixen pigments que els permeten una alimentació autòtrofa. Els organismes del zooplàncton que ingereixen aquests protozous aconsegueixen incrementar en més d’un 50% el rendiment de les cadenes tròfiques planctòniques.
El fet que la major part de les espècies del zooplàncton es concentrin a les capes fòtiques arriba a plantejar un problema de denso-dependència. Aquesta implica que les diferents espècies es reparteixen —moltes vegades mitjançant moviments migratoris— al llarg de la columna d’aigua, ocupant nivells sovint molt discrets. Per exemple, estudiant la distribució vertical d’algunes espècies de copèpodes a la mar Negra, es va observar que cada espècie situava el nucli central de la seva població a una determinada profunditat. Així Acartia clausi se situava entre 9 i 11 m, Paracalanus parvus entre 4 i 5 m i Oithona nana entre 5 i 7 m de profunditat. La denso-dependència també crea problemes de competència tròfica. Una solució va ser, en el cas de larves de diverses espècies de peixos estudiades a les costes britàniques, seleccionar el tipus de presa segons la mida i el nivell de distribució de les espècies. Així, mentre una espècie s’especialitzava a depredar preferentment nauplis d’una determinada espècie de copèpode, altres larves s’alimentaven de petits copèpodes d’una altra espècie. Aquest procés de selecció de presa a petita escala (a menys de 60 m de profunditat) sembla que és vital per a la supervivència de comunitats zooplanctòniques tan variades com les de les aigües superficials.
Altres exemples de compartimentació de l’hàbitat són la distribució a diferents nivells de profunditat dels eixams d’eufausiacis a l’ecosistema de Benguela, o la selecció per part dels amfípodes hipèrids de diferents espècies de zooplàncton gelatinós (situats a diferents fondàries) com a substrat on s’estableixen, i del qual s’alimenten quan els recursos de l’ambient escassegen. A les comunitats zooplanctòniques les relacions interspecífiques són molt més subtils del que es creia fins fa poc. Un cas seria el de la compartimentació de l’hàbitat entre les larves d’anxova i sorell a l’Atlàntic nord. Les larves d’anxova són menys mòbils i, en tenir la boca petita, capturen preses també petites, però que formen eixams grans. Les larves de sorell, més actives, capturen preses grans, més diluïdes però més rendibles energèticament.
Les relacions interspecífiques del zooplàncton
Les relacions interspecífiques a petita escala i a mesoescala queden reflectides en la constitució de les cadenes tròfiques zooplanctòniques. En les comunitats zooplanctòniques, altrament, els processos biològics estan acoblats a processos físics de la mateixa escala.
Les relacions tròfiques
Les relacions tròfiques generalment són força complexes, ja que no tan sols s’ha de considerar l’abundància de preses i depredadors, sinó també la possibilitat que ambdues puguin trobar-se en medi planctònic. Un exemple poden ser les relacions tròfiques establertes a les costes de l’illa de Vancouver.
Un exemple de la complexitat de les relacions tròfiques en el sistema planctònic pot ser el d’una comunitat composta per 1 000 individus per m3 del copèpode Pseudocalanus minutus (d’una mida d’1 mm), 400 del copèpode Calanus plumchrus (4 mm) i 10 de l’eufausiaci Euphausia pacifica (20 mm), que competeixen per pasturar sobre una població de diatomees Chaetoceros (23 mm). Els que consumeixen més són els eufausiacis, que esbiaixen la població de Chaetoceros en més d’un 50%, i les que resten queden a disposició dels copèpodes. Entre aquests, C. plumchrus compensa la seva dieta quan troba un eixam d’un flagel·lat d’una mida de 10 mm, mentre que l’altra espècie de copèpode s’alimenta només de Chaetoceros. Al cap d’uns quants dies es produeixen grans quantitats d’ous d’eufausiacis, que en tres o quatre setmanes donaran lloc a una generació de larves furcília. Les larves furcília competeixen amb l’eixam de C. plumchrus per alimentar-se dels flagel·lats. D’a-questa manera, les dues espècies tenen una taxa de producció més alta que P. minutus. El desenvolupament més gran dels eixams de furcília i d’aquest darrer copèpode davant la població de P. minutus fa que siguin detectats més fàcilment per les larves del salmó Oncorhynchus gorbuscha, de fins a 8 mm de longitud. Un possible depredador secundari de les larves de salmó podrien ser les meduses (considerades com a voraces depredadores de larves de peixos), sempre que els eixams d’ambdós grups coincideixin. Al mateix temps, les meduses poden competir amb èxit amb les larves de peix, ja que consumeixen grans quantitats de copèpodes. Arribat a aquest punt, les larves de salmó prefereixen els copèpodes (i les fases larvals) de l’espècie més petita, mentre que la medusa agafarà els de l’espècie més gran. Estudis recents duts a terme a Chesapeake Bay han demostrat que en un dia una població de l’escifomedusa Chrysaora quinquecirrha pot reduir la comunitat de copèpodes quasi en un 90%.
Els processos biològics i els processos físics
El desenvolupament d’una espècie des que s’origina fins que arriba a la maduresa sexual segueix una evolució espàcio-temporal que depèn de l’estructura i la dinàmica de les masses d’aigua en què es va trobant al llarg de la seva vida.
Un exemple podria ser el cicle biològic de l’areng (Clupea harengus) a les costes escoceses. Els adults fan la posta en una zona de la plataforma continental on les condicions de turbulència ajuden a la flotabilitat dels ous, que suren. Els ous són arrossegats cap al nord per un corrent, i quan eclosionen les larves poden disposar d’aliment, ja que es troben prop de les taques de més alta densitat de fitoplàncton. A mesura que les larves van creixent són arrossegades per la mateixa massa d’aigua que conté una rica comunitat zooplanctònica. Els copèpodes calanoides creixen al mateix temps, i donen diverses generacions que permeten a les larves de mida més gran poder-se trobar amb eixams de preses també més grans. Quan el corrent que les desplaça es desvia de la costa, les larves ja tenen la mida i la forma de juvenils i migren cap al fons tornant a l’àrea d’origen, nedant activament. Durant el seu desenvolupament, la massa d’aigua que les transporta els permet estar protegides de molts depredadors i no allunyar-se de les fonts d’aliment.
El zooplàncton de fondària
La vida a mar oberta i en profunditat és una mica diferent a tot el que s’ha explicat. La zona de l’oceà situada per sota d’un quilòmetre de profunditat representa quasi el 75% de l’espai on es desenvolupa vida a la terra. És un espai ampli, mancat de qualsevol tipus de llum, fins i tot de l’ultraviolada, i amb unes condicions de temperatura i salinitat molt semblants a escala oceànica. En aquesta zona de quietud de les masses d’aigua es desenvolupen uns tipus d’organismes poc coneguts, ja que són difícils d’atrapar amb les tècniques de mostreig tradicionals.
Estudis recents amb submarins han permès observar un sistema de vida molt més variat del que se suposava. Els organismes estan força diluïts enmig de l’oceà i es desplacen lentament, a excepció d’alguns crustacis. Els organismes gelatinosos, com els sifonòfors, les meduses, les salpes, els ctenòfors, els mol·luscs pteròpodes, són els més freqüents. Són de mida molt gran comparats amb les formes d’aigües superficials, i tenen una esperança de vida de diversos anys. Es tracta de cossos delicats i transparents, ja que no els cal protegir-se de la llum ultraviolada. Presenten flotabilitat neutra i, tot i que són carnívors, tenen la capacitat de passar força temps sense alimentar-se. En altres casos es tracta de grans nedadors, capaços de desplaçar-se força quilòmetres cada dia.
L’escassetat de recursos ha fet que desenvolupin tècniques de captura molt sofisticades. Per exemple, una colònia de sifonòfors fisonèctides no més gran de mig metre estén una complexa xarxa de fins tentacles que pot arribar a fer més de 20 m de diàmetre. Altres desenvolupen relacions interspecífiques, com els amfípodes hipèrids, que viuen associats o bé dintre els cossos gelatinosos de les meduses. Així aprofiten part de les preses que captura la medusa, a part d’alimentar-se de la mateixa medusa quan els cal. Molts d’aquests organismes gelatinosos, especialment els sifonòfors fisonèctides, que presenten gas en els pneumatòfors, formen poblacions denses, i la seva distribució a l’oceà profund coincideix amb l’anomenada capa de reflexió profunda. El fet que siguin tan comuns ha portat a parlar d’una zona de depredadors de la capa mitjana de l’aigua, estesa a través dels oceans.