Un ambient fàcil i perillós alhora
Mancant-hi els refugis que proporcionen el substrat i la cobertura, els organismes que habiten a l’ambient pelàgic es troben voltats d’aliment, però alhora estan exposats a ser menjats. Tal com s’ha dit, a més del plàncton permanent, hi ha molts ous i larves que en formen part de manera temporal; no és infreqüent que els estats larvals d’alguns predadors de mida gran siguin, al seu torn, consumits pels adults de les seves preses habituals. La major part dels organismes passen per diferents etapes en què són consumidors primaris quan s’alimenten de fitoplàncton, secundaris quan canvien la seva alimentació a zooplàncton i terciaris o de grau superior quan arriben a l’edat adulta.
Tot i que que en general hi ha més diversitat d’organismes pelàgics a les regions tropicals que a les temperades i en aquestes darreres més que a les fredes, en particular sobre les plataformes continentals, la diversitat específica dels organismes pelàgics de cada regió biogeogràfica és més petita que la del seus equivalents bentònics. La diversificació de formes està molt relacionada amb l’heterogeneïtat del medi; com més gran és el nombre de microhàbitats en un ambient, més gran és la quantitat relativa de formes que es poden desenvolupar en aquest ecosistema. La relativa homogeneïtat del medi pelàgic no es presta a la diversificació.
El fet que hi hagi poques formes possibles provoca que la selecció natural sigui innegablement més severa, ja que no es pot defugir la competència interspecífica a través de l’especialització. En canvi, l’abundància dels peixos pelàgics és notablement més gran que la de les espècies bentòniques, ja que estan relacionats íntimament amb la capa de productors primaris i secundaris més importants i perquè l’absència d’una gran diversitat afavoreix la concentració de biomassa en unes poques espècies.
Els principals grups taxonòmics representats al nivell de consumidors secundaris o terciaris a l’ambient pelàgic són els peixos, alguns crustacis (en especial eufàsids i galatèids), alguns mol·luscs (calamars i sípies), ocells i mamífers.
Les estratègies de supervivència
La major part de les estratègies que permeten als organismes bentònics d’incrementar les seves probabilitats de vida i deixar descendència (adoptar una coloració que els permeti de confondre’s amb el medi, ensorrar-se o protegir-se entre les roques, cobrir-se amb d’altres organismes que els permetin de confondre’s amb el substrat, etc.) no són possibles a l’ambient pelàgic. En aquest medi, les úniques diferències apreciables en un punt donat de la columna d’aigua a la capa eufòtica són els colors clars (amunt) i els colors obscurs (avall), matisats per reflexos i onades; l’única coloració críptica possible és la manca de color o l’anomenat “countershading”, que consisteix a tenir la superfície dorsal obscura i matisada per tal de ser confosos amb el fons quan es veuen des de dalt i la superfície ventral clara i reflectora per tal de confondre’s amb la superfície quan es veuen des de baix. Paral·lelament, aquest tipus de coloració permet fer menys notable la presència de l’organisme en compensar amb el color obscur dorsal la il·luminació que li arriba des de dalt i viceversa. La majoria dels organismes que habiten a la capa eufòtica de l’ambient pelàgic tenen una coloració semblant. Altres estratègies de defensa es basen en la velocitat de desplaçament i en l’agrupació de molts individus en taques compactes. Malgrat tot, la predació és intensa i la major part dels organismes pelàgics és devorada abans d’atènyer l’edat adulta. En aquestes condicions d’exposició a la predació, les poblacions requereixen, naturalment, estratègies que assegurin un mínim de supervivència suficient per a la reproducció; en l’actualitat només formen part de l’ecosistema pelàgic aquelles espècies que han aconseguit tenir aquest tipus de respostes a la selecció intensa.
Algunes d’aquestes respostes a nivell poblacional es basen en el creixement ràpid i en la producció de nombrosos descendents, combinades en diferents graus en espècies menors, mitjanes i algunes de més grans. Una altra estratègia específica consisteix a ser grans des del primer moment i a protegir els alevins. Tant els taurons com els cetacis disposen d’aquest conjunt d’estratègies de supervivència. Finalment, l’ambient pelàgic atrau un grup d’organismes que hi participa de manera parcial, alimentant-se dels seus pobladors permanents i reproduint-se fora d’aquest medi tan atzarós; entre els que han adoptat aquesta modalitat hi ha els ocells oceànics i els pinnípedes.
Els peixos purament pelàgics solen produir una gran quantitat d’ous; les postes de milions no són estranyes en aquest grup d’organismes. Els peixos petits creixen ràpidament i maduren al més aviat possible, per tal de poder acomplir la seva funció reproductiva abans de ser menjats: la major part dels peixos petits tenen longevitats menors als sis anys i comencen a reproduir-se abans dels tres. Els calamars i les sípies estrictament pelàgics solen viure només un any i la major part de les femelles moren després de la reproducció. Altres peixos mitjans i grans tendeixen a créixer ràpidament i a assolir mides grans, que contribueixen a reduir molt considerablement llur predació per part d’altres espècies. Algunes tonyines creixen fins a atènyer un pes mil milions de vegades superior al que tenien en sortir de l’ou. En termes generals, la transferència d’energia a l’ambient pelàgic segueix el principi ecològic que els organismes incrementen la seva mida d’adults i redueixen la seva biomassa a mesura que s’allunyen dels productors primaris a la cadena tròfica. Aquest principi, que té nombroses excepcions a qualsevol ambient, troba les més notables justament a l’ecosistema pelàgic: els organismes més grans que mai no hagin existit al planeta escurcen la cadena tròfica i s’alimenten bàsicament de zooplàncton.
Les migracions també són una característica comuna a l’ambient pelàgic; la major part de les espècies nectòniques migra d’una manera o altra. Les migracions verticals diàries es presenten en una gran proporció d’organismes, particularment entre els de les regions tropicals oceàniques. Els peixos mesopelàgics són un exemple de migració vertical de gran importància. Tot i així, les migracions horitzontals són les més espectaculars. Les distàncies que poden cobrir alguns dels organismes més grans de l’ambient pelàgic són impressionants: hi ha registres propers a les 10 000 milles (uns 18 000 km). En altres casos la precisió de la migració és sorprenent, en especial en aquells organismes que es reprodueixen exactament al mateix indret on eclosionen. Tenint en compte que es tracta d’un ambient sense cap punt de referència visible, aquesta és una gran proesa.
Migracions de peixos pelàgics
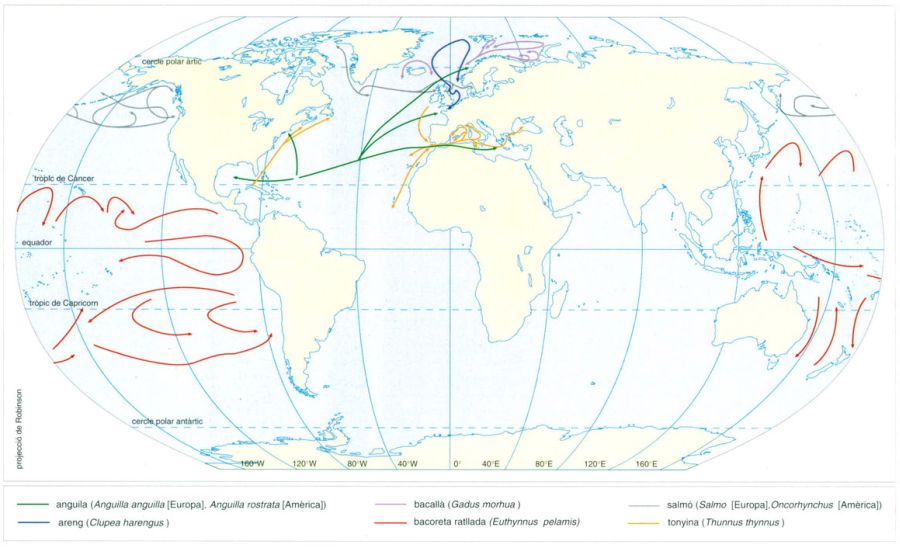
Rutes de migració d’alguns peixos pelàgics.
Editrònica, a partir de fonts diverses
En una o altra mesura, la majoria dels peixos pelàgics són migradors a grans distàncies. Hom anomena oceanòdromes les espècies que limiten les seves migracions al medi marí i diàdromes les que abasten també el medi fluvial. El mapa mostra les rutes migratòries de quatre peixos oceanòdroms, l’areng (Clupea harengus), el bacallà (Gadus morhua), la tonyina (Thunnus thynnus) i la bacoreta ratllada (Euthynnuspelamis), i dos grups de peixos diàdroms, les anguiles i els salmons. Les anguiles, tant l’europea (Anguilla anguilla) com l’americana (A. rostrata), són catàdromes, és a dir, que es reprodueixen a la mar i creixen i s’alimenten a les aigües dolces. Els salmons, per contra, tant els de l’Atlàntic (Salmo) com els del Pacífic (Oncorhynchus), són anàdroms, és a dir, es comporten a l’inrevés.
La productivitat dels sistemes pelàgics
Tal com ja s’ha dit, la distribució de la productivitat és molt poc uniforme a tot l’oceà, per més que superficialment totes les mars semblin iguals. En termes generals, les plataformes continentals són molt més riques que l’oceà obert, les mars d’altes latituds són més productives que les tropicals i les àrees de major productivitat són les zones d’aflorament. Les àrees més pobres de l’ambient pelàgic són, és clar, els centres dels grans girs, un a cada hemisferi i a cada oceà. Les més riques són aquelles en què els afloraments afavoreixen una productivitat extraordinàriament elevada. Les espècies pelàgiques són especialment abundants en aquestes regions en què la productivitat és força elevada.
L’eficiència ecològica (bàsicament la proporció d’energia que s’acumula d’un nivell tròfic al següent) també canvia molt; les zones d’aflorament són molt menys eficients que les tropicals; la producció és tan alta en aquestes que no hi ha mecanismes efectius d’aprofitar-la cabalment, i una part important es perd cap als fons marins. L’existència d’estrats abundants de restes de diatomees i de dipòsits de fosforita fa evident aquesta fuga considerable de la producció. A l’extrem oposat, el cas dels ecosistemes tropicals oceànics és totalment diferent; amb una productivitat primària molt menor, l’eficiència ecològica és molt més alta i tota la matèria orgànica s’aprofita al capdamunt de la columna d’aigua, que és molt més profunda: la producció que s’origina a la capa eufòtica és consumida, degradada i remineralitzada en els 500 m de la capa superficial. Els fons de les regions centrals dels girs oceànics pràcticament no presenten restes orgàniques. Per més que els coeficients de transferència entre nivells siguin tres vegades més grans als ecosistemes pobres, la diferència en productivitat primària entre els ecosistemes rics i els pobres sol ser força important.
Els cicles de producció
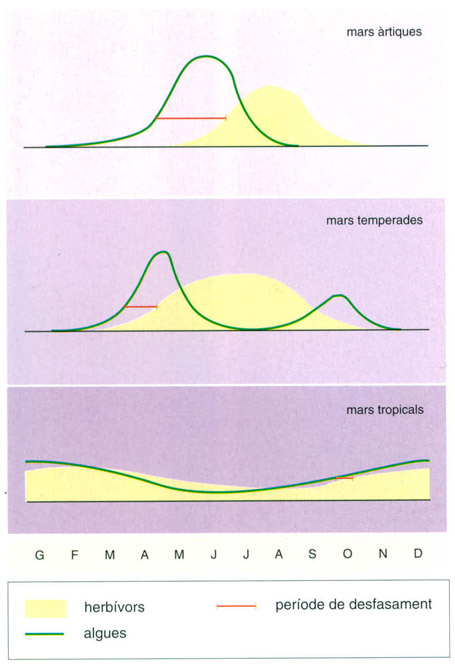
Cicles de producció anual en diferents regions oceàniques. Els cicles estacionals de producció primària i secundària tenen una estacionalitat més acusada a les latituds polars. A les altes latituds àrtiques o antàrtiques hi ha un únic pic de producció primària a l’estiu, seguit d’un pic de producció secundària, encara a l’estiu, però lleugerament desfasat. A les latituds mitjanes hi ha dos pics de producció primària (el més important a la primavera i un de secundari a la tardor) i un màxim d’estiu de la producció secundària dels herbívors. A les latituds equatorials no hi ha pics estacionals, sinó oscil·lacions molt lleugeres dins de la tendència a mantenir una producció uniforme al llarg de tot l’any, tant pel que fa als productors primaris (algues) com als herbívors.
Editrònica, a partir de dades de Cushing,1959
L’abundància i la reproducció de les espècies pelàgiques depenen no sols de la regió de l’oceà, sinó també dels cicles anuals de producció de les espècies de les quals s’alimenten. Els cicles de producció són de gran amplitud (és a dir, que hi ha una diferència important entre la productivitat mitjana mínima i la màxima al llarg de l’any) a les regions de latituds altes, on els períodes de productivitat màxima són de curta durada. Els períodes de reproducció dels organismes pelàgics en aquestes zones són molt breus; en el cas de l’areng (Clupea harengus), per exemple, només dura de dues a tres setmanes. La sincronia de la reproducció és clau, doncs, en un ambient en què la producció alta, que afavoreix una bona alimentació per a les larves, dura poc.
A les àrees temperades d’afloraments, l’alta productivitat s’estén durant un període molt més llarg i la temporada reproductiva de les espècies pot ser molt més àmplia. La màxima activitat de fresa de la sardina de Califòrnia (Sardinops caerulea, S. sagax), per exemple, pot presentar-se dins un període de tres a quatre mesos. És més, segons l’àrea de fresa de la població, pot haver-hi una sola màxima en tot l’any (per exemple, a la regió meridional de les aigües que confronten amb l’estat nord-americà de Califòrnia) o dues (per exemple a la badia Magdalena, al SW de la península de Baixa Califòrnia). Els anys freds, la fresa a la primera de les regions esmentades pot quedar restringida a unes quantes setmanes, però els anys càlids pot incloure pràcticament tots els mesos.
A les zones tropicals, la fresa se sol realitzar durant tot l’any, ja que el cicle de reproducció té una amplitud mínima al llarg dels diferents mesos. En aquest cas, la fresa pot estar restringida a una àrea dins el gran gir oceànic corrent amunt de les zones més adients per al creixement de les larves, permetent que la deriva larval porti aquests organismes fins a les zones on tenen més probabilitats de supervivència.
L’explotabilitat de l’ambient pelàgic
A mitjan segle XX, quan la pesca creixia a un ritme més gran que la població mundial, hi havia un optimisme generalitzat sobre la idea que l’oceà representava la gran reserva d’aliments per a la humanitat. Investigadors seriosos calcularen que les captures mundials podrien atènyer els 200 milions de tones anuals abans de l’any 2 000. Avui dia aquest somni sembla molt llunyà. Recentment s’ha fet un recàlcul dels potencials i s’ha considerat que un volum anual d’uns 140 milions de tones només seria possible si s’administressin de manera adequada les captures actuals i s’explotessin els recursos tradicionals encara no plenament utilitzats.
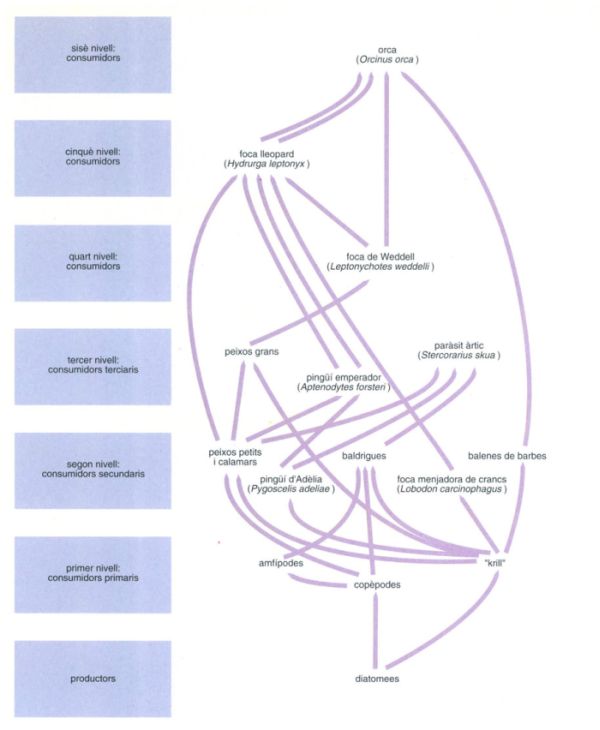
Esquema simplificat de les xarxes tròfiques de l’oceà Antàrtic, probablement un dels més complexos i amb més llargues cadenes tròfiques que es pugui descriure en un medi pelàgic. S’ha prescindit d’alguns fets que el complicarien en excés, com ara que molts cefalòpodes poden alimentar-se de peixos petits i són, per tant, consumidors terciaris, i que foques i ocells exporten part de la producció fora del medi pelàgic, i fins i tot en el cas del paràsit gros (Stercorarius skua), que migra d’una zona polar a l’altra, fora de l’hemisferi austral.
Editrònica, a partir de fonts diverses
Tot i que és evident que la potencialitat de l’ambient pelàgic és molt més gran, la possibilitat d’explotar volums més grans no és tan senzilla com semblava. Algunes espècies no són accessibles de manera econòmica amb les tecnologies actuals. Encara que es desenvolupessin tecnologies noves que donessin a la humanitat accés a aquests recursos no tradicionals, les societats humanes es trobarien davant una conjuntura incòmoda; haurien de buscar la manera d’incrementar les seves captures canviant les espècies objectiu a nivells tròfics més baixos? La necessitat de primeres matèries i d’aliments podria obligar a prendre una decisió d’aquesta mena. En efecte, la gran majoria dels peixos que consumim es troben en nivells tròfics superiors; la reorientació de la pesca tindria repercussions importants, car els humans competirien per l’aliment amb aquestes espècies. Potser arribaríem a canviar 1 kg de tonyina per 10 kg de mictòfids.
Potser, a llarg termini, seran les noves i imaginatives generacions que trobaran maneres pràctiques de trencar la paradoxa oceànica d’una capa eufòtica il·luminada, però pobra de nutrients, reposant sobre una capa rica de nutrients però sense llum.
Els consumidors pelàgics típics
Les cadenes tròfiques de l’ambient pelàgic són, en termes generals, curtes. A més, excepte en alguns casos extrems, són pobrament definides. Els peixos més petits, per exemple, són consumidors primaris i secundaris, ja que s’alimenten de fitoplàncton i de zooplàncton. Els menors d’entre la següent categoria (els anomenats pelàgics mitjans) són essencialment zooplanctòfags, per més que també consumeixen peixos juvenils. Els més grans d’aquest grup són definitivament ictiòfags que s’alimenten dels pelàgics menors.
Els calamars i les sípies contribueixen a la pobra definició de les cadenes tròfiques als ambients pelàgics: consumeixen peixos i plàncton gros i, al seu torn, serveixen d’aliment a una ampla varietat de consumidors de nivell més alt. Un cas particular de cadena tròfica es dóna a les àrees tropicals on els peixos mesopelàgics (en especial els mictòfids) s’alimenten de zooplàncton i serveixen d’aliment a les tonyines. La major part dels investigadors es refereixen a la producció de peixos zooplanctòfags i a llurs consumidors com a producció terciària; sense ser estrictament cert, és una aproximació pràctica i convenient.
Els peixos i els crustacis petits
Les larves de moltes espècies de peixos, tant pelàgiques com bentòniques, habiten al plàncton durant una part breu però important de llur existència. La seva enorme abundància té com a resultat una participació considerable en el flux energètic de l’ecosistema.
A banda aquests organismes, que normalment s’estudien com a part del zooplàncton, els peixos adults de mida petita més significatius per la seva abundància a l’ambient pelàgic es redueixen a uns pocs tipus: alguns clupeïformes (arengs, sardines i anxoves), les espècies mesopelàgiques, que pertanyen en la seva majoria als mictofiformes (peixos llanterna), i els trumfaus i espècies afins (escomberesòcids). Indubtablement, a l’ecosistema pelàgic hi ha multitud d’altres espècies de peixos petits, però la seva abundància és molt menor i la seva importància dins el flux energètic de la comunitat no és comparable.
Juntament amb alguns crustacis i amb d’altres organismes del micronècton, els peixos petits representen la baula més significativa entre la productivitat primària i secundària a la resta de la cadena tròfica dins l’ambient pelàgic. Comparats amb la resta d’espècies, són molt abundants i constitueixen les preses fonamentals dels peixos de mida més gran, tant de l’ambient pelàgic com de moltes espècies del bentònic. Part de l’explicació rau en el fet que aquests peixos petits (en particular els clupèids) poden alimentar-se en part del fitoplàncton gràcies a la seva associació simbiòtica amb bacteris intestinals que són capaços de produir cellulasa. L’escurçament de la cadena tròfica resultant d’aquesta propietat permet a aquestes poblacions assolir biomasses molt altes.
Les sardines i les anxoves
Tot i que hi ha més de 300 espècies de clupèids agrupats en 80 gèneres reconeguts a la literatura especialitzada, els dos principals, Sardinops i Engraulis (sardines i anxoves respectivament), representen més del 60% de les pesques mundials de clupeïformes, les quals, al seu torn, arriben a prop del 20% del total de les captures mundials de peix, tant marí com d’aigua dolça. Tots dos gèneres inclouen diverses espècies o poblacions de sardines i anxovetes d’aigües tèbies, que habiten a les àrees on es barregen aigües temperades i tropicals. La major part d’aquestes àrees es troben a les barreres orientals dels grans girs dels corrents oceànics on se situen les zones d’aflorament: Califòrnia, Perú-Xile, les Canàries i Benguela. De tota manera, una de les regions més importants on viuen aquests clupèids se situa al voltant del Japó, on no es presenten afloraments. En cada una d’aquestes zones principals d’abundància, les poblacions de tots dos gèneres es presenten com els peixos petits dominants del sistema. Altres gèneres de clupèids, com Sardina o Sprattus, ocupen la posició de Sardinops al sistema del corrent de Canàries i a la Mediterrània, amb abundàncies també considerables.
Una de les característiques més sorprenents d’aquests peixos és que la seva abundància varia dràsticament en temps molt curts. L’exemple més impressionant i conegut és el de l’anxoveta peruana (Engraulis ringens), les pesques de la qual es precipitaren des d’un nivell proper als 13 milions de tones anuals fins a pràcticament zero en menys d’una dècada; les poblacions de sardines (Sardinops) al Japó i a Perú-Xile, en canvi, creixeren de manera que la combinació de les seves pesques arribà als més de 10 milions de tones en menys de deu anys. L’abundància relativa d’un i altre gènere s’ha contraposat sovint als registres històrics, de manera que quan les sardines eren abundants, les anxovetes eren escasses i viceversa. Malgrat tot, és més interessant el fet que aquestes alternances es produeixen paral·lelament als diferents sistemes, allunyats entre ells per oceans de distància. La coherència en el comportament sincrònic de les poblacions de sardina i anxoveta al Japó, a Xile-Perú i a Califòrnia, i els canvis que es produeixen al mateix temps (encara que desfasats, ja que l’anxoveta és abundant quan les sardines són abundants a la resta de sistemes i viceversa) al corrent de Benguela sembla demostrar que hi ha algun factor cíclic d’abast planetari que determina la dominància d’una o altra població.
En alguns dels sistemes d’aflorament hi ha àrees de fons pràcticament anòxics que preserven especialment bé les restes orgàniques. L’anàlisi d’aquests fons ha demostrat que les escates de molts peixos es conserven en perfecte estat en estrats de deposició, els anomenats sediments laminats. Aquest tipus de sediments s’ha estudiat a les costes de l’estat de Califòrnia, a les de Baixa Califòrnia, tant a l’oceà com al golf de Califòrnia, i també a Sud-àfrica. Actualment també s’estan estudiant mostres d’altres regions. Tot i que anteriorment s’havia considerat que el factor determinant per a l’abundància relativa de les poblacions d’anxovetes i sardines era l’explotació pesquera, l’anàlisi de les escates trobades als sediments laminats de diversos sistemes de corrents ha demostrat que aquestes grans fluctuacions d’abundància ja havien estat característiques d’aquestes poblacions molt abans de ser explotades pels humans. Independentment del fet que la pesca acceleri i magnifiqui els canvis d’abundància, aquests s’han produït al passat i segurament continuaran produint-se al futur, com a reflex de canvis profunds que afecten tot l’ecosistema encara mal coneguts.
Els arengs
Els arengs (Clupea harengus) són particularment abundants a l’Atlàntic nord-oriental, on constitueixen una de les espècies més tradicionals en la pesca. Es reprodueixen prop del substrat i dipositen els seus ous als fons de grava de prop de la costa. En eclosionar, les larves s’incorporen al plàncton. La fresa d’aquesta espècie està molt estretament lligada amb les condicions ambientals; la data de la fresa màxima anual pot calcular-se per a cada població amb una variació mitjana d’una setmana. A més, s’ha demostrat que la majoria dels peixos tornen a fresar al mateix indret on ells mateixos eclosionaren, en un 80% de les vegades; la resta de peixos permeten la recombinació entre poblacions. També hi ha arengs al Pacífic septentrional, tot i que la seva abundància sembla considerablement menor que a l’Atlàntic. Els arengs són els clupèids de biomassa més important que aprofiten els períodes i les àrees d’alta productivitat a les mars temperades-fredes.
Altres clupèids
Els clupèids són costaners i canalitzen la productivitat primària i secundària de les àrees més productives, les quals es troben sobre la plataforma continental. A les regions més tropicals també hi ha clupèids, tot i que les seves biomasses són molt inferiors a les de les poblacions d’aigües temperades i temperades-fredes, de les quals procedeixen al voltant del 80% de totes les pesques de clupèids. Els gèneres Brevoortia (les “menhaden” o “lachas” de les costes del golf de Mèxic i del SE dels Estats Units), Sardinella (els seitons de l’Atlàntic, l’Índic i el Pacífic) i Opisthonema (l’“arenque de hebra” de les costes tropicals del Pacífic americà) es troben entre els més abundantment representats.
Els peixos mesopelàgics petits
En altres regions més allunyades de la costa, els principals peixos petits són espècies mesopelàgiques, oceàniques, que viuen a la capa intermèdia de les mars profundes. Normalment habiten en profunditats d’entre 200 i 1 000 m, a la capa eufòtica (epipelàgica) que queda limitada per la penetració de l’1% de la llum incident i la màxima penetració de la llum. Se solen distribuir enllà del talús que voreja cada continent i illa. Aquestes zones comprenen un 22,5% del total de la superfície oceànica del planeta.
Moltes espècies realitzen migracions verticals diàries, i pugen fins a la superfície cap al tard per a alimentar-se a les aigües més riques de zooplàncton. Algunes fins i tot arriben a les capes superficials de l’oceà durant la nit. En sortir el Sol tornen a descendir a les seves profunditats habituals. Són tan abundants que formen la major part de la capa de dispersió profunda, un registre que apareix als sondeigs a considerable profunditat i que es mou verticalment d’acord amb la sortida i la posta del Sol. Els seus ous i les seves larves formen part del meroplàncton i constitueixen la biomassa més alta de qualsevol tipus de vertebrats al plàncton oceànic. Constitueixen reserves de milions de tones no explotades.
Aquest conjunt de peixos, el més important contingent de biomassa a la zona mesopelàgica, és format per unes 700 espècies a tot el món. Són peixos d’ulls relativament grossos que els permeten de veure-hi en les condicions crepusculars de la capa mesopelàgica; molts els tenen tubulars, per concentrar l’escassa llum. Les seves mandíbules poden obrir-se de manera extrema, i els permeten capturar preses o ingerir trossos d’animals morts o cadàvers sencers d’una mida que pot arribar a ser comparable a la seva. Posseeixen nombrosos fotòfors sobre la superfície ventral, que s’ha dit que confonen els predadors que els busquen per sota contra l’escassa llum que penetra des de la superfície de la mar.
Ocasionalment s’han dut a terme algunes pesques limitades d’aquestes espècies, especialment a Sud-àfrica. La major part s’han processat per a obtenir farines i olis; aquests darrers, però, són tan abundants que no resulten fàcils d’extreure amb els procediments convencionals perquè obturen la maquinària. El consum directe d’alguns exemplars de les espècies de mida més gran no ha tingut una acceptació gaire entusiasta, principalment perquè algunes tenen una gran concentració de ceres i esters a la carn. Entre els problemes que planteja llur possible explotació hi ha les seves petites dimensions (la major part de les espècies fa entre 5 i 10 cm) i també que es troben molt dispersos (un individu per 1 000 m3), al contrari del que succeeix amb els clupèids, que formen moles molt compactes i, per tant, d’explotació rendible.
Els peixos mesopelàgics s’alimenten bàsicament de zooplàncton i constitueixen, al seu torn, l’aliment principal de nombroses espècies més grans, com ara les tonyines, els verats, els calamars, els pinnípedes i els cetacis, especialment alguns rorquals i molts dofins.
Els peixos mesopelàgics formen part de l’escala tròfica, la qual transporta activament l’energia cap al fons. Més que l’enfonsament passiu de cadàvers i productes de rebuig, que en la seva gran majoria són consumits en la mateixa capa on s’originen o en la immediatament inferior, aquesta forma de transferència és basada en la migració diària d’organismes de les capes profundes cap a la superfície per a alimentar-se, on, al seu torn serveixen d’aliment per als organismes de la capa subjacent i aquests a la de més ensota, etc. La visió original d’una capa eufòtica produint biomassa excedent que baixa lentament i és consumida pels diferents organismes de fons és difícilment compatible amb el dinàmic reciclatge dels nutrients dins de cada nivell de la llarga columna d’aigua a l’oceà profund.
Els escomberesòcids, com ara el trumfau (Scomberesox saurus) i altres espècies afins, són peixos pelàgics que quan arriben a la forma adulta fan entre 20 i 35 cm de longitud. Per la seva abundància constitueixen una baula important a la cadena tròfica, i connecten la productivitat secundària del zooplàncton amb els peixos més grans, especialment les tonyines i molts ocells. Si bé no arriben als volums dels dos grups precedents, la seva importància rau en el fet que viuen prop de la superfície i allunyats de la costa. Hi ha escomberesòcids tant al Pacífic com a l’Atlàntic. Enfront de la costa del Pacífic dels Estats Units se n’ha calculat una biomassa de prop de 500 000 tones. El Japó ha estat un dels principals pescadors d’aquestes espècies, de les quals, en anys recents, s’han registrat captures de prop de 250 000 tones. Els escomberesòcids pertanyen a l’ordre dels beloniformes, que agrupa també d’altres espècies igualment presents a l’ambient pelàgic com ara els ocellets (hemiràmfids) i els peixos voladors (exocètids), relativament importants en aigües tropicals costaneres.
A l’oceà Antàrtic hi ha dues espècies de peixos petits que ocupen un nivell tròfic equivalent: Pleurogramma antarcticum i Notolepis coatsi. Tots dos són relativament abundants, consumeixen zooplàncton i són presa d’animals més grans, el primer de pingüins i foques, el segon especialment de rorquals.
Els microcrustacis pelàgics
En el mateix nivell tròfic de consumidors de fitoplàncton i zooplàncton es troben alguns microcrustacis pelàgics. Euphausia superba, el “krill” de l’Antàrtic, se sol estudiar com un component del zooplàncton i, per tant, no serà considerat aquí. En canvi alguns galatèids pelàgics, com ara el llagostí xilè (Munida gregaria) de Xile i Nova Zelanda o la llagosteta de Mèxic (Pleuroncodes planipes), són molt abundants i de gran importància a la cadena tròfica d’algunes àrees.
Durant els seus primers anys de vida, aquests petits crancs rogencs, anomenats llagostins a Xile, tenen una fase pelàgica i més endavant es transformen en bentònics. Durant la seva etapa pelàgica formen concentracions extraordinàriament abundants, i en ocasions arriben a embarrancar a les platges en quantitats apreciables. Tenen la propietat d’alimentar-se no sols de zooplàncton, sinó també de fitoplàncton; aquesta és la raó per la qual són equivalents als clupèids en el sentit de no ser canal de la productivitat cap a nivells tròfics superiors. El llagostí xilè s’ha explotat comercialment a Xile, on es processa per a obtenir les relativament petites cues, que s’exporten a bon preu.
Les llagostetes de Mèxic són un aliment preferent de les tonyines durant la part de la seva migració anual en què penetren al corrent de Califòrnia, i arriben a constituir fins a un 85% de l’aliment que consumeixen en aquesta àrea. També són dels pocs organismes que consumeix la balena grisa (Eschrichtius robustus) durant la seva estada a les aigües temperades-càlides de la Baixa Califòrnia, a les llacunes costaneres on neixen les seves cries durant l’estació hivernal.
Els peixos mitjans
Els peixos mitjans constitueixen la categoria menys definida de tota la cadena tròfica en l’ambient pelàgic. N’hi ha que tenen una mida amb prou feines més llarga que la dels peixos petits, però la seva alimentació inclou zooplàncton, micronècton, juvenils d’altres peixos i també calamars. Els gèneres de peixos mitjans més importants per la seva abundància pertanyen a la família dels escòmbrids, com els verats (Scomber) i les “caritas” (Scomberomorus), o a la dels caràngids, com els sorells (Trachurus) i les sèrvies (Seriola).
Altres gèneres importants que no pertanyen a cap d’aquelles dues famílies són Mallotus, de la família dels osmèrids, i Micromesistius, al qual pertany la maire (M. poutassou), de la dels gàdids. Micromesistius, una excepció entre els gadiformes, normalment d’hàbits bentònics, és principalment pelàgic, però tendeix a formar capes extenses més que moles denses. El “capelín” (Mallotus villosus) és un osmèrid principalment àrtic, que es distribueix pel N del front polar, incloent-hi les costes del Canadà i de Noruega i la mar de Barentsz. Es pesca al N i al SE de l’Atlàntic. Les pesques de cada un d’aquests dos gèneres han excedit les 100 000 tones anuals, valors que corresponen a les seves abundàncies. Molts altres gèneres d’aquestes dues darreres famílies tenen representants a l’ambient pelàgic, especialment sobre les plataformes continentals de les mars tropicals.
Una altra família que participa de manera especial dins aquesta categoria és la dels esfirènids, les barracudes (Sphyraena), depredadors de mida mitjana que s’alimenten bàsicament de peixos petits i de calamars.
Els peixos batipelàgics
La subdivisió batipelàgica (compresa entre els 1 000 m, la màxima penetració de la llum, i els 6 000 m) i l’abissal (que s’estén fins als 10 km) constitueixen la gran majoria de l’ambient pelàgic. L’ecosistema d’aquestes mars profundes és quasi totalment heterotròfic, dominat pel catabolisme. És al·lòcton, en el sentit que l’energia necessària per a la seva existència ha de venir de fora.
A més de ser un sistema obscur, la major part de l’aigua és freda i les variacions estacionals de temperatura són quasi imperceptibles. Les característiques físiques i químiques de l’aigua d’aquestes capes reflecteixen aquelles de les altes latituds on es formaren. Es necessita molt de temps perquè l’aigua originada a les àrees circumpolars es distribueixi cap a l’equador als fons marins. L’aigua profunda de l’Atlàntic, per exemple, té probablement una antiguitat de 200 a 300 anys, mentre que la del Pacífic segurament en té uns 1 000. Les variacions de l’ambient dels oceans profunds només té sentit a escala geològica.
Així com a la superfície de la mar els afloraments tenen una enorme importància perquè eleven aigua rica de nutrients cap a la capa eufòtica, a les mars profundes els fenòmens que semblen ser més rellevants són els corrents de terbolesa; cascades gegantines que arrosseguen sediments cap als fons pobres, portant amb ells restes orgàniques i ensorrant part de la fauna profunda.
S’ha demostrat que l’abundància d’organismes disminueix exponencialment (però no regularment) d’acord amb la profunditat. Hi ha una tendència marcada a la reducció de la densitat de l’organisme segons la profunditat, amb el probable resultat d’incrementar l’eficiència metabòlica i de conservar energia, en disminuir la necessitat de mantenir-se surant en un nivell determinat. El contingut d’aigua en els peixos s’incrementa amb la profunditat, comparant amb proteïnes, hidrats de carboni, lípids i esquelet.
Els peixos batipelàgics tenen les modificacions més profundes de tots els peixos que habiten en les diferents profunditats. Tenen grans mandíbules i dents, i molts posseeixen barbetes per a atreure les preses. Els ceratioideus, per exemple, tenen un apèndix amb fotòfors al front. En general els seus ulls són molt petits. Moltes femelles semblen tenir la propietat d’emetre feromones per atreure els mascles, que al seu torn posseeixen un olfacte molt fi; això facilita la unió dels sexes en un món sense llum. El dimorfisme sexual és comú i arriba a situacions extremes: els mascles de Ceratias són molt més petits que les femelles; quan les localitzen s’hi fermen i degeneren gradualment fins a transformar-se en un mer sàcul amb els òrgans reproductors. Els peixos batipelàgics sofreixen transformacions fins i tot més profundes que els bentopelàgics, molt més relacionats amb el fons. Aquests adopten formes molt més familiars en apariència, com passa amb les quimeres (quimèrids), els titolots i altres macrúrids, algunes rajades, etc.
Els calamars i les sípies
Els cefalòpodes, en especial els calamars (Loligo, Todarodes, Alloteuthis, Abralia, Illex, etc.) i les sípies (Sepia) constitueixen un component molt important dels ecosistemes pelàgics, per més que només una part mínima de les seves poblacions és explotada pels humans. La gran majoria de la pesca del calamars es realitza en aigües de profunditat inferior als 200 m, i aquestes representen una part mínima del total de l’oceà. Els calamars de mida més gran són comuns a profunditats més importants que l’abans esmentada.
L’abundància de les espècies de calamars de mida més gran (Architeuthis) s’ha estimat parcialment a través de l’anàlisi de les seves restes (les mandíbules especialment) als estómacs dels catxalots. S’ha calculat que només aquests grans mamífers consumeixen anualment uns 100 milions de tones de calamars; és a dir, més que la captura pesquera total a tot el món. A l’Antàrtic, on s’ha estudiat la depredació de calamars per part dels cetacis, s’ha calculat que el consum de calamars només per part dels catxalots representa al voltant del 35% del total consumit per diversos depredadors.
Els calamars pelàgics s’alimenten principalment de peixos petits, per més que també poden consumir alguns crustacis, inclosos copèpodes i galatèids, així com pteròpodes i altres calamars. La conversió d’aliment (l’eficiència alimentària) dels calamars es troba entre les més altes enregistrades: al voltant del 50% del pes fresc consumit s’incorpora al creixement de l’organisme.
Els peixos grans
El subordre escombroïdeus inclou peixos mitjans i grans, depredadors actius d’alt nivell tròfic: tonyines (Thunnus) i bonítols (Sarda), emperadors (Luvarus) i verats (Scomber), sabres (Trichiurus), marlins (Tetrapturus) i peixos espasa (Xiphias). Tots ells estan adaptats al moviment permanent i a la velocitat; el peix vela (Istiophorus platypterus) ha estat cronometrat a velocitats superiors als 110 km/h. Són els més hidrodinàmics de tots els peixos, fins al punt que les seves aletes s’arrepleguen en canaladures especials i que els ulls formen una superfície llisa amb la resta del cap. No poden deixar de nedar per dues raons: la primera és que depenen de l’oxigen que transporta el corrent d’aigua que passa a través de la seva boca permanentment oberta, ja que han perdut la major part dels mecanismes que permeten a altres peixos de bombar aigua a les brànquies; la segona és que, si no neden, s’enfonsen, perquè pesen més que l’aigua, en part pel fet que no tenen bufeta natatòria.
El seu moviment permanent requereix una despesa energètica considerable; algunes espècies han de consumir fins a un 25% del seu propi pes cada dia. Aquest intens metabolisme genera una gran quantitat de calor que fa de les tonyines en particular animals de sang calenta. Els mecanismes fisiològics que posseeixen per dissipar la calor excessiva són únics i inclouen una intrincada xarxa de fins vasos sanguinis: l’anomenada rete mirabilis. Com a predadors eficients, tenen l’oïda molt fina, detectors químics sensibles i una visió estereoscòpica que els permet de mesurar distàncies. També són molt fecunds: les femelles alliberen uns 100 000 ous per quilogram de pes; així, una femella de 50 kg produeix uns 5 000 000 d’ous petits. Originada en un ou de tot just un mil·límetre de diàmetre, la tonyina comuna (Thunnus thynnus) multiplica el seu pes uns mil milions de vegades fins a assolir un pes superior als 700 kg. L’agulla negra (Makaira ampla) arriba a pesar més de 1 300 kg; sens dubte, el peix ossi més gran de totes les mars. Són notablement migratoris; la bacora (Thunnus alalunga) migra des de la costa de Califòrnia fins al Japó, uns 8 500 km, i s’han enregistrat distàncies més grans per a altres espècies del mateix grup.
Altres peixos grans són les llampugues o “mahi-mahi” (Coryphaena) i el peix gall (Nematistius), molt coneguts però comparativament poc abundants. Un grup particularment important a l’ambient pelàgic i en especial a l’oceà obert són els taurons; no hi ha dubte que es troben entre els depredadors més importants de tot l’oceà.
Els consumidors pelàgics al·lòctons
Peixos i crustacis pelàgics són animals pròpiament marins. Però a les cadenes alimentàries de les aigües marines participen també animals típicament continentals, com ara ocells, rèptils i mamífers carnívors, especialitzats a explotar els estocs íctics. La gamma d’estratègies abastada per aquests grups és molt gran, i va des de la pesca costanera fins a la captura en alta mar, des de la vida continental amb incursions oceàniques a l’adequació permanent a les condicions marines. No cal sinó pensar en gavines i en petrells, en llúdries de mar o en cetacis.
Les tortugues marines

Tortugues de les mars del món. La tortuga babaua (Caretta caretta) viu a les zones tropicals de tots els oceans, menja mol·luscs i crancs i la seva longitud d’adult és d’entre 96 fins a 114 cm. La tortuga verda del Pacífic oriental (Chelonia agassizii) viu a les mars de Mèxic, Perú i les illes Galápagos; menja algues i pot mesurar de 71 a 91 cm de llargària. No tots els zoòlegs estan d’acord en separar les dues espècies de tortugues verdes. La tortuga plana d’Austràlia (Chelonia depressa) viu a les mars del nord d’Austràlia, menja holotúries i altres invertebrats, i pot assolir els 90 cm de longitud. La tortuga llaüt (Dermochelys coriacea) cria als tròpics però també se la pot trobar en aigües temperades i, fins i tot, en aigües subàrtiques; menja meduses i pot assolir les llargàries més grans d’entre els rèptils marins: entre 152 i 178 cm. A la tortuga golfina (Lepidochelys kempi) se la pot trobar al golf de Mèxic i a l’Atlàntic nord; menja crancs i mol·luscs i mesura de 58 a 66 cm. La tortuga carei (Eretmochelys imbricata), tan explotada per la seva closca, viu en oceans tropicals prop d’esculls coral·lins o rocosos i pot mesurar de 28 a 36 cm. La tortuga verda (Chelonia mydas) viu a tots els oceans tropicals, excepte el Pacífic est; menja algues i fanerògames marines i mesura de 90 a 110 cm. I, per últim, la tortuga bastarda (Lepidochelis olivacea) viu als oceans tropicals, principalment al Pacífic est, l’Índic i el sud de l’Atlàntic; menja crustacis, ous de peixos i vegetació. La seva llargària pot fer de 58 a 66 cm.
Anna Maria Ferrer
Les tortugues marines pertanyen a dues famílies: la dels dermoquèlids, amb un sol gènere (Dermochelys), i la dels quelònids, amb 4 gèneres (Chelonia, Caretta, Lepidochelys, Eretmochelys). La tortuga llaüt (Dermochelys coriacea), un dels rèptils més grans que existeixen avui dia, pertany al primer grup. Els adults es diferencien fàcilment per la seva closca coriàcia, sense escates. És la de distribució més extensa i habita en aigües més fredes que les de la resta de tortugues marines. Nia a la tardor i a l’hivern i es desplaça en grups molt nombrosos. Les principals platges on nien es troben a la costa occidental de Mèxic (80 000 nius l’any), a les Guaianes (10 000 a 15 000), a Costa Rica (5 000) i a Trinitat i Tobago (1 000); les costes de Malàisia també havien estat molt importants, tot i que actualment el nombre de zones d’arribada ha caigut molt. A la resta de les costes tropicals els nombres de nius són molt inferiors. L’altra família, la dels quelònids, reuneix espècies importants, com la tortuga babaua (Caretta caretta), la tortuga verda (Chelonia mydas), la tortuga bastarda (Lepidochelys olivacea), la tortuga golfina (L. kempi), la tortuga plana d’Austràlia (Chelonia depressa) i la famosa tortuga carei (Eretmochelys imbricata), que havia estat explotada per la seva closca. Aquesta darrera és la més tropical de les tortugues i es distribueix a l’Atlàntic central i a l’Indo-Pacífic; nia a la primavera i a l’estiu.
Els mamífers marins
Els mamífers marins inclouen els consumidors més grans no sols de les mars, sinó de tot el planeta, i no sols dels actuals sinó també de tots aquells dels quals es té coneixement. La gran majoria dels mamífers marins es distribueixen a les àrees circumpolars o a les zones temperades d’alta productivitat, per afloraments o per mescla d’aigües. En qualsevol dels casos, és evident que aquests organismes s’han desenvolupat de manera especial a les àrees que tenen riquesa i disponibilitat d’aliment. Un grup de mamífers, els pinnípedes, utilitza l’ambient pelàgic de manera semblant a com ho fan els ocells: s’alimenta dins seu, però defuig els seus aspectes més perillosos sortint a terra a l’hora de reproduir-se.
No tots els mamífers marins s’alimenten fonamentalment d’organismes pelàgics. Ben al contrari, molts d’ells són consumidors preferencials d’espècies de fons, incloent-hi diverses foques (per exemple Erignathus), les morses (Odobenus), alguns dofins com ara el narval (Monodon) i fins i tot una balena, la grisa (Eschrichtius). En termes generals, entre els mamífers marins que habiten l’Àrtic, hi ha moltes espècies que s’alimenten d’organismes del fons a causa de l’amplitud de les plataformes continentals d’aquella regió; tot i així, les espècies que habiten a l’Antàrtic són pràcticament en la seva totalitat dependents de l’ambient pelàgic per a la seva alimentació, ja que no hi ha pràcticament disponibilitat de fons soms.
Els dofins són consumidors actius d’organismes pelàgics, especialment de peixos petits i de calamars. Hi ha nombroses espècies de dofins i es distribueixen a pràcticament totes les mars, encara que la seva abundància és considerablement més gran a les zones d’alta productivitat. Els organismes més grans de tots, les balenes franques (família dels balènids), han escurçat la cadena tròfica alimentant-se essencialment de zooplàncton i, en alguns casos, d’animals pelàgics menors. L’oceà Antàrtic i la mar Àrtica són les àrees d’alimentació més importants d’aquests colossos durant l’estiu. L’Antàrtic, en especial, té quatre vegades més balenes que tot l’hemisferi nord. L’atractiu especial d’aquesta regió es deu a l’enorme abundància de zooplàncton i micronècton que creix amb rapidesa extraordinària i forma enormes taques d’alta densitat.
Entre les balenes que obtenen el seu aliment a l’ambient pelàgic, les formes d’alimentació són dues: la primera és característica de les balenes d’aleta (família dels balenoptèrids), i consisteix a prendre una gran glopada d’aigua (fins a 60 m3 en alguns casos) i filtrar-la a través de les barbes, les quals retenen el zooplàncton. L’altra, característica de les balenes franques (família dels balènids), consisteix a nedar prop de la superfície amb la boca oberta i les barbes tancades, de manera molt semblant al funcionament d’una xarxa. Totes dues maneres requereixen concentracions denses d’aliment per a compensar la considerable despesa energètica i deixar un romanent suficient no sols per a la despesa metabòlica normal, sinó també per al creixement i l’acumulació de greix.
Els ocells marins
Els ocells marins ocupen una posició importantíssima entre els depredadors de l’ambient pelàgic. En alguns indrets, el guano dipositat per aquests animals a les illes de la costa ha estat font considerable de fertilitzants i explotat intensament, en especial en àrees d’aflorament com ara el corrent de Humboldt o el de Benguela.
Hi ha nombroses espècies d’ocells que s’alimenten essencialment a l’ambient pelàgic; gavines, cormorans, pelicans i pingüins. Tot i que moltes s’alimenten d’organismes bentònics com ara els crancs i els mol·luscs, en termes de volum d’aliment no hi ha dubte que els pelàgics petits són la font més important del seu manteniment. De fet, una part important dels efectes d’El Niño (el fenomen d’escalfament d’origen equatorià al Pacífic) es reflecteix en l’àmplia mortalitat d’ocells marins, ja que les moles del seu aliment principal, l’anxoveta, es distribueixen a més profunditat de la normal i no els són accessibles.
Rutes migratòries d’alguns ocells marins
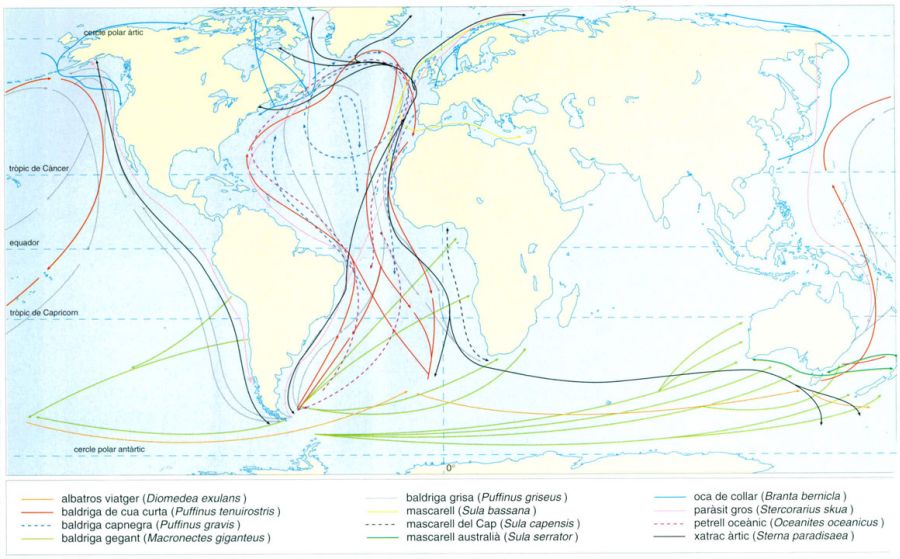
Editrònica, a partir de fonts diverses
A desgrat que tots els ocells marins crien a terra i molts romanen de manera permanent en un territori relativament restringit, n’hi ha que fan migracions de milers de quilòmetres entre les seves àrees de nidificació i les d’hivernada. El xatrac àrtic (Sterna paradisaea), les baldrigues (Puffinus) i els paràsits (Stercorarius) pràcticament arriben, en les seves migracions, de pol a pol. Altres, com el petrell oceànic (Oceanites oceanicus), limiten els seus desplaçaments a les regions temperades d’un i altre hemisferi. Altres encara, com l’albatros viatger (Diomedea exulans) i el fulmar gegant (Macronectes giganteus) aprofiten els vents dominants per a fer grans desplaçaments, en el sentit dels paral·lels, als oceans australs. Els mascarells (Sula) i l’oca de collar (Branta bernicla), finalment, acostumen a fer desplaçaments latitudinals, però sempre dins d’un mateix hemisferi o, en el cas del mascarell del Cap (Sula capensis), sense ultrapassar gaire la línia equatorial.
El grau d’adaptació a l’ambient pelàgic es reflecteix en gran mesura en la forma d’alimentació de cada grup d’ocells. L’espectre de variació és molt ampli; hi ha espècies que pertanyen a grups taxonòmics netament terrestres, per exemple l’àguila pescadora (Halichoerus), la qual obté les seves preses durant el seu vol rasant sobre la superfície, i també n’hi ha d’altres que, tot i estar plenament identificades amb l’ambient marí, no se submergeixen ni neden, sinó que aconsegueixen el seu aliment arrabassant-lo a d’altres ocells que els capturen directament, com ara les fragates (Fregata). Les gavines (Larus) i els pelicans (Pelecanus) obtenen el seu aliment nedant a la superfície o penetrant dins l’aigua a escassa profunditat; només aquelles espècies que solen nedar molt superficialment són al seu abast. Les primeres, en especial, a més de peixos consumeixen quantitats considerables d’altres organismes de fons. Els corbs marins (Phalacrocorax) estan molt més adaptats a cabussar-se per a caçar les seves preses i poden penetrar a considerable profunditat perseguint-les; tot i així, no hi ha dubte que l’extrem de l’adaptació a la cabussada marina són els pelicans (Pelecanus).
Si bé la distribució mundial dels dipòsits de guano revela que les àrees d’aflorament són les més importants pel que fa a biomasses d’aus que s’alimenten de peixos i nien a terra, és indubtable que l’Antàrtic, amb la seva rica fauna de pingüins, és la regió on és més evident la participació d’aquests organismes a les cadenes tròfiques de l’ambient pelàgic (i per això són tractades al volum 9 d’aquesta mateixa obra). La biomassa de les set espècies de pingüins que viuen a l’Antàrtic s’ha calculat en unes 500 000 tones, mentre que la resta d’ocells marins a penes arriben a 50 000 tones. El consum de peixos petits, “krill” i calamars que consumeix aquesta quantitat d’ocells s’ha calculat en uns 30 milions de tones anuals, més de la tercera part de les captures pesqueres mundials.