
Planta jove d’angiosperma monocotiledònia (A) i seccions esquemàtiques longitudinals de la gemma apical (A’) i transversal de la tija (A’’), que mostra una estructura primària. 1 Con vegetatiu, 2 primordis foliars, 3 primordis de les gemmes axil·lars, 4 feixos conductors, 5 epidermis, 6 còrtex primari, 7 medul·la, 8 floema, 9 xilema.
Mercè Cartañá, a partir de fonts diverses
En contraposició a la rel, la tija és un òrgan habitualment aeri que creix cap a la llum. S’encarrega de suportar les fulles i les flors (que poden distribuir-se, d’aquesta manera, en l’espai) i esdevé el veritable eix del corm, a través del qual té lloc el transport de nutrients entre rel i fulles. No són rares, tanmateix, les tiges subterrànies ni les que acumulen productes de reserva.
Els espermatòfits que viuen només un any o dos tenen les tiges tendres i verdes. Aquest és el cas de la civada (Avena sativa), entre les plantes anuals, i del cascall marí (Glaucium flavum), entre les biennals. Algunes de les plantes que viuen molts anys tenen també les tiges aèries anuals i de consistència herbàcia, però en aquest cas disposen d’òrgans subterranis pluriennals que cada any, generalment amb l’arribada del bon temps, renoven els brots aeris. Entre les liliàcies (alls i cebes, del gènere Allium), les ranunculàcies (el buixol, Anemone nemorosa), etc., trobem bons exemples d’aquestes plantes, anomenades vivaces. Però habitualment els espermatòfits de vida llarga tenen les tiges llenyoses i longeves; són les plantes perennes, tan freqüents a la nostra flora. La durada de la tija a les plantes superiors va des d’uns quants dies, com passa a certes herbes dels deserts que apareixen amb les pluges, fins a més de 4500 anys, com a la conífera americana Pinus aristata, de Califòrnia.
Morfologia i estructura
L’allargament de la tija és possible gràcies a la gemma apical situada al seu àpex, on són produïdes les cèl·lules que, a la llarga, donaran lloc a noves branques i fulles. La gemma consta d’un con vegetatiu de caràcters meristemàtic que es protegeix amb alguns dels resultats més recents i tendres de la seva activitat, els primordis foliars. Aquestes fulles joveníssimes, encara en formació, ultrapassen el meristema i l’embolcallen perquè creixen més per la cara inferior que no pas per la superior. La gemma no és altra cosa, doncs, que un lluc condensat on comencen a diferenciar-se la tija amb les seves fulles, i que conté ja els primordis de gemmes que hi ha a l’axil·la d’aquests. Tanmateix, és un òrgan tendríssim i delicat, particularment sensible a les condicions climàtiques extremes, perquè els primorfis foliars, com el con vegetatiu, són formats per cèl·lules de paret molt fina. Per aquesta raó, quan arriba l’hivern, els arbres i els arbusts la protegeixen mitjançant unes fulles especials, els catafil·les, que actuen com un hivernacle aïllant. Aquestes gemmes de resistència són els borrons, que ja no brostaran fins a l’arribada del bon temps.
La tija s’estira per dessota de la gemma, mentre els primordis foliars se separen i es converteixen en fulles. Només surten fulles d’unes zones determinades de la tija, els nusos, separats per espais sense fulles anomenats entrenusos. De vegades, algunes branques tenen creixement limitat, com passa a les rames que porten els fascicles de fulles als cedres (Cedrus) i es parla de braquiblasts. Les branques normals, que creixen sense límits predeterminats, reben el nom de macroblasts. L’estirament dels entrenusos, d’altra banda, no sempre és prou clar. Les plantes anomenades acaules semblen mancades de tija perquè els entrensos són tan curts que les fulles surten totes juntes fent una roseta. En són exemples alguns plantatges (Plantago) i l’atzavara (Agave americana), entre d’altres. En canvi, hi ha plantes de tiges llarguíssimes, com ara alguns eucaliptus (Eucalyptus), que son els arbres més alts, i les palmeres enfiladisses del gènre Calamus, que viuen a l’Índia i poden arribar als 300 m.
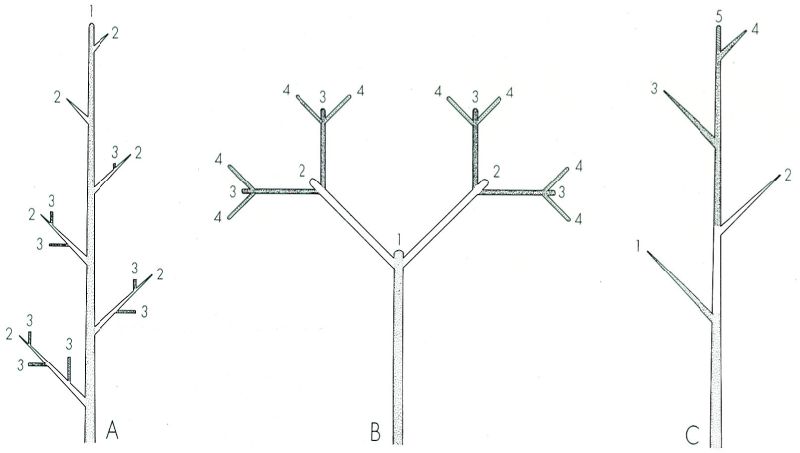
Esquemes de ramificació monopòdica (A) i simpòdica (B, en dicasi, C en monocasi). Els números indiquen l’ordre d’aparició de les rames. Convé remarcar la similitud que pot arribar a haver-hi entre un monopodi (A) i un monocasi (C), no obstant les diferències en els processos respectius de ramificació.
Mercè Cartañá, a partir de Nultsch, 1975
Per bé que algunes plantes fan només una tija senzilla, la majoria dels espermatòfits es ramifiquen, és a dir, tenen una tija principal de la qual s’originen tiges secundàries. Aquesta ramificació no és endògena com a la rel, sinó que resulta del desenvolupament de les gemmes axil·lars). Les branques que es formen s’anomenen branques secundàries o branques laterals, i les fulles de l’axil·la de les quals arrenquen són les fulles tectrius); les branques secundàries poden ramificar-se posteriorment i donar lloc a les terciàries, aquestes a les quaternàries, i així successivament. Si, com en el cas de l’avet (Abies alba), la tija principal creix indefinidament per efecte de l’activitat d’una única gemma apical que resta funcional any rere any, es parla de ramificació racemosa), o monopòdica); les branques laterals creixen en aquest cas menys que la tija principal i, encara, les branques velles, basals, són més llargues que les joves, situades a les zones altes. Més sovint, en l’anomenada ramificació simpòdica), la gemma apical actua només durant un any. Arribat el següent període vegetatiu, és substituïda en la seva funció per una o més gemmes axil·lars, precisament les més properes a l’àpex, les quals seran novament reemplaçades al cap d’un any. Aquest relleu de protagonisme comporta petits canvis direccionals que, en el cas dels arbres, donen com a resultat un eix principal que fa ziga-zagues més o menys acusades.
Les cèl·lules que resulten de l’activitat del punt vegetatiu es diferencien en allunyar-se de les gemmes i s’organitzen de manera aproximadament semblant (per bé que amb diferències significatives entre monocotiledònies i dicotiledònies) a totes les tiges que es troben en el seu primer any de vida. Llur estructura, anomenada primària, consta essencialment d’una epidermis cutinitzada al dessota de la qual apareix un parènquima clorofíl·lic que encercla, a la zona central, els vasos conductors i la medul·la. A partir del segon any, però, la majoria de les tiges perennes incrementen el seu gruix i modifiquen la seva constitució. La nova estructura, anomenada secundària, comporta una suberificació de les capes cel·lules externes i una lignificació de les més internes, formades bàsicament pel xilema vell. El creixement en gruix és degut principalment a l’activitat d’un meristema de neoformació, situat entre floema i xilema, que cada any origina noves capes de cèl·lules, sobretot conductores, a un costat i a l’altre. Precisament els anells concèntrics o anells de creixement que s’observen a molts arbres tallats corresponen, essencialment, a la producció anual de xilema, que comença a la primavera amb vasos prims de llum ampla per tal d’afavorir la circulació de la saba i acaba amb vasos més estrets i gruixuts, tardorals, que se situen a la perifèria dels anteriors. Tot plegat permet de diferenciar tres grans zones a bona part dels troncs vells: l’escorça, suberificada, a l’exterior; l’albeca, blanquinosa i tendra, situada immediatament cap endins i formada pels teixits conductors actius; el cor o duramen, central, dur i de color bru, constituït per elements inaactius impregnats de tanins, resines, etc. La fusta del duramen és molt apreciada industrialment per la seva duresa i perquè difícilment es podreix. No obstant això, els arbres anomenats de fusta blanca no fan duramen; aquest és el cas dels pollancres (Populus), dels til·lers o tells (Tilia) i dels aurons (Acer), entre altres.
Només una part de les gemmes axil·lars de les plantes perennes arriben a brostar, de manera que les plantes llenyoses tenen menys branques que les que podrien arribar a produir. L’explicació d’aquest fet és senzilla: si totes les gemmes brostessin, les branques corresponents es farien nosa i, a més, la densitat excessiva de fulles rebaixaria l’efectivitat fotosintetitzadora de cadascuna d’elles. Aquest mecanisme d’autocontrol, que tendeix a estalviar la producció de materials innecessaris, no arriba a anul·lar la capacitat meristemàtica de les gemmes que no es desenvolupen, les quals resten en estat dorment fins que és necessària la seva reactivació. Així s’explica l’aparició de tanys als calcinals, a les soques velles i als arbres viaris dels nostres carrers i jardins, periòdicament decapitats per podes rigorosíssimes. La brostada de les gemmes no és un fet a l’atzar, sinó que en cada cas n’afecta unes de concretes, fins al punt de determinar el port o hàbit de la planta: el port arbori, per exemple, s’explica perquè en els arbres acostumen a brostar cada any només les gemmes situades a l’extrem de les branques; en els arbusts, en canvi, es veuen afavorides les gemmes de la base de la tija. En el primer pas, la planta sol presentar una tija principal o tronc que es ramifica a una certa alçada, mentre que els arbusts es ramifiquen pràcticament des de terra.
De manera general, els vegetals de tija lignificada es poden separar en tres grups de límits poc precisos, segons el seu port i les seves dimensions. Les mates o subarbusts, els arbusts i els arbres. Aquests últims fan més de cinc metres i solen tenir un únic tronc que comença a ramificar-se a una certa alçada; els roures i les alzines (Quercus), els pins (Pinus), etc. en són bons exemplars. Els arbusts, com el boix (Buxus sempervirens) i el baladre (Nerium oleander), que fan entre un i cinc metres, són també abundants a la nostra flora. Les mates són arbusts petits, com el romaní (Rosmarinus officinalis), el timó (Thymus vulgaris) o el mateix eriçó (Erinacea anthyllis), que no solen superar el metre d’alçada.
La presència de pèls o tricotes a la superfície de les tiges i, en general, dels òrgans aeris de les plantes superiors es relaciona habitualment amb la seva economia hídrica. Els tricomes són prolongacions de l’epidermis dels vegetals formades per una o més cèl·lules, vives o mortes; de vegades es tracta de pèls simples, però en certs casos són ramificats, i en altres posseeixen cèl·lules secretores (pèls glandulars). Els pèls vius serveixen per a facilitar la circulació de l’aigua perquè incrementen la superfície de transpiració, però a les nostres plantes és més freqüent que els pèls siguin morts, i en aquest cas s’utilitzen per a limitar les pèrdues d’aigua. La quantitat i la qualitat dels pèls que cobreixen els òrgans vegetals s’indiquen mitjançant un vocabulari específic, del qual comentem seguidament els mots més usats: glabre vol dir sense pèls, en contraposició a pilós; parlem de pubescent quan els pèls, poc densos, són tous, curts i suaus; vellutat si hi ha pèls molt junts que donen un tacte com el del vellut; sedós significa que els pèls, molt fins i lluents, semblen de seda; hirsut s’aplica si els pèls són drets i rígids; híspid vol dir que els tricotes són gairebé punxents; quan són molt atapeïts i curts com formant borra, cal parlar de tomentós.
Modificacions estructurals
L’estructura i la morfologia d’algunes tiges poden separar-se manifestament dels esquemes fins ara descrits. De vegades són petites variacions, com a les tiges fistuloses del cebollí (Asphodelus fistulosus), amb l’eix buit en desaparèixer la medul·la central. O bé com a les tiges en canya, pròpies de les gramínies, que són com tiges fistuloses de nusos plens. I, encara, les tiges prostrades o ajagudes, que creixen aplicades a terra sense enlairar-se, com en el cas de la verdolaga (Portulaca oleracea), en contraposició a les plantes erectes, de tiges dreçades. Però ben soviet les modificacions de la tija relacionades amb adaptacions a determinats ambients i gèneres de vida peculiars són tan profundes que reben el nom de metamorfosis. En són exemples els bulbs, els estolons, els rizomes, etc., dels quals parlarem seguidament.
Sovintegen les tiges subterrànies carregades de substàncies de reserva, de vegades asociades a fulles també modificades. És el cas dels rizomes, dels tubercles caulinars i dels bulbs, característics de les plantes vivaces, que solen tenir gran importància en la multiplicació vegetativa. Els rizomes són tiges de llargada i de gruix variables, que creixen horitzontalment a profunditats diverses segons les espècies. Als nusos porten fulles menudes, del tipus catafil·le, i cada any produeixen rels que s’endinsen en el sòl i tiges aèries de vida curta. El gram (Cynodon dactylon), com moltes altres gramínies, i el lliri blau (Iris germanica) són exemples de plantes amb rizoma. Si el rizoma produeix tiges aèries molt atançades diem que la planta és cespitosa. Les gespes no són altra cosa que plantes cespitoses esteses per superficies amplies. Els tubercles caulinars, més gruixuts que els rizomes, se’n diferencien també perquè tenen creixement limitat, no presenten habitualment rels i solen durar només un sol període vegetatiu. Molts dels tubercles són utilitzats en la nutrició humana, entre els quals els de la patatera (Solanum tuberosum), i es fan servir per a la multiplicació de la mateixa planta: n’hi ha prou amb un bocí de patata (que, això sí, ha de tenir un ull o gemma com a mínim) per a sembrar-la. Els bulbs són òrgans en la formació dels quals intervenen una tija ampla, generalment molt curta, i diverses fulles que l’embolcallen: són, doncs, gemmes subterrànies especialitzades. Els bulbs de les tulipes (Tulipa), del marcòlic (Lilium martagon), etc., porten fulles del tipus catafil·le, mentre que els dels alls i les cebes (Allium), per exemple, queden envoltats per les beines tancades i engruixides de fulles que ja no són funcionals. En el primer cas es parla de bulbs esquamosos i, en el segon, de bulbs tunicats. Tant els uns com els altres produeixen petits bulbs que, en descompondre’s el bulb vell, donaran lloc a nous individus. Els bulbs són particularment freqüents a les liliàcies i famílies properes.
Les tiges aèries metamorfosades són molt diverses. Els estolons són brots llargs, originals a la base de les tiges, que arrelen als nusos. Com que en els nusos es formen també fulles i branques, cada vegada que es trenqui l’estoló s’independitzaran nous individus. Aquesta capacitat s’utilitza en la multiplicació de les maduixeres (Fragaria vesca). D’estolons, però, en presenten també altres plantes de la Nostra flora, com és ara el gram negre (Potentilla reptans) . Algunes plantes, com l’arç blanc (Crataegus monogyna) o l’argelaga (Genista scorpius), produeixen branques curtes acabades en punxa que reben el nom d’espines. Però no tots els vegetals que punxen tenen espines: l’esbarzer (Rubus ulmifolius) i els rosers (Rosa) presenten aculis, també anomenats agullons, que són formacions només epidèrmiques que es desprenen fàcilment. Mentre que les espines semblen destinades sobretot a dissuadir els herbívors, els aculis s’utilitzen alhora com a sostenidors de les tiges llargues i poc consistents de certes plantes enfiladisses. Els circells caulinars, presents sobretot a les lianes, són rames transformades en filaments capaços d’entortolligar-se al voltant del suport; en tenen els ceps (Vitis vinifera), per exemple.
De vegades, les tiges han d’actuar en substitució d’unes fulles nul·les o bé tan reduïdes que, a la pràctica, ja no són funcionals. Una de les modificacions més espectaculars és la formació de fil·locladis, que no són altra cosa que tiges aplatades amb aspecte de fulla. En els del galzeran (Ruscus aculeata) es reconeix molt fàcilment el seu origen caulinar perquè porten, de primer, les flors i, finalment, els fruits. També són tiges aplatades les pales de la figuera de moro (Opuntia), anomenades cladodis per raó de la seva mida més gran.
Les tiges suculentes emmagatzemen aigua en parènquimes aqüifers especialitzats. Són freqüents en moltes plantes d’ambients àrids, com són ara les cactàcies, en les quals les fulles han quedat reduïdes a espines. El port d’aquestes plantes, anomenat cactiforme, és observable també en algunes euforbiàcies, asclepiadàcies, etc., en un procés de convergència adaptativa que ha donat com a resultat morfologies similars en el si de famílies molt allunyades filogenèticament.