Els protozous són protists heteròtrofs, que no constitueixen un grup natural. Una definició acceptable dels protozous, diu que són organismes eucariòtics essencialment unicel·lulars. Això vol dir, a grans trets, que en la seva majoria cada individu és una cèl·lula solitària i independent, bé que en algunes espècies hi ha una organització colonial en la qual els individus depenen mútuament els uns dels altres però sense arribar a especialitzar-se en funcions diferents, com passa, en canvi, amb les cèl·lules que formen els teixits i els òrgans dels metazous, les metàfites i els fongs. No són ni animals ni vegetals, però alguns mostren unes afinitats clares amb els primers mentre que d’altres s’assemblen més, biològicament parlant, als segons. Per aquesta raó, tradicionalment, les obres generals solen incloure entre les plantes els que mostren afinitats vegetals i reserven el nom de protozous exclusivament per als que presenten tendències animals, criteri que s’ha adoptat també en la present obra. La mateixa etimologia de la paraula «protozou», és a dir, «animal primitiu», col·labora a establir aquest criteri, que, realment, és força arbitrari.
Organització i estructura
Constitueixen un grup nombrós d’organismes, del qual existeixen segons que hom calcula, unes 25 000 espècies vivents (i un nombre semblant de fòssils), d’una importància biològica molt gran. En primer lloc perquè l’evolució dels protozous no s’ha dirigit vers la formació de teixits i òrgans sinó vers l’especialització funcional de diverses regions cel·lulars, de manera que en la seva única cèl·lula s’han desenvolupat estructures molt variades que no es troben en les cèl·lules dels organismes pluricel·lulars. En segon lloc, perquè la seva condició d’organismes aquàtics i les seves petites dimensions els han permès de colonitzar qualsevol ambient ric en aigua, des de les aigües dolces i marines fins als líquids interns dels organismes pluricel·lulars. I finalment també perquè gairebé la meitat dels protozous són paràsits de l’home i dels animals, i alguns els difonen greus malalties.
De les consideracions anteriors hom dedueix que els protozous constitueixen un grup ben heterogeni, els diferents representants del qual no tenen gaires trets en comú. No obstant això, hom pot consignar unes certes característiques força generals. La seva mida és molt petita i generalment són microscòpics: alguns mesuren escassament un micró i la majoria fan entre 10 i 50 microns. No obstant això, n’hi ha de relativament gegantins, com per exemple moltes gregarínes i la majoria de les espècies de foraminífers, que són visibles al primer cop d’ull; alguns, ja extingits, com els nummulits, tenen esquelets fòssils que arriben a fer uns quants centímetres. Això sense comptar els nombrosos casos de protozous colonials, en els quals una colònia, és a dir, una unitat biològica, pot mesurar uns quants mil·límetres, o els protozous polienèrgids, formats per un citoplasma amb nombrosos nuclis, que es podem mesurar en mil·límetres o fins i tot en centímetres. En els protozous típics, cada individu dels quals equival a una cèl·lula, el cos s’organitza com un tot, i podem considerar-hi presents les parts que reconeixem en una cèl·lula, és a dir, la membrana cel·lular i el citoplasma, amb els seus orgànuls i enclavaments, de vegades semblants i altres molt diferents dels que trobem a les cèl·lules animals o vegetals.
La membrana
La part que constitueix pràcticament la totalitat del cos, és a dir, el citoplasma, queda separada del medi extern per la membrana, és a dir, una fina capa bilipídica amb inclusions proteiques, que s’ajusta al model molecular del mosaic fluid, i que sol rebre el nom de plasmalemma o membrana citoplasmàtica. Aquesta membrana pot ésser reforçada per microtúbuls, microfibril·les o vesícules membranoses i aplanades, estructures que engruixeixen i donen solidesa a aquesta regió del citoplasma, que en aquest cas rep diverses denominacions com periplast, pel·lícula, cutícula o escorça. Quan la capa limitant és constituïda exclusivament pel plasmalemma, que és el cas més freqüent, és fàcilment deformable i el citoplasma és capaç de desplaçar-se mitjançant pseudopodis (expansions lobulars del cos), que serveixen per al moviment. D’altra banda, en d’altres protozous, com els flagel·lats i els ciliats, l’escorça és la base on s’implanten altres orgànuls de moviment, els flagels i els cilis. En la seva superfície externa, el plasmalema pot ésser recobert, com passa amb les amebes, per un glicocàlix o capa mucopolisacàrida, equivalent al que es troba en moltes cèl·lules animals; però també pot ésser reforçat amb formacions esquelètiques exteriors a la cèl·lula.
El citoplasma
En els diversos protozous, el citoplasma té característiques diferents, ja que pot ésser més o menys consistent i més o menys ric en orgànuls i inclusions, segons els grups taxonòmics. Però és freqüent, especialment en les formes ameboides, que es trobi dividit en dues zones: una d’exterior, més fluida i d’aspecte més homogeni, l’ectoplasma, i una altra de més viscosa, d’aspecte granular i més rica en inclusions, l’endoplasma.
Les inclusions citoplasmàtiques
En el si del citoplasma dels protozous hi ha inclusions molt diverses, entre les quals cal distingir, a grans trets, els orgànuls, amb funcions fisiològiques diverses, i les inclusions homogènies en el citoplasma, les quals són producte de l’activitat fisiològica del protozou. Hom parlarà en primer lloc dels orgànuls; alguns es troben a tots o a gairebé tots els protozous, mentre que d’altres són específics d’uns grups determinats.
El reticle endoplasmàtic o ergastoplasma, en primer lloc, forma un conjunt de cisternes membranoses amb la superfície externa revestida de ribosomes en els quals es fa la síntesi de proteïnes. La importància que pren aquest reticle endoplasmàtic és molt diferent en uns i altres protozous, però també varia segons l’estat fisiològic de l’organisme. L’aparell de Golgi, que es troba en tots els protozous sense excepció, elabora i segrega diverses substàncies, i pot presentar característiques diverses en els diferents protozous: bé que generalment pren la forma de dictiosomes aïllats i escassos (de vegades un de sol, com en els tripanosomes), també pot presentar-se com un dictiosoma múltiple (en els flagel·lats de l’intestí dels tèrmits), i llavors rep també el nom d’aparell parabasal. Els mitocondris també són presents en gairebé tots els protozous. Generalment tenen la mateixa estructura que els de les cèl·lules dels metazous i es componen d’una coberta feta de dues membranes, la interior de les quals s’invagina i forma unes crestes transversals; però en alguns, com en els ciliats, les crestes no són planes sinó tubulars, com els dits d’un guant, i en d’altres, com els esporozous, són vesiculars, en forma d’ampolla. En general, els mitocondris són nombrosos dins la cèl·lula, però també hi ha protozous que en tenen molt pocs. Hi ha, a més, alguns protozous, gairebé tots paràsits, que no en tenen (com per exemple els tricomonàdids, les amebes paràsites del gènere Entamoeba i els microsporidis). D’altra banda, algunes característiques hereditàries dels protozous són producte de l’expressió genètica del DNA contingut en els mitocondris; cal recordar el cas dels tripanosomes i els seus afins, que tenen un sol mitocondri, una part del qual és diferenciada en forma d’un corpuscle molt visible, el cinetoplast, on es concentra tot el DNA mitocondrial, amb una organització molecular especial.
El nucli conté quasi tot el DNA, i varia segons el grup de protozous, tant de forma i de nombre com d’organització. En general, en els protozous hi ha un sol nucli, que pot ésser ben arrodonit, gros i vesicular, proveït d’un corpuscle central (cariosoma), o bé ésser més petit, i de vegades de forma irregular, més o menys compacte i homogeni. Però també és freqüent, especialment en protozous de grans dimensions (com les opalines), que hi hagi molts nuclis tots-semblants; llavors es diu que són protozous polienèrgids, en els quals cada nucli forneix l’RNA missatger als ribosomes del territori citoplasmàtic circumdant (enèrgida). Una altra excepció important a la regla general de posseir un sol nucli per individu és el cas dels protozous ciliats, que tenen un aparell nuclear doble, amb dos tipus de nucli: el macronucli, gros, i el micronucli, més petit. En general, en un protozou ciliat n’hi sol haver un de cada mena, però també n’hi ha que tenen diversos macronuclis i, encara més freqüentment, més d’un micronucli. La diferència entre uns i altres nuclis no consisteix exclusivament en les dimensions: el micronucli és diploide, es divideix per mitosi i no produeix RNA; en canvi, el macronucli és poliploide, es divideix per amitosi i produeix l’RNA necessari per a la síntesi de proteïnes en el citoplasma. Els macronuclis es formen a partir dels micronuclis durant els processos sexuals.
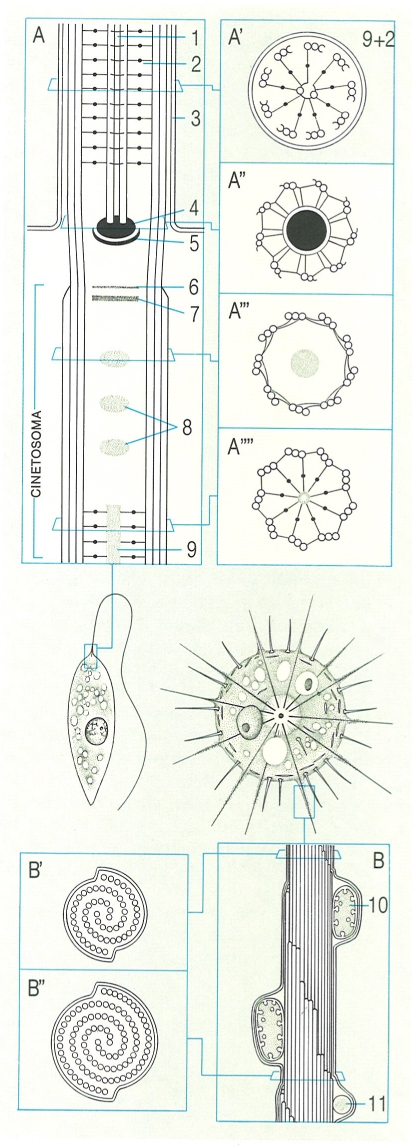
Estructura dels undulipodis o orgànuls vibràtils en els protozous: A Tall longitudinal i talls transversals del flagel i el cinetosoma del flagel·lat Leptomonas: A tall longitudinal; A’ tall transversal del flagel; A" tall transversal de la zona de transició a l’altura del cinetosoma; A"’ i A"" talls transversals del cinetosoma a l’altura mitjana i a la zona de roda de carro. B Esquema de l’axopodi de l’heliozou Echinosphaerium nucleofilum; noteu, sota la membrana citoplasmàtica, els dos mitocondris i els grànuls. B’ i B" Disposició dels microtúbuls en seccions transversals. 1 Microtúbuls, 2 doblets de l’axonema, 3 membrana cel·lular, 4 axosoma, 5 septe, 6 placa addicional, 7 placa terminal, 8 grànuls axials, 9 eix de la roda de carro, 10 mitocondri, 11 grànul.
Biopunt, a partir de fonts diverses
Els undulipodis o orgànuls vibràtils constitueixen un dels mecanismes més freqüents de moviment dels protozous. Filiformes i implantats a l’escorça cel·lular, poden ésser de dos tipus molt semblants, els cilis i els flagels, uns i altres molts difosos en els diferents grups de protozous. No tenen diferències substancials, però els flagels solen ésser llargs i poc abundants, mentre que els cilis són més curts, molt nombrosos i, sobretot, es disposen al llarg de l’escorça fent unes llargues files anomenades cinèties, que són les veritables unitats fisiològiques del moviment en els protozous ciliats. D’altra banda, els flagels es troben en tots els grups de flagel·lats, en alguns rizòpodes i actinòpodes, i també en algunes fases del cicle biològic dels esporozous. Estructuralment, tant els cilis com els flagels són constituïts per un eix de microtúbuls anomenat axonema, revestit d’una membrana finíssima que és una continuació del plasmalemma. Els microtúbuls estan distribuïts formant nou doblets perifèrics, amb dos microtúbuls sencers i un d’incomplet (braços) cadascun; els nou doblets envolten una parella de microtúbuls centrals. Els microtúbuls de l’axonema acaben en un orgànul, sempre present als cilis i als flagels, anomenat cinetosoma, una formació constituïda per nou triplets curts de tres microtúbuls cadascun, sense microtúbuls centrals; és a dir: cada triplet del cinetosoma es prolonga amb un doblet de l’axonema. Tant els cilis com els flagels són formats per moltes proteïnes diferents, les més importants de les quals són les tubulines, que constitueixen els microtúbuls de l’axonema i del cinetosoma, i la dineïna, que forma els «braços» adosats als doblets de l’axonema. A la base dels cilis i els flagels, i en relació amb el cinetosoma, hi ha unes formacions fibril·lars de naturalesa variada (microtubular, microfibril·lar o d’estructura periòdica). Un cili o un flagel, amb el seu cinetosoma i les seves formacions fibril·lars adosades, constitueix la unitat de moviment, que rep el nom de cinètida.
El citoplasma dels protozous és recorregut, com passa també en moltes cèl·lules animals, per uns sistemes fibril·lars que en conjunt reben el nom de citosquelet. La naturalesa i les funcions d’aquestes fibres són variades i moltes vegades són específiques d’uns determinats grups de protozous. En general, es pot dir que les principals funcions que compleixen estan en relació amb la conservació de la forma, la locomoció, la contractibilitat i el transport de partícules o d’orgànuls a través del citoplasma.
Alguns d’aquests sistemes fibril·lars es localitzen a l’escorça dels protozous, mentre que d’altres es troben al citoplasma superficial (ectoplasma) i d’altres al citoplasma profund (endoplasma). Com a exemples més representatius dels elements del citosquelet hom pot esmentar els sistemes fibril·lars d’actina-miosina que es troben en les amebes i que són responsables del moviment ameboide, i les fibril·les proteiques de l’epiplasma, dels ciliats, probablement relacionades amb el manteniment de la forma. Els abundants sistemes microtubulars que es troben en el citoplasma d’una gran quantitat de protozous mereixen també una menció especial: per exemple, en els esporozous i els flagel·lats com els tripanosomes, hi ha una capa molt atapeïda de microtúbuls subpel·liculars, paral·lels, que es dirigeixen de davant cap enrere i que probablement serveixen per a mantenir la forma del cos i els seus moviments de contracció. En els axopodis dels heliozous i altres actinòpodes, un eix d’abundants microtúbuls, organitzats segons uns models complicats i propis de cada espècie, serveix per a donar-los rigidesa i moviment. Ja hem vist que, en l’escorça dels ciliats, diverses parts de la cinètida són fibres microtubulars relacionades amb el cinetosoma; aquestes fibres també són part del citosquelet, igual que les fibres periòdiques de la mateixa cinètida, bé que és dubtós que s’hagin de considerar constituents del citosquelet els mateixos microtúbuls del cinetosoma i del cili. També en relació amb els cilis hi ha paquets microtubulars anomenats nemadesmes, que es dirigeixen cap a la profunditat del citoplasma; en molts zooflagel·lats, com els diplomonàdids, hi ha unes llargues fibres, de naturalesa microfibril·lar i associades als cinetosomes dels flagels. Finalment, cal recordar que en els ciliats suctors, que s’alimenten mitjançant uns tentacles xucladors, aquests estan proveïts d’uns sistemes longitudinals de microtúbuls semblants als que recorren l’eix dels axopodis dels heliozous. Les fibres del fus que es forma durant la mitosi també són de naturalesa microtubular; és més, formen una part important del citosquelet cel·lular i participen en els variats processos mitòtics dels protozous. Les fibres proteiques contràctils o mionemes, que serveixen per a fer diversos moviments del cos en molts protozous i que de vegades són molt visibles, també formen part del citosquelet; han estat molt estudiades en els acantaris, i també en diversos ciliats, com Stentor, Spirostomum, Vorticella i d’altres.
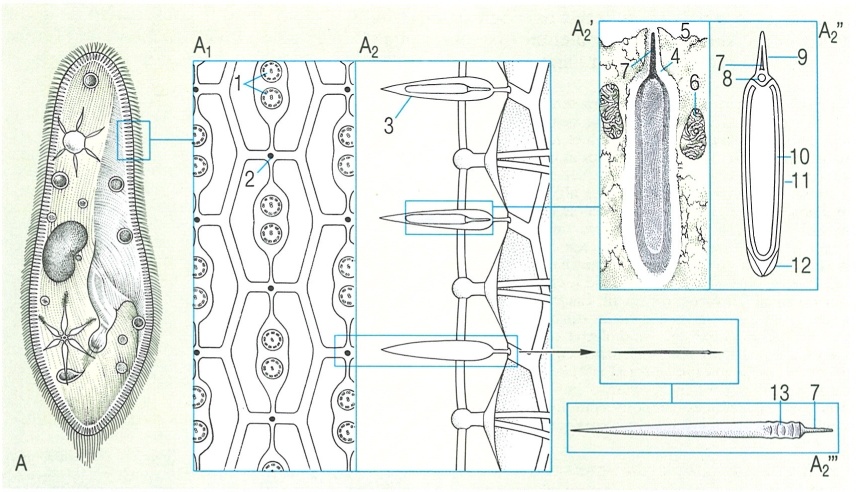
Estructura dels extrusomes (tricocists) en un parameci. A1, A2 Esquemes de l’escorça ampliada del protozou, en secció transversal i longitudinal respectivament, per a veure com es distribueixen els porus dels tricocists (2) entre els cilis (1). i detalls de tricocists (en morat) continguts en l’escorça, abans de disparar-se (A2’ i A2"), i un cop disparats a l’exterior (A2"’). Presents en els protozous més grossos, els extrusomes es disparen per estimuls químics o mecànics, i aboquen a l’exterior el seu contingut. Hom ha assenyalat en el dibuix: 1 cilis, 2 porus del tricocist, 3 càpsula del tricocist, 4 vesícula, 5 plasmalemma, 6 mitocondri, 7 punta, 8 grànul, 9 extrem de la vesícula, 10 tija, 11 membrana, 12 membrana del tricocist, 13 estries.
Biopunt, a partir de fonts diverses
Els extrusomes, molt característics dels protozous, es poden definir com uns orgànuls proveïts d’una membrana i situats en la superfície del cos, els quals, per l’acció d’estímuls químics o mecànics, poden disparar-se i evacuar instantàniament el seu contingut a l’exterior.
S’han descrit molts tipus diferents d’extrusomes, però la majoria encara estan molt mal definits i se’n desconeix la utilitat. Molts han estat descrits en flagel·lats d’afinitats vegetals i de vegades hom ha donat diferents noms a les mateixes estructures o a estructures molt semblants. Els extrusomes més coneguts i difosos són els mucocists, els tricocists, els toxicists i els haptocists. Els mucocists es troben en els ciliats i els flagel·lats. En els primers apareixen en files regulars entre les cinèties i són formacions corticals en forma de sac voltat d’una membrana; a través d’un petit porus de la pel·lícula, descarreguen el seu contingut mucós paracristal·lí, que s’estén per la superfície del ciliat sense que aquest perdi la seva estructura; és possible que els mucocists tinguin un paper en la formació del cist dels ciliats. Els tricocists són extrusomes grossos, també de contingut paracristal·lí, que es disposen regularment a l’escorça d’alguns ciliats de mida grossa (com Paramecium i Frontonia) i que desemboquen en porus regularment alineats entre les cinèties. Quan el tricocist es dispara explosivament, llença a l’exterior el seu contingut en forma d’una llarga cinta estriada que acaba en una punta d’aspecte compacte i homogeni. Els tricocists es formen al reticle endoplasmàtic i emigren després fins a l’escorça; la seva funció continua avui essent enigmàtica. Els toxicists són extrusomes llargs, prims i tubulars, carregats de fosfatases àcides, que es troben concretament en determinats ciliats depredadors que s’alimenten d’altres ciliats (com Litonotus i Dileptus), alineats al llarg d’una «trompa» situada a la part anterior del cos; sembla que disparant els toxicists, el protozou depredador aconsegueix de paralitzar i fins i tot d’iniciar la citòlisi de la presa. Els haptocists són semblants als toxicists, es troben a l’extrem dels tentacles xucladors dels suctors i als axopodis dels heliozous, i serveixen per a paralitzar i capturar les preses.
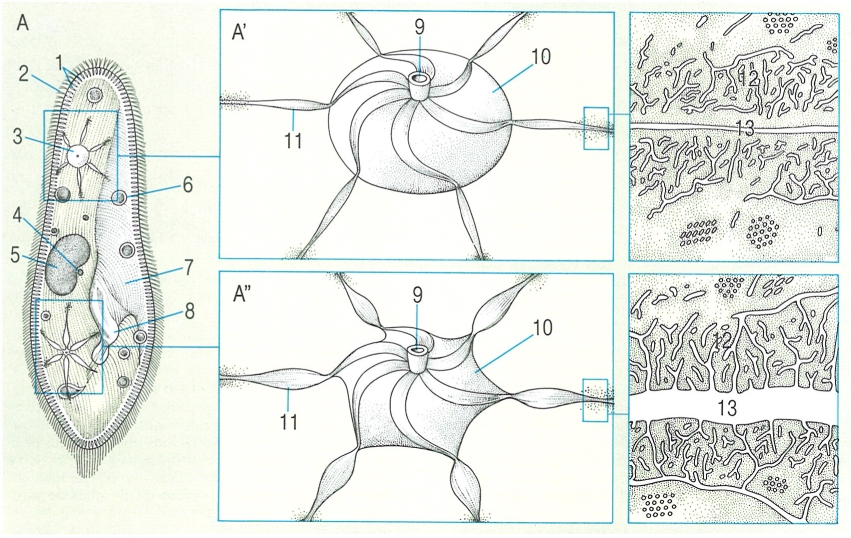
Estructura i funcionament dels vacúols pulsàtils en un parameci (A): en el seu funcionament, el vacúol és ple de líquid en la fase de diàstole (A’) i buit en la de sístole (A"). 1 Càpsules dels tricocists, 2 cilis, 3 vacúol pulsàtil o contràctil, 4 micronucli, 5 macronucli, 6 vacúol digestiu, 7 vestíbul, 8 citòstoma, 9 porus excretor, 10 dipòsit, 11 ampolla, 12 reticle endoplasmàtic, 13 canal radial.
Biopunt, a partir de fonts diverses
La majoria dels protozous que menen una vida lliure a l’aigua dolça, principalment rizòpodes, flagel·lats i ciliats, tenen uns orgànuls molt peculiars que no es troben en altres cèl·lules animals o vegetals: els vacúols contràctils, també anomenats vacúols pulsàtils. Es tracta d’unes cavitats plenes d’aigua, la paret de les quals es contrau rítmicament per tal d’evacuar el seu contingut a través d’un porus. En molts protozous n’hi ha un de sol, però tampoc és estrany que n’hi hagi més, i fins i tot poden ésser nombrosos en un mateix individu. En el seu moviment rítmic, el vacúol contràctil té dues fases: la diàstole, quan és ple de líquid o en vies d’omplir-se, i la sístole, quan es buida; en general, la diàstole és lenta i la sístole ràpida, però el ritme del vacúol depèn de tota una sèrie de variables, entre les quals hi ha l’estat fisiològic del protozou, la temperatura ambiental i la concentració de sals dissoltes en l’aigua.
Aquest vacúol sol ésser d’estructura senzilla i, quan és així, únicament es compon d’una fina membrana a la qual arriben uns canalets connectats amb el reticle endoplasmàtic, que només es fan perceptibles al microscopi electrònic; en d’altres protozous, pot ésser més complicat i pot tenir un porus permanent, i, en alguns ciliats, com en Paramecium, cadascun dels vacúols contràctils d’un mateix individu s’envolta de diverses ampolles des d’on l’aigua passa al vacúol per a ésser evacuada; la paret d’aquestes ampolles és envoltada per uns llargs microtúbuls que intervenen en la seva contracció i, en general, en el funcionament del vacúol contràctil. Encara que no es pot descartar que pel vacúol s’evacuïn substàncies solubles com a producte final del metabolisme cel·lular, sembla que es tracta d’un orgànul essencialment extractor d’aigua: expulsa l’aigua que entra al citoplasma per osmosi a través de la membrana cel·lular, o amb vacúols fagocitaris; així, el vacúol es comporta com un orgànul osmoregulador, encarregat d’eliminar l’aigua que hi ha en excés.
Les inclusions citoplasmàtiques més o menys inertes són principalment, com ja ha estat dit, productes de l’activitat metabòlica cel·lular que resten un cert temps al citoplasma amb la finalitat d’ésser utilitzats després (materials de reserva) o per a ésser eliminats definitivament (materials d’excreció). Els materials de reserva generalment són polisacàrids o lípids, i es troben principalment en els flagel·lats d’afinitats vegetals, on són freqüents grans dipòsits de midó o d’altres polímers de sucres; aquests materials també es troben sovint en protozous paràsits, com els esporozous, amb forma de grànuls d’amilopectina. Pel que fa als productes d’excreció, solen ésser inclusions de naturalesa inorgànica, insolubles, com els cristalls de carbonat de bari que apareixen en el citoplasma dels foraminífers.
Els elements esquelètics

Exemples de diferents tipus d’esquelet en els protozous: testa o conquilla calcària de foraminífer (a dalt); teca silicia del rizòpode Euglypha (a baix).
Index, Pilar Gracia
Els elements esquelètics són estructures generalment rígides, de naturalesa inorgànica o formades per compostos orgànics, que apareixen en molts protozous fonamentalment amb la missió de mantenir la forma de l’organisme o de defensar-lo contra les inclemències del medi.
En alguns casos se situen a l’interior del protozou i llavors solen ésser estructures inorgàniques, rígides i ben visibles, cosa que trobem gairebé exclusivament en alguns actinòpodes, als quals serveixen (igual que els elements del citosquelet) per a mantenir la forma; per exemple, és el cas de les grans espícules de sulfat d’estronci que caracteritzen el grup dels acantaris.
Són més característics, però, els esquelets interns dels radiolaris (sovint anomenats closques), d’estructura molt complexa, compostos de diversos elements, espines o plaques de sílice amorfa, que se solden o s’articulen entre si i en conjunt formen una estructura buida que acull al seu interior una gran part del citoplasma. És semblant el cas de les testes o closques calcàries dels foraminífers, que generalment es componen de diverses cambres dins les quals s’allotja una gran part del citoplasma, mentre una altra gran part surt pels foradets de la mateixa testa i l’envolta per fora, de manera que aquesta queda convertida en una estructura esquelètica interna. Un cas contrari és el de les anomenades amebes testàcies, en les quals la testa o closca conté tot el citoplasma excepte una petita part que surt per un únic orifici i que es pot retraure cap endins, de manera que segurament té la funció de protegir la cèl·lula del medi extern: aquesta testa pot ésser de naturalesa orgànica (quitinoide) o inorgànica (especialment, calcària) però també pot ésser formada per escates segregades o per elements minerals del medi, que l’ameba aglomera. Altres formacions esquelètiques perifèriques dels protozous són incompletes i proporcionen a la cèl·lula una protecció menys eficaç i no tan permanent. Es tracta de les llorigues, una mena de closques de naturalesa orgànica (de vegades endurides amb grans de sorra) que formen una casa més que una coberta adherent, ja que el protozou hi viu ficat a dins i hi té completa llibertat de moviments. La lloriga és amplament oberta per la seva part anterior i pot ésser de formes molt variades (tubular, de campana, de bot, etc.). Podem trobar llorigues en espècies de diversos grups, com per exemple, en algunes euglenes, alguns coanoflagel·lats, determinats ciliats perítrics, i tots els ciliats tintínnids. Finalment, cal esmentar com un element més de protecció de la cèl·lula la membrana cística, que envolta els estadis de resistència (cists) i és de naturalesa orgànica complexa i força resistent.
Biologia dels protozous
Els protozous, ja que no constitueixen un conjunt natural d’organismes, no tenen tampoc gaires coincidències quant a la seva biologia. No obstant això, el fet d’ésser unicel·lulars i fins i tot la seva mateixa talla microscòpica, fa que hi hagi algunes característiques biològiques que els sòn comunes. La petita mida dels protozous fa que siguin extremament vulnerables a la dessecació, la qual cosa vol dir que no poden viure si no és en medis aquosos: es pot afirmar que un protozou extret del seu medi es mor instantàniament perquè perd tota l’aigua del citoplasma per evaporació a través de la membrana plasmàtica. L’aigua és, doncs, la primera condició de vida dels protozous: habiten tant les aigües dolces i marines com els líquids corporals dels animals i fins i tot l’interior de les cèl·lules. Això, amb tot, té la seva excepció en el cas dels cists, que, com hem dit, presenten una resistència extrema a la dessecació. La segona condició per a viure dels protozous és la presència d’oxigen al medi, ja que la majoria són aerobis. N’hi ha alguns, però, que poden viure en ambients amb una tensió d’oxigen escassa, com per exemple al fons de basses profundes o en els llots sapropèl·lics. És especialment interessant el cas de la gran ameba Pelomyxa palustris, que en lloc de mitocondris conté uns bacteris simbionts que empren l’oxigen per a la respiració del protozou.
El moviment
En general, els protozous es mouen d’una manera molt activa i es desplacen per cercar l’aliment o per trobar condicions ambientals adequades. Són molt marcats i coneguts els tactismes que presenten, especialment el quimiotactisme, el tigmotactisme i el geotactisme. Molts no tenen uns orgànuls específics per al moviment, i aleshores es mouen per contraccions i flexions del cos, amb la participació dels microtúbuls de la pel·lícula o dels mionemes de l’escorça; però la majoria tenen uns orgànuls específics que, en molts casos, també serveixen per a recollir l’aliment del medi. En general, es poden distingir tres tipus d’orgànuls per al moviment: els pseudopodis, els flagels i els cilis.
Els pseudopodis són formacions més o menys transitòries que, en general, es formen i desapareixen ràpidament; són característiques principalment dels rizòpodes i els actinòpodes. Hom n’ha descrit tres tipus fonamentals. En primer lloc, hi ha els anomenats pseudopodis lobats o lobopodis, que es donen sobretot en les amebes, i que serveixen també per a capturar partícules alimentàries sòlides. Són unes eminències o evaginacions que sorgeixen i desapareixen de la superfície de la cèl·lula sobtadament, gruixudes i arrodonides, en les quals s’aprecia una zona externa clara, ectoplasmàtica, que n’envolta una d’interna més fosca, endoplasmàtica. La seva formació obeeix a una transformació local en sol del gel citoplasmàtic, amb la qual cosa, en aquest punt, el citoplasma «vessa» cap a la membrana de la cèl·lula; llavors, aquest citoplasma fluid és pressionat cap enfora per la contracció dels filaments d’actina-miosina de l’ectoplasma. Algunes amebes (del tipus límax) es desplacen per mitjà d’un sol lobopodi anterior, mentre que d’altres (del tipus pròteus) emeten simultàniament diversos pseudopodis lobats i es desplacen cap on es dirigeixen la majoria. En segon lloc hi ha els pseudopodis filosos o fil·lopodis, que tenen algunes amebes testàcies i alguns radiolaris; són expansions d’ectoplasma llargues i finíssimes que no s’uneixen amb les veïnes, expansibles i retràctils, que també funcionen per a capturar partícules alimentàries. Poden executar el moviment per la seva mateixa contractibilitat o bé si estan dotats d’un moment flagel·liforme: estan proveïts d’unitats microtubulars que es traslladen al llarg del pseudopodi. En tercer lloc distingim els pseudopodis anomenats granuloreticulosos o també reticulopodis, que són propis dels foraminífers i d’alguns radiolaris. Són unes fines expansions citoplasmàtiques perifèriques que es ramifiquen molt i s’anastomosen amb les veïnes, de manera que el conjunt és com una xarxa finíssima i complicada, que serveix tant per a la locomoció com per a la captura de preses. El citoplasma d’aquests pseudopodis és molt granulós, i els seus grànuls són conduïts passivament per uns corrents cítoplasmàtics que els recorren en ambdós sentits, moviment en el qual probablement hi ha implicades algunes molècules contràctils. Finalment, els axopodis, típics dels actinòpodes, tenen la particularitat de posseir un eix microtubular resistent, que els confereix una rigidesa característica. Aquest eix és format per un o més paquets paral·lels de microtúbuls que es disposen simètricament segons un model determinat que varia en cada família i en cada gènere. Diferentment d’altres pseudopodis, no són gens versàtils, de manera que constitueixen formacions més o menys permanents de la cèl·lula, i la seva missió principal és la de conservar-ne la forma, bé que també poden ajudar a capturar les preses, ja que en el citoplasma que els envolta s’observa la presència d’haptocists capaços de paralitzar les víctimes; aquest citoplasma perifèric és molt mòbil, gràcies a la presència de molècules d’actina.
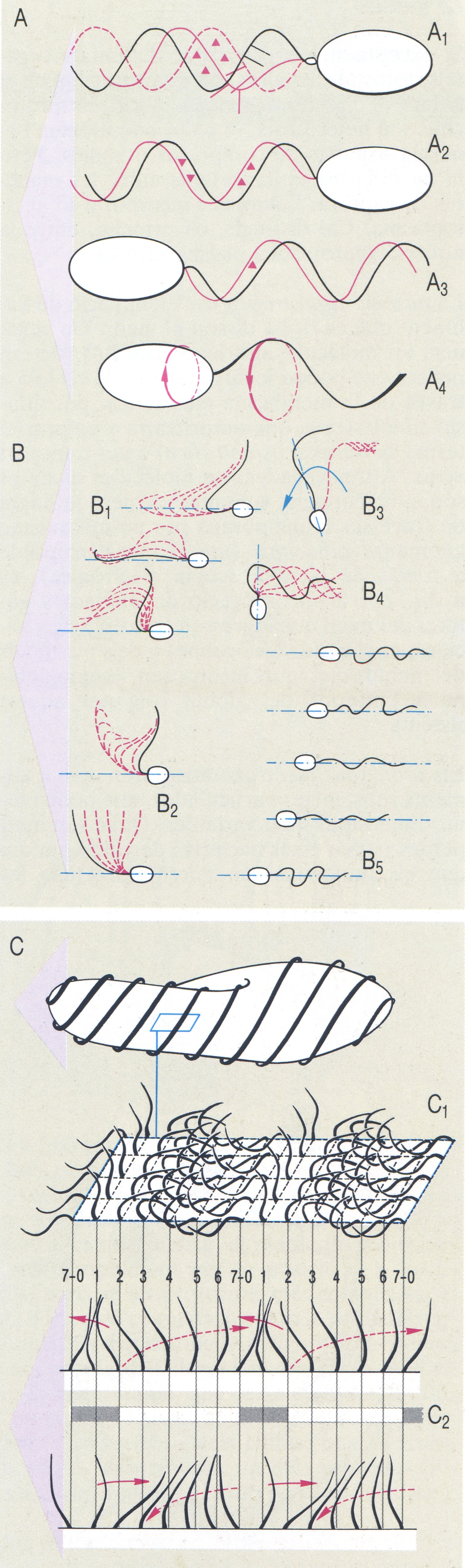
Diversos tipus de moviment (en vermell) que permeten els flagels i els cilis dels protozous. Les fletxes indiquen la direcció del moviment. A Moviments flagel·lars: A1 i A2 model de moviment amb el flagel anterior (A1 amb mastigonemes); A3 i A4 model de moviment amb el flagel posterior (en A4, ones helicoïdals). B Moviments flagel·lars en diverses espècies de Monas: B1 moviment ràpid, B2 moviment moderat, B3 moviment de gir, B4 moviment lateral, B5 moviment cap enrere. C Moviment metacrònic en els ciliats (Paramecium) C1 Esquema que mostra la coordinació del moviment de batuda dels cilis; C2 detall de l’anterior que mostra la metacronia simplèctica (a dalt) i antiplèctica (a baix). Les fletxes vermelles contínues indiquen moviments ràpids efectius; les discontínues, moviments lents de recuperació.
Biopunt, a partir de fonts diverses
Els flagels i els cilis, a l’estructura dels quals ja ens hem referit anteriorment, es mouen per un principi que probablement és molt semblant a escala molecular. L’element actiu és l’axonema microtubular, especialment els doblets de microtúbuls perifèrics i els apèndixs laterals de cada doblet, que reben el nom de «braços». Aquests braços de dineïna, estimulats per la despolarització produïda localment per l’entrada d’ions de calci (equilibrada per la sortida d’ions de potassi), canvien de conformació i formen ponts amb el doblet veí, la qual cosa fa que uns doblets microtubulars llisquin sobre els altres i, per tant, hi hagi una deformació, en sentit lateral, del cili o el flagel. D’altra banda, però, els cilis i els flagels no es mouen igual: els flagels es mouen per ondulacions que es desplacen per tota la seva longitud, mentre que els cilis realitzen el moviment en dos temps clarament diferenciats, un d’«eficaç», en el qual el cili es doblega ràpidament i bat amb energia l’aigua del voltant, i un altre de «recuperació», pel qual el cili es redreça lentament fins que recupera la seva posició erecta de partida; el moviment eficaç és el que determina l’acció motriu del cili. Cal dir que l’acció dels flagels sol ésser individual, mentre que els cilis estan coordinats en el seu moviment col·lectiu, i precisament en aquest sentit la cinètia pot ésser considerada com la unitat de moviment ciliar. En efecte, en els ciliats es pot observar que al llarg de cada cinètia els cilis successius estan, en un determinat moment, en una fase del seu moviment lleugerament desplaçada respecte del cili anterior, de manera que es pot veure com una ona de batuda ciliar recorre la cinètia; aquest moviment metacrònic és la causa que la superfície d’un ciliat en moviment sigui semblant a un camp de blat quan les espigues es mouen per l’acció de bufades de vent successives.
La nutrició
Si exceptuem els protozous d’afinitats vegetals, fotosintetitzadors, els protozous són heteròtrofs, és a dir, adquireixen l’energia a partir de compostos orgànics. Però hi ha diferències importants entre les maneres de capturar l’aliment i incorporar-lo al citoplasma. Cal distingir, en principi, entre la nutrició saprotròfica i la fagotròfica.
La nutrició saprotròfica és l’assimilació de l’aliment que es troba dissolt al medi. De vegades, les molècules nutrients dissoltes són tan petites que poden incorporar-se a la cèl·lula a través de la membrana plasmàtica, per difusió simple (cosa que autoritzaria a emprar el terme de nutrició osmotròfica) o per transport actiu. Altres vegades, les molècules dissoltes són suficientment voluminoses perquè hagin de travessar la membrana per un mecanisme anomenat pinocitosi (únicament perceptible a través de la microscòpia electrònica), és a dir, per la incorporació de diminutes gotetes del medi que ingressen en forma de vesícules; és el cas de les opalines o dels plasmodis del paludisme, que incorporen l’hemoglobina a l’interior del glòbul roig per aquest sistema.
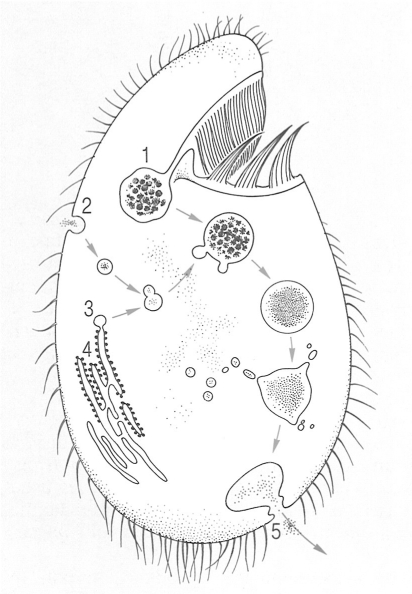
Formació d’un vacúol digestiu en un ciliat (Tetrahymena pyriformis). Paral·lelament a la formació del vacúol, les vesícules pinocítiques es fusionen amb els lisosomes i descarreguen després els enzims a l’interior del vacúol. Aquest entra llavors en la fase digestiva i seguidament les substàncies nutritives s’escampen pel citoplasma per mitjà de petits vacúols; posteriorment les restes són expulsades a l’exterior de la cèl·lula. 1 Vacúol digestiu, 2 vesícula pinocítica, 3 lisosoma, 4 reticle endoplasmàtic rugós, 5 citopigi.
Biopunt, a partir de fonts diverses
Els protozous fagòtrofs incorporen com a aliments substàncies particulades, que poden tenir dimensions molt variables. Quan són molt petites (solen ésser bacteris i detrits) hom diu que els protozous són micròfags. Quan les partícules són grosses hom parla de protozous macròfags, —com passa amb els ciliats que viuen al rumen dels remugants, que sovint ingereixen fragments vegetals tan grossos que els arriben a deformar el cos. Altres protozous són tan voraços que poden ingerir preses senceres: són els depredadors, com els ciliats del gènere Didinium, que poden engolir un parameci sencer més gros que ells mateixos, diverses vegades al dia. Els protozous fagòtrofs poden ingerir les partícules de diverses maneres: en alguns casos, com en les amebes, els pseudopodis lobats es dirigeixen cap a la presa i l’engloben juntament amb una petita gota d’aigua; en d’altres, com en els foraminífers, la presa queda atrapada a la xarxa formada pels pseudopodis granuloreticulosos; i encara, en d’altres, la partícula capturada entra en la cèl·lula per mitjà d’una regió especialitzada, la boca. Aquesta boca pot ésser simplement un orifici circular o bé una fenedura, i pot estar dotada d’una sèrie d’estructures destinades a fer més efectiva la ingestió (varetes per a mantenir obert l’orifici bucal o citòstoma, fibres intracel·lulars destinades a conduir l’aliment, flagels o cilis especialitzats que provoquen corrents per a portar les partícules a la boca). Sempre que hi ha una alimentació fagotròfica, la partícula ingerida acaba essent englobada dins una cavitat anomenada vacúol digestiu, en la qual és digerida efectivament; en aquest vacúol, que conté la presa i l’aigua ingerida simultàniament, hi van a parar els enzims digestius continguts en els lisosomes produïts al reticle endoplasmàtic, que degraden els compostos nutritius. Després, per un procés molt semblant a la pinocitosi, anomenat endocitosi, el contingut del vacúol passa al si del citoplasma dividit en gotetes minúscules, mentre el vacúol va disminuint de volum fins a desaparèixer o bé expulsa, convertit en vacúol fecal, les seves restes indigeribles a l’exterior; aquesta expulsió pot fer-se per qualsevol punt de la cèl·lula, com passa en les amebes, o bé per una estructura permanent que s’hi ha especialitzat, el citopigi, com passa en molts ciliats.
La reproducció
La majoria dels protozous es reprodueixen fonamentalment per un procés asexual, pel qual l’individu transmet als seus descendents una dotació genètica idèntica a la seva. Però també és extraordinàriament freqüent que en la vida dels protozous es produeixin fenòmens de transferència genètica entre individus que no tenen una relació de progenitor i descendent, és a dir, de sexualitat; aquestes transferències de vegades són semblants al que passa en animals superiors i d’altres, en canvi, se’n distancien enormement.
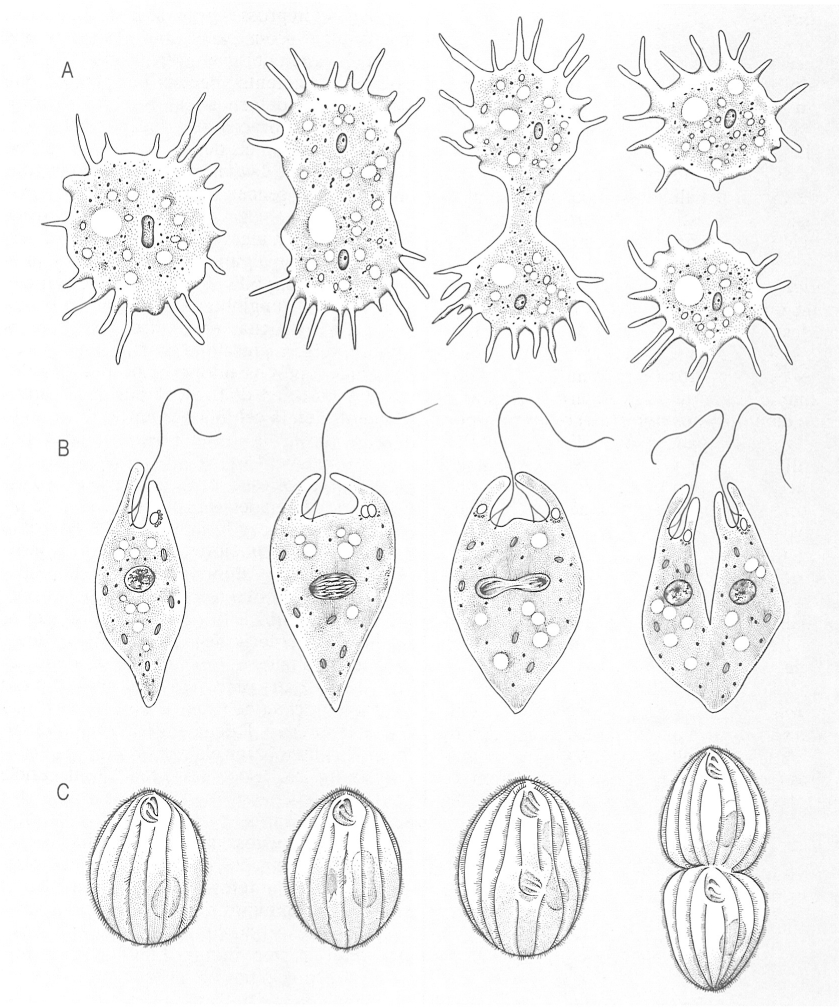
L’escissió binària és el tipus de reproducció asexual més freqüent en els protozous. El dibuix en recull tres casos diferents: A Escissió binària en les amebes (Amoeba); B escissió longitudinal en els flagel·lats (Euglena); C escissió transversal en els ciliats (Tetrahymena).
Biopunt, a partir de fonts diverses
La reproducció asexual pot ésser molt senzilla i es pot comparar a la reproducció de les cèl·lules dels teixits d’animals i plantes: el protozou multiplica el seu nucli per mitosi, duplica les seves altres estructures i orgànuls i després s’escindeix en dos individus fills, semblants en tot a l’individu matern. Però dins aquesta anomenada escissió binària hi ha moltes variacions. En primer lloc, atesa l’heterogeneïtat dels grups de protozous, no és estrany que hi hagi diferents tipus de mitosi, que difereixen per l’estructura i el comportament dels cromosomes, per la disposició i el moment d’aparició dels microtúbuls del fus acromàtic i, sobretot, per la permanència o no de la membrana nuclear, la qual sol conservar-se durant tota la mitosi, bé que en alguns casos arriba a desaparèixer totalment.
Una diferència notable entre els protozous i altres organismes a l’hora de fer aquesta simple escissió és la posició del pla de divisió entre les dues cèl·lules filles. En les amebes, mancades de simetria, el pla de divisió es defineix simplement per la posició dels nuclis fills; però en els protozous més complicats, el pla d’escissió es relaciona amb la resta dels orgànuls. Es pot distingir la divisió simetricogènica, pròpia dels flagel·lats, on el pla de divisió és longitudinal, que deixa els individus fills a un costat i l’altre, de la divisió homotetogènica, característica dels ciliats, que és transversal, de manera que un dels individus fills és anterior (pròter) i l’altre posterior (opíster).
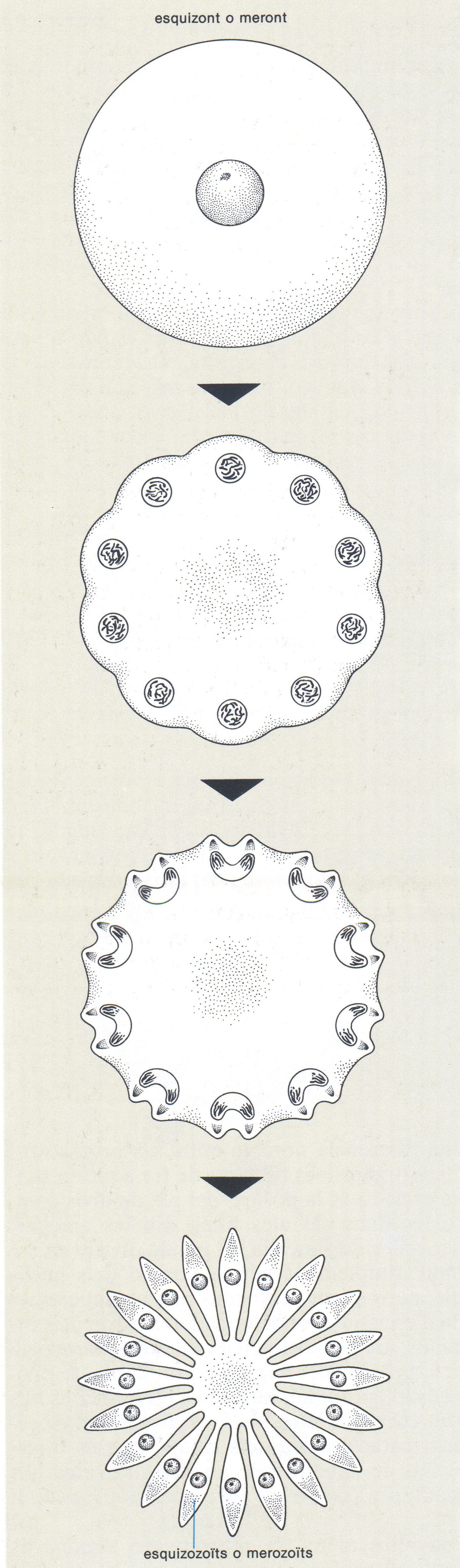
L’escissió múltiple, també anomenada esquizogònia o merogònia, és un procés de reproducció asexual, típic dels esporozous i els foraminífers, pel qual, a partir d’una única cèl·lula mare (esquizont o meront, a l'esquerra) s’obtenen nombroses cèl·lules filles (esquizozoïts o merozoïts, a la dreta).
Biopunt, a partir de fonts diverses
Una altra mena de reproducció asexual és l’escissió múltiple, procés durant el qual la cèl·lula es divideix en diversos descendents, de vegades en un nombre molt elevat. En aquest procés, també anomenat esquizogònia o merogònia, la cel·lula que està per dividir-se ( esquizont o meront) multiplica diverses vegades el seu nucli, de manera que al final es converteix en una cèl·lula polinucleada. Al voltant de cadascun dels nuclis es diferencia una part de citoplasma acompanyat dels orgànuls imprescindibles (mitocondris, reticle endoplasmàtic) i els diferents individus així formats (esquizozoïts, merozoïts) s’emancipen de la cèl·lula mare, de la qual generalment no queda més que una escassa part de citoplasma residual inviable. Aquest tipus de reproducció asexual múltiple es dona sobretot en els esporozous i en els foraminífers.
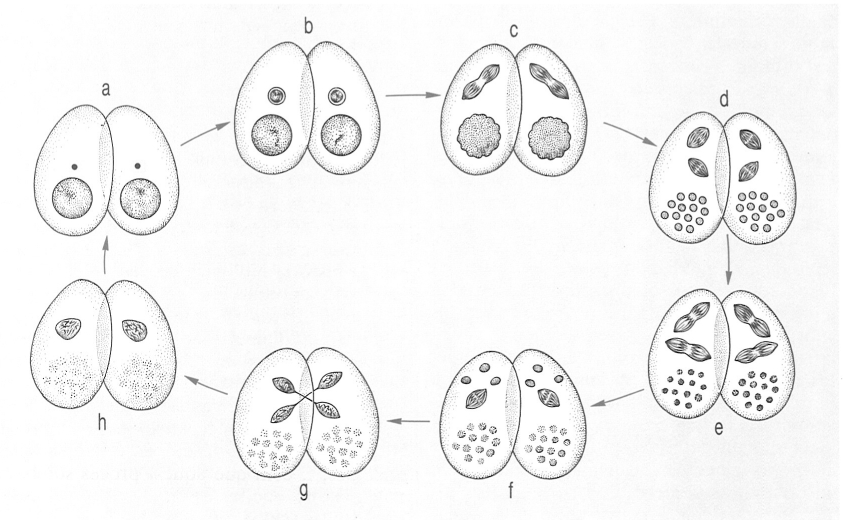
La conjugació és un tipus de reproducció sexual peculiar dels ciliats, un procés en el qual no hi ha una unió completa de gàmetes sinó solament una fecundació creuada entre dos individus (conjugants), cadascun dels quals es comporta després com un zigot. El procés s’inicia amb un creixement del macronucli (a), el qual acaba desapareixent per reabsorció; entretant, el micronucli es divideix dues vegades (c-e) i ocorre la reducció corresponent del nombre de cromosomes. Així, es generen quatre nuclis fills, dels quals tres es reabsorbeixen i el darrer es torna a dividir (f). Aquesta divisió dona dos nuclis gamètics haploides, dels quals un resta dins l’individu mare original (nucli estacionari) i l’altre emigra a l’altre conjugant (nucli migrador); dins de cada cèl·lula es fusionen el nucli estacionari de l’una amb el nucli migrador de l’altra i formen un sincàrion diploide (h). Acabat aquest procés, els dos individus exconjugants se separen i cadascun es divideix mitòticament en dos nuclis, dels quals l’un es convertirà en micronucli i l’altre en macronucli.
Biopunt, a partir de fonts diverses
Els processos reproductors sexuals es produeixen en molts grups de protozous i fins i tot és possible que la sexualitat sigui present en totes les espècies. Com en la resta dels éssers vius, el procés sexual comporta una fecundació o unió de les dotacions genètiques de dos individus diferents. De vegades aquesta fecundació significa, com en els animals o les plantes pluricel·lulars, la unió completa de les dues cèl·lules portadores dels genomes (gàmetes), que, en molts casos (la majoria dels esporozous), són de morfologia diferent; en aquests casos es tracta d’una veritable còpula, en la qual la cèl·lula resultant o zigot reuneix els genomes d’ambdós gàmetes. En d’altres casos no hi ha una unió de gàmetes: és la conjugació dels ciliats, en la qual els protagonistes del procés sexual (conjugants) s’uneixen temporalment i s’intercanvien material genètic, de manera que es produeix una fecundació creuada; quan els conjugants se separen, cadascun es comporta com un zigot i, per tant, no hi ha unió de dues cèl·lules, sinó exclusivament un intercanvi genètic entre elles dues.
Els cicles biològics
Hi ha alguns protozous, bé que no gaires, en els quals la reproducció és exclusivament asexual, de manera que un individu dona naixement a d’altres, exactament iguals, que donaran descendents també idèntics. En aquest cas, la història biològica de l’espècie és summament monòtona, ja que es redueix a la repetició indefinida d’un mateix model biològic; és el cas de l’ameba de les basses Amoeba proteus, en la qual no s’ha observat fins a l’actualitat cap altre sistema de reproducció que l’escissió binària. El cas més general, però, és que al llarg de les successives generacions de la mateixa espècie s’intercalin diferents tipus biològics, i, com que aquest procés sol ésser cíclic, es diu que les espècies tenen cadascuna el seu corresponent cicle biològic.
Una primera complicació apareix quan s’intercala, en la successió de formes idèntiques sorgides per reproducció asexual, una forma de resistència anomenada cist. El cist neix quan una forma vegetativa, obeint a un estímul determinat, passa per uns canvis fisiològics i morfològics i secreta una membrana d’estructura complexa relativament resistent a la dessecació (membrana cística), que tanca a dins el protozou en un estat especial de vida latent anomenat criptobiosi; quan es tornen a restablir en l’ambient les condicions adequades de vida, el cist s’obre i deixa sortir el mateix individu que es va encistar, que ha recuperat la seva viabilitat. Aquest és el cas, per exemple, de l’ameba paràsita (no patògena), de l’intestí de l’home lodamoeba buetschlii, i també el de molts ciliats. D’altres protozous tenen un cicle una mica més complex, com l’ameba patògena de l’intestí humà Entamoeba histolytica, o el ciliat Colpoda cucullus, en els quals el cist no solament és de resistència sinó també de reproducció, perquè el protozou s’hi reprodueix a dins, de manera que al moment d’obrir-se el cist surten quatre petites amebes o quatre petits ciliats, respectivament.
Però el més normal és que en els cicles biològics es produeixin fenòmens de sexualitat que acompanyin la reproducció asexual, cosa que fa que els cicles siguin realment molt més complicats i donin lloc a moltes més formes. Per a comprendre’ls adequadament, cal recordar que s’hi produeixen dos processos d’una importància excepcional des del punt de vista genètic: la fecundació i la meiosi. Aquests dos processos determinen l’existència de cèl·lules haploides i diploides o bé, dit d’una altra manera, la distribució del cicle biològic en dues fases: la fase diploide, que comença amb la fecundació i acaba amb la meiosi, i la fase haploide, que comença quan la meiosi redueix a la meitat el nombre de cromosomes i acaba quan la fecundació restableix la doble dotació cromosòmica. Ara bé, la durada relativa de les fases haploide i diploide varia molt d’uns cicles a altres. Els tres cicles més clars són el dels ciliats, el dels foraminífers i el dels esporozous.
Els cicles biològics dels ciliats
Els dos nuclis de la cèl·lula del ciliat, el macronucli i el micronucli, es divideixen en l’escissió binària habitual: el macronucli, poliploide, ho fa per amitosi, i el micronucli, diploide, ho fa per mitosi; així, a través de generacions successives asexuals es va desenvolupant una població que constitueix la fase diploide del cicle. En un moment donat, per unes determinades circumstàncies fisiològiques i ambientals, dos individus de la fase diploide fan una conjugació: el micronucli es divideix diverses vegades, una de les quals és una meiosi, desapareixen diversos productes, i el resultat és l’aparició en cada conjugant de dos micronuclis haploides (pronuclis o nuclis gamètics), mentre que el macronucli desapareix. Aquests nuclis gamètics representen la fase haploide del cicle, ja que immediatament es restableix l’estat diploide: dels dos pronuclis de cada conjugant, un (el pronucli emigrant) passa a l’altre conjugant per unir-se a l’altre pronucli (pronucli estacionari). La conseqüència d’aquesta doble fecundació és que en separar-se immediatament els dos individus, cadascun té un únic nucli diploide, el qual, per un variat procés de divisions, dona al final la dotació típica del ciliat, amb un micronucli diploide i un macronucli poliploide. Per tant, en els ciliats, la meiosi és gamètica (es fa immediatament abans de la fecundació), la fase haploide és molt curta (afecta exclusivament els nuclis gamètics) i la diploide, molt llarga (dura pràcticament tot el cicle).
Els cicles biològics del foraminífers
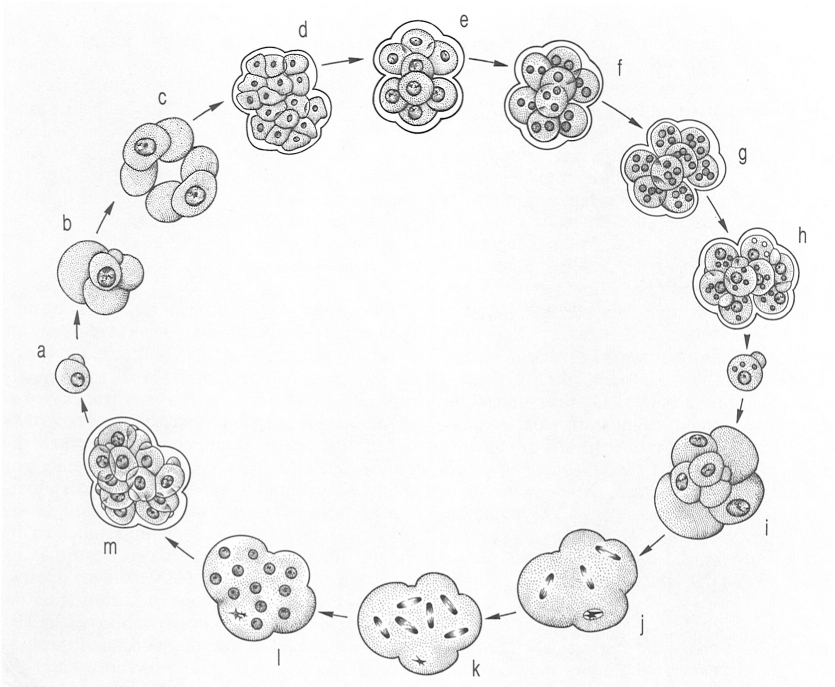
Cicle biològic dels foraminífers. Generalment, comprèn una fase haploide, o gamònia (en groc, en el dibuix), i una fase diploide o agamònia (en taronja, en el dibuix). El del dibuix correspon a Metarotaliella parva. En la fase haploide, els gamonts (b), uninucleats, s’aparellen (c) i formen, per divisió múltiple, gàmetes (d) els quals finalment donen lloc als zigots uninucleats (e). Cada zigot divideix el seu nucli dues vegades (f-h) fins a convertir-se en agamont adult o esquizont (i) diploide. Tres dels seus quatre nuclis donen, per meiosi, 12 nuclis haploides, mentre que el quart comença a desaparèixer (j-l); després, per esquizogònia (m), l’agamont es divideix en 12 embrions haploides (a).
Biopunt, a partir de Weber, 1965
En els foraminífers, el cicle encara que és variable segons les espècies, sol comprendre formes diploides i formes haploides que es van alternant. En l’espècie Elphidium crispum, per exemple, hi ha una forma diploide, l’anomenat agamont o esquizont, que al principi només té un sol nucli, però que al llarg de la seva vida el multiplica per mitosi i, quan ateny la maduresa, ja té un estat plurinucleat; llavors, l’agamont desenvolupa a cadascun dels seus nuclis una meiosi, que els converteix en haploides, i seguidament passa per un procés d’esquizogònia pel qual es disgrega en una multitud de petits organismes haploides, els embrions, semblants al seu progenitor. Cadascun d’aquests embrions haploides creix fins a convertir-se en un gamont, organisme semblant a l’esquizont però uninucleat i haploide. En la maduresa del gamont es produeix una altra divisió múltiple, que produeix uns petits gàmetes flagel·lats. Finalment, la unió d’aquests gàmetes dona origen a un zigot que creix fins a convertir-se en un agamont diploide, que al principi és uninucleat. Tot i que no sempre en els foraminífers es produeix aquesta alternança de generacions tan esquemàtica, sempre hi ha unes formes haploides i unes altres de diploides, ja que en el seu cicle biològic els processos de fecundació i la meiosi estan molt allunyats.
Els cicles biològics del esporozous
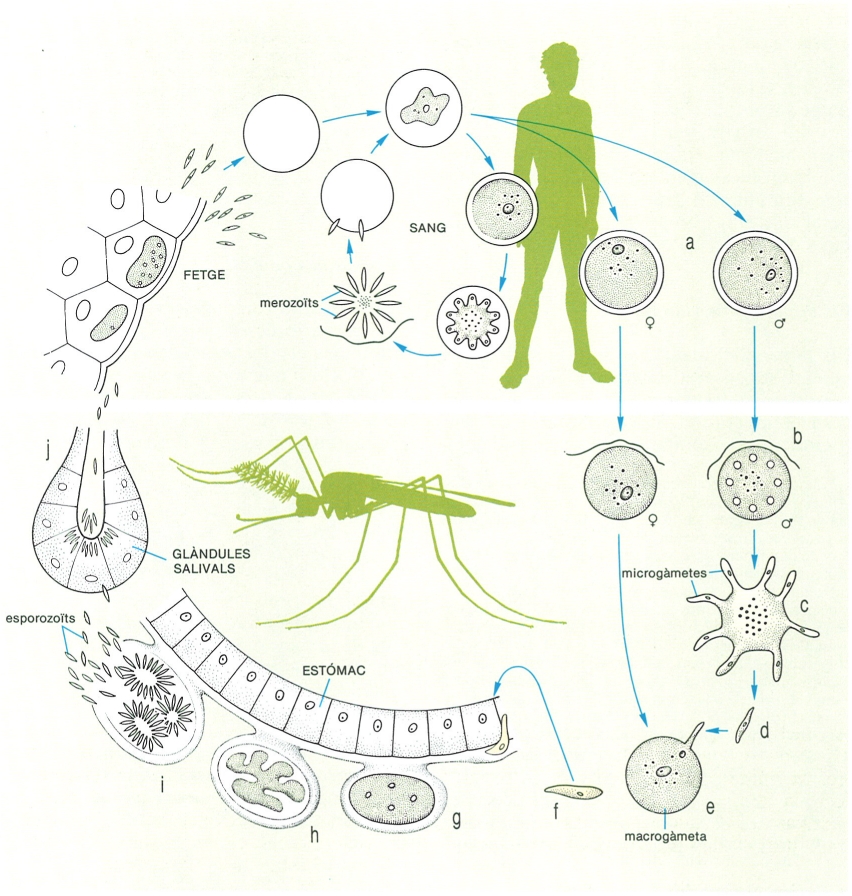
Cicle biològic de l’esporozou Plasmodium vivax, agent del paludisme, el desenvolupament del qual passa per l’alternança de generacions i l’alternança d’hostes. Els gamonts masculins i femenins (a) formats dins els eritròcits de la sang de l’home, hi poden restar durant diverses setmanes. Quan passen al mosquit, penetren al seu estómac, i allà el gamont femení es converteix en macrogàmeta, mentre que el masculí divideix tres vegades el seu nucli i dona lloc a vuit microgàmetes (b-d). Aquests s’uniran (e) formant un zigot mòbil o oocinet (f). Dins el mosquit, aquest zigot travessa la paret de l’estómac a l’interior del qual forma esporocists (g-i), els quals donen lloc, per divisió, als esporozoïts. Aquests esporozoïts s’escampen pel cos del mosquit, i concretament els que van a parar a les glàndules salivals (j) són els que poden passar a l’home. Quan això s’esdevé, s’obren pas fins a les cèl·lules del fetge, on es multipliquen ràpidament i penetren després dins els glòbuls rojos. Totes les fases són haploides, excepte la de zigot, que és diploide.
Biopunt, a partir de fonts diverses
En els esporozous, la meiosi és zigòtica i, per tant, totes les formes del cicle biològic són haploides excepte el zigot, que és diploide. Però en aquest cas hi ha diversos processos esquizogònics i, per tant, les formes vegetatives poden ésser variades; a més, els esporozous són tots paràsits i unes formes poden aparèixer en un hoste i unes altres en un altre, cosa que complica i diversifica molt els cicles.
Hom es referirà únicament al cicle d’un dels agents del paludisme, Plasmodium vivax. El cicle s’inicia quan un mosquit infectat (una femella d’Anopheles) pica una persona. El mosquit infectat té a les glàndules salivals una multitud d’individus d’una fase diminuta del plasmodi, l’esporozoït, que són introduïts en la sang de la persona. L’esporozoït, com totes les altres formes excepte el zigot, és haploide, i penetra en una cèl·lula del fetge, on va creixent fins que es converteix en una altra forma, l’esquizont, que després es divideix per un altre procés esquizogònic en una gran quantitat de merozoïts o esquizozoïts; aquests són els que penetren un glòbul roig, dins el qual creixen fins a convertir-se també en esquizonts, que passen una altra esquizogònia que dona lloc novament als merozoïts. Aquest procés es pot repetir unes quantes vegades, de manera que generacions successives de merozoïts nascuts als glòbuls roigs van infectant-ne cada vegada més. Alguns d’aquests merozoïts, no obstant això, no es convertiran en esquizonts sinó en unes formes sexuades que no es reprodueixen i que resten a l’interior dels hematies fins que es moren o fins que un mosquit les reculli i puguin recomençar el seu cicle biològic en l’insecte. Aquestes formes són els gamonts. En l’estómac del mosquit es desenvolupa la fase anomenada gamogònica: els gamonts masculins passen tres mitosis successives i donen lloc a vuit gàmetes masculins cadascun; els gamonts femenins no experimenten canvis nuclears i es limiten a arrodonir-se. Cal fer notar que, com que els gamonts ja són haploides, els gàmetes es formen sense que hi hagi prèviament una meiosi. També dins el mosquit, s’esdevé la fecundació, que dona origen a un zigot diploide i mòbil, anomenat oocinet, el nucli del qual es divideix per meiosi i després per mitosi fins a donar, per un altre procés d’escissió múltiple, una multitud d’esporozoïts. Cal dir que en el cicle biològic de la majoria dels esporozous, moltes de les formes són protegides per cobertes cístiques, com passa amb els esporozoïts, que gairebé sempre es troben en un nombre reduït dins una càpsula o membrana que rep el nom d’espora, paraula de la qual procedeix el nom d’esporozoït.
Ecologia dels protozous
El fet que els protozous puguin viure en qualsevol medi que contingui una quantitat suficient d’aigua líquida no vol dir que cada espècie no tingui unes exigències característiques quant als factors del medi; efectivament, la majoria mostren una capacitat extraordinària per acomodar-se a diferents medis específics. La seva presència sol ésser constant als medis adequats, ja que, igual que altres microorganismes, com els bacteris, els protozous són ubics. A grans trets es poden considerar dos grans grups de protozous: els que fan vida lliure i els que són paràsits.
Els protozous de vida lliure
Els factors del medi que influeixen decisivament sobre els protozous de vida lliure són diversos, però destaquen per la seva importància la temperatura, la concentració de sals, l’acidesa i la quantitat i qualitat de l’aliment.
La majoria dels protozous únicament poden viure dins un marge estret de temperatures, bé que en estat de cist poden resistir temperatures extremes; el límit inferior és la temperatura de congelació del citoplasma, i el superior, la que inactiva els enzims responsables del metabolisme de l’organisme. D’altra banda, la tolerància de les variacions de temperatura no és la mateixa en totes les espècies, ni tampoc dins una mateixa espècie, ja que depèn de la resta de factors del medi: l’acidesa, en general, disminueix la tolerància i l’alcalinitat l’augmenta; passa el mateix amb la presència d’aliments: s’ha observat que alguns ciliats, en presència d’aliment abundant, tenen menys resistència a la temperatura que quan passen gana. Alguns protozous poden resistir temperatures molt extremes, fins al punt que alguns viuen en les aigües termals a temperatures de fins 56°C, i, com s’ha demostrat en experiments, hi ha tripanosomes que reprenen l’activitat normal després d’ésser congelats en nitrogen líquid (– 196°C). A part l’acció directa de la temperatura sobre la velocitat de les reaccions metabòliques, aquesta exerceix una acció indirecta, perquè la solubilitat de l’oxigen en l’aigua varia amb relació inversa a la temperatura.
La concentració de sals en el medi circumdant també exerceix una influència important sobre els protozous lliures, els quals no solen tolerar gaires variacions de la salinitat. Això vol dir que la majoria de les espècies són o bé d’aigua dolça o bé d’aigua marina, però també n’hi ha algunes que poden viure indiferentment en una o en l’altra. En el laboratori s’ha aconseguit habituar una espècie d’aigua dolça a concentracions creixents de sal i sembla que el vacúol contràctil fa un paper molt important en aquesta adaptació, ja que el seu ritme de contracció disminueix gradualment a mesura que s’eleva la concentració de sal fins que el mateix vacúol arriba a desaparèixer, quan s’ha arribat a un nivell de salinitat molt elevat; el fet que els protozous marins no tinguin vacúol contràctil dona suport a la hipòtesi de la seva naturalesa osmoreguladora.
L’acidesa o pH també és important, i, bé que hi ha protozous que toleren un ventall de variació del pH de l’ambient força ampli, la majoria prefereixen uns valors determinats, generalment pròxims a la neutralitat o lleugerament més àcids.
La quantitat i la qualitat d’aliment influeixen, òbviament, en el creixement i la multiplicació dels protozous. Com que, al seu torn, l’aliment sol dependre de les característiques físiques i químiques del medi, aquestes defineixen en gran mesura la densitat de protozous. Per exemple, en les aigües molt contaminades de matèria orgànica, que són extraordinàriament riques en bacteris, els protozous bacteriòfags (amebes, alguns flagel·lats i molts ciliats) són molt nombrosos; en canvi, en les aigües pobres de matèria orgànica però relativament riques de sals minerals i ben il·luminades, on es reprodueixen abundants algues unicel·lulars, els més abundants són els ciliats fitòfags. Això fa que en una massa d’aigua gran, com pot ésser un riu o un llac, la distribució de les poblacions de protozous sigui molt heterogènia, ja que per als protozous hi ha una gran quantitat de microambients diferents des del punt de vista de l’alimentació: les espècies presents i llurs densitats poden ésser molt variables a diferents llocs d’una mateixa massa d’aigua.
Els protozous paràsits
Entre els protozous paràsits d’animals hi ha tota una sèrie de situacions de diferent intimitat entre els dos éssers associats, des de la perpetració d’una autèntica agressió amb un perjudici per a l’organisme que allotja el protozou (parasitisme), fins al benefici mutu (mutualisme), passant pel benefici unilateral del protozou, relació en què aquest s’aprofita de l’hoste per a alimentar-se o per a protegirse, però sense fer-li cap mal (comensalisme). Però moltes vegades és difícil de determinar la veritable naturalesa de la relació mútua, per la qual cosa s’utilitza el terme «paràsit» quan el protozou viu a l’interior del cos d’un animal, sense entrar en una anàlisi més detallada de la relació que té amb l’hoste (tret, és clar, que sigui un cas ben conegut d’un altre tipus de relació); aquest és el sentit que hom donarà ací al terme «paràsit».
En l’ecologia dels protozous paràsits hi ha dos fets principals: la necessitat que té el protozou d’arribar a l’interior del cos del seu hoste, i el fet que el medi on s’instal·la és un medi viu.
Per atènyer el cos de l’hoste els protozous es valen de tres sistemes. En primer lloc, poden accedir-hi directament a través de la pell o de les mucoses, un sistema molt difós entre els virus i els bacteris però poc emprat pels protozous: només ho fan els que tenen hostes aquàtics, com per exemple, algun flagel·lats paràsits de la pell o de les brànquies dels peixos. En segon lloc, poden entrar-hi per la boca, sistema que utilitzen els flagel·lats i les amebes intestinals, molt eficaç però que n’exigeix la transformació en cist resistent perquè la sortida del tub digestiu (mecanisme de dispersió per a infestar un altre hoste) els obliga a mantenir-se un temps determinat en un medi diferent, no sempre humit. I finalment, poden accedir-hi per mitjà d’un determinat vector, un animal dins el qual el paràsit completa una part del seu cicle biològic; és l’únic sistema assequible quan el protozou s’ha adaptat a viure a l’interior de cèl·lules o teixits sense comunicació amb l’exterior (com els tripanosomes en el plasma sanguini o els plasmodis en els glòbuls roigs dels vertebrats).
El medi dels protozous paràsits pot ésser fins a un cert punt semblant al dels protozous de vida lliure, com passa en el cas de les amebes intestinals, que es troben en un medi ric en matèria orgànica i en bacteris, amb aigua abundant i poc oxigen, medi relativament semblant a les aigües amb fons sapropèlics; però és diferent per la temperatura, que a l’intestí dels mamífers i els ocells és constant. Els que viuen en la sang o en la intimitat dels teixits i les cèl·lules es troben en un ambient diferent del dels protozous lliures. En primer lloc, es tracta d’un ecosistema molt senzill, que no té més que dos elements: el medi intern de l’hoste i el mateix protozou. En segon lloc, aquest medi, molt complex des del punt de vista químic i de característiques físiques molt constants (temperatura, grau d’humitat, pH, llum, etc.) es troba sotmès a canvis que deuen influir sobre el paràsit (per exemple, l’acció de les hormones o dels mateixos metabòlits d’origen alimentari). Però el que fonamentalment caracteritza el medi dels protozous paràsits és la capacitat de reacció de l’hoste davant la seva presència, és a dir, els fenòmens immunitaris que es desencadenen i que determinen tan decisivament l’èxit o el fracàs d’una invasió parasitària. Finalment, cal destacar el fet que, contra el que passa en els de vida lliure, no es tracta d’organismes cosmopolites sinó que generalment només es troben en unes determinades regions, on coincideixen les distribucions dels hostes i els vectors amb condicions ecològiques adequades.
Evolució i sistemàtica dels protozous
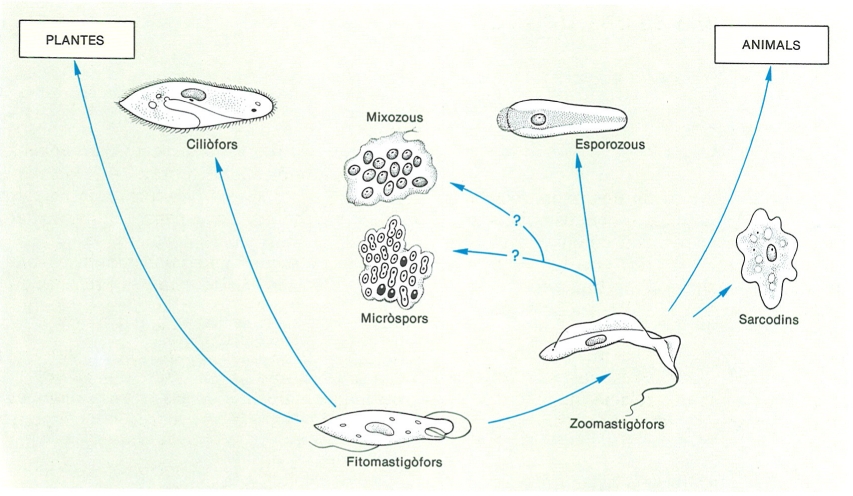
Esquema de les relacions filètiques que relacionen els grans grups de protozous.
Biopunt, a partir de fonts diverses
Pel fet de no formar un grup natural des del punt de vista de l’evolució, els protozous formen un conjunt polifilètic. Es fa difícil de seguir la història dels diferents grups que el formen i de veure les relacions de parentiu que estableixen entre ells. Una primera conseqüència és la dificultat de dissenyar un arbre genealògic raonablement versemblant, i això pot explicar els canvis que hom ha introduït en la seva sistemàtica en les darreres dècades.
Hi ha, però, una circumstància predominant, que permet d’entreveure unes tendències evolutives de caràcter més o menys general, que es presenten en tots els grups de protozous: la unicel·lularitat fonamental que presenten tots. De fet, la naturalesa unicel·lular dels protozous és la causa d’una diferència essencial respecte de les cèl·lules dels metazous, que té una gran trascendència evolutiva. Generalment, la diferenciació dels metazous durant el desenvolupament es produeix perquè les diferents estirps cel·lulars assumeixen unes funcions fisiològiques determinades; en canvi, en els protozous, on cada cèl·lula equival a un organisme complet que assumeix totes les funcions, s’hi diferencien uns orgànuls intracel·lulars que, aquests sí, poden ésser molt especialitzats. Això fa que, mentre que el procés de la diferenciació cel·lular porta, en els metazous, a la formació de teixits i sistemes, en els protozous mena a una gran complicació estructural de l’única cèl·lula que els compon; és a dir, en els primers es produeix una diferenciació «histològica», mentre que en els segons es duu a terme una diferenciació «citològica».
Són dues les principals tendències evolutives que s’observen en els diferents grups de protozous. La més característica i més generalitzada és la multiplicació progressiva de les estructures que en principi són úniques o poc nombroses, procés que es coneix amb el nom de polimerització. Aquest fenomen es manifesta de diferents maneres. En la més senzilla, l’augment del nombre no afecta sinó els orgànuls d’una sola mena, mentre que la resta es queden en nombre singular; això passa, per exemple, en alguns zooflagellats: en els tricomonàdides, el gènere Trichomitus, que té quatre flagels, ha donat origen a Trichomitopsis, que en té cinc, i a Pentatrichomitus, que en té sis. Altres vegades s’observa que el procés de polimerització afecta dos o més orgànuls, però sense que augmenti el nombre de nuclis, i, en aquest cas, la cèl·lula es fa estructuralment molt complexa però no perd la seva individualitat; és el cas d’alguns flagel·lats hipermastígides, que tenen flagels i aparells parabasals abundants però mantenen un sol nucli. Finalment, hi ha molts casos en què la polimerització afecta també el nombre de nuclis i es produeixen els anomenats protozous multinucleats, com passa en les opalines. En aquest cas, sorgeix la segona tendència evolutiva important dels protozous: la tendència a la pluricel·lularitat, que es manifesta en principi en l’augment de dotacions genètiques nuclears, però que pot afectar d’altres aspectes; en el cas dels ciliats, les cèl·lules tenen diverses dotacions genètiques en el macronucli a més de la dotació diploide del micronucli.
Establir quin és l’origen dels protozous és, en el fons, resoldre el problema de l’origen de la cèl·lula eucariòtica, del qual no se sap res amb seguretat. Actualment comença a prendre força la teoria de l’endosimbiosi seqüencial, que situa l’origen de la cèl·lula eucariòtica en l’associació simbiòtica de diversos elements procariòtics. Pel que fa a les relacions filogenètiques dels protozous, l’única teoria generalitzada fins ara és la que suposa que els coanoflagel·lats tenen un origen comú a les esponges; però com que, al seu torn, les esponges són un grup molt aïllat, queda encara per resoldre l’origen de tota la resta dels metazous. Tampoc no es pot assegurar res pel que fa a les relacions filogenètiques entre els diversos grups de protozous, bé que hom ha descrit algunes hipòtesis.
Tradicionalment, i fins fa pocs anys, els protozoòlegs, bé que reconeixien més o menys tàcitament l’origen polifilètic d’aquests organismes, admetien l’existència del fílum dels protozous (Protozoa). Actualment, els protozous no són considerats en conjunt com un fílum sinó com un agregat de set fílums sense sentit filogenètic, que es podria considerar com un subregne del regne dels protists (Protista) o com un subregne del regne dels animals (Animalia). Aquesta proposta de classificació és la que, amb algunes variacions menors, continua vigent en l’actualitat.
L’antic fílum dels protozous havia estat descrit per August Goldfuss el 1818 i defensat més tard (1845) per Ernest von Siebold. Tanmateix, un segle i mig més tard, la classificació que va publicar oficialment el comitè de taxonomia i problemes taxonòmics de la Society of Protozoologists (1964) encara admetia l’existència d’aquest fílum, que, de fet, constitueix una declaració explícita del monofiletisme dels protozous. No obstant això, el 1974, aparegueren dues noves classificacions del fílum dels ciliòfors (Ciliophora), i, el 1980, un altre comitè de la mateixa societat féu pública per primera vegada la classificació que regeix actualment.
Els grans grups de protozous i les seves característiques principals
| CLASSIFICACIÓ DELS PROTOZOUS | PRINCIPALS CARACTERÍSTIQUES DE CADA GRUP | ||||
|---|---|---|---|---|---|
| FÍLUM SARCOMASTIGÒFORS (Sarcomastigophora) | Ampli conjunt de protozous que tenen pseudopodis o flagels, o ambdós tipus d’orgànuls locomotors. | ||||
| Subfílum mastigòfors (Mastigophora) | Flagel·lats proveïts típicament d’un o més flagels. Divisió simetricogènica. | ||||
| Cl. fitomastigoforis (Phytomastigophorea) | Flagel·lats proveïts de pigments fotosintètics. | ||||
| Cl. zoomastigoforis (Zoomastigophorea) | Grup polifilètic de flagel·lats proveïts d’un nombre variable de flagels (des d’un fins a una gran quantitat). Nutrició holozoica. | ||||
| O. coanoflagèl·lides (Coanoflagellida) | Flagel·lats amb un sol flagel, envoltat d’un collar citoplasmàtic tènue. Ex: Monosiga. | ||||
| O. diplomonàdides (Diplomonadida) | Flagel·lats «dobles», amb dos nuclis i una dotació de flagels doble. Paràsits. Ex: Giardia. | ||||
| O. tricomonàdides (Trichomonadida) | Amb diversos flagels anteriors i un de recurrent amb membrana ondulant. Paràsits. Ex: Trichomonas. | ||||
| O. hipermastígides (Hipermastigida) | Molts flagels. Simbionts de l’intestí dels tèrmits. Ex: Trichonympha. | ||||
| O. cinetoplàstides (Kinetoplastida) | Un o dos flagels. Generalment membrana ondulant. Lliures o paràsits. Ex: Trypanosoma. | ||||
| O. opalínides (Opalinida) | Molts flagels. Generalment, molts nuclis. Ex: Opalina. | ||||
| Subfílum sarcodins (Sarcodina) | Tenen pseudopodis. De vegades poden tenir flagels en alguna fase del seu cicle biològic. | ||||
| SpCI. rizòpodes (Rhizopoda) | Tenen pseudopodis no distribuïts simètricament. | ||||
| Cl. lobosis (Lobosea) | Pseudopodis lobats (lobopodis). | ||||
| SCI. gimnamebis (Gymnamoebia) | Amebes nues. Sense esquelet. Ex: Amoeba. | ||||
| SCI. testaceolobosos (Testaceolobosa) | Amebes amb testa o closca. Ex: Arcella. | ||||
| Cl. filosis (Filosea) | Pseudopodis fil·losos (fil·lopodis). Ex: Euglypha. | ||||
| Cl. granuloreticulosis (Granuloretículosea) | Són els foraminífers, amb pseudopodis granuloreticulosos, closca generalment proveïda de diverses cambres i perforada per a donar sortida als pseudopodis. Ex: Elphidium. | ||||
| Cl. micetozoïs (Mycetozoea) | Són els micetozous o mixomicets. Són petites amebes que s’aglomeren en plasmodis amb cossos fructífers. | ||||
| SpCI. actinòpodes (Actinopoda) | Pseudopodis distribuïts simètricament. Gairebé sempre tenen esquelet inorgànic. | ||||
| Cl. acantaris (Acantharea) | Esquelet de sulfat d’estronci, amb espícules disposades segons una llei. Marins. Ex: Acanthometron. | ||||
| Cl. radiolaris (Radiolaria) | Esquelet silici, amb formes variables. Marins. Ex: Astracantha. | ||||
| Cl. heliozoïs (Heliozoea) | Generalment sense esquelet. Axopodis abundants. D’aigua dolça, excepcionalment marins. Ex: Actinosphaerium. | ||||
| FÍLUM ESPOROZOUS (Sporozoa) | Protozous tots paràsits, que tenen a l’extrem anterior un complex apical, només visible al microscopi electrònic. Reproducció sexual i asexual, aquesta darrera, múltiple. Cicles biològics complicats, haploides, amb meiosi cigòtica. | ||||
| Cl. gregarinis (Gregarínea) | Paràsits del tracte digestiu o de la cavitat general del cos d’invertebrats. Gamonts grossos. Gàmetes masculins i femenins semblants. Les formes infectants (esporozoïts) es formen a dins d’espores resistents. Ex: Monocystis. |
||||
| Cl. coccidis (Coccidea) | Paràsits d’invertebrats o de vertebrats. Gamonts petits, intracel·lulars. Gàmetes masculins petits i mòbils; els femenins, grossos, es formen a cada gamont. Esporozoïts dins d’espores. Ex: Eimeria. | ||||
| Cl. hemosporidis (Haemosporiidea) | Paràsits que desenvolupen el seu cicle a la sang d’un vertebrat i tenen un artròpode vector. Gamonts petits, dins dels hematies. Gàmetes masculins escassos; els femenins, un per gamont. Sense espores. Ex: Plasmodium, Theileria. | ||||
| FÍLUM MICRÒSPORS (Microspora) | Protozous paràsits intracel·lulars de tots els grups d’animals. Cicle biològic que comprèn una espora, amb un filament espiral evaginable. Ex: Nosema. | ||||
| FÍLUM MIXOZOUS (Myxozoa) | Paràsits de molts grups d’animals. Espores d’origen pluricel·lular, amb diverses càpsules polars cadascuna, amb un filament espiral evaginable. Ex: Ceratomyxa. | ||||
| FÍLUM CILIÒFORS (Ciliophora) | Protozous dotats de cilis agrupats en cinèties. Amb micronucli diploide i macronucli poliploide. Reproducció asexual per escissió homotetogènica. Transferència genètica per conjugació. Lliures o paràsits. | ||||
| Cl. cariorelictis (Karyorelictea) | Són els ciliats més antics, amb macronuclis que no es divideixen. Ex: Loxodes. | ||||
| Cl. espirotriquis (Spirotrichea) | La ciliació especialitzada de la boca és una «zona adoral de membranel·les» (ZAM), composta d’unes formacions de tres o quatre files de cilis que es disposen en espiral vers l’orifici bucal. | ||||
| SCI. heterotriquis (Heterotrichia) | La ciliació somàtica es compon de cinèties de cilis senzills. Ex: Stentor. | ||||
| SCI. coreotrlquis (Choreotrichia) | Marins, amb lloriga. Ex: Tintinnus. | ||||
| SCI. esticotrlquis (Stichotrichia) | La ciliació somàtica és formada per cirrus, que són conjunts compactes de cilis. Ex: Oxytricha. | ||||
| Cl. prostomacis (Prostomatea) | Boca anterior, sense una ciliació especialitzada. Ciliació somàtica senzilla. Ex: Coleps. | ||||
| Cl. litostomacis (Litostomatea) | Boca generalment lateral, dilatable o amb ciliació bucal de sincilis formats per nombrosíssimes cinèties paral·leles. Ciliació somàtica senzilla. | ||||
| SCI. haptoris (Haptoria) | Boca terminal o lateral, dilatable, amb toxocists. Carnívors. Ex: Litonotus. | ||||
| SCI. tricostomacis (Trichostomatia) | Boca amb sincilis. Paràsits o simbionts de l’aparell digestiu de vertebrats. Ex: Ophryoscolex. | ||||
| Cl. fil·lofaringis (Phyllopharingea) | Boca amb una petita nansa faríngia composta d’unes barnilles cargolades fent espiral, o sense boca. Ciliació somàtica més o menys reduïda. | ||||
| SCI. fil·lofaringis (Phyllopharingia) | Ciliació somàtica reduïda a la cara dorsal. Boca a la cara ventral. Nedadors. Ex: Chilodonella. | ||||
| SCI. conotriquis (Chonotrichia) | Ciliació somàtica molt reduïda a tot el cos. Boca al fons d’un embut. Sedentaris. Ex: Spirochona. | ||||
| SCI. suctoris (Suctoria) | Sedentaris. Ciliació absent a l’adult fix i solament present a la «larva» nedadora. Sense boca. Nutrició per mitjà d’uns tentacles xucladors. Ex: Acineta. | ||||
| Cl. nassoforis (Nassophorea) | Boca amb una nansa grossa i recta o amb nemadesmes o amb una ZAM. | ||||
| SCI. nassoforis (Nassophoria) | Nansa, quan n’hi ha, grossa i recta, i com a mínim, nemadesmes a la boca. Ex: Nassula. | ||||
| SCI. hipotriquis (Hypotrichia) | Boca amb ZAM i ciliació somàtica només a la cara ventral, formada per cirrus. Ex: Euplotes. | ||||
| Cl. oligohimenoforis (Oligohymenophorea) | Ciliació bucal formada per tres membranel·les petites a l’esquerra de la boca i una llarga «membrana paroral» a la dreta. | ||||
| SCI. himenostomacis (Hymenostomatia) | Ciliació somàtica ben desenvolupada. Ex: Colpidium. | ||||
| SCI. peritriquis (Peritrichia) | Ciliació somàtica molt reduïda. Ciliació bucal molt aparent al voltant de la boca. Generalment, sedentaris, fixats per un peduncle que pot ser contráctil. Ex: Vorticella. | ||||
| SCI. astomacis (Astomatia) | Sense boca ni ciliació bucal. Ciliació somàtica ben desenvolupada. Paràsits de l’intestí d’oligoquets. Ex: Anoplophrya. | ||||
| Cl. colpodis (Colpodea) | Ciliació bucal composta de dos «polibraquicinèties» formades per nombroses cinèties paral·leles. Ciliació somàtica ben desenvolupada. Ex: Colpoda. |
Els grans grups de protozous
Els flagel·lats
Els coanoflagèl·lides sòn uns petits flagel·lats marins o d’aigua dolςa, sedentaris, fixos, que viuen de vegades formant colònies, i que tenen un sol flagel anterior envoltat d’un «collar» delicat, compost d’uns finíssims tentacles anastomosats entre ells. Les espècies marines poden tenir lloriga. La semblança del seu collar i el seu flagel amb les mateixes estructures dels coanòcits de les esponges ha fet pensar que hi tinguin una relació filogenètica. En sòn dos gèneres característics Monosiga i Codosiga.
Els diplomonàdides són paràsits que tenen gairebé tots els orgànuls duplicats i adquireixen l’aspecte d’organismes dobles. El més ben conegut, Giardia intestinalis, és un paràsit freqüent en l’intestí humà i de vegades pot produir greus transtorns intestinals (giardiasi); presenta dos grups de tres flagels, dos nuclis i una ventosa ventral per mitjà de la qual s’adhereix a la mucosa intestinal.
Els tricomonàdides són paràsits intestinals o d’altres òrgans de molts animals, invertebrats o vertebrats. Els mès típics són piriformes, amb un pinzell de diversos flagels dirigits endavant i un flagel sol dirigit enrere que aixeca un plec de la pel·lícula (membrana ondulant). També té un eix antero-posterior de naturalesa microtubular, l’axòstil. Trichomonas vaginalis és una espècie simbiont, no patógena, de la vagina femenina humana; en canvi, una altra de molt semblant, paràsita del tracte genital del bestiar boví, Tritichomonas foetus, és l’agent patogen de l’avortament epizoòtic de les vaques. També és patogen Histomonas meleagridis, paràsit de l’aparell digestiu dels paons, als quals provoca una malaltia greu anomenada «blackhead». Però la majoria són comensals de l’intestí de molts animals; fins i tot en els cas dels tèrmits, hi ha certs tricomonàdides (al costat dels hipermastígides) mutualistes, que digereixen per als seus hostes la fusta que aquests no poden degradar.
Els hipermastígides són flagel·lats simbionts de tèrmits i altres insectes que mengen fusta. Tenen molts flagels pero un sol nucli, i són de grans dimensions.
Els cinetoplàstides són flagel·lats petits, amb un orgànul mitocondrial especial, el cinetoplast, molt ric de DNA. Hom considera l’existència de dos grans grups: els bodònides (per exemple, Bodo saltans), la majoria dels quals són lliures i tenen dos flagels, i els tripanosomàtidcs, amb un sol flagel, paràsits i patògens d’un gran nombre de vertebrats. Aquests darrers són polimorfs; les seves formes principals són l’amastigot, arrodonit i amb un flagel molt curt, i el tripomastigot, lanceolat i amb un flagel dirigit endavant i unit a una membrana ondulant. El gènere Leishmania es presenta en forma d’amastigot i és paràsit de gossos i humans, als quals produeix la malaltia del kalaazar i el botó d’Orient. El gènere Trypanosoma és paràsit de bòvids i humans i produeix la greu malaltia de la son; els tripanosomes es transmeten als mamífers a través de dípters que s’alimenten de sang —generalment del gènere Glossina (mosca tse-tse)— en els quals es troben en forma de tripomastigot.
Els opalínides són protozous amb molts flagels situats en llargues files longitudinals. En alguns gèneres (Protoopalina) hi ha dos nuclis i en els altres (Opalina, Cepedea) els nuclis són nombrosos. Les opalines habiten quasi exclusivament la cloaca dels amfibis anurs.
Els rizòpodes


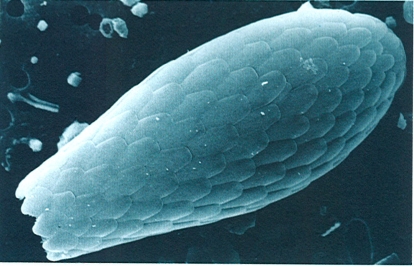
Els rizòpodes més característics són les amebes, que formen un gran grup d’espècies d’una alta diversitat ecològica, totes capaces de produir pseudopodis. N’hi ha de desproveïdes de teca, com la de la fotografia (Amoeba), i n’hi ha de tecades. La de la fotografia viu en les aigües dolces. Els foraminífers són importants rizòpodes marins, posseïdors d’abundants reticulopodis i d’una característica conquilla calcària perforada. La fotografia, feta al microscopi electrònic d’escandallatge, correspon a la conquilla de l’espècie Elphidium macellum.
Miquel Blasi
Les amebes nues tenen lobopodis, però no tenen testa. Habiten en l’aigua i al sòl, on són molt abundants (Mayorella, Acanthamoeba, Chaos), però algunes (Entamoeba) són paràsites de l’intestí de mamífers i poden ésser patògenes. Entre les amebes testàcies, és a dir, proveïdes de testa, n’hi ha que tenen lobopodis (Arcella, Difflugia) i d’altres que tenen fil·lopodis (Euglypha); són freqüents en l’aigua, el fang i al sòl.
Els foraminífers tenen reticulopodis molt llargs, ramificats i anastomosats, que formen en conjunt una xarxa extensa amb la qual capturen les preses. La seva closca té una sola cambra o diverses cambres, comunicades per fines perforacions (foramina), a través de les quals surten els pseudopodis. Tenen uns cicles biològics complicats, en els quals s’alternen les formes haploides (gamonts) i les diploides (agamonts). La gran majoria dels foraminífers (Globigerina, Elphidium, Discorbis, etc.) són politalàmics, és a dir, tenen diverses cambres a la closca, i són marins.
Els actinòpodes
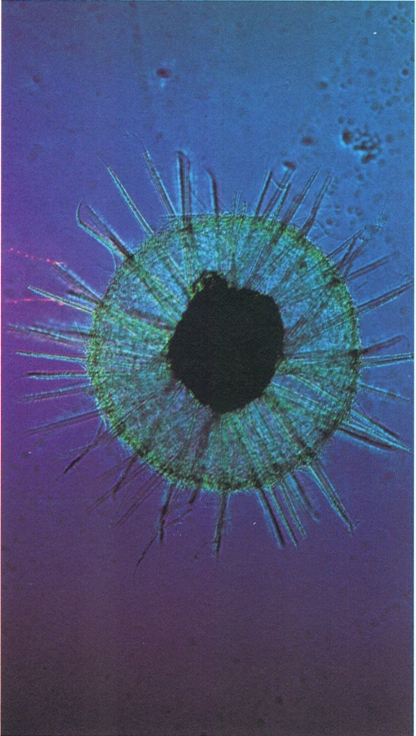
Dins els actinòpodes, destaca el grup marí dels radiolaris, proveïts d’un esquelet silici molt complex. La fotografia correspon al gènere Aulacantha.
Claude Carré
Els acantaris, el primer gran grup d’actinòpodes, tenen simetria radial, amb un endoplasma granular que conté el nucli i un ectoplasma vacuolar amb reticulopodis, fil·lopodis i axopodis. La majoria tenen una membrana esfèrica, la càpsula central, que separa les dues zones del citoplasma. Estan dotats d’un esquelet axial format per deu espícules de sulfat d’estronci, regularment espaiades, que es creuen al centre i travessen la càpsula central; algunes espècies tenen vint espícules que es troben al centre de l’organisme. Són marins i fagòtrofs, però al seu ectoplasma quasi sempre hi ha abundants zooxantel·les simbionts. Una espècie molt coneguda és Acanthometron elasticum.
Els radiolaris també tenen simetria radial; l’endoplasma i l’ectoplasma són separats per una càpsula central, que en aquest cas és més consistent i perforada. Tenen fil·lopodis i axopodis, i l’esquelet silici que presenten és molt variat, generalment molt complex, format per espícules radials (que no s’uneixen al centre), espícules tangencials o esferes buides concèntriques i amb perforacions de diverses formes. Són marins i la majoria conviuen amb zooxantel·les al seu ectoplasma. La majoria dels gèneres són solitaris (Phaeodina, Aulacantha), però també n’hi ha de colonials (Collozoum).
Els heliozoïs són actinòpodes molt petits; la majoria són d’aigua dolça, encara que alguns són marins. No tenen càpsula central i els seus pseudopodis són axopodis que neixen d’un grànul central o del nucli i poden tenir flagels. L’esquelet és inconstant i pot ésser silici o orgànic, format per escates tangencials o una càpsula perforada. Són planctònics o sedentaris, i, en aquest cas, es fixen per un peduncle. De vegades tenen zooxantel·les. n’és un bon exemple Actinophrys sol.
Els esporozous
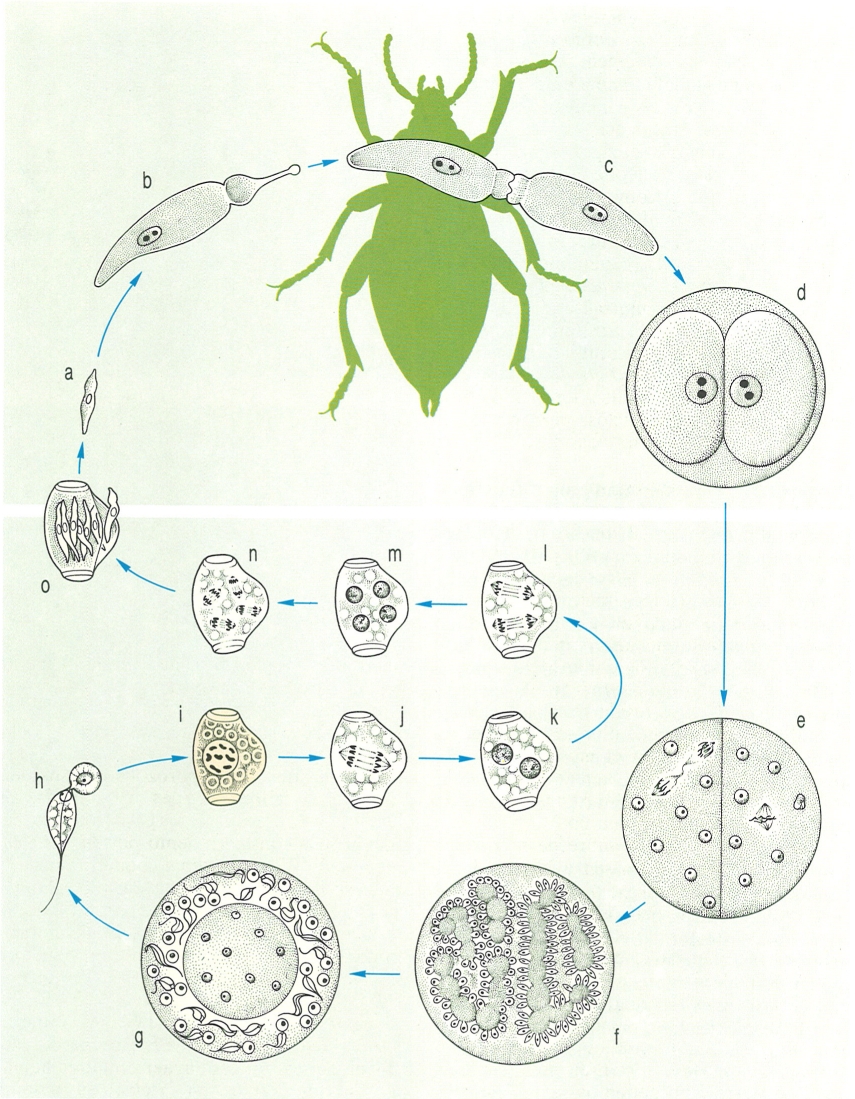
Els esporozous formen un extensíssim grup de protozous paràsits, de cicles biològics complexos i especialitzats. Hom en considera dos grups: les gregarines, paràsits d’invertebrats, en les quals tant els gamonts masculins com els femenins donen lloc a molts gàmetes; i els coccidis, paràsits d’invertebrats o de vertebrats, el gamont femení dels quals dona lloc a un sol gàmeta. L’esquema representa el cicle de la gregarina Stylocephalus longicollis, una part del qual s’esdevé a l’intestí d’un coleòpter tenebriónid (Blaps mortisaga). Els esporozoïts (a) hi penetren amb l’aliment, i es transformen en gamonts (b). Dos d’aquests gamonts s’acosten l’un a l’altre (c) i acaben formant un cist gamòntic (d), que és expulsat a l’exterior amb els excrements de l’escarabat. Un cop fora, dins el cist s’esdevé una multiplicació nuclear (e) i s’inicia la formació del gàmetes masculins i femenins (f-h). Per mitjà de la fertilització es forma el zigot (i), l’única fase diploide del cicle, zigot que inicia posteriorment la reducció del nombre de cromosomes (j) i l’esporogònia (k-n), que dona lloc a 8 esporozoïts (o). Compareu aquest cicle amb el corresponent al coccidi Plasmodium vivax.
Biopunt, a partir de Léger i Grell
Els gregarinis són esporozous paràsits, especialment de l’intestí dels insectes. L’esporozoït creix fora de les cèl·lules intestinals, a l’interior del tub intestinal, però fixat a una cèl·lula, fins que es converteix en un gamont gros, que s’alimenta per pinocitosi dels líquids de l’intestí. Quan ateny la maduresa es deixa anar i s’uneix estretament a un altre gamont, en una associació que rep el nom de sizígia.
Aquesta s’envolta d’una membrana cística, que converteix el conjunt en un gametocist, dins el qual es produeixen els gàmetes, molt nombrosos i d’ambdós sexes, i on es dona la fecundació; després que cada zigot s’ha envoltat d’una membrana esporal, aquest esporocist sofreix una meiosi i unes quantes mitosis, que donen lloc a espores, de les quals neixen generalment quatre esporozoïts. L’esporocist i les espores són una protecció excel·lent per als esporozoïts, que, a l’exterior, esperen l’ocasió d’envair un altre hoste. Gregarina, que viu a l’intestí d’ortòpters, Monocystis, al celoma d’oligoquets, i Stylocephalus, a l’intestí de coleòpters, són gèneres ben coneguts.
Els hemosporidis són esporozous que no fan espores, ja que en cap moment del seu cicle no queden expostos al medi extern, i, per tant, els són innecessàries; són paràsits de les cèl·lules de la sang i d’altres teixits de mamífers i ocells, on arriben gràcies a la intervenció d’artròpodes hematòfags vectors, en els quals es duu a terme una part del seu cicle biològic. El cicle biològic d’un dels causants de la malària, Plasmodium vivax, s’inicia amb els esporozoïts, que resideixen a les glàndules salivals de la femella d’un mosquit del gènere Anopheles. Quan el mosquit pica una persona, els esporozoïts passent al torrent circulatori i després s’introdueixen a les cèl·lules del fetge, on experimenten una esquizogònia que produeix un cert nombre de merozoïts. Aquests són els que passen a la sang i envaeixen els glòbuls roigs, dins els quals passen una altra esquizogònia, que produeix nous merozoïts que penetren a d’altres hematies per recomençar el cicle reproductor esquizogònia Aquesta esquizogònia eritrocítica es repeteix diverses vegades, i després, en un moment determinat, alguns dels merozoïts que han envaït els hematíes, en lloc de multiplicar-se, es converteixen en gamonts, destinats a morir si no poden passar a l’estómac d’un altre mosquit per mitjà d’una picada. A l’estómac del mosquit passa la gamogònia, amb producció de vuit gàmetes masculins i un sol gàmeta femení. Produïda la fecundació dins la cavitat estomacal del mosquit, el zigot, que és mòbil i rep el nom d’oocinet, emigra cap als teixits que envolten l’estómac i allà passa la meiosi i les mitosis subsegüents, que donen lloc a un gran nombre d’esporozoïts; aquests viatgen fins a les glàndules salivals de l’insecte, on resten en espera d’atènyer la sang d’un nou hoste. Els hemosporidis genuïns són del gènere Plasmodium, a més d’unes quantes formes més, que també s’hi relacionen; són també els responsables de la malària de l’home i de la de l’aviram. En l’actualitat també es consideren hemosporidis els esporozous anomenats piroplasmes, dels quals hom parla a continuació.
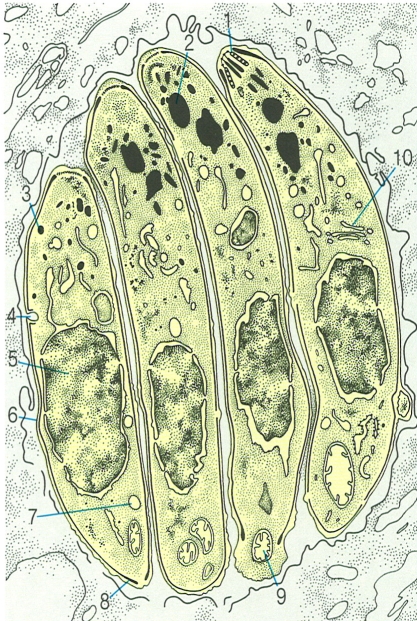
Merozoïts del coccidi Toxoplasma gondiï (en groc), inclosos en una cèl·lula de l’hoste. Mostren els caràcters morfològics típics dels estadis infectius dels esporozous: 1 conoide, 2 roptria, 3 micronema, 4 micròpor, 5 nucli, 6 pel·lícula, 7 grànul d’amilopectina, 8 anell polar, 9 mitocondri, 10 complex de Golgi.
Biopunt, a partir de fonts diverses
Els coccidis són paràsits de l’intestí de nombrosos vertebrats i invertebrats que poden produir malalties greus. L’esporozoït es desenvolupa a l’interior d’una cèl·lula de l’intestí, on es converteix en esquizont; aquest sofreix esquizogònia i dona molts merozoïts, els quals reprodueixen aquest procés una o més vegades i envaixen nombroses cèl·lules intestinals. Alguns merozoïts es converteixen després en gamonts intracel·lulars, que desenvoluparan la gamogònia; aquesta es fa d’una manera diferent en els gamonts de cada sexe: el gamont masculí dona nombrosos gàmetes petits i biflagel·lats i el femení creix fins a convertir-se en un gàmeta femení únic. Alliberats al lumen intestinal, els gàmetes s’ajunten en la fecundació i a continuació es crea la coberta cística que envolta el zigot o oocist. Dins l’oocist, cada zigot s’envolta al seu torn d’una altra coberta que el converteix en esporocist o espora, dins el qual es fa la meiosi i unes quantes mitosis que produeixen un nombre variable d’esporozoïts (generalment entre 2 i 8 dins cada espora). Eimeria tenella és un coccidi que causa molta mortalitat entre els pollastres joves, i E. stiedae també produeix greus epizoòties en els conills domèstics; un altre coccidi, Toxoplasma gondii, és un paràsit molt comú en els teixits d’un gran nombre de mamífers, l’home inclòs, però només és patogen en determinades ocasions.
Els piroplasmes són esporozous paràsits dels hematies i d’altres cèl·lules dels bòvids, als quals arriben a través de les paparres. En l’actualitat se’ls considera hemosporidis. El cicle biològic de Theileria parva comença quan els seus esporozoïts, que s’estaven a les glàndules salivals d’una paparra, penetren a la sang de la vaca. L’esporozoït envaeix el limfòcit, on desenvolupa una esquizogònia productora de molts merozoïts; aquests envaeixen un eritròcit cadascun, dins el qual es multipliquen per un procés semblant a l’escissió binària, que dona dos descendents. Els merozoïts nascuts en aquest procés envaeixen altres hematíes, dins els quals es converteixen en gamonts masculins i femenins. Quan una nimfa de paparra pica l’animal infectat, una part dels gamonts habitants dels eritròcits s’escapen de morir a l’intestí de l’artròpode i passen a través de la paret de l’intestí cap a la cavitat general del cos, on passa la gamogònia, amb la producció de 4 gàmetes masculins i un de femení. Finalment, té lloc la fecundació i la formació d’un oocinet, que emigra fins a les glàndules salivals, on, després de la meiosi i les mitosis consecutives, apareix una multitud d’esporozoïts. Són aquests esporozoïts els que esperen a ésser inoculats a la sang d’un nou animal, emmagatzemats dins les glàndules salivals de la paparra adulta.
Els microsporidis o micròspors
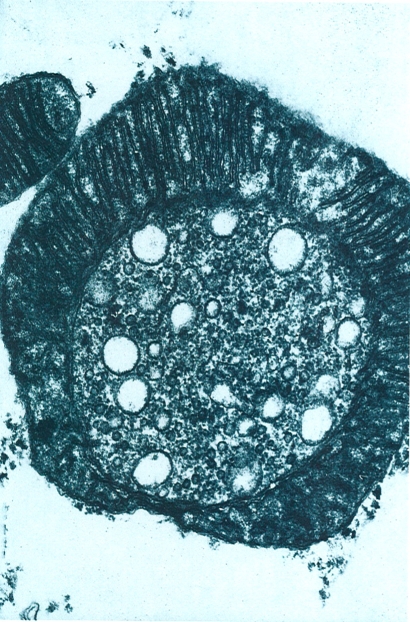
Trofozoït del microsporidi Unicarion myticolae, hiperparàsit de les cèl·lules intestinals del copèpode Myticola intestinalis, embolcallat per un mitocondri.
Mercè Durfort
Són organismes unicel·lulars que mostren escasses afinitats amb els altres grups de protozous. Són molt nombrosos i es troben en molts animals com a paràsits intracel·lulars, mancats de mitocondris. El seu cicle biològic, que encara és desconegut en moltes espècies, parteix d’una forma infectant diminuta (de vegades té un diàmetre inferior a 1 μm), l’ esporoplasma, amb un o dos nuclis. Altres estadis del seu cicle són formes plurinucleades, anomenades esquizonts, i una espora unicel·lular, que és l’estadi de difusió i que conté l’esporoplasma; el més notable d’aquesta espora és que té un llarg filament enrotllat en espiral que s’estén abans de donar sortida a l’esporoplasma dins el cos de l’animal hoste. Nosema bombycis és un microsporidi paràsit dels cucs de seda, als quals causa una malaltia contagiosa anomenada pebrina.
Els mixosporidis o mixozous
Són uns organismes paràsits d’invertebrats i de vertebrats inferiors, que es caracteritzen pel fet de tenir un cicle biològic en el qual hi ha espores d’origen pluricel·lular, amb cèl·lules valvars (que donen origen a les valves) i cèl·lules capsulars (que originen uns elements semblants als nematocists dels cnidaris), proveïdes d’un filament enrotllat en espiral i extensible; aquestes càpsules fixen l’espora als teixits de l’hoste, mentre que les valves protegeixen sobretot el germen infectant, que és un esporoplasma binucleat. En el cicle biològic dels mixosporidis hi ha un estadi plurinucleat, plasmodial, on es diferencien les cèl·lules que formaran l’espora. Es tracta d’un grup amb nombrosíssimes espècies. El gènere Sphaeromyxa en comprèn moltes que parasiten la vesícula biliar de molts peixos. Actualment es comença a dubtar de la seva condició de protozous, perquè s’ha descobert que en el plasmodi de S. sabrazesi es diferencien cèl·lules «somàtiques» de diferents estirps; per aquesta raó és preferible el nom de mixozous al de mixosporidis.
Els ciliats o ciliòfors
Els cariorelictis són ciliats proveïts d’un micronucli i un macronucli diploide, que no es divideix quan ho fa el ciliat sencer; d’aquesta manera, el micronucli es divideix a cada cèl·lula filla i origina un macronucli i un altre micronucli. Tenen una ciliació bucal senzilla. Són petits i viuen a la mar o en l’aigua dolça. Tracheloraphis i Loxodes són dos gèneres ben coneguts.


Ciliat heteròtric del gènere Spirostomum, de vida lliure a l’aigua dolça.
Els ciliats espirotriquis estan proveïts de cirrus que els serveixen per a nedar i també per a desplaçar-se pel damunt de substrats sòlids. La fotografia mostra un individu del gènere Euplotes, de vida lliure i comú a les aigües dolces. Noteu els cirrus i la zona adoral de membranel·les, ben desenvolupats.
Miquel Blasi (a dalt), Pilar Gracia (a baix)
Sota la denominació d’espirotriquis es coneixen des de temps antics tota una sèrie de ciliats que en les classificacions modernes ocupen llocs diversos i tenen un diferent tractament taxonòmic, bé que aquest nom serveix per a fer-ne una ràpida identificació. Tots es caracteritzen per posseir a l’esquerra de la boca una gran formació membranel·lar anomenada zona adoral de membranel·les (ZAM) composta de nombroses membranel·les curtes i paral·leles, formades al seu torn per tres o quatre fileres de cilis; i per tenir, a la part dreta de la boca, una formació paraoral ciliar, amb els cilis ordenats de manera variable. Els de la subclasse dels heteròtrics tenen una zona adoral de membranel·les i una ciliació somàtica ben desenvolupades; n’hi ha de lliures, d’aigua dolça (Stentor, Spirostomum, Blepharisma i Metopus) i de paràsits de l’intestí d’insectes i vertebrats inferiors (Nyctotherus). Els tintínnids (una altra subclasse) són semblants als heteròtrics, i tenen una zona adoral de membranel·les molt grossa i una ciliació somàtica reduïda a alguns grups de cilis; quasi tots tenen lloriga i són importants com a habitants de les aigües marines, on s’han diferenciat molts gèneres (Tintinnus i Codonella són potser els més abundants). Finalment, també són espirotriquis els de les subclasses dels esticòtrics i els hipòtrics, que també han rebut conjuntament el nom d’hipòtrics, encara que entre ells hi ha certes diferències. En tots dos grups la zona adoral de membranel·les està ben desenvolupada i la ciliació somàtica és present només a la cara ventral, on els cilis es reuneixen en uns pinzells anomenats cirrus, que varien de nombre i distribució en cada espècie; els cirrus són orgànuls molt robusts i serveixen per a «caminar» sobre superfícies sòlides, perquè es mouen com potes, de manera que aquests ciliats, que neden bé gràcies al moviment de les membranel·les i dels mateixos cirrus, també poden ésser ciliats caminants quan cerquen l’aliment per sobre de les algues o d’algun altre objecte submergit. Les diferències entre ambdues subclasses rauen, a part algunes particularitats ultrastructurals de la ciliació, en el fet que les seves formacions paraorals són diferents i que els esticòtrics tenen dos o més macronuclis arrodonits i la cara dorsal nua, mentre que els hipòtrics tenen un sol macronucli molt allargat, en forma de salsitxa, i la cara dorsal amb unes cinèties de cilis gruixuts (sedes). Entre els esticòtrics destaquen els gèneres Stylonychia i Oxytricha, i entre els hipòtrics, Euplotes i Aspidisca.
Els gimnostòmides són ciliats que no tenen una ciliació bucal especialitzada i que en la classificació actual s’inclouen en les classes dels prostomats i litostomats. Tots són petits i senzills, i tenen la ciliació somàtica en forma de cinèties meridianes. La majoria són depredadors i de vida lliure, com Prorodon, Coleps, Litonotus, i d’altres.
Els tricostomats són ciliats, paràsits principalment de mamífers, de ciliació somàtica molt abundant i amb una ciliació bucal formada per polibraquicinèties o sincilis. Reuneixen dos ordres: els vestibulíffers, dels quals pot ésser un bon exemple Balantidium, un gènere paràsit de l’intestí dels amfibis i de molts altres animals; i els entodiniomorfs (Entodinium, Epidinium i Ophryoscolex), que són extraordinàriament notables per les seves formes extravagants i per la singularitat del seu hàbitat: el rumen dels remugants i el cec dels èquids.
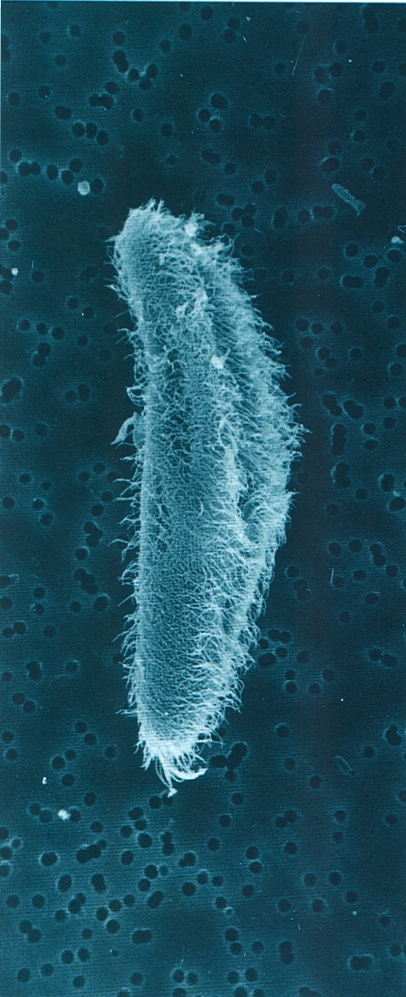
Parameci (Paramecium) vist al microscopi electrònic d’escandallatge. Noteu el profús revestiment ciliar.
Pilar Gracia
Els peniculins tenen una ciliació bucal molt complexa, amb llargues formacions de quatre files de cilis (panicles), que recorren la cavitat bucal, molt profunda. La ciliació somàtica és uniforme i abundant. El gènere més conegut és Paramecium. Hom els considera de la classe dels nassoforis per les seves particularitats ultrastructurals.
Els cirtofòrides s’inclouen en la subclasse dels fil·lofaringis i són uns petits ciliats herbívors, d’aigua dolça o marins, amb ciliació preferentment ventral; a la boca, situada també a la cara ventral, hi ha unes barnilles microtubulars en forma de banya buida per dins. Chilodonella és el gènere més conegut.
Els conotriquis són petits ciliats que fan vida lliure, sedentària, i que presenten una ciliació molt reduïda. Tenen el cos perllongat en forma d’embut, al fons del qual hi ha la boca. Un gènere comú és Spirochona.
Els suctoris també son coneguts com a tentaculífers, perquè els individus adults no tenen boca i s’alimenten per mitjà d’uns tentacles xucladors que, armats d’uns feixos de microtúbuls, tenen haptocists al seu extrem; tampoc no tenen cilis i són sedentaris: viuen fixats per un peduncle a un substrat sòlid, però produeixen per gemmació unes larves ciliades, que, després d’emigrar i fixar-se, donen lloc als adults. Tokophrya i Acineta són gèneres molt comuns, que apareixen al costat d’altres ciliats sedentaris, dels qual s’alimenten.
Els nassoforis es caracteritzen pel fet de tenir una ciliació somàtica uniforme i una boca amb una nansa grossa. Formen part de la subclasse dels nassoforis, dins la qual també s’inclouen els peniculins.
Els himenostòmides tenen una ciliació somàtica ben desenvolupada i una ciliació bucal formada per una membrana paraoral a la dreta de la boca i tres membranel·les de forma i desenvolupament desiguals a l’esquerra. Són molt freqüents a les aigües dolces, i es poden considerar els gèneres més conspicus Tetrahymena, Colpidium i Glaucoma.
Els astomacis són paràsits de l’intestí dels cucs de terra i de molts altres animals, com ara anèl·lids poliquets, equinoderms o amfibis, i es caracteritzen pel fet de no tenir ni boca ni ciliació bucal, encara que ostenten una ciliació somàtica ben desenvolupada. Es reprodueixen per escissió binària, però també per una gemmació múltiple que origina cadenes d’individus. Anoplophrya és paràsit de Lumbricus; Haptophrya s’ha trobat en amfibis.
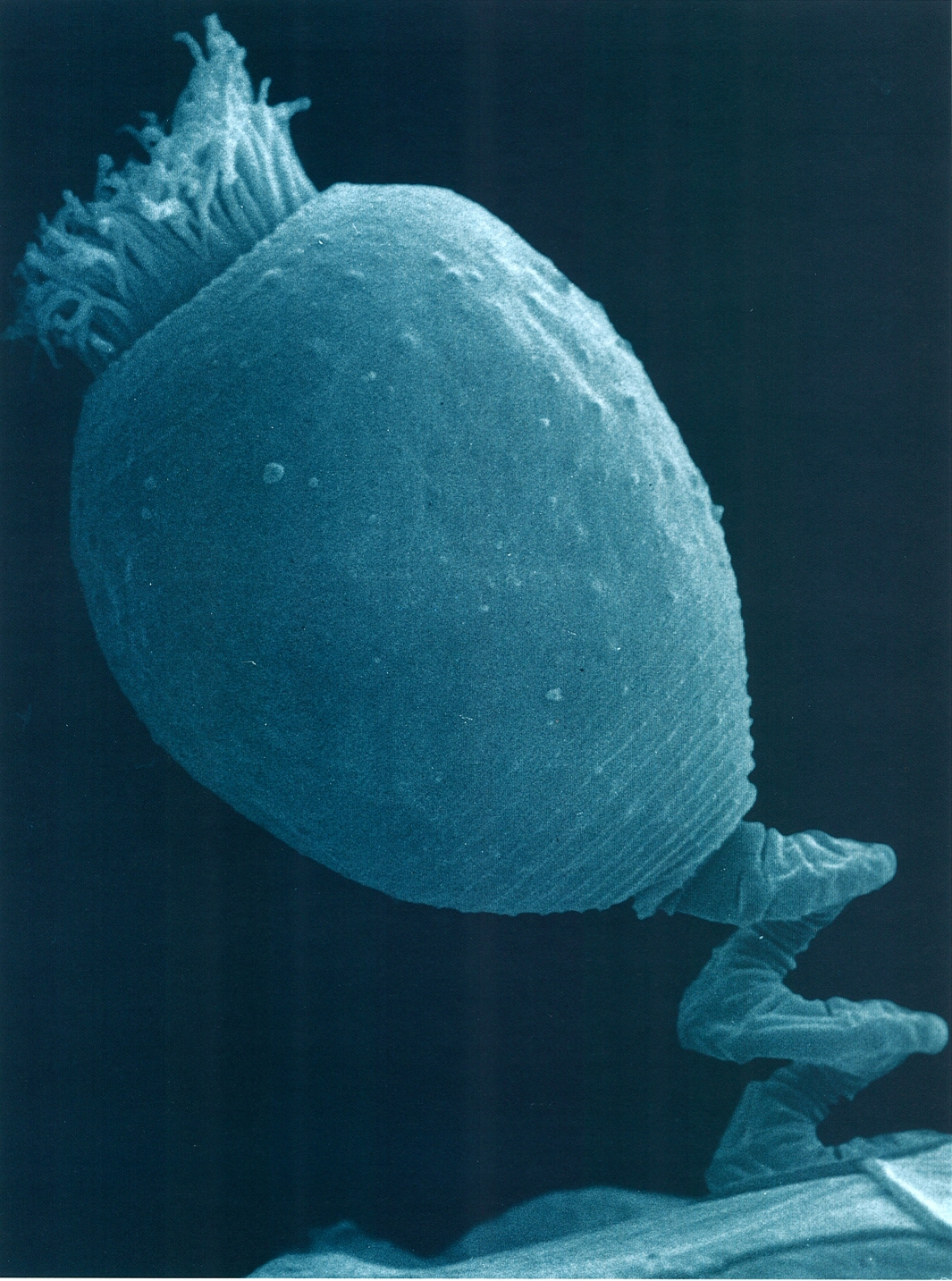
Vorticèl·lid (Vorticella), fixat pel peduncle damunt el cos d’una puça d’aigua (Daphnia pulex). Noteu el panicle de cilis bucals i la nul·la ciliació de la resta de la cèl·lula. És comú a les aigües dolces.
Mercè Durfort
Els peritriquis es caracteritzen pel fet de tenir una ciliació somàtica molt escassa. La ciliació bucals és constituïda per uns panicles llarguíssims i enrotllats en espiral, de manera que fan una volta molt ampla al voltant de la boca abans d’enfonsar-se cap endins. Molts són fixats per un peduncle que pot ésser contràctil o no; el peduncle pot ramificar-se i llavors es forma una colònia arboriforme. Són propis de les aigües dolces contaminades i n’hi ha molts que poden viure a les brànquies i altres apèndixs d’artròpodes d’aigua dolça, o també marins. Vorticella és coneguda pel seu peduncle contràctil, Charchesium és colonial i Epistylis viu sovint sobre metazous.
Els colpodis són ciliats que moltes vegades presenten forma de ronyó, i tenen una ciliació somàtica molt conspícua. La boca se situa al fons d’un vestíbul i és flanquejada per dos polibraquicinèties relativament petites. Un dels gèneres més freqüents és Colpoda.
Els protozous als medis naturals
Els protozous d’aigua dolça
Protozous comuns a les aigües dolces: 1 Stentor polymorphus, 2 Acanthamoeba castellani, 3 Arcella vulgaris, 4 Colpoda cucullus, 5 Dileptus anser, 6 Actinophrys sol.
Biopunt, a partir de fonts diverses
Les aigües dolces continentals són els biòtops més característics dels protozous, i també d’altres organismes, alguns macroscòpics, però principalment organismes d’una talla semblant a la seva, amb els quals solen establir relacions tròfiques; aquests organismes diminuts, que la majoria de vegades els serveixen d’aliment, solen ésser bacteris i les algues unicel·lulars. En les aigües dolces es troben pràcticament tots els grups de protozous lliures (amb l’excepció dels que són exclusivament marins), però els més freqüents són els flagel·lats, les amebes, els heliozous i els ciliats. Entre els protozous que viuen en les masses d’aigua dolça, podem distingir a grans trets els planctònics i els bentònics, i també entre els habitants d’aigües amb aliment abundant i els d’aigües menys fèrtils; és igualment ben divers el paper ecològic que hi fan: els uns es nodreixen de bacteris, els altres d’algues, d’altres de protozous, etc.
Les aigües temporànies, és a dir, basses originades per les pluges, generalment petites i de poc fons, sobre el sòl o la fullaraca, experimenten grans variacions de temperatura: un gruix tan petit d’aigua s’escalfa ràpidament amb el sol, i es refreda quan baixa la temperatura atmosfèrica, es gela, etc. Això fa que en aquestes aigües hi visquin protozous especialment preparats per a resistir aquests canvis, és a dir, capaços de fer cists de resistència. D’altra banda, aquestes aigües acostumen a ésser pobres d’aliment, per la qual cosa els protozous no hi solen fer poblacions importants. S’hi poden trobar amebes nues (Acanthamoeba, Mayorella, Hartmanella), amebes testàcies (Arcella, Difflugia), heliozous (Actinophrys) i alguns ciliats (Colpoda).
En les aigües permanents o en les aigües corrents, les condicions són d’una altra mena. D’altra banda, el fet d’atènyer profunditats més grans fa que moltes vegades s’estableixin condicions diferents a la superfície i al fons. A la superfície i fins a una fondària moderada hi ha llum abundant i una tensió d’oxigen relativament alta que, en condicions de contaminació modesta, depèn de la temperatura. Al fons dels llacs, en canvi, la llum arriba filtrada i la tensió d’oxigen pot ésser molt baixa; a més, fins i tot en condicions de poca contaminació, el fons sol ésser cobert d’un llim de matèria orgànica fonamentalment vegetal en estat de descomposició, llot sapropèl·lic, que desprèn gasos de fermentació reductors, especialment àcid sulfhídric. Això estableix una primera diferència entre els protozous de la superfície i els del fons, és a dir, entre els planctònics i els bentònics. Entre els protozous planctònics hi ha, a més de nombroses algues unicel·lulars fotosintetizadores (Chlamydomonas, Gonium, Euglena), afavorides per la claror, alguns heliozous (Actinophrys), i, sobretot, abundants ciliats, que s’alimenten de bacteris (Paramecium, Spirostomum, Tetrahymena), d’algues (Chilodonella) o d’altres protozous (Litonotus, Dileptus, Didinium). Aquests ciliats són gairebé tots nedadors, bé que a la zona superficial no falten ciliats fixos (Vorticella, Carchesium), d’altres que poden ésser fixos o surants (Stentor), i alguns hipòtrics, que caminen amb els cirrus ventrals per damunt de superfícies sòlides (Euplotes); quan viuen a la superfície, tots aquests ciliats se sostenen damunt de filaments d’algues i altres partícules. Els protozous bentònics d’aquestes aigües s’han adaptat a viure en un ambient reductor, pobre d’oxigen, on conviuen amb algunes algues decaracterístiques semblants, com alguna espècie d’Euglena. Entre aquests ciliats destaquen l’ameba gegant Pelomyxa i alguns heteròtrics (Caenomorpha, Metopus), com també peritriquis fixos (Vorticella, Epistylis) i suctors, també fixos i pedunculats (Tokophrya, Acineta); igual que a la superfície, són freqüents al fons els hipòtrics caminants (Euplotes, Aspidisca) i algunes amebes testàcies (Euglypha, Difflugia). Cal dir que en aquestes aigües hi ha, accidentalment, cists de protozous paràsits i fins i tot patògens (Entamoeba histolytica, per exemple); algunes amebes de vida lliure que es troben a les aigües dolces poden passar al cos humà i comportar-s’hi com a patògenes: és el cas de les petites amebes límax dels gèneres Acanthamoeba i Naegleria, que sovint viuen als rius, als llacs i fins i tot a les piscines, on s’alimenten de bacteris; Naegleria, que té una fase amebiana i una altra de flagel·lada, pot entrar accidentalment pel nas i passar a l’encèfal, on desencadena una malaltia greu, gairebé sempre mortal, anomenada meningoencefalitis amebiana primitiva; no obstant això, és poc freqüent.
En les aigües contaminades, aquelles conditions estan alterades. Es pot establir una classificació de les aigües contaminades, que les divideix en cinc grups, corresponents a cinc graus creixents de saprobietat, és a dir, de quantitat de contaminació orgànica; l’aigua de cadascun d’aquests graus té una fauna de protozous característica.
Les aigües xenosapròbies encara no són contaminades per matèria orgànica; són aigües de fonts i rierols, molt oxigenades, amb una demanda biològica d’oxigen (DBO) d’1; són, doncs, aigües d’una qualitat excel·lent i solament tenen algunes algues vermelles i crisofícies unicel·lulars. Les aigües oligosapròbies, dels llacs d’alta muntanya o de trams més baixos de rierols, són d’una qualitat semblant, però amb una DBO lleugerament més alta (entre 1 i 2,5) i amb més sais minerais, la qual cosa afavoreix el creixement de desmidiàcies i diatomees, que apareixen cobertes de protozous, com amebes testàcies i alguns ciliats nedadors, bé que s’hi desenvolupen fent poblacions modestes. Les aigües betamesosapròbies són les dels rius i basses, amb una quantitat mitjana de matèria orgànica i riques en oxigen, que posseeixen una activa flora bacteriana que descompon aquella matèria, i una DBO que oscil·la entre 3 i 5; en aquestes aigües, hi continuen vivint algues verdes, però també hi ha flagel·lats incolors i pròsperes poblacions d’amebes i ciliats bacteriòfags. El quart grau és constituït per les aigües alfamesosapròbies, que presenten més matèria orgànica en descomposició bacteriana i una certa escassesa d’oxigen, amb una DBO entre 10 i 20; gràcies a l’abundància d’aliment, ací hi ha un gran nombre d’espècies de protozous, especialment dels que s’alimenten de bacteris: zooflagel·lats, ciliats nedadors, ciliats sedentaris, heliozous i amebes nues; quantitativament, solen formar poblacions importants. Finalment, les aigües polisapròbies presenten una forta contaminació, amb xifres de DBO que arriben a 50 o fins i tot a més: el medi és microaeròfil, i hi apareixen bacteris filamentosos (Sphaerotilus), rotífers, nematodes i altres organismes de vida més o menys anaeròbia; els protozous hi presenten poblacions més modestes i, en general, es tracta de les mateixes espècies capaces de viure en medis més o menys anaerobis que ja hem esmentat en parlar dels fons de llacs i rius, ja que les condicions biològiques d’ambdós medis són força semblants.
Però encara hi ha, a més, un altre biòtop, que és el més contaminat per la matèria orgànica, i que és originat exclusivament per l’activitat humana: es tracta de les aigües residuals, anaeròbies, amb xifres de DBO situades entre 400 i 500, que es pot considerar com el darrer grau de saprobietat, conegut amb el nom d’aigües isosapròbies, en les quals també hi ha uns protozous específics: Metopus es, Trimyema compressa, Vorticella microstoma i Euglypha alveolata. La presència d’aquests i d’altres protozous en les aigües residuals és important pel paper que fan en el procés de la depuració, a les plantes de tractament d’aigües: en efecte, se sap que a les aigües i els llots activats d’aquestes plantes, els protozous, especialment els ciliats, col·laboren a netejar l’aigua, en primer lloc per la seva gran activitat com a devoradors de bacteris, i també perquè ajuden a la floculació d’alguns materials dissolts, en forma de flòculs que llavors s’eliminen més bé. Aquesta activitat millora la qualitat de l’aigua efluent i fins i tot es pot determinar per mitjà de l’estudi dels protozous presents quin és el rendiment en qualitat de la planta depuradora i la marxa del procés.
Els protozous del sòl
Protozous comuns a les comunitats del sòl, que viuen en els espais intersticials existents entre les partícules de sòl. 1 Arcella, 2 Bodo, 3 Mayorella, 4 Colpoda, 5 Chilodonella, 6 Euglypha. En el dibuix apareixen representats altres organismes de la comunitat: bacteris, hifes de fongs, actinomicets, etc.
Biopunt, a partir de fonts diverses
Els protozous representen una biomassa petita en comparació amb d’altres animals que formen la fauna del sòl (cucs de terra, larves d’insectes, col·lèmbols, etc.), però s’hi poden trobar en un nombre molt considerable. Aquesta fauna de protozous es troba predominantment als horitzons superficials i s’associa a les restes vegetals en descomposició; no obstant això, cal no oblidar que els protozous són organismes que viuen a l’aigua, i per tant, també al sòl requereixen humitat. Ara bé, l’aigua del sòl es troba sotmesa a unes fluctuacions extremes, especialment a l’horitzó superficial, on s’entolla quan plou però també s’evapora ràpidament; per això, els protozous solen trobar-se en un horitzó lleugerament inferior, que té l’aigua que puja per capil·laritat a les temporades seques, provinent de nivells inferiors. Per tant és important la porositat del sòl, ja que les partícules del sòl són recobertes d’una primíssima pel·lícula d’aigua que hi resta adherida per capil·laritat; el gruix d’aquesta làmina d’aigua, que és on habiten les poblacions de protozous, varia a cada moment segons l’estat d’humitat del sòl i segons la mida dels porus.
La majoria dels grups de protozous tenen representants en la fauna del sòl, però n’hi ha que són particularment importants, com els flagel·lats bodònids (Bodo), les amebes nues (especialment les petites límax), les amebes testàcies (Arcella, Difflugia) i molts ciliats (el més important és Colpoda, que pot constituir entre el 50 i el 95% dels protozous edàfics). Aquests protozous conviuen a l’esmentada pel·lícula d’aigua amb d’altres microorganismes, especialment bacteris i micelis de fongs; generalment, els petits metazous del sòl, com els nematodes, no s’hi troben.
Entre els protozous habitants del sòl es distingeixen dos tipus nutricionals: els micròfags, que mengen bacteris, i els que s’alimenten de microalgues i, especialment, de fragments vegetals en descomposició. Els primers són fonamentalment les amebes nues i els ciliats; els segons, les amebes testàcies. Les xifres d’abundància d’uns i altres varien segons l’abundància de l’aliment, que, al capdavall, depèn de la quantitat d’aigua; en efecte, als sòls amb menys aigua, la relació entre amebes testàcies i ciliats pot ésser d’ l:l, mentre que als sòls més humits, pot arribar a ésser de 10:1. Però en general es pot afirmar que tots els protozous del sòl estan molt adaptats a les fluctuacions d’humitat i de temperatura que hi ha a les capes superficials, adaptació que es revela principalment en la capacitat de formar cists i en la rapidesa de multiplicació que resulta de la necessitat d’explotar ràpidament un hàbitat més o menys efímer; aquesta rapidesa de multiplicació té com a conseqüència que en general la talla dels protozous del sòl sigui menor que la dels de la seva mateixa espècie que viuen a l’aigua.
El paper ecològic dels protozous del sòl no és uniforme, perquè, d’una banda les amebes testàcies col·laboren amb eficàcia amb els fongs actinomicets i mixomicets i amb els bacteris a la descomposició de la matèria vegetal, i, de l’altra, els ciliats bacteriòfags haurien de fer disminuir el nombre de bacteris i entorpir el procés de descomposició de matèria orgànica; de tota manera és possible que aquests ciliats col·laborin a la floculadó dels materials orgànics dissolts i afavoreixin així la seva degradació bacteriana. Una altra funció que s’ha atribuït als protozous del sòl en general és la de la descomposició de substàncies de degradació difícil (la lignina és degradada per les amebes testàcies) mitjançant el reciclatge d’elements a la xarxa tròfica. Igualment, se’ls ha atribuït una acció mecànica que pot afavorir l’acció bacteriana.
Els protozous de l’aigua marina
Els biòtops que hi ha en una massa d’aigua tan vasta i de característiques geogràfiques tan diverses com la mar són, naturalment, molt diferents: hi ha les aigües superficials de l’alta mar, els fons marins, la plataforma continental revestida d’algues, els estuaris amb aigües salabroses i amb fortes entrades de material orgànic, les platges sorrenques desproveïdes de vegetació, etc. Entre tots aquests biòtops hi ha una gran diversitat de faunes. Aquí ens limitarem a donar alguna informació sobre faunes de protozous molt diferents, que ens poden servir d’exemple per a mostrar la diversitat biològica d’aquests biòtops marins.
Els protozous intersticials
La microfauna de les platges sorrenques ha estat objecte d’estudis detallats. Aquest biòtop té una característica que el fa, en certa manera, semblant al sòl: és format per partícules inorgàniques (grans de sorra) entre les quals queda l’aigua quan es retiren les onades o baixa la marea. En aquests intersticis hi viuen, a més de bacteris i diatomees, uns petits animals que en conjunt justifiquen el nom de fauna intersticial amb què se’ls coneix, i entre els quals hi ha cnidaris, turbel·laris, gastròtrics, arquianèl·lids i ciliats. Tots comparteixen diverses característiques: tenen els cilis com a principal o únic mitjà de locomoció (turbel·laris, gastròtrics, ciliats); són molt petits i allargats, per a desplaçar-se més bé pels intersticis (per exemple, Remanella gigas, un cariorelíctide, mesura fins un 1 mm del llargada) i no semblen afectats per la naturalesa química dels grans de sorra sinó únicament per la grossària dels grans, la qual determina, naturalment, la magnitud dels intersticis. En aquest biòtop, hi ha representats diversos gèneres de protozous, tots de cos allargat (Remanella, Loxodes, Tracheloraphis, Holphrya, etc.), entre els quals predominen els cariorelictis. No s’ha trobat cap ciliat sedentari entre aquesta fauna intersticial, cosa ben lògica atesa la inestabilitat del substrat.
Els protozous oceànics
Dels protozous oceànics n’hi ha tres grans grups, que predominen tant per la seva importància numèrica com pel seu paper ecològic: els foraminífers, els actinòpodes i els tintínnids.
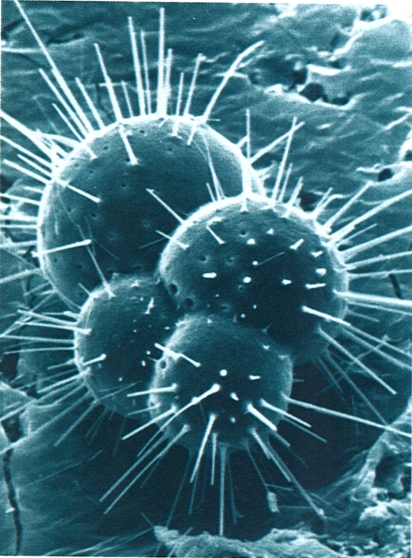
Els foraminífers marins que hom engloba dins el gènere Globigerina són segurament un extens complex d’espècies i fins i tot gèneres diferents, difícils de separar per raó de la seva gran variabilitat morfòlogica. Estan proveïts d’una conquilla calcària perforada. sovint dotada d’agulles finíssimes. com és el cas de l’espècie fotografiada. L’acumulació d’aquestes conquilles és important en la formació de sediments marins i té una gran transcendència per a les investigacions paleontològiques.
Lluís Vallmitjana / Marta Estrada / R. Fontarnau
Els foraminífers formen un conjunt de protozous marins d’una gran importància. Són organismes bentònics típics que llisquen per damunt de superfícies sòlides, principalment en fons no gaire profunds, sobretot damunt les algues de les zones costaneres, encara que n’hi ha alguns que es troben a grans fondàries, i d’altres que són pelàgics. S’alimenten amb una gran voracitat gràcies a la seva xarxa de reticulopodis, que reté qualsevol mena d’organisme: diatomees, peridinis, larves de crustacis i d’equinoderms, etc., per la qual cosa fan un paper de consumidors importants en la xarxa tròfica. També són molt interessants les closques dels foraminífers. Encara que n’hi ha de naturalesa orgànica, la majoria són calcàries i silícies, i fossilitzen molt bé. Els actuals són d’una mida relativament gran (entre 0,5 i 1 mm, aproximadament), però els del gènere Camerina, ja extingits, eren gegants (feien fins a 10 cm). L’acumulació i cimentació de les closques d’uns foraminífers que visqueren a l’Era Secundària, ha donat lloc a les potents calcàries nummulítiques; una altra roca, la creta, és formada també per closques de foraminífers. Han estat, doncs, uns importants agents geològies, i encara ho són avui, ja que als fons situats entre 150 i 800 m s’acumulen ingents quantitats de closques de foraminífers, principalment dels gèneres Globigerina i Globorotalia; constitueixen un extensíssim i potent dipòsit que rep el nom de «fang de globigerines». L’elaboració de la closca dels foraminífers depèn molt de la temperatura i de la composició química de l’aigua ambiental, per la qual cosa quan fossilitzen poden indicar amb precisió les condicions ambientals del moment, que es poden relacionar amb d’altres esdeveniments geològies. Com que, a més, són molt abundants als dipòsits marins, el seu estudi és actualment molt útil per a investigar l’existència i les condicions dels jaciments petrolífers.

Acantari planctònic marí, proveït d’un esquelet de vint varetes de sulfat d’estronci. Els acantaris generalment tenen algues simbionts al seu interior, però també actuen com a fagòtrofs.
Claude, Carré
Els actinòpodes més interessants de la mar són els acantaris i els radiolaris, dotats d’un esquelet mineral fossilitzable, com els foraminífers; però, a diferència d’aquests, són tots planctònics. Els acantaris i els radiolaris són els elements del plàncton marí més importants, al costat de les diatomees, les dinofícies (que representen els elements productors d’aquesta comunitat) i els tintínnids. Els acantaris i els radiolaris són fagòtrofs i, per tant, representen, al costat de molts animals planetònics i nectònics, macroscòpics i microscòpies, organismes consumidors. Però és important de tenir en compte que la presència de zooxantel·les és molt freqüent en aquests organismes, i això vol dir que la seva nutrició no és exclusivament holozoica. La major importància dels radiolaris en la mar es deu al fet que, a més de servir d’aliment a molts animals planctònics i nectònics, són uns agents geològics importants, igual que ho eren els foraminífers, perquè l’acumulació dels seus esquelets dona lloc en l’actualitat a extensos dipòsits d’un material anomenat «fang de radiolaris». En terrenys geològics més antics, les espícules que formen aquests esquelets silicis es varen dissoldre i varen tornar a precipitar en forma de sílice amorfa, constituint nòduls com els que es troben a la creta.
Finalment, els tintínnids també són components importants del plàncton oceànic i es troben en quantitats enormes en determinats llocs de les aigües marines. Es mouen gràdes a les membranel·les de la seva zona adoral, que són molt grosses i baten constantment contra la superfície de l’aigua. S’alimenten de bacteris i petites algues, i són presa de nombrosos animals, entre els quals hi ha peixos d’interès comercial.
Els protozous paràsits
Amastigotes de l’agent productor de la greu leismanias de l’home, Leishmania infantum, parasitant el citoplasma d’un macròfag de medul·la òssia de gos.
J.G. Esteban
Es calcula que de les 30 000 espècies de protozous vivents que es coneixen, unes 10 000 són simbionts. Els protozous paràsits són freqüents en metazous aquàtics i, especialment, a la superfície del cos, en contacte amb l’aigua ambiental. Així, en trobem que viuen com a comensals a les brànquies dels artròpodes (alguns ciliats peritriquis i d’altres que poden causar lesions locals d’una certa gravetat a la pell dels peixos, com el flagel·lat in color Ichtyobodo necator (=Costia necatrix) o el ciliat Ichtyophthyrius multifiliis.
Els protozous paràsits de l’aparell digestiu dels vertebrats han aconseguit un progrés cap a la intimitat de les relacions amb l’hoste; són d’aquest tipus, per exemple, els ciliats Opalina, Balantidium i Nyctotherus, que viuen a la cloaca de les granotes, les quals infecten quan encara són cap-grossos, per la qual cosa la transmissió d’un hoste a un altre es pot fer sense cap mena de formes de resistència ni vectors. Quan els hostes són terrestres, la transmissió sol ésser obligatòriament per cists, com passa en les amebes intestinals humanes. De les amebes humanes, Entamoeba coli i Endolimax nana són inòcues; en canvi, Entamoeba hístolytica és causa de la greu disenteria tropical humana; cal advertir que, malgrat el seu nom, la distribució d’aquesta disenteria és molt àmplia i s’estén pràcticament a tots els països temperats, inclosos els nostres.
Entre els protozous del tub digestiu hi ha casos de mutualisme. Són ben coneguts els flagel·lats tricomonàdides i hipermastígides que viuen a l’intestí dels tèrmits i d’altres insectes que mengen fusta, com les paneroles (dictiòpters blatodeus) del gènere Cryptocercus; dirigeixen la cel·lulosa i subministren nutrients a l’hoste en canvi d’allotjament i aliment. Un altre cas és el dels entodiniomorfs (Epidinium, Entodinium, Ophryoscolex, Polyplastron) i els vestibulífers (Isotricha), que viuen al rumen dels remugants, i el dels entodiniomorfs que viuen al cec dels èquids (Cycloposthium); tant el rumen, en un cas, com el cec, en l’altre, són cambres de fermentació on actuen les cel·lulases que l’hoste no pot produir; els simbionts són fonamentalment bacteris i llevats, però també ho són els protozous esmentats, que no es troben a cap altre lloc.
De vegades, tot i que l’entrada a l’hoste es fa a través de l’aparell digestiu, el protozou emigra cap els teixits, on viu i desenvolupa la seva activitat patògena fins que emigra novament al lumen de l’intestí on, en forma de cist, pot tornar a envair un altre hoste de la mateixa o d’una altra espècie; és el cas de les espècies d’Eimeria, que són patògenes per a pollastres o conills, i de Toxoplasma, els oocists del qual, carregats d’esporozoïts, apareixen a l’intestí gros del gat i d’allí passen a l’home.
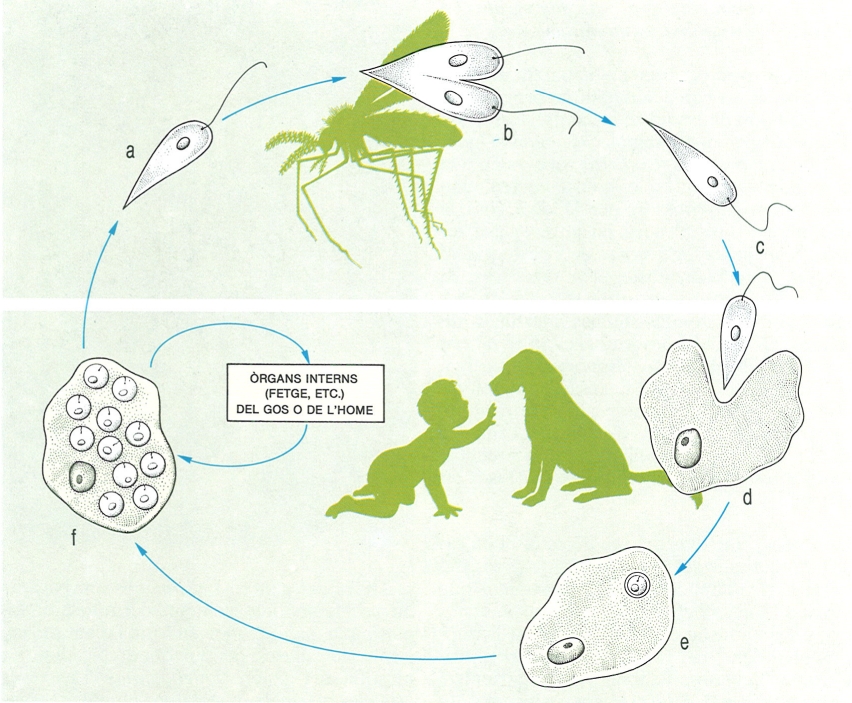
Cicle biològic de l’agent productor del kala-azar infantil, Leishmania infantum. El flagel·lat, en l’estat de promastigot (a) es multiplica (b) en l’intestí del mosquit Phlebotomus i és inoculat (c) en l’home o el gos per mitjà d’una picada. Quan penetra en el cos del mamífer, entra en un mascròfag (d), on adquireix l’estat d’ amastigot (e), i es multiplica en aquest estatfins a omplir tot el citoplasma del macròfag (f), amb el qual passa als diversos teixits del gos o de l’home, on estableix la malaltia del kala-azar. El mosquit s’infecta en picar un malalt (a), amb la qual cosa s’inicia un nou cicle.
Biopunt, a partir de fonts diverses
Un grau més complicat de parasitisme és el d’aquells protozous que habiten als teixits dels vertebrats, on arriben per mitjà d’un artròpode vector; els més interessants són els tripanosòmides i els hemosporídides. Entre els primers, considerarem els gèneres Leishmania i Trypanosoma, agents de greus malalties de l’home i dels animals. Els cinetoplàstides del gènere Leishmania són paràsits intracel·lulars dels elements del reticle endoplasmàtic, on es troben en forma d’amastigot, i són transmesos per un dípter picador, un mosquit del gènere Phlebotomus. Les seves espècies produeixen malalties humanes, alguna de les quals es dona als nostres climes. L. donovani és agent del kala-azar, una malaltia que es produeix en els adults de l’Àsia meridional; una varietat d’aquesta espècie, que alguns anomenen L. infantum, és l’agent responsable del kala-azar infantil, una malaltia que s’ha patit als Països Catalans, i que es manifesta per una gran inflamació del fetge i la melsa. L. donovani també afecta els gossos, als quals produeix una leishmaniasi canina molt greu; són precisament els gossos els que transmeten el paràsit als mosquits flebòtoms, i aquests el transmeten als nens. Una altra espècie patògena de Leishmania és L. tropica, agent del botó d’Orient, afecció de la pell que no es dona als nostres climes.
Les espècies de Trypanosoma resideixen, en forma de tripomastigot, a la sang de diversos mamífers, i algunes són greument patògenes. La majoria són transmeses per una mosca picadora del gènere Glossina, la mosca tse-tse de l’Àfrica tropical. T. gambiense i T. rhodesiense produeixen la greu malaltia de la son, en la qual els tripanosomes infecten no solament la sang sinó també el fetge, els ganglis limfàtics, el líquid cerebrospinal i la melsa. T. brucei i T. vivax, també transmesos per Glossina, a l’Àfrica produeixen malalties del bestiar. Hi ha d’altres tripanosomes que no són transmesos per la mosca tse-tse sinó pels tàvecs o per contagi directe, per la qual cosa són cosmopolites i les malalties que produeixen poden afectar el nostre país. Es tracta de T. evansi, causant de la surra dels equins, i de T. equinum, que causa el mal de malucs als mateixos animals, tots dos transmesos per tàvecs, i de T. equiperdum, agent d’una veritable malaltia venèria dels cavalls, l’anomenada durina, que es transmet per contacte sexual. Aquestes tres espècies de tripanosomes poden produir algun cas de malaltia a les nostres latituds però no són gaire freqüents. S’ha de fer una menció especial de T. cruzi, agent d’una important tripanosomiasi humana que dévasta grans extensions del continent americà, on produeix la greu malaltia de Chagas, transmesa per diversos insectes hemípters triatómids (Triatoma infestans i altres congèneres, Panstrongylus megistus i altres del mateix gènere i Rhodnius prolixus), que en conjunt es coneixen a l’Argentina amb el nom vulgar de «vinchuca» i a Brasil com a «barbeiro».
Finalment, esmentarem els esporozous hemosporidis, que viuen als glòbuls roigs de la sang dels vertebrats; per a nosaltres, els més importants són els agents de la malària, malaltia que durant segles va assolar els països mediterranis i que encara és un gran flagell en grans extensions d’Àfrica i Àsia. Aquesta malaltia és causada per diverses espècies del gènere Plasmodium i és transmesa per mosquits del gènere Anopheles. Es caracteritza per inferir greus danys al fetge i als òrgans hematopoètics i per la destrucció periòdica dels hématies circulants, que es manifesta en un accès febril periòdic, violent però curt (d’algunes hores). Són quatre les espècies causants de la malària humana: P. vivax, P. malariae, P. ovale i P. falciparum, totes cosmopolites excepte P. ovale, que és pròpia de l’Àfrica tropical. Tres d’aquestes espècies (P. vivax, P. ovale i P. falciparum) provoquen accessos de febre cada 48 hores, que es coneixen amb el nom de febres tercianes (perquè apareixen el primer i el tercer dia); els accessos de febre en la malaltia produïda per P. malariae són cada 72 hores, i per això s’anomenen febres quartanes (perquè apareixen el primer i el quart dia). En l’actualitat, s’ha aconseguit erradicar la malària de l’Europa méridional i el nord d’Àfrica, on va ésser un flagell devastador. Però continua essent-ho en alguns països, especialment de la resta d’Àfrica i d’Àsia, entre altres coses perquè l’espècie que allà predomina, P. falciparum, es fa constantment resistent als medicaments antipalúdics i també per la dificultat de preparar vacunes específiques. D’altra banda, gràcies a la facilitat actual de viatjar, de tant en tant apareixen brots de malària «importada» en països que l’havien erradicada completament.