
Les peces bucals mastegadores, típiques dels coleòpters, són ben visibles en el cap d’aquesta saperda (Saperda carcharias), un banyarriquer que ataca els xops o pollancres debilitats: les femelles, amb les mandíbules, fan incisions a l’escorça d’aquests arbres i hi dipositen un o dos ous. Les larves que en neixen s’alimenten dels teixits vegetals, cosa que les mena a fer galeries cada cop més profundes en el tronc afectat.
Rafael Campillo.
Els coleòpters o escarabats constitueixen el grup d’insectes més diversificat i més nombrós. Actualment, hom en coneix més de 350 000 espècies, que viuen en els ambients més variats, i aquest nombre probablement encara és baix si pensem en la quantitat de regions que encara cal estudiar.
Potser la característica més conspícua dels coleòpters, la que ens permet de reconèixer-los fàcilment, és la duresa del primer parell d’ales, els èlitres i la seva disposició en la posició de repòs: posades al damunt del cos, com formant un estoig, i amagant a sota el segon parell d’ales (poden ser reduïdes o, fins i tot, mancar) membranoses i aptes per al vol. Hi ha molts altres caràcters, però, que els diferencien dels altres insectes: al cap, les peces bucals mastegadores; al tòrax, el primer segment o protòrax molt més desenvolupat que els altres, que queden, juntament amb l’abdomen, més o menys coberts pels èlitres; i, a la base dels èlitres, una peça triangular anomenada escudet. D’altra banda, els coleòpters, com la resta dels oligoneòpters, són holometàbols, és a dir, tenen metamorfosi complicada en el seu desenvolupament, i també hi ha alguns casos d’hipermetamorfosi. Les larves poden tenir formes molt diverses, però sempre tenen el cap diferenciat. Les pupes són molt poc o gens mòbils, de vida lliure o semilliure, de vegades protegides pels coneguts capolls o cel·les nimfals.
La majoria dels coleòpters són terrestres i menen una vida lliure, però tampoc manquen exemples d’espècies paràsites, sigui d’animals o de vegetals. També hi ha coleòpters que han retornat a la vida aquàtica i, així, trobem espècies aquàtiques en tot el seu cicle i d’altres que en són només durant les fases larvals. El règim alimentari dels coleòpters és d’allò més variat, però cal remarcar la importància de la seva acció alimentària, ja sigui com a predadors d’altres insectes perjudicials, com a pol·linitzadors, com a col·laboradors en la degradació de la matèria orgànica, o, també, com a flagells de conreus i boscos, minadors de fusta i productes emmagatzemats, etc.
Hom ha pogut especular sobre l’origen dels coleòpters gràcies a l’aparició de fòssils, provinents del Permià, a l’Era Primària. A partir d’aquest període, el grup es diversificà extraordinàriament, fins a constituir, d’una manera progressiva, el conjunt més divers i amb el nombre d’espècies més gran de tots els insectes.
Morfologia
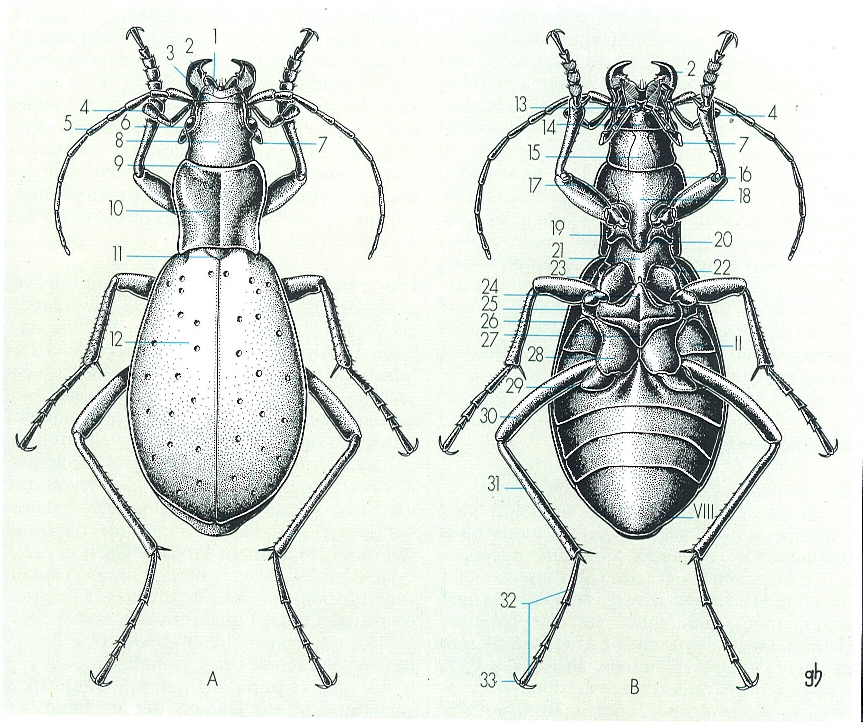
Aspecte general extern d’un coleòpter (×5), en visió dorsal (A) i ventral (B). En l’esquema, hom hi ha indicat: 1 labre, 2 mandíbula, 3 clipi, 4 palp maxil·lar, 5 antena, 6 ull compost, 7 palp labial, 8 front, 9 vèrtex, 10 pronot, 11 escudet, 12 èlitres, 13 maxil·la, 14 llavi, 15 gula, 16 episternita del protòrax, 17 sutura notopleural, 18 prostèrnum, 19 epimerita del protòrax, 20 apòfisi prosternal, 21 mesostèrnum, 22 episternita del mesotòrax, 23 epimerita del mesotòrax, 24 metastèrnum, 25 episternita del metatòrax, 26 epipleura dels èlitres, 27 peça premetacoxal, 28 coxa, 29 trocànter, 30 fèmur, 31 tíbia, 32 tars, 33 pretars (format per dues ungles).
Gustavo Hormiga, del natural
Entre els coleòpters, predominen els de dimensions petites i mitjanes: de menys d’1 mm fins uns quants centímetres de llargada. Algunes espècies tropicals atenyen fins a 15 cm (Goliathus regius, Megasoma elephas, Macrodontia cervicornis), les quals, al costat d’algunes papallones, també tropicals, constitueixen els insectes actuals de dimensions més grans. La forma del cos també és molt variada: alguns són esvelts i delicats, d’altres arrodonits i robusts, i hi ha tota una gamma de formes entre aquests dos extrems que es combina amb un desenvolupament variable dels apèndixs. Tot i que predominen les espècies de colors poc vistents, principalment marrons, negres i grisos, alguns tenen reflexos metàl·lics, verdosos, tornassolats, etc, i també colors llampants fent dibuixos als èlitres i a la resta del cos.
La cutícula, en general, és molt endurida i constitueix una veritable cuirassa. Sovint porta pèls i relleus en forma de tubercles, depressions, reticles, punts, o bé costelles, estries i dibuixos de tota mena. Alguns tenen apòfisis amb forma de banya, o d’esperó, a diferents regions del cos, les quals els donen aspectes molt peculiars. Aquestes apòfisis moltes vegades són caràcters sexuals secundaris, i solen tenir un desenvolupament més gran en els mascles.
El cap dels coleòpters sol ser prògnat, però també pot ser ortògnat. Moltes vegades es prolonga endavant per una mena de rostre o trompa, com passa en els curculiònids, i, en d’altres, s’encaixa cap ensota del protòrax, però en general queda lliure i ben visible. Els ulls compostos poden ésser absents, en alguns casos excepcionals, igual que passa amb l’ocel·le o els dos ocel·les que poden tenir. Les antenes tenen diverses formes i dimensions, i són constituïdes per un nombre d’artells generalment variable entre 9 i 11, però algunes vegades inferior (2 en els paússids) o superior (27 en alguns cerambícids). Les peces bucals són de tipus mastegador, amb un grau de desenvolupament variable en relació amb el règim alimentari de cada espècie; en algunes, les mandíbules es troben hipertrofiades, és a dir, són molt més grans que normalment, i constitueixen un tret característic del dimorfisme sexual, com és el cas de l’escanyapolls (Lucanus cervus). En d’altres, les mandíbules i tot el complex maxil·lar perden la típica funció mastegadora i es transformen en un aparell per a perforar, xuclar i llepar.
Dels tres segments que formen el tòrax, el primer o protòrax és el més desenvolupat i té una gran mobilitat. Per contra, el mesotòrax i el metatòrax, fusionats i el primer molt reduït, no són visibles dorsalment, ja que queden amagats sota els èlitres i, d’altra banda, quan hi ha una reducció o atròfia del segon parell d’ales, el segment corresponent també la manifesta; la seva cara dorsal és dividida en diverses esclerites, una de les quals forma l’escudet (corresponent al mesotòrax), que és visible entre els èlitres.
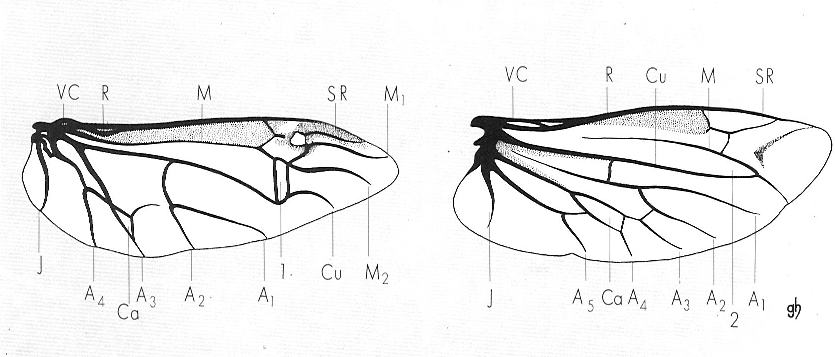
Estructura i nervació de les ales dels coleòpters, tipus adèfag (a l’esquerra), i polífag (model cantaroïdeu, a la dreta). Noteu l’existència d’una cel·la tancada, l’anomenat oblòngum (1), en els adèfags, contra la cel·la oberta, apèrtum (2), dels polífags. VC costal, R radial, M mediana, SR sector radial, Cu cubital, VA anal, J joval, Ca cèl·lula anal.
Gustavo Hormiga, original de Crowson/Forbes
Dels dos parells d’ales típiques, el primer, que forma els èlitres, s’articula al mesotòrax, i el segon, membranós, o en alguns casos reduït, ho fa al metatòrax; en cas que aquest segon parell sigui reduït o absent, els èlitres queden íntimament imbricats. La configuració i el desenvolupament dels èlitres són variables: poden ser plans o convexos, puntuats, estriats o llisos, glabres o peluts, etc.; a la regió dorsal moltes vegades porten estries longitudinals, que hom creu que corresponen a l’antiga nervació de les ales. La llargada dels èlitres també pot variar força: en general cobreixen tot l’abdomen, però hi ha vegades que són tan curts que amb prou feines tapen la regió tergal del tòrax, i encara, en alguns casos excepcionals, són atrofiats. Les ales del segon parell, les membranoses, son més llargues que els èlitres, però en posició de repòs queden plegades sota d’aquests; la nervació d’aquestes ales té una gran importància per a interpretar la filogènia del grup, i hom distingeix dos tipus bàsics: el tipus adèfag, més primitiu (característic dels arcostemats i dels adèfags), i el tipus polífag (característic dels mixòfags i dels polífags), que alguns autors consideren representat per dos models: l’estafilinoide i el cantaroide. Durant el vol, que en la majoria dels coleòpters és lent i poc àgil, els èlitres es mantenen oberts en forma de V per sobre del cos per tal de fer possible l’extensió de les ales posteriors, que són les responsables del vol.
Les potes s’articulen als tres metàmers toràcics i, ultra la seva funció per a caminar i córrer, poden mostrar adaptacions per a excavar (en els escarabèids, alguns caràbids i altres), nedar (en els ditíscids, els girínids, alguns hidrofílids, etc.), saltar (alguns crisomèlids, per exemple), etc. Els artells que les componen són els típics de tots els insectes, bé que els diferents grups poden tenir adaptacions peculiars, sigui a les coxes i les cavitats coxals o, sobretot, en el nombre d’artells que formen el tars (els tarsòmers), caràcter aquest que és molt útil per a identificar algunes famílies i per a situar-les dins de la successió evolutiva: hom considera el nombre de cinc tarsòmers com a condició primitiva, però pot reduir-se fins a tres; d’altra banda, els tarsòmers poden ésser dilatats, especialment en els mascles, com un caràcter sexual secundari, mentre que el darrer generalment porta dues ungles terminals, que constitueixen el pretars.
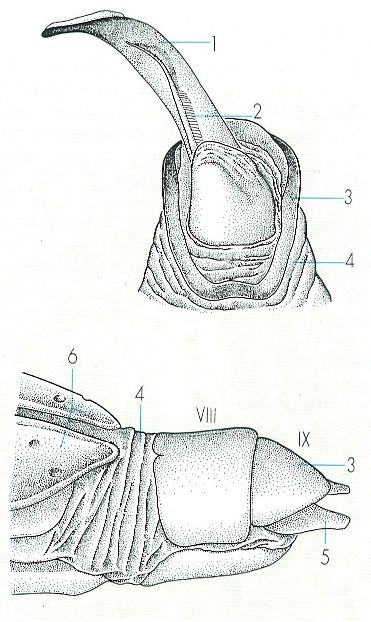
Esquema de l’extrem de l’abdomen i la genitàlia dels coleòpters, en el cas d’un mascle, en posició ventral (a dalt), en el qual s’observa l’edeagus semievaginat, i en el d’una femella, en posició lateral (a baix), amb la genitàlia també evaginada. Hom hi ha indicat: 1 lòbul mitjà de l’edeagus o òrgan copulador, 2 estils de l’edeagus, 3 segment genital (IX abdominal), 4 membrana intersegmentària, 5 gonapòfisi, 6 èlitres.
Gustavo Hormiga, del natural
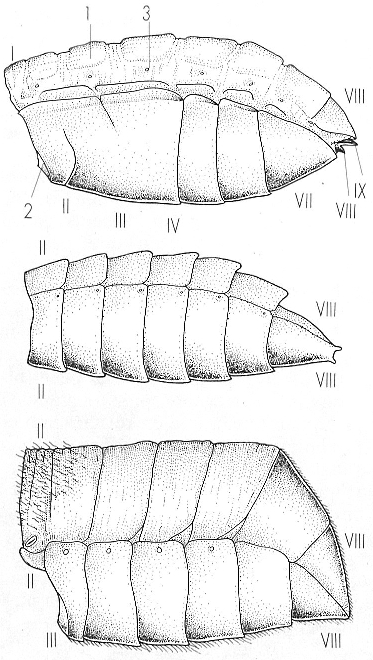
Esquema de diferents models d’estructura abdominal en els coleòpters. Tot i que l’abdomen es compon sempre de deu segments, no tots són visibles per causa de l’atròfia dels primers i de la invaginació i modificació dels darrers. En l’abdomen adèfag (a dalt), el més característic és l’atròfia de la primera esternita i, la fusió de les esternites segona, tercera i quarta; l’hologastre (al mig) té la segona esternita semblant a les següents; l’haplogastre (a baix), com l’anterior, característic d’alguns polífags, té la segona esternita molt reduïda i que queda tapada sota el tercer parell de potes. Un model derivat és el criptogastre o simfiogastre, en què desapareix la segona esternita abdominal. 1 tergita, 2 esternita, 3 estigma.
Gustavo Hormiga, a partir de Jeannel & Paulian
L’abdomen, de forma i dimensions molt variables, és constituït en els coleòpters per 10 segments o metàmers, encara que no tots són visibles a causa de l’atròfia d’alguns dels anteriors i de la invaginació dels terminals; a això s’afegeix el fet que el desè, anomenat metàmer anal, és molt reduït. Segons quin sigui el nombre d’esternites visibles hom distingeix quatre tipus d’abdomen: adèfag, criptogastre (o simfiogastre), haplogastre i hologastre, cadascun característic d’un grup determinat de coleòpters, i sobre la seva estructura hom fonamenta actualment tota la filogènia del grup. A l’abdomen se situa, també, l’orifici genital: just per darrere de la novena esternita en els mascles, i en la novena esternita mateix en les femelles. En relació amb els segments genitals es diferencia la genitàlia o conjunt d’estructures, que hom considera apèndixs modificats, relacionades amb la reproducció. L’estructura de la genitàlia és d’una gran utilitat en la determinació de les espècies de coleòpters. En els mascles, hom anomena edeagus el conjunt de peces que componen la genitàlia i, segons el desenvolupament i la disposició de cadascuna, hom defineix quatre models bàsics: el trilobulat, el vaginat, el de genet i l’articulat. A la còpula, una part de l’edeagus, l’anomenat sac intern, s’evagina i s’introdueix a les vies genitals femenines, mentre que altres parts s’encarreguen de subjectar la femella i d’obrir l’orifici genital. La genitàlia femenina és més simple: tot i que no tenen ovipositor, en les femelles de les espècies que ponen els ous dins de fissures i foradets en diversos medis (com alguns cerambícids i edemèrids, entre altres), els metàmers terminals poden ser retràctils i tubulars, proveïts de membranes intersegmentàries molt desenvolupades que permeten que es telescopitzin, i fan les mateixes funcions que un ovipositor. En molts casos, la posta va precedida d’una tasca preparatòria d’excavació, que la femella porta a terme amb l’ajut d’alguns dels components de la genitàlia (les hemisternites).

El dimorfisme sexual és prou comú entre els coleòpters, com el cas de l’escanyapolls (Lucanus cervus); a la dreta es pot observar un mascle, amb les seves característiques i enormes mandíbules, ben diferent de la femella, situada a l’esquerra. Els lucànids, família a la qual pertany l’escanyapolls, tenen antenes colzades i acabades en una maça pectinada, cosa ben patent en la femella il·lustrada.
Jordi Berthold
El dimorfisme sexual, com ja hem comentat, és força corrent entre els coleòpters. Les diferències morfològiques entre els mascles i les femelles solen residir en les dimensions i els colors, o en el grau de desenvolupament d’alguns apèndixs i en la presència d’unes determinades apòfisis. En alguns casos, les diferències són absolutament exagerades, tant, que ningú diria que el mascle i la femella pertanyen a una mateixa espècie. n’és un bon exemple el cas de les anomenades cuques de llum (lampírids), que també es dóna en altres espècies, en el qual el mascle té un aspecte típic de coleòpter mentre que la femella adopta forma de larva.
Biologia
La reproducció
La reproducció sexual és l’habitual en els coleòpters. Només hi ha alguns casos, excepcionals, de partenogènesi entre alguns escarabats estafilínids (els mirmecòfils del grup dels aleocarins) i en algunes marietes de muntanya (coccinèl·lids), als quals les larves acabades de néixer ja els falta poc per a la nimfosi. En aquest cas i en d’altres, com en alguns coleòpters cavernícoles, el desenvolupament postembrionari és escurçat i, per tant, tenen menys estadis larvals que els típics del grup. Tenen metamorfosi complicada (holometabolia) i es coneixen alguns casos d’hipermetamorfosi.
L’apropament i el reconeixement dels individus mascles i femelles per a l’aparellament és afavorit per l’emissió de feromones per part de les femelles i pel reconeixement d’aquestes per part d’òrgans olfactoris especials dels mascles, i també per l’emissió de sons que els mascles capten per mitjà dels òrgans auditius; igualment, la visió intervé també en el reconeixement, en darrer terme. El mètode d’apropament més eficaç, efectiu i estès és el primer, i, en general, els mascles són molt més actius en la recerca de parella que no les femelles, tret d’algunes rares excepcions. De vegades, el mecanisme d’apropament és indirecte, a través de l’olor que desprenen les fonts d’aliment, damunt de les quals la femella haurà de pondre els ous, que afavoreixen el trobament de la parella. En altres casos, els adults no abandonen mai el substrat alimentari que ha permès el seu desenvolupament, cosa que afavoreix que es formin poblacions nombroses d’una mateixa espècie damunt d’un substrat determinat.
L’aparellament sol ésser breu i moltes vegades va precedit d’un festeig, més o menys sofisticat. Durant l’acoblament, el mascle s’enfila damunt la femella i introdueix el sac intern del seu penis dins de la vagina de la femella, mentre la subjecta amb les potes per mitjà dels tarsos, generalment dilatats per a fer aquesta funció. La fecundació és, doncs, interna, i generalment els escarabats s’aparellen una sola vegada en tota la vida.
La posta, generalment, es fa immediatament després de l’aparellament o bé amb un lleuger retard, durant el qual, els espermatozoides resten emmagatzemats dins del receptacle seminal fins que els òvuls són madurs. Les femelles usualment fan una sola posta, ja que els adults viuen poc temps, però hi ha excepcions, i per a fer-la cerquen el substrat alimentari adequat per a alimentar les futures larves; en algunes espècies, la pròpia femella ha de preparar aquest substrat: els escarabats piloters (Scarabaeus sacer) preparen les pilotes de fem, el banyarriquer (Saperda populnea) i alguns Iberodorcadion fan petits tallets als troncs als quals viuran les larves per tal de fer que s’assequin o es debilitin sincrònicament al seu desenvolupament; alguns curculiònids i crisomèlids protegeixen les larves a base de modificar les fulles i els fruits als quals deixen els ous. De vegades, els adults protegeixen directament els ous i les larves, com fa Helochares, que transporta els ous al dors, mentre que d’altres estableixen una relació directa amb les cries, com fa Xyloterus, un escarabat escolítid, que vigila l’alimentació de les larves. Alguns caràbids, com Carterus, Abax, o Molops, també vigilen les cries: Carterus calydonius, per exemple, recull petites llavors d’apiàcies (del gènere Daucus) per tal d’alimentar les larves acabades de néixer. Són nombrosos els casos de vigilància de les cries entre els escarabats.
Els ous de la posta generalment són ovalats, blancs i llisos a la superfície, encara que en algunes espècies són acolorits i prenen formes geomètriques especials. El nombre d’ous de cada posta oscil·la entre algunes desenes i uns quants milers. Algunes famílies tropicals els reuneixen dins d’ooteques semblants a les que fan les paneroles (blàtids).
El desenvolupament

Diferents fases del desenvolupament d’un coleòpter crisomèlid, concretament de l’escarabat de la patata (Leptinotarsa decemlineata): posta i larves acabades de néixer (a dalt, a l’esquerra), larves en un ulterior estadi de desenvolupament, actives devoradores de fulles (a baix, a l’esquerra), larva prepupal i pupa desprenent-se de l’exúvia larval (a dalt, a la dreta), i individus adults, també devoradors de fulles (a baix, a la dreta). Tot el cicle d’aquesta espècie té lloc sobre les fulles de patatera i altres solanàcies.
Rafael Campillo.
El desenvolupament embrionari dels coleòpters sol ésser ràpid, de manera que dels ous no triguen a aparèixer les primeres larves, que passaran per diferents estadis larvals separats per mudes, del darrer dels quals s’originarà la pupa, i d’aquesta, l’adult. El nombre d’estadis larvals que componen el desenvolupament postembrionari oscil·la entre un i quinze, i generalment les diferents larves tenen totes un aspecte semblant (holometabolia), bé que hi ha també el cas contrari: l’aparició d’estadis larvals amb característiques molt diferents que caracteritza l’hipermetamorfisme és present en alguns coleòpters paràsits d’animals, com en els melòids.
La morfologia de les larves és força uniforme en alguns caràcters bàsics estructurals: la càpsula cefàlica ben diferenciada i proveïda de peces bucals de tipus mastegador n’és la principal. En canvi, l’aspecte és d’allò més variable, i hi ha una gran heterogeneïtat pel que fa al grau de desenvolupament de les potes: algunes tenen els tres parells dels adults, i per tant, es poden desplaçar ràpidament pel substrat, d’altres són completament àpodes i tenen, en canvi, unes protuberàncies ambulacrals. Al final del cos, algunes larves hi tenen cercs, urogomfs o pigopodis. Hi ha larves molt actives, depredadores, i d’altres que en són molt poc, que solen ser fitòfagues o sapròfagues; les àpodes es desenvolupen sempre dins del substrat alimentari, sigui fusta, excrements, bolets, cadàvers, etc. Hom ha descrit aproximadament una dotzena de tipus de larves d’escarabat diferents.
Quan les larves del darrer estadi arriben al moment de pupar, cerquen un lloc adequat, tant per les condicions climàtiques com per les de protecció. En alguns casos construeixen capolls i cel·les nimfals amb materials diversos per tal de preparar el seu període inactiu de pupa, durant el qual s’esdevindrà tot el conjunt de fenòmens fisiològics que porten a l’estructuració de l’adult. Les pupes són molt poc mòbils o, fins i tot, són totalment immòbils. La majoria són exarates, és a dir, tenen els apèndixs i les ales plegats dins d’unes fundes anomenades pteques, que es marquen a la superfície de la pupa; també n’hi ha, però, d’obtectes, que tenen ales i apèndixs recoberts per la cutícula del cos, i no es diferencien externament. En ambdós casos, es tracta de pupes adèctiques, que no fan moviments importants. Quan acaba l’etapa pupal, emergeix l’adult o imago, amb la morfologia típica de l’espècie, però amb la cutícula tova i mancat de capacitat reproductora, el qual esdevé adult al cap de poc temps.
La fase d’adult no acostuma a superar els dos anys de vida, i el més corrent és que duri només algunes setmanes. Durant el període d’adult, els individus no creixen i, per tant, no experimenten cap més muda. La seva funció és la de reproduir-se. El cicle complet dels coleòpters generalment és anual, és a dir, s’acompleix dins d’un any, però en molts casos, la fase de desenvolupament postembrionari o larval pot durar anys, més que no pas la vida d’adult.
Ecologia i etologia
L’estratègia biològica
Els coleòpters s’han adequat, al llarg de la seva història evolutiva, a gairebé tots els ambients naturals, fins i tot els subterranis. La majoria són terrestres, però alguns han colonitzat el medi aquàtic: famílies senceres d’escarabats, com els ditíscids, els higròbids, els halíplids, els girínids i els hidrofílids, viuen gairebé exclusivament al medi aquàtic continental i han adquirit formes hidrodinàmiques que els permeten nedar més bé. No obstant això, la majoria dels coleòpters aquàtics també tenen ales, que només fan servir per a passar d’un sistema aquàtic a l’altre. S’han especialitzat en funció de la natura i les condicions del medi aquàtic: hom pot distingir un grup característic de llacs i basses grans d’aigua dolça (els ditíscids), un altre propi de rius i rieres d’aigües més oxigenades (els psefènids, els driòpids, els èlmids, etc.), i altres propis de llacunes salabroses o salades, tant litorals com de l’interior (d’origen endorreic), i són capaços de suportar concentracions de sal molt altes (escarabats halòfils i halobis, com els Octhebius) i un altre que inclou els escassos coleòpters marins. Entre els coleòpters no hi ha, en canvi, espècies pelàgiques o que visquin a mar oberta, sinó que no passen de viure submergits una part del dia a les aigües costaneres, com Aepopsis, Aepus i Octhebius, entre altres.
Els coleòpters terrestres viuen al mateix lloc on troben bones condicions per a alimentarse i reproduir-se. En el cas de les espècies caçadores, l’alimentació de les quals depèn de la presència d’altres organismes, els factors climàtics i, especialment, el grau d’humitat ambiental, influeixen molt en la seva distribució; la majoria viuen sota les pedres (lapidícoles), on troben refugi i condicions ambientals de temperatura i humitat més estables; altres ho fan a la vora de l’aigua, entre les pedres mig submergides (espècies ripícoles), i unes altres a les molses (muscícoles) o entre la fullaraca humida (humícoles).

Els coleòpters cavernícoles presenten diverses singularitats morfològiques, com és evident en aquest caràbid del gènere Aphaenops, endèmic de les coves pirinenques: cos i apèndixs molt estilitzats, despigmentació, manca d’ulls, etc.
Jordi Berthold.
Alguns coleòpters petits penetren a les capes profundes de l’escorça terrestre; són els hipogeus, que poden ser, a son torn, endogeus o cavernícoles. Els endogeus viuen als intersticis del sòl, i es caracteritzen, globalment, per les seves dimensions, molt petites, i pel fet de tenir els apèndixs molt curts. Els cavernícoles tenen el cos i els apèndixs més esvelts i generalment són més grossos. Uns i altres, però, tenen en comú el fet de no tenir ulls i d’ésser despigmentats. Els cavernícoles, també anomenats troglobis, troglòfils o trogloxens segons de quin grau sigui la seva adequació a aquest medi, viuen dins de les coves i avenes i en l’anomenat medi subterrani superficial; de tots, els troglobis són els més especialitzats i els que viuen exclusivament en el medi cavernícola. Els caràbids, els catòpids, els pselàfids, els histèrids i els curculiònids, entre d’altres famílies, són els que tenen més espècies hipogees, moltes de les quals són endèmiques d’àrees petites a causa de la seva escassa capacitat de dispersió.

Els ambients molt secs i calorosos indueixen modificacions, com les d’aquest tenebriònid dels arenys litorals del migjorn català (Pimelia interjecta), que presenta una pigmentació intensa, un tegument molt endurit i els èlitres estretament juxtaposats per tal d’evitar la deshidratació; en aquestes condicions són corrents les formes de vida crepuscular i nocturna, a fi de defugir les hores d’insolació forta.
Enric Curto.
Altres grups de coleòpters s’han adequat a viure en climes extremament secs, àrids i calorosos. Són els anomenats xeròfils o esteparis, d’entre els quals encara hom pot discernir els sabulícoles, que viuen preferentment en arenys i dunes, tant del litoral com de l’interior. Els gipsòfils i els halòfils viuen en terrenys extremament salats de la costa o de les conques endorreiques de l’interior. Tots aquests grups xeròfils són molt resistents a la dessecació i han desenvolupat mecanismes fisiològics i morfològics que els permeten de sobreviure en condicions tan extremes com les dels deserts. Algunes de les seves característiques són la duresa i gruixària del tegument, en general negre, i els seus costums nocturns, la llargada de les potes, que els permet mantenir-se separats del terra, i la presència de llargs pèls als tarsos, que els donen la possibilitat de caminar àgilment per la sorra; d’altra banda, en general el cap, el tòrax, l’abdomen i els èlitres encaixen perfectament per tal d’evitar al màxim l’evapotranspiració. Entre les espècies xeròfiles hi ha molts tenebriònids, cicindèlids, i caràbids. També hi ha coleòpters que viuen en zones extremament fredes i humides, com és el cas dels nivícoles, que hom pot veure al límit de les neus, a l’alta muntanya; aquests escarabats tenen el tegument molt prim i fi, de colors clars, per tal de compensar l’ambient sobresaturat d’aigua al qual viuen.
A més dels factors climàtics, hi ha altres factors que condicionen la presència de coleòpters en un substrat determinat, i és l’existència de l’aliment adequat. Els insectes copròfags viuen a les proximitats dels excrements de tota mena d’animals, de vegades perquè se n’alimenten, i altres vegades perquè allà troben els organismes realment copròfags que els fan de presa; són copròfags molts escarabèids, hidrofílids, estafilínids i histèrids. Uns altres, els necròfags, s’alimenten d’animals en descomposició o de larves d’altres insectes (especialment dípters) que viuen dins dels cadàvers, com és característic de molts sílfids, dermèstids, clèrids i histèrids, entre altres. Els sapròfags, o més ben dit, sapròfits i saproxilòfags s’alimenten de matèria orgànica vegetal en descomposició; generalment es tracta de coleòpters de dimensions molt petites. Els micòfags o micetòfags mengen gairebé exclusivament bolets i altres fongs, i alguns viuen sota les escorces dels arbres (són corticícoles), on troben uns fongs determinats que els serveixen d’aliment.
Un gran nombre de coleòpters són fitòfags, especialment cerambícids, crisomèlids, curculiònids, brúquids, antríbids, bostríquids, anòbids, alguns escarabèids i representants de moltes altres famílies. Els fitòfags s’alimenten de matèria vegetal, sigui viva o morta i, en aquest darrer cas, contribueixen a llur descomposició. D’entre els fitòfags, hom distingueix els xilòfags, que s’alimenten de fusta de diferents menes, sigui dels troncs dels arbres o de la fusta morta de les construccions humanes; altres mengen fulles o brots tendres (fil·lòfags) i arrels (rizòfags) de diferents plantes, a les quals poden perjudicar notablement. També hi ha espècies florícoles, que s’alimenten d’algunes parts de les flors o de les substàncies que hi troben, com el nèctar i el pol·len; això no obstant, de vegades viuen a les flors per a caçar-hi altres organismes que sí que són veritablement florícoles. Algunes de les espècies fitòfagues poden atacar considerablement els conreus i produir danys a les fustes i a diversos productes emmagatzemats d’origen vegetal, com ara llegums, gra, etc. (espermòfils). Les espècies anomenades carpòfiles s’alimenten de fruits. Finalment, altres poden atacar la farina i les substàncies amilàcies.
Hi ha encara el cas particular dels coleòpters que viuen associats amb d’altres animals, tot establint relacions de parasitisme, simbiosi o comensalisme. Un cas és el dels mirmecòfils, que viuen dins dels nius de formigues, on també passen el seu període de desenvolupament; la mirmecofília és freqüent, entre d’altres, en els estafilínids Atemeles i Lomechusa, en Sternocoelis (un histèrid), en els tenebriònids del gènere Oochrotus, en els paússids Paussus, en el gènere Claviger (un pselàfid), en Clytra (un crisomèlid), i en l’espècie Amorphocephalus coronatus, l’únic brèntid present a la fauna europea. D’aquestes, algunes viuen en simbiosi amb les formigues, les quals els protegeixen per tal d’obtenir-ne a canvi unes substàncies ensucrades que segreguen per mitjà d’unes glàndules especials, mentre que d’altres actuen com a autèntics paràsits ja que s’alimenten en realitat de les larves del niu de les formigues. Altres coleòpters són paràsits dels nius d’altres insectes, especialment himenòpters (melòids, clèrids, etc.), i, fins i tot, algunes espècies exòtiques s’alimenten a expenses dels tèrmits (termitòfils). Alguns altres viuen als nius de micromamífers i d’ocells, i poden establir amb els seus hostes relacions de parasitisme o de comensalisme; Leptinus (un leptínid) és un dels pocs coleòpters ectoparàsits de vertebrats, que parasita petits mamífers, enganxat al seu pèl, des d’on s’alimenta de les desquamacions de la pell, restes de menjar i excrements del seu hoste. També es comporten com a comensals els anomenats coleòpters foleòfils, que viuen dins dels cataus de mamífers, i els nidícoles, que viuen dins dels nius d’ocells; en ambdós casos s’alimenten de restes de matèria orgànica que s’hi acumulen, sense perjudicar aquells qui els hostatgen; la majoria dels coleòpters foleòfils i nidícoles són histèrids, catòpids, ptínids, estafilínids, trògids i escidmènids.
Els hàbits alimentaris
Els coleòpters, com tota la resta dels insectes, han desenvolupat una sèrie de models de conducta peculiars, en relació amb el tipus de vida i els hàbits alimentaris. Els seus sistemes de predació i de defensa marquen les característiques etològiques de les espècies, juntament amb els de tipus sexual.

L’atac a productes alimentaris emmagatzemats, sobretot els de natura cel·lulòsica o amilàcia, és corrent per part de certs coleòpters. És el cas d’aquest tenebriònid (Tenebrio), les larves del qual són significativament anomenades cucs de la farina. Hom cria cucs de la farina també com a aliment per a animals carnívors de laboratori i com a material per a recerques genètiques i fisiològiques.
Ramon Torres

Els coleòpters florícoles són corrents a la nostra fauna com és el cas d’aquest buprèstid (Anthaxia hungarica, tanmateix, de larva xilófaga). Els coleòpters florícoles, al contrari que els himenòpters o els dípters, no són bons pol·linitzadors, car devoren els òrgans reproductors de les flors que visiten o els insectes pol·linitzadors que hi acudeixen.
Enric Curto
Molts escarabats són actius depredadors, tant en la vida larval com en l’adulta. Ataquen altres invertebrats més petits, sense que generalment tinguin gaires preferències per espècies concretes, però en canvi, sí que trien grups determinats. Alguns gèneres, com Pterostichus, Staphylinus, Bembidium i Nebria, capturen amb les mandíbules qualsevol presa que es posi al seu abast; d’altres, per contra, com els dels gèneres Cychrus, Phosphuga i Lampyris, cacen exclusivament cargols terrestres, Cicindela cerca petits dípters que caça al vol, Notiophilus captura només col·lèmbols, Dyschirius ataca larves d’estafilínids del gènere Bledius, etc. Tots els coleòpters carnívors fan la digestió extraoral, és a dir, regurgiten els sucs digestius dins de les preses i després n’absorbeixen els sucs interns i els teixits ja mig digerits. En alguns casos, no gaire corrents, les mandíbules poden tenir un canal intern que és connectat a una glàndula secretora de substàncies tòxiques o digestives, el qual introdueixen al cos de les preses, cosa típica del gènere Staphylinus i dels escarabats aquàtics Dytiscus, d’algunes espècies de cantàrids, drílids i lampírids, per exemple. Alguns escarabats molt petits, de les famílies dels escidmènids i pselàfids, s’han especialitzat a caçar àcars diminuts. Altres, com els coccinèl·lids i els cerilònids només cacen pugons i còccids (homòpters), de vegades amb una gran especificitat. Hi ha comportaments d’un grau elevat de sofisticació, com per exemple el d’algunes cleroïdeus, que emeten feromones que atrauen les seves preses.

Els hàbits depredadors són corrents en moltes espècies de coleòpters, com és el cas del caràbid Chlaenius velutinus, activíssim caçador d’invertebrats diversos, i habitant de llocs pedregosos immediats a riuets i punts d’aigua, del litoral a la muntanya mitjana. Segreguen productes tòxics per a defensar-se, a llur torn, d’altres depredadors.
Xavier Moreno.
El comportament alimentari dels escarabats fitòfags i sapròfags és molt més simple, atès que no necessiten altres mecanismes que els de localitzar la font d’aliment. En aquests casos, la digestió és interna, és a dir, es fa a dins del seu tub digestiu. Les espècies xilòfagues, que s’alimenten de fusta, tenen fongs i bacteris simbionts (micetomes i bacteriomes) dins dels tubs de Malpighi, al teixit adipós, en algunes cèl·lules intestinals i a les gònades, els quals produeixen substàncies necessàries per al seu metabolisme i enzims que afavoreixen o fan possible la utilització de determinats tipus d’aliment, com és la cel·lulosa, i que moltes vegades són transmesos d’una manera regular a la descendència.
Malgrat que, com hem vist, entre els coleòpters hi ha espècies causants de danys per als conreus, boscos i productes emmagatzemats, cal destacar el seu efecte útil com a pol·linitzadors o com a depredadors d’altres espècies perjudicials. El gènere Calosoma, per exemple, s’alimenta especialment de les erugues de les processionàries del pi i de la cua peluda del suro, per la qual cosa és utilitzat per a combatre aquests flagells. També alguns coccinèl·lids són actius depredadors d’àfids i còccids, i en regulen les poblacions; altres, com alguns clèrids, cacen les larves d’altres insectes, especialment xilòfags. D’altra banda, els insectes xilòfags que minen els troncs caiguts poden ésser considerats útils, ja que col·laboren a la disgregació de la fusta i acceleren el retorn de la matèria orgànica al sòl; una tasca semblant duen a terme els copròfags i els necròfags, que descomponen la matèria orgànica d’origen animal. Tots col·laboren al manteniment d’un equilibri energètic entre els productors i els destructors.
Els sistemes de defensa

La defensa per mimetisme no és rara en els coleòpters. Així, aquest cerambícid (Clytus arietis) dels alzinars, rouredes i fagedes (a la fotografia apareix sobre una faja) pren l’aparença d’un himenòpter picador (vespa) per tal de dissuadir possibles predadors.
Josep M. Barres.
A més dels sofisticats sistemes d’atac, destinats gairebé exclusivament a la captura de les preses, els coleòpters han desenvolupat també uns sistemes de defensa independents dels hàbits de caça, que poden ser de natura química en uns casos o passius en d’altres, com passa a les espècies críptiques i mimètiques. Sigui quin sigui el sistema de defensa, la seva finalitat és evitar caure preses d’altres insectes predadors i, especialment, d’alguns vertebrats insectívors, com ocells, rèptils i micromamífers. Un sistema de defensa molt generalitzat entre els escarabats és el de regurgitar enzims digestius per la boca, que són corrosius i tòxics per a altres organismes, fenomen que és corrent en molts caràbids, sílfids i melòids, entre molts altres. Un mecanisme de defensa especialitzat, de natura química, és el conegut amb el nom d’autohemorrea, que consisteix a emetre hemolimfa, que en aquest cas conté productes tòxics, a través de determinats punts del cos: per les articulacions, el prostèrnum, la línia dorsal (en algunes larves), etc.; presenten aquesta facultat les marietes (coccinèl·lids), els melòids, els crisomèlids, els tenebriònids, els lampírids, etc. Un mecanisme semblant és el d’exhalar tòxics o repel·lents per tot el cos, cosa en què s’han especialitzat alguns coleòpters que tenen unes glàndules especials, generalment pigidials o anals, que expulsen i projecten aquelles substàncies corrosives a distància; és el cas dels gèneres Carabus, Calosoma, Dytiscus, Blaps, etc.; aquesta ejecció pot fer-se d’una manera explosiva, com passa en diferents gèneres de braquinins (Brachynus, Aptinus, Pheropsophus) i altres escarabats, coneguts amb el nom d’escopeters o petadors. La finalitat d’aquests mecanismes sempre és la mateixa: espantar i allunyar els seus possibles depredadors.
Pel que fa als sistemes de defensa passius, el més simple és la immobilització reflexa, que consisteix a fer-se el mort com a reacció davant les situacions de perill: es deixen caure a terra i hi resten totalment immòbils. Un altre mecanisme del mateix tipus és la cripsi, és a dir, l’intent de passar desapercebuts, per la qual cosa imiten els colors del substrat sobre el qual viuen o bé mostren coloracions disruptives, amb la finalitat de rompre la silueta o el contorn del seu cos per tal de quedar dissimulats sobre el fons; és el cas de la rosalia (Rosalia alpina), d’algunes Cicindela, de molts coccinèl·lids, clèrids, etc. Obeint al mateix principi, els fitòfags solen ésser acolorits d’acord amb els colors de les plantes, i els terrícoles solen ser de colors foscos. Un altre sistema de passar desapercebuts és el d’amagar-se en el substrat durant els períodes d’inactivitat, construir-se refugis o col·locar-se al damunt de materials que puguin camuflarlos. Són especialment curiosos els casos dels escarabats que combinen la cripsi amb l’homotípia, com fan alguns coleòpters tropicals, que imiten la forma de les fulles. El mimetisme és un altre sistema de defensa passiu que té la finalitat contrària: fer-se notar. En aquest cas, es tracta d’escarabats molt ben acolorits i vistents que pretenen advertir les preses de la seva perillositat, ja que porten a l’hemolimfa substàncies que els són tòxiques; aquest cas és poc freqüent entre els coleòpters, però s’observa, per exemple, en els lícids. Altres s’aprofiten d’aquests símbols de perillositat, i, sense ésser perillosos, imiten la forma del cos, els colors i fins i tot la conducta dels insectes advertidors, per tal d’ésser confosos amb ells pels depredadors; són freqüents aquests mimetes o imitadors entre els coleòpters: molts cerambícids (els de la tribu dels Clytini) copien els dibuixos i la conducta dels himenòpters de les flors, per exemple.
Els sistemes de reclam: fotoemissió i estridulació
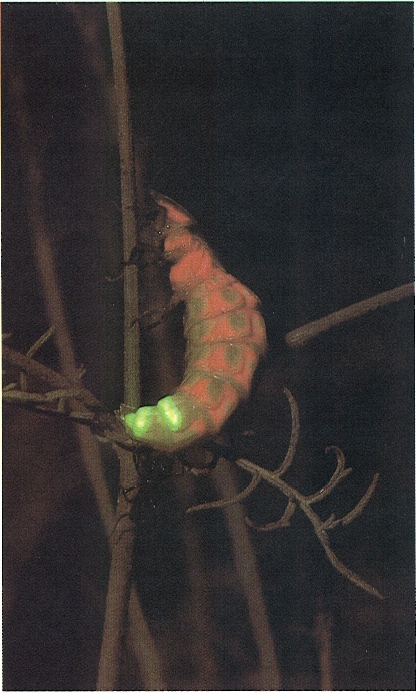
L’emissió de llum caracteritza els lampírids, justament anomenats cuques de llum o lluernes, proveïts d’òrgans fotoemissors a l’extrem de l’abdomen. Presenten aquesta facultat tant les larves com els adults, bé que és més accentuada en aquests darrers, car es tracta d’un recurs que facilita l’aparellament. L’exemplar de la fotografia correspon a una femella o una larva (són d’aspecte molt semblant) car els mascles són alats i tenen la fesomia normal d’escarabat.
Oriol Alamany.
Un cas curiós és la capacitat que tenen alguns coleòpters d’emetre llum, que només presenten, dins de la nostra fauna, els lampírids o cuques de llum, també anomenats lluernes. L’emissió de llum s’interpreta com un reclam sexual, que afavoreix el trobament dels mascles i les femelles. Alguns coleòpters tropicals també emeten llum, la intensitat i la durada de la qual és més notable que en els nostres lampírids. També hi ha coleòpters capaços d’emetre sons, hom creu que com un mecanisme d’alarma o per a localitzar la parella o, també, per a delimitar el territori particular; els emeten picant el substrat amb alguna part del cos, fregant dues parts endurides del cos, o bé per mitjà d’òrgans estriduladors especials. Tenen capacitat d’emetre sons alguns cerambícids, elatèrids, caràbids, i escarabèids, entre altres.
Sistemàtica i filogènia
La classificació que emprem en aquesta obra és l’establerta per Crowson (1955) i modificada posteriorment (1981) pel mateix autor. Cal observar que de les 167 famílies que ell establí, únicament esmentarem les que es troben representades dins de l’àmbit geogràfic dels Països Catalans. Alhora, hom ha introduït algunes modificacions a la classificació esmentada. L’ordre dels coleòpters comprèn, doncs, quatre subordres: els arcostemats (Archostemata), els adèfags (Adephaga), els mixòfags (Myxophaga) i els polífags (Polyphaga). Alguns autors han considerat que dins dels polífags podien separar-se dos subordres: els heterogastres (Heterogastra) i els haplogastres (Haplogastra), segons com fos l’estructura abdominal, cosa que posteriorment fou desmentida per d’altres autors.
Hi ha dues teories que destaquen d’entre les moltes que hom ha arribat a pensar sobre l’origen dels coleòpters. La primera els relaciona amb els blatoïdeus (Blattoidea), ja que hom dóna més importància a la forma del cos i a l’estructura dels èlitres; la segona, els relaciona amb els neuropteroïdeus (Neuropteroidea), i es fonamenta en els tipus de larves i de desenvolupament postembrionari. Actualment, és més acceptada la segona, segons la qual cal cercar relacions entre coleòpters i megalòpters.
El fòssil més antic que hom pot atribuir als coleòpters és Tshekardocoleus, procedent del Permià inferior, material que segons Crowson (1960), representa el pas intermedi entre els coleòpters del tipus cupèdids (Cupedidae, dels arcostemats) i els megalòpters del tipus Corydalis, la qual cosa referma la segona de les dues teories esmentades.
Hom considera els quatre subordres com a línies evolutives independents originades a partir d’un ancestre comú. De les quatre línies, els arcostemats són els més primitius: hom en coneix fòssils del Permià. Les línies restants ja s’havien diferenciat al Triàsic, donat que durant el Juràssic els adèfags convivien amb els polífags més antics; hom considera que en aquesta època ja s’havien diferenciat la majoria de les famílies que actualment coneixem. Al Terciari, i concretament a l’Oligocè, els coleòpters ja mostraven una gran diversitat ecològica i alimentària, i, a partir d’aquest període, encara es diversificaren més, fins a adquirir la seva importància numèrica actual.