Els vegetals i la producció primària
En contrast amb la costa, les aigües d’enfora, especialment les més blaves, semblen particularment desproveïdes de vida. El contrast entre masses d’aigua de diferent aspecte (més blaves i més verdoses, per exemple), i especialment quan aquest caràcter s’associa amb la presència o, amb l’alternació de les poblacions de peixos (com quan les «aigües blaves» amb les tonyines s’apropen a la costa per sant Josep, o bé hi ha anys bons o dolents, amb relació a la sardina, el seitó, o altres peixos, o si hi ha més o menys meduses en certs anys i indrets, etc.) són coses que fan pensar que l’aigua no és solament un receptacle, sinó també una font de vida que, naturalment, ha de tenir una base microscòpica. Aquest caràcter microscòpic de la vida es fa visible quan la concentració dels organismes és extraordinària. A la primavera, al llarg de les nostres costes, l’aigua sovint es veu rosada a conseqüència de l’acumulació de Noctiluca, un organisme semblant a un «préssec amb cua de porc», de mig mil·límetre o menys, que té tendència a flotar i que, segons el moviment de l’aigua, s’acumula prop de la costa. Aquesta és una manifestació modesta del que sovint s’anomenen «aigües roges» o «marea roja» o «purga de mar» (a Galícia), produïdes per l’acumulació d’organismes acolorits en quantitats molt grans (milers o desenes de milers de cèl·lules per centímetre cúbic) i que criden més l’atenció si són vermells que si són verdosos o groguencs, com s’escau més sovint, per exemple a l’interior de ports, com és ara el de Barcelona, on organismes dels gèneres Scrippsiella, Prorocentrum i Chattonella i també d’altres, fan tèrbola l’aigua. Al port de Barcelona s’han arribat a medir concentracions de clorofil·la de fins 34 mg/m3. Tot això és conegut d’antic i Ramon Muntaner ja en parla, tot referint-se a una observació d’aigües roges a la mar Egea, atribuïda a la sang dels màrtirs innocents i, segons sembla, amb suposades propietats remeieres, ja que la gent de mar procurava d’empapar-ne mocadors per a treure’n algun benefici.
L’estudi del fitoplàncton
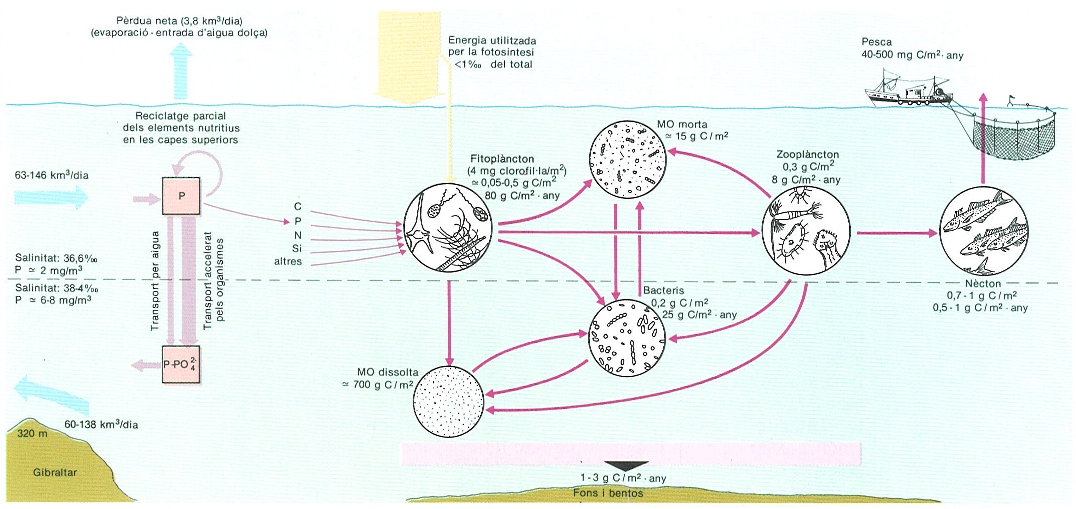
Model general de l’intercanvi de la mar Mediterrània amb l’oceà Atlàntic i del sistema de producció a la Mediterrània. Les xifres són orientadores i aproximades.
Ricardo Génova, a partir de fonts diverses.
De fet, l’estudi del plàncton entrà a la ciència en el curs del segle passat, durant el qual es recolliren molts organismes, i es descriviren i es dibuixaren, sovint amb minuciositat exquisida, en obres de gran valor, fins i tot per a bibliòfils no científics. Pel que fa als organismes vegetals són més petits que els animals, de manera que solament es podien recollir emprant xarxes en forma d’embut fetes de tela molt fina. Durant molt de temps, la millor tela de què es podia disposar era la de seda destinada a tamisar farina, de manera que, sovint, distincions proposades pels planctòlegs, com ara la distinció entre categories de plàncton per la seva mida (microplàncton, nanoplàncton, etc.) eren més relacionades amb la tècnica farinera que amb la lògica científica. De fet, durant molt de temps s’usaven malles entre 0,06 i 0,1 mm; ara n’hi ha de més fines, de 0,04 mm. Les xarxes construïdes amb aquest material deixaven passar molts organismes de mida menor; però una part d’aquests quedaven, malgrat tot, retinguts, ja que altres organismes majors anaven tapant el teixit i ho permetien; però, evidentment, la representació relativa no era correcta. Tampoc no podia ser correcta qualsevol avaluació de l’aigua filtrada, a base de multiplicar la secció de la boca de la xarxa o del colador per la longitud recorreguda, perquè no es tenia en compte la filtració parcial i el reflux de l’aigua. Amb finalitat taxonòmica, l’estómac de les salpes i d’altres animals pelàgics es podia utilitzar per a cercar organismes molt petits, que s’escapaven de les xarxes dels investigadors, però no dels aparells filtradors de captura d’aquells animals. Amb aquestes tècniques primitives no solament es pogueren descriure molts organismes planctònics, sinó que també es començà a tenir una idea de com les diferents espècies —el mateix que en la vegetació terrestre— reflectien les propietats de l’entorn, i s’aprengueren a diferenciar tipus d’aigua i tipus de plàncton. Per exemple, les diatomees es multiplicaven i es trobaven en quantitat a les mostres d’hivern o amb intensa barreja vertical, mentre que els grossos i bonics dinoflagel·lats eren més freqüents relativament —encara que llur quantitat total fos baixa— en aigües blaves i estratificades.
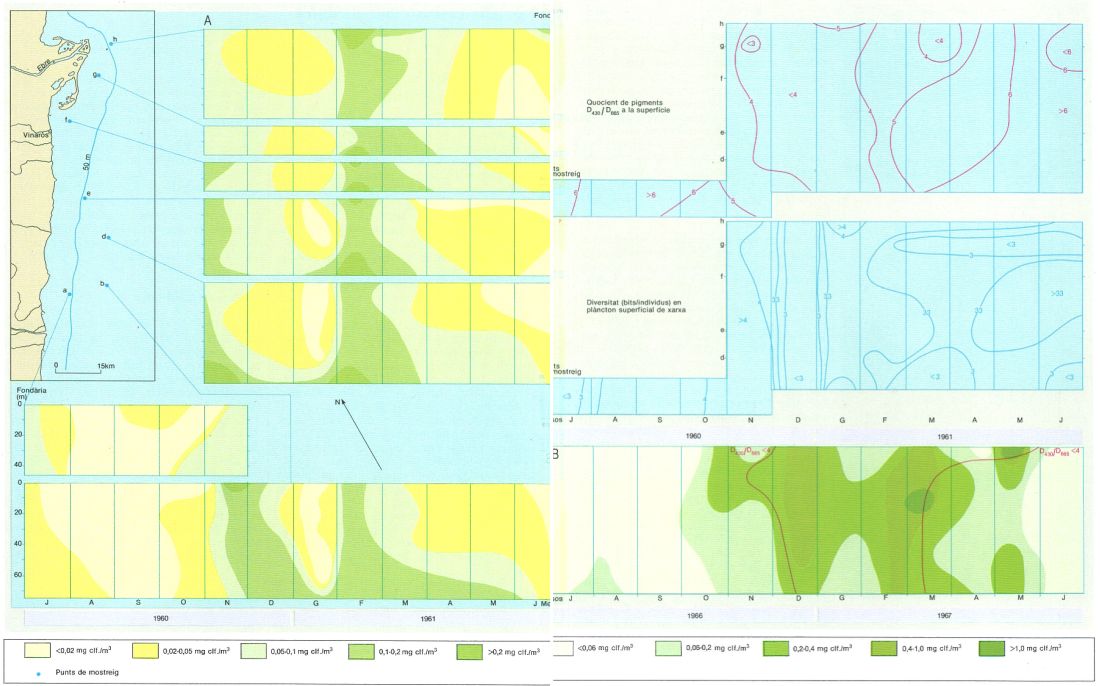
Cicle anual del desenvolupament del fitoplàncton, en mil·ligrams de clorofil·la/m3, a la zona marina del sud de l’Ebre indicada en el mapa (A) i, a la dreta, canvis de diversitat i del quocient D430/D665 associats. A sota (B) cicle anual del desenvolupament del fitoplàncton en aigües no gaire allunyades de la costa de davant de Barcelona. En els gràfics A s’observen dos màxims de concentració de clorofil·la: un a la tardor (novembre-desembre), quan es trenca la termoclina, i un altre a l’hivern (febrer), que s’ha relacionat amb l’aportació de nutrients deguda a l’aflorament costaner. En el gràfic B s’observa la presència de màxims de clorofil·la a la tardor (desembre), l’hivern (febrer-març) i la primavera (maig).
Carto-Tec, original de R. Margalef i J. Castellví.
Generalment, la recerca oceanogràfica des de la costa s’orientava a fer estudis per a determinar la fertilitat de la mar i la capacitat per a mantenir poblacions de peixos explotables. Dintre el programa entrava estudiar periòdicament mostres o «lots» de peixos, obtinguts sovint en el mercat local, i sortir periòdicament per fer algunes observacions físico-químiques i capturar plàncton amb una o diverses xarxes: per exemple, una xarxa de poc diàmetre i de malla fina per al fitoplàncton, i una xarxa de major diàmetre, fins a un metre o més, i de malla més grollera (0,4 mm o més) per al zooplàncton. L’anàlisi d’un seguit d’aquestes mostres permetia de descriure un «cicle anyal» del plàncton local. Aquesta ha estat una pràctica molt general, que ha donat alguns resultats de valor sinòptic, i que, a la nostra àrea, s’ha fet a Banyuls de la Marenda, Mallorca, Castelló, Vinaròs, Blanes, Barcelona, i encara d’altres localitats. Les grans expedicions oceanogràfiques complementaren aquesta visió, molt costanera i localista, amb una altra visió «venint de mar», amb més o menys fortuna. A la nostra mar, les expedicions daneses, entre el 1908 i el 1930, aportaren resultats i descripcions aprofitables i fins i tot, hipòtesis més o menys encertades, però, almenys, suggeridores de noves recerques.
Després, principalment la introducció del microscopi invertit o del mètode d’Utermöhl, científic alemany que morí el 1985, permeté d’afinar més. Aquest procediment implica la sedimentació total del fitoplàncton que hi ha en un petit volum d’aigua (generalment de 20 a 100 ml), després d’una fixació adient (generalment amb iode), de manera que el fitoplàncton es pot examinar i comptar, en la seva totalitat, sobre el fons d’una cubeta o un cilindre que ha rebut la mostra. Molts milers de mostres s’han observat a la Mediterrània occidental, fins al punt que el coneixement que es té de la composició taxonòmica del nostre fitoplàncton pot ser encara incomplet, però no és negligible.
Podríem dir que hem arribat a un moment en què la rutina oceanogràfica de l’estudi del fitoplàncton s’ha estabilitzat entorn dels vèrtexs d’un triangle. Un d’ells és representat per l’estudi taxonòmic de les mostres, la composició en termes d’espècies i la quantitat de cadascuna d’elles. Un altre vèrtex és la concentració de pigments importants en la fotosíntesi i, principalment, de la clorofil·la; el darrer vèrtex es refereix a mesures de la producció, de la capacitat de síntesi de matèria orgànica manifestada per altres mostres paral·leles. Aquesta representació en forma de triangle és útil, perquè permet de reunir entorn de cada vèrtex diverses modalitats tècniques que cerquen una mateixa finalitat. És clar que l’estudi del fitoplàncton agafat amb xarxa se situa al costat de l’estudi del plàncton total al microscopi invertit i que aquestes operacions són, fins a un cert punt, complementàries, perquè la xarxa permet de detectar la presència d’espècies que són massa rares perquè es trobin en petits volums d’aigua, però que, tanmateix, poden tenir una gran significació per la seva presència. L’estudi dels pigments es pot fer per extracció, emprant dissolvents adients; però, també, per mesura directa de la fluorescència de l’aigua i del plàncton que porta en suspensió. La producció primària es mesura generalment amb un traçador radioactiu, el 14C (isòtop de pes atòmic 14 del carboni); però és avaluable també, si el fitoplàncton és abundant, mesurant els bescanvis d’oxigen en mostres confinades, i fins per mètodes indirectes, com és ara la taxa de consum de nodriment en el medi.
La mateixa representació triangular és bona per a recordar que les relacions entre cada parell de magnituds no es poden considerar com a factors de pas. Ens hem de guardar de voler deduir la quantitat de cèl·lules a partir de la quantitat de clorofil·la, o bé d’avaluar la capacitat productiva a partir del nombre de cèl·lules o de la quantitat de pigments. De fet, les relacions entre cada parell de magnituds són summament indicatives de les característiques de les poblacions. Les cèl·lules velles tenen menys clorofil·la que les joves; amb relació al volum, les més grosses tenen menys clorofil·la que les petites i, sovint, les cèl·lules que viuen amb poca llum tenen més concentració de clorofil·la. Com a xifra per a fixar la imaginació, podríem dir que el valor mitjà podria ser a la ratlla d’1 picogram (1 microgram és un milionèsim de gram, i un picogram un milionèsim de microgram), però tant pot ser una mica menys com cent vegades més. Semblantment, la quantitat, en grams, de carboni fixat per un gram de clorofil·la durant una hora a la llum pot arribar a 11 com a màxim, però valors de 3 a 5 ja són alts i, generalment, es troben xifres inferiors, especialment si la llum és feble. La relació entre la producció i el nombre de cèl·lules s’estableix, naturalment, en funció del nombre de divisions per dia i, d’una manera general, hom podria escriure:
d = (1/0,69) ln[( (hores de fotosíntesi per dia / relació pes carboni: clorofil·la) × Z) ] + 1
on d és el nombre de divisions per 24 hores i Z els grams de carboni assimilats per gram de clorofil·la i hora. Les hores de fotosíntesi per dia corresponen, aproximadament, a les hores de llum, i la relació ponderal carboni/clorofil·la sol variar entre 30 (plàncton molt productiu, diatomees) i 100 (plàncton més parsimoniós i, principalment, dinòfits o dinoflagel·lats).
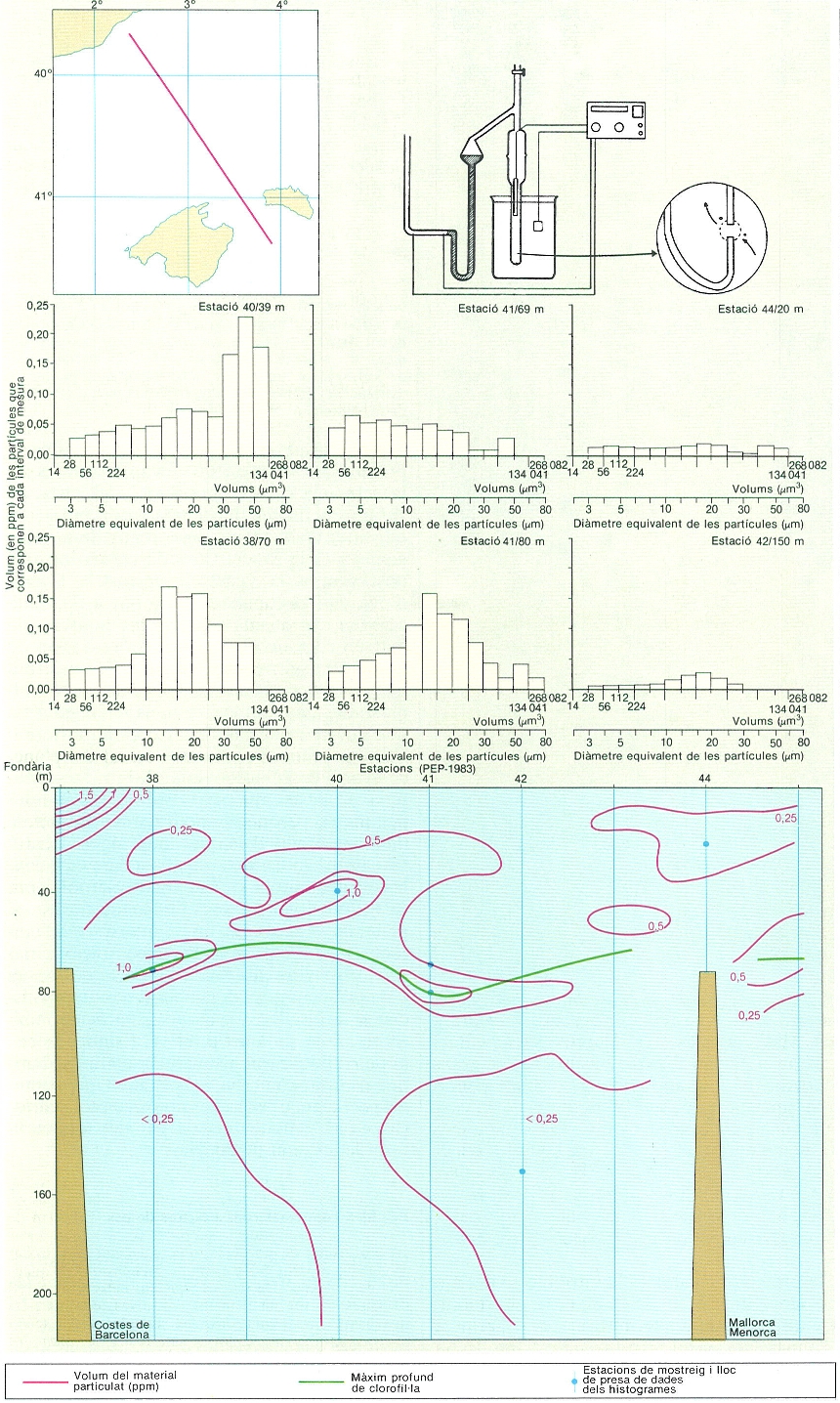
Fonament del comptador Coulter (a dalt a la dreta) i mesures de distribució de partícules en el transsecte que indica el mapa (a dalt a l’esquerra), realitzat durant la campanya PEP («Producció Estival Profunda»), de 1983. La mostra continguda en el vas (a dalt, a la dreta) és xuclada cap a dins del tub submergit a través d’un orifici calibrat (detall). A dins del tub i del vas de la mostra hi ha elèctrodes. L’aigua que passa per l’orifici arrossega partícules que desplacen un volum d’electròlit (aigua de mar) igual al volum de la partícula que passa; això produeix una pulsació elèctrica proporcional al volum desplaçat, pulsació que es compta i classifica segons la grandària. Al mig (histogrames) es mostra com es reparteix el volum total del material particulat (en parts per milió en volum, ppm) segons la grandària de les partícules constituents. Cada histograma correspon a una mostra indicada per un punt en la secció vertical (a baix) feta entre Barcelona i Mallorca. Les escales, en els histogrames, són dobles: la primera indica el volum (en µm3) de les partícules que cauen en els extrems dels intervals de grandària, mentre que la segona indica el diàmetre equivalent de les partícules (en µm) que és el que tindria una partícula del mateix volum que fos esfèrica. Cal fixar-se que en cada interval el volum de la partícula mitjana és el doble del de l’interval anterior. Sobre el transsecte (a baix) s’observa que hi ha un màxim de volum de sèston prop de la costa, a nivells superficials, i un altre aigües enfora a nivells més profunds, acompanyant el màxim profund de clorofil·la.
Carto-Tec, original de Jordi Flos
L’acumulació gradual de molta informació permet de formar-se una idea de com les característiques més importants del plàncton varien en funció del temps i de l’espai. Hi ha, indubtablement, una regularitat, però aquesta és lluny del tipus de regularitat que observem a la vegetació terrestre. Canvis d’una entitat comparable als que a terra necessiten uns quants anys, es produeixen a la mar en cosa de dies, si no d’hores. L’oceà i la vida pelàgica tenen l’aparent irregularitat que és característica dels canvis del temps atmosfèric, però també el tipus de regularitat que associem al pas dels fronts atmosfèrics. En línies generals i a la nostra regió, la concentració de cèl·lules varia entorn d’una vintena o poc menys per mil·lilitre (20 milions per metre cúbic, que podrien ser entorn de 0,02 mg de clorofil·la per m3 d’aigua) i entre 50 i 100 vegades més (alguns milers de cèl·lules per mil·lilitre i 1,5 o 2 mg de clorofil·la per m3). Però la major part de l’aigua conté poc plàncton, s’acosta al nivell inferior que, per tant, és molt a prop de la concentració mitjana, i els valors alts tenen l’aspecte de taques o de pulsacions relativament discontínues. És clar que els llocs confinats i particularment fèrtils, com és ara l’interior d’alguns ports, poden allotjar un plàncton molt més dens, que es fa aparent per la coloració que comunica a l’aigua; però això no és comú i, en tot cas, la superfície afectada per tals fenòmens és pràcticament negligible.
Factors i mecanismes de producció als ecosistemes pelàgics
El plàncton, com tots els ecosistemes, s’organitza entorn d’un eix vertical definit per les direccions de la llum i de la gravetat. El gradient de llum defineix la possibilitat de producció; la gravetat assenyala la direcció de sedimentació, d’exportació natural de matèria orgànica assimilada.
El gradient de llum
A mitjan segle XIX, l’astrònom jesuïta Secchi va introduir un mètode molt senzill per a mesurar la transparència de l’aigua, consistent en l’observació de la fondària fins on era visible un disc blanc (s’usen de 30 cm de diàmetre) penjat horitzontalment al cap d’una corda.
A la Mediterrània, d’una manera especial a l’aigua blava, el disc pot ser visible fins a més de 40 m de llargada del cordill. S’accepta que a 2,7 vegades la fondària de visió del disc de Secchi arriba fins a 1’1% de la radiació de la superfície, que pot ser una bona estima del límit de producció per al plàncton. Per tant, la producció neta del plàncton, la seva assimilació en excés sobre la respiració, es podria produir fins a uns 40 × 2,7 = 110 m, aproximadament.
La concentració de nutrients
De fet, com que durant una bona part de l’any gairebé no queda nutrient (fosfat, compostos de nitrogen) a les aigües superficials, la producció es dóna, i les cèl·lules s’acumulen, just en el límit entre la caiguda de la llum i l’augment cap avall del nutrient, un límit fluctuant, almenys a causa del ritme diari de producció i regeneració, límit que va a parar a una fondària considerable, més avall dels 60 m. Allí hi ha sovint un estrat més aviat de poc gruix, on el fitoplàncton no és tan rar com en estrats immediats. Aquest màxim profund de clorofil·la s’ha trobat en mars diverses, especialment a les que són relativament pobres o oligotròfiques, com és ara l’oceà Pacífic, i s’ha reconegut a la nostra mar. Val la pena de fer remarcar que la seva troballa ha estat relativament tardana, perquè generalment no es pretenia mai de pescar fitoplàncton en fondària per mitjà de xarxes. Més endavant, en agafar mostres d’aigua amb ampolles, la normalització habitual demanava ampolles a 60, 80, 100 m, però no a fondàries intermèdies, que és on, precisament, se situava el màxim profund de clorofil·la, estructura que, d’altra banda, sovint és subtil o de poc gruix.
Observacions a la mar i una infinitat d’experiments mostren com la producció és en funció, alhora, de la llum i de la concentració de nutrients, bàsicament de la disponibilitat de fosfat. Pràcticament, la producció es relaciona amb el producte de la llum per la concentració de nutrients, tal com queda molt clar en comparar el màxim profund de clorofil·la amb les respectives distribucions de la llum i dels elements limitants més importants.
La sedimentació del plàncton i el reciclatge
Sovint, la relació entre fitoplàncton i concentració de fosfat —o de compostos de nitrògen— no és fàcil d’establir o de reconèixer. De vegades, tot depèn de l’escala amb la qual es fan les observacions. Per exemple, és indubtable que, en general, quan hi ha més nutrients o on n’hi ha més el plàncton creix més, i hi ha més fitoplàncton. Però si ens ho mirem amb una dimensió més petita, per exemple en una anàlisi detallada al llarg d’una trajectòria feta amb cura, podrem trobar claps on hi ha molt plàncton i pocs nutrients i d’altres amb concentracions més elevades dels elements necessaris i amb poc plàncton. Ho expliquem en el sentit que, en uns llocs, hi ha fitoplàncton que ja ha consumit o ha absorbit els elements que tenia a la seva disposició, mentre que, en altres llocs els elements són encara a l’aigua, perquè el plàncton no ha tingut temps de créixer, cosa que pot ser certa. No obstant això, el detall d’aquesta distribució i dels núvols o les petites estructures que reflecteix ens fan pensar que el món del plàncton és essencialment dinàmic, com una mar de núvols, on els processos biològics creen contínuament estructures que s’esvaeixen més o menys aviat, com en un meravellós castell de focs artificials.
El fitoplàncton es nodreix, en certa manera, de materials que pugen d’una fondària relativa, de formes diverses; d’altra banda, el fitoplàncton té una certa, certíssima, tendència a sedimentar-se. Però aniríem errats si volguéssim fer un balanç molt senzill amb els materials entrats de fondària i amb totes les cèl·lules o bé que se sedimenten o bé que són consumides per animals. De fet, les capes superiors il·luminades de l’oceà són el lloc d’un reciclatge important. Molts organismes del fitoplàncton són poc mengívols i mantenen la seva posició perquè neden activament, com els dinoflagel·lats en general. Les mateixes diatomees excreten matèria orgànica a la part alta, i si els animals se les mengen, una part del material corresponent retorna a l’aigua, a través de les excrecions dels mateixos animals. Això vol dir que, si per exemple, un àtom de fòsfor és introduït de capes profundes a la capa fòtica o il·luminada, abans que aquest mateix àtom retorni a l’aigua profunda i fosca pot haver circulat unes quatre vegades —una xifra raonable per a la Mediterrània— pel cos d’algun productor primari. Aquest reciclatge local fa, doncs, més dispensable la fertilització, i com més voltes representi, més gran serà la producció real amb relació a l’aportament inicial de nodriment.
Un concepte tan important per a entendre el funcionament del sistema productiu del plàncton ha donat lloc a fer una distinció entre producció nova i producció reciclada, però és difícil d’estudiar experimentalment. Si tenim mesures bones de la producció i podem avaluar la fracció d’aquesta que abandona les capes superiors il·luminades cap al fons, podrem fer un model conceptual consistent; però és tan difícil de mesurar la sedimentació! Cal instal·lar i deixar en el lloc, a diferents fondàries, unes «trampes de sediment» de característiques adients, generalment en forma de cilindres o de pots fondejats a profunditats fixes, deixar-les un temps i després tancar-les, recuperar-Íes i analitzar-ne el contingut. Informació d’aquesta mena és pràcticament inexistent a la nostra àrea, de manera que les vegades que recicla el material en superfície s’han d’estimar per evidència indirecta i en comparació amb el que pot passar en altres mars de característiques semblants. Generalment, hom accepta que a les mars més fèrtils hi ha una major contribució de la població nova, i que a les mars més pobres, més oligotròfiques, una fracció relativament més gran de la producció que es mesura té el seu origen en el reciclatge de materials. Però aquesta distribució depèn molt del curs de l’any i també del lloc. És evident que en els moments de grans pulsacions de producció a càrrec de diatomees, la fracció de producció nova és més gran i que una gran part del material produït se sedimenta o és menjat per animals que migren cap avall. També en relació amb el front marí de què parlarem aviat, es pot dir que és versemblant que la producció nova tingui relativament més importància prop dels llavis del front, mentre que el reciclatge pot esdevenir relativament més notable en allunyar-se del front, amb relació, també, a canvis corresponents en la composició del fitoplàncton.
Pel que fa a aquest tema, i per acabar, cal dir que el tipus de reciclatge a la capa fòtica té molta importància en l’alimentació del bentos profund i, naturalment, del plàncton de fondària. Un sistema sota una certa tensió, que té molta producció nova i que també sedimenta molt, permet una vida més rica en aigües profundes; un sistema que produeix poc i encara la major part és fruit de reciclatge, no deixa gairebé res per a altri. Precisament, una hipòtesi que caldrà comprovar és que la posició del nostre front paral·lel a la costa deu quedar reflectida en la riquesa de la fauna de fondària, naturalment en el sentit que sota el front hom deu esperar una vida més rica.
La seqüència ideal
És obvi que la descripció que es pot fer ara de la producció i de la distribució del plàncton vegetal a la nostra Mediterrània ha de ser forçosament molt incompleta i provisional. És un fenomen amb tanta riquesa que qualsevol informació resulta parcial. És particularment sensible la manca d’informació sinòptica amb un grau suficient de precisió. De manera que solament es pot oferir una descripció provisional, encara que creiem que és prou raonable. Potser el més senzill és d’oferir una descripció de com es veuen o es veien els «cicles» des de la costa, que serà complementada més tard.
Els processos de fertilització
El cicle anual
Totes les observacions costaneres coincideixen a distingir un semestre productiu, que va de novembre a maig, i un semestre estèril, durant el qual hi ha poc plàncton i sembla que no passi res, en l’àmbit del fitoplàncton, s’entén. Les poblacions de fitoplàncton se succeeixen amb una gran diversificació durant el semestre productiu, en el curs del qual es mesuren produccions notables, fins a mig gram de carboni assimilat per m2 i dia. Hi ha una explicació hidrogràfica senzilla dels diferents episodis de floració de fitoplàncton que se succeeixen. El primer acostuma a donar-se pel novembre i coincideix amb el trencament de la termoclina, que permet una barreja vertical de la columna d’aigua local, en aigua no gaire fonda, amb la mobilització de la petita quantitat de nutrients que s’hagin pogut acumular en aigües de 50 i més metres de fondària. Es donen proliferacions de diatomees i, generalment, d’Asterionella glacialis, Thalassiothrix frauenfeldii, Thalassionema nitzschioides, i alguns Chaetoceros. És un episodi que no sol durar més d’un mes. Després, per Nadal, acostuma a venir una pausa. Més tard, en general al començament de febrer, s’observa la màxima producció de l’any, que coincideix amb vents forts de garbí, associats a situacions meteorològiques que es produeixen regularment en aquesta època. El resultat és un moderat aflorament d’aigua d’una certa fondària sobre la costa. L’èxit d’aquesta fertilització depèn de la riquesa de l’aigua aflorada i aquesta, segons sembla, és molt relacionada amb la intensitat de barreja vertical pel desembre precedent, sota l’acció del mestral i al llarg del front paral·lel a la costa. Si la barreja ha estat intensa, si s’ha produït aigua prou enriquida i a una fondària assequible, i si el vent és fort, hi ha pulsacions d’una producció extraordinària, que pot arribar fins al doble de la de novembre. El resultat són denses poblacions de diverses espècies dels gèneres Chaetoceros, Bacteriastrum i algunes Rhizosolenia, com R. stolterfothi, com també diversos flagel·lats i dinoflagel·lats que poden ser abundants. Aquest episodi és també de curta durada i per sant Josep ja ha arribat a la fi.
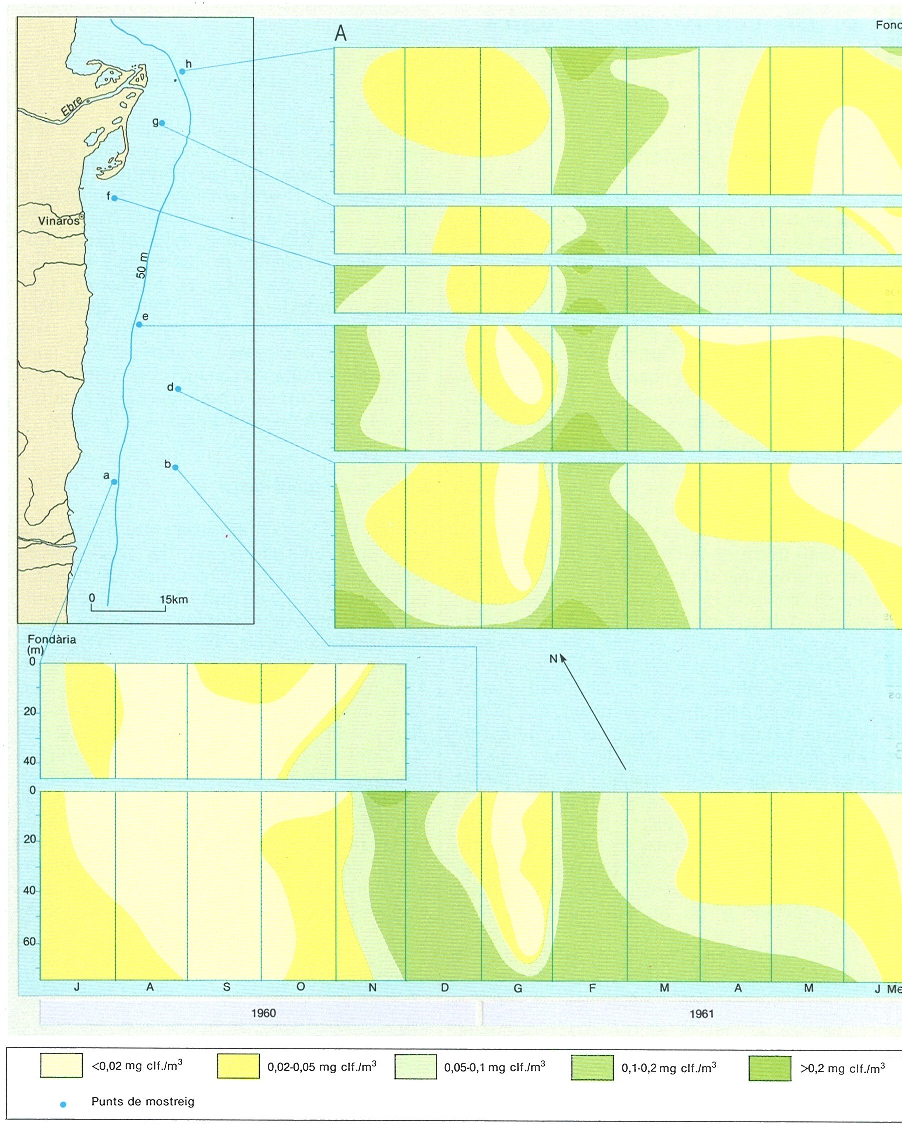
Cicle anual del desenvolupament del fitoplàncton, en mil·ligrams de clorofil·la/m3, a la zona marina del sud de l’Ebre indicada en el mapa (A) i, a la dreta, canvis de diversitat i del quocient D430/D665 associats. A sota (B) cicle anual del desenvolupament del fitoplàncton en aigües no gaire allunyades de la costa de davant de Barcelona. En els gràfics A s’observen dos màxims de concentració de clorofil·la: un a la tardor (novembre-desembre), quan es trenca la termoclina, i un altre a l’hivern (febrer), que s’ha relacionat amb l’aportació de nutrients deguda a l’aflorament costaner. En el gràfic B s’observa la presència de màxims de clorofil·la a la tardor (desembre), l’hivern (febrer-març) i la primavera (maig).
Carto-Tec, original de R. Margalef i J. Castellví.
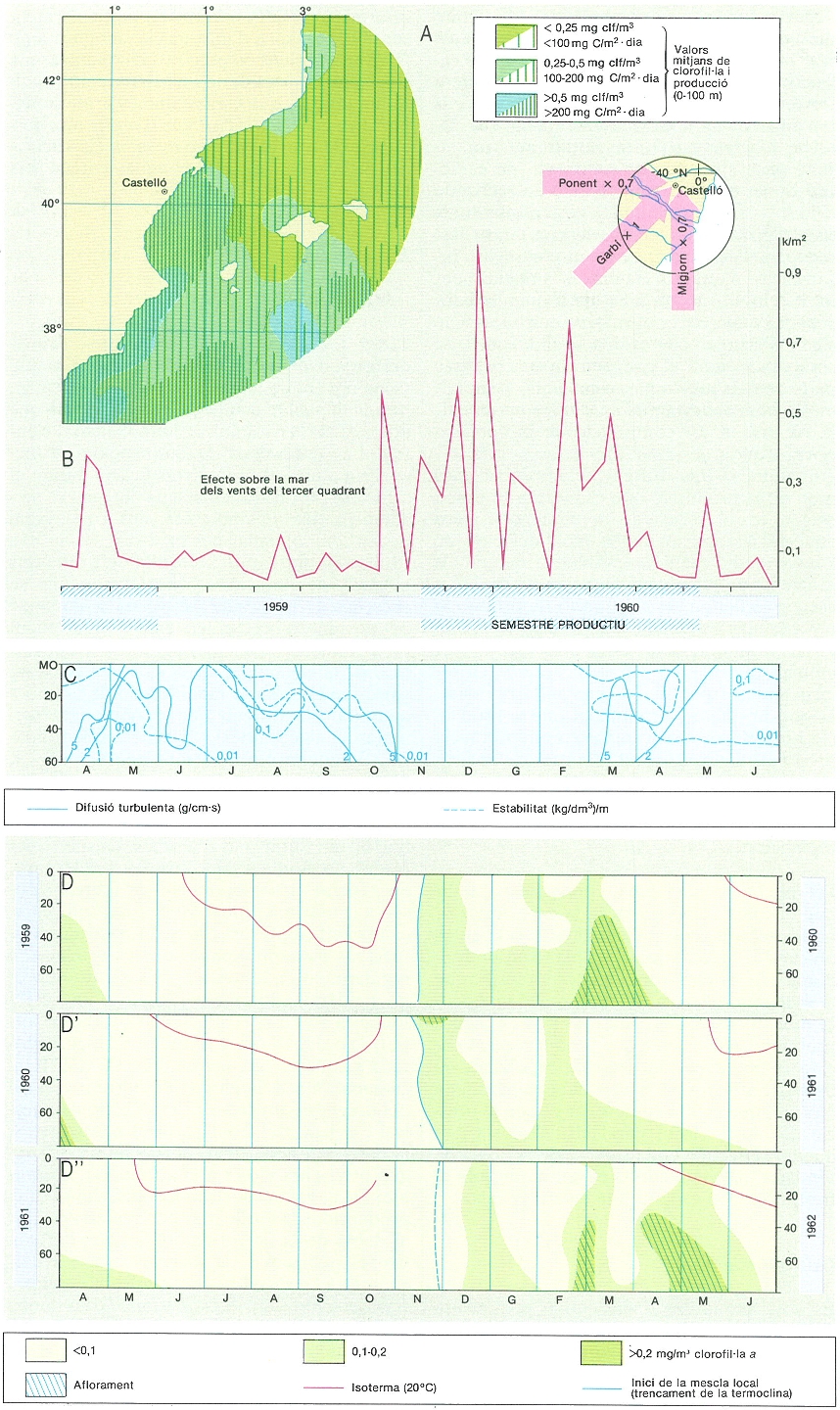
La distribució del fitoplàncton i de la seva producció no solament és desigual geogràficament, com mostra el dibuix A (corresponent a una situació estudiada a l’octubre de 1976), sinó que també varia amb el temps, com mostra l’esquema B (aigües costaneres del sud de l’Ebre), on la productivitat és més alta en el semestre que va de desembre a maig: la termoclina ha desaparegut, la qual cosa facilita la barreja vertical de l’aigua, que és promoguda principalment per vents del tercer quadrant (representats per mitjanes decennals de les observacions fetes, estimades en kg/m2 passant a través de l’escala de Beaufort i multiplicant per 1 els de garbí, més efectius en separar l’aigua de la costa, i per 0,7 el de migjorn i ponent; són xifres aproximades i cal tenir present que la resultant de tots els vents és del primer quadrant). En l’esquema C, l’època productiva correspon a valors alts de vents adequats i de difusió turbulenta i baixos d’estabilitat. Els gràfics inferiors (D, D’ i D"), del cicle del fitoplàncton en una estació representativa (a poca fondària davant de Castelló), al llarg de tres anys, assenyalen els moments del trencament de la termoclina i l’inici de la barreja vertical de l’aigua, amb l’aflorament (febrer-març) de les aigües d’origen profund, i, finalment, quan s’acaba el període d’alta producció, l’augment de la transparència de l’aigua, amb valors del disc de Secchi de més de 20 m.
Carto-Tec, original de l’autor
Precisament en aquest moment, dies més, dies menys, acostuma a produir-se un canvi que consisteix en l’arribada d’aigües blaves, que els pescadors coneixen bé, i que porten noves condicions de producció. Representen, en certa manera, una inversió de la pauta de circulació prevalent fins aleshores. Normalment, l’aigua costanera circula de NE a SW; ara hi ha una inversió que no sol durar molt de temps: les aigües noves contenen una indubtable barreja d’aigües atlàntiques i porten amb elles una concentració de fosfat que, si bé és baixa, ja, per consum dels organismes, amb relació a les aigües superficials de l’Atlàntic, és encara considerable si es compara amb els nivells generals a la Mediterrània. En aquestes aigües es multipliquen principalment diatomees d’una etapa avançada de la successió, com ara diverses Rhizosolenia, principalment alata, i les Nitzschia enllaçades en llargues cadenes.
Els nutrients aportats pels rius
Els rius contribueixen a la fertilitat marina. A la nostra mar tenim dos rius importants amb una forta influència, el Roine (cabal d’uns 50 km3/any), i l’Ebre (cabal entre 15 i 19 km3/any), i d’altres de menors. La circulació costanera a l’hemisferi nord, en equilibri amb la presència d’aigua dessalada prop de la costa, es dirigeix vers la dreta, de NE a SW, de manera que, en aquesta direcció, cal esperar la influència dels rius. La influència directa del Roine sobre la hidrografia local arriba almenys fins a Banyuls, i és ben marcada en la distribució del plàncton. Una tal influència es manifesta també en l’àrea de l’Ebre.
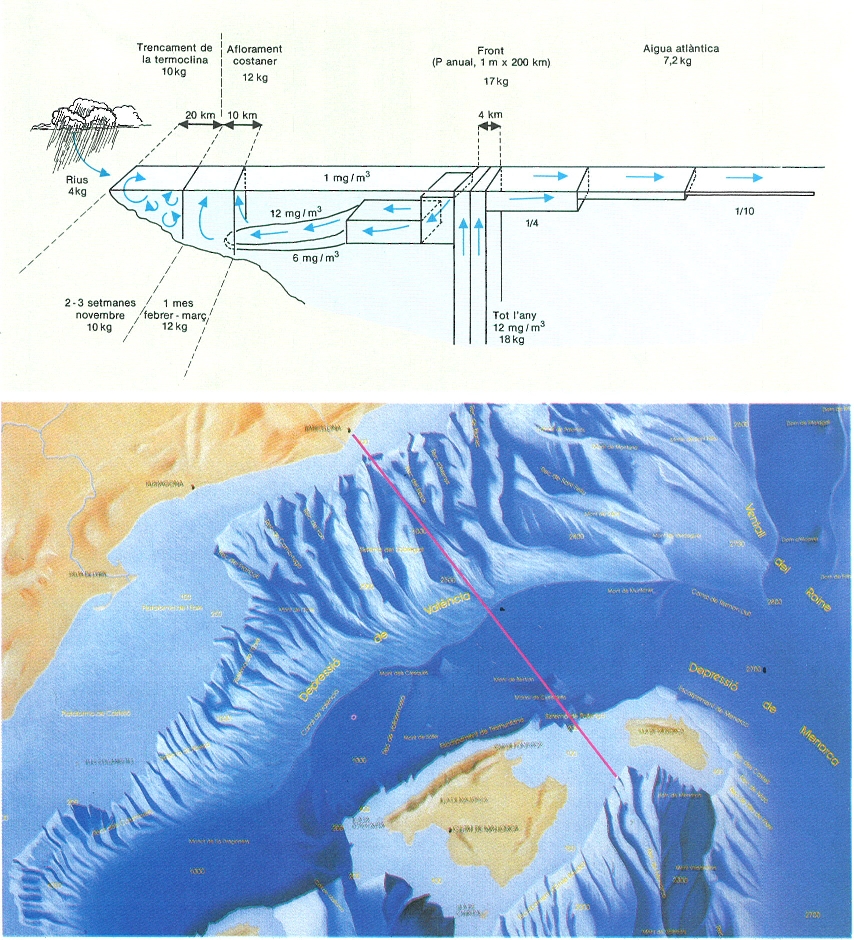
Representació idealitzada (a dalt) de la contribució de diversos fenòmens i mecanismes al subministrament de fòsfor «nou» a la capa fòtica de la nostra mar, representada ací, com a simplificació, per una trinxa d’un metre de gruixària que s’estendria des de la costa barcelonina fins a la mar balear (a baix), i de la superfície fins al fons. La suma de totes les aportacions de fòsfor dóna un total de 50 kg de l’element per a la superfície total de la trinxa.
Ricardo Génova i T&N&T comunicació / Cedit pel Servei de Medi Ambient, Diput. Barcelona.
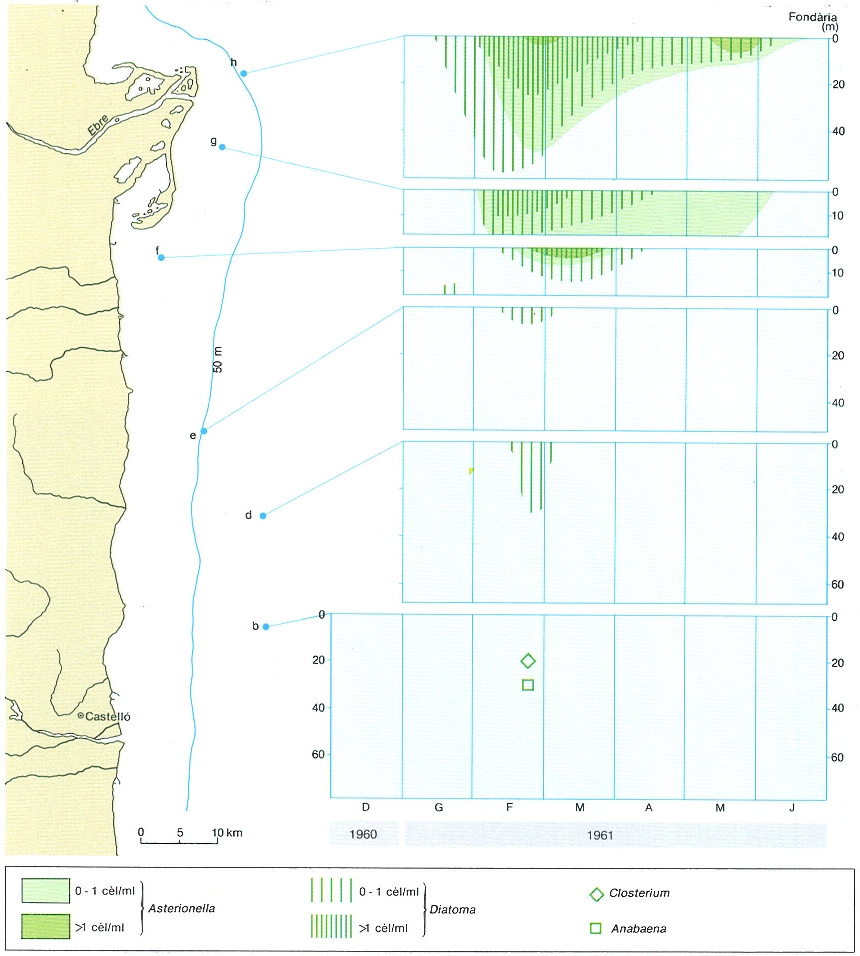
Distribució de les algues d’aigua dolça arrossegades per l’Ebre cap a la mar a les estacions indicades al mapa i en un període que va de desembre de 1960 a juny de 1961. Per a les estacions d-h s’indiquen les concentracions de cèl·lules de dues espècies, Asterionella formosa i Diatoma elongatum, que formaven gairebé la totalitat de les algues d’aigua dolça presents. Aquestes espècies no es trobaven a l’estació b, a la qual s’assenyala la presència d’algues d’aigua dolça pertanyents a d’altres gèneres (Closterium i Anabaena). Les estacions de mostreig i les dades corresponen als estudis duts a terme els anys 60 per l’equip d’oceanògrafs del Centre d’Investigacions Pesqueres de Barcelona (ara Institut de Ciències del Mar) i publicats a la revista «Investigacions Pesqueres».
Carto-Tec.
La influència del riu es reconeix també en la dispersió de les algues procedents dels embassaments de l’Ebre a les aigües de la Mediterrània. És interessant de conèixer la quantitat de fòsfor que les aigües del riu porten a mar, i com aquest fòsfor es reparteix a mesura que l’aigua del riu es dilueix en la mar. Les aigües de l’Ebre, del Ter, del Llobregat, contenen fins a 40 mg de fòsfor per m3; però, en arribar a mar, una part d’aquest fòsfor desapareix sense induir increments corresponents en el plàncton. Sembla que una bona part es precipita, tot contribuint al que es reconeix, més o menys immobilitzat, en els sediments costaners (davant de Castelló). Per això, es pot pensar que l’efecte fertilitzant dels rius es deu més a l’efecte de barreja que determina l’elevació d’aigua marina a partir d’un nivell relativament profund, que no pas a una contribució directa en nodriment. Ultra incidir sobre les estructures hidrogràfiques, especialment en temps de crescuda, creiem que la contribució dels rius és de segon ordre, malgrat que sovint es parla dels rius com a factors de contaminació i d’aportació de nodriment produït per l’activitat humana.
Un front marí com a font de nutrients
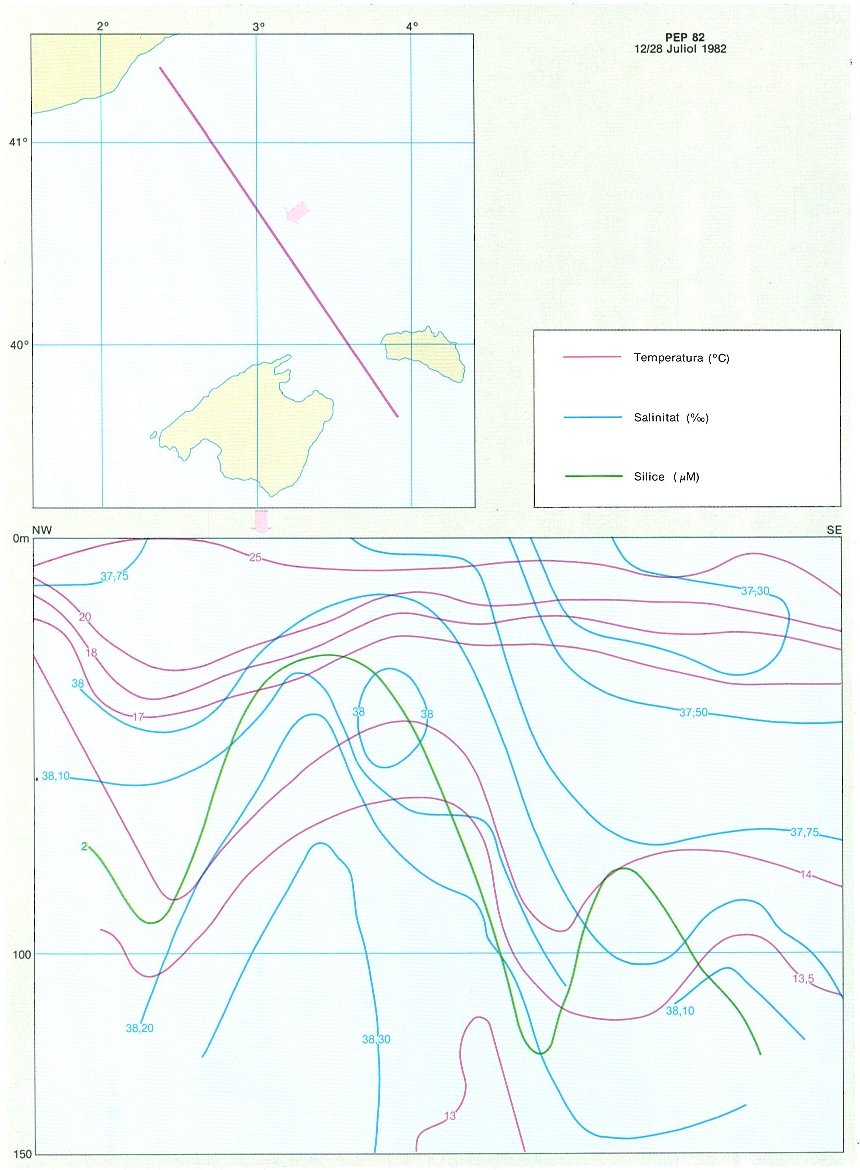
Secció hidrogràfica entre Barcelona i Balears que mostra estructures ascendents centrals ben clares en les distribucions de la salinitat i la concentració de silicat. Les línies isohalines mostren l’existència d’inestabilitats (30 ‰). La figura s’ha compost amb dades de la campanya PEP-82 (Productivitat Estival Profunda), del 12-28 de juliol de 1982.
Carto-Tec, original de l’autor.
Els episodis més importants que contribueixen a la producció anyal de fitoplàncton tenen el caràcter de pulsacions asimètriques, és a dir, d’inici brusc i de gradual dissipació. Sembla que són independents els uns dels altres i, per això, es pot especular que la variació de la producció total podria ser menor que la variació de cadascun dels episodis components, estabilitzant, en certa manera, la producció. Però, malgrat tot, hi pot haver considerables diferències d’un any a un altre, de vegades en forma d’anticipació o de retard de cadascun dels episodis, que sembla que donen una producció màxima segons el grau amb què fan coincidir els diferents factors favorables (vegeu-ho, més endavant, a propòsit de la successió). Aquests episodis esmentats s’escauen dins el que hem anomenat semestre productiu, que, hidrogràficament, es caracteritza molt bé per valors elevats de la difusió turbulenta i manca d’estratificació a l’aigua. Es comprèn que, en aquestes condicions, sigui possible la barreja vertical de l’aigua o que la manca d’estratificació indiqui ascensió d’aigua d’origen profund. Només les poblacions que vénen amb les aigües atlàntiques es poden estendre, vers la costa, damunt d’una aigua que s’estratifica o ja estratificada. L’altre semestre, que podríem anomenar pobre o estèril, cobreix els mesos de l’any en què l’aigua és fortament estratificada i les aigües de superfície pràcticament exemptes de fitoplàncton. Com es manté l’ecosistema pelàgic? Té algun sistema d’acumulació, que permeti de fer la viu-viu amb la «corda» rebuda durant el curs del semestre productiu? Tanmateix, semblaria un xic massa.
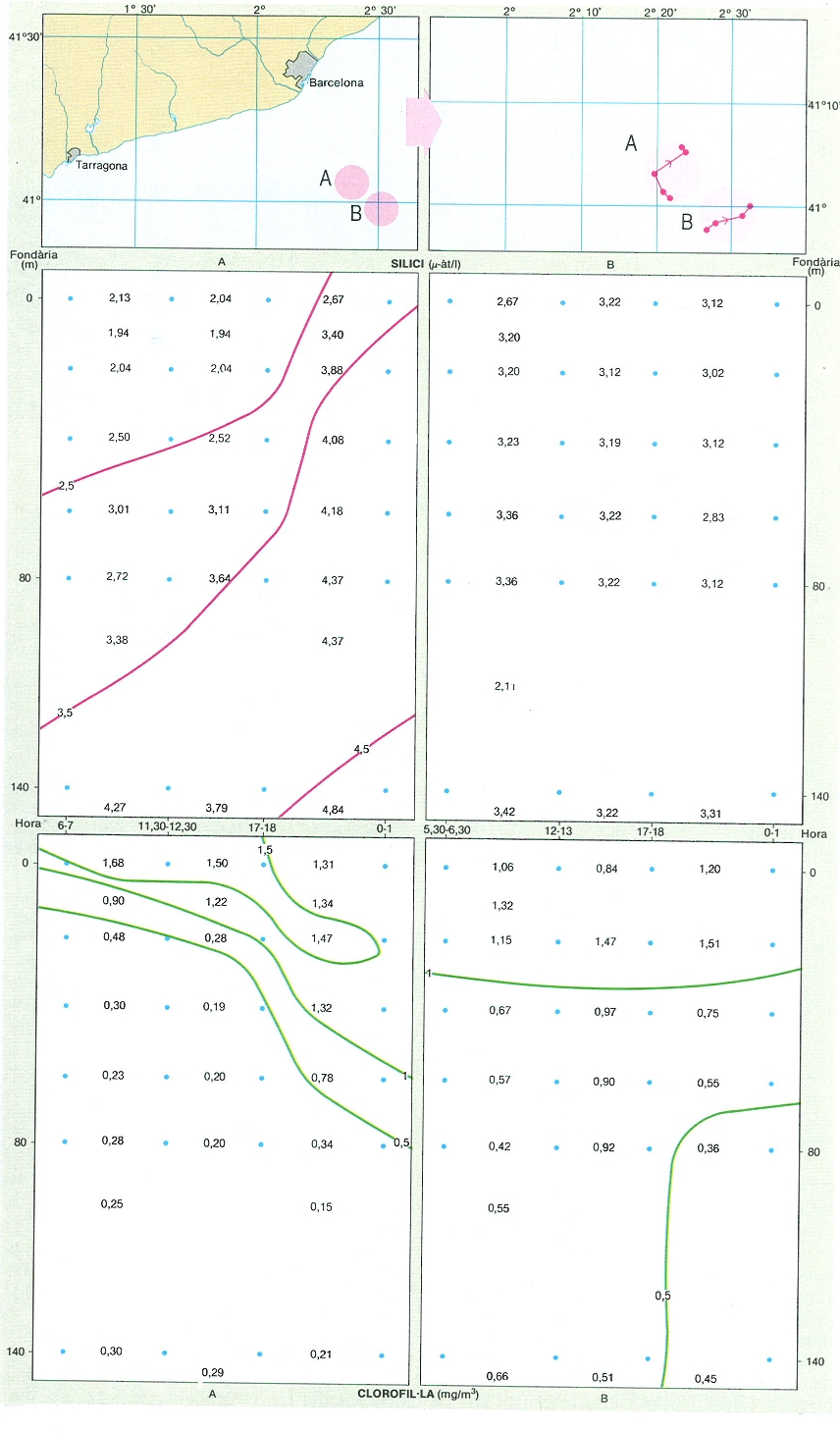
En els llocs actius del front catalano-balear hi ha una dinàmica molt ràpida que es reflecteix en la distribució de les característiques hidrogràfiques i en la dels organismes que es multipliquen molt ràpidament. Aquesta figura representa les distribucions del silici i de la clorofil·la en dues seccions properes al front (A i B) del 15-26 de març de 1985. En la secció més propera al front (B) la concentració de silici és alta i uniforme, i la de clorofil·la és encara baixa. Una mica més enfora del front (A) el silici és emprat en les capes més superficials i, en l’aigua menys moguda, la corofil·la augmenta i es va estratificant.
Carto-Tec, original de l’autor.
Un estudi més precís dels requeriments per a explicar la biologia local, enfront dels subministraments de nodriment, feia creure que l’estiu no era una estació tan poc productiva com semblava. Sota la termoclina —que, en certa manera, fossilitza una estructura força complicada i profunda— es reconeix un màxim profund de clorofil·la, amb concentracions fins d’1,5 a 2 mg de clorofil·la/m3 en un estrat més aviat prim, que pot contribuir a la producció primària d’una manera considerable. Però l’estudi d’aquest màxim profund serví per a reconèixer que tenia arrels en un front marí actiu durant tot l’any, i que constitueix una font de nodriment que, en els primers estudis, basats en la costa i no venint d’alta mar com hauria d’haver estat fet, havia passat inadvertida. En el cas del màxim profund de clorofil·la era òbvia la asimetria entre la part d’enfora respecte al front i les aigües potser més barrejades i irregulars i, en tot cas, amb un major component de moviment vers el SW, entre el front i la costa. A l’estiu, pensem que la inestabilitat creada en el front produeix volums d’aigua enriquida que rellisquen al nivell que els correspon per la seva densitat, aigües enfora, i entre altres paquets o llentilles d’aigua formades amb anterioritat. El plàncton del màxim profund de clorofil·la no és de característiques gaire peculiars. Solament de manera local s’hi troben petits cianobacteris que, en altres llocs, s’han considerat com a importants contribuïdors a la producció primària oceànica i, especialment, a fondàries considerables. També cal esmentar algunes espècies que s’han qualificat sovint de «plàncton de penombra», com Planktoniella sol i Ceratium platycorne, dues espècies precisament molt característiques i de gran interès per la seva biologia cel·lular.
El front, tal com es reconeix en una secció entre Barcelona i Mallorca, es mostra com una singularitat molt clara, reflectida en la distribució de tots o gairebé tots els descriptors físics (temperatura, densitat), químics (salinitat, silicat, etc.) i biològics (distribució d’organismes). Aquest front s’estén des del nord de Còrsega, on ha estat identificat per diversos investigadors francesos (Prieur 1979) fins a Eivissa com a mínim, en un traçat més o menys paral·lel a la costa i sempre seguint aproximadament la línia de màxima fondària (de més de 2000 m en bona part del seu trajecte). Es reconeix amb característiques de divergència en totes les compilacions de topografia geopotencial. Precisament aquesta mena d’informació mostra que l’estructura es pot reconèixer fins a les majors fondàries, cosa que significa la gran energia potencial associada amb les distribucions, que garanteix llur persistència. Si bé hi ha desviacions en la posició del front a la superfície, sembla que aquesta deriva superficial produïda per l’impuls de corrents estacionals (pel març, per exemple, el front pot estar més a prop de la península) no s’estén en fondària, com si el front es mogués com una cortina al revés, fixa en el fons i oscil·lant a la superfície. L’energia associada al front fa que aquest sigui absolutament persistent, encara que, algunes vegades, per exemple al març, l’aigua superficial d’enfora pot estendre’s per damunt del front, en direcció a la costa, en una capa prima, sempre de menys de 35 m de gruix, i en aquesta forma es detecta en imatges des de l’espai, que no són les més adients per a formar-se una idea del traçat del front. En altres temporades, especialment durant l’hivern, els llavis del front són més nets a la superfície i més ben detectables des de l’espai. Al front entren en contacte una aigua costanera, que es mou de NE a SW (ciclònicament), i una aigua central, que circula lentament, o bé en sentit anticiclònic. Les masses d’aigua, una enfront de l’altra, es deuen renovar amb una relativa rapidesa per mantenir el contrast de les qualitats de l’aigua i de llurs poblacions. El notable és que la velocitat de circulació de l’aigua, al llarg d’una secció perpendicular a la costa, no variï gradualment fins a invertirse, sinó que mostri el desenganxament o trencament sobtat que constitueix el front. Per la circulació descrita i amb relació a la rotació de la Terra, es comprèn que es generi una divergència, i que l’aigua de fondària sigui impulsada cap amunt, dintre del mateix front. En conseqüència, podríem dir que el front es fa doble i entre els dos fronts hi ha una aigua diferent que es mou cap amunt, i aquesta aigua, que puja enriquida en nodriment, és la més important des del punt de vista biològic, perquè és un important factor de fertilització. Es comprèn que els autors francesos, amb referència a les aigües lígurs, parlin de dos fronts paral·lels, encara que, possiblement, el front és més ample i complex a l’extrem NE que a les aigües de la mar Catalana.
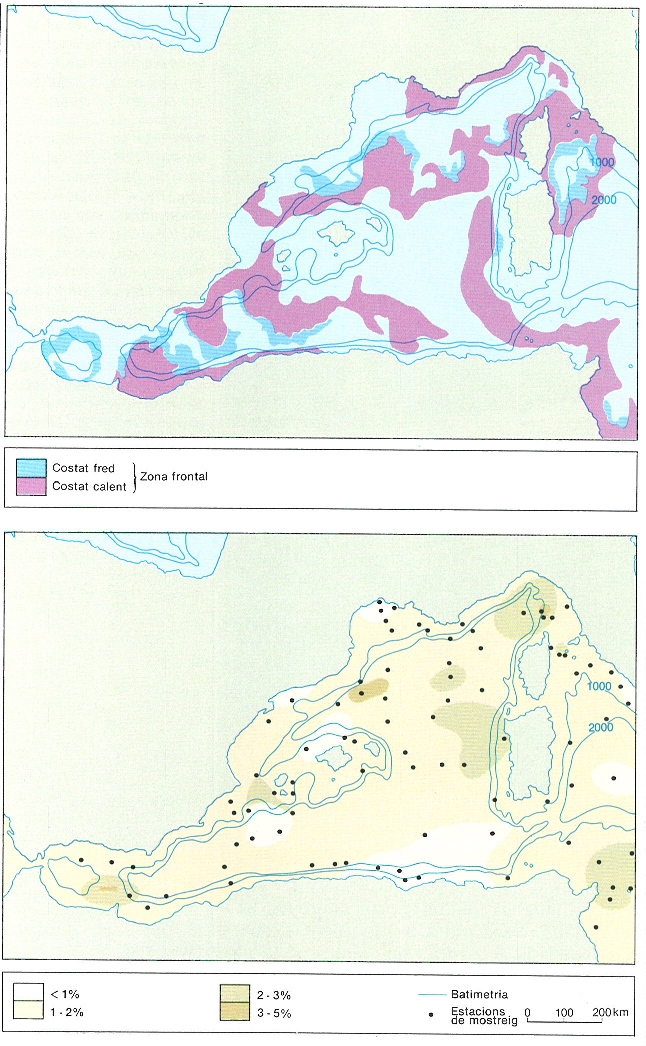
Possible localització dels fronts marins segons tècniques diverses. A dalt, límits tèrmics de caràcter possiblement frontal deduïts de radiometries de satèl·lit (CMS Lannion, 14/X/84). A baix, % de sílice (possiblement, restes de diatomees i altres organismes pelàgics) en la part superior dels sediments, segons dades d’Emelyanov i Shinkus. Cal suposar que aquestes restes són més abundants per sota de les àrees habitualment més productives.
Carto-Tec.
La major complexitat del front vers el NE es pot relacionar, també, amb un altre fenomen característic del nostre racó de la Mediterrània. Al golf del Lleó, quan bufa el mestral i la tramuntana, en ple hivern, es formen unes veritables xemeneies o remolins ciclònics que arriben a gran fondària i que són un factor preponderant en la barreja vertical de l’aigua durant l’estació freda, i, per tant, de l’enriquiment de les aigües de superfície. Havíem cregut que la intensitat d’aquesta barreja, més gran els anys més freds, condicionava el nivell a què es trobava aigua enriquida a la vora de la plataforma de la costa i, per tant, l’èxit del moderat aflorament del febrer sota l’acció del vent. El que sí que es pot afirmar és la importància d’aquests cilindres de barreja per a la vida de la Mediterrània occidental i el fet important que tots ells es formen o van a raure a l’eix del front —o entre els dos fronts geminats, si preferim aquesta interpretació—, i, d’aquesta manera, circulen vers el SW i s’esvaeixen o queden reduïts a bosses d’inestabilitat entre dues aigües. Temps enrere, es pensava que, en lloc de front, hi podia haver cúpules ciclòniques aïllades; no ho neguem, en el sentit que el front és més o menys irregular i, sobretot, és recorregut per ones importants al seu llarg. És com si diguéssim que les cúpules visibles no són més que els anells de la gran serpent de mar, que també descriu meandres horitzontals.
Aquest front no és una estructura única en el seu gènere. Sembla que es reconstrueix al llarg de l’E i el SE de la Península i en direcció a Gibraltar. També a la Tirrena es reconeixen estructures semblants. Certament, constitueixen exemples magnífics d’ergoclines, llocs on l’energia cinètica de les masses d’aigua marina desenvolupa un treball, que es manifesta en una elevació del nivell de producció biològica.
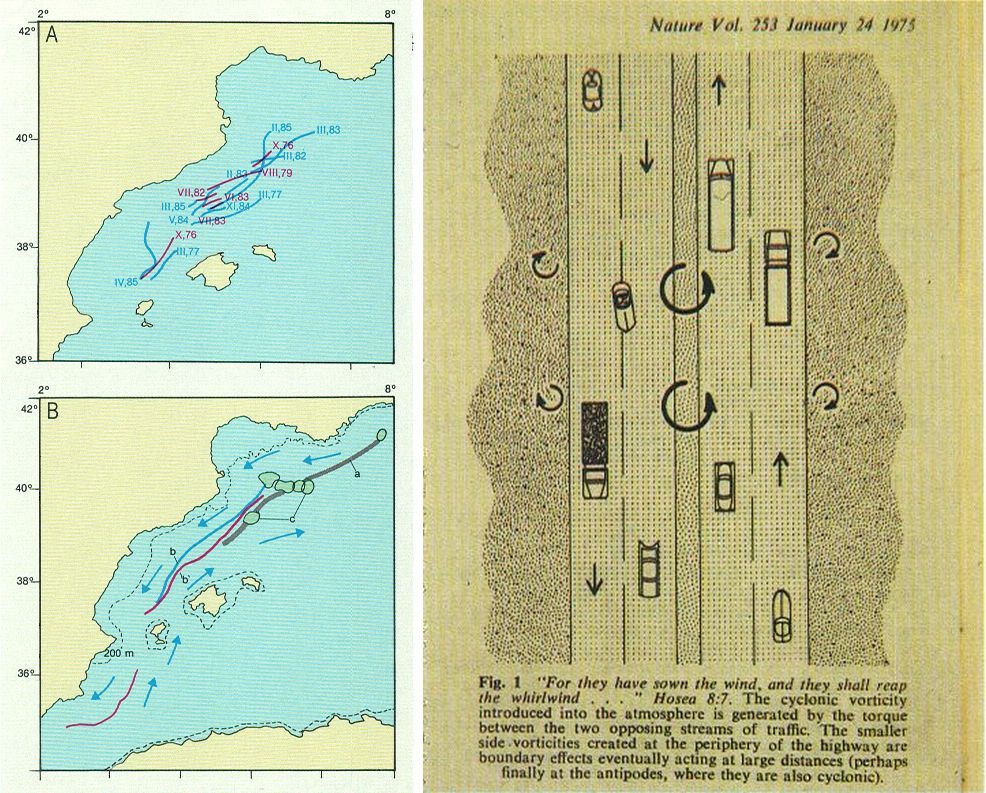
El front marí catalano-balear. A Imatge sobre un mapa, amb les posicions del front en diferents creuers, segons Font (s’indiquen el mes i l’any de cada observació). B Posició general del front, a l’hivern (b) i a l’estiu (b’), i posició de la divergència segons Ovchinnikov (a); la c indica posicions observades de xemeneies de barreja vertical durant l’hivern. A la dreta, la il·lustració de la interpretació faceciosa d’Isaacs i els seus col·laboradors, de la introducció de vorticitat a l’atmosfera pel tràfic de carreteres. En el cas del front és diferent, ja que més aviat l’existència dels llavis del front absorbiria energia del vent i donaria lloc als cilindres d’eix vertical o xemeneies de circulació (marcades en el dibuix B) característiques del golf del Lleó.
Ricardo Génova
La part del front al llarg de les costes catalanes és coneguda, entre els que l’hem estudiada, amb la designació de «l’autopista de la Mediterrània». Val la pena d’explicar-ne la raó, perquè és instructiva i divertida. El 1975, John D. Isaacs, un respectat biòleg marí, treballant a la Jolla (Califòrnia), amb altres col·laboradors, publicà un estudi que argumentava que la circulació a les carreteres, seguint la dreta a l’hemisferi nord, podia crear gèrmens de vorticitat ciclònica que, introduïts a l’atmosfera i amplificats per aquesta, podien contribuir a la generació d’estats de mal temps. L’article pogué semblar una enginyosa facècia, però ara veiem que conté una aplicació directa per a la nostra situació. La circulació en el front sembla particularment apropiada per a transferir energia del vent a l’aigua, on pren la forma de remolins, que faciliten i acceleren la circulació, com si fossin les boles d’un coixinet de rodament. Una altra manera de veure-ho és acceptar que l’eix de la divergència exposa al vent una aigua que és particularment sensible a la pèrdua de flotabilitat. En aquestes condicions, l’efecte del vent es transmet fàcilment més avall.
Un estudi acurat de qualsevol secció del front mostra clarament la divergència central, amb l’elevació de les estructures a l’eix del front. El front és un lloc d’elevada producció, i les poblacions es desplacen i es dispersen a una banda i a l’altra. És notable de veure a l’eix una població encara poc densa, on es troben força organismes que dipositen material calcari, com els foraminífers i els coccolitoforals (la presència de coccolitoforals en abundància ha estat reconeguda com a característica de fronts marins en altres llocs), i, estratificant-se successivament, unes capes damunt les altres, poblacions variades i successives i, en conjunt amb una major biomassa, als costats del llavi. Aquesta imatge recorda molt les distribucions que s’observen entorn dels centres d’aflorament, com ara els de les costes del Sàhara i és natural que sigui així, ja que es tracta, essencialment, del mateix fenomen.
L’eix del front és, doncs, un lloc de producció molt alta, que funciona tot l’any, i que completa el balanç de la producció precisament amb la quantitat que trobàvem a faltar. Probablement, el fosfat (i el silicat, com també els compostos de nitrogen) que porta a la regió il·luminada el nostre front cobreix una tercera part del que fa falta per a explicar la producció observada. De manera que el semestre pobre no és tan pobre. El front funciona contínuament i, a més, alimentat en part pel mateix front, tenim el màxim profund de clorofil·la. De fet, aquest màxim es veu perquè la difusió és petita o feble al seu nivell. Més amunt, també hi ha producció, per exemple, amb moltes diatomees i coccolitoforals, però és més dispersa. El que caldria conèixer és la quantitat total de fòsfor injectada per grau de latitud, que depèn de la distribució de les velocitats ascensionals entorn del front, calculables a partir de les distribucions de les característiques hidrogràfiques. Velocitats ascensionals d’un a cinc metres per dia són possibles i suficients.
Cicles d’elements químics i estratificació
D’una manera general, sempre s’ha fet referència al fòsfor com a element limitant final. El fòsfor, per les seves característiques químiques, circula sempre en forma de fosfat, que fàcilment dóna minerals insolubles amb el calci, el ferro, l’alumini, etc. En canvi, hi ha una gran reserva de nitrogen en forma de gas (N2) a l’atmosfera i, per tant, en dissolució a l’aigua.
Entre els organismes i l’entorn físic, el nitrogen pot circular per tres vies diferents, el conjunt de les quals té una enorme significació en el funcionament de la biosfera. En els organismes, el nitrogen es troba reduït (per exemple, en els aminoàcids), en forma d’amines i amoni. Si surt així al medi, pot ser reabsorbit sense gaire despesa energètica. Ordinàriament, aquests compostos són oxidats —en gran part, per acció bacteriana— i trobem nitrat i nitrit al medi, l’assimilació dels quals, que comporta la reducció, és relativament costosa. Finalment, una part es veu reduïda i desproveïda d’oxigen en el procés de desnitrificació, que té lloc en ambients amb poc oxigen: el resultat és nitrogen gasós (dinitrogen). La seva fixació és energèticament molt cara i solament poden fer-la procariotes.
El cicle de desnitrificació i fixació de nitrogen és com un fre o regulador que tendeix a limitar la taxa de renovació dels ecosistemes. La seva importància relativa és una mesura de la tensió sota la qual treballen els ecosistemes; potser un 2% del nitrogen circula per aquesta via més cara als ecosistemes terrestres; bastant més, almenys un 5%, als ecosistemes d’aigües epicontinentals, que sovint treballen forçats o sota estrès, i força menys, potser un 1%, als sistemes oceànics actuals. La possible significació del nitrogen inorgànic combinat —nitrat, nitrit, amoníac—, com a limitant als ecosistemes marins, s’ha de veure sota aquest aspecte. Hi ha, certament, desnitrificació a la mar, probablement poca a la Mediterrània, però sí en llentilles d’aigua pobres en oxigen en àrees d’aflorament i, naturalment, als sediments litorals i de maresmes. Hi ha, també, en el domini pelàgic, organismes fixadors de nitrogen, sovint associats —com a simbionts— amb altres, com els cianòfits Richelia, que es troben a l’interior de diatomees del gènere Rhizosolenia, en una simbiosi que s’observa sovint a la Mediterrània a l’estiu, o com altres rivularials que viuen damunt les algues brunes flotants a la mar dels Sargassos. En experiments d’enriquiment fets expressament, a la Mediterrània occidental i a les capes superficials durant l’estiu, sembla que hi ha, positivament, un efecte enriquidor més manifest del nitrogen que del fòsfor, i tot fa pensar que el primer és, almenys en certes condicions, més limitant que el segon. Però, de fet, les condicions canvien molt en el curs de l’experiment, i sempre es pot pensar que queden afavorits organismes «nitròfils» que usen i fan reciclar les petitíssimes quantitats de fòsfor que hi havia a les cèl·lules dels dinoflagel·lats grossos que es trobaven a l’aigua inicial i que, generalment, moren durant l’experiència. En tot cas, a l’estiu ens trobem en unes condicions d’extremada pobresa de nodriment, en general.
A les mars estratificades se sol trobar un estrat prim, relativament ric en nitrits, per sota de la termoclina i vers el límit de la llum útil. En una campanya d’estudi de la Tirrena fórem dels primers a precisar-la i a remarcar l’interès que ofereix la seva topografia, amb ondulacions i forats que reflecteixen tant els accidents hidrogràfics (influència de fronts) com l’activitat biològica. Es pot interpretar com una conseqüència de la sedimentació del fitoplàncton, en el sentit que les cèl·lules que van davallant, en arribar en un nivell on hi ha nitrat però on ja no hi ha llum, poden reduir el nitrat a nitrit amb l’energia de què disposen, però, per a anar més enllà —reduir el nitrit— necessitarien llum, i no en tenen, de manera que el nitrit ja no pot ser transformat i retorna al medi. Així, es forma el màxim de nitrit, com a resultat d’aquest efecte «seguit, seguit, d’aquest colpeig repetit, d’estalactita que plora», de les cèl·lules del fitoplàncton en la seva davallada fatal.

Histogrames de la distribució mensual (1950) de freqüències de l’amplada màxima del solc (longitud del solc, en µ) en Ceratium tripos de la Mediterrània occidental (a dalt), i variació al·lomètrica de la forma de les cèl·lules (a baix). Observeu que els mesos de temperatures més altes apareix una població de cèl·lules més petites.
Amadeu Blasco, amb dades de Juan J. López.
Aquest és un dels molts efectes que se sumen per a generar una forta estratificació del fitoplàncton, especialment persistent i visible quan l’aigua és tèrmicament estratificada, durant l’estiu. Les necessitats fisiològiques de les diverses espècies que hi persisteixen no són coincidents, i això fa que unes predominin en uns llocs, i unes altres en altres. Hi ha, per exemple, un petit grup de dinoflagel·lats que caracteritzen les aigües superficials, com ara Ceratium furca, C. declinatum, C. fusus, Protoperidinium brochi, etc. Encara que no s’ha demostrat, és probable que formes suposadament diferents i que, àdhuc, han rebut designacions diverses, de les quals una es troba més amunt i una altra més avall (principalment en Ceratium), siguin formes de reacció d’uns mateixos genotips, i sempre en el sentit que les formes que viuen a més fondària són de coberta més gruixuda i de relleus (carenes, espinetes) més marcats. Hi ha, doncs, en aquest sentit i en d’altres, una variabilitat considerable en les característiques dels individus amb relació a les propietats de l’ambient, l’abast de la qual no és prou conegut, si no és la idea molt general que unes mateixes espècies taxonòmiques donen, generalment, formes més petites quan la temperatura és més elevada. Diferències de mida porten associades diferències en les proporcions de la cèl·lula, de tipus al·lomètric, cosa que és especialment perceptible en els organismes proveïts d’apèndixs molt llargs, com ara els Ceratium. La variabilitat en la mida s’estén sovint per la combinació amb fenòmens de poliploïdia. Val la pena d’esmentar que Jörgensen, un dels planctòlegs que més contribuí a l’estudi del nostre plàncton, per mitjà de l’anàlisi de les col·leccions aplegades pels vaixells danesos «Thor» i «Dana», s’adonà de la variabilitat dels Ceratium en dependència dels fenòmens indicats i d’altres i suposà que les poblacions mediterrànies es refeien més o menys periòdicament a través de l’estret de Gibraltar, i que la morfologia de les cèl·lules anava «degenerant» (dimensions menors i banyes relativament més curtes, en aigua més càlida) a mesura que s’avançava vers l’Orient.
Caràcter discontinu de la producció
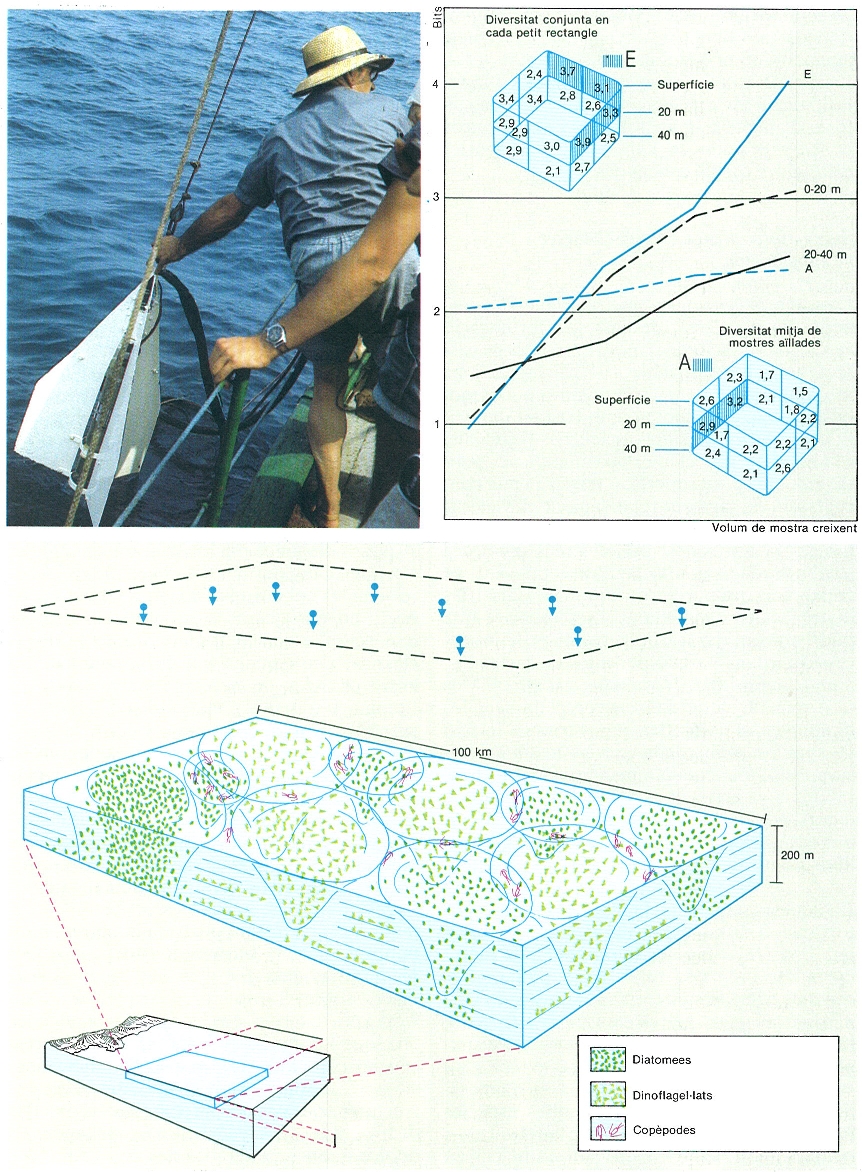
La composició del plàncton reflecteix l’organització dinàmica del sistema fluid en què és immers, com els núvols reflecteixen la dinàmica atmosfèrica. Un estudi acurat vol analitzar centenars o milers de mostres, i això es pot fer solament aspirant mostres d’aigua contínuament amb una bomba i un tub de recol·lecció, de manera que es controla la posició d’entrada. La part baixa d’un aparell d’aquest tipus a la «Nika» es veu a la fotografia de l’esquerra. La diversitat del plàncton en bits és una mesura utilitzable i a la dreta es veuen les diversitats en mostres separades (a baix a la dreta) i en mostres compostes (a dalt a l’esquerra) en un bloc estudiat prop de Castelló (final de l’estiu). Entre 0 i 20 m, la diversitat augmenta en mostres més extenses, la qual cosa indica una organització més rica, entre 20 i 40 m a l’oest; en aigua més barrejada la diversitat varia poc. Aquestes diferències es poden representar com a espectres de diversitat. A baix, interpretació provisional del món del plàncton: un nombre indefinit de pertorbacions (vent, etc.) i de distribució i intensitats irregulars afecten contínuament la massa d’aigua: la resposta genera petites successions que centrífugament es perden en un fons de motiu de distribució molt complex, en sistemes de baixa energia i relativament poc mesclats.
Ramon Margalef; Ricardo Génova.
És clar que el sistema pelàgic, i ja a partir dels seus elements productors, el fitoplàncton, no és pas un sistema homogeni. No es podria comparar mai a una suspensió de cèl·lules uniforme en una solució nutritiva molt diluïda. I la mateixa afirmació val per a totes les característiques de les poblacions. Per això es fa tan difícil d’avaluar d’una manera correcta la producció primària de la regió. Diem que, en general, la producció és d’entorn 80 g C assimilats per m2 i any, un valor relativament baix amb relació a l’oceà en general, però encara per damunt dels valors mesurats a la resta de la Mediterrània. Ara bé, això és el resultat d’experiments de fixació de 14C fets en flascons —és a dir, sota condicions molt diferents de les que hi ha a la mar—, de mostres agafades en fondàries diferents i en llocs diferents, amb la dificultat d’entendre bé les causes de les diferències i, per tant, d’avaluar el significat de la variabilitat de les dades i ferne una integració adient. Però el relatiu optimisme, en acceptar aquells valors, és confirmat per la coherència entre els resultats dels experiments i el que caldria per a explicar altres particularitats de la vida de la nostra mar. Però és evident que hi ha grans diferències en l’espai i en el temps.
De moltes, ja en coneixem la causa. La barreja vertical en trencar-se la termoclina, els afloraments costaners sota el vent, són fenòmens sobtats que inicien un procés de producció i una successió ecològica. El front persistent de «l’autopista de la Mediterrània» funciona de manera més seguida, però la dinàmica de les poblacions té també una discontinuïtat, just arran del front, i a partir d’allí s’estenen de manera centrífuga, i així la successió ecològica agafa, fins a cert punt, una projecció damunt l’espai. En un cas i en l’altre, tenim exemples d’una de les regles més generals de la biologia del plàncton —i podríem dir de tota l’ecologia—: les comunitats d’organismes no es desenvolupen en cicles regulars i més o menys simètrics, sinó a partir de pertorbacions inicials de diferent entitat i de caràcter òbviament asimètric, amb una forta entrada inicial d’energia (els processos de barreja o d’aflorament, discontinus a la costa, menys discontinus al front) i una gradual «estabilització» amb organització pausada de comunitats que es van substituint les unes a les altres fins a un estat final de peresa extrema i de gran confusió, on sovint es troben organismes curiosos, de règims de vida complicats i, sovint, molt variats. Probablement, la hipòtesi, més aviat gratuïta, que el plàncton es desenvolupava segons cicles regulars ha influït negativament en la programació de la recerca de molts laboratoris i, fins i tot, en l’organització d’expedicions d’estudi.
De fet, la mar repeteix a l’infinit i en totes les combinacions allò que es veu en l’experiment tan senzill d’agafar una mostra d’aigua, barrejar-la i posar-la en un flascó a la llum. Sempre hi ha un canvi sobtat inicial de la població —que podríem considerar certament indesitjable en els experiments fets per a mesurar la producció primària—, seguit de canvis graduals de disminució de la productivitat, fins a una situació relativament poc activa. És clar que no podem considerar senzillament que el sistema pelàgic és fet d’un seguit de flascons de cultiu disposats els uns als costat dels altres. En realitat, cada petit volum d’aigua es mou contínuament, sovint cap amunt i cap avall, i, a més, es difon gradualment en els volums d’aigua immediats.
Creiem que la idea d’un sistema que tendeix a disminuir la productivitat (P/B) en el seu conjunt, i que és pertorbat, ací i allà, de manera irregular i contínua, per «desastres» locals, que tenen, generalment, la forma d’una introducció d’energia externa, és perfectament vàlida. Almenys, és coherent amb totes les observacions, i explica la dificultat de sistematitzar-les en un model massa senzill. Els models demogràfics clàssics que ignoren l’espai són molt poc aplicables a les poblacions marines i fou, principalment, Riley qui intentà produir nous models quantitatius de fitoplàncton. El 1949, jutament amb altres científics, publicà un estudi modèlic sobre ecologia de plàncton on, de manera natural, s’assenyala el paper de la velocitat de sedimentació de les cèl·lules i de la difusió turbulenta en el medi. És basant-nos en estudis semblants que podem pensar en la asimetria fonamental de la dinàmica de les poblacions del fitoplàncton marí. Els períodes de desorganització d’una estructura vella i de ràpid augment de la productivitat són relativament breus i van sempre seguits d’una successió pausada, on l’energia de turbulència en el medi disminueix, la segregació ecològica augmenta —pot incrementar-se el grau d’estratificació de les poblacions— i la productivitat (P/B) disminueix. Aquest és el procés natural de la successió ecològica, que no és pas uniforme. En el medi pelàgic estem sempre exposats que una tempesta o un cop de vent determinin localment un retorn enrere de la successió de manera sobtada, fenòmens que, com és natural, no afecten de la mateixa manera tots els components del sistema pelàgic: el fitoplàncton, més a prop del lloc d’ingrés de l’energia, reacciona més de pressa i pot ser més alterat, mentre que els animals que es poden moure d’una banda a una altra en són menys, i els animals de vida relativament llarga, com els peixos, gens. Però hi ha canvis dins de canvis, i pot haver-hi diversos anys seguits on predominin certs estils de canvi i, aleshores, fins i tot les poblacions dels animals de vida més llarga resulten també afectades, augmentant o disminuint la densitat total. D’aquesta mena són les fluctuacions en la relació entre les poblacions de sardina i de seitó, o d’alatxa, a la nostra mar, i altres canvis de llarg període en l’abundància d’espècies distingides o més conegudes.
Successió ecològica i tipus biològics de fitoplàncton
La successió ecològica que podríem dir «normal» des d’un estat de barreja i fertilització fins a una situació final més empobrida és bastant ben caracteritzable. Al principi, hi ha moltes diatomees, que són organismes no mòbils i que se sedimenten amb bastant de facilitat, però també petits organismes flagel·lats, i la mida de les cèl·lules acostuma a ser petita, amb una gran superfície relativa i una concentració de pigment relativament elevada; si hi ha una ascensió sensible de l’aigua, les diatomees poden ser de cèl·lules més grosses, i sovint produeixen mucílag. Més endavant, va creixent la proporció de dinoflagel·lats i les diatomees es van reduint a les espècies que podríem considerar característiques de la successió tardana —final per a les diatomees—, amb Rhizosolenia, Hemiaulus, i d’altres. Més tard encara, desapareixen completament les diatomees, si no són les que entren en relació de simbiosi amb ciliats que les mantenen en suspensió, tot actuant a manera de motors forabord, i va creixent el nombre d’espècies de dinoflagel·lats, incloent-hi moltes formes més o menys heterotròfiques, o amb altres modalitats de nutrició especial.
En aquesta seqüència, la biomassa total del fitoplàncton disminueix, fins al punt que hi ha una bona relació entre la presència de diatomees i la biomassa total. Els dinoflagel·lats són més constants en formar el fons de la vida marina pelágica. En el curs de la successió, també, el quocient producció/biomassa disminueix i la riquesa i la diversitat d’espècies sol augmentar: en particular, es passa d’una situació en què hi pot haver moltes espècies juntes en una aigua fortament barrejada —i rica en nodriment— a una altra situació en què les poblacions tendeixen a una major segregació espacial, de manera que, en un clap molt petit, potser es troben poques espècies, però sobre un espai gran, les petites taques locals d’un plàncton molt heterogeni donen una representació d’una gran diversitat. Aquesta estructuració bàsica del fitoplàncton es reflecteix en nombroses observacions locals, de les quals no es tenen pas les que es desitjarien, precisament perquè és molt laboriós d’aconseguir-les, encara que estiguin a l’abast de tothom. D’altra banda, cal recordar que diferències de temperatura poden definir que siguin unes espècies o unes altres les que ocupin una determinada «funció» en una situació determinada. I no es pot dir mai que falti la «llavor»: en un estudi minuciós, fet a l’estiu, es troben exemplars de la major part de les espècies presents a la regió durant tot l’any, si bé moltes d’elles representades per un nombre molt reduït d’individus, que no es veuen en un estudi menys detallat; aquestes cèl·lules són allà, a tot arreu, en espera de condicions adients per a multiplicar-se. Si pensem en la petitesa de les cèl·lules del fitoplàncton i en la seva enorme superfície relativa, comprendrem que el fitoplàncton treu profit ràpidament de qualsevol enriquiment eventual del medi —per aflorament, barreja, cop de vent, etc.—. Aleshores, les espècies que es multipliquen més són les que, més endavant, considerem com a característiques de les primeres etapes de la successió, principalment flagel·lats menuts i diatomees (Chaetoceros, Skeletonema, Thalassionema, etc.).
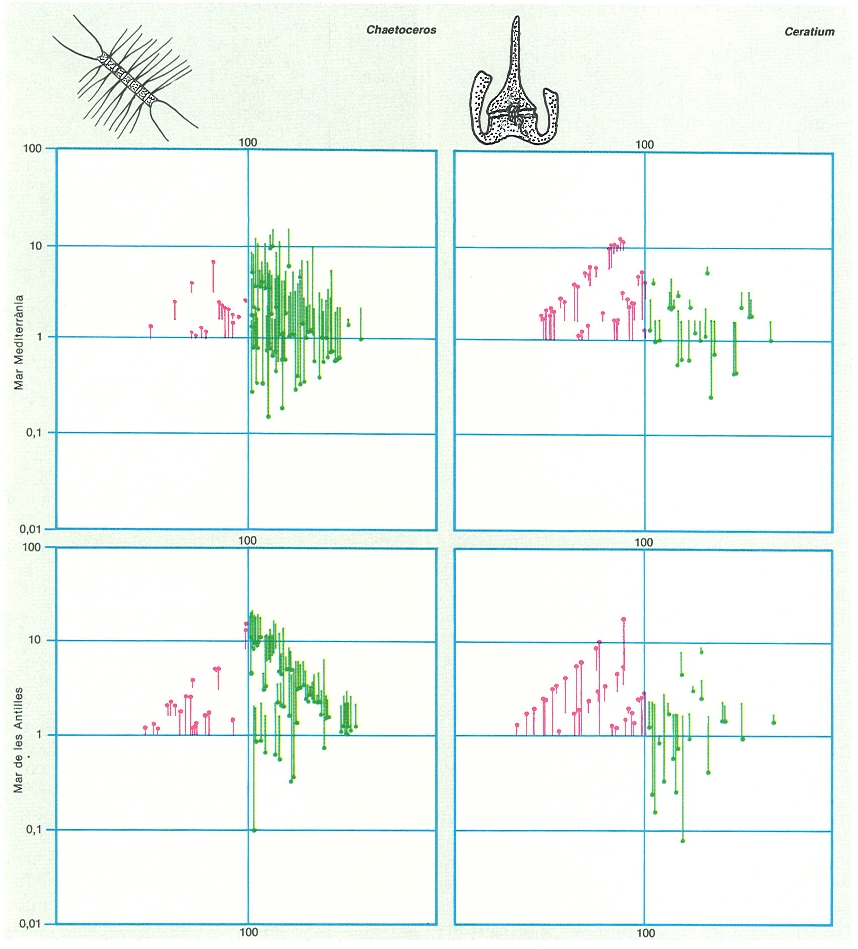
La teoria ecològica dóna molta importància a l’associació entre espècies d’un mateix gènere. Poden trobar-se juntes si tenen necessitats comunes; però fins i tot en aquest cas cal esperar que la competència porti a distribucions no coincidents. Tota aquesta problemàtica es retroba en el plàncton. El dibuix mostra l’exemple corresponent a dos gèneres diferents, una diatomea (Chaetoceros) i un dinoflagel·lat (Ceratium), cadascun estudiat en dues mars diferents. Les ratlles representen la distància que hi ha entre la probabilitat d’associació, calculada suposant que es distribueixin a l’atzar i calculada sobre les freqüències respectives, i la freqüència real observada de la seva presència conjunta. Les ratlles verdes corresponen a combinacions binàries de freqüència real superior a l’esperada; les vermelles, a les combinacions binàries d’ocurrència inferior a la que caldria esperar de les freqüències independents de cadascuna de les espècies, és a dir, que es dóna segregació o repulsió. És clar que, en general, i de manera semblant tant en la Mediterrània com en la mar de les Antilles, les espècies de Ceratium tendeixen a excloure’s i anar cadascuna pel seu costat, mentre que les de Chaetoceros s’associen en grups congenèrics amb més freqüència, certament perquè depenen molt d’aigües agitades. Les xifres en ordenades són percentatges de mostres (en escala logarítmica). Per fer més senzill el gràfic sense que perdi el seu valor demostratiu, s’ha prescindit de les combinacions que es podien esperar rares perquè corresponen a espècies poc freqüents, que en realitat tenien poca probabilitat de trobar-se juntes, i que de fet mai no coincidiren o solament ho feren una vegada.
Ricardo Génova, original de R. Margalef.
Aquesta dinàmica tan intensa, on les cèl·lules s’arriben a dividir més d’una vegada en 24 hores i on les poblacions canvien totalment d’aspecte en uns quants dies, produeix imatges que no es concilien fàcilment amb la teoria clàssica, que demana una forta competència entre espècies de necessitats semblants. El fitoplàncton sembla massa divers i hom es pregunta fins a quin punt valdria la pena d’estudiar espècie per espècie, superant els problemes tècnics que aquest projecte podria plantejar. A més de reconèixer les grans estructures relacionades amb els episodis de fertilització, i els seus canvis, i altres estructures menors, com les fonamentades en contrastos en l’abundància relativa de diatomees i dinoflagel·lats, a una escala més petita —de centenars de metres a pocs quilòmetres— on també hi ha una estructura òbvia, probablement no podem fer més que reunir informació sobre transsectes o seqüències d’informació, tal com ens poden donar algunes tècniques disponibles —mesures de fluorescència, comptadors de partícules en suspensió—, i, després, formar hipòtesis sobre estructures tridimensionals compatibles amb les distribucions observades al llarg de línies. És dubtós que les fotografies des de l’aire, o des de l’espai, puguin contribuir al reconeixement de la distribució del plàncton; en part, perquè sols es noten les temperatures o les concentracions de pigment properes a la superfície, mentre que aspectes importants de les distribucions, com és ara el màxim profund de clorofil·la, resten invisibles. El més important que aporten les imatges tretes d’enfora és el reconeixement de freqüents remolins i vòrtexs, que tenen una estructura sovint espiral, l’origen dels quals és ben interpretable hidrogràficament; però el seu significat biològic per a la ulterior evolució de les poblacions es desconeix.
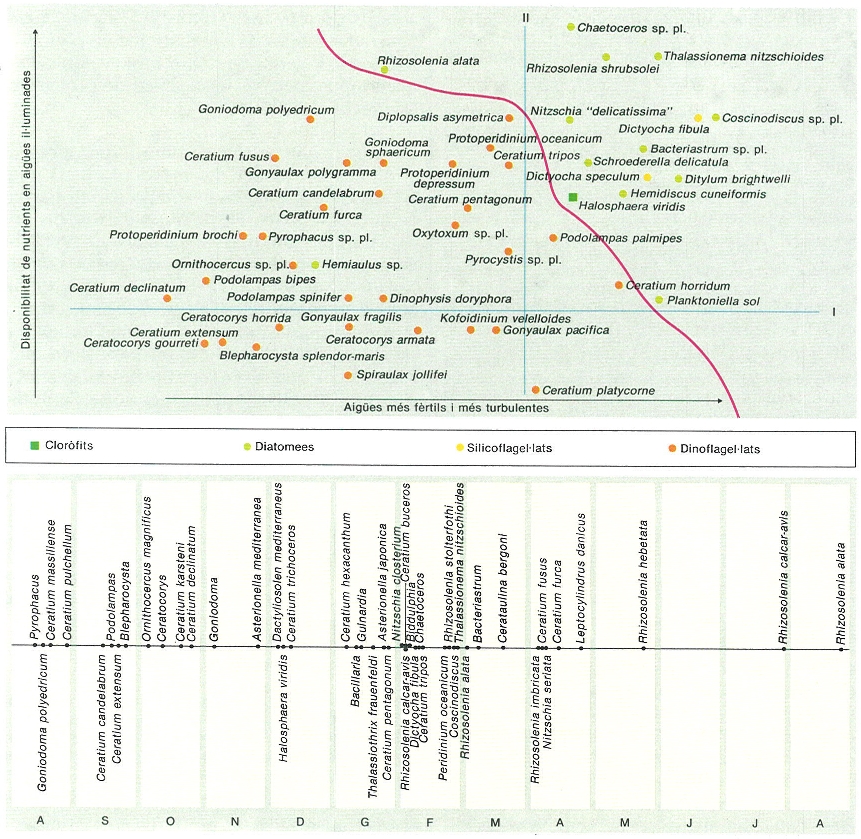
Distribució de les espècies fitoplantòniques en un espai ecològic (a dalt) basat en la probabilitat de trobar-les juntes o amb abundàncies correlades: dues espècies que en el gràfic apareixen pròximes, sovint viuen juntes; i, al contrari, és molt petita la probabilitat de trobar associades espècies que se situen en punts allunyats en el diagrama. Aquesta distribució correspon bastant bé a una interpretació dels tipus ecològics de fitoplàncton com a resposta evolutiva a condicions de vida caracteritzables principalment per l’energia mecànica de l’entorn (turbulència) i per la presència de nutrients en la zona fòtica. El diagrama, que hem anomenat «el mandala del fitoplànton», permet una sistematització còmoda de les comunitats fitoplanctòniques. Les espècies ocupen posicions característiques al llarg de les successions i ho van repetint en successions d’anys diferents. A baix, seqüència mitjana (expressada per la posició del centre de gravetat de la distribució de cada espècie) en anys consecutius, per a mostres corresponents a àrees pròximes a Barcelona i a Castelló.
Carto-Tec, original de l’autor.
Potser val la pena d’assenyalar que les anteriors observacions menen a una síntesi conceptual que representa un cert progrés amb relació a interpretacions massa simplistes de la vida planctònica. Combinant els valors de sedimentació, de capacitat d’assimilació dels diversos nutrients pels organismes, amb les condicions de turbulència i subministrament de nutrients —lligades a la turbulència— que ofereix l’entorn, s’arriba a establir la correspondència possible o la compatibilitat entre determinats organismes i determinades situacions. Diferències mínimes en la propensió a sedimentar-se són decisives. Adaptació precisa a certes temperatures o altres condicions d’entorn semblen molt secundàries, amb relació al paper preponderant de la coincidència de condicions favorables de producció (prou concentració d’elements químics necessaris en zona fòtica) i turbulència o energia externa. Això es pot representar de manera senzilla en un gràfic —el «mandala del plàncton»— que sintetitza una sèrie de relacions ja assenyalades. A més, ens permet de veure que hi pot haver altres situacions molt instructives. Una és la combinació de molta energia externa amb poca disponibilitat de nodriment, en la qual el plàncton és negligible i es dispersa. Aquesta situació es dóna en condicions de barreja molt intensa quan s’estén fins a nivells que són fora de la llum, com en les xemeneies de barreja vertical del golf del Lleó, que són pràcticament exemptes de plàncton que, no obstant això, creix rapidíssimament en tota la vora. Aquesta relació fou reconeguda a les costes de Noruega i de Maine, i fou estudiada, més tard, quantitativament; en una versió d’aplicació òbvia, diu que el fons posa un límit a la barreja vertical de l’aigua i que la productivitat és major, o el seu màxim es dóna abans en el temps en aigua menys profunda. Recordem que la poca producció dels oceans, en general, l’explicàvem en part per la seva extrema fondària, que imposava un retorn molt lent; també és la probabilitat de pèrdua d’una part de les poblacions cap a aigües sense llum. Certament, la producció primària és més elevada en mars de poca fondària, especialment si la llum pot arribar prop del fons i si són relativament agitats, com ara la mar del Nord. A les nostres costes, la plataforma continental és relativament estreta, de manera que l’efecte esmentat no és important i la producció és baixa. Això es combina amb el règim de circulació general a la Mediterrània que, com sabem, és descendent en la seva resultant i tendeix a empobrir-la.
L’angle oposat, que correspon a les condicions de poca turbulència i concentració alta de nodriment, és una situació inestable, si no hi ha un subministrament seguit d’elements necessaris a la vida en superfície, com s’escau en badies i estuaris, úniques situacions que permeten la persistència de les poblacions característiques que anomenem «aigües roges». Tanmateix, concentracions semblants, visibles, també, com les aigües roges, es poden observar a la banda més estratificada d’un front que és turbulent i ric en nodriment a l’altra banda. En aquest cas, l’acoloriment de l’aigua no es deu solament a organismes vegetals autònoms, sinó també a Mesodinium, un ciliat que conté simbionts fotosintetitzadors, que no són altra cosa que cèl·lules incompletes de criptomonadals.
L’estudi de la distribució de les espècies, una per una, basada en les llistes de la composició del plàncton en les diverses estacions d’estudi, permet d’examinar l’associació estadística entre parells d’espècies. Hi ha espècies, fins i tot del mateix gènere, que tenen tendència a viure juntes en certs volums d’aigua. Altres espècies s’exclouen en la pràctica. Això es pot interpretar per les semblances en requeriments ambientals i també en la relació que hi ha entre requeriments ambientals i forma biològica, aquelles característiques que diferencien, per exemple, les diatomees dels dinoflagel·lats, i els dinoflagel·lats aplanats i amb poc pigment, propis d’aigües pobres, dels dinoflagel·lats arrodonits i rics en clorofil·la de les aigües roges. Noteu que aquests tipus biològics són anàlegs als que es descriuen a la vegetació superior, com ara arbres, herbes, teròfits, etc. Si s’escull una representació on les espècies siguin representades per punts, i la distància entre aquests punts sigui inversament proporcional a la probabilitat de trobar-les juntes, dintre d’aquesta representació es pot trobar un pla que reculli una part important de la variància. En línies generals trobem que un pla damunt del qual les espècies es destrien bé, coincideix amb la representació del «mandala» del plàncton. És el que cal esperar si les espècies són diversament característiques de condicions singulars.
Canvis a llarg termini: fluctuacions
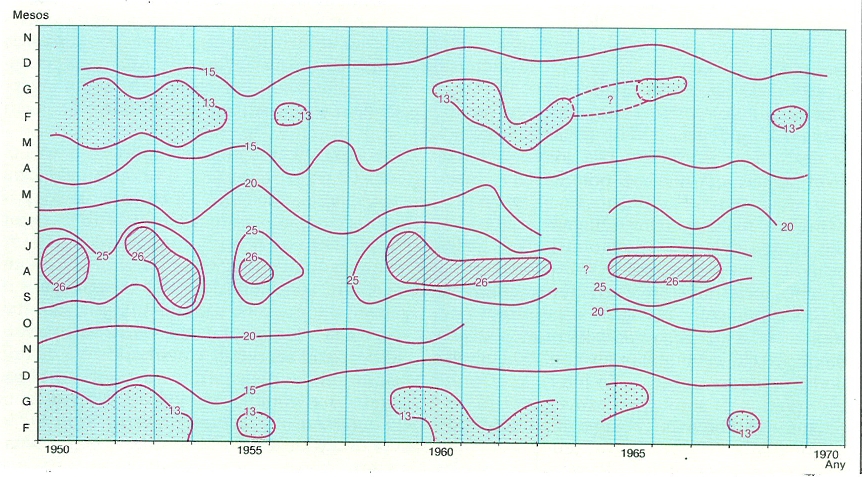
La fluctuació d’uns anys a uns altres en la dinàmica de la nostra mar es pot veure en el canvi de temperatures i salinitats, que mostra unes «ones estadístiques» cada 3-7 anys, que són comparables als «cicles» o fluctuacions observats a El Niño, i que es relacionen amb canvis en les condicions meteorològiques globals. El dibuix recull l’evolució de la temperatura de l’aigua de superfície en l’àrea marina de Castelló. El puntejat indica hiverns excessivament freds, i el ratllat oblic, estius anormalment càlids.
Carto-Tec, original de l’autor.
Amb tantes espècies i amb l’heterogeneïtat de llur distribució, ja podem imaginar que la complicació del plàncton es podria comparar amb la complicació del bosc. Senzillament, el bosc és més estàtic, el temps hi passa més lentament; el plàncton sembla com si estigués subjecte a una agitació termal i es modifica amb rapidesa. Això no obstant, és meravellós com, any rere any, i malgrat els canvis calidoscòpics de les èpoques de barreja i producció, en determinats moments, per exemple, a l’estiu, es reconstrueixen comunitats d’espècies d’una semblança sorprenent. No és com en un bosc que visitem de tant en tant; a la mar, entre una observació i una altra han pogut ocórrer canvis extraordinaris, amb destrucció i reconstrucció d’un sistema de gran complexitat. Però, al llarg dels anys la situació no sempre és tan repetitiva.
Bàsicament, el fitoplàncton de la Mediterrània varia poc. Es pot dir que no ha canviat en els quaranta anys que duren les nostres observacions, malgrat les molècules nocives i recalcitrants a la descomposició que l’home hi va abocant. Certament, la Mediterrània no és pas morta, de cap manera; el que passa és que la vida no hi és gaire densa i que els grans fons són molt més pobres que els grans fons d’altres mars; però gairebé sempre ha estat així, i no és pas conseqüència de contaminacions recents. Si hi ha petites diferències, són fluctuants i irregulars i, de moment, s’escapen a qualsevol explicació: entre les espècies més freqüents a l’àrea de Castelló, algunes són comunes del 1960 al 1967, però, en aquest temps, algunes espècies de Chaetoceros i Cyclotella han cedit la preeminència a Thalassionema nitzschioides, Chaetoceros insignis i Plagioselmis. En el plàncton estival, en aquests darrers anys (1982-1985) Ceratocorys gourreti sembla més freqüent que en períodes anteriors. Es podrien reunir moltes notes d’aquest tipus, però és molt difícil de provar-ne qualsevol significació.
Els canvis que es refereixen a la producció i a la biomassa poden ser més importants que no pas els que afecten la composició florística de les comunitats. Hi ha alts i baixos; els hiverns freds són més productius i, generalment, mostren una certa periodicitat, potser cada 6 o 7 anys (1957, 1963, 1970, 1977, 1985, etc.). Cal entendre que no són períodes regulars, sinó que representen màxims la distància entre els quals no és constant, però que varia poc, i depèn d’algun tipus d’oscil·lador, com ara el bescanvi d’energia entre l’atmosfera i l’oceà, que podria convertir una sèrie d’entrades que variessin aleatòriament, entorn d’un cert valor, en períodes d’aparença més regular. Es tracta, en resum, de fenòmens molt freqüents, dels quals el corrent del Niño, o l’envaïment per aigües càlides d’un extrem del sistema d’aflorament del Perú —o, més generalment, de qualsevol sistema d’aflorament—, n’és el tipus. La Mediterrània no n’és exempta i els períodes dels pics o màxims més notables són, si fa o no fa, semblants arreu, entre 4 i 8 anys.
Com a resum, proposem d’imaginar la nostra mar com un ample sistema líquid, productor i conservador de vida, poblat per organismes microscòpics que tenen el lloc de les plantes a la superfície dels continents. Però la mar és un sistema fluid, amb una dinàmica tan complicada o més, però afortunadament —per a l’observació— més lenta que la de l’atmosfera. Les distribucions de biomassa del fitoplàncton i de la producció primària no són uniformes. Podríem comparar-Íes a una bassa coberta d’escuma verda d’algues unicel·lulars, on aquesta escuma és irregular, fa línies i es pot acumular prop de les vores. A la mar, la producció del fitoplàncton va a glopades en certes àrees, en funció d’interaccions entre atmosfera i oceà, i en altres llocs s’associa amb estructures d’una tenacitat més considerable. Això poden ser els fronts marins. També, la topografia del fons, prop les costes, resulta un factor important de producció en interferir en la inèrcia de corrents marins. És notable la producció entorn de les illes i és interessant de tenir diferents illes per veure com el seu contorn i el relleu dels respectius fons perifèrics condicionen situacions variables de producció. En resum, el conjunt de la nostra mar es podria comparar a un prat animat per vegetals microscòpics, que viuen en un gruix d’uns cent metres al costat de la superfície i que, per damunt de les diferències locals, unes de molt transitòries, altres de més permanents, manté una producció anyal mitjana de 80 gC/m2, que són 200 g de matèria orgànica seca per la mateixa superfície i el mateix temps. Com hem dit, mundialment l’oceà produeix tant com la tercera part de la Terra. El fitoplàncton de la Mediterrània seria comparable, salvant aquestes proporcions, a un prat. L’herba d’un prat de muntanya pot assimilar l’equivalent de 600 g de pes sec per m2, o 6 t/ha. La nostra mar, repetim-ho, en produeix la tercera part; és a dir, queda, efectivament, al límit del desert.
Seria desitjable que l’anàlisi dels fenòmens i dels mecanismes implicats en els processos de producció ens portés a formular un programa que permetés una simulació, és a dir, que a la pantalla de l’ordinador poguéssim tenir una representació de la densitat i de la producció del plàncton en els diversos punts de la nostra mar, i que aquesta representació fos realista, tant en distribucions sinòptiques com en la variació al llarg del temps (núvols que apareixen discontínuament en certs llocs, que s’esvaeixen transferint massa a altres punts, amb altres densificacions més persistents i evolucionant de manera realista). Avui, això es podria aconseguir fins a un cert punt, si prescindim del detall a l’escala més petita, que és tan impossible de predir com la configuració de cada núvol.
El fitoplàncton és la base de la vida pelàgica. L’energia de la seva producció es dissipa, mantenint una munió d’organismes (zooplàncton, peixos, bacteris) que s’estenen molt més enllà dels estrats il·luminats; finalment se’n va tota sota una forma de longitud d’ona més llarga i de calor, i els elements químics retornen al medi i reprenen el cicle.
Els organismes heteròtrofs
L’excedent de matèria orgànica produïda sobre la respirada, per part del plàncton vegetal a les capes il·luminades de la mar, és consumit i respirat per altres organismes. Altrament, s’acumularia incessantment. Aquests altres organismes són els anomenats heteròtrofs i, en línies generals, formen dos grups molt importants. Uns —principalment bacteris— són petits, absorbeixen materials orgànics a través de la seva superfície, que és relativament molt extensa (osmòtrofs), a conseqüència de les dimensions petites dels individus i, en certa manera, es poden considerar poc eficients, perquè usen molta energia per a mantenir una organització relativament limitada. Moltes «algues» del plàncton i, entre elles, una fracció dels dinoflagel·lats més gran del que s’acostuma pensar són també heterotròfiques. Els altres heteròtrofs són els animals, més grossos, que s’alimenten ingerint i confinant a l’interior del cos partícules de matèria orgànica o que contenen matèria orgànica (fagòtrofs), i en els quals sembla com si l’energia fos més ben utilitzada, perquè sosté estructures complexes i sorprenents, seu d’una activitat variada que es pot estendre fins al nivell cultural.
Bacterioplàncton
La bacteriologia ha arrossegat, durant molt de temps, el prejudici de considerar els bacteris gairebé només com a causa de malalties. Així doncs, l’estudi dels bacteris que formen la flora marina normal ha estat negligit massa temps, i encara avui en sabem ben poca cosa. Pensem, però, que la biomassa bacterial marina és sovint entre 0,1 i 2 mg de carboni per m3, del mateix ordre que el fitoplàncton o poc menys, i que s’estén en fondària, fora dels nivells fòtics. Pensem també que més de la meitat de la producció neta del fitoplàncton a l’últim és utilitzada pels bacteris. De fet, una visió que contrasta fortament amb la tradicional, i que ara sembla més a prop de la realitat, ens presenta l’oceà com un món en el qual els heteròtrofs més importants són bacteris i, solament en segon lloc, veiem un plàncton animal (zooplàncton) que apareix i es va diversificant, tot adquirint una importància relativa progressivament més gran, però sempre damunt del fons constituït per la població bacteriana. Certament, hi ha una mena de simbiosi entre el zooplàncton i els bacteris: d’una banda, tot enllaç tròfic facilita el desenvolupament de bacteris, entorn de la boca, en els residus no menjats, a l’interior del tub digestiu, als excrements. D’altra banda, materials sòlids produïts o compactats pels animals, excrements i mudes, són el suport de denses poblacions bacterianes que en modifiquen contínuament la qualitat i també el valor nutritiu, si s’escau, i que són ingerits, de nou, per ells mateixos o per altres animals.
L’activitat dels bacteris és important en la transformació final de moltes molècules orgàniques que formen el que anomenàvem la necromassa o la necrosfera, i són les molècules recalcitrants a la descomposició bacteriana, o bé molècules resistents que resulten de l’activitat dels mateixos bacteris, les que formen la gran quantitat de matèria orgànica, morta i més o menys inert, que es troba dissolta en l’aigua de la mar. L’observació directa del plàncton mostra sempre més material mort, partícules detrítiques, en forma de mudes, organismes morts, excrements i altres residus que no pas plàncton viu.
Entre aquest material detrític hi ha coses sorprenents, com ara fibres procedents de paper i esteses per tots els oceans, mucilag, petites fibres col·loïdals de pocs nanòmetres de llargada, etc. En certes condicions, aquest material inert, incloent-hi de vegades plàncton viu o moribund, s’agleva o s’aglomera formant una mena de flocs, anomenats neu marina. En aigües sumament quietes, a gran profunditat, l’aglomeració es pot produir simplement per forces físiques entre partícules que passen a prop les unes de les altres o es toquen; de totes maneres, té interès perquè les partícules més grosses que en resulten sembla que augmenten la velocitat de sedimentació. Però també es produeix neu marina en altres situacions, per mitjà de mecanismes poc coneguts. Un exemple: pel febrer del 1957, bussos actius prop de Castelló notaren un enterboliment de l’aigua que els impedia de treballar; la causa n’era una pèrdua de transparència generalitzada a tota l’àrea de pesca d’aquell port, que durà del 19 al 28, amb un màxim el 23 de febrer. A l’aigua es trobava un material floculat, format d’una trama mucilaginosa en la qual eren englobades partícules minerals, bacteris, plàncton viu i, principalment, plàncton mort, però sense restes de zooplàncton pròpiament gelatinós, com ara les salpes o els ctenòfors. Contribuïen a compactar aquest material espècies de Chaetoceros (alguna amb mucilag propi), acantaris socials de voluminosa calima (coberta mucilaginosa), agulles de Globigerina i coccolitoforals. La part de cendres era del 65 al 72% del pes sec, cosa que correspon a diatomees i a una part considerable de sèston mineral. Sembla que es tractava del desenvolupament de microorganismes —bacteris principalment— sobre una massa de material detrític i plàncton morent. Però, quina en podia ser la causa? Era un moment d’aflorament local, amb forta producció. Potser algun canvi brusc, potser tèrmic, ja que la temperatura a la superfície passà a 15°C en un parell de dies, després d’uns quants dies prop de 13°C, determinà la mort de molt de plàncton, que anava sedimentant-se en més aigua ascendent, tot fent de nuclis de multiplicació d’organismes variats. El fenomen desaparegué ràpidament. No hi ha dubte que altres formacions comparables de neu marina —que no són totes d’una causa comuna— es produeixen ací i allà, de tant en tant.
Hi ha una altra relació entre el material detrític en suspensió —eventualment la neu marina— i els bacteris marins, en el sentit que s’ha debatut sovint si hi ha més bacteris lliures, en suspensió, o si la majoria és associada al material detrític. Durant un quant temps es pensava això darrer; ara hi ha força dades que semblen indicar que la major part dels bacteris poden trobar-se separadament en suspensió. Probablement depèn de condicions locals i potser cal esperar una major propensió a associar-se a partícules de sèston, si l’aigua té una capacitat nutritiva més baixa.
La proporció pot arribar i arriba sovint a deu vegades més de material detrític que de plàncton viu. Doncs bé, la fracció de matèria orgànica dissolta és encara més elevada, i és ordinàriament entre 0,5 i 1,5 mg de carboni orgànic per litre a la Mediterrània. Podríem precisar més i acceptar valors de 1,2 a 2 mg en aigües superficials, entre 0,5 i 0,7 mg en aigües fondes, i potser valors més alts prop de la costa. La mar conté, doncs, entorn de 25 g de carboni inorgànic, i d’1 g de carboni orgànic dissolt per metre cúbic. La concentració de la vida és molt més baixa. A la nostra mar els valors més alts de fitoplàncton són de 100 mg de C/m3 com a valors màxims, fins a concentracions negligibles en aigües fondes, i concentracions de zooplàncton que van entre 5 i 8 mg de C/m3. Quant als bacteris, poblacions que consten de 10 milers de cèl·lules/ml són equivalents a 1 mg de C/m3, una mica menys que el fitoplàncton en zona fòtica, però més que el fitoplàncton de més avall.
Zooplàncton
Els heteròtrofs representen una organització més o menys jerarquitzada que explota els productors primaris. Ordinàriament, es parla de cadenes o xarxes tròfiques, o d’una organització en forma de piràmide, seguint la regla general que «el peix gros es menja el petit». En una situació persistent sembla que el nombre d’individus ha de ser menor si els individus són més grossos. De fet, relacions d’aquest tipus són molt generals a la vida marina i permeten una certa sistematització de la vida pelàgica, o almenys són en relació amb les característiques que deuen tenir els diferents estris emprats per a la captura i l’estudi. Si les classes de dimensions s’estableixen segons una escala logarítmica, o progressió geomètrica, sovint les biomasses de les diferents classes tenen una distribució d’allò més regular. Regularitats semblants s’estenen a la distribució del sèston en general, és a dir, a les partícules molt petites, en les quals no es pot discriminar bé entre vives i mortes. Així es determina amb els comptadors dimensionals de partícules basats en les pertorbacions que les partícules introdueixen en la conductivitat elèctrica de l’aigua (comptador Coulter). Semblants espectres, quan hom ha reunit certa experiència —experiència que, de fet, ja existeix a la Mediterrània i per les nostres aigües— tenen un considerable valor indicador, amb relació a la procedència de les masses d’aigua i dels fenòmens biològics que s’hi han esdevingut en un passat pròxim.
No és senzill de fer i de portar a terme un programa adequat de recerca per al zooplàncton. Es tracta d’animals de mides diferents, que s’estenen fins a les màximes fondàries marines, que es mouen de manera diversa —alguns són tan àgils que s’escapen fàcilment de les xarxes, com ara molts eufausiacis, que sovint són subrepresentats en el material recol·lectat—, i que, sovint, tenen una distribució sumament irregular, formant eixams, taques o bancs.
Mentre que el fitoplàncton marí no es pot dir que sigui gaire més variat que el fitoplàncton de les aigües dolces o continentals, al zooplàncton hi ha diferències considerables. El zooplàncton animal de les aigües dolces és sorprenentment pobre en comparació del marí. Al zooplàncton marí hi ha representats un nombre molt gran de grups zoològics i hom pot creure que el domini pelàgic ha estat un escenari important en l’evolució dels animals. També es troben al zooplàncton costaner larves pelàgiques de molts animals que, en ser adults, s’incorporen a la vida bentònica. Com que només es troben al plàncton una part de la seva vida, s’anomenen meroplàncton, que vol dir plàncton a temps parcial. Òbviament les reconstruccions orgàniques que pressuposa aquesta mena d’adaptació (per exemple, les larves i els adults en un bàlan, en un eriçó de mar, en un mol·lusc) poden ser una via de diferenciació i evolució a la qual la naturalesa ha recorregut probablement més d’un cop.
Una distinció òbvia en el zooplàncton es fa entre els organismes unicel·lulars (foraminífers, radiolaris, acantaris, ciliats) i els pluricel·lulars (cnidaris, quetògnats, tunicats, crustacis; amb menor representació relativa, ctenaris, anèl·lids, mol·luscs). Una diferència és la mesura: els unicel·lulars fan d’una fracció de mil·límetre a uns quants centímetres (algunes formes colonials de radiolaris, com Collozoum, no rars a la nostra mar), mentre que els pluricel·lulars, o metazous, són més grossos, encara que també n’hi ha de fracció de mil·límetre. Alguns organismes que, per la seva afinitat d’organització, anirien més amb el fitoplàncton, són, en realitat, animals, com ara diversos dinoflagel·lats, tals com Noctiluca, Scaphodinium, i altres. Juntament amb la mesura, hi ha una altra diferència important que, en cert sentit, va associada a la mesura: la taxa de renovació de les poblacions. Els ciliats, per exemple, poden ser molt abundants i constituir el 20% de la biomassa del zooplàncton total; però, agafant aquest exemple com una situació que s’esdevé sovint, podem dir que llur importància en la producció és molt major, ja que les poblacions de ciliats es poden renovar en qüestió de dos dies a una setmana, mentre que el zooplàncton pluricel·lular i més gros renova les seves poblacions en setmanes o mesos. Així doncs, la contribució del zooplàncton unicel·lular a la producció pot ser desproporcionadament gran.
Animals planctònics unicel·lulars
L’organització molt simplificada dels protozous unicel·lulars no sempre fou reconeguda. El 1826 d’Orbigny descrivia els foraminífers com si fossin petits cefalòpodes (sèpies, pops i ammonítids), i Ehrenberg, el 1833, podia referir-se a molts unicel·lulars com a organismes perfectes i complets (que ho són) i de complicació comparable als animals superiors (que no la tenen).
Els foraminífers, de closca calcària perforada i, per tant, conservables en estat fòssil, són molt importants en paleontologia i, especialment, en la paleontologia aplicada a l’estudi dels estrats petrolífers. Aquesta utilitat ha fet que l’estudi dels foraminífers hagi disposat de recursos notables i estigui força avançat, cosa que ha beneficiat també el coneixement dels foraminífers planctònics. La composició isotòpica de l’oxigen de les closquetes de fora minífers pelàgics, conservades al sediment marí, ha permès conclusions interessants sobre la temperatura de l’aigua en èpoques passades (Emiliani). Cal remarcar que els foraminífers no són fòssils que es conservin igualment en totes les condicions. La naturalesa calcítica de les seves closques fa que es dissolguin naturalment en arribar a una certa fondària (entre 3800 i 4200 m, l’anomenada lisoclina, amb referència a lisi, desfer), segons la tensió local d’anhídrid carbònic i el grau de saturació del calci. Això vol dir que a les grans fondàries oceàniques no s’acumulen foraminífers, sinó només restes silícies d’altres organismes, com ara radiolaris i diatomees. Hi ha poques espècies de foraminífers planctònics, i en les acostumades llistes de plàncton se simplifica encara més llur taxonomia, ja que es distingeix solament entre Orbulina universa, Hastigerina pelagica, i «Globigerina» que, en realitat, és un complex de diverses espècies —que avui dia es distribueixen en gèneres de noms diferents (Globigerinoides, Globorotalia, etc.)—, que necessiten una cura considerable per a la seva diferenciació. I no obstant això, seria important de separar-les, perquè hi ha proves que canvis climàtics, sobre dècades o segles, es reflecteixen en la representació relativa de les diverses formes. La dificultat en la determinació precisa de les espècies es complica amb la variabilitat intraspecífica. Trobem globigerines amb agulles o sense agulles calcàries damunt la closca, i aquesta pot ser més o menys gruixuda i, sovint, envoltada per una coberta d’aspecte gelatinós. El malaguanyat biòleg francès Jean Le Calvez estudià Orbulina a la nostra Mediterrània i notà com en capes superiors (50 m) la closca globigeriniforme es conserva encara adossada a l’interior de la gran cambra globosa, però que, més avall, va desapareixent a mesura que els animals es fan més vells. Tot fa creure que es reprodueix a fondàries de 200 a 400 m, i els joves pujarien cap amunt. D’altra banda, Emiliani trobà que les Orbulina grosses donen una temperatura isotòpica (160/180) diferent de les petites, i en el sentit postulat. Diferències verticals en característiques reproductives, mesura i calcificació de les globigerines tenen un caràcter diferent just en el front de la mar catalano-balear i als costats del front; a més, les globigerines són més freqüents al front, i això suggereix que el moviment ascensional de l’aigua no solament pot fornir nutrients, sinó també alterar equilibris que poden tenir influència en la deposició de carbonat de calci.
Els rizòpodes tenen, sovint, algues en simbiosi; als foraminífers és conegut de fa temps i ara mereix una atenció creixent. Als acantaris és molt manifest. Els acantaris són rizòpodes de fins uns quants mil·límetres de diàmetre, amb un esquelet de forma variable, a base de varetes radials que contenen una gran quantitat de sulfat d’estronci. Això no ens ha de meravellar, ja que l’aigua de mar conté normalment uns 8 mg/l d’estronci, que és el doble de la concentració normal de bor i quatre vegades la concentració normal de silici. L’acumulació d’estronci pels acantaris va fer que, després de la guerra de 1939-45, hom tractés d’utilitzar-los per a descobrir la presència de l’estronci radioactiu (89Sr, vida mitjana de 51-53 dies; 90Sr, vida mitjana 20-29 anys; l’estronci normal és una barreja dels isòtops 88Sr (82%), 86Sr, 87Sr i 84Sr). Amb aquesta finalitat férem algunes recol·leccions d’acantaris al llarg de la nostra costa, on, sovint, es troben en grans quantitats, i les enviàrem; però sembla que els resultats no foren sensacionals. L’ocasió s’aprofità per a fer mesures de la quantitat d’algues que els mateixos acantaris portaven en simbiosi, cèl·lules que pertanyen al grup dels dinoflagel·lats. La participació de les algues, en termes de quantitat i qualitat de pigments, és semblant en els coralls hermatípics (madrèpores) i en els acantaris pelàgics, almenys de la Mediterrània (Margalef 1961), de 0,15 a 0,19 mg de clorofil·la per gram de matèria orgànica seca, sense cendres. Una tal concentració garanteix una autonomia pràcticament total a la simbiosi.
Els radiolaris són organismes interessantíssims, ben coneguts per la bellesa dels seus esquelets silicis, reproduïts des de les làmines de «Kunstformen der Natur» de Häckel, fins a les fotografies obtingudes amb el microscopi d’escandallatge. D’aquestes espècies, no en tenim gaire informació referida a la nostra mar, on els gèneres més abundants són formes colonials a les quals diverses càpsules individuals són incloses en una massa comuna. Aquests (Sphaerozoum, Collozoum) tenen algues en simbiosi i també mengen; aquests dos recursos deuen cobrir llurs necessitats meitat i meitat. Sticholonche zanclea és un organisme molt curiós, freqüent a la Mediterrània, i també en altres mars, que, per la seva organització, s’apropa als radiolaris, encara que té un aspecte força diferent. És molt característic per unes espícules silícies allargades, disposades en feixos, cadascuna amb una base eixamplada en forma de paleta i una part distal de vores denticulades. Espícules despreses són un objecte freqüent a les mostres de plàncton, i són origen de confusions per al qui comença.
Els ciliats són un component important del zooplàncton. A la Mediterrània són força nombrosos, com mostra la distribució de les seves densitats en una sèrie de mostres estudiades procedents de les nostres costes.
Amadeu Blasco, original de l’autor.
Els ciliats constitueixen un grup molt important, en tota mena de medis, però al plàncton, encara que nombrosos, pertanyen a un grup de tipus d’organització més restringit: oligòtrics i perítrics. Els perítrics són més escassos, però no són rares grans colònies ramificades nedadores de Zoothamnion i algunes Vorticella, inserides usualment damunt de colònies de grossos Chaetoceros (com Ch. coarctatus). Entre els oligòtrics, en tenim tres tipus, cadascun dels quals té el seu interès particular. Mesodinium, amb «mitges» algues en simbiosi, pot arribar a ser tan nombrós que fa vermella l’aigua. Les algues permeten aconseguir un balanç metabòlic positiu. Lohmannella i altres organismes semblants són cònics, petits i pràcticament nus. L’extens grup dels tintínnids es caracteritza perquè les cèl·lules són envoltades o protegides per una coberta inerta resistent. Són organismes freqüents i amb cobertes molt belles; la seva variabilitat ha determinat una subdivisió excessiva de les espècies. Aquestes cobertes es construeixen amb gran rapidesa i d’una manera encara un xic misteriosa. Sovint, un cop enllestides, s’hi adhereixen partícules minerals, o peces dures de les cèl·lules de què s’alimenten (coccòlits). La mateixa càpsula, o lorica, pot contenir una fracció considerable de silici. Hom ha reunit força informació sobre ciliats del plàncton marí a les nostres costes, i s’han estudiat, recentment i amb detall, les poblacions de la regió del golf del Lleó. La relació quantitativa entre ciliats i fitoplàncton és entorn d’1:50 en nombre de cèl·lules i d’1:10 en biomassa, cosa que permet de reconèixer la importància dels ciliats com a elements de la cadena tròfica pelàgica. Les cèl·lules nues o gairebé nues es poden consumir més fàcilment i es multipliquen més de pressa que els tintínnids loricats, de manera que es reconeix una ordenació ecològica de les espècies al llarg d’un eix d’estratègies reproductives i potencialment evolutives. Si bé la presència de lorica és una defensa enfront de depredadors dels ciliats, és clar que és un llast per a l’animalet. La lorica buida de Stenosemella pesa fins a 0,25 μg i la cèl·lula viva del mateix ciliat entre 0,02 i 0,08 μg. Quan les cèl·lules es divideixen, una de les filles hereta la vella lorica i l’altra en fa una de nova, i hi ha raons per a creure que la primera té tendència a anar cap avall i l’altra a mantenir-se en capes superiors, on sovint troba més aliment i es divideix més de pressa. El resultat és que hi ha una estratificació, no sols de les espècies de tintínnids, sinó també de formes diferents dintre de cada espècie, una mica com en els foraminífers, encara que, potser, per altres raons. Els tintínnifd mengen bacteris, petits flagel·lats, coccolitofòrids, dinoflagel·lats i petites diatomees (Thalassiosira). A les nostres costes, les màximes densitats són de més de 10 000 cèl·lules de ciliats per litre; acostumen a ser més baixes i disminueixen gradualment de la superfície fins al fons. La proporció de ciliats loricats és petita, potser només el 5% del total. Els ciliats nus són els que poden tenir més interès en l’alimentació de les larvetes de peixos, que moltes vegades no se sap què mengen. És clar que si s’ingereixen tintínnids loricats, les cobertes queden com a testimoni en el contingut digestiu, però els ciliats nus es desfan pràcticament com una mandonguilla de carn o com un caramel, i fan més difícil tot intent d’avaluació de llur importància en el cicle de la vida marina. Alguns tintínnids són associats habitualment amb diatomees vives. En aquesta relació, el ciliat les manté en suspensió i és possible que alguna cosa de les algues, si més no, bacteris que viuen a prop, arribi a alimentar el ciliat.
Un problema persistent en el coneixement dels ciliats del plàncton és que passen a través de les xarxes que s’usen per a recollir el zooplàncton, i els fitoplanctòlegs, que acostumen a sedimentar llurs mostres, no es preocupen dels ciliats, perquè no és el seu ofici. És, certament, una especialitat diferent i el seu estudi és molt necessari, si més no, com s’acaba d’indicar, per a entendre la supervivència de les etapes més jovenívoles de la vida dels peixos.
Animals planctònics pluricel·lulars

La mida i la velocitat dels organismes, en relació amb la viscositat de l’aigua, condicionen la vida dels animals del plàncton, com mostra aquest diagrama de C.H. Mortimer. En hidrodinàmica, s’usa el «nombre de Reynolds» (velocitat x alguna dimensió lineal característica de l’organisme/viscositat de l’aigua), que en el dibuix és situat al costat de les ratlles inclinades interrompudes. A partir d’un cert límit, és inevitable un règim turbulent, on la viscositat cinemàtica (v) s’hauria de substituir pels valors més alts que correspondrien a una altra definició.
Ricardo Génova.
La major part dels nostres coneixements sobre les característiques, la biologia i la distribució dels animals planctònics pluricel·lulars, que són generalment més grossos, entre 0,5 i 50 mm, s’han obtingut a base de material pescat amb xarxes còniques de malla de 0,14 o de 0,4 mm. Com que les poblacions de zooplàncton són poc denses, cal combinar el diàmetre de la xarxa i la llargada de la pesca per filtrar una quantitat d’aigua suficient (uns quants metres cúbics com a mínim). És clar que si s’empra una xarxa d’obertura petita cal remolcar-la més temps, i això pot integrar poblacions que viuen al llarg del trajecte recorregut i es perd la possibilitat de destriar diferències més locals. D’altra banda, si la xarxa avança lentament, molts animals s’escapen davant i, si va més ràpida, cal assegurar una bona relació entre la superfície filtrant i la boca. Cal que la corda que serveix per a remolcar la xarxa no sigui tesada, perquè en aquest cas bona part de l’aigua rellueix pel davant i la filtració no és quantitativa. Es pot instal·lar un comptador de paletes per avaluar la quantitat d’aigua realment filtrada. Avui dia, s’han ideat dispositius força enginyosos per obrir i tancar les xarxes a fondàries determinades i per augmentar el control sobre el que realment s’extreu. Tot això ha complicat força l’aspecte i el maneig d’un dispositiu que, essencialment, es començà a usar sistemàticament en biologia marina fa més de 150 anys (almenys des del 1828 per V. Thompson a Irlanda) i que, en una forma semblant, ja es veu en còdexs asteques que representen la pesca de petits crustacis i insectes aquàtics usats en l’alimentació. Per a l’estudi dels elements més petits i nombrosos del zooplàncton, com ara els copèpodes (poblacions de 0,5 a 5 individus/1 en aigües de les capes superiors), es poden utilitzar ampolles ordinàries d’hidrografia de cabuda suficient (fins a 20 l) que permeten d’obviar moltes de les diverses dificultats que comporta l’ús de les xarxes.
Els organismes del zooplàncton es poden detectar, també, amb el sonar, usant freqüències elevades. Partícules d’1 mm demanen ones de 3 mm, que, amb una velocitat de propagació del so de 1500 m/s serien de 500 kHz; però, si el zooplàncton és abundant o forma eixams, és perceptible als detectors de freqüència més baixa usats pels pesquers, d’entre 10 i 30 kHz; aquests permeten de veure la migració vertical del zooplàncton en forma de les onades d’ecos que es van aproximant vers la superfície al capvespre i que retornen cap avall abans de clarejar.
Del zooplàncton formen part organismes meravellosos, però generalment delicats, ja que les parts dures es troben reduïdes al mínim; sovint hi ha embolcalls de contextura gelatinosa i és molt freqüent l’existència d’apèndixs flexibles i retràctils, que, dins de l’aigua, es mouen amb una elegància que no sobreviu a la captura i a la fixació (generalment amb formol) del material. És per això que l’observació directa dels organismes, en immersió, o per mitjà de fotografia fixa o animada, va resultant un complement indispensable per a l’estudi del plàncton.
Tradicionalment, hom feia una distinció entre plàncton, que són els organismes petits que deriven a la mercè de l’aigua, i nècton, que fa al·lusió a la capacitat de nedar, i que comprèn crustacis grossos, cefalòpodes, peixos i cetacis. Pot semblar que la distinció no és correcta, perquè hom podria fer un enfilall de tota mena d’organismes, classificats per grandàries, des del bacteri fins al cetaci més gros. Però la distinció sí que és correcta si hom té en compte les condicions dinàmiques. En hidrodinàmica, hom considera alguns nombres, sense dimensió, que caracteritzen propietats dels fluids; per exemple, el nombre de Reynolds, que és igual a una llargada característica de l’objecte o organisme que es considera, multiplicat per la velocitat i dividit el producte per la viscositat cinemàtica del fluid. Per a organismes passius i petits, el nombre de Reynolds és molt inferior a la unitat, i la viscositat de l’aigua els confina en un món tot especial. Per a organismes més grossos, arriba un moment que les forces d’inèrcia esdevenen més importants que la viscositat que s’oposa al moviment i, aleshores, a petits augments de l’energia invertida en el moviment correspon un augment molt més gran de la velocitat. Som en el domini de copèpodes i zooplàncton petit. Més enllà, en el veritable nècton, arribem a valors del nombre de Reynolds que s’apropen als 10 000 o els superen. Ací, els remolins que els organismes deixen darrere seu, la forma ajustada a les exigències de la hidrodinàmica, tenen una importància creixent. Hi tenim la forma típica de peix, arrodonida pel davant i més afuada pel darrere, les aletes flexibles, que tendeixen a destruir els remolins importants del darrere, o la pell flexible i elàstica dels cetacis que absorbeix les pertorbacions del fluid i, en minvar la formació de remolins posteriors, millora, a l’ensems les condicions de propulsió. Sovint, hi ha carenes laterals i, lògicament, tota l’organització s’emmotlla a l’estratègia de la mobilitat. Les velocitats més freqüents del nècton acostumen a quedar entre 1 i 4 m/s, a les tonyines fins a 20, en general, entre 2 i 10 vegades la longitud de l’individu.
Tipus biològics de zooplàncton
La mobilitat i la mateixa conformació del cos permeten distingir dos grups molt importants en el plàncton. D’una banda, els crustacis i, potser, alguns animals d’altres grups, com ara anèl·lids i quetògnats, que es mouen amb força rapidesa i migren activament en la dimensió vertical. Potser, els més importants són els copèpodes i els eufausiacis. Els copèpodes tenen unes dimensions mil·limètriques, i una velocitat de migració de 9 a 30 m/h. Els eufausiacis tenen entorn d’1 cm i la seva velocitat arriba a uns 90 m/h. Aquests grups corresponen, doncs, a escales de significat diferent, i el seu conjunt constitueix sovint del 60 al 70% de la biomassa del zooplàncton. Hi ha motius per a creure que aquest zooplàncton essencialment migrador és adaptat a l’explotació, a contracorrent, d’un fitoplàncton en forma de taques discontínues de gran productivitat, on poden predominar les diatomees. És clar que hi ha, també, moltes espècies carnívores, del mateix i d’altres grups, que formen part d’aquest complex.
En línies generals, el plàncton que podem anomenar «gelatinós» consisteix en organismes de cos molt empapat d’aigua i, sovint, diàfan (densitat entre 1,024 i 1,07), poc nutritiu i amb baixa intensitat respiratòria, i amb una mobilitat que no es pot considerar extraordinària. Uns filtren (tunicats, com ara les salpes o el llepó), i d’altres son carnívors (meduses, sifonòfors, ctenòfors i alguns mol·luscs). Aquest plàncton gelatinós, a les mars actuals, mai no sembla ser dominant, però és possible que, en oceans del passat, alguns organismes que, funcionalment, hi eren molt a prop, com una sèrie de cefalòpodes amb closca parcialment plena de gas i, per tant, amb la possibilitat de flotar entre dues aigües, hi fossin realment molt abundants. Es pot dir que el plàncton gelatinós és sota el patrocini d’Arquimedes; de fet, en alguns casos, hi ha cavitats aeríferes o, més ben dit, gasíferes que en faciliten la flotació, com en els cefalòpodes suara esmentats (Nautilus actuals i formes fòssils) i en els sifonòfors proveïts d’una bombolla de gas en els individus que encapçalen la colònia. Tot això contrasta amb els més enèrgics nedadors, per als quals la resistència i la viscositat de l’aigua són factors molt importants.
En una visió mundial de la vida pelàgica, el plàncton ric en copèpodes i eufausiacis és molt més dens entorn de les àrees de surgència, vers els corrents orientals (Sàhara i Namíbia, a l’Atlàntic), mentre que el plàncton gelatinós té una representació relativa més important vers els límits occidentals i en els girs del centre dels anticiclons dels mateixos oceans. Es pot suposar que en èpoques de major estratificació oceànica, durant la història de la Terra, el plàncton gelatinós o «arquimedià» fou més freqüent que no pas ara.
Encara caldria fer una distinció vertical important en el plàncton, diferenciant aquells components que viuen sempre associats a la superfície de l’aigua. Una tal relació admet graus diferents. Hi ha copèpodes, generalment de color blau, que sovint es mouen prop la superfície, en eixams, i alguns es llancen contra la pel·lícula superficial de l’aigua, fins al punt que, amb mar calma, fa l’efecte que plou, per les petites deformacions amb què sembren la llisa pel·lícula del líquid. Altres floten, senzillament, damunt de l’aigua, com Velella i Physalia, que son celenteris colonials molt característics i coneguts. El segon és el «carallo a la vela» dels pescadors gallecs, nom substituït per l’eufemisme «caravel·la» o «caravel·la portuguesa». Aquests organismes deriven sota l’impuls del vent i són construïts de tal manera que ho fan cap a la dreta o cap a l’esquerra, segons llur organització. Sembla que sempre són produïts en igual quantitat els dos tipus i, si les poblacions s’associen amb remolins de direcció anticiclònica, com passa a la Mediterrània occidental, les que tenen més probabilitat d’arribar a la costa són les sinistres. De fet, aquestes arribades es produeixen, sovint, a la primavera, quan una prima capa superficial d’aigua passa pel damunt del front i s’estén fins a la costa: les platges queden, aleshores, cobertes d’una munió d’animalets blaus i cartilaginosos de fins un parell de centímetres de llargada. Uns mol·luscs gasteròpodes, blaus com les colònies de Velella, naveguen units a la superfície per un flotador que es fabriquen i que conté bombolles, com ara certes formes industrials de plàstic, i s’alimenten de Velella; són els del gènere Janthina. Els flotadors de Janthina, restes de Velella i, fins i tot, terrossos flotants d’hidrocarburs, són colonitzats per Lepas, crustacis sedentaris i altament modificats del mateix grup dels peus de cabra.
Grups zoològics presents en el plàncton marí
En aquesta mateixa obra, i en l’ordre acceptat de la taxonomia zoològica, es presenta una informació considerable referent a elements molt importants del zooplàncton marí. Hi remetem el lector, però no volem deixar de consignar aquí alguns aspectes d’especial interès en termes ecològics.
Entre els cnidaris hidraris, distingim els grups següents: les antomeduses (gimnoblasts, atecats), de les quals són freqüents Phialidium, Zanclea i Sarsia, que pot retenir unes preses amb els tentacles mentre va menjant les altres; les leptomeduses (caliptoblasts, tecats), amb Laodicea i Obelia, i Aequorea, que arriba a 17 cm de diàmetre; les traquimeduses, com Rhopalonema, Lyriope, Geryonia, Aglaura i Aglantha digitale, que és una de les meduses petites més freqüents a la nostra mar; les narcomeduses, amb Solmaris i Cunina; i les pteromeduses, un grup molt limitat, representat per un animal de forma extraordinària: Tetraplatia volitans; les limnomeduses, amb Olinidas, i Odessia maeotica, que entra en aiguamolls, i els condrofòrids, flotants i que naveguen amb el vent, com ara l’esmentada Velella, i Porpita, que és més rara i de forma circular. Un altre grup de celenteris és el dels sifonòfors, organismes colonials, en els quals s’inclouen els ordres següents: els cistonèctids, que són les boniques Physalia que floten a la superfície; els fisonèctids, estesos en forma de llargs filaments, amb elements de caça que simulen copèpodes (un exemple de mimetisme agressiu) i amb un flotador que conté gas, com a Forskalia i Physophora; i els calicofòrids, de forma més recollida i sovint amb aspecte de peces de cristall tallat, dels quals són freqüents espècies dels gèneres Muggiaea, Chelophyes i Bassia. La classe de les grans meduses o escifozous, amb una complexitat estructural més gran, conté quatre grans ordres. Les cubomeduses, que es poden considerar gairebé com una classe diferent, atès que tenen característiques pròpies molt peculiars, són representades per Charybdea; en aquest grup s’inclou la medusa més verinosa del món, responsable de bastants accidents mortals, í que viu a Austràlia: Chironex. Els coronats, figuren al plàncton amb Periphylla, una bonica medusa de fondàriaria que no és rara, i Nausithoe. Els semeostomes són meduses molt abundants, dels gèneres Aurelia, Pelagia i Chrysaora. Aurelia és costanera, blavissa i amb una mena de dibuix en forma de creu; Chrysaora té tot un cercle de marques fosques en forma de compassos oberts per fora; Pelagia té puntets vermells, és força lluminosa i potser és l’espècie més implicada en les alarmes periòdiques sobre abundàncies excepcionals de meduses, gràcies al fet que es pot reproduir directament per ous, sense passar la fase bentònica de pòlip. Sovint s’acumulen en rengleres al llarg de les convergències associades amb les cèl·lules o bandes de Langmuir. Els rizostomes són meduses que, secundàriament, han modificat la boca i han passat a ser filtradores de plàncton petit (poden menjar copèpodes); Rhizostoma és una gran medusa blava, ben coneguda des de temps antic (el «pulmo marinus» dels autors del Renaixement).
Els ctenòfors tenen vuit fileres meridianes de paletes nedadores d’aspecte irisat (a causa de la seva estructura de laminetes; és el mateix principi físic dels teguments d’insectes o de les escates de les papallones, o dels filtres d’interferència); els de l’ordre dels nus (Nudae), representats per Beroe, no tenen tentacles. Els ordres restants tenen tentacles que funcionen a manera de palangres; són els tentaculífers, amb Hormiphora, Ciddipe, Callianira, com glòbuls de gelatina de la mesura entre un cigró i una cirera; els cintiformes, amb l’original cinturó de Venus (Cestus veneris); i els lobulats, que solen ser intensament lluminosos, com Eucharis.
Entre els cucs pelàgics, són notables algunes famílies d’anèl·lids poliquets (alciòpids, tomoptèrids) que han arribat a tenir una forma semblant i convergent, amb els paràpodes eixamplats.
Els quetògnats constitueixen un grup zoològic, altament característic, d’animals carnívors en forma de petit torpede (1-3 cm), sageta o míssil, amb diverses aletes fines estabilitzadores i nedadores i una mena de peces dures o garfis a la boca. Són molt abundants i el gènere més comú n’és Sagitta, amb diverses espècies.
Els mol·luscs pelàgics son gasteròpodes i cefalòpodes; aquests darrers, per la seva mesura i mobilitat, es consideren, generalment, com a part del nècton. Dos grans grups de gasteròpodes són representats al plàncton: els estreptineures i els eutineures. Aquests noms es refereixen a detalls de l’anatomia, però els representants dels dos grups al plàncton són força diferents. Els estreptineures són representats pels heteròpodes, profundament modificats per a l’adaptació a la vida pelàgica, amb el cap allargat com una trompa i el peu convertit en una mena d’aleta vertical. La closca pot ser present (Atlanta), rudimentària (Carinaria) o nul·la (Pterotrachea). Tanmateix, arriben a una llargada de més d’un pam. Entre els eutineures tenim el grup dels opistobranquis, amb els pteròpodes, en els quals es distingeixen dos ordres, uns amb closca, en forma d’escut (Clio), paperina (Creseis) o cargol (Limacina). Les closques de pteròpodes formen una part considerable dels sediments en certes mars (potser en més del 0,5% de l’extensió dels oceans mundials). També té una closca, petita i morada, un gasteròpode d’un altre grup, Janthina, ja esmentat.

Les apendiculàries són tunicats planctònics en forma de cap gros que viuen dins de «cases» gelatinoses segregades per la mateixa epidermis de l’animal. El dibuix mostra, a l’esquerra, l’aspecte de la «casa» amb l’animal dins (el conjunt fa només alguns mil·límetres), i, a la dreta, una interpretació més «tècnica» del funcionament. L’aigua de mar (fletxes morades) entra per 1 i surt per la cambra 2, que porta a l’esfínter de sortida (3); és impulsada per les oscil·lacions de la cua de l’animal, dins la cambra 4, que és intercalada al circuit. En el trajecte, l’aigua es filtra (fletxes blaves) en el sistema 5, i el material retingut en el filtre (fletxes vermelles) segueix cap al tub bucal (6) i al cos de l’animal (7).
Amadeu Blasco, original de R. Fenaux.
Els tunicats pelàgics més coneguts, almenys pels pescadors, són les salpes, transparents, amb anells musculars, una obertura anterior i una altra de posterior, i una mena de filtre inclinat en el pas de l’aigua. Arriben a fer 10 cm de llarg i poden formar llargues cadenes, per gemmació. Les vísceres són acumulades en una massa vermella o violada. En són exemples: Thalia democratica, Salpa maxima i S. fusiformis. Els del gènere Doliolum són més petits i tenen forma de barral. En el gènere Pyrosoma, un gran nombre d’individus elementals, d’organització fonamental semblant a la de les salpes, s’uneixen lateralment tot formant una mena de tub o sac que pot arribar a tenir dimensions considerables, especialment en algunes espècies que prefereixen les grans fondàries. Tal com recorda el seu nom genèric, Pyrosoma són fortament luminiscents. Un altre grup de tunicats pelàgics tenen la forma de cap-grossos i són semblants a les larves nedadores dels ascidis, que són tunicats sedentaris quan són adults. Però les apendiculàries no passen d’aquesta forma i, per tant, no són gaire grosses, fins a uns quants mil·límetres generalment, i la seva organització, simplificada d’una banda, l’han adquirida junt amb la capacitat de produir, per secreció de la superfície del cos, una mena de casetes d’aspecte gelatinós que són, ensems, dispositius de filtració; quan són vives, es veuen talment com si fossin glòbuls de gelatina, dintre els quals hi ha l’apendiculària amb una cua ondulant irisada. Els moviments d’aquest apèndix fan circular aigua a través d’un sistema preestablert, del qual formen part uns filtres, on es retenen les partícules que després ingerirà l’animal. A intervals d’unes quantes hores l’apendiculària deixa la casa vella i n’infla una de nova, que ja era «estampada» pel mateix epiteli. Els tunicats pelàgics es reprodueixen ràpidament per gemmació asexual —cadenes de salpes—, procés que alternen amb la producció d’ous d’origen sexuat. La reproducció asexuada és particularment curiosa en els doliòlids, on les gemmes produïdes en un lloc migren per anar a ordenar-se després damunt un estoló d’origen separat.
La producció d’individus molt semblants per reproducció asexuada, és una estratègia molt comuna quan es tracta d’ocupar ràpidament un medi de característiques uniformes, com ara les aigües estivals d’un llac (rotífers, cladòcers), una determinada planta (pugons) o el cos d’un hoste determinat (certs paràsits). És notable que a la mar aquesta estratègia no sigui comuna en el plàncton. Llevat del cas dels tunicats, i especialment de les salpes, que poden formar llargues cadenes amb una activitat filtradora considerable, solament n’hi ha exemples en alguns rotífers que viuen generalment en ports o, almenys, en aigües costaneres, i en algun cladòcer —el grup de les puces d’aigua de les nostres basses— que a l’estiu forma eixams prop de la superfície en aigües càlides (Penilia). El fet d’esmentar aquest cladòcer ens introdueix en el món dels crustacis del plàncton. Els cladòcers són pocs; tampoc no són gaire freqüents els ostràcodes, representats en la regió pelàgica per famílies característicament nedadores (Conchoecia, etc.), que contrasten amb els ostràcodes d’aigua dolça que, per la seva organització, són tots característicament bentònics.
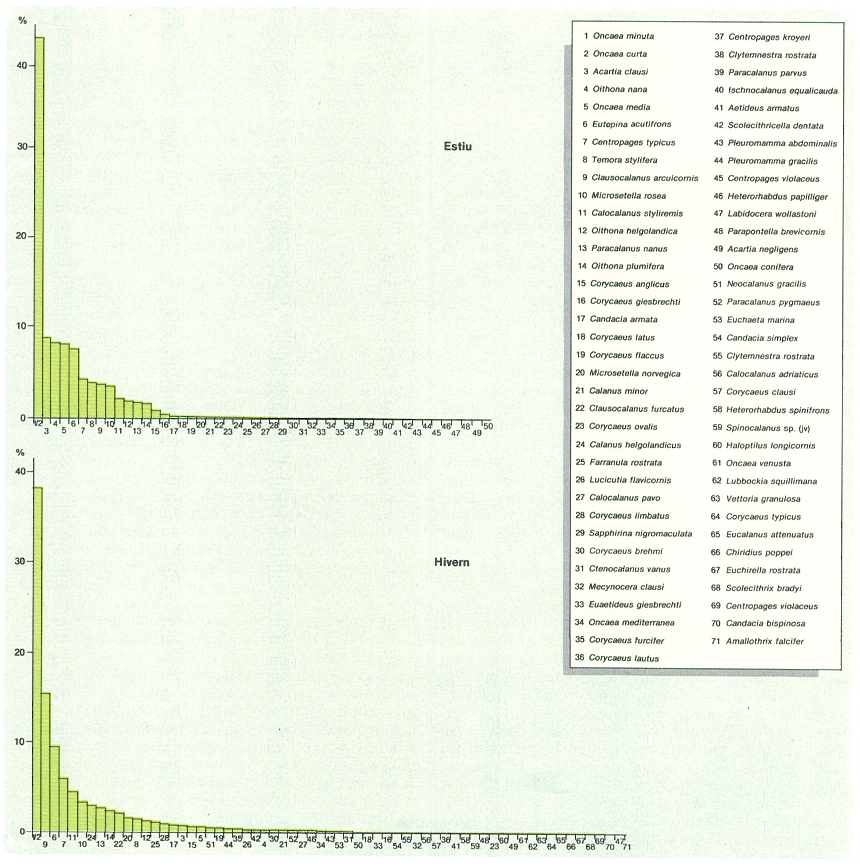
Composició relativa del plàncton de copèpodes i representació de les diverses espècies en les aigües costaneres de Catalunya i el País Valencià. La barreja vertical és la causa de la diversitat més gran de les poblacions en l’època freda.
Carto-Tec, original de F. Vives.
Els copèpodes són els crustacis més importants en el plàncton. Representen, a la mar, el que els insectes representen a terra, i el seu nombre d’individus és immens. En una gran part de l’extensió marina, en els primers 200 m d’aigua se’n poden comptar uns quants milers per metre quadrat. Cada copèpode sol contenir entre 6 i 50 µg de carboni orgànic. En el plàncton marí es troben representats calanoides, ciclopoides i harpacticoides, en aquest ordre de freqüència descendent, i és notable que fins i tot els harpacticoides, profundament transformats per a viure damunt d’un substrat, hagin donat formes pelàgiques que, precisament, es troben entre els elements més característics del nostre plàncton estival de superfície (Microsetella, etc.). Els més abundants són calanoides (Centropages typicus, Calanus helgolandicus, Paracalanus parvus, Clausocalanus arcuicornis, Temora stylifera, etc.) i alguns ciclopoides (Oithona, Corycaeus). Encara que petits, els copèpodes són organismes d’unes «performances» molt notables en tots els aspectes. El seu cos porta molts elements sensoris, gairebé comparables a la línia lateral dels peixos. El conjunt d’apèndixs al servei de la boca envolta la cavitat bucal a manera de cistella, amb una gran diversificació de funcions: alguns, simplement, llancen petits organismes contra uns filtres que els retenen i els porten a la boca —però que poden rebutjar-los si cal— i altres poden ensumar l’aliment a distància i agafar-lo amb manipulacions més complicades. En fi, hi ha animals de «cullera», que poden alimentarse amb relativa passivitat en aigües riques en fitoplàncton, i animals de «forquilla» que persegueixen i capturen altres animalets, sense oblidar una munió d’espècies paràsites, algunes, fins i tot, d’animals pelàgics. Normalment, un copèpode filtrador de mida mitjana clarifica al dia de 10 a 30 ml d’aigua. Una cinquantena de copèpodes en un litre d’aigua, densitat elevada però no impossible, repassen cada dia tot el volum. Es comprèn l’enorme influència que aquests organismes tenen en la dinàmica de tot el fitoplàncton. Els copèpodes tenen sexes separats, fecundació amb espermatòfors i diferenciacions en alguns apèndixs per a portar a terme la funció de retenir la femella i passar-li el paquet d’elements fecundants. En els ciclopoides, els ous es porten en petits sacs transparents penjats de l’abdomen de la femella. Els calanoides generalment els dispersen en l’aigua. De fet, l’hàbit de portar els ous fins que esclaten és molt més freqüent entre els copèpodes d’aigüa dolça que en els marins, i deu tenir alguna conseqüència en la demografia i en la genètica de les respectives poblacions. Si els copèpodes porten els ous en saquets es presten a un bon control de llurs postes, com en el cas dels ocells, i no és gens estrany que un treball molt suggeridor en el domini de les interaccions entre genètica i demografia, de Hutchinson (1951), portés el títol: «Copepodologia per als ornitòlegs».
Els copèpodes neixen en una forma larval molt diversa de l’adult i necessiten una dotzena de mudes per a adquirir la forma definitiva que, en línies generals, és un miler de vegades més grossa que l’ou o la primera larva. És, tanmateix, un canvi profund, comparable al de molts insectes, encara que és, en certa manera, més gradual; però l’augment progressiu de les dimensions i la complicació dels apèndixs bucals fan que del naupli a l’adult, passant per les fases que s’anomenen de copepodit, els copèpodes ingereixin aliments bastant diversos. Una espècie de copèpode, en conseqüència, equival a moltes espècies segons la forma amb què actua sobre l’aliment del seu entorn. Les femelles dels copèpodes deixen els ous en el punt culminant de la migració diària, de manera que els nauplis acostumen a trobar-se en aigües superficials riques en fitoplàncton menut i, després, van descendint progressivament. Fet i fet, la renovació mitjana de les poblacions de copèpodes demana entre 10 i 100 dies, encara que la vida individual màxima pot ser considerablement més llarga. Una secreció del tub digestiu dels copèpodes queda envoltant els excrements com si fossin un embotit. Aquests paquets o pilotetes fecals són considerablement compactades i, per això, se sedimenten ràpidament a una velocitat de 40 a 400 metres per dia. Contenen bacteris que, finalment, desfan la pel·lícula orgànica que envolta el paquet fecal, com una bomba de temps, després d’un cert nombre d’hores. Durant aquest temps, el material dels excrements ha pogut salvar ràpidament una distància considerable. Aquest dispositiu és una de les fonts d’alimentació més importants en aigües profundes. Sembla que tampoc no és menyspreada per membres de la mateixa confraria, és a dir, per altres copèpodes de fondària, caracteritzats pel fet de tenir el tub digestiu anormalment llarg, com si fos adaptat a treure tot el suc d’una menja evidentment molt pobra. L’excedent s’acumula en els copèpodes en forma de lípids, que no tenen la composició habitual en altres animals, sinó que són ceres.També es troben en el plàncton representants d’altres grups de crustacis, que poden arribar a ser molt importants. Els amfípodes pelàgics són sovint fantàstics i bons exemples d’al·ometria o transformació, segons algunes regles senzilles, d’unes formes en unes altres, cosa que és molt suggeridora des del punt de vista d’interpretacions evolutives. A les nostres aigües són molt notables les formes que viuen associades amb plàncton gelatinós, sovint instal·lades dintre el mantell dels tunicats que s’han menjat (Phronima, Vibilia). Molt de passada cal recordar alguns isòpodes paràsits de peixos; els cumacis, que solen viure al fons i de nit poden pujar a formar part d’un meroplàncton rítmic; els misidacis, més aviat associats al litoral, però amb alguns més planctònics i fins batipelàgics; i, sobretot, els eufausiacis, que representen un grup molt important, amb una problemàtica paral·lela a la dels copèpodes, però construïts a una escala més grossa. D’eufausiacis, també n’hi ha formes més aviat filtradores (Euphausia) i altres de més carnívores (Thyssanoessa, Meganyctiphanes). Són l’anomenat «kriil», o aliment preferit de les balenes, i es troben en grans quantitats en aigües fredes; però no manquen a la Mediterrània, en eixams que es congreguen on l’aliment és més abundant, i constitueixen una part molt important de l’aliment dels peixos. Tenen fins a 2 o 3 cm i la seva agilitat fa que no sempre estiguin ben representats a les pesques. Acumulen diatomees o altres aliments entre les potes toràciques, que porten pèls llargs i fan una mena de cistell en les espècies d’aquest règim alimentari. Entre les seves espècies, es molt general la producció de llum. El seu lloc de densificació són les termoclines i, en general, totes les ergoclines o fronts. Finalment, cal esmentar que hi ha un nombre considerable de decàpodes nedadors pelàgics, que corresponen al tipus «gamba», amb una sèrie de gèneres representatius a la nostra mar, com ara Aristeomorpha, Solenocera, Sergestes, Lucifer.
El balanç de la vida pelàgica
Per a entendre el funcionament de l’ecosistema pelàgic, la primera condició és la de quadrar una mena de balanç, per veure si l’excedent representat per la producció neta del fitoplàncton es distribueix a bacteris i animals, tenint en compte les necessitats metabòliques d’aquests heteròtrofs i llur densitat i distribució. Obviament, hi ha d’haver encara un excedent net, destinat a explicar la persistència de la vida en els fons on no arriba llum i, també, en proporció variable, per a explicar l’exportació de matèria orgànica dels sistemes pelàgics als sistemes bentònics costaners. A més, tot això ha d’estar d’acord amb el consum d’oxigen a les capes profundes i amb l’alliberament paral·lel de nutrients que s’acumulen en certs nivells o són reutilitzats segons les possibilitats de transport. Una manifestació d’aquesta dinàmica són les relatives biomasses, fluxos d’energia i velocitat mitjana de renovació per a cadascun dels components principals del món pelàgic.
Una fracció del material sintetitzat pel fitoplàncton passa a l’entorn com a matèria orgànica dissolta, que es va transformant i degradant progressivament, per l’activitat de bacteris, però el resultat és que tant la concentració del dit material orgànic com la dels bacteris que l’utilitzen va disminuint a mesura que ens allunyem de l’origen del material, és a dir, de la superfície de la mar. Com que la Mediterrània no és gaire productiva, aquest consum gradual utilitza només una fracció relativament petita de l’oxigen present a l’aigua.
A continuació, la taula presenta valors orientatius de biomassa, producció i temps mitjà de renovació per al plàncton i el nècton de la Mediterrània occidental. Atesa la migradesa de la informació quantitativa, cal remarcar que les xifres es donen només a tall d’orientació.
Biomassa, producció i temps mitjà de renovació del plàncton i el nècton
| ORGANISMES | BIOMASSA (g C/m2) |
PRODUCCIÓ (g C/m2 ∙ any) |
TEMPS MITJÀ DE RENOVACIÓ (dies) |
| Fitoplàncton | 1 | 70 | 5 |
| Bacteris | 0,6 | 35 | 2 |
| Zooplàncton | 5 | 10 | 150 |
| Nècton (peixos principalment) | 0,7 | 1 | 200 |
| Dades elaborades per l’autor a partir de fonts diverses |
Migració vertical i biofotogènesi
Ecograma que mostra la posició de superfícies reflectants formades per l’acumulació d’organismes planctònics. Seguintles durant un temps es poden detectar moviments d’ascensió i descens que s’integren dins del fenomen general de migració vertical.
Jordi Vidal; font: Institut de Ciències del Mar
L’adaptació del zooplàncton a aquest fons de fitoplàncton i bacteris comporta una peculiaritat dels animals pelàgics que consisteix en la seva propensió a situar-se a nivells més propers a la superfície durant la nit i a moure’s cap avall durant el dia. És un fet conegut, des de fa més d’un segle, que les pesques de zooplàncton prop de la superfície són més fructíferes durant la nit. La migració afecta, en grau divers, la major part dels components del plàncton, però més els crustacis (eufausiacis i copèpodes) que no pas altres. I afecta de manera paral·lela habitants de diversos nivells: per exemple, hi pot haver una migració entre la superfície i, posem, 250 m per a molts copèpodes pobladors dels estrats relativament superiors, i una altra migració, que afecta altres crustacis i peixos batipelàgics, entre uns 500 i uns 150 m de fondària. Es notable que la migració afecti també espècies dintre d’una zona on no hi ha pràcticament llum, de manera que la llum no és l’inductor principal del comportament. A més, els organismes migradors comencen a pujar abans de la posta de sol i inicien la davallada molt abans de clarejar. Sembla, més aviat, un ritme intern en sincronia amb el cicle diari, però la capacitat de reacció és molt gran. De fet, s’han aprofitat eclipsis totals de sol per veure la reacció del zooplàncton i els resultats satisfan totes les opinions; en amagar-se el sol i minvar la llum, unes espècies reaccionaven com en fer-se fosc, i unes altres «no es deixaven enganyar».
La migració vertical és òbvia a totes les mars i, per tant, a la Mediterrània, però no tenim estudis prou profunds per a precisar-la, o per a precisar les peculiaritats que el fenomen pugui revestir a la nostra mar. Però, com que la Mediterrània és una mar poc productiva, la densitat de les poblacions pelàgiques profundes (batipelàgiques) és massa baixa perquè es pugui seguir còmodament la propagació de les ones de migració cap a nivells profunds.
El mecanisme o els mecanismes fisiològies directes de la migració són complexos: no és una reacció senzilla a la llum, sinó que sovint hi deu haver un ritme intern que es posa a l’hora d’acord amb un sistema complex de percepció de l’ambient. En bona teoria evolutiva, l’adquisició i el manteniment del comportament migratori, que no és gratuït, demana una contrapartida, una utilitat, que hagi tingut com a conseqüència la selecció dels mecanismes fisiològics adients. S’ha dit que el zooplàncton podia escapar-se durant el dia de les aigües il·luminades, on els peixos podien veure’l i menjar-se’l més fàcilment. Segons Hardy, la migració vertical dóna al zooplàncton unes «botes de set llegües» si, com passa sovint, les aigües de superfície es mouen amb relació a les de fons sobre un pla horitzontal. El plàncton que puja i baixa pot explorar i explotar successivament, sense gaire esforç, taques o núvols de fitoplàncton que poden ser disperses, en el sentit que els animals poden romandre més temps on les condicions d’alimentació són millors. Cal remarcar, a més, que la senzilla activitat filtradora de molts crustacis pot portar al seu moviment ascensional, de manera que pujarien quan mengen (nit) i baixarien quan paren de «treballar». Ara bé, les cèl·lules del fitoplàncton tenen una càrrega elèctrica negativa, que és més forta si assimilen activament; és més difícil de separar, acumular i ingerir les partícules més carregades que les de menys càrrega, de manera que els copèpodes poden filtrar amb més rendiment si les algues no assimilen, és a dir, a les fosques; una altra raó per a explicar la migració. Algun autor ha fet notar que, tanmateix, el fet de passar una part de la vida en aigua profunda, que és més freda, pot contribuir a disminuir el metabolisme i la despesa d’aliment, raó que pot tenir una part de veritat, però que no és totalment convincent, perquè sotmetre un organisme a temperatures fluctuants —pujant i baixant entre uns estrats superiors més calents i uns altres inferiors, més freds— més aviat fa augmentar el metabolisme. Com és natural, en aigües més o menys agitades o de moviments irregulars, com ara a la part compresa entre la costa i el front principal, la migració vertical mai no és tan clara.

Migració vertical del zooplàncton expressada segons les mesures de la quantitat de nitrogen (un copèpode mitjà pesa uns quants micrograms, i conté uns quants micrograms de N) entre Barcelona i el front catalano-balear, al juliol de 1982. Aquest exemple de migració té la particularitat que, a causa de la gran fondària a què es troba el plàncton vegetal, la coincidència més gran entre zooplàncton i fitoplàncton es dóna de dia i no de nit, com és corrent.
Carto-Tec, original de M. Alcaraz.
D’acord amb una interpretació tròfica del reialme pelàgic, cal relacionar la migració amb dos fets: els animals mengen preferentment a dalt i excreten preferentment a baix; i, tant els copèpodes com els eufausiacis produeixen excrements compactes, que adquireixen una gran velocitat de sedimentació, almenys durant les primeres hores (desenes de metres per hora). El resultat és que hi ha una acceleració del transport de dalt a baix de la producció primària, que té diferents conseqüències per a la vida pelàgica. És clar que això acumula més ràpidament materials en fondària i retarda el cicle de producció. La migració vertical fa el sistema més oligotròfic. Altrament, distribueix de manera més equitativa, en fondària i horitzontalment, el material produït en superfície pel fitoplàncton. Sense la migració vertical, encara arribaria menys aliment del que arriba a les grans fondàries marines. Aquesta funció igualitzadora i d’estabilització solament té sentit a escala de tot l’ecosistema i, com és evident, no és utilitzable com una raó per explicar l’adquisició de la forma de comportament que comentem.
Si la dimensió vertical, segons la llum i la gravetat, dóna una organització més o menys elàstica o dinàmica que es pot reconèixer en l’estratificació de diferents espècies segons les fondàries i també en la migració vertical, la distribució del zooplàncton segons el pla horitzontal és molt més aleatòria. És evident que hi ha diferenciacions i acumulació de plàncton animal en certs indrets, com és ara al llarg del nostre front, o bé, d’una manera més transitòria, en relació amb cèl·lules de Langmuir o altres estructures comparables, però, en general, no es pot parlar més que d’heterogeneïtat horitzontal, manifestada en tots els estudis estadístics una mica acurats que impliquin la comparació de mostres agafades en diferents punts. Com que aquesta heterogeneïtat es veu com una distribució de contagi, en el sentit que, entorn dels llocs on les densitats de plàncton són altes és probable trobar-ne d’altres també elevades, els planctòlegs parlen contínuament de «patchiness», o de taques o bancs, potser pensant en els peixos, però sense voler dir res de particularment clar.
Evidentment, si pensem que el plàncton té una estructura vertical molt ben definida —cada columna d’aigua es podria considerar com una corda tensa i ben temprada— el conjunt d’aquestes columnes ideals paral·leles és recorregut per ones horitzontals de diferent importància i direcció, a les diverses fondàries, especialment en aquelles on hi ha major discontinuïtat física, com ara a les termoclines. La major difusió horitzontal de materials a les termoclines, materials que poden ser atractius per a altres individus, i el fet que les mateixes capes de discontinuïtat tèrmica i de densitat constitueixin bons canals per a les ones sonores i ultrasonores, les converteixen en superfícies preferents de comunicació i acumulació per als animals. Sembla, efectivament, que la sardina forma bancs quan hi ha termoclina, i precisament a la termoclina, com a major referència.
Organismes lluminosos en el plàncton i llur detecció. A l'esquerra, una instal·lació emprada en el «Mechelen», en el golf de Cadis. A la dreta, freqüència amb què s’observen guspires de llum en el mesurador a través del qual flueix aigua amb plàncton. Noteu que els organismes lluminosos (en groc) s’observen solament de nit, sigui perquè de dia estan més avall, sigui com a conseqüència del ritme endogen que limita la seva lluminositat a les hores de fosca, la qual cosa és ben comprovada.
Ramon Margalef; Ricardo Génova, original de l’autor
Molts organismes lluminosos formen part de l’ecosistema pelàgic. Els més coneguts són unicel·lulars, com els dinoflagel·lats Noctiluca, Pyrocystis, Gonyaulax i Ceratium, o bé animals com ara meduses, ctenòfors, poliquets, cefalòpodes, crustacis, tunicats i peixos. Òbviament, hi ha bacteris lluminosos —perquè la producció de llum és una funció biològica primordial, que potser té el significat de desempallegar la cèl·lula d’un excés d’oxigen— i, de fet, si molts animals fan llum és perquè porten bacteris lluminosos associats, en una simbiosi molt íntima. En altres animals, els elements lluminosos consisteixen en bacteris molt transformats.
Els organismes eucariotes han trobat noves funcions a la producció de llum: en els unicel·lulars s’ha dit que esvera els possibles depredadors i en els crustacis, els cefalòpodes i els peixos pot atreure preses, servir de reconeixement per a interacció social o sexual i, fins i tot, dissimular la presència del peix. S’ha dit que peixos amb grans plaques lluminoses ventrals, com els Argyropelecus i Sternoptyx, que migren verticalment i que de nit s’apropen fins a un centenar de metres de la superfície, resulten menys visibles des de baix per la seva mateixa lluminositat. Hom pensa en els edificis amb les finestres il·luminades vistos sobre un cel de crepuscle: l’edifici sembla calat i perd els senyals normals d’identificació; una cosa semblant pot succeir amb els peixos. La llum sempre és blanca verdosa, encara que és modificable per cromatòfors acolorits que s’estenen, en alguns casos, davant de l’òrgan productor de llum que, si és diferenciat, té gairebé l’estructura d’un ull amb funció invertida (envia llum i no en rep).
Entre els dinoflagel·lats, hi ha espècies congèneres que es diferencien en la producció de llum: de dos Ceratium molt vulgars, C. fusus fa llum i C. furca no en produeix. La llum s’estimula per factors químics o mecànics —agitació— i sovint mostra un ritme endogen: dinoflagel·lats de les aigües superficials produeixen més llum de nit que de dia, i aquest ritme persisteix en el laboratori com un rellotge, almenys durant uns dies, i en contra de possibles diferències d’il·luminació. Molts peixos lluminosos, especialment els que tenen poques llanternes, generalment a les galtes, a la llengua o en algun apèndix a manera de canya de pescar com si fos l’esquer (i realment sembla que fa aquesta funció) ho deuen a l’associació amb bacteris. Els bacteris són més integrats i han esdevingut orgànuls del peix en aquelles nombroses famílies que tenen fotòfors o òrgans lluminosos en quantitat i ordinàriament en dues sèries laterals inferiors, com ara estomiatoides i mictòfides, que són peixos batipelàgics i mesopelàgics, generalment petits i d’allò més freqüents. Amb sensors adients es pot registrar la llum d’origen animal i, d’aquesta manera, obtenir perfils verticals de la seva distribució. Així es detecta la migració vertical d’aquests animals i s’afegeix una altra evidència a la que dóna el sonar.
Biogeografia del plàncton
Els animals del plàncton tenen molta més facilitat per a moure’s que els animals marins de fons o que els animals terrestres. Això comporta un bescanvi de gens, i la mateixa motilitat de les aigües marines pot portar a una contínua reestructuració de les poblacions. La tendència a la formació d’espècies i la segregació geogràfica són mínimes en el món pelàgic. Evidentment, hi ha diferències entre les àrees d’aflorament i les àrees on predomina plàncton gelatinós, o entre les regions del centre dels grans remolins anticiclònics oceànics i les regions fredes. A aquesta mena de diferències s’han d’afegir les que resulten del meroplàncton, el plàncton passatger unit a cada costa i que específicament mostra una diferenciació geogràfica més gran de l’ecosistema bentònic.
S’han publicat molts treballs i s’ha intentat de sistematitzar la distribució geogràfica del plàncton marí per distingir-hi diversos elements. Pel que fa a la nostra mar, és clar que la Mediterrània ha rebut el seu poblament actual de l’Atlàntic, després de la crisi messiniana. Es parla, per tant, d’una regió biogeogràfica marina atlàntico-mediterrània. Per la seva posició, la Mediterrània, en temps antics, pogué rebre elements d’afinitat més índics i, fins i tot, pacífics, però foren extingits i, si actualment hi ha espècies que mostren aquestes afinitats, com a llevant (Israel) o al llarg de les costes africanes, es tracta d’espècies d’àmplia distribució, sense excloure la possibilitat d’un pas recent a través del canal de Suez (que ara és més travessable que abans, quan els llacs salats interposats eren de salinitat encara no diluïda), o altres introduccions degudes més directament a la mà de l’home, ben evidents en alguns crustacis i mol·luscs, i potser en l’únic corall hermatípic de la nostra mar.
Sovint es pensa que la introducció d’elements procedents de l’Atlàntic no fou contínua. És evident que les fluctuacions glacials afavoriren alternativament espècies que demanen temperatures diverses; algunes s’extingiren certament més tard. Espècies d’afinitats òbviament nòrdiques, com Nephrops, alguns Gadus, i l’eufausiaci pelàgic Meganyctiphanes norvegica —molt important actualment a la nostra mar— pogueren expandir-se especialment en èpoques fredes, i avui són més o menys confinades a les capes profundes, on l’aigua és més freda. Ampliant aquests punts de vista, hom podria associar nombroses introduccions més recents amb el moviment de l’aigua superficial de l’Atlàntic, i el mecanisme d’aquestes entrades quedaria confirmat amb la presència accidental d’espècies que normalment ja no s’hi queden. És en aquest sentit que hom ha parlat sovint d’indicadors d’aigües atlàntiques a la Mediterrània i, de fet, s’han publicat llistes de copèpodes i d’altres animals que tindrien aquest significat. Es podrien utilitzar, eventualment, per descobrir si alguns anys les aigües de l’Atlàntic avancen més endins sense barrejar-se, o ho fan en remolins o altres formes discontínues.
Aquest sentit d’organismes indicadors sols el tenen si no persisteixen, si la seva presència correspon a una àrea d’expatriació, on les poblacions no es mantenen ja indefinidament per reproducció pròpia; de vegades poden ser relíquies de distribucions passades. En un altre sentit, són indicadores les espècies més o menys ubiqües i de ràpida multiplicació que apareixen en quantitat allà on troben condicions particularment favorables. L’estudi taxonòmic del plàncton, ultra distingir combinacions particulars d’espècies i llur distribució vertical i horitzontal, ha de ser receptiu a la possible presència d’indicadors de les dues menes.
El nècton: peixos i cefalòpodes
Els peixos típicament nectònics no comprenen gaires espècies (potser menys de 400 en tot el món), pertanyen a moltes famílies força diverses, acostumen a ser de distribució geogràfica àmplia i són més aviat d’aigües pobres, on hi ha menys competència d’una fauna litoral o costanera en constant tendència a l’expansió. Aquest grup nectònic comprèn peixos voladors, com l’oranyola (Exocoetus volitans), l’oreneta (Hirundichthys rondelettii) i els diversos voladors del gènere Cheilopogon, els grans túnids («tonyines») dels gèneres Auxis, Euthynnus, Sarda i Thunnus, el peix espasa (Xiphias gladius), el marlí (Tetrapterus belone), l’emperador (Luvaris imperialis), les llampugues (Coryphaena), la sèrie dels verats (Scomber) i sorells (Trachurus, Caranx) de dimensions ja menors, uns quants peixos de forma curiosa, com el bot (Mola mola), el rei (Regalecus glesne) i els pocs taurons pelàgics i planctòfags com el pelegrí (Cetorhinus maximus), peixos dependents d’altres, com el peix ventosa (Echeneis naucrates), o el pilot (Naucrates ductor), sense oblidar els joves sorells, que sovint s’associen amb celenteris pelàgics.
En general, els peixos més abundants a les aigües dels estrats superiors s’associen a àrees molt productives, no necessàriament costaneres, com la sardina (Sardina pilchardus), sinó més enfora, com l’alatxa (Sardinella aurita) o el seitó (Engraulis encrasicolus). Aquests peixos són petits, de vida curta i de forta reproducció, de manera que llurs poblacions fluctuen ràpidament seguint els canvis ambientals. Aquesta és llur estratègia de supervivència. Encara es pot reconèixer una diferència important, manifesta, per exemple comparant les poblacions d’ací amb les d’aigües atlàntiques, on la producció mitjana és més alta i les fluctuacions relativament majors. Doncs bé, la vida d’aquestes espècies petites és relativament més llarga ací i, potser, les fluctuacions de producció menys marcades, encara que ben perceptibles. Precisament, les fluctuacions de sardina i seitó, tal com s’aprecia en els desembarcaments dels pescadors, mostren una certa alternació i, per a cada espècie, uns màxims separats per uns quants anys de distància (de 5 a 9) fan pensar que llurs poblacions reflecteixen el funcionament del sistema pelàgic enter. Hi ha ací una gradual transició d’uns dominis de la biologia marina a uns altres. La sardina, per exemple, és força relacionada per la seva posta i per l’èxit de la supervivència dels joves a les condicions heterogènies del medi pelàgic, tal com es donen prop de la desembocadura de rius. La sardina, que just abans formava bancs més o menys grossos, es dispersa vers el fons quan la termoclina es trenca a la tardor. Poc després, s’inicia la preparació de la posta, que es fa durant uns quants mesos, freds, i prop de les aigües heterogènies influïdes per l’aiguabarreig dels rius. En altres clupèids, com ara l’amploia (Sprattus sprattus) i, naturalment, les sabogues i guerxes (Alosa), l’atracció dels individus que van a reproduir-se per les aigües dessalades és molt més evident. El seitó, en canvi, pon a l’estiu en aigües estratificades i a la superfície i, generalment, més lluny de la costa. És natural que les fluctuacions hidrogràfiques del període mitjà, com la del Niño, o més o menys septennals, puguin influir en la partició entre la sardina i el seitó dels recursos tròfics de la mar. Molts peixos són més o menys meroplanctònics, en el sentit que els joves es crien a una distància més gran de la costa, a base de recursos del món pelàgic, i més tard, o bé són peixos bentònics o de fons, o fan vida semipelàgica, és a dir, a les aigües costaneres, però sense integrar-se pròpiament mai en la comunitat de l’aigua blava.
Altres peixos pelàgics, especialment els que neden bé i fan llargues migracions horitzontals, com les tonyines, van d’una banda a una altra i, com a nòmades, van explotant recursos locals. Resulta molt interessant de comparar les dues estratègies, la dels peixos petits de poblacions fluctuants lligats a l’explotació de recursos locals, i la dels peixos grossos, de vida llarga, que en llurs migracions aprofiten la producció d’extenses àrees marines i poden combinar regions allunyades, en el sentit que si els en falla una un any, en poden explotar una altra. Aquestes espècies es poden comparar als ocells que migren; els que freqüenten les maresmes acudeixen de seguida a la ment de l’oceanògraf, com a terme de comparació. És clar que les condicions que ofereixen a l’explotació humana són diverses. Les espècies fluctuants i de vida curta permeten una intensa explotació, i són sotmeses a fluctuacions que l’home pot agreujar, però que no pot modificar substancialment. En canvi, els peixos grossos i migradors, de vida llarga, tenen menys enemics a la mar i acusen d’una manera alarmant la pressió de la pesca que l’home exerceix, generalment en forma de palangres i també —en temps passats— amb xarxes fixes com les que s’instal·laven, no fa encara gaires anys, a l’Hospitalet de l’Infant. En una situació intermèdia tenim els verats i els sorells; en canvi, la resta dels peixos veritablement pelàgics, a les regions oligotròfiques d’aigua blava, tenen molt poca importància comercial, llevat potser de la llampuga i el peix espasa, que es troben, especialment, al centre de la conca occidental de la Mediterrània, vers les illes, tant les Balears, com Sicília i altres.
Els peixos pelàgics que mengen plàncton o animals petits colen l’aigua que entra per la boca a través d’unes pintes formades per barnilles. Aquests dispositius foren adquirits secundàriament i separadament al llarg de l’evolució, ja que els peixos planctòfags són formes aparentment derivades de formes primitives que sembla que eren més aviat de fons. També el pelegrí (Cetorhinus), de fins 12 m, és filtrador i hom el troba flotant mandrosament prop de la superfície. Els peixos carnívors i depredadors tenen formes irreprotxables des del punt de vista de la ciència hidrodinàmica: cos llis, més arrodonit pel davant, més afuat pel darrere, sovint amb carenes laterals, i amb les aletes dissenyades per reduir al mínim les pertorbacions i la turbulència que el moviment comporta o per treure’n el màxim de profit. La velocitat de les tonyines pot ser de fins 20 longituds del cos per segon, el seu pes pot arribar a 730 kg, la concentració d’hemoglobina en la sang és com la de l’home i tenen temperatura elevada, uns 10 °C per damunt de l’aigua circumdant, generada en les masses de teixit muscular roig, que s’insereix com un tascó entre els músculs de carn més blanca. La pèrdua de calor, que podria ser molt elevada a través de les brànquies, es minimitza per l’acció d’un sistema de contracorrent (la sang que va a un òrgan perifèric cedeix calor a la sang que en torna, en uns òrgans vascularitzats anomenats rete mirabile, i, certament, es tracta d’un dispositiu admirable). Els pescadors han sabut des de sempre que les tonyines tenen la sang relativament calenta, però els savis sembla que en tingueren la primera notícia amb les observacions de John Davy, fetes a la Mediterrània el 1835. Després se n’oblidaren fins fa unes dècades, que, volent instal·lar registradors de temperatura damunt de tonyines per reconèixer, mitjançant els registres, les aigües per on passaven en les seves migracions, s’adonaren de la interferència causada per la temperatura pròpia dels peixos. L’interès econòmic —i científic— de les tonyines ha fet que s’aboquessin bastants recursos a la seva investigació. S’han marcat molts exemplars (generalment amb una mena de fideu de plàstic —dintre el qual hi ha el missatge— implantat en una aleta dorsal), cosa que ha permès de reconèixer l’amplitud oceànica dels seus viatges, i és sorprenent que, a vegades, semblen escollir per criar llocs de producció baixa, tal com s’esdevé amb la tonyina vera o roja (Thunnus thynnus) a la Mediterrània, cap al SE de la península. Les migracions horitzontals dels peixos són de període anual, mentre que les migracions verticals, àdhuc les dels peixos pelàgics, són de període diari.
La densitat dels teixits dels peixos pelàgics ultrapassa la de l’aigua de mar, de manera que la veixiga natatòria és un òrgan benvingut per a equilibrar el peix i disminuir una mica l’esforç necessari. Però la veixiga natatòria resulta molt identificable per als animals —principalment els cetacis— que usen òrgans de detecció basats en el mateix principi del nostre sonar, és a dir, per mitjà del ressò o el retorn d’infrasons dirigits. Per això, el benefici en el sentit de facilitar la flotació és contraposat per la facilitat de detecció. L’evolució de la bufeta ha estat sotmesa a una pressió contradictòria de selecció. De joves tots en tenen; més tard es pot conservar en animals grossos que no tenen res o poc a témer, com ara en diversos Thunnus; en canvi, els verats i altres espècies de tonyines —en sentit ampli— la perden.
D’altra banda, el peix espasa (Xiphias) és bastant diferent de les tonyines i els marlins; té menys massa relativa de teixit muscular roig, però disposa d’un sistema de calefacció especial per al cervell, i és un animal que, segons sembla, caça especialment a l’aguait i, sovint, a gran fondària. La seva espasa és realment el suport d’un equip força complex d’òrgans perceptors d’ones de pressió a l’aigua, per a la detecció de preses.
Cal afegir que hi ha un nombre considerable de calamars que formen part del nècton d’aigües superficials, (Ommatostrephes, Loligo), on es mouen a reacció, per projecció d’un raig d’aigua a través de l’embut, amb la contracció de la cambra pal·leal; així, poden ser projectats obliquament a través de la superfície de la mar, talment com peixos voladors, a alçades 10 vegades la llargada de l’animal i a distàncies longitudinals cinc vegades més grans (50 vegades la llargada de l’animal). També hi ha altres cefalòpodes més petits i menys mòbils que gairebé «són» plàncton, com ara els famosos Argonauta les femelles dels quals tenen una elegant closca separada, on deixen els ous, o els encara més estranys Ocythoe, un parent de l’argonauta el mascle del qual es fa una caseta amb un mantell de salpa, igual que la de Phronima. Les espècies batipelàgiques, que viuen a centenars de metres de fondària, són sovint gelatinoses i probablement menys freqüents a la Mediterrània que en altres mars. Els «becs» dels cefalòpodes es poden identificar específicament i, com que són molt duradors, es reconeixen en el contingut digestiu dels depredadors i també en el sediment, amb les conseqüents implicacions per a la recerca.
El món dels peixos pelàgics de les aigües superiors és duplicat, en fondària, per una fauna paral·lela, constituïda per peixos poc corpulents, d’esquelet i teixits febles, sovint de pell negrosa i gairebé sempre amb fotòfors o òrgans lluminosos. Rarament són cecs; si hi ha algun peix cec, més aviat és de fons, o és litoral. Però els ulls dels peixos que viuen a gran fondària poden estar transformats: de vegades, són brillants, com ara els de Chlorophthalmus (ulls verds), que tenen un tapetum lucidum reflectant com el dels gats; altres vegades són sortints i molt miops, malgrat que reben la denominació, certament inadequada, d’ulls telescòpics, com els d’Argyropelecus. Són animalets petits: la majoria fan entre 1 i 10 cm. Els òrgans lluminosos són freqüents, generalment en forma de fileres de puntets brillants, en algun cas amb l’aspecte de plaques o finestres majors, com en Scopelus o Argyropelecus. Quina n’és la funció? Atreure preses? Permetre el reconeixement d’espècies i sexes? Això és probable, perquè la disposició és característica, i algunes espècies els poden usar per a fer senyals (apagant-los, per exemple). Aquests peixos es poden moure entre nivells diferents, en un àmbit entre 200 i 800 metres, cosa que permet de fer molta maniobra, i de nit arriben fins més amunt. Ja sia per als seus moviments, ja sia per als dels seus depredadors, el fet és que aquests peixos lluminosos formen una fracció no negligible de l’aliment dels túnids epipelàgics.
Els peixos mesopelàgics de la Mediterrània han estat estudiats en diversos indrets. A les nostres costes, s’han fet diversos creuers de recerca, i d’altres s’han estès per tota la Mediterrània. El nombre d’espècies és molt més gran que el d’espècies epipelàgiques i hi són ben representades les famílies dels gonostomàtids (Cyclothone, Ichthyococcus, Maurolicus, Vinciguerria), esternoptíquids (Argyropelecus), cauliodòntids (Chauliodus), estomiàtids (Stomias), evermannèl·lids (Evermannella), mictòfids, una família molt important (Diaphus, Electrona, Hygophum, Lampanyctus, Lobianchia, Myctophum, Notoscopelus, alguns d’ells coneguts genèricament pels pescadors amb el nom d’anxova de fons). La taxa de renovació d’aquesta biomassa és relativament lenta, de manera que no es pot pensar en una explotació comercial d’aquests animals, ultra les dificultats tècniques que presentaria. Són poblacions molt diferents de les de la sardina o del seitó, que són transportadors eficacíssims d’energia transformada. Els peixos mesopelàgics es reprodueixen preferentment en llocs més rics, com ara a prop de costes profundes i, segurament, a nivell del nostre front mediterrani.
Certament, no queda esgotada amb això la fauna pelàgica de peixos, però aquesta visió sintètica pot complementar el seu tractament taxonòmic d’aquesta mateixa obra. Quant a llur biomassa i a llur producció, cal dir, francament, que la informació de què disposem és molt inferior a la desitjable i, sovint, més reduïda que la que tenim sobre fitoplàncton i zooplàncton. De fet, els valors que es donen de la biomassa i la producció de peixos se solen basar en les estimacions d’aquells esglaons o anells que els precedeixen en la cadena tròfica marina. Però no tots els peixos graviten sobre el plàncton. Hi ha la dita «el peix gros es menja el xic» i, de fet, hom pot reconèixer relacions d’aquest tipus entre diversos grups de peixos. Potser, l’únic que cal dir és que la relació en pes entre el depredador i la presa acostuma a ser de 100 a 1 i, per tant, les corresponents relacions pel que fa a longitud, s’apropen a 4,6. Per exemple, la sèrie de longituds 100 - 21,3 - 4,6 -(1)- 0,2 cm, correspon a la tonyina, el verat, la sardina i, saltant-nos un terme (perquè la sardina és filtradora), el copèpode, que fa entre 1 i 2 mm. Aquest exemple, que podria tenir algun punt discutible, s’insereix amb sentit pedagògic, per fer veure que la productivitat dels sistemes marins a escala de peixos grossos, que són els més apreciats, davalla ràpidament. Si el fitoplàncton produeix 100, el zooplàncton produeix 10, la sardina 1, i arribem a 0,01 quant a la tonyina. La producció de tonyina, o de lluç, representa el 0,01% de la producció dels vegetals, mentre que, per a una vaca, aquella proporció pot ser superior al 5%.
Si tractem d’integrar la informació existent sobre la pesca a les nostres aigües marines, reduint el producte de la pesca a carboni orgànic, i si el referim a la superfície on se suposa que s’ha generat el material orgànic que ha trobat el seu camí vers els peixos extrets, arribem a xifres aproximades als 30 kg de carboni orgànic per km2 de mar i per any. És clar que s’hi inclou també peix costaner, que gravita, en part, sobre sistemes bentònics litorals; però cal no oblidar que aquests sistemes reben una bona aportació o un subsidi del sistema pelàgic i que, en conjunt, es pot dir que el sistema pelàgic contribueix en un 95%, si més no, a la producció marina, sobre un model mundial. La producció primària de la nostra Mediterrània és propera a 80 t de carboni orgànic per km2 i per any (80 g C/m2∙any). La pesca extreta, doncs, si fa o no fa, és un 0,3‰ de la producció primària, valor que sembla baix. I no obstant això, no ho és, si es té en compte que l’extracció pesquera mundial és del mateix ordre —les estadístiques donen uns 70 milions de tones de peix fresc per any, suposant que siguin incompletes, augmentades a 100 milions de tones de peix fresc, que equivalen a 10 milions de tones de carboni orgànic, i dividides per l’extensió dels oceans, de 361 milions de km2, resulten a 1/36 g de C/m2, per a una producció primària mitjana més elevada que la de la Mediterrània (100 g C contra 60 g C/m2∙any)—. Això vol dir que els nostres pescadors s’ho enginyen bé; però, al mateix temps, que no hi ha esperances d’augmentar sensiblement el rendiment. També sobre una base mundial, la pesca total es distribueix de la manera següent: pesca en sistemes costaners i bentònics, 41%; pesca en sistemes d’aflorament, 52%; pesca pròpiament pelàgica (tonyines, calamars), 5%; marisc, etc., la resta.
La pesca en regions d’aflorament és majoritària, i s’hi inclouen les pesqueries d’anxoveta al Perú, que, per elles soles, fluctuen entre els 2 i els 14 milions de tones anyals de pes fresc. Al cap i a la fi, el millor indicador de la disponibilitat de peixos en una àrea és la densitat de vaixells de pesca, que, evidentment, es pot apreciar en fotografies fetes des d’una gran alçària. I és evident que una pressió d’explotació equilibrada vol disminuir el nombre d’embarcacions a mesura que aquestes esdevenen més poderoses, que és el que passa amb l’aplicació de les modernes tecnologies (motors més potents, xarxes més perfeccionades, sonar i altres mètodes avançats d’exploració i de navegació).
La consideració de les tècniques de pesca porta a la conclusió que l’home mateix no s’escapa de les diverses tècniques de captura que empra la natura: els animals filtradors o micròfags, els animals de «cullera» que agafen indiscriminadament aliment de mida petita, com ara molts copèpodes i els peixos amb branquispines, com la sardina, practiquen una tècnica de captura relativament indiscriminada, com els sardinals o les xarxes de ròssec. Les tècniques dels depredadors actius, o els que utilitzen formes específiques de comportament de llurs víctimes, es poden comparar a l’ús de nanses, de diverses formes de palangres i curricans, o a la persecució i la caça de cetacis.
Vertebrats superiors
No són els peixos els únics vertebrats de la regió pelàgica. Hi ha tortugues, no hi falten ocells marins i els cetacis hi tenen un paper important. Diverses espècies de tortugues marines tenen grosses escates: la tortuga babaua (Caretta caretta), que és la més freqüent, i Eretmochelys imbricata, Lepidochelys kempii i la tortuga verda (Chelonia midas), que es veu que són molt més rares. La tortuga llaüt (Dermochelys coriácea), que s’agafa de tant en tant, pot ser molt més grossa i la seva coberta amb relleus longitudinals consisteix en ossets immersos en una mena de cuir. Els ocells marins han estat ben estudiats en una altra part de l’obra, però ací ens plau de remarcar, en línies generals, l’existència de dos conjunts: els ocells planadors, de mars estratificades, que floten en l’aigua i mengen a la superfície, sovint deixalles (gavines, cues de jonc en països tropicals), i els ocells que es cabussen per pescar, com el mascarell (Sula), de llocs més productius, que agafen peix viu en fondària. Molts altres ocells marins es poden associar amb algun d’aquests tipus, i altres queden entremig, tremig, menjant a la superfície, i alguns menjant plàncton. La distinció és interessant perquè correspon a la del plàncton. El plàncton gelatinós d’aigües estratificades i caldejades correspon a corrents de convecció que permeten de planar. Les regions de fronts i d’aflorament, riques en plàncton i en peixos, porten a la superfície aigua relativament freda, que genera una circulació atmosfèrica descendent, on no es pot planar. Ací hi ha ocells que es cabussen per agafar llur presa. Encara no tenim prou dades relatives a l’observació d’ocells marins en alta mar; però, en tot el món, són sempre més abundants en les proximitats dels fronts.
Cal fer una curta referència als nostres cetacis, ja presentats en aquesta mateixa obra. La biomassa present de cetacis reposa sobre les disponibilitats d’aliment i, en darrer terme, de producció primària de l’àrea. Es pot dir que l’existència del front mediterrani assegura una producció contínua d’aliment que explica que, entorn del front, mai no siguin gaire rars els cetacis. Hi ha prou aliment, principalment peix i crustacis. També en el front, són particularment abundants i grossos els eufausiacis, germans dels que formen el «krill» en aigües més fredes de l’oceà mundial. Els cetacis han evolucionat a partir de mamífers carnívors i, bàsicament, continuen essent carnívors com els dofins i els catxalots; els catxalots mengen, per exemple, peix i grans cefalòpodes mesopelàgics. En un altre grup de cetacis l’evolució ha portat al desenvolupament d’uns òrgans absolutament nous, en forma de làmines desflocades de matèria còrnia que pengen del paladar i serveixen per a retenir el plàncton i el petit nècton quan expulsen les glopades d’aigua que agafen amb la boca oberta. És extraordinàriament interessant aquest canvi de règim, d’animal macròfag o «de forquilla» a animal mícròfag o de «cullera», ja que rarament es dóna al llarg de l’evolució. Això vol dir abreujar de cop la cadena tròfica, i si en un catxalot o en un dofí teníem una productivitat de 100:10:1:0,1 i fins a 0,01, en passar del fitoplàncton al final, en una balena tenim 100:10:1, més o menys com en la sardina, malgrat que es tracta d’un animal molt gros, però que menja eufausiacis, teròpodes o peixets, els quals mengen o poden menjar directament fitoplàncton. Però, tot avantatge en aquest sentit pot tenir l’inconvenient de les fortes fluctuacions en la producció que són característiques dels sistemes més productius. Això queda compensat per la capacitat de migració de les balenes que, pràcticament, integren la producció sobre àrees molt extenses. Però cal pensar que un sistema regular, com el de la mar Catalana, encara que no gaire productiu, podria mantenir permanentment algunes balenes. S’han fet força treballs sobre cetacis de la nostra mar, però és encara difícil estimar-ne la biomassa. Les nostres balenes, que poden pertànyer a diverses espècies, potser mengen peixos petits, seitons, etc., com a dieta preferida o que tenen més a l’abast. Des del punt de vista de l’explotació de la mar, potser la forma més eficient o econòmica d’extracció és obtenir la producció primària concentrada per les balenes. Pero caldria que passés un quant temps, deixar que es refessin unes poblacions importants i, aleshores, explotar-les amb prudència. I no es pot negar que matar sistemàticament animals tan sensitius i lliures ens produeix més repugnància que no pas abatre les víctimes habituals dels nostres escorxadors.
Senyals de les activitats de la vida i dels humans: el risc de les contaminacions

Encara que els mecanismes biològics en el món pelàgic de la Mediterrània no hagin canviat gaire sota l’acció de l’home, la constància i l’increment d’aquesta acció fan preveure danys ben aviat, que ja es manifesten més clarament en la proximitat de les costes. La pressió dels que en patim les conseqüències i la preocupació de les administracions han portat a prendre acords i crear organismes per millorar (la fotografia n’és un de tants testimonis), si es pot, la situació i garantir un futur acceptable. És obligació de tots que les propostes d’acció i de capteniment no quedin en paper mullat.
Jordi Vidal.
Avui dia, el problema de la contaminació preocupa, potser més en l’aspecte propagandístic que no pas com un convenciment general, reflectit en l’actuació tant dels ciutadans individualment com de les grans organitzacions, incloent-hi els estats. Però, de fet, s’ha arribat a diversos acords i s’han creat organitzacions amb l’objecte de limitar els possibles estralls d’una manipulació incorrecta dels recursos naturals que, en aquest cas, consisteix principalment en l’abocament de materials i de substàncies que tenen alguna influència negativa sobre la vida natural, o que afecten aspectes sanitaris o estètics als quals la humanitat és més sensible. Hi ha una sèrie de Convencions Internacionals aplicables a la Mediterrània, i potser val la pena d’esmentar-ne específicament la Convenció de Barcelona del 1976 per a la protecció de la mar Mediterrània. Les decisions explícites o implícites s’han pres en un grau divers i, sovint, poc satisfactori.
Es comprèn que la contaminació, la introducció de tota mena de materials, es fa més aviat prop de les costes, encara que mar enfora hi ha sempre el risc de pèrdues i d’abocament de materials —hidrocarburs— relacionats amb l’enorme tràfic de carburants. Cada any s’extreuen entre 3500 i 5000 milions de tones de petroli, i cada any creuen la Mediterrània uns 115 milions de tones. Sovint es diu que les pèrdues per rentatges de tancs i altres causes, són entre el 0,5 i 1’1%, i que la Mediterrània rep relativament força més petroli i materials semblants que els altres oceans. Això és cert, perquè és al centre d’una àrea molt transitada. La contaminació petroliera anyal és entre 500 000 i un milió de tones i correspon a 0,2 - 0,4 g de petroli per metre quadrat. És un valor modest, en comparació amb el cicle de la vida i, com que la major part del material es reintegra a un cicle biològic —del qual, d’altra banda, procedeix el petroli—, sembla que seria assimilable sense un mal major. Però els residus de petroli i de quitrà s’acumulen prop de la superfície, fan lleig i poden crear problemes locals. Els altres residus de l’activitat humana s’afegeixen més a prop de les costes i també poden quedar en segon terme dintre de la quantitat considerable de matèria orgànica que la mar rep normalment dels continents.
Les molècules sintetitzades per la vida, i les molècules orgàniques sintetitzades per la indústria, una vegada immerses en l’entorn natural, tenen una vida mitjana que pot ser llarga. A poc a poc, es van reciclant i descomponent. Però hi ha molècules recalcitrants a la descomposició, de més durada, i aquestes poden ser nocives si es van acumulant, i els seus efectes sobre la vida són de caràcter negatiu. Així passa amb les molècules d’hidrocarburs clorats i d’altres pesticides. En això, l’home segueix la mateixa estratègia dels altres organismes, que també empren molècules resistents, en la fusta, en missatgers químics, etc.
Realment, la Mediterrània no és gaire alterada. No ha sofert tants estralls com, per exemple, la Bàltica. I no és pas per mèrit dels habitants de les seves costes, sinó per virtut de la mateixa mar. El fet que la Mediterrània rebi, a la superfície, aigua procedent de l’Atlàntic, que ja hi entra amb pocs nutrients, i que dintre de la nostra mar aquesta aigua sigui sotmesa a un procés parcial de separació, amb sedimentació d’una part dels materials i evacuació cap a l’Atlàntic en profunditat, manté la mar poc productiva, en perjudici dels pescadors, però relativament neta i d’aigües transparents, en benefici dels turistes. És a dir, al contrari que a la mar Bàltica, on la circulació és inversa i la mar, tancada, acumula nutrients, augmenta la seva productivitat —es diu que s’eutrofitza— i és d’aigües més verdoses i menys transparents, amb una major facilitat per a causar problemes secundaris, com és ara l’esgotament de l’oxigen dissolt a l’aigua d’algunes conques.
La Mediterrània és oligotròfica, poc productiva, i això és un avantatge en aquests temps en què la càrrega de tota mena de materials augmenta. Principalment resulta favorable en les àrees centrals, on la vida planctònica és exactament igual a la que hi havia fa 50 anys. D’altra banda, la compartimentació de les aigües, per causa de corrents i fronts marins, pot limitar les vies i els efectes de la contaminació. Així, la procedent de costes franceses es mou més fàcilment cap a les nostres costes i, potser, queda relativament confinada en el corrent costaner.
Aquests darrers temps, hom s’ha preocupat força de les concentracions anormalment altes d’alguns metalls, que poden tenir efectes específicament negatius sobre la vida. Però, sovint els valors exageradament alts que s’han divulgat són simple conseqüència de tècniques poc acurades, cosa que no vol dir que se n’hagi de negligir el control, per més que l’aigua de mar té tas característiques que qualsevol excés d’elements serà probablement eliminat en forma sòlida. El mercuri planteja un problema especial; la seva concentració, que pot arribar a límits perillosos, es fa per via biològica i seguint les cadenes tròfiques.
Els problemes sanitaris, relacionats amb bacteris i virus, es limiten pràcticament a la costa, amb relació a desguassos, i més amb l’aigua propera al sediment i la que es troba entre les partícules del sediment que no pas amb les aigües lliures. L’ús d’aigua per refrigeració de centrals productores d’energia determina alteracions tèrmiques que són de poca importància. Només n’hi ha prou a calcular la calor evacuada i comparar-la amb l’intercanvi natural aigua/atmosfera per veure que els efectes no poden ser més que locals. Té més importància el tractament de l’aigua de refrigeració amb substàncies destinades a inhibir el creixement d’organismes a l’interior de les canonades.