
Els ecosistemes terrestres presenten característiques força diferents dels aquàtics. En ells els organismes depenen majoritàriament d’uns productors primaris que, situats a la interfase entre atmosfera i litosfera, ocupen un espai ben reduït (només un centenar de metres en el cas dels boscos més alts, i molt menys normalment). Alguns animals terrestres cerquen l’aliment al medi aquàtic i altres poden enlairar-se considerablement durant un temps curt, però el marc de la vida terrestre és molt més restringit que el de la marina, que arriba a colonitzar permanentment les grans fondàries. Les plantes terrestres, necessàriament fixes, poden tenir una llarga vida, i això, amb les característiques del relleu i dels sòls, fa que aquests sistemes conservin molt més profundament les marques de la història prèvia que no pas els medis fluids, i aquesta història condiciona molt llur funcionament actual. D’altra banda, la incidència de l’home, en els sistemes terrestres, és molt més aparent: arriba a organitzar-ne molt l’estructura en benefici propi, com passa als conreus d’aquesta fotografia, de l’horta valenciana.
Jordi Vidal.
Ens introduirem, ara, en el coneixement de la manera com funcionen els ecosistemes terrestres dels Països Catalans, començant amb algunes característiques generals de l’ecologia en el medi terrestre i passant, després, a analitzar les limitacions més importants que el nostre medi físic imposa a la producció vegetal i a l’organització dels sistemes ecològics a partir de l’entrada d’energia que aquesta producció representa. Entrarem també a considerar les solucions d’adaptació de l’organisme en el cas de les plantes, en tant que aquestes solucions condicionen l’esmentada organització dels ecosistemes, i alguns aspectes rellevants de les relacions entre animals i plantes. Acabades aquestes pàgines generals, tractarem les comunitats en una perspectiva inclinada cap als aspectes funcionals i d’adaptació, i no tant a la descripció detallada del paisatge. Algunes de les dades que donarem es publiquen per primer cop. Aquest esforç especial de recerca ha estat necessari perquè, per desgràcia, els coneixements que tenim de molts aspectes de l’ecologia terrestre als Països Catalans són encara força escassos i, sobretot, se centren en uns pocs ecosistemes, de manera que hauria estat impossible de donar una visió global amb una bona quantitat de dades pròpies. Això no obstant, el lector sabrà fer-se càrrec que l’ecologia terrestre és una ciència encara molt jove entre nosaltres, que ha disposat de molt pocs conreadors. Ens hem limitat a proposar un punt de vista sistèmic, si bé no equiparable al que podem tenir dels ecosistemes aquàtics, molt més documentats, sí prou elaborat per a servir de base inicial de comprensió i com a punt de partida.
Al llarg de les pàgines que segueixen ens ocuparem, doncs, dels aspectes que ens han semblat més essencials dels nostres ecosistemes terrestres, donant tants exemples propis com ens sigui possible. No podrem, però, oferir un tractament sistemàtic exhaustiu, primer perquè ens caldrien diversos volums i, segon, perquè ens mancaria molta informació. La selecció dels exemples i dels temes estarà inevitablement desequilibrada pel que fa a diversos aspectes. Així, hem preferit de referir-nos als ecosistemes mediterranis, ja que els de caràcter medioeuropeu, atlàntic o boreal tenen una bibliografia més extensa. D’altra banda, disposem de més informació sobre els productors primaris que no pas sobre els animals, fongs o bacteris. En zoologia, han estat més freqüents els estudis sistemàtics, biogeogràfics i etològics que no pas els ecològics, i són relativament rares a casa nostra, per la majoria de grups, les dades sobre censos, dinàmica poblacional, alimentació, selecció d’ambients, relacions depredador-presa, competència, etc. Els estudis del paper que tenen els fongs i els bacteris als ecosistemes terrestres són encara més rars, per no dir quasi inexistents, fora d’alguns casos de parasitisme en espècies útils a l’home. En definitiva, hem fet una presentació que sabem que és parcial, tant pel que fa als ecosistemes tractats, com a les temàtiques que s’hi toquen. Esperem, no obstant això, que hi hagi una coherència en el plantejament del conjunt i que el lector vulgui excusar les moltes llacunes que, inevitablement, apareixeran en l’exposició relativa a cada ecosistema.
Organització i estructura
Estructura i funcionament dels sistemes productius
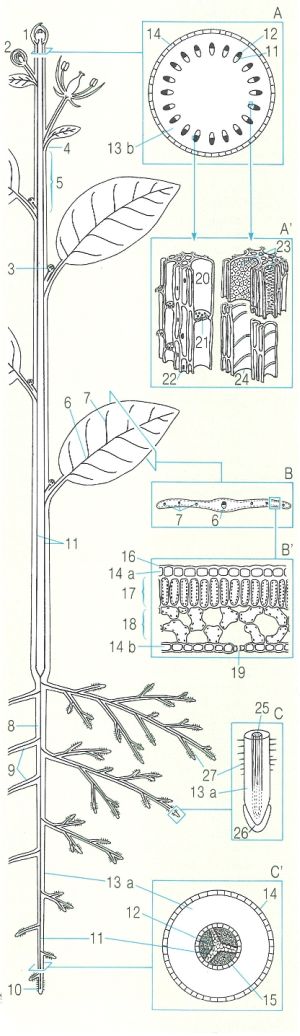
Organització general d’una planta i detalls de l’àpex caulinar, en un tall transversal (A) i longitudinal (A’), de la fulla (B, B’) i de les rels (C, C’). amb indicació de les parts principals. Les plantes vasculars han estat capaces de colonitzar el medi terrestre, gràcies a un conjunt d’adaptacions que els permet de mantenir un flux continu d’aigua amb nutrients des del sòl fins a les fulles, on es realitza la producció de matèria orgànica. Les fulles s’estenen en l’aire, un medi dessecant, en recerca de la llum. Pèls absorbents, vasos conductors o estomes són elements destacats d’aquesta organització adaptativa que garanteix l’aliment i el manteniment de l’estat hidratat dels teixits. 1 Àpex caulinar i borró terminal, 2 borró floral, 3 borró axil·lar, 4 nus, 5 entrenus, 6 nervi central, 7 nervi lateral, 8 rel principal, 9 rels laterals, 10 àpex radical, 11 xilema, 12 floema, 13 còrtex (a radical, b caulinar), 14 epidermis (a superior, b inferior), 15 endodermis, 16 cutícula, 17 parènquima en palissada, 18 mesofil·le esponjós, 19 estoma, 20 tub floemàtic, 21 placa cribrosa, 22 cèl·lules companyes nucleades, 23 vasos xilemàtics amb perforacions, 24 vasos xilemàtics amb engruiximents espiralats, 25 estela, 26 còfia, 27 pèls radicals.
Mercè Cartañá, a partir de fonts diverses.
Fins ara s’ha donat una visió comparativa molt general sobre els ecosistemes terrestres i aquàtics. La vida al continent presenta unes característiques netament diferenciades de les que imposa el medi aquàtic, tant en aspectes molt importants de les solucions adaptatives que han assolit evolutivament els diferents tipus d’organismes, com en l’organització a nivell de l’ecosistema. En el medi terrestre, la vida es localitza en una fina pel·lícula que es troba a la interfase entre el medi sòlid, constituït per la litosfera, i el medi gasós atmosfèric. Aquesta pel·lícula té, com a màxim, un centenar de metres de gruix. Alguns animals, com ara els ocells, i molts tipus de diàspores poden allunyar-se’n, però hi resten, de fet, vinculats per llur dependència de l’aigua i dels aliments, o es veuen limitats a romandre en formes de resistència. Un punt clau és que la vida terrestre activa continua associada a l’aigua, on molts organismes del sòl estan, de fet, immersos, i que els de vida aèria l’han incorporada d’alguna manera al seu medi interior.
El flux d’aigua i l’estructura vegetal
La dificultat principal per a la vida en el medi terrestre és, precisament, el caràcter dessecant del medi aeri, que dificulta el manteniment de l’estat d’imbibició del protoplasma cel·lular i, per tant, la continuïtat de les funcions vitals. En això el sòl té un paper decisiu. En efecte, dins el sòl hi ha una reserva d’aigua i de nutrients minerals de què depenen quasi tots els autòtrofs terrestres, és a dir la immensa majoria de les plantes verdes. Dins el sòl, però, no hi entra la llum. Les plantes vasculars han de captar la llum fora del sòl, però treuen l’aigua i els nutrients minerals del sòl. Això és determinant per a llur estructura. A diferència del que passa a la mar, no fóra possible, a terra, un fitoplàncton format per petits organismes aerovagants. A l’aire trobem, com dèiem, formes de resistència (espores, llavors, fruits, pol·len, etc.) però no autòtrofs actius, ja que les cèl·lules exposades a l’intercanvi gasós (indispensable per a la fotosíntesi) es dessecarien en el desert aeri. Les plantes terrestres són, com a conseqüència, totes fixes i tenen diferenciades una part aèria i una de subterrània, que representen proporcions variables de la biomassa total, segons les condicions específiques de cada indret i els resultats de l’evolució concreta de cada espècie vegetal. No tenen dificultat a aconseguir el subministrament d’oxigen i de diòxid de carboni, a partir de l’aire. En canvi, entre plantes que conviuen hi ha, molt sovint, competència per la llum, l’aigua i els nutrients (sobretot nitrogen i fòsfor).
Les plantes vasculars mantenen entre les fulles (òrgans fotosintetitzadors exposats a la llum i realitzadors de l’intercanvi de gasos) i el sòl un flux continu d’aigua que és absorbida per les arrels i conduïda al llarg de tiges i fulles fins a les cèl·lules del parènquima clorofíl·lic i fins als estomes. Es tracta d’un sistema vascular complex i perfeccionat, amb algunes variants menors que serveixen per a facilitar l’adaptació a medis de condicions diferents.
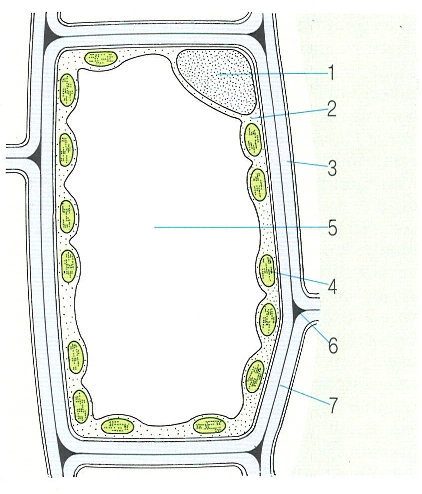
Organització d’una cèl·lula vegetal. Potser el caràcter més notable de la cèl·lula vegetal típica és la presència d’un o diversos vacúols centrals grans. L’estat hídric del protoplasma és regulat pels sucs vacuolars, que constitueixen un veritable medi interior. Això evita que el protoplasma estigui sotmès directament a les fluctuacions més importants del medi extern. Com un embassament fa per a nosaltres, el vacúol actua com una reserva reguladora del subministrament d’aigua a les parts actives. 1 nucli, 2 citoplasma, 3 paret cel·lular, 4 cloroplast, 5 vacúol, 6 espai aeri intersticial, 7 membrana cel·lular.
Mercè Cartañá, a partir de fonts diverses.
També les cèl·lules de les fulles de les plantes superiors tenen característiques peculiars d’adaptació al medi terrestre. Es tracta de cèl·lules grans en què la major part del volum és ocupat per un gran vacúol, de vegades subdividit per envans fins; el protoplasma queda reduït a una fina làmina perifèrica, en contacte amb la membrana. L’estat d’imbibició del protoplasma es manté per l’intercanvi amb els sucs continguts al vacúol, i, per això, cal que la relació volum vacuolar/volum protoplasmàtic sigui molt alta. El vacúol actua com una comporta que regula la hidratació de les parts actives i constitueix un veritable medi interior, controlat pel mateix vegetal; per això les plantes vasculars amb cèl·lules vacuolades típiques s’anomenen homeohídriques.
Finalment, l’intercanvi gasós que permet l’entrada de diòxid de carboni per realitzar la fotosíntesi implica que també surti vapor d’aigua des del medi més hidratat (els teixits vegetals) fins al més eixut (l’atmosfera). Per a estalviar pèrdues excessives en els moments de més capacitat evaporativa de l’aire, o de menor disponibilitat d’aigua per a les arrels, els vegetals tenen les fulles cobertes d’una cutícula que pot reduir força la transpiració, i gran part de l’intercanvi es fa a través dels estomes, que funcionen com a vàlvules de control en el procés. Segons quin sigui el seu estat hídric, la planta obrirà o tancarà els estomes, i, d’aquesta manera, l’economia de l’aigua condicionarà l’absorció i la fixació de carboni, és a dir, la producció.
No obstant això, no totes les plantes terrestes responen a aquesta estructura que acabem de repassar succintament. N’hi ha que són capaces de sobreviure en el medi aeri sense mantenir un flux continu d’aigua des del sòl; es tracta sobretot d’algues, briòfits i líquens, i també d’algunes falgueres i fanerògames. Un exemple de fanerògama d’aquesta mena és l’orella d’ós (Ramonda myconii); entre les falgueres que viuen en medis rocosos podem esmentar diversos gèneres, com Ceterach, Notholaena, Cheilanthes, etc. Aquestes plantes normalment no absorbeixen aigua per les arrels, si és que en tenen, sinó que ho fan especialment a través de l’epidermis de les fulles. Si la humitat atmosfèrica és elevada, són capaces d’absorbir aigua i les cèl·lules s’hidraten. Aleshores, poden activar els processos de la fotosíntesi. El secret adaptatiu d’aquestes plantes es troba en algunes característiques anatòmiques i fisiològiques: tenen cèl·lules petites i gairebé sense vacúols; llur protoplasma pot resistir, reduint al mínim l’activitat vital, llargues fases de deshidratació, de forma que la fotosíntesi es produeix només en condicions d’humitat atmosfèrica molt alta o aprofitant la rosada que es diposita damunt els òrgans fotosintetitzadors. Les plantes d’aquesta mena reben el nom de poiquilohídriques. Poden sobreviure en ambients molt adversos, com ara els deserts o les roques (les superfícies de les quals gairebé no retenen aigua), aprofitant la humitat de la matinada i les pluges que hi puguin caure. Són, de tota manera, més freqüents als llocs d’ambient humit, però, com que l’activitat productiva es restringeix als moments favorables, no poden competir, sinó molt limitadament, amb les homeohídriques, l’avantatge decisiu de les quals és precisament el fet d’haver assolit una independència considerable respecte de les condicions ambientals de cada moment.
Entre la forta variabilitat d’hidratació que suporta el protoplasma de les plantes poiquilohídriques i el manteniment de valors constants hi ha tota una gamma de situacions. Algunes plantes homeohídriques tenen poca capacitat de regulació, sense arribar a l’extrem de les poiquilohídriques, i el contingut hídric fluctua força i en poc temps (per això en diem hidrolàbils). Estan adaptades a aquests canvis, de forma que poden considerar-se eurihídriques. A l’extrem oposat hi ha plantes que tenen òrgans de reserva d’aigua: són suculentes o mantenen recursos abundants als troncs i a les branques, i tenen un bon sistema radical i una regulació estomàtica efectíva. Són les anomenades hidrostables. Tots els arbres s’inclouen en aquest grup, si bé alguns són més hidrostables que altres.
La llum i l’estructura vegetal
La competència per la llum és un punt central en l’organització dels ecosistemes terrestes. La llum és la font de l’energia metabòlica i esdevé un element limitant per a la vida vegetal quan la seva intensitat es redueix molt (per exemple, dins un bosc dens, la llum que arriba al sòl pot ser només un 1% de la que incideix a la part superior de les capçades dels arbres i esdevenir insuficient per al creixement de moltes plantes). Això vol dir que es produeix una pressió selectiva a favor de les plantes capaces d’assegurar-se la llum necessària d’una forma o altra. Per tant, la competència per la llum tendeix a afavorir la formació d’estructures verticals, allunyant els òrgans fotosintetitzadors, les fulles, de la superfície del sòl: si una planta alça les fulles per damunt de les plantes veïnes obté la llum necessària i, alhora, en priva les altres. La carrera cap a la llum té uns límits d’alçària imposats pel fet que cal continuar pujant aigua i nutrients fins a les fulles. En termes generals, l’alçada màxima que assoleix una determinada comunitat vegetal es relaciona amb les disponibilitats d’aigua i nutrients del lloc. Més endavant ens ocuparem de diverses qüestions relatives a la transcendència de la llum per a la producció. Tradicionalment hom distingeix entre plantes heliòfiles, que funcionen òptimament a altes intensitats de radiació, i esciàfiles, pròpies d’ambients de radiació reduïda.
Parts productives i parts estructurals
Les fulles són les parts productives de les plantes. El seu metabolisme és molt més actiu que el de les parts de sosteniment o conductores (troncs, branques), el creixement és més ràpid, la composició química és diferent, i es renoven més de pressa, de manera que llur vida mitjana és relativament breu. En contrast, a les parts amb una funció estructural, a més d’un metabolisme més pausat, hi ha un predomini de materials que donen persistència (cel·luloses, lignines) i resistència o protecció superficials (escorces).
La proporció entre parts actives i productives i parts estructurals varia d’uns ecosistemes a uns altres, i pot ser un índex útil per a identificar certes característiques funcionals de cada ecosistema. Això no obstant, no ens hem d’interessar només per les relacions en termes de masses o volums, sinó també per les que hi ha en termes de superfícies, ja que les proporcions entre superfícies d’intercanvi (l’àrea foliar, sobretot) i volum o pes poden modificar-se també sensiblement. Algunes plantes, com ara les cactàcies i moltes euforbiàcies, perden evolutivament les fulles o les redueixen molt. Les tiges són, aleshores, verdes i fotosintetitzadores. L’àrea activa i transpiradora es redueix molt amb relació al volum del cos. Sense recórrer a aquests casos extrems, plantes com la ginesta també mostren tiges verdes fotosintetitzadores i una forta reducció de la superfície d’intercanvi. En definitiva, la proporció entre parts actives i parts estructurals en termes de superfície o de biomasa depèn, sobretot, dels factors ambientals, essencialment pel que fa a proporcions relatives i a quantitats absolutes dels recursos (aigua, llum, nitrogen, fòsfor, etc.), i de la pressió exercida pels herbívors o per factors mecànics (com el vent), com també del resultat de la selecció sobre les característiques genètiques en el marc d’aquests condicionaments ambientals i biòtics i al llarg de la història.
Si considerem ara les parts subterrànies, podem retrobar la diferenciació entre, d’una banda, unes parts més actives, les arrels més fines, amb molts pèls absorbents, sense càmbium vascular i amb el creixement limitat, sovint associades a fongs que n’incrementen la superfície d’intercanvi (micorizes), de taxa de renovació elevada (la producció d’arrels fines és comparable, i de vegades superior, a la de les fulles); i, d’altra banda, unes parts sobretot conductores i de reserva, les arrels gruixudes. Per desgràcia, hi ha molta menys informació sobre biomasses, proporcions relatives i produccions d’aquestes parts subterrànies, per problemes metodològics fàcils d’entendre. Alguns autors han mirat de trobar una relació entre la superfície foliar (índex d’àrea foliar) i la de rels fines, però hi ha moltes dificultats.
Les parts que anomenem estructurals, és a dir, troncs, branques, arrels gruixudes i estructures com bulbs, tubercles i d’altres, tenen una important funció d’emmagatzemament d’aigua i de substàncies (com el midó) que poden mobilitzar-se quan convé per obtenir noves produccions o refer-se de pèrdues. A més, totes aquestes estructures asseguren als vegetals el control dels fluxos verticals. Aquests, en el medi aquàtic, depenen de turbulències o afloraments d’aigua profunda, el control dels quals és extern als organismes. En el medi terrestre, en canvi, la pujada dels nutrients a les parts productives es fa per l’interior del cos dels vegetals. Aquest control intern del transport, juntament amb el menor recorregut dels cicles materials (ja hem dit que ens movem en un interval vertical d’uns 100 m, mentre que a la mar aquest gruix és molt més gran) expliquen precisament la major eficiència productiva dels ecosistemes terrestres, que poden fixar una mitjana anual d’uns 300 g C/m2, enfront dels 100 g que fixen els sistemes aquàtics.
Centrem-nos ara un xic més en les parts productives. L’àrea foliar, és a dir, la superfície de fulles per unitat de superfície de terreny, és un paràmetre molt utilitzat per a descriure els diferents ecosistemes. L’índex varia, i pot arribar a valors de 15 o 20 en boscos aciculifolis de coníferes, però normalment és al voltant de 4 en ecosistemes molt diferents, des d’un prat a un bosc de planifolis. Ací trobem una altra diferència entre els ecosistemes aquàtics i els terrestres. En una làmina d’aigua, n’hi ha prou amb una concentració de 400 mg/m2 de clorofil·la per a absorbir pràcticament tota la llum. Aquesta és la concentració usual també a les fulles, però amb una àrea foliar de 4 hi hauria quatre cops més de pigment per unitat de superfície que no pas a l’aigua. Els sistemes terrestres sembla que deixen passar la llum, i l’usen amb més eficàcia, gràcies a reflexions entre les fulles a l’interior de la capçada. És clar que aquest factor pot influir en la major eficiència productiva del medi terrestre.
Ultra la superfície total de fulles, cal considerar també les dimensions de cada fulla, la disposició que adopten en l’espai (angle respecte de terra), les formes de les fulles amb el limbe més o menys dividit i les vores més o menys regulars, les característiques de llurs superfícies, etc. Tots aquests factors influeixen en els balanços d’aigua i de calor de les fulles i determinen les possibilitats adaptatives a condicions específiques del medi físic.
La fotosíntesi i l’eficiència fotosintetitzadora
La fotosíntesi és el procés bàsic de la producció. Per ella, les plantes fixen l’energia radiant de la llum en enllaços químics que són el ciment amb el qual es construeixen les complicades molècules orgàniques a partir de materials més senzills: àtoms de carboni, oxigen i hidrogen procedents del diòxid de carboni atmosfèric i de l’aigua, i elements com nitrogen, fòsfor, sofre, calci, potassi, magnesi i molts d’altres procedents, sobretot, de les sals minerals del sòl.
Normalment, la fotosíntesi condueix a la formació inicial de molècules d’hidrats de carboni amb sis àtoms de carboni, que s’escindeixen en dos d’àcid 3-fosfoglicèric. Això és el que s’anomena via C3 de fixació del CO2, on aquest és fixat inicialment per un acceptor, la ribulosa 1-5 difosfat (RuDP).
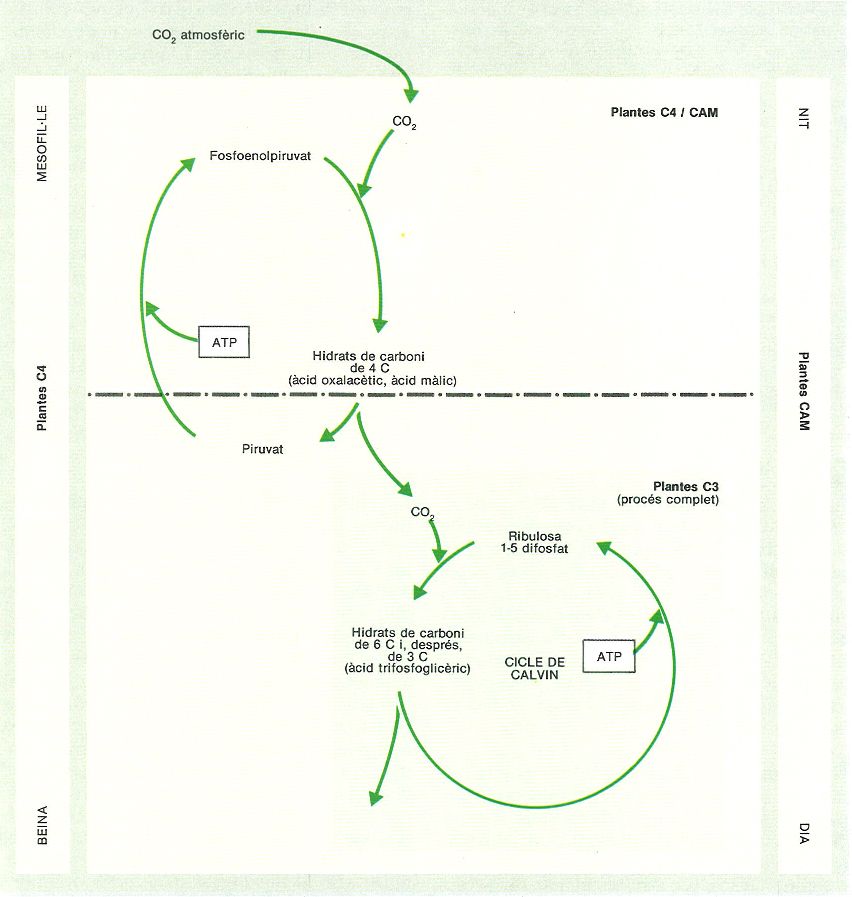
Tipus metabòlics dels vegetals. Les plantes C3 integren el diòxid de carboni absorbit de l’aire a la ribulosa 1-5 difosfat per obtenir hidrats de carboni de sis carbonis, que després passen a àcid trifosfoglicèric (cicle de Calvin). Les plantes C4 i CAM tenen un cicle previ en què el diòxid de carboni és fixat en oxalacètic i màlic; després, es torna a alliberar per passar al cicle de Calvin normal. L’avantatge de la separació en dos cicles és que permet separar el procés de fixació del diòxid de carboni del de producció d’hidrats de carboni, ja sigui en estructures diferents (C4, en el mesofil·le i la beina) o en moments diferents (CAM, nit i dia). Les plantes C4 aprofiten el diòxid de carboni que resulta de la pròpia respiració, per a augmentar la producció neta, i les CAM poden absorbir-lo de nit i tancar els estomes de dia per reduir el consum hidric.
Román Montull, a partir de fonts diverses.
Això no obstant, no totes les plantes segueixen el mateix procediment bioquímic. Algunes gramínies, amarantàcies, quenopodiàcies i altres fanerògames, d’origen tropical, que mostren rendiments fotosintètics sorprenentment elevats i poden funcionar amb concentracions de CO2 a l’aire molt més baixes que les normals, segueixen una via modificada. L’acceptor del CO2 és ara el fosfoenolpiruvat (PEP), i es formen hidrats de carboni de quatre àtoms de carboni en lloc de tres: primer, àcid oxalacètic i després àcid màlic. Finalment, es regenera el piruvat i queda lliure altre cop el CO2, que inicia ara un cicle similar al normal C3. Aquestes plantes reben la denominació de plantes C4, i llur sistema metabòlic, més complicat, sembla força més eficient. Solen presentar també diferències anatòmiques amb els feixos vasculars de les fulles voltats per una capa de cèl·lules mesofíl·liques on es concentren els cloroplasts, els quals també tenen diferències ultrastructurals amb els de les C3. Tot i això, les vies C3 i C4 es poden donar en espècies del mateix gènere, com passa a Atriplex i Euphorbia, que fins i tot formen híbrids amb caràcters intermedis.>
Altres plantes encara, com passa amb moltes de suculentes (cactàcies, crassulàcies, orquidàcies, bromeliàcies, etc.), usen també el fosfoenolpiruvat i el pas per àcid màlic, però es caracteritzen perquè fan la fixació de CO2 durant la nit, que és quan obren els estomes. Després, amb el CO2 absorbit, es formen àcids orgànics que s’emmagatzemen als vacúols. De dia, amb els estomes tancats, els àcids van als plastidis de les cèl·lules que volten els feixos. conductors, i contínua el procés com a les C4. És l’anomenat metabolisme àcid de les crassulàcies. Aquesta separació entre la fixació de carboni (i, per tant, l’intercanvi gasós) i la fase lluminosa de la fotosíntesi estalvia, òbviament, moltes pèrdues d’aigua.
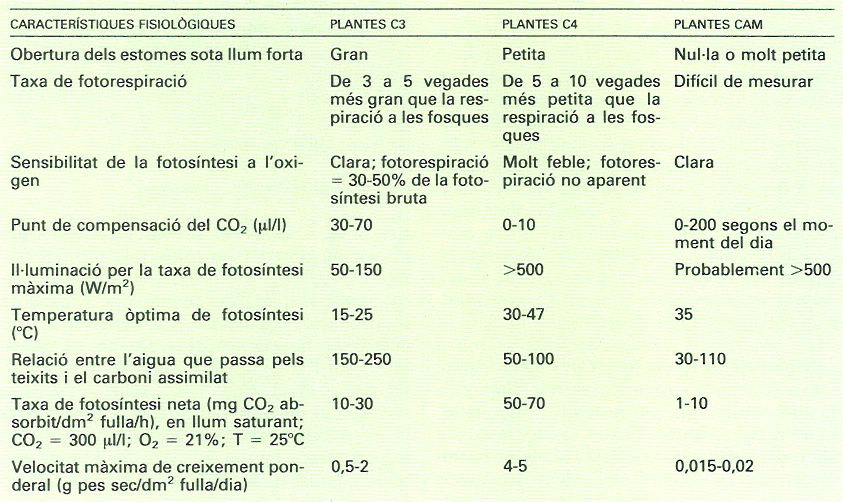
Característiques fisiològiques comparades per als tres tipus de metabolisme de les plantes (C3, C4 i CAM), segons A. Moyse. Noteu les diferències, molt importants: per exemple, les C4 redueixen molt la fotorespiració i això els permet tenir una velocitat màxima de creixement en pes més alta, però també tenen requeriments superiors de llum i temperatura; les CAM, en canvi, destaquen per una eficiència més gran en l’ús de l’aigua per a la producció, però llur velocitat màxima de creixement és molt baixa.
Informació d'A. Moyse, cedida per Pere Montserrat
Les diferents característiques metabòliques de les plantes determinen avantatges per a unes o altres en diferents ambients. Les plantes C4 tenen una temperatura òptima per a la fotosíntesi més alta i fan un ús més eficient del CO2 (poden aprofitar àdhuc el que es produeix per fotorespiració, amb els estomes quasi tancats). La proporció de plantes C4 augmenta dels països freds als càlids, i són especialment importants damunt de sòls salins i a les praderies tropicals. En conjunt, gasten molta menys aigua per unitat de matèria produïda. Les plantes amb el metabolisme àcid de les crassulàcies (també dites plantes CAM, per les inicials angleses d’aquesta denominació) són pròpies de llocs on cal suportar períodes molt llargs d’eixut. És interessant de notar que hi ha diversitat de comportament entre les plantes C3 i C4 respecte dels isòtops de diferents elements. Les plantes C3 prenen una proporció menor de l’isòtop pesat C
Economia global dels productors
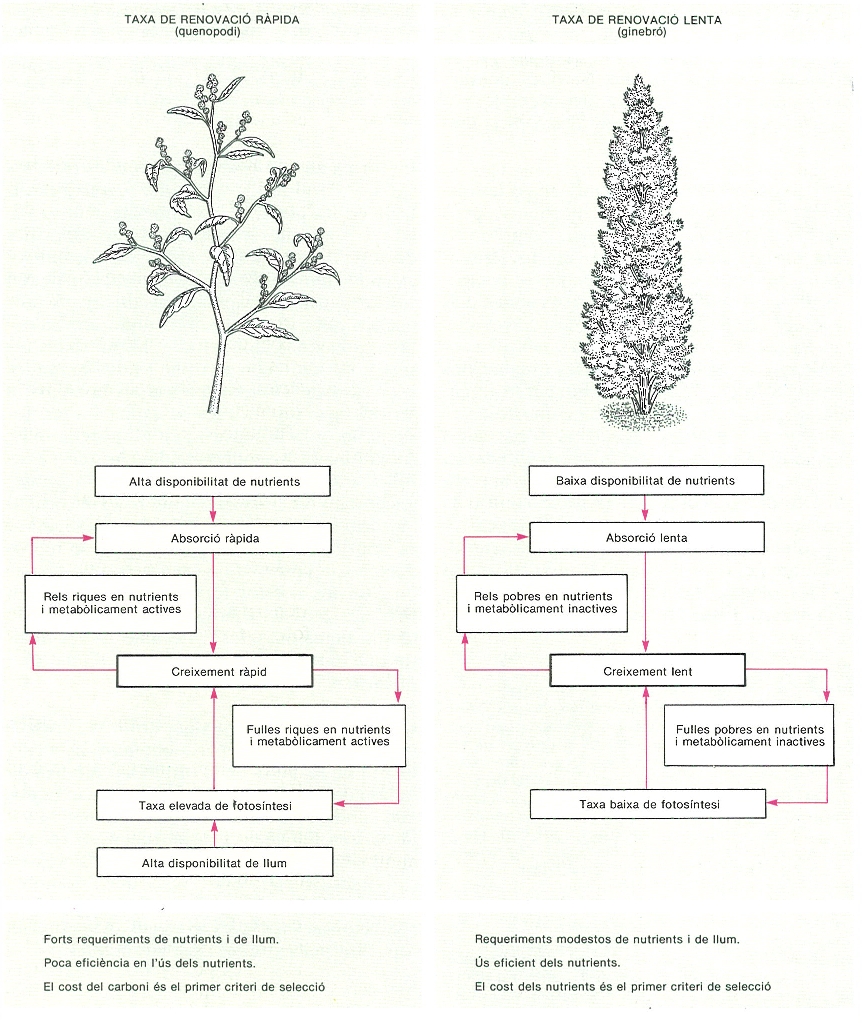
Dues estratègies oposades en la utilització de recursos (aigua, nutrients) per les plantes, que condicionen diversos aspectes de llur desenvolupament i llurs característiques ecològiques. L’esquema compara les estratègies de taxa de renovació ràpida i lenta, exemplificades respectivament per una planta ruderal, com pot ser un quenopodi (Chenopodium), i un arbust de creixement lent, com un ginebre (Juniperus communis).
Mercè Cartañá, a partir de fonts diverses.
Les plantes terrestres constitueixen un sistema productiu complet, que comprèn llocs de fabricació, vies de transport, magatzems i estructures protectores, com també òrgans de reproducció. Aquest sistema depèn d’uns recursos o primeres matèries presents a l’entorn (aire, sòl). Els materials produïts poden servir per al creixement del sistema o per a la substitució de parts envellides o consumides (i, en ambdós casos, són reinvertits en el propi sistema), o bé per a la creació de nous sistemes similars, és a dir, per a la reproducció i per a la dispersió.
La producció elaborada per la planta (que podem expressar per la quantitat de carboni o de matèria orgànica, o en termes d’energia) s’assigna a unes o altres finalitats (estructures productives, magatzems, estructures de suport i conducció, estructures reproductives i de disseminació, etc.) segons uns criteris que podríem considerar econòmics. Com a resultat d’aquesta atribució diferencial, la forma d’un vegetal pot variar d’uns ambients a uns altres, encara que la pauta de creixement té un determinisme genètic essencial. A l’adaptació evolutiva, que és el resultat de la selecció exercida per l’ambient al llarg de moltes generacions, s’hi superposa, per tant, una resposta fenotípica resultant de les circumstàncies que afecten localment l’«economia del carboni».
Carboni
En el balanç global del carboni de què disposa una planta, una part es perd en diòxid de carboni per respiració de carbohidrats fotosintètics, en el procés de construcció dels teixits; però, se’n consumeix una quantitat molt més gran en el manteniment d’aquests teixits. El cost de manteniment varia amb el tipus de teixit i amb les condicions ambientals. Les plantes acumulen carbohidrats, sobretot midó, al parènquima de la part interior de l’escorça i de l’albeca, sobretot als troncs, a les soques i les rels més gruixuts. Aquestes reserves es mobilitzen quan s’inicia el creixement d’eixos (tiges o rels) a la primavera, i també poden emprar-se en la resposta a una defoliació o a la destrucció de tota la part aèria (per exemple, per un incendi): com més gran és la reserva de midó, més gran és el dany que pot ser resistit, i és freqüent que un arbre pugui aguantar més de dues defoliacions successives. Un roure, per exemple, pot tenir reserves per a construir tres cops la biomassa foliar.
En l’assignació de recursos influeixen les disponibilitats d’aigua, llum i nutrients, però també els factors d’explotació, com són el vent, els herbívors o el foc. En efecte, els herbívors consumeixen plantes senceres o algunes parts, el vent pot trencar parts aèries dels vegetals i el foc pot suprimir-les; altres factors encara, com les sals o la presència de contaminants atmosfèrics, les fortes fluctuacions tèrmiques o els transports horitzontals intensos associats a processos geomorfològics (substrats inestables, pendents forts, etc.) constitueixen motius de pèrdues de biomassa, és a dir, d’explotació. L’economia de la planta serà determinada per la relació entre la producció permesa per la disponibilitat de recursos i l’explotació més la mortalitat natural, i també per les possibilitats, genèticament fixades, d’aprofitar els recursos i respondre als esdeveniments de l’entorn.
Per mostrar un exemple dels criteris econòmics que regeixen l’assignació de recursos a unes estructures o unes altres, ens referirem a les fulles i els corresponents suports immediats (branquillons, pecíols, raquis, etc.). Aquests suports demanen una despesa mesurable d’energia i de materials, que podem resumir pel pes sec de matèria orgànica que representen aquestes estructures totes juntes. Si es redueix al mínim la inversió metabòlica per als teixits de sosteniment no productius, quedaran recursos disponibles per a ser invertits en processos reproductius, per a ser emmagatzemats o per a ser usats per al creixement de tiges, rels, etc. Quan la planta ha de competir amb altres plantes veïnes i li cal créixer ràpid, o quan la possibilitat de producció és limitada, és avantatjós de reduir els costos que representa la construcció d’aquestes estructures: la selecció tendeix a afavorir la màxima superfície fotosintetitzadora amb el mínim cost metabòlic d’elements passius. Ara bé, la minimització d’aquests costos no pot depassar certs límits. En primer lloc, cal mantenir una capacitat de transport proporcional a la superfície activa, i, en segon lloc, cal garantir la resistència mecànica de les fulles, que han d’aguantar la força del vent i la pluja, els xocs per caiguda de materials i el pes d’animals (insectes, aus, etc.). Així, doncs, cal optimitzar la relació entre l’economia de medis-seguretat i la persistència.
La mateixa mena de raonament econòmic s’imposa a l’hora de considerar els tipus de fulles. Els costos metabòlics de construir i mantenir una fulla han de ser proporcionals a la capacitat d’aquesta fulla per produir al llarg de la vida. Les plantes presenten variacions, tant en la composició de les fulles (més o menys riques en elements nutritius, més o menys esclerificades per la presència de molècules com la cel·lulosa i la lignina), com en la seva durada (menys d’un any, entre un i dos anys o bé molts), per tal d’ajustar-se a les possibilitats del medi per a la producció i als ritmes estacionals en què es donen els moments favorables. No obstant això, el cost total de fabricació i manteniment d’una fulla caduca o perenne sembla que pot ésser similar, i l’opció per una estratègia perennifòlia dependrà de la necessitat, imposada externament, d’allargar el període de vida, de l’assimilació de cada fulla individual, de l’avantatge d’acumular una major àrea foliar superposant fulles de diferents edats, de la millora d’ús dels nutrients minerals i, potser, de la reducció, per via de l’esclerofil·lisme, de les pèrdues per herbivorisme i lixiviació. Els caducifolis eviten l’eixut estival o el fred de l’hivern, segons els casos, i, en compensació, maximitzen la taxa fotosintetitzadora.
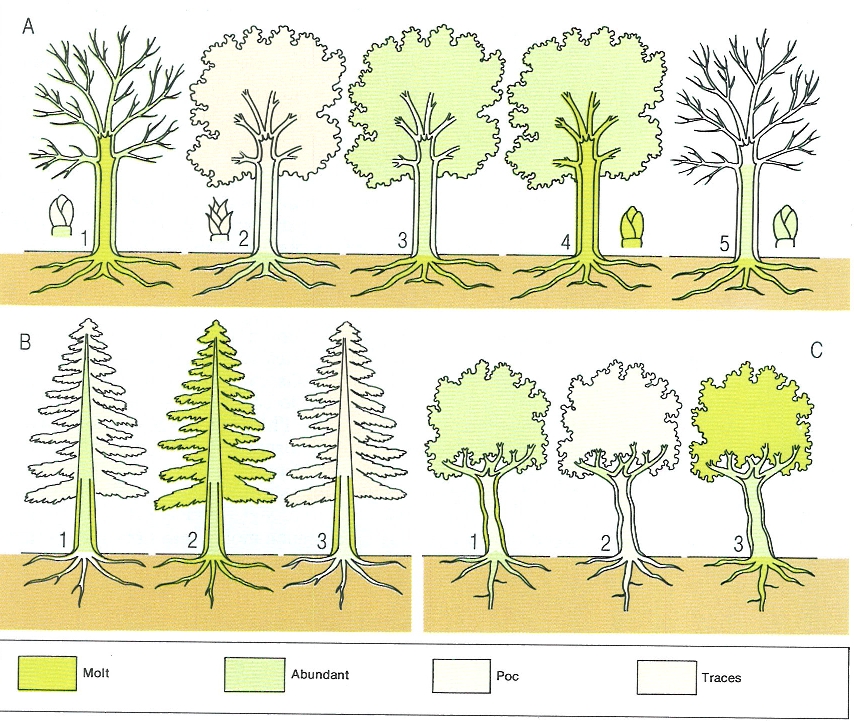
Variació de la distribució de reserves a les plantes al llarg de l’any, que mostra l’evolució del contingut de midó en un caducifoli (A), una conífera (B) i un perennifoli esclerofil·le (C). Els números indiquen, per al faig (Fagus sylvatica): 1 primavera, abans de sortir les fulles; 2 durant el desenvolupament de les fulles; 3 estiu; 4 tardor, abans de la caiguda de les fulles; i 5 hivern, amb glaçades. Per a l’avet (Abies alba): 1 primavera, durant el desenvolupament, 2 final de l’estiu; 3 hivern, amb gelades. Per a l’olivera (Olea europaea): 1 primavera, durant el desenvolupament; 2 ple estiu, amb eixut; 3 hivern, sense gelades.
Mercè Cartañá, original de W. Larcher.
La forma en què es distribueixen les reserves varia al llarg de l’any i també segons l’estratègia de les plantes. Això no solament és vàlid per a aquelles que tenen òrgans específics de reserva, com els tubercles que acumulen el midó durant l’època desfavorable i poden construir després ràpidament els òrgans productius quan les condicions milloren: en arbres caducifolis, com el faig, les màximes reserves de midó són al tronc i a les branques i, en avançar l’hivern, passen als borrons i a les rels, per tornar a les branques i a les noves fulles a la primavera; en els perennifolis, com l’olivera, s’acumula midó a les rels i a les fulles, que fan, així, de magatzem; les coníferes boreals tenen molt poques reserves durant l’estació freda. La producció de fulles sempre és prioritària. En canvi, els òrgans de reproducció són costosos i és per això que els arbres de les nostres terres només en fan en períodes de 2 a 10 anys. Aquest interval de temps augmenta a les zones més fredes mentre que al tròpic pot haver-hi més d’una fructificació a l’any.
Un altre cas en què podem utilitzar l’aproximació econòmica és per a entendre el repartiment de recursos en la producció relativa de fulles i de rels fines. S’ha vist que, quan es donen condicions d’eixut moderat o quan al sòl hi ha una deficiència de nitrogen, el creixement de les tiges és inhibit i es transfereixen carbohidrats cap a les rels, que aleshores creixen amb més força. És evident que, en les condicions esmentades, convé d’augmentar la capacitat per extreure recursos edàfics.
Nutrients
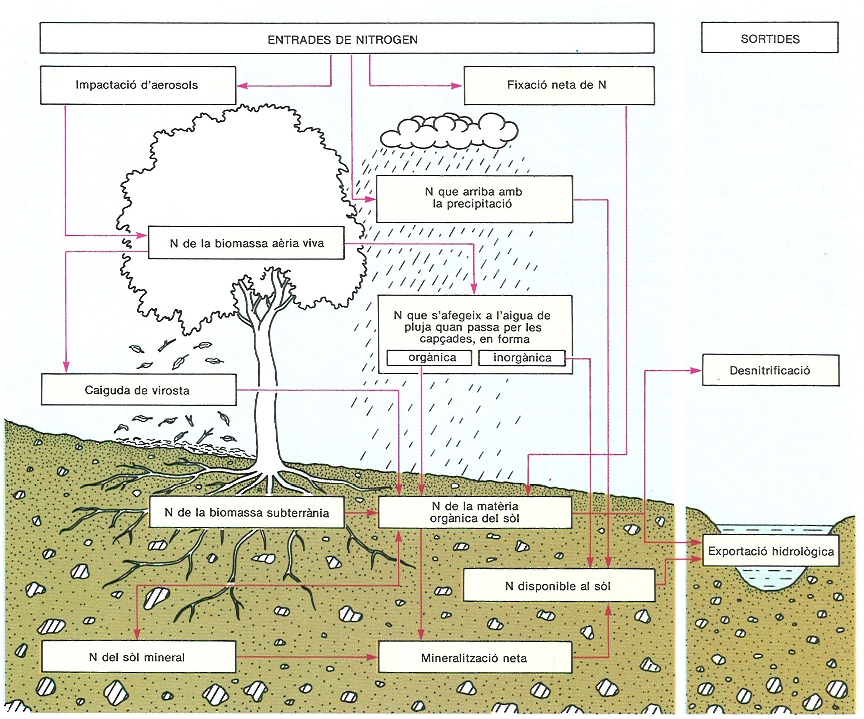
Cicle del nitrogen en un ecosistema forestal, segons un model simplificat que mostra les entrades (impacte d’aerosols, precipitació, fixació) i les sortides (desnitrificado, exportació hidrológica) principals, els compartiments interns i els fluxos. La quantificació completa del nitrogen implicat en cada procés o magatzem demana estudis molt laboriosos, però aporta una informació indispensable per a comprendre el funcionament del sistema i les conseqüències de les pertorbacions. El nitrogen és particularment important, ja que molt sovint limita la producció primària.
Mercè Cartañá, a partir de fonts diverses.
Fins ara ens hem referit a l’economia del carboni, però també es donen fenòmens d’assignació de recursos si considerem altres elements. Hem vist que les possibilitats productives són determinades per l’aigua, la llum, i també per la disponibilitat de nutrients minerals. La noció de factor limitant s’introduí per explicar que la producció vegetal d’un cultiu és determinada per aquell factor que es troba en menor quantitat en relació amb les necessitats de la planta. Si aleshores afegim certes quantitats de l’element limitant, obtindrem un ràpid increment de la producció. La noció de factor limitant s’ha estès també als ecosistemes naturals, però la situació esdevé sovint més complicada. Les comunitats que viuen sobre els sòls menys fèrtils no són, però, les que responen més intensament a l’addició de nutrients. Diverses raons ho expliquen. En primer lloc, la proporció del fertilitzant que s’adsorbeix i queda immobilitzat, o que és capturat pels microorganismes, pot ser molt diferent en sòls pobres i en d’altres de més fèrtils; de fet, els microbis competeixen amb avantatge amb les plantes, ja que es trobem limitats per la baixa qualitat de la virosta produïda (teixits pobres en nutrients i rics en fenols): si s’hi afegeixen adobs, s’activa la producció microbiana i n’augmenta la biomassa en detriment de l’increment esperat en les plantes. En segon lloc, les espècies que viuen en uns llocs són diferents de les d’altres llocs i tenen capacitats de creixement genèticament diverses: les plantes de llocs més pobres tenen una taxa menor de creixement damunt de sòls rics o adobats; per contra, les espècies de llocs més rics serien eliminades competitivament damunt els sòls més pobres. De fet, és normal que s’observin més situacions de limitació nutritiva entre les plantes de llocs rics.
La disponibilitat de nutrients és afectada per l’activitat dels animals. Si hi ha un grau alt d’explotació per a pastura, el cicle de retorn cap al sòl s’accelera i el material retornat és més ric que la virosta, perquè els herbívors mengen de forma selectiva, no ingereixen només fulles senescents i en la digestió es concentren elements en formes fàcils de reutilitzar. Afegim a això que, en estimular la producció, com veurem després, la pastura ajuda que es produeixin fulles amb elevades concentracions d’elements nutritius. Ara bé, aquests efectes positius no es dónen damunt sòls gaire pobres on, en general, les plantes són pobres en elements nutritius i presenten adaptacions defensives contra el consum i altes concentracions de lignines i tanins. Un increment de l’explotació sol conduir, en aquest cas, a una reducció de la producció.
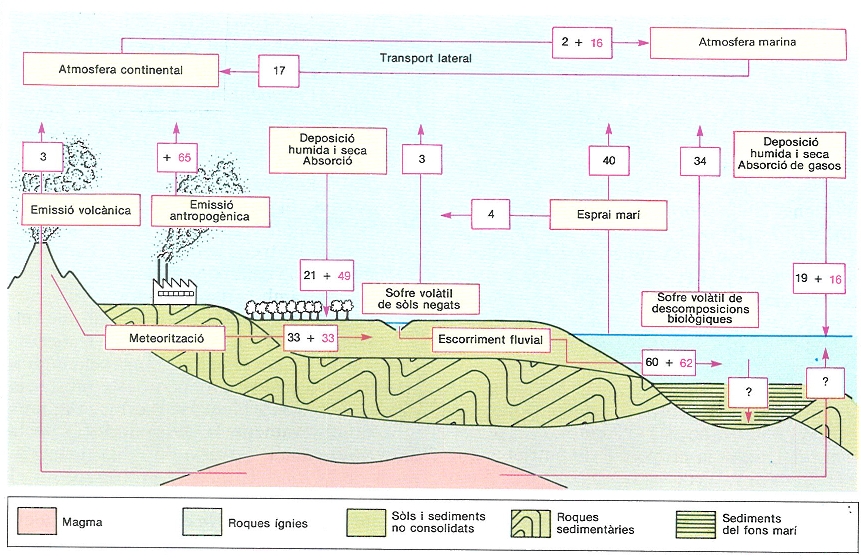
Cicle global del sofre, un dels cicles biogeoquímics més profundament marcats per la influència humana i que han despertat més preocupació. L’esquema presenta valors estimatius, en milions de tones l’any, per als fluxos més importants. Les xifres en negre corresponen als valors previs al període de forta incidència de l’activitat humana i les xifres en vermell, a la contribució d’aquesta activitat.
Mercè Cartañá, original de C. Granat i altres.
La concentració de nutrients de les diferents parts dels vegetals pot variar molt, i també hi ha canvis al llarg del temps en un mateix òrgan. Així, doncs, pel que fa als recursos minerals també hi ha qüestions d’assignació que cal tenir en compte. Les fulles solen ser més riques en nitrogen, fòsfor i potassi que no pas les estructures llenyoses. Els fruits acumulen potassi, fòsfor i sofre. Normalment, als troncs i a les branques hi ha concentracions relativament elevades de calci. Les composicions varien també dins un mateix òrgan amb el temps, tant en resposta als canvis estacionals (del medi o de l’activitat biològica de la planta) com per l’envelliment de l’òrgan. A les fulles joves acabades de brotar hi ha inicialment una gran aportació de nitrogen, potassi i fòsfor; després, les concentracions d’aquests elements es dilueixen a causa de la producció de materials estructurals.
En una planta individual, la major part dels nutrients essencials (nitrogen, fòsfor, sofre i, sobretot, potassi) es consumeixen en la fabricació dels òrgans de renovació ràpida; concretament, les fulles, que tenen la major concentració de nitrogen, les rels fines i els fruits, particularment rics en potassi, s’enduen la part més gran dels nutrients absorbits per les rels. No obstant això, és cert que no tots aquests recursos tornen al sòl amb la caiguda d’aquests òrgans. Durant la senescència, bona part dels nutrients acumulats a les fulles són retirats i translocats a òrgans permanents, on s’emmagatzemen per a tornar-se a usar a l’estació de creixement següent. Per a altres nutrients el comportament és menys conservador. El calci o el ferro, per exemple, més aviat tendeixen a acumular-se a les fulles a mesura que envelleixen; el calci té les concentracions màximes a l’escorça i no pas a les fulles. Naturalment, aquestes diferències es relacionen amb el paper específic que té cada element en el metabolisme del vegetal i amb una adaptació per a retenir dins les pròpies reserves aquells elements que més sovint esdevenen limitants per a la producció. Assenyalem també que la proporció de nutrients que es retransloca des de les fulles pot variar d’uns llocs a uns altres, i que les concentracions dels diferents elements que trobem en un determinat òrgan no són completament fixes per a cada espècie sinó que depenen de la riquesa del sòl en cada lloc.
Els mecanismes de retenció no solament es limiten a la fisiologia individual sinó que en l’ecosistema n’hi ha de molt eficients. L’associació d’herbes amb microorganismes (les fabàcies amb simbionts fixadors de nitrogen i les gramínies amb fixadors que viuen a la rizosfera) permet, a més, d’incrementar les entrades de l’element més decisiu per a la producció. Encara que no es tracta de mecanismes de retenció, sinó d’absorció, convé de recordar-los ací per a tenir una visió més completa de l’economia dels nutrients a nivell de l’ecosistema. Si hi afegim també l’acció de bacteris i cianofícies lliures, trobem que, en certs casos, el nitrogen obtingut per fixació és fins cinquanta o cent cops més important que el que arriba amb la deposició seca (pols, aerosols) i amb aigües de pluja (les altres úniques entrades al sistema). No obstant això, sovint també és molt menys, ja que sembla que els fixadors són molt poc actius en sòls massa pobres en nitrogen i que tenen requeriments relativament elevats de fòsfor, molibdè, ferro i cobalt.
La reserva més gran de nitrogen i fòsfor és la que queda retinguda a la matèria orgànica en forma d’humus i a les argiles (és a dir, al complex organicomineral del sòl). En el cas del nitrogen, la seva conversió en amoni per organismes descomponedors ajuda a aquesta retenció en forma de catió. Altres elements segueixen vies diferents, però el resultat és que l’ecosistema es mostra, en conjunt, estalviador dels nutrients més importants, almenys mentre no hi hagi excedents respecte de la capacitat de produir. S’ha pogut constatar que, en boscos vells en què ja no hi ha acumulació activa de nova biomassa, les aigües de drenatge són més riques en elements nutritius, que es perden així per al sistema. A més del nitrogen, l’oxigen i el diòxid de carboni, també pot entrar en forma gasosa el diòxid de sofre en certes quantitats. No obstant això, la major part dels elements que s’incorporen a l’ecosistema ho fan dissolts en l’aigua de la pluja o amb la deposició seca a través del vent.
Aigua
Un cop en contacte amb les fulles i les branques, l’aigua de pluja pot canviar la seva composició, sobretot perquè s’enduu per rentatge substàncies i ions que eren a les parts aèries de les plantes (procedents de la deposició seca o dels propis teixits vegetals respectivament). L’aigua arriba al sòl a través de la capçada (trascol) o lliscant pels troncs (escolament cortical). Els canvis químics continuen al sòl, principalment per intercanvi amb el complex col·loïdal i per absorció biològica.
Quan l’aigua surt de l’ecosistema, per escorriment o per infiltració profunda, té una composició molt diferent que quan hi entra, ja que una part dels nutrients que duia s’han incorporat al sòl o a la vegetació, i una altra ha passat, de les plantes, el sòl o la roca, a l’aigua. D’altra banda, les plantes absorbeixen nutrients del sòl per les rels i n’hi retornen amb la caiguda de virosta, la mortalitat de rels fines i, en part, per lixiviació i per sortida de substàncies orgàniques per les rels (exsudació). Els herbívors, els carnívors i els detritívors completen aquest retorn en forma de restes molt diverses, com després veurem. Tot això constitueix el cicle bàsic de la matèria dins l’ecosistema.
Aquest cicle bàsic respon a alguns principis essencials. En primer lloc, és clar, obeeix a la llei de conservació de la matèria. El que es treu d’un lloc es retroba en un altre. Els organismes presenten determinades composicions elementals, que poden presentar diferències importants entre espècies distintes o entre òrgans d’un mateix individu, sobretot entre les parts més actives i les de sosteniment. Els elements més importants són, amb diferència, el carboni, l’hidrogen i l’oxigen, el nitrogen, el fòsfor i el sofre, el calci, el potassi, el magnesi i, de vegades, el silici. Quan una planta fotosintetitza ha d’obtenir materials de fora per crear nous compostos. Per tant, la síntesi orgànica i el metabolisme poden estar limitats per la disponibilitat d’elements nutritius. Els elements usats pel vegetal poden passar als consumidors o tornar al sòl per a ser, altre cop, emprats en nous processos de síntesi. La capacitat de les espècies per obtenir els elements varia d’unes a les altres, i, això, determina diferències competitives entre elles i fa que unes puguin sobreviure en llocs on altres no poden fer-ho.
Els vegetals terrestres tenen una important característica química, lligada a l’existència d’una forta proporció de teixits de sosteniment i a la necessitat de respondre evolutivament a la pressió dels herbívors mantenint estructures durables. És tracta de l’abundància de polisacàrids, com la cel·lulosa, i de substàncies amb enllaços aromàtics, com la lignina i els tanins. Aquests materials són difícils de descompondre i mineralitzar, com també ho són els compostos estructurals dels animals, que conformen els esquelets (ossos, quitina), la qual cosa té una gran transcendència en els cicles biogeoquímics de l’ecosistema.
Un poderós sistema de control del transport permet a les plantes d’aprofitar pràcticament tota l’aigua en medis on escasseja. En un mateix indret, i sense modificar la fitomassa, pot ser que en un any sec tota o quasi tota l’aigua de pluja sigui evapo-transpirada (i no hi hagi, doncs, escorriment superficial ni gaire infiltració profunda), i que en un any humit, en canvi, l’escorriment i la infiltració al subsòl siguin importants. Les rels exploren el sòl, de vegades fins a fondàries o distàncies lineals molt considerables, de 15 o 20 m.
La capacitat de reciclatge i transport des de les rels és incrementada per la presència de micorizes, que augmenten molt la capacitat d’absorció de recursos edàfics. En alguns casos, com s’ha demostrat en selves plujoses tropicals, les hifes dels fongs connecten directament la virosta amb les rels, i són capaces de prendre els nutrients encara en forma orgànica, sense passar per una mineralització prèvia. Això escurça extraordinàriament el cicle i l’«interioritza» d’una manera completa en els organismes. De fet, aquesta forma de nutrició es dóna també en plantes sapròfites, com certes monotropàcies o orquidàcies. Per tant, es tracta de fenòmens que podrien tenir més importància de la que fins ara es pensava. S’ha dit que espècies de sòls molt pobres, com el pi roig (Pinus sylvestris) o la bruguerola (Calluna vulgaris), o espècies com l’avet roig (Picea abies), amb una capacitat extraordinària per absorbir nitrogen, podrien usar directament la virosta mitjançant les micorizes. En general, les micorizes són indispensables per a les coníferes, perquè substitueixen els pèls absorbents, que no hi havia en aquest grup, i incrementen la superfície d’absorció i l’espai explotat per les rels. El creixement de les micorizes és estimulat pel subministrament d’hidrats de carboni des de la planta i, en canvi, les hifes es mostren molt efectives en el transport i l’absorció de nitrogen i fòsfor. L’augment de la superfície absorbent en alguns casos pot ser de fins mil cops la que tindria la rel sense micoriza.
Els descomponedors
Virosta i descomposició
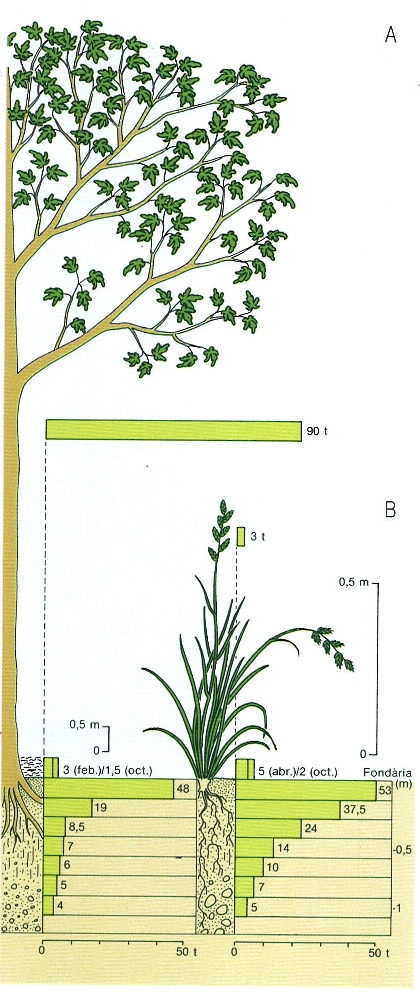
Distribució de la matèria orgànica en dos tipus d’ecosistemes terrestres, un bosc jove de caducifolis (A) i un prat (B), tots dos sobre sòls ben drenats i similars, de la zona temperada. S’han indicat la biomassa aèria en peu, la virosta i la matèria orgànica al sòl. Observeu la importància de la matèria orgànica al sòl en ambdós casos, i sobretot al prat, que fa que els valors totals siguin força semblants en un cas i altre (167 t en el cas del bosc, i 150 en el del prat). En el prat, la quantitat de matèria orgànica morta més la biomassa subterrània és quasi cinquanta vegades més gran que la biomassa aèria, i encara les rels en són una part mínima, de manera que quasi tot és matèria orgànica morta. Al bosc, molta matèria orgànica incorporada als arbres constitueix estructures de suport i transport, mentre que la biomassa aèria activa, la de les fulles, pot oscil·lar entre 3 i 6 t/ha, és a dir, té valors semblants a la del prat.
Mercè Cartañá, original de W. Foth.
Deixant, ara, de banda els casos en què pugui produir-se el curt circuit entre restes orgàniques i rels per les hifes, l’obtenció de nutrients per les plantes depèn de la presència d’ions inorgànics fàcils d’absorbir. Aquests ions procedeixen de la descomposició de la matèria orgànica morta o bé de la meteorització de la roca. El primer procés sol ser el més important en un ecosistema complex. No obstant això, l’estudi dels processos de descomposició al sòl presenta notòries dificultats.
S’ha fet la mesura de les «entrades» de restes de les parts aèries, és a dir, el que anomenem la caiguda de virosta al sòl, en un ampli conjunt d’ecosistemes de tot el món i s’ha pogut observar que hi ha una correlació positiva entre la caiguda de virosta i la producció primària. En un sistema equilibrat, tot el que cau correspon a materials que han de ser renovats (fulles, branquillons, esquames, fruits, llavors, etc.). Aquesta virosta no és, però, el total de les entrades de matèria orgànica morta al sòl. Hi ha també aportacions de les parts subterrànies, que poden ser molt importants (recordem l’elevada taxa de renovació de rels fines), com també sortida de substàncies orgàniques per les rels, i tot això és molt difícil de mesurar. També cal comptar amb els troncs i les soques d’arbres morts, els excrements, els cadàvers i altres restes (com les mudes d’insectes, els aliments no consumits totalment, etc.). La quantitat de matèria orgànica present en un moment determinat és el resultat de restar a la virosta preexistent en aquestes entrades la matèria orgànica que es va mineralitzant en els processos de descomposició. A cada ecosistema hi ha una evolució històrica d’aquest balanç d’entrades i sortides. En alguns casos, les quantitats presents de matèria orgànica morta poden arribar a ser molt considerables i, en el conjunt dels ecosistemes terrestres, es pot acceptar que n’hi ha fins deu cops més que no pas de biomassa, encara que entre els uns i els altres hi ha molta variació. En un alzinar, biomassa viva i matèria orgànica morta són de valors similars. No cal dir que molta d’aquesta matèria orgànica es troba en forma d’humus.
La massa de virosta present a la capa superior del sòl varia entre 1 i més de 50 tones per hectàrea. Dins de zones climàtiques semblants, la variació continua essent molt gran, fins al punt que es fa molt difícil de cercar relacions entre aquest caràcter i les diferències de tipus climàtic. La forta variabilitat en bona part és perquè les taxes de descomposició no són només influïdes pel clima, sinó que depenen molt de la qualitat de la virosta (per exemple, com més alta és la proporció de lignina, més difícil és la descomposició) i de la disponibilitat de nutrients. S’ha vist que, en general, les formacions perennifòlies acumulen masses de virosta superiors a les caducifòlies, perquè la virosta és de qualitat inferior i hi ha una major entrada subterrània, perquè les rels fines es renoven més de pressa.
L’activitat descomponedora es tradueix en una respiració global del sòl, que pot ser mesurada per l’alliberament de CO2. En una situació equilibrada, la respiració del sòl és aproximadament igual a la quantitat de CO2 assimilada per les plantes (producció primària bruta) menys les quantitats emprades per animals i plantes epigeus en llur respiració. Pot haver-hi algunes pèrdues per dissolució de CO2 en l’aigua de pluja, però, en conjunt, l’activitat respiradora del sòl és correlada amb la producció. Hi ha variacions temporals, perquè l’activitat microbiana és influïda per canvis d’humitat i temperatura. Això és un fet important en els sistemes mediterranis, en els quals sobretot la humitat pateix fortes fluctuacions. Els valors solen ser màxims a les estacions plujoses i també varien diàriament: són més grans de nit que de dia i en dies ennuvolats que en dies clars.
Els organismes del sòl
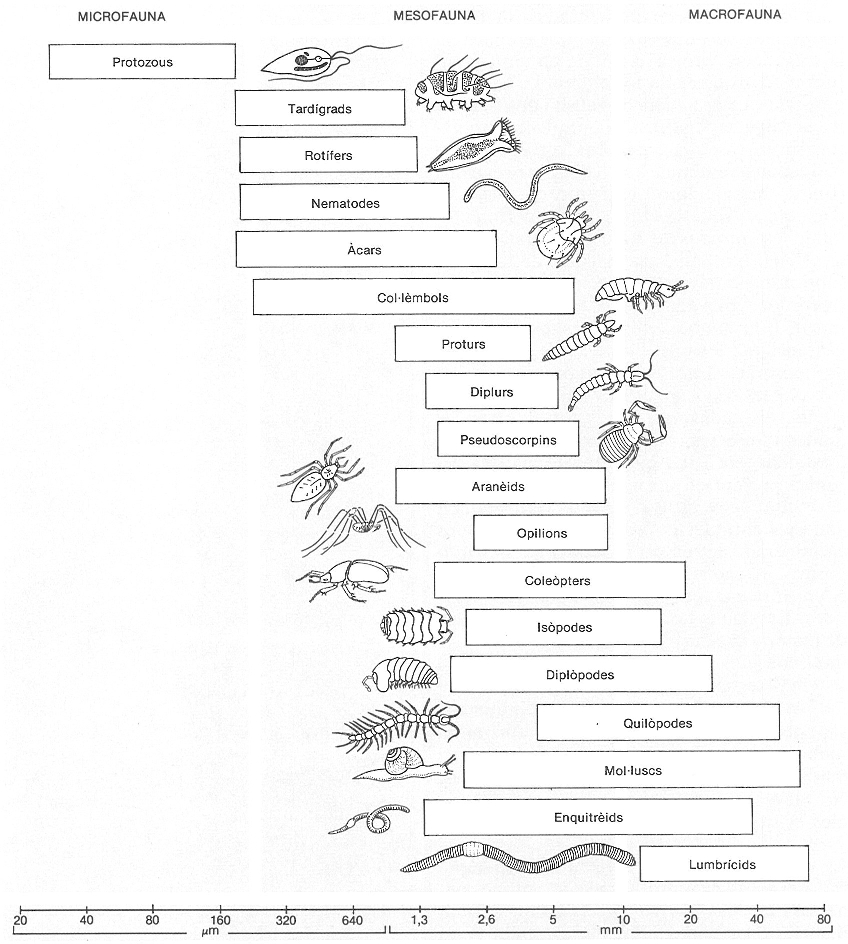
Fauna d’un sòl temperat, classificada per grandàries. És important veure l’ampla varietat de grups presents i entendre que l’esglaonament de dimensions correspon a una distribució de funcions en la degradació de matèria orgànica dins d’aquest subsistema heteròtrof, com també en les relacions de depredació i altres que apareixen dins el mateix subsistema.
Amadeu Blasco, original de J.A. Wallwork.
Al sòl no hi ha sols matèria orgànica morta, és clar; també s’hi troben organismes. Sense comptar les rels de les plantes, la biomassa d’organismes hi pot representar una proporció petita però sensible de la matèria orgànica total. En un prat de regions temperades que contenia un 4% de matèria orgànica, s’avaluà la biomassa viva sense les rels en un 20% d’aquesta matèria orgànica. Els grups majoritaris són sempre fongs i bacteris, però la proporció relativa dels uns i dels altres varia molt d’un lloc a l’altre; també s’hi troben protozous, els quals s’alimenten de formes diverses, però, sobretot, de bacteris incorporats a les partícules que ingereixen; nematodes que, com els anteriors, són essencialment aquàtics, tenen capacitat per a enquistar-se quan el sòl s’asseca, poden ser paràsits pràcticament de tots els animals i plantes i estan enormement diversificats; col·lèmbols, menjadors de restes vegetals; larves de dípters, també preferentment sapròfagues i capaces de barrejar les restes orgàniques amb el sòl; cucs de terra (anèl·lids), que són els que fan un major treball d’aquest tipus; també hi ha abundants àcars, miriàpodes, rotífers, tardígrads, mol·luscs, crustacis i vertebrats, i als sòls tropicals són molt importants els tèrmits.
La varietat d’estructures de la virosta, la presència d’aigua i la important entrada total d’energia permeten que al sòl hi hagi, en definitiva, un subsistema molt complicat i ric. Entre els artròpodes, a més dels sapròfags hi ha també una enorme varietat de depredadors, paràsits i hiperparàsits. Són també importants els que s’alimenten de micelis i d’estructures reproductives dels fongs, com és el cas de molts àcars oribàtids, abundantíssims a terra. Aquesta diversitat de grups té un significat funcional. Tot i que la biomassa principal correspon als descomponedors directes, la fauna edàfica té un paper prou important: modifica les propietats físiques, químiques, bioquímiques i biològiques del sòl, com ara la porositat, l’estabilitat estructural, la capacitat de retenció d’aigua, la velocitat dels processos d’humificació, la mineralització del nitrogen, la disseminació dels microorganismes, etc.
Els consumidors
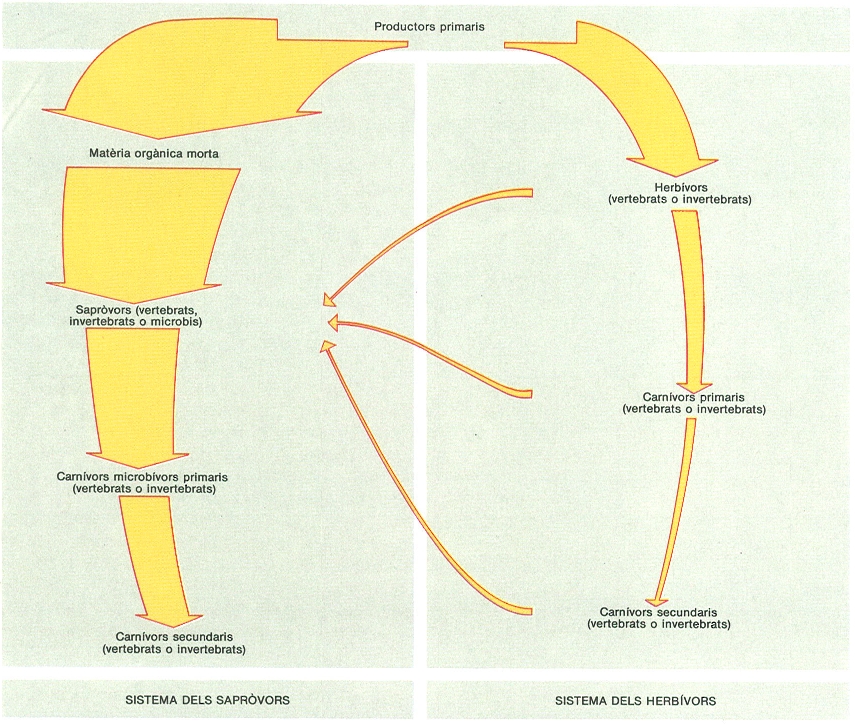
Doble via tròfica als ecosistemes terrestres. Encara que el model més difós presenta la cadena tròfica «productors primaris-herbívors-carnívors», als sistemes terrestres la major part de l’energia circula per un altre camí; els consumidors usen preferentment matèria orgànica morta, i tenim una cadena alternativa que anomenem sistema dels sapròvors. En molts sistemes terrestres, fins el 90% de l’energia captada pels vegetals segueix aquesta via. Això explica, en part, la importància dels processos que tenen lloc al sòl.
Amadeu Blasco.
Fins ara, ens hem ocupat de producció i descomposició. L’existència de processos de construcció i degradació és, evidentment, la base de tot funcionament, i amb aquesta manera d’introduir-nos als ecosistemes terrestres volem posar de manifest un aspecte molt característic del seu «metabolisme». L’energia captada pels productors primaris pot seguir dues vies diferents: un cicle més llarg, que dels herbívors passa per diversos nivells de carnívors i sapròfags i, finalment, arriba als descomponedors; i un segon cicle més curt, que va directament a consumidors de matèria orgànica morta (sapròfags) i a descomponedors. Normalment, als sistemes terrestres, entre el 75% i el 90% de la matèria orgànica incorporada no és ingerida viva per consumidors, sinó que passa a aquesta segona via, que anomenem detrítica. La proporció és molt diferent de la que es dóna als ecosistemes aquàtics, on els consumidors usen quasi la totalitat de la producció. Aquesta forma diferent de canalitzar el flux energètic a través del sistema va lligada al gran control que mantenen els vegetals al medi terrestre. Es cert que, a les pastures, els herbívors arriben a usar entre el 40% i el 60 % de l’energia fixada per les plantes, i l’home, en les seves explotacions, també n’aconsegueix proporcions elevades; però, en conjunt, la via detrítica, el cicle curt, és molt dominant.
En un bosc, el consum directe de biomassa viva realitzat pels herbívors sol ser inferior al 10% de la producció vegetal; la resta va a parar d’una forma o altra al sòl: fulles mortes, branques, troncs d’arbres morts, rels, tota la producció de flors, fruits i llavors (la majoria de les quals no donaran lloc a noves plantes), grans de pol·len, exsudats de rels, etc. Aquest flux enorme d’energia i materials en diverses formes justifica la complexitat del subsistema edàfic. En herbassars molt pasturats o en conreus els animals i l’home arriben a tenir força més control del sistema. Les plantes sotmeses a explotació no poden generar estructures tan complexes de sosteniment i de transport, tenen més parts actives amb relació al pes total, més producció amb relació a la biomassa i una taxa més alta de renovació. En aquests casos, pot donar-se una considerable organització i diversificació dels herbívors, que exploten els recursos productius de l’herbassar amb una fina divisió de funcions. És el cas de les sabanes africanes, els «llanos» veneçolans, les grans praderies nord-americanes i la pampa argentina.
Herbivorisme
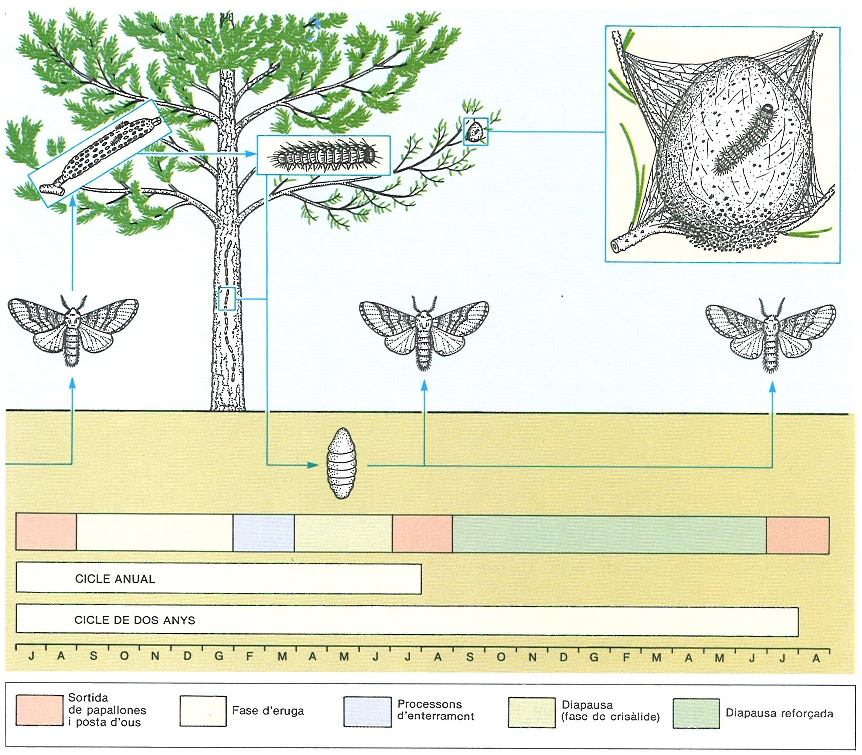
Cicle de vida de la processionària del pi (Thaumetopoea pityocampa). Aquesta papallona provoca considerables pèrdues a les pinedes i ataca especialment algunes espècies exòtiques, com Pinus radiata, però també pins indígenes, sobretot en anys o llocs particularment difícils. Molts aspectes de la biologia d’espècies que produeixen plagues, de llurs enemics naturals, dels productes químics que s’empren per a combatre-les (insecticides, fungicides, feromones atractives, etc.) i els etectes que tenen sobre l’entorn són encara per estudiar, i demanen un considerable esforç de recerca.
Mercè Cartañá, a partir de fonts diverses.
Els herbívors presenten en els ecosistemes terrestres, malgrat el que acabem de dir sobre llur importància, relativament limitada en molts casos, una àmplia diversitat de formes i prou motius d’interès. Artròpodes, alguns cucs, mol·luscs, ocells i mamífers són els grups més importants, per no dir els únics, d’herbívors i consumeixen des de l’estrat muscínic fins als arbres, des dels sucs cel·lulars fins als fruits. La importància dels herbívors no pot mesurar-se només per la quantitat d’energia que canalitzen. Els rosegadors, per exemple, tenen una forta influència en el sistema per llur activitat excavadora, amb la construcció de túnels (quasi 8 km/ha en una roureda mixta estudiada a Rússia) i la renovació de la terra (prop de 3 m3/ha excavats en el mateix bosc). Aquesta activitat facilita el drenatge de l’aigua de pluja i l’aireig, pot alterar l’estructura i la composició del sòl remogut i ajudar a la formació de mull. El consum de llavors, en principi, dificulta la regeneració del bosc. Però això no sempre és tan senzill. Les glans de roures i d’alzines només germinen si han estat prèviament enterrades, i el costum d’alguns animals, com el gaig i diversos rosegadors, d’enterrar-les per tenir reserves emmagatzemades d’aliments és un ajut a la germinació de les que després no són consumides per una raó o altra.
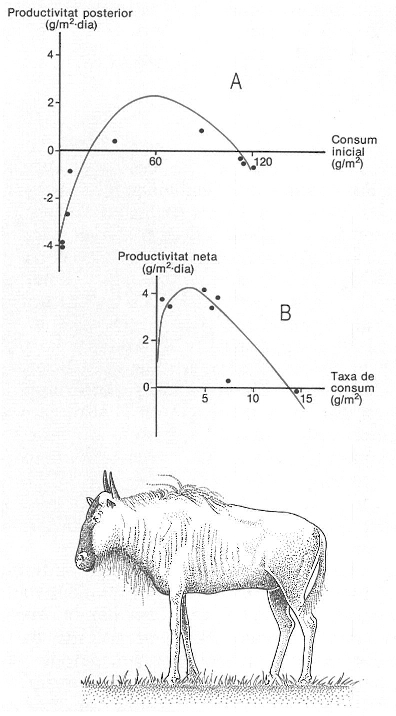
L’herbivorisme pot actuar com un estímul per a elevar la producció primària d’un sistema. El dibuix que serveix d’exemple s’ha fet amb dades procedents d’un estudi de McNaughton a les sabanes del Parc Nacional del Serengeti (Tanzània), però s’obtenen resultats semblants a les pastures de zones temperades, per l’efecte dels ramats domèstics. A Evolució de la productivitat després del pas d’un ramat de nyús, segons el consum inicial. B Increment de la productivitat neta d’acord amb la taxa de consum. Ambdues corbes mostren un màxim quan els valors de consum són intermedis.
Amadeu Blasco
Els invertebrats fitòfags solen associar-se estretament amb plantes específiques: al més sovint s’alimenten d’una o de poques espècies, i només més rarament són polífags. Fins i tot s’especialitzen a viure sobre parts determinades del vegetal hoste, sigui l’escorça, el tronc, la capçada o el sistema radical. Les poblacions d’insectes són sotmeses a control per depredadors, com són altres insectes, ocells, etc. S’ha destacat en aquest sentit, per exemple, el paper de la formiga roja als boscos centreuropeus. Les relacions entre les dinàmiques poblacionals dels insectes fitòfags i llurs depredadors són un camp d’estudi apassionant, però ací ens és impossible de deturarnos-hi.
Alguns invertebrats produeixen plagues capaces de destruir boscos sencers o de menjar-se’n totes les fulles, que tenen efectes profunds en el metabolisme de tot el sistema. Durant una plaga massiva d’una eruga menjadora de fulles, com la de l’eruga de l’alzina (Lymantria dispar), s’altera la fisiologia de la planta, la qual cosa es reflectirà després en les característiques químiques i morfològiques de les fulles formades a la rebrotada següent; es modifiquen, també, les relacions entre la planta i el sòl i, de retruc, algunes característiques d’aquest, i s’altera el microclima en destruir-se les fulles. De cop es produeix, també, un enorme increment de l’aliment disponible per a depredadors i paràsits dels insectes plaga. Sembla que aquesta alteració fisiològica en alguns arbres pot ser detectada ja en arbres veïns, els quals modifiquen també el seu metabolisme per a produir substàncies defensives i fulles amb proteccions especials (més coriàcies, dentades, etc.). Com a resultat d’una llarga evolució conjunta, les plagues ataquen més fàcilment els vegetals debilitats, que tenen menys respostes defensives. Això fa que, sovint, les causes de mortalitat en els vegetals se superposin i es confonguin. Moltes plagues forestals, com les d’escolítids, actuen com a causa secundària, i apareixen en condicions d’estrès provocades, per exemple, per un llarg període d’eixut o per un problema de contaminació.
Els vertebrats poden actuar diferentment segons la disponibilitat d’aliments, els canvis d’estat successional, les condicions ambientals, etc. Poden consumir fruits, fulles, branquetes, escorça, plantes senceres, etc., i la dieta es pot modificar d’acord amb els recursos que es trobin més abundants en cada moment. És conegut el cas de l’elefant, que estacionalment completa la proporció de fibres de la seva alimentació menjant escorces, que arrenca amb els ullals, dels troncs de les acàcies; produeix, així, ferides als arbres, que obren el pas a coleòpters perforadors i a infeccions micòtiques, i són causa de mortalitat.
L’acció dels herbívors sobre les pastures ha estat objecte de molts estudis. En general, el consum moderat sembla determinar un increment subsegüent de la producció primària i una inhibició de la floració (ja que la planta dedica els recursos assimilats a les necessitats de creixement, desviant-les de les de reproducció). Els pics de producció poden ser molt considerables. Al Parc Nacional Serengeti (Tanzània), s’han mesurat valors situats entre 20 i 40 g/m2·dia. En plantes no pasturades, es produeix, després de la maduració de les llavors, una senescència foliar, s’emmagatzemen reserves a les rels i hi ha altres translocacions. La pastura inverteix aquests processos i estimula el creixement de les tiges; però si és massa intensa, la producció decau. Naturalment, els nivells de consum que estimulen la producció depenen de les condicions ambientals i varien marcadament amb les estacions de l’any.
L’estímul de la producció és determinat per diverses causes. La taxa de fotosíntesi és inhibida, si no hi ha consum, per l’acumulació de productes fotosintètics de reserva, com el midó, en fulles actives. A més, es produeix inhibició, també, per l’ombra entre fulles. Un punt notable és que les fulles de les gramínies creixen des de la base, per meristemes intercalars, i, com que el consum és des de dalt, s’eliminen els teixits més vells i menys eficients. L’equilibri hormonal es modifica amb la defoliació i es produeix més flux d’hormones de creixement als meristemes residuals. La pèrdua d’àrea foliar redueix la transpiració. Els nutrients procedents dels excrements i l’orina dels animals estimulen el creixement. Finalment, s’ha comprovat que la saliva dels remugants té efectes directes d’estímul del creixement.
S’han fet estudis molt interessants de la manera com es reparteixen els recursos els diferents herbívors del Parc Nacional Serengeti (un dels millors exemples dels ecosistemes amb més diversitat d’espècies de grans herbívors). Cadascun té una estratègia pròpia d’alimentació, i això els diferencia ecològicament i permet que puguin conviure. L’hàbit migratori o resident de cada espècie té també un paper molt important en aquesta diferenciació ecològica, ja que les espècies es reparteixen l’ús dels recursos en l’espai i en el temps. Usant diversos tipus de fulles o parts de fulles (més verdes, més seques, etc.) i canviant de lloc en migracions esglaonades, els herbívors poden ser altament diversificats.
Entre els mamífers herbívors hi ha una distinció important a fer, segons que siguin remugants o no. L’estratègia del remugant es basa en una alta eficiència en l’ús de les proteïnes, però, per contra, al remugant li cal ingerir i processar grans quantitats d’aliment i seleccionar-ne components vegetals rics en proteïnes. Els no remugants, com ara el cavall, són menys eficients en l’ús de les proteïnes, però el material passa dos cops més ràpid per l’estómac, la qual cosa els permet d’emprar una dieta més pobra en proteïnes de la que podrien acceptar els remugants. El cavall, la zebra o el nyu són molt menys selectius que el bou, l’ovella, el búfal o el bisó.
Carnivorisme
L’estratègia dels carnívors és, en general, ben diferent de la dels herbívors. Les característiques dels aliments són, en primer lloc, molt diferents. El carnívor pren un aliment molt més fàcil d’assimilar, cosa que comporta un tub digestiu proporcionalment molt més curt. La depredació pot tenir lloc per mètodes diferents, que van des de la caça, relativament més passiva, mitjançant trampes (la teranyina, per exemple), fins a la persecució i mort de la presa, sia per depredadors individuals o en grups (com fan els llops). Alguns viuen d’animals morts, com fan els carronyers (necròfags). Per extensió, també podem incloure, en aquesta categoria general dels carnívors, els animals hematòfags (com els mosquits) i diverses menes de paràsits dels animals. Cada una d’aquests variants, i d’altres que podríem trobar, dóna lloc a un conjunt especial d’adaptacions per a cada fase de la depredació, des de la recerca o l’espera de la presa fins a la seva ingestió i digestió, passant per la persecució i la captura. L’exposició d’aquestes adaptacions donaria lloc a tot un tractat d’història natural.
Des del punt de vista energètic, també ací podem adoptar una perspectiva econòmica. El depredador ha d’obtenir suficient retribució alimentària de la presa per a compensar tota l’energia que li ha calgut gastar en la depredació. Una presa més gran vol dir més energia, més aliment, però també costa més de capturar i és menys abundant (cal buscar més), i d’altra banda, la caça amb trampes no pot aplicar-se a preses poc abundants. Mida i nombre de les preses, doncs, són factors clau per a definir l’estratègia del depredador. Pensem, per exemple, en les diferències entre dos animals voladors homeoterms, les rates-pinyades, que s’alimenten d’insectes que cacen al vol localitzant-los, durant la nit, gràcies al sonar de què estan dotades, i les àguiles o els falcons, que mengen molt menys sovint animals molt més grossos. Si les preses són grans, el depredador ha de moure’s per un territori molt extens, adoptar una forma de vida molt activa i gastar molta energia en l’activitat de recerca i captura de l’aliment. Aquestes tendències solen conduir a un control molt més gran del sistema i a tenir més facilitat per a fugir de les fluctuacions de l’entorn i per a mantenir poblacions més constants. Però també és cert que les densitats de població són baixes i això, juntament amb la destrucció d’hàbitats i preses o la del mateix depredador per l’home, posa els grans depredadors i carronyers al límit de llur desaparició.
Els carronyers, i no diguem els paràsits, poden esdevenir més petits que llurs preses. No obstant això, alguns carronyers són molt grossos, com és el cas dels voltors, i alguns depredadors grossos adquireixen fàcilment hàbits carronyers, com és l’ós polar. L’atac a un cadàver gros és, però, quasi sempre l’acció combinada de diversos necròfags (ocells, mamífers i insectes). En la depredació, de vegades la presa s’ingereix sencera (com fa una granota en empassar-se un mosquit); però sovint no és així i en queden restes (de vegades la presa és regurgitada per la boca sense passar per l’intestí) que serviran d’aliment a comensals dels depredadors o a altres necròfags. Des d’un punt de vista energètic, depredadors i carronyers presenten poques diferències; però l’acció dels uns i dels altres és molt diferent en la regulació de les poblacions de les espècies que els serveixen d’aliment. En aquest sentit, la depredació té un paper molt rellevant. En el medi terrestre, les formes de vida carnívora estan repartides a quasi tots els grups filogenètics. Deixant de banda els microorganismes, i l’enorme varietat de paràsits pluricel·lulars, són carnívors, necròfags, copròfags o hematòfags terrestres els aràcnids, molts insectes dels diferents ordres, adults o en fases larvals, quilòpodes i diplòpodes, amfibis, rèptils (tots, menys les tortugues), molts ocells i mamífers.
Les relacions de mutualisme
Els sistemes naturals generalment ofereixen una tossuda resistència als intents d’imposarlos esquemes de classificació. No obstant això, en el cas de les interaccions entre organismes, els ecòlegs van establir, ja fa moltes dècades, una classificació que amb lleugeres variants i alguna discrepància s’ha mantingut inalterada fins avui. Segons aquesta classificació, els organismes poden interactuar de tres maneres fonamentals, segons els efectes (positius o negatius) que la interacció tingui per a cada participant. En les relacions de competència entre dues espècies d’organismes, la interacció té conseqüències negatives per a ambdós participants. En les de predació, l’efecte és negatiu per a un d’ells (presa) i positiu per a l’altre (depredador). El tercer tipus d’interacció és el mutualisme, en què ambdós participants obtenen un benefici. En resum, el mutualisme pot definir-se com una interacció +/+, mentre que la competència i la predació serien, respectivament, −/− i −/+ (per completar aquest esquema podríem incloure el comensalisme, que seria una relació de tipus +/0).
Dels tres tipus fonamentals d’interacció que s’acaben de descriure, la competència i la predació han estat estudiades amb profunditat durant el segle actual. En canvi, el mutualisme, encara que mai no ha estat completament absent del pensament ecològic, ha rebut una atenció ben marginal, essent considerat per alguns autors com una relació tròfica de tipus col·lateral.
A la conca mediterrània, els sistemes naturals actuals són segurament només un pàl·lid reflex dels que existiren abans que l’activitat humana no hi reduís dràsticament l’extensió dels boscos limitant-los a petits retalls aïllats, n’ocupés les terres més fèrtils per a aprofitaments agrícoles i usés el foc com a arma enfront de la vegetació. És de suposar, per tant, que la diversitat biològica, i amb ella la riquesa d’interaccions entre espècies, ha experimentat una notable degradació en els darrers mil·lennis. Tot i així, els ecosistemes terrestres mediterranis constitueixen encara un camp privilegiat per a l’estudi dels fenòmens de mutualisme, sobretot en aquelles àrees en què han estat menys alterats per l’acció humana. Algunes d’aquestes relacions han estat ja estudiades amb un cert detall a la península Ibèrica, com ara les interaccions entre algunes plantes i els animals que les pol·linitzen o dispersen llurs llavors. Altres relacions encara no han rebut gairebé l’atenció dels investigadors com són les que s’estableixen entre certes plantes i les formigues que disseminen llurs llavors o defensen llurs flors, les relacions entre formigues i pugó, formigues i larves de lepidòpters, plantes i fongs (micorizes), flora microbiana i remugants, insectes micòfags i fongs superiors, formigues i plantes amb nectaris extraflorals, etc.
La pol·linització de les flors pels insectes
La gran majoria de les plantes llenyoses i bona part de les herbàcies dels ecosistemes ibèrics són pol·linitzades per insectes. Excepcions notables són les coníferes (pins i avets) i moltes de les frondoses (faigs, alzines i roures), en les quals el transport de pol·len és fet pel vent (anemofília). Malgrat aquestes excepcions, les espècies pol·linitzades per insectes (entomòfiles) predominen en la majoria de localitats mediterrànies. Per exemple, la proporció d’espècies llenyoses que són entomòfiles és del 83% al «matorral» del Parque Nacional de Doñana, del 70% en una localitat de la serra de Cazorla (Jaén) a 1100 m d’altitud, i del 86% en una localitat de la baixa vall del Guadalquivir; a aquest grup pertanyen, per esmentar alguns exemples familiars, les espècies de cistàcies, labiades, caprifoliàcies i ramnàcies.
Parlant en termes d’economia humana, en la relació de mutualisme entre la planta i el pol·linitzador s’intercanvia un «bé» per un «servei». El servei a la planta és donat pel pol·linitzador: es tracta del transport de pollen d’una flor a l’estigma d’una altra flor de la planta. El bé el dóna la planta al pol·linitzador, generalment en forma d’aliment. El nèctar floral n’és potser la modalitat més coneguda, però no l’única. De fet, nombroses plantes entomòfiles mediterrànies no secreten nèctar o ho fan en quantitats mínimes. En aquests casos, el mateix pol·len, produït en abundància, és el principal «premi» del pol·linitzador. Comparades amb comunitats vegetals d’altres latituds, sembla que les plantes entomòfiles mediterrànies es caracteritzen, en general, per una incidència més gran de les espècies que ofereixen sobretot pol·len com a premi a llurs pol·linitzadors.
Són característiques d’aquest grup les espècies de cistàcies (estepes i heliantems) i moltes fabàcies (ginestes, ginestells i gatoses, dels gèneres Cytisus, Genista, Ulex) i ericàcies, com el bruc (Erica arborea). En contrast amb aquest grup, les espècies de lamiàcies, caprifoliàcies, ramnàcies, timeleàcies i altres ericàcies com l’arboç (Arbutus) o la bruguerola (Calluna) donen nèctar als pol·linitzadors per pagar llurs serveis. Una característica del nèctar de la majoria d’aquestes plantes, sobretot de les labiades, és l’elevada concentració de sucres, si es compara amb la de plantes d’hàbitats més septentrionals o, fins i tot, del tròpic sec. Són habituals, per exemple, concentracions entre el 60% i el 75% (pes de sucre/pes de dissolució) per al barballó (Lavandula latifolia) o el romaní (Rosmarinus officinalis) en determinades dates i determinats llocs. Es coneix de temps la influència de les disponibilitats hídriques sobre la concentració de nèctar. El dèficit hídric crònic que el clima mediterrani imposa a les plantes, unit a una intensa evaporació causada per temperatures elevades, pot servir per a explicar l’existència d’aquests nèctars extremament concentrats. Teòricament, els nèctars molt concentrats tenen una rendibilitat energètica molt alta per als insectes que els consumeixen, però llur extracció comporta despeses també molt altes a causa d’increments desproporcionats de la viscositat a mesura que la concentració augmenta. Hom pot esperar, per tant, que l’ús dels nèctars molt concentrats i viscosos d’algunes plantes mediterrànies demani certa especialització dels insectes que les consumeixen.
Segons el seu sistema reproductiu, plantes d’espècies diferents depenen de forma també diferent dels serveis dels pol·linitzadors. Les espècies dioiques, en què les flors masculines i femenines són produïdes per individus diferents, o les espècies amb flors hermafrodites, però que tenen algun mecanisme d’incompatibilitat, necessiten per força la intervenció d’un agent pol·linitzador per a la producció de les llavors. Fins i tot les espècies de flors hermafrodites sense mecanismes d’incompatibilitat demanen l’activitat d’un pol·linitzador per produir llavors si l’autogàmia espontània no es produeix amb freqüència. Els sistemes reproductius de les plantes ibèriques no s’han començat a estudiar amb detall fins fa poc temps, però el coneixement preliminar que en tenim actualment permet d’assenyalar diverses característiques interessants des del punt de vista de la relació mutualista planta-pol·linitzador en sistemes naturals mediterranis. En moltes formacions vegetals, una proporció apreciable (25-30%) de les espècies llenyoses són dioiques; quasi tota la resta són hermafrodites (la proporció d’espècies monoiques i ginodioiques és molt baixa). Tot i que encara no en sabem prou per a generalitzar sobre els sistemes reproductius de les hermafrodites, es pot dir que una família tan característica com la de les cistàcies té mecanismes d’autoincompatibilitat força estrictes, i que moltes espècies de lamiàcies (per exemple, l’espígol i el romaní), tot i que són autocompatibles, no mostren autogàmia espontània. La impressió que s’obté en considerar tota aquesta informació és que en les formacions vegetals mediterrànies predominen les espècies que depenen forçosament dels insectes per a produir llavors, és a dir que, lluny de ser un mutualisme facultatiu (del qual hom pot prescindir), la relació de moltes plantes amb el conjunt de llurs pol·linitzadors és de tipus obligat (per a produir llavors). No és aquesta l’aplicació habitual del concepte de mutualisme obligat, que se sol reservar per a situacions en què un mutualista no pot sobreviure sense l’altre. Però en termes evolutius, la «fitness», és a dir, l’eficàcia reproductora d’una planta que no produeix llavors, és idèntica a la «fitness» d’una planta morta, i per aquest motiu una planta que necessiti interaccionar amb una sèrie d’insectes per a produir llavor mor evolutivament si aquesta interacció desapareix. (Quina fóra la «fitness» d’un individu d’una espècie entomòfila dioica que cobríssim durant totes les èpoques de floració de la seva vida amb una malla fina que no deixés passar els insectes?)
Hi ha nombrosos grups d’insectes implicats en relacions de pol·linització amb les plantes mediterrànies. Els més destacats són, per ordre decreixent d’importància, abelles (àpids; totes les famílies), mosques (dípters; diverses famílies, sobretot sírfids i bombílids), papallones (lepidòpters ropalòcers; totes les famílies), escarabats (coleòpters; nombroses famílies, però sobretot escarabèids, dasítids, mordèlids i malàquids) i volianes (lepidòpters heteròcers; esfíngids, noctúids). De tots, les abelles són sens dubte el grup que més depèn dels recursos recol·lectats en les flors i el que ha evolucionat en relació més estreta amb les plantes. Tant les abelles socials (àpids) com les solitàries (megaquílids, halíctids, andrènids, antofòrids) depenen inexcusablement del pol·len i del nèctar per a reproduir-se, ja que aquest és llur aliment en la fase larval. En certa manera, per tant, també les abelles com a grup tenen un mutualisme obligat amb la flora.
La conca de la Mediterrània és una de les regions del món més riques en espècies d’abelles, fins i tot més que el tròpic humit. Tot i no haver-se’n fet estudis exhaustius ni recol·leccions detallades, la fauna ibèrica d’abelles recollida al catàleg de Ceballos totalitza 1043 espècies, la xifra més alta de tots els països europeus. En l’àmbit mundial, aquesta diversitat només és equiparable a la que hi ha a l’estat nord-americà de Califòrnia (1985 espècies) i a tot el continent australià. La intensa especiació que ha tingut lloc entre les abelles de la Mediterrània occidental no sembla relacionada amb la diversitat florística regional, sinó més aviat amb característiques climàtiques i cdàfiques especialment favorables. Independentment del seu origen, això no obstant, aquesta fauna d’abelles tan rica en espècies persisteix gràcies a les relacions de mutualisme que les abelles mantenen amb les plantes de les quals s’alimenten i que, alhora, pol·linitzen.
Entre tots els restants grups d’insectes pol·linitzadors, només els dípters bombílids s’acosten a les abelles pel que fa a llur nivell de dependència respecte dels recursos florals. Els insectes adults d’aquest grup, tant de les espècies amb peces bucals llargues (consumidores de nèctar) com de les que tenen peces bucals curtes (consumidores de pol·len), són visitants freqüents i pol·linitzadors de les flors de moltes espècies. En llur fase larval, totes les espècies són paràsites en els nius d’altres insectes, segons sembla sobretot en els nius d’abelles solitàries (amb subministraments de nèctar i pol·len). En certa manera, doncs, aquest grup de dípters depèn també, per a la seva reproducció, dels recursos florals, encara que sigui mitjançant una relació de parasitisme. La seva diversitat específica és, això no obstant, molt més modesta que la de les abelles, ja que els bombílids només tenen unes 200 espècies a tota la península Ibèrica.
La dispersió de les llavors pels vertebrats
Com en el cas de la pol·linització, també la relació mutualista planta-animal que gira al voltant de la dispersió de llavors es fonamenta en l’intercanvi d’un bé per un servei. La planta produeix un fruit carnós (baia, drupa, etc.) que un animal (ocell, mamífer) usa com a aliment. Després d’ingerir el fruit sencer (és a dir, la polpa més les llavors del seu interior), l’animal digereix la polpa i n’extreu els nutrients; després expulsa les llavors (per defecació o regurgitació) intactes i en condicions adients per a la germinació, a certa distància de la planta mare. El benefici que obté l’animal d’aquest sistema és evident: tots els fruits tenen a la polpa certa quantitat d’hidrats de carboni, lípids, proteïnes minerals o vitamines que són de valor immediat per a qualsevol consumidor. Els avantatges per a la planta són menys evidents o més difícils de quantificar, però tenen a veure amb el fet que la llavor sigui allunyada de la planta mare; aquest allunyament pot ser favorable com una manera d’escapar d’un risc quasi cert de predació, o per a reduir la competència per llum i nutrients que es desencadenaria entre la mare i les plàntules germinades sota seu. Altres avantatges suplementaris poden ser un increment del flux genètic i una possibilitat més gran de colonitzar nous sectors de l’hàbitat, tot i que aquests són més difícils d’encaixar en un model purament darwinista de selecció. Des del punt de vista funcional, són equivalents a fruits tota una ampla gamma d’estructures que comparteixen la qualitat d’estar integrades per una o més llavors acompanyades d’estructures accessòries de tipus nutritiu. S’inclouen ací, per exemple, els «fruits» de gimnospermes (teix, savines i ginebres, èfedres), en els quals la part carnosa activa deriva de bràctees modificades; els de moltes rosàcies, en què la part carnosa deriva del receptacle floral (la maduixa, el cinoròdon del roser); o el cas del fruit que representa realment un agregat de fruits individuals (la móra dels esbarzers).
A la península Ibèrica, prop de 200 espècies de plantes, pertanyents a gairebé 40 famílies, produeixen fruits carnosos i llurs llavors són disseminades per vertebrats (espècies endozoòcores); pel que fa al nombre d’espècies, les famílies més importants són rosàcies, caprifoliàcies, liliàcies i ramnàcics. En el conjunt de la flora ibèrica, les espècies dispersades per vertebrats representen una proporció relativament petita; però en comunitats vegetals locals, llur importància és normalment considerable, ja que representen entre el 40% i el 70% del nombre total d’espècies llenyoses i entre el 50% i el 80% del recobriment total. Pertanyen a aquest grup, per exemple, les espècies d’aladerns (Phillyrea), arboç (Arbutus), llentiscles (Pistacia), savines i ginebres (Juniperus), ullastres (Olea), arços (Cralaegus) i rosers (Rosa).
En formacions vegetals de tipus mesofític (fagedes, rouredes), les plantes endozoòcores són escasses a l’interior del bosc madur i es concentren sobretot en clarianes naturals, pertorbacions i vores; es tracta, doncs, generalment d’espècies relativament pioneres que desapareixen en progressar la successió. En les formacions mediterrànies, al contrari, les plantes endozoòcores són abundantment representades en els estadis successionals avançats, mentre que les espècies pioneres són dispersades pel vent o no tenen mecanismes especials de disseminació. Aquests patrons successionals són, al capdavall responsables de l’elevat recobriment proporcional de les espècies dispersades per vertebrats a les comunitats vegetals mediterrànies poc alterades que abans hem esmentat. En termes de la relació mutualista planta-animal, aquest patró successional té una traducció immediata: als boscos mesofítics «temperats», el mutualisme per a la dispersió de llavors és un procés espacialment localitzat, que té lloc en clapes intersticials, aïllades, de natura generalment efímera, mentre que pràcticament no n’hi ha a l’interior del bosc; a les formacions mediterrànies no gaire pertorbades, en canvi, la relació es produeix de forma extensiva al llarg d’amples superfícies de terreny, sense estar limitada a clapes localitzades, i les úniques discontinuïtats existents són precisament les clapes produïdes per pertorbacions, on s’estableixen ràpidament les espècies pioneres no endozoòcores.
Ocells i mamífers són els principals agents dispersadors de les llavors de plantes endozoòcores. Entre els mamífers, els frugívors i dispersadors de llavors més importants pertanyen a l’ordre dels carnívors, encara que sembli paradoxal. Les espècies més importants actualment són la marta (Martes martes), la fagina (M. foina), el gat mesquer (Genetta genetta) i la guineu (Vulpes vulpes), però no és difícil d’imaginar la importància que tingué l’ós (Ursus arctos) abans que les seves poblacions fossin virtualment liquidades. L’ecologia alimentària d’aquest grup, i particularment el seu frugivorisme, és menys coneguda que la dels ocells frugívors, especialment en hàbitats on ni llurs densitats poblacionals ni el recobriment vegetal no han estat modificats per l’actuació humana. Hi ha evidències que, en aquestes circumstàncies «naturals», el frugivorisme de certs carnívors és quelcom més que una activitat conjuntural i esporàdica: en certs llocs la fagina i la guineu són predominantment frugívors cinc mesos a l’any; en aquest temps, dispersen les llavors de dues dotzenes d’espècies vegetals i en són els dispersadors principals o quasi exclusius de la meitat. En el cas de la dispersió de llavors per ocells, n’hi ha prou amb unes poques desenes d’hectàrees de vegetació acceptablement conservada per a comprendre la dinàmica i els mecanismes del sistema de mutualisme planta-dispersant. Hom creu que la relació de mutualisme planta-carnívor fou extraordinàriament important per a totes aquelles espècies vegetals que produeixen fruits relativament grossos prop del sòl o que cauen després de madurar, que fan una olor apreciable per al nostre olfacte de mamífers, que d’alguna manera defensen de les dents llurs llavors i que tenen fruits que avui es podreixen al sòl dels molts llocs on els carnívors han estat delmats o exterminats, com per exemple, el margalló (Chamaerops humilis), el cirerer d’arboç (Arbutus unedo), Malus sylvestris, i el server (Sorbus domestica).
Entre els ocells frugívors i dispersadors de llavors més significatius hi ha diverses espècies de Turdus, sobretot la merla (T. merula), el tord comú (T. philomelos) i l’ala-roig (T. iliacus), i de tallarols, principalment el tallarol de casquet (Sylvia atricapilla), el gros (S. borin) i el capnegre (S. melanocephala), com també el pit-roig (Erithacus rubecula). Altres espècies, com el mastegatatxes (Ficedula hypoleuca) o la cotxa cua-roja (Phoenicurus phoenicurus) solen fer un paper molt secundari respecte de les anteriors. Tots aquests ocells ingereixen fruits sovint i defequen o regurgiten les llavors intactes, de manera que són autèntics mutualistes. El tallarol de casquet dispersa les llavors de més de 40 espècies diferents de plantes. Entre els ocells frugívors es troben espècies més o menys sedentàries com ara la merla, hivernants com són el tord comú o l’ala-roig i migradores de pas com és el tallarol gros. Per això, i perquè la producció de fruits per les diferents espècies de plantes té lloc al llarg d’una part important de l’any (de juny a febrer), l’època de fructificació d’una planta determina en bona mesura quins seran els seus dispersadors. La majoria de les espècies maduren llurs fruits entre setembre i novembre, i en moltes els fruits madurs persisteixen tot l’hivern. Per això, els frugívors hivernants i els migradors de tardor solen ser els dispersadors més importants a la majoria de llocs. No obstant això, hi ha importants variacions altitudinals relacionades tant amb diferències en la fenologia de les plantes com en l’estacionalitat de les comunitats d’ocells.
Pel que fa al nombre d’espècies, els frugívors són generalment un grup minoritari en les nostres comunitats d’ocells; tanmateix, en termes de nombre d’individus sovint són majoria, ja que les espècies frugívores tendeixen a tenir abundàncies mitjanes més grans que les insectívores o granívores dels mateixos hàbitats. Una de les causes d’aquest fet és l’abundant disponibilitat de fruits durant bona part de l’any, particularment a la tardor i a l’hivern. En hàbitats mediterranis poc alterats, les plantes produeixen anualment entre 100 000 i 3 000 000 de fruits per hectàrea (10-300 fruits/m2), la qual cosa representa de 15 a 400 kg/ha (en pes fresc) d’un recurs alimentari fàcil d’obtenir, conspicu i sovint altament nutritiu. El contingut en lípids de la polpa dels fruits d’algunes espècies arriba a ser superior al 30% del pes sec; per exemple, el llentiscle (Pistacia lentiscus), la noguerola (P. terebinthus), l’ullastre (Olea europea var. silvestris) o el llorer (Laurus nobilis). La gran disponibilitat d’un recurs molt rendible explica no solament l’abundància dels ocells frugívors, sinó també alguns aspectes de llurs hàbitats migratoris i de llur fisiologia digestiva.
Tot i que encara és difícil d’avaluar de forma general, podem afirmar que la importància del mutualisme planta-dispersador per a cada participant és més petita que en el cas de la relació planta-pol·linitzador. Des de la perspectiva de la planta, no produir llavors fa la seva «fitness» igual a zero, però deixar de dispersar-les, encara que pugui tenir un efecte negatiu molt pronunciat, no anul·la necessàriament la seva «fitness». En el cas dels frugívors, encara que cap espècie no usi els fruits per a alimentar les seves cries, aquest recurs alimentari té una gran importància per a la supervivència dels adults en certes èpoques de l’any, tant entre els mamífers com entre els ocells. En el cas d’aquests darrers, l’acumulació de greix subcutani que fan a l’hivern i abans dels viatges migratoris, tan important per a llur supervivència, és facilitada per l’elevat contingut de lípids de molts dels fruits que consumeixen.
Mutualisme i funcionalitat de l’ecosistema
Prenent com a exemple les relacions de pol·linització i dispersió de llavors, intentarem de fer més explícita la idea que les relacions de mutualisme constitueixen un element important en la xarxa d’interaccions que formen la base de la funcionalitat de l’ecosistema. Considerarem en particular el significat del mutualisme com a generador i sustentador d’estructures ecològiques, i el seu paper com a catalitzador de bona part de la productivitat primària i secundària de molts ecosistemes terrestres.
El mutualisme com a generador d’estructura
El grau d’estructuració de l’ecosistema es pot mesurar en termes de la seva diversitat, la seva estructura tròfica, la circulació de nutrients, etc.; totes són propietats emergents del conjunt però no pas de les parts (poblacions, espècies). Una altra propietat emergent, la connectància, mesura la proporció del total d’interaccions interspecífiques teòricament possibles que es donen realment. La connectància, com a mesura d’estructura, depèn, doncs, de la «densitat» d’interaccions entre espècies. Les relacions de mutualisme incrementen la diversitat específica i la connectància, tant directa com indirectament.
No podem fer experiments per augmentar artificialment la incidència de les relacions de mutualisme en la natura i observar després els efectes sobre la diversitat. Però sí que podem predir les conseqüències de la desaparició de les relacions de mutualisme. Del miler d’espècies d’abelles, quantes en continuarien existint després d’un canvi en què totes les espècies de plantes entomòfiles es tornessin de cop autògames, deixessin de produir nèctar i reduïssin dràsticament el nombre de grans de pol·len per flor fins a aconseguir quocients pol·len/òvul pròxims a 1? Quant de temps es mantindria el nombre actual d’espècies de plantes entomòfiles si totes es tornessin de cop anemòfiles, amb la conseqüent necessitat d’ una estreta proximitat física entre els individus i amb la inevitable saturació dels estigmes florals amb pol·len aliè? Continuarien existint gaires insectes frugívors altament especialitzats («plagues» en els conreus humans, però genuïna diversitat biològica en els ecosistemes naturals), com són ara els dípters tefrítids, si la dispersió de llavors per vertebrats fos de cop substituïda per l’anemocòria i els fruits carnosos desapareguessin? Finalment, sobreviurien les aranyes de la família dels tomísids, especialistes en la cacera del pol·linitzador, a la desaparició de les flors entomòfiles i de llurs visitants? Aquest exercici d’imaginació biològica podria prosseguir indefinidament amb altres exemples que il·lustressin, com els precedents, que la diversitat biològica (la riquesa en espècies) depèn sovint estretament de l’existència d’una relació de mutualisme.
Els dos mutualismes que ens serveixen d’exemple, pol·linització i dispersió, tracten de la provisió de productes nutritius (nèctar, pol·len, polpa dels fruits). Els animals mutualistes són els usuaris «legítims» d’aquests recursos, però no pas els únics. Aquest aspecte és important per a entendre la influència potencial del mutualisme sobre la connectància, com ho demostra la següent sèrie d’exemples: els coleòpters de la família dels brúquids, en els seus estats larvals, són depredadors especialitzats de llavors, sobretot de fabàcies, i els adults (és a dir, els que pondran els ous als llegums) s’alimenten de pol·len i nèctar de les flors entomòfiles; els himenòpters bracònids i els dípters taquínids són parasitoides obligats d’altres insectes, sobretot de lepidòpters, i els adults s’alimenten de nèctar floral; aquesta situació és similar a la de moltes vespes solitàries (esfècids, eumènids) que nodreixen llurs cries amb larves o adults d’insectes, mentre que el nèctar i el pol·len son l’aliment bàsic dels adults. A les nostres latituds no hi ha exemples tan nítids com els anteriors per al mutualisme planta-dispersador. No obstant això, podem esmentar diferents espècies d’ocells granívors de la família dels fringíl·lids que s’alimenten de les llavors contingudes als fruits sense dispersar-les; per exemple el verdum (Carduelis chloris), el pinçà comú (Fringilla coelebs), el durbec (Coccothraustes coccothraustes) o els dípters de la família dels tefrítids abans esmentada, els quals estan especialitzats a alimentar-se de polpa de fruits gràcies a una estreta simbiosi amb microorganismes.
El que volen il·lustrar els exemples anteriors és que la fracció de connectància (o estructura, si la mesurem amb aquesta magnitud) que es pugui deure localment a la relació brúquids-lleguminoses, parasitoides-preses, vespes-preses, etc. és mantinguda de forma indirecta per la relació del mutualisme planta-pol·linitzador, i més concretament pel recurs alimentari que aquesta relació fa assequible per a molts organismes, a més dels mutualistes legítims. Altres vegades l’increment de connectància és a causa de relacions molt més directes, com és el cas de les relacions de predació que es basen precisament en els pol·linitzadors. Molts invertebrats s’han especialitzat a caçar pol·linitzadors a les flors o prop seu, com fan alguns dípters de la família dels asílids, els màntids i les aranyes tomísides. Entre els vertebrats, l’abellerol (Merops apiaster) n’és un exemple familiar.
El mutualisme com a catalitzador
A efectes del seu estudi i anàlisi, molts ecòlegs conceben els ecosistemes com a sistemes compartimentats en nivells tròfics, connectats entre ells i amb el medi físic per fluxos de matèria i energia. La quantificació de la magnitud dels compartiments i dels fluxos configura els objectius i la metodologia de l’ecologia de sistemes. En el context d’aquesta concepció de l’ecosistema, el major o menor significat de les diferents relacions entre organismes és determinat per les respectives magnituds de les transferències d’energia i nutrients associades amb cadascuna d’elles. En aquest esquema, les relacions del tipus +/– com l’herbivorisme o la predació adquireixen el paper més destacat perquè per aquestes vies es realitzen la major part de les transferències entre compartiments. Tot i que no tenim prou informació quantificada, sembla fora de dubte que les relacions de mutualisme representen magnituds petites en el conjunt de fluxos materials (encara que no menyspreables en alguns casos, com mostren les xifres de producció de fruits esmentades en un apartat anterior). Igualment, llur participació directa en els processos de producció primària i secundària deu ser poc destacada en termes de «quantitats».
Parlant quantitativament, les magnituds significatives d’un procés de síntesi química industrial a gran escala són les quantitats de primera matèria i de producte elaborat final. No obstant això, la magnitud del producte final depèn quasi sempre de la intervenció en el procés de petites quantitats de substàncies que acceleren i fan possible la síntesi. Es tracta dels catalitzadors. Moltes relacions de mutualisme, encara que poc importants en termes de xifres absolutes, actuen efectivament com a catalitzadors de la productivitat dels ecosistemes. Es tracta en essència de relacions de baixa energia amb implicacions quantitatives a escala molt més gran. Aquest és el sentit de l’analogia entre mutualisme i catàlisi, que és il·lustrat breument en els exemples següents. La productivitat primària que representen els fruits i les llavors de totes les plantes entomòfiles d’un ecosistema ha estat «catalitzada» mesos o setmanes abans per la relació mutualista entre les dites plantes i llurs pol·linitzadors. La productivitat secundària d’ocells i mamífers granívors que s’alimentin d’aquestes llavors és igualment conseqüència de la mateixa catàlisi. En la mesura que els productors primaris d’un ecosistema (plàntules, plantes adultes) procedeixen de llavors que sorgiren d’una pol·linització entomòfila i/o foren disseminades per un ocell o un mamífer, llur considerable biomassa i productivitat primària són el resultat final d’una interacció catalítica de baixa energia en què intervenen un o més mutualismes.
El significat ecològic del mutualisme
Els apartats anteriors volen ser un petit cop al pèndol que manté l’atenció dels ecòlegs insistentment penjada de les relacions de competència i depredació, en un intent de desplaçar-lo cap a posicions més centrals i equidistants, i no pas de fer-lo bascular cap a un nou extrem. Totes les relacions que els ecòlegs reconeixen tradicionalment entre els organismes existeixen a la natura, cadascuna amb la seva dinàmica i amb implicacions peculiars, i no es tracta, per tant, de donar més significat a unes que a les altres. El fet que remarquem ací la importància del mutualisme no obeeix a la idea que sigui una relació més important que les altres sinó, més aviat, a la necessitat d’assenyalar que és una relació amb un interès similar a la resta, però que, per raons poc científiques, ha estat infreqüentment estudiada. El mutualisme no és una relació rara ni exclusivament tropical. En tenim la caricatura al costat nostre, en el cas de les abelles que visiten les flors o en les merles que mengen fruits als parcs. És patent i ubiqua en els ecosistemes naturals, com hem demostrat, sovint més que no la competència o la predació: veiem més sovint insectes a les flors que esparvers caçant ocells o espècies en competència. Explica una part substancial de la diversitat biològica, genera estructura als ecosistemes i és responsable, en darrer terme, d’una fracció important de la productivitat. No hi ha, per tant, raons objectives per a seguir mantenint la relació mutualista en una posició marginal dins de l’ecologia, solament perquè sembla difícil d’encaixar dins la poderosa «ecologia de la competència i la predació», que hem heretat.
El nostre coneixement actual sobre l’evolució de les relacions de mutualisme, la seva influència en la dinàmica poblacional, els mecanismes estabilitzadors, etc., és molt rudimentari, sobretot en els aspectes teòrics. No hi ha un cos teòric específic per a les relacions de mutualisme, per causa, sobretot, de l’escàs interès que tradicionalment ha despertat el tema, però també per la gran varietat de formes en què es manifesta. Això fa difícil la necessària simplificació que tota teoria requereix i alhora crida l’atenció sobre la necessitat de conèixer amb detall suficient la història natural dels mutualistes. La història natural d’organismes, sistemes i interaccions és la base empírica que ens cal abans de fer qualsevol teoria ecològica. L’estudi del mutualisme no ha anat encara gaire més enllà, però serà més fàcil progressar en el seu coneixement si comencem a admetre’n l’existència com a fenomen pròxim i generalitzat.