Consideracions generals

Les parts fondes de les cavitats constitueixen el medi més característic de la fauna troglòbia. Les condicions ambientals hi són remarcablement estables, i els recursos tròfics, poc espectaculars però força regulars. La fotografia, presa a la cova de la Fou de Bor (Cerdanya), mostra alguns biòtops típics, com la base amarada d’aigua de les colades estalagmítiques, i els petits bancs d’argila.
Lluís Auroux.
Durant molt de temps les cavitats subterrànies es consideraren desproveïdes d’interès biològic, i les primeres troballes d’animals cavernícoles, fetes entre els segles XVIII i XIX, van ser acollides per la comunitat científica com a simples curiositats que no mereixien cap més atenció. El 1905, però, el naturalista Emil G. Racovitǎ, esperonat per les seves descobertes faunístiques a les coves del Drac de Manacor (Mallorca Oriental) i ajudat pel seu deixeble R. Jeannel, fundà l’organització anomenada Biospeologica, amb la finalitat de sistematitzar la investigació dels organismes de les coves, establint les bases de la moderna biospeleologia, o estudi biològic de les cavitats subterrànies. En el temps transcorregut, la biospeleologia ha deixat de ser una simple curiositat per a esdevenir una disciplina científica amb personalitat pròpia, com ho demostra l’espectacular volum de dades obtingudes, moltes de les quals ultrapassen el nivell de la taxonomia bàsica per a incidir en àrees tan diverses de les ciències que estudien la vida com són la fisiologia, l’ecologia, la genètica o l’evolució.
El medi
Les cavitats subterrànies presenten una sèrie de peculiaritats ambientals que han condicionat les estratègies biològiques dels organismes que hi viuen. La característica més evident és l’absència de llum; d’altra banda, la humitat hi és molt elevada, gairebé fins a la saturació, i la tempertura, força constant al llarg de l’any, és propera a la mitjana anual exterior d’aquell indret.
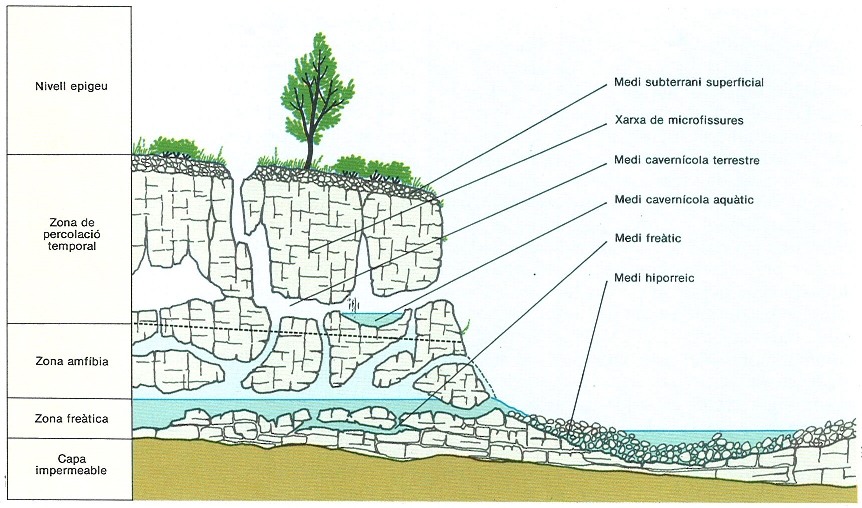
L’extensió del medi cavernícola no es limita a les coves i als avencs accessibles a l’home. Diversos medis subterranis, com els esquematitzats a la figura, poden allotjar la mateixa fauna que hom pot trobar a les cavitats accessibles.
Josep Nuet i Badia, a partir de fonts diverses
Cal afegir que aquestes característiques es refereixen a les parts fondes de les cavitats: tot al llarg hi ha un gradient de tots els factors ambientals fins a arribar a les condicions de relativa estabilitat suara esmentades. Convé aclarir també que, àdhuc en aquestes parts més profundes, l’estabilitat aparent respon tan sols a una primera aproximació, ja que els canvis estacionals també s’hi fan sentir, bé que atenuats i més o menys retardats. Altrament, alguns estudis fets amb gran precisió de mesura a prop de les parets i el terra de les coves han demostrat que es donen fenòmens de microventilació i d’evapo-condensació que provoquen variacions climàtiques petites però freqüents, la qual cosa influeix en el comportament dels animals cavernícoles. Pel que fa a la delimitació del medi cavernícola, cal esmentar que les coves han estat definides des d’una perspectiva antropocèntrica, és a dir, hom considera la cova circumscrita en l’espai accessible a l’home. Els invertebrats que hi viuen, però, rarament ultrapassen alguns mil·límetres i poden desplaçar-se lliurement per la xarxa de petites fissures adjacents. Els descobriments fets en aquest decenni sobre la fauna que havia estat considerada com a estrictament cavernícola en el medi subterrani superficial (nivell de roca fragmentada que se situa entre la roca mare i el nivell edàfic) han demostrat que aquesta fauna pot anar de la cova a aquest medi passant per la xarxa de fissures, i a la inversa. En el domini aquàtic, es dóna una situació similar de continuïtat entre les aigües cavernícoles, el medi freàtic i altres medis propers a la superfície, com l’hiporreic, és a dir, el medi intersticial constituït per la grava que serveix de llit de determinades aigües superficials. Tot això suggereix que cal considerar el medi cavernícola en sentit ampli i no restringir les observacions únicament a les coves accessibles. L’estudi de la fauna de les fissures i dels petits intersticis és, justament, un dels objectius prioritaris en la investigació biospeleològica actual.
Els factors tròfics
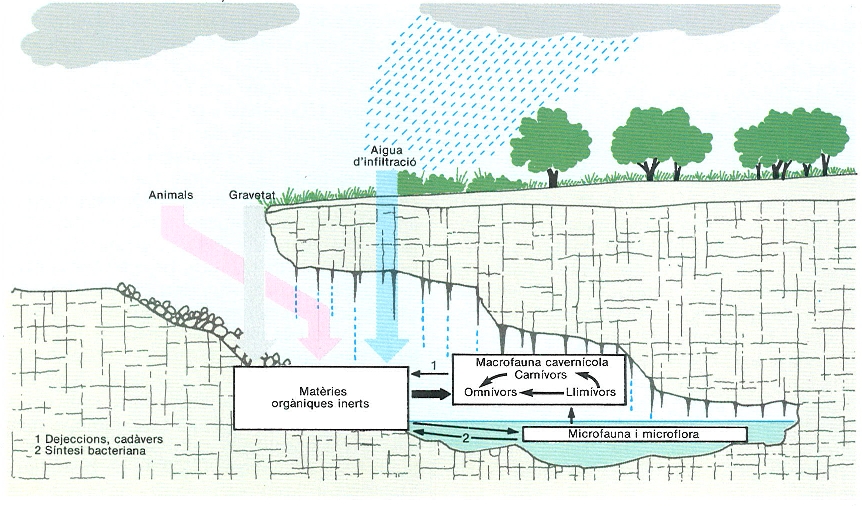
La major part dels recursos tròfics de les cavitats procedeix de l’exterior, i s’hi introdueix sobretot a través de l’aigua d’infiltració, dels animals que les visiten circumstancialment i de la gravetat. La producció primària es limita pràcticament a l’activitat quimiosintetitzadora de determinats microorganismes que viuen a l’argila.
Josep Nuet i Badia, a partir de fonts diverses
Pel que fa als factors tròfics, i atesa l’absència de llum que impedeix l’existència d’organismes fotosintetitzadors, la producció primària autòctona és molt limitada, i es redueix pràcticament a l’activitat quimiosintetitzadora de determinats microorganismes autòtrofs (particularment tiobacteris i ferrobacteris) que viuen sobretot a l’argila.
La major part dels recursos tròfics procedeixen de l’exterior i s’introdueixen a les cavitats per vies diferents. Les més aparents són: l’aigua de pluja, que en filtrar-se a les coves a través de la xarxa de fissures arrossega la fracció orgànica que ha incorporat en travessar la capa de sòl subjacent; la gravetat, que fa introduir matèria orgànica de textura més gran; i els animals que en visitar circumstancialment les cavitats aporten els excrements i sovint els seus cadàvers. Les rates-pinyades constitueixen potser l’exemple més il·lustratiu d’aquesta darrera via, com ho demostren algunes cavitats amb notables acumulacions de guano que serveixen de base a tota una munió d’invertebrats molt especialitzats per a aquest biòtop.
Els organismes i les comunitats
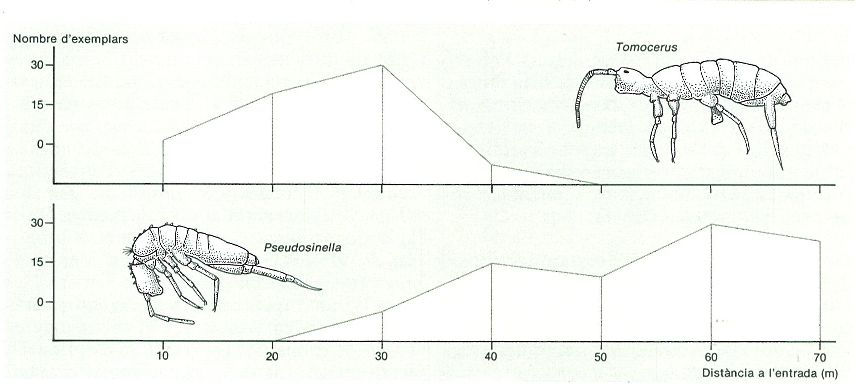
La fauna de la franja de transició que s’estableix entre la zona d’entrada i la zona profunda de les cavitats, es caracteritza perquè conté elements d’ambdues zones, la qual cosa es tradueix en un augment de la diversitat. L’esquema representa la distribució en l’espai de dues espècies de col·lèmbols en una galeria rectilínia d’una mina calcària, a Pontons (Alt Penedès). Tomocerus és troglòfil i Pseudosinella és troglòbia. Noteu que, amb la profunditat, disminueix el nombre d’exemplars de Tomocerus i augmenta el de Pseudosinella, i que ambdues espècies coincideixen a les estacions de mostreig situades a 30 i a 40 m de l’entrada, en la franja de transició.
Amadeu Blasco, original de Xavier Bellés.
No tots els animals que es poden trobar en una cova són estrictament cavernícoles; ans al contrari, hom hi acostuma a observar una mescla heterogènia d’espècies amb diversos graus d’adaptació respecte a aquest medi. El naturalista J.R. Schiner proposà el 1854 una classificació dels cavernícoles en tres categories: troglobis (estrictament confinats a les cavitats subterrànies), troglòfils (freqüents, però no estrictament confinats) i cavernícoles ocasionals. Per a aquests darrers, Emil Racoviţǎ proposà el 1907 el terme de trogloxens. Aquesta classificació tan simple ha esdevingut la més emprada, malgrat que posteriorment n’han estat publicades d’altres aparentment més precises, però que per llur sofisticació han resultat poc operatives. Per a la fauna aquàtica, S. Thienemann va proposar el 1926 els termes estigobi, estigòfil i estigoxè (de «styx» un dels rius de l’infern, segons la mitologia grega), i que són equivalents als de Schiner i Racoviţǎ. Cal aclarir que es tracta de categories força difícils d’aplicar en sentit estricte, atesa la poca informació disponible sobre la biologia de les espècies trobades en coves, però el cert és que se’n fa un ús molt generalitzat, encara que utilitzant criteris més o menys intuïtius o basats en determinats trets morfològics que veurem tot seguit.
Els animals estrictament cavernícoles o troglobis (tots invertebrats, en les nostres latituds) mostren una sèrie de peculiaritats morfològiques i fisiològiques que, encara que no els són pas exclusives, acostumen a diferenciar-los de les espècies afins que viuen a l’exterior. Pel que fa a la morfologia, les més importants són la proliferació i l’especialització de la dotació sensorial, l’allargament dels apèndixs, la despigmentació i la regressió de les ales i de l’aparell de visió, que pot arribar a la anoftalmia total. Des del punt de vista fisiològic, tendeixen a una fecunditat més baixa, a un augment de la talla i de les reserves dels ous i a una durada més gran de totes les fases del cicle biològic, compresa la d’adult; és a dir, tenen una longevitat més gran. D’altra banda, els ritmes biològics resulten molt desdibuixats, particularment els circadiaris. En general, reprenent els termes de Mac Arthur i Wilson, hom ha pogut observar una bona mostra de situacions entre aquelles espècies que segueixen l’estratègia de la r (oportunistes) i les que són típiques estrategues de la K (especialistes), amb una clara tendència, però, al predomini d’aquestes darreres.
El desenvolupament dels diferents intents d’elaborar una tipologia de les comunitats de les coves ha anat lligat a una descripció prèvia d’una sèrie de biòtops cavernícoles, més o menys característics. En una primera aproximació, i considerant simplement els factors ambientals, hom pot constatar fàcilment l’existència de dues zones ben diferenciades a les cavitats: una és la d’entrada (amb una llum solar atenuada i amb unes condicions climàtiques similars a les de l’exterior, però sense ser tan variables), i l’altra és la zona profunda (sense llum i amb les condicions de relativa estabilitat esmentades abans). Ambdues zones mostren un tipus de fauna clarament diferenciat i estan separades per una franja de transició caracteritzada perquè presenta una composició faunística en què intervenen elements de totes dues, la qual cosa es tradueix en un increment de la diversitat, com hom pot observar de manera més o menys espectacular en gairebé totes les coves. A part d’aquesta divisió bàsica, diversos autors han intentat de definir biòtops més concrets. El problema consisteix en el fet que molts són redundants i difícils de delimitar. Malgrat tot, hom pot arribar a reconèixer-ne alguns de prou característics i menys resistents a la definició; vegem-los tot seguit.
Les comunitats de la zona d’entrada

La zona d’entrada de les cavitats es caracteritza per la presència de plantes verdes típiques de llocs ombrívols i humits, i perquè mostra unes condicions climàtiques peculiars, encara que notablement influïdes per les de l’exterior. La fauna d’aquesta zona és composta fonamentalment d’elements higròfils i obscurícoles, no estrictament cavernícoles. El vestíbul de la cova del Gel, al massís del Montsec, pot servir com a exemple de les característiques d’aquesta zona d’entrada.
Lluís Auroux.
La zona d’entrada, tant el vestíbul de les coves com el pou d’accés als avencs, es caracteritza per unes condicions climàtiques força influïdes per les de l’exterior, però amb unes fluctuacions més atenuades. La major part de la matèria vegetal prové de l’exterior i es troba en forma de restes de textura grossa (fullaraca, fusta en descomposició, etc.) que es mesclen amb la terra rica en matèria orgànica i amb els esbaldregalls, que són particularment abundants en aquest sector.
D’altra banda, la llum permet l’existència de plantes verdes que es distribueixen d’acord, sobretot, amb els factors del relleu i ambientals, entre els quals destaquen, de lluny, la llum i la humitat. Hom ha constatat en diverses coves dels Pirineus centrals l’existència de tres nivells florístics: en el primer dominen les criptògames superiors, especialment les falgueres, que són força exigents, de llum i humitat; els briòfits, més exigents d’humitat, defineixen aproximadament un segon nivell, situat a una distància més gran de l’entrada però encara prou il·luminat (entre un 5% i un 2% de la llum de l’exterior); al tercer nivell, hi arriben a penes uns pocs luxs i només pot ser colonitzat per alguns líquens, uns pocs fongs i, sobretot, cianofícies. Les algues cianofícies són dominants en aquest tercer nivell, al llindar de la foscor absoluta, ja que poden experimentar adaptacions cromàtiques i sintetitzar pigments, ultra la clorofil·la, que fan possible la captació de llum amb un millor rendiment. A més, les formes més senzilles, amb menys capacitat d’adaptació cromàtica, poden associar-se tot optimitzant la captació de llum. Un cas il·lustratiu en aquest sentit és el de Cueva Vieja de Villanúa, als Pirineus centrals. A la zona d’entrada es desenvolupen diferents Gloeocapsa, Chroococcus i d’altres espècies de cianofícies, que mostren pigments un xic diferents cadascuna i que donen bandes d’absorció de llum complementàries. Sembla clar que aquesta situació és el resultat de l’efecte de competència entre les espècies, però fa la impressió com si el conjunt funcionés d’acord, per tal de cobrir una màxima regió d’absorció de llum que cap espècie sola no podria aconseguir.
Pel que fa a la fauna, aquesta zona tan peculiar acull una sèrie d’elements higròfils i obscurícoles, que poden trobar-se també en altres ambients críptics i humits, no necessàriament cavernícoles, de l’àrea en qüestió. Així, doncs, entre els invertebrats no artròpodes, són freqüents els mol·luscs del gènere Oxychilus i els oligoquets lumbrícids. D’aràcnids, hom n’acostuma a trobar diferents aranèids (Leptoneta), pseudoscorpins (Chthonius, Neobisium, Roncus) i opilions (Scotolemon, Nemastoma, Astrobunus grallator). Els crustacis són representats pels isòpodes terrestres típicament humícoles (Cylisticus esterelanus, diverses espècies de Porcellió) o bé per diferents triconíscids endogeus; també són molt típics els diplòpodes polidèsmids, especialment l’espècie Polydesmus coriaceus, i diferents quilòpodes humícoles del gènere Lithobius. Entre els insectes, hi ha una gran profusió de col·lèmbols, principalment entomòbrids (Tomocerus, Heteromurus, Pseudosinella) i, amb menys abundància, també hi són representats els diplurs (campodèids i japígids); en gairebé tots els casos es tracta d’elements endogeus o humícoles. De l’ordre dels coleòpters, trobem tan aviat espècies endogees (caràbids escaritins i anil·lins) com humícoles (caràbids del gènere Trechus), o bé muscícoles (catòpids del gènere Bathysciolà).
En conjunt, es tracta d’una mescla molt heterogènia d’elements, molts circumstancials i, d’altres, força característics, en què dominen els saproxilòfags, però amb una bona representació de depredadors. La diversitat global és relativament alta, però això cal atribuir-ho més aviat a la presència de moltes espècies accidentals i també al fet que es mesclen associacions molt localitzades que semblen essencialment diferents però que costen dedelimitar, atesa la considerable intensitat dels gradients de elstots els paràmetres amb incidència ecològica.
Altrament, aquestes comunitats varien considerablement en el temps, d’acord amb les variacions pròpies de les diferents èpoques de l’any. L’estudi de la cova de la Torre, a prop de Sant Feliu de Pallerols (Garrotxa), ha posat de manifest que, al final de la tardor, quan el riu que travessa la cova perd cabal i afavoreix la deposició d’una abundant matèria vegetal al vestíbul, les comunitats d’aquest sector estan compostes principalment per formigues (Myrmecina, Myrmica), àcars trombídids i coleòpters estafilínids (Trogophloeus, Gabrius). En canvi, al començament de la primavera, quan el riu assoleix el seu màxim cabal i neteja la cova de la major part de les restes dipositades anteriorment, la composició faunística té un perfil absolutament diferent, on dominen els col·lèmbols tomocèrids i els glomèrids del gènere Rypharomeris. Veiem, doncs, com les fluctuacions de la matèria orgànica fan canviar dràsticament l’estructura d’aquesta comunitat, tot seguint uns cicles anuals molt condicionats per les fluctuacions estacionals del cabal del riu.
Les comunitats parietals

Les aranyes són elements típics de les comunitats parietals i fan les seves teranyines a les anfractuositats de la roca, on atrapen les seves preses, principalment mosquits i microlepidòpters, també parietals. Hom pot veure a la fotografia un exemplar de l’aranèid Meta bourneti penjat dels fils de la seva tela a la cova del Far, a Susqueda (Selva).
Xavier Bellés
A les voltes i a les parets de les cavitats, especialment a la zona d’entrada i a la franja de transició, es troba un tipus de fauna molt característic compost per diverses espècies, sobretot per aràcnids i insectes.
Entre els aràcnids, destaquen diferents aranèids, que fan les seves teranyines a les an-fractuositats de la roca (diferents espècies de Meta, Pholcus, Tegenaria, Nesticus), i opilions, normalment frondícoles, de potes extraordinàriament llargues (com Leiobunum rotundum, Gyas titanus o Cosmobunus granarius) i que es disposen molt a prop els uns dels altres, entrellaçant les potes i, de vegades, formant veritables masses de nombrosos individus.
Quant als insectes, cal destacar la presència freqüent en nombroses cavitats pirinenques i prepirinenques de dues espècies d’ortòpters del gènere Dolichopoda: D. linderi (Garrotxa, Ripollès, Osona, Selva) i D. bolivari (Berguedà, Solsonès, Pallars, Alt Urgell). Ultra aquests ortòpters, les espècies d’insectes més freqüents en les comunitats parietals s’inclouen en els ordres dels dípters, tricòpters i lepidòpters. La seva estada a les coves sembla relacionada amb factors estacionals, ja que hi ha espècies que es refugien exclusivament a l’hivern, mentre que d’altres ho fan a l’estiu. A més, en alguns casos hom ha demostrat que aquest període cavernícola té determinades conseqüències fisiològiques, com per exemple la interrupció de l’activitat reproductiva. Entre els dípters hi ha espècies hivernants i estiuejants. La més típica a les nostres cavitats és potser el limònid Limonia nubeculosa, que és estiuejant i, de vegades, es troba en gran nombre (s’han observat densitats de prop de 40 exemplars per metre quadrat en determinats racons preferencials de coves pirinenques). Els tricòpters es refugien en les cavitats quan són adults i durant el període d’estiu; les espècies observades pertanyen als gèneres Mesophylax, Stenophylax i Micropterna, dins la família dels limnofílids. Els lepidòpters acostumen a trobar-se més freqüentment a l’hivern i dominen les espècies de noctúids (Scoliopteryx, Pyrois, Autophila, Apopestes) i de geomètrids (Triphosa). Sembla que algunes d’aquestes espècies experimenten una veritable diapausa durant el període cavernícola, com per exemple Scoliopteryx libatryx, que sempre es troba absolutament quiescent; d’altres, com Triphosa dubitata, fan desplaçaments curts per les parets i àdhuc sortides a l’exterior de la cova durant la nit.
Les comunitats guanòfiles

El guano de rata-pinyada constitueix una de les fonts d’energia més importants de les coves. El forat d’en Ferràs, prop d’Orpesa de Mar (Plana Alta), és un bon exemple de cavitat amb notables acumulacions de guano que suporten una fauna molt especialitzada a aquest substrat i poc diversa.
Tere Cuñé
Com hem esmentat abans, els excrements dels animals que visiten les coves circumstancialment, especialment les rates-pinyades, són una de les fonts d’energia més importants. Les dejeccions aïllades són sovint aprofitades per les espècies sapròfagues estrictament cavernícoles, però les grans acumulacions de guano suporten una fauna molt diferent, altament especialitzada per a aquest substrat.
Els elements dominants són determinats col·lèmbols (com Nesachorutes ojcoviensis), àcars micetòfags i depredadors, i dípters copròfags pertanyents sobretot als fòrids, esferocèrids i heleomícids. D’aquesta darrera família cal esmentar la mosca del guano (Thelida atricornis), que ha estat capturada en algunes cavitats pirinenques. Sol viure sobre les masses de guano fresc i hom pot observar totes les fases del seu cicle biològic durant tot l’any. Els coleòpters són representats per espècies copròfagues (com per exemple, l’histèrid Gnathonchus nannetensis), micetòfagues (com el colídid Aglenus brunneus) i depredadores, com el caràbid Pristonychus terricola, o diferents estafilínids dels gèneres Atheta, Quedius o Conosoma. També són depredadors, a les nostres contrades, els pseudoscorpins del gènere Allochernes, A. masi i A. wideri. Ocasionalment, es troben també artròpodes ectoparàsits de les rates-pinyades que s’han desprès dels hostes, com ara diferents dípters o àcars parasítids o ixòdids.
L’estat del guano influeix en gran manera sobre l’estructura i la dinàmica de les comunitats d’artròpodes que hi viuen. Quan és permanentment fresc, com és el cas d’aquelles cavitats on hi ha concentració de rates-pinyades tot l’any, trobem comunitats constituïdes per un gran nombre d’exemplars, però pertanyents a unes poques espècies (particularment de dípters i àcars), és a dir, caracteritzades per una diversitat molt baixa. En canvi, si la cavitat és abandonada temporalment per les rates-pinyades, per exemple després d’una hibernació, el guano perd aigua i experimenta una sèrie de transformacions químiques, la qual cosa afavoreix la invasió per altres espècies, amb l’augment consegüent de la diversitat; hom pot observar, aleshores, determinades modificacions en l’estructura de la comunitat guanòfila, com són la desaparició dels paràsits despresos de les rates-pinyades, la disminució de les larves de dípters copròfags, l’augment dels coleòpters histèrids i dels col·lèmbols i l’aparició d’espècies sapròfagues diverses. Amb el retorn de les rates-pinyades, les comunitats guanòfiles tornen a la situació de baixa diversitat d’espècies, esmentada al principi i típica del guano fresc. Com ja ha estat suggerit per diversos autors, les comunitats de les masses de guano tenen molt poc a veure amb la fauna pròpiament cavernícola. De fet, les comunitats que trobem a les coves són pràcticament iguals a les que hom pot observar en les acumulacions de guano d’altres medis no cavernícoles, per exemple dels medis urbans.
Les comunitats estrictament cavernícoles
A una certa distància de l’entrada, allà on les característiques climàtiques i tròfiques aconsegueixen les condicions de relativa estabilitat que són necessàries per a definir el medi pròpiament cavernícola, s’estableixen les comunitats troglòbies.

Les característiques d’un animal estrictament cavernícola són sovint peculiars per la notable longitud dels apèndixs, la despigmentació i la regressió de l’aparell ocular i de les ales membranoses. L’exemplar de la fotografia correspon a l’espècie Paraphaenops breuilianus, un coleòpter troglobi que és endèmic de les cavitats del Caro, als ports de Beseit
Xavier Bellés
Nombrosos ordres d’artròpodes terrestres hi són representats. Entre els aràcnids dominen els aranèids (Leptoneta, Leptyphantes, Troglohyphantes, Speleoharpactea levantina, endemisme del Montsià i el Baix Maestrat, o l’interessantíssim Telema tenella, localitzat en unes poques cavitats d’ambdues vessants dels Pirineus orientals), i els pseudoscorpins (Ephippiochthonius, Parablothrus, o les remarcables relíquies Spelyngochthonius heurtaultae, d’una cova del Tarragonès, o Troglobisium racovitzai, repartida sobretot pel Garraf). Entre els crustacis trobem una gran diversificació d’isòpodes terrestres, particularment de la família dels triconíscids (Spelaeonethes, Catalauniscus, Oritoniscus, Trichoniscoides). Els diplòpodes més freqüents són els júlids (Tarracoblaniulus, Typhloblaniulus, Paratyphloiulus), però també s’hi poden trobar craspedosòmids (Cranogoma, Psychrosoma, o bé l’endemisme de la Cerdanya Hispaniosoma racovitzai). Entre els quilòpodes poden observar-se alguns Lithobius força modificats per adaptació al medi cavernícola (com, per exemple, L. ambulotentus, a la vall de Ribes). Entre els insectes apterigots, dominen de lluny els col·lèmbols, especialment entomòbrids del gènere Pseudosinella, encara que també es troben altres ordres, com diplurs campodèids dels gèneres Plusiocampa, Podocampa i Paratachycampa; o bé tisanurs, com Coletinia capolongoi, endemisme de la Ribera Alta. Els pterigots són representats sobretot pels coleòpters, tan aviat depredadors (diferents caràbids dels gèneres Geotrechus i Aphaenops a l’àrea pirinenca, Duvalius i Paraphaenops en algunes comarques tarragonines, Duvalius i Trechopsis a Mallorca, o bé els interessants endemismes Zariquieya troglodytes, propi de l’Alt Empordà i la Garrotxa, Henrotius jordai de Mallorca, o l’extraordinari Ildobates neboti de la Plana Alta i el Baix Maestrat), com detritívors, representats particularment per una notable quantitat d’espècies de catòpids de la subfamilia dels batiscins pertanyents sobretot als gèneres Speonomus i Troglocharinus.
En el domini aquàtic hi ha diversos crustacis, principalment isòpodes (Stenasellus del grup virei al Principat, Typhlocirolana moraguesi a les Balears) i amfípodes (Pseudoniphargus del grup africanus). Acostumen a trobar-se en aigües embassades, com tolls i llacs (sobretot quan hi ha llim al fons) més que no pas en aigües corrents (rius subterranis). Diguem, de passada, que, en aquest ambient cavernícola saturat d’humitat, queden molt desdibuixats els límits entre el domini terrestre i l’aquàtic, perquè s’hi poden observar tant espècies terrestres que travessen un toll per sota l’aigua (per exemple, determinats isòpodes triconíscids), com altres de típicament aquàtiques (Stenasellus, posem per cas) que deambulen per l’argila humida adjacent a l’aigua.
Deixant a part els efectes deguts a la fecunditat i la mortalitat específiques, i atesa la relativa constància ambiental, l’estructura i la dinàmica de les poblacions cavernícoles poden interpretar-se partint principalment de les interaccions entre les espècies. Però se sap ben poca cosa sobre aquestes interaccions, sobretot pel que fa a la fauna terrestre, i els diferents especialistes no acaben de posar-se d’acord sobre quina importància cal atribuir a cadascun dels diferents tipus d’interacció (competència, predació, parasitisme, mutualisme). És probable que tots els tipus coexisteixin en una mateixa comunitat, donant una xarxa complexa. En el cas de la fauna aquàtica dominen els fenòmens de competència, i possiblement de mutualisme, atès que els depredadors són pràcticament absents a les nostres cavitats. En els poquíssims casos on hi ha depredadors grans, la petita fauna aquàtica esdevé francament rara: per exemple, en el riu subterrani que travessa la cova d’Aso, a la vall d’Añisclo (Pirineus centrals), viu una colònia cavernícola del tritó pirinenc (Euproctus asper) i, malgrat les successives prospeccions de fauna aquàtica que s’hi han realitzat, tan sols s’hi han trobat invertebrats (com per exemple larves de dípters quironòmids, l’isòpode Stenasellus virei) en el tub digestiu dels tritons.
Sobre la dinàmica poblacional d’espècies cavernícoles concretes, hom disposa d’una mica més d’informació, i un dels grups més estudiats en aquest sentit és el dels coleòpters catòpids de la subfamilia dels batiscins. Per exemple, la distribució espacial de Speonomus delarouzeei ha estat estudiada en diferents estacions dins l’avenc de Talaixà, cavitat d’uns 180 m2 de planta, situada a prop de Terrades (Garrotxa). Els resultats permeteren de censar la població (10 309 exemplars, a partir del marcatge de 1652 exemplars, dels quals es retrobaren 259 entre un total de 1350 recollits a la campanya de recaptura), calcular les probabilitats de transició entre les diferents estacions estudiades, estimar la dinàmica espàcio-temporal de la població, i quantificar determinades preferències ecològiques de l’espècie; cal destacar, per exemple, que el temps de retorn resultà més curt per al cas d’aquelles estacions més riques en matèria orgànica i més estables des del punt de vista climàtic. També d’aquesta mateixa espècie s’han estudiat mensualment les fluctuacions poblacionals al llarg d’un any a les coves de Rialb, de la vall de Ribes (Ripollès) i s’ha vist que, en les estacions climàticament més inestables, es dóna una certa correlació entre determinats paràmetres físics (temperatura, humitat, pluviositat i ventilació) i les variacions numèriques de la població; per exemple, la davallada de la humitat relativa i l’augment màxim de temperatura que es produeix al mes d’agost es corresponen amb els valors poblacionals més baixos de l’any. Així, doncs, en aquestes estacions, els factors abiòtics semblen determinants per a interpretar les fluctuacions de la població, i sembla que les condicions ambientals desfavorables poden propiciar la migració de les espècies d’Speonomus a d’altres indrets de la cavitat més estables. Les observacions fetes en zones menys pertorbades climàticament donen també fluctuacions poblacionals notables, que no poden interpretar-se a partir de fenòmens d’immigració-emigració determinats per factors climàtics i que suggereixen l’existència de ritmes de reproducció, també postulats per altres autors, principalment G. Racoviţǎ. Aquests ritmes estarien condicionats, d’una manera més subtil i indirecta, per factors estacionals (sobretot els fenòmens cíclics de pluviositat amb l’aportació consegüent de matèria orgànica) que, malgrat ser atenuats i retardats, es deixarien sentir en aquests sectors aparentment estables.
En definitiva, i generalitzant molt, l’ecosistema estrictament cavernícola, cal considerar-lo com a relativament estable i madur, amb uns recursos tròfics majoritàriament exògens, però que arriben amb una regularitat remarcable. Tots aquests factors fan que la diversitat sigui alta, contràriament al que podria semblar a primer cop d’ull en un ecosistema aparentment poc favorable per al desenvolupament de la vida. Alhora, però, es tracta d’un ecosistema fàcil de pertorbar i, de fet, hom ha pogut comprovar en diverses de les nostres cavitats com l’acció de l’home (abocadors «controlats» en massisos càrstics, avencs usats com a pous negres o per a desfer-se del bestiar mort, àrees de gran volum amb les aigües d’infiltració —que van a parar a les cavitats i després a les fonts— contaminades per residus industrials de tota mena, etc.) ha fet canviar de forma difícilment reversible el seu estat natural.