Consideracions generals sobre els conreus com a ecosistemes
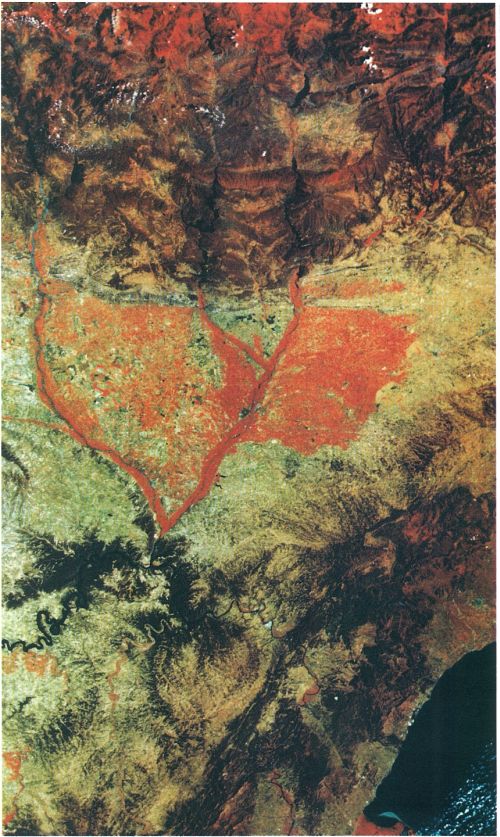
L’eficàcia energètica dels ecosistemes agrícoles és funció de la seva capacitat fotosintetitzadora. En aquesta imatge en fals color obtinguda per un satèl·lit artificial equipat amb sensors MSS («Multispectral Scanner») es posa en evidència l’alta capacitat per a capturar energia solar dels conreus irrigats de les valls del Segre i del Cinca, del Camp de Tarragona o del delta de l’Ebre (superfícies vermelles), superior a la dels boscos prepirinencs i pirinencs (part superior de la imatge). Els secans que els envolten (tons marronosos) tenen una productivitat baixa, comparable a la dels boscos i les bosquines mediterranis (meitat inferior de la imatge).
Institut Cartogràfic de Catalunya
Hom ha dit que els ecosistemes agrícoles s’han de considerar a mig camí entre els ecosistemes naturals, com ara boscos o aiguamolls, i altres d’artificials, com són els ecosistemes urbans. En ells també és l’energia del sol el flux d’entrada que permet tant el funcionament instantani del sistema com el manteniment en el temps de la seva estructura. Cal dir que la seva posició més pròxima a uns o a altres ecosistemes depèn en gran manera del grau d’industrialització del conreu. Encara que els ecosistemes agrícoles s’han de considerar autotròfics, mentre que els urbans són heterotròfics, els fluxos energètics de certs conreus (energia total consumida per unitat de superfície) arriben a ser deu vegades més grans que en els ecosistemes naturals, i prop dels que es donen a les ciutats. Però, en altres aspectes, els ecosistemes agrícoles tenen les seves particularitats.
En primer lloc, gaudeixen d’una energia auxiliar que no prové directament del sol i que en els conreus més primitius consisteix en l’esforç dels animals domèstics i del mateix home, mentre que en els sistemes més moderns és a base de combustibles que, certament, es poden considerar energia solar fòssil, amb un paper fonamental en la producció d’aquests ecosistemes.
En segon lloc, s’hi afavoreix una o unes poques espècies, les quals donen el producte que es conrea, i així la diversitat de components de la comunitat d’organismes es redueix a conseqüència de la intervenció de l’home: les plantes i els animals propis d’un lloc se substitueixen per unes poques espècies originàries d’altres contrades; això representa, a més de la desaparició de l’estructura pròpia de la comunitat natural, l’eliminació de la seva xarxa d’interaccions tròfiques; per a l’organització del nou sistema cal una nova relació entre producció i consum que no serà pas aliena als patrons d’homogeneïtat i heterogeneïtat de l’espai.
En tercer lloc, les plantes dominants en els ecosistemes agrícoles s’han de considerar sotmeses a una selecció artificial més que no pas a la selecció natural en què estan sotmesos els organismes que formen els ecosistemes naturals; des del punt de vista taxonòmic, és curiós que quasi la totalitat dels conreus són formats per unes dues dotzenes d’espècies vegetals, que representen una mínima part sobre el total de vora 350 000 espècies de plantes superiors.
En quart lloc, mentre que en els ecosistemes naturals el control del sistema en conjunt es pot considerar intern, mitjançant una sèrie de mecanismes reguladors molt generals en tots els nivells de la jerarquia de sistemes vius, en els ecosistemes agrícoles el control general es fa des de fora, molt sovint des de centres de decisió molt allunyats del conreu. En altres temps el pagès es podia considerar en certa manera un element més del control intern del sistema, amb capacitat per a respondre als nombrosos mecanismes de regulació, positius i negatius, del mateix conreu. La tendència és cap a altres elements de control, que generalment l’exerceixen tot cercant de maximitzar la producció a curt termini.

L’ús d’hivernacles fets de materials plàstics (del Maresme, a la fotografia), a més de procediments de reg i fertilització força eficients, permet obtenir condicions microclimàtiques diferents del medi extern, amb les quals s’accelera el cicle de desenvolupament de les plantes i, per tant, l’aportació als mercats de productes vegetals en moments de mínima competència. Els conreus forçats són els sistemes de producció que més s’assemblen als industrials, ja que suprimeixen la incertesa tradicional de la consecució final del producte.
Manel Crespo
En les darreres dècades hi ha hagut increments importants de les aportacions d’energia auxiliar als conreus, les quals fins a la meitat dels anys seixanta tenien repercussions notables en la producció, mentre que d’aleshores ençà es pot parlar d’una situació d’una certa aturada. En relació amb això resulta força descoratjadora la comparació del creixement de l’energia continguda a l’aliment consumit les darreres dècades amb el creixement corresponent a les energies auxiliars que han fet possible d’obtenir aquest aliment: mentre que la primera s’ha duplicat en un període de 30 anys assolint gairebé 1,5 bilions de megajoules (MJ), la segona s’ha triplicat i n’assoleix els 10 bilions. La crisi energètica ha portat a l’elaboració de models de conreus que permeten de fer estimacions de balanços entre les entrades d’energia (solar més auxiliar) i les sortides (producció del conreu). En alguns dels esquemes de balanços energètics d’ecosistemes agrícoles es fa una distinció entre subsidis energètics directes (combustibles, electricitat i treballs fets pel bestiar o l’home) i els indirectes, com ara els adobs, les llavors, els biocides, la maquinària i l’aigua, que es poden avaluar en termes d’energia d’una manera que, tot i ser aproximada, permet una visió quantificada del funcionament del conreu. D’algunes de les entrades al sistema resulta senzilla l’estimació de balanços si tenim en compte les sortides; és el cas de l’aigua i dels elements químics que constitueixen els adobs (nitrogen, potassi, fòsfor, etc.). L’existència d’una energia auxiliar que no prové directament del sol no és exclusiva dels agroecosistemes, però sí que ho és el caràcter no natural d’aquesta aportació feta per l’home. En altres ecosistemes hi ha energies, com la de les marees, la barreja vertical de les aigües, el vent i el foc, que en general tenen un efecte desorganitzador sobre l’estructura del sistema, el qual comporta un renovellament molt sovint associat a un notable increment de la producció. Aquest principi no és aliè a l’home, que tendeix a simplificar els ecosistemes naturals en recerca de la seva explotació i d’aquesta manera els rejoveneix i n’aconsegueix un augment de la producció, especialment en la del producte que específicament desitja.
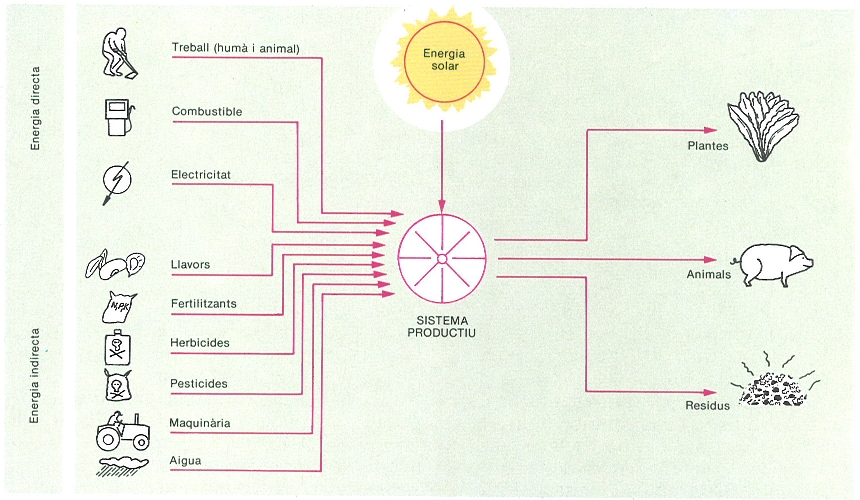
La producció agronòmica sempre ha requerit l’aportació d’algun tipus d’energia auxiliar. Sense aquesta energia es dóna un procés autoorganitzatiu que porta cap a sistemes dels quals l’home treu poc profit.
Eduard Clavero, original de l’autor
La quantificació del magatzem i del flux d’aigua i de nutrients en els ecosistemes agrícoles no solament té un interès teòric sinó que permet de corregir alguns dels problemes creats per una agronomia massa industrial, que ha arribat a perdre de vista aspectes ecològics importants. Els balanços que es poden calcular en aquests diagrames reflecteixen el mal funcionament d’aquests sistemes. Així, per exemple, el càlcul d’un balanç entre l’aigua aportada per la pluja i el regatge, en una conca de vora 4000 ha de conreus, a la vall de l’Ebre, ha permès de saber que el nivell de regatge era excessiu i, a més a més, donava lloc a la mobilització de 76 000 tones de sals a l’any, que en gran part provenen de la dissolució dels guixos, i que contribueixen a la salinització dels rius receptors i dels conreus que s’hi reguen. L’anàlisi química de l’aigua d’un gran nombre de pous de l’Horta de València ha posat de manifest concentracions molt altes de nitrats, moltes de les quals estan per damunt del límit de potabilitat i s’atribueixen en gran part a la continuada aportació en els últims anys d’un excés d’adobs nitrogenats.
Aquest enriquiment en nitrogen dels sòls i els aqüífers més superficials és un fet molt general als països amb una agronomia intensiva, i fins i tot, en alguns casos, continua la migració d’aquest nivell d’alta contaminació en nitrats cap als aqüífers de més gran fondària, la contaminació dels quals tindrà efectes imprevisibles.
Un altre factor peculiar dels agroecosistemes és la introducció de substàncies tòxiques utilitzades com a insecticides o com a herbicides, moltes de les quals tenen una gran persistència, bé com a molècules bé com a fragments, molt sovint també tòxics, que poden passar a incorporar-se a la xarxa tròfica i a acumularse en els organismes de nivells tròfics superiors. La interferència de l’ús excessiu de biocides en els ecosistemes naturals localitzats a prop d’agroecosistemes es dóna en moltes ocasions als arrossars, els quals solen estar envoltats d’aiguamolls on es produeixen acumulaments faunístics importants, sobretot ocells.
A continuació, la taula compara diverses característiques, abiòtiques i biòtiques, dels ecosistemes naturals i dels de conreu que influeixen directament en llur propensió a acumular nutrients i a persistir al llarg del temps (font: E.T. Odum).
| CARACTERÍSTIQUES | ECOSISTEMES | |
|---|---|---|
| NATURAL | DE CULTIU | |
| ABIÒTIQUES | ||
| Taxes d’infiltració | alta | baixa |
| Taxa d’escorriment superficial | baixa | alta |
| Erosió | poca | molta |
| Pèrdua d’aigua del sòl per transpiració | alta | baixa |
| Quantitat de col·loides del sòl | molts | pocs |
| Pèrdua d’aigua per lixiviació | baixa | alta |
| Temperatura del sòl | baixa | alta |
| BIÒTIOUES | ||
| Reciclatge intern de nutrients per les plantes | alt | baix |
| Sincronia de l’activitat de les plantes i els microorganismes | alta | baixa |
| Diversitat temporal de l’activitat dels organismes | alta | baixa |
| Activitat de les plantes/activitat dels microorganismes | 1 | |
| Diversitat estructural de les plantes | alta | baixa |
| Diversitat genètica | alta | baixa |
| Potencial reproductiu | alt | baix |
Els efectes negatius d’una agricultura molt més abocada cap a la immediata rendibilitat econòmica que no pas a cercar el manteniment d’un cert nivell de producció a llarg termini, va generant un corrent d’interès cap a cultius més d’acord amb els principis ecològics. Els darrers anys, centres de recerca ecològica de prestigi, que abans estaven exclusivament dedicats a l’estudi d’ecosistemes naturals, han començat a incloure en els seus programes de recerca els agroecosistemes. Entre les nombroses recomanacions per a aconseguir que els agroecosistemes s’assemblin més als ecosistemes naturals convé destacar les referents a l’increment de la retenció de nutrients i el seu reciclatge; la disminució de l’ús d’herbicides i insecticides, especialment els d’ampli espectre i la reducció de l’erosió i la preservació, en la mesura que pugui, de la qualitat de les aigües de regatge. En aquesta nova orientació de l’agricultura, fins i tot tècniques com ara la llaurada són qüestionades.
A continuació, la taula mostra dades de producció de diverses espècies de conreu, obtingudes als Estats Units (Califòrnia, Colorado, Washington) i publicades per R.S. Loomis i P.A. Gerakis.
| ESPÈCIE | PRODUCCIÓ (t/ha·any) |
|---|---|
| Blat de moro (Zea mays) | 26,6 |
| Melca (Sorghum) | 46,6 |
| Remolatxa (Beta vulgaris) | 33,8 |
| Alfals (Medicago) | 29,7 |
| Blat (Triticum) | 29,8 |
| Patatera (Solanum tuberosum) | 22,0 |
| Arròs (Oryza sativa) | 22,4 |
El cas dels tarongerars
Molts autors han reconegut la manca d’un enfocament ecològic en l’estudi dels conreus, malgrat la seva extensió, que com més va és més important. Els ecòlegs rarament han estat interessats en l’estudi d’aquests ecosistemes artificials i així, mentre que hi ha dades abundants sobre l’ecofisiologia de les espècies conreades i dels seus enemics —plagues animals i males herbes— i sobre els mètodes de lluita per a aconseguir conreus purs i sans, en poques ocasions els cultius han estat estudiats com una agrobiocenosi —és a dir com a comunitats d’organismes que viuen tots plegats— en la qual s’estableixen una sèrie d’interaccions entre els organismes, similars a les existents als ecosistemes naturals. Les arrels d’aquest desinterès es podrien, potser, cercar en aquesta concepció de relacions força deterministes entre els elements de la biocenosi, com un símptoma de perfecció que es donaria, sobretot, en les comunitats clímax.
Una de les poques excepcions del rebuig existent a iniciar l’estudi de la biocenosi de conreus s’ha donat, precisament, en el tarongerar saguntí. Alguns aspectes de la dinàmica dels nutrients en aquest agroecosistema han estat aclarits molt recentment. Així, per exemple, amb la tècnica de l’isòtop 15N s’ha pogut comprovar que l’absorció de nitrogen per les rels és mínima a l’hivern i va creixent a la primavera fins a un màxim que coincideix amb la maduració del fruit. També s’ha posat de manifest que les reserves d’aquest element als òrgans vells (el 90% de les quals es troba entre les rels i les fulles) contribueixen amb un 70% al nitrogen que contindran els òrgans nous. També s’ha fet palès que els aminoàcids (sobretot asparagina, prolina i arginina) són la forma principal de nitrogen transportable, de la qual representen el 90%. Aquest tipus de dades permet el disseny de mètodes de regatge i adobament més eficients que els emprats fins ara i que, a més d’estalviar adobs, són menys perillosos per a la contaminació dels aqüífers amb nitrats.
En tarongerars adults s’han calculat balanços hídrics d’uns 720 o 750 mm/any, i eficiències de regatge entre el 50% i el 85%. L’alternativa de reg gota a gota dóna valors de producció molt similars als de reg amb inundació. Es calcula que, de mitjana, un taronger adult produeix uns 75 kg de fruita (pes fresc), i algunes estimacions molt provisionals sobre l’esporgada donen valors entre 60 i 70 kg de pes fresc per arbre i any.
L’ornitocenosi del tarongerar saguntí
Els cultius arboris tenen un paper interessant en la conservació d’espècies d’ocells i, en conjunt, apareixen com a sistemes més diversos i estructurats que els conreus herbacis. Als camps de tarongers s’ha pogut observar un total de vora cinquanta espècies d’ocells, incloent-hi les que hi viuen tot l’any, les de passada i, fins i tot, d’altres de més lligades a l’habitatge humà o que, pel fet de tenir un territori de vol molt ampli, sobrevolen sovint els tarongerars. Sembla que la colonització d’ocells en aquest cultiu arbori introduït no ha estat pas un problema. Així també ho han observat ornitòlegs francesos que comparaven l’avifauna de boscos naturals i de repoblacions, i arribaren a la conclusió que per als ocells són més importants les estructures que la taxonomia dels arbres, i en ambdós casos presentaven comunitats d’una complexitat comparable.
En les anàlisis de les comunitats, hom sol insistir en la manera com les poblacions que les constitueixen són capaces de dur a terme un cert paper ecològic més o menys característic, i en el fet que el conjunt d’aquests papers diversos es complementa per a assolir un ús millor dels recursos existents. En general, es parla de tres dimensions fonamentals en les quals té lloc la distribució de recursos: temps, espai i aliment.
Fenologia
Una idea de la distribució de la dimensió temporal entre els elements principals de l’ornitocenosi es pot tenir comparant, en l’interval total de nidificació, el nombre de nius actius de les quatre espècies dominants: el gafarró (Serinus serinus), el pardal comú (Passer domesticus), la merla (Turdus merula) i el verdum (Carduelis choris). Encara que estigui lluny d’una distribució modèlica que eviti qualsevol mena de superposició entre les èpoques de nidificació, la distribució obtinguda al tarongerar del Camp de Morvedre mostra poques superposicions dels pics entre les diferents espècies. El fet que el nombre total de nius en activitat variï proporcionalment menys al llarg del temps indicat per a les quatre espècies per separat mostra una distribució més regular en la comunitat que no pas en les poblacions de cada ocell.
Si es consideren les altres espècies nidificants, menys abundants, es pot veure també que n’hi ha que comencen la nidificació al març i l’acaben al juny, com la cadernera (Carduelis carduelis) o el trist (Cisticola juncidis), unes altres que nidifiquen entre l’abril i el juliol, com el cruixidell (Miliaria calandra), la mallerenga carbonera (Parus major) i el gratapalles (Emberiza cirlus) i, finalment, els que van del maig al juliol o a l’agost, com el rossinyol (Luscinia megarhynchos), la tórtora (Streptopelia turtur) i el papamosques gris (Muscícapa striata).
Distribució dels nius
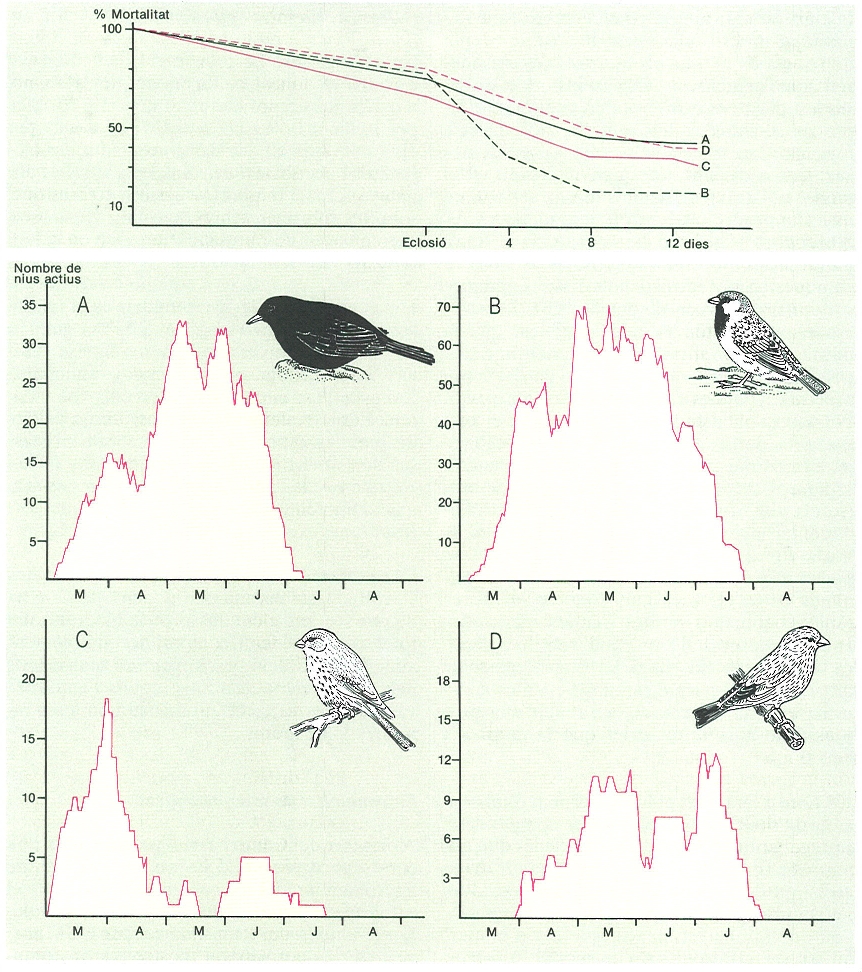
Corbes de mortalitat i nombre de nius de les quatre espècies dominants d’ocells en un tarongerar del Camp de Morvedre, segons les dades obtingudes a l’estació reproductora de 1977: A merla (Turdus merula), B pardal comú (Passer domesticus), C gafarró (Serinus serinus), D verdum (Carduelis chloris).
Amadeu Blasco, amb dades de J.A. Gil Delgado
En l’estudi de la distribució de l’espai en l’ornitocenosi veurem la distribució dels nius de les diferents espècies tant als diversos punts de l’estructura vertical del conreu com al lloc que ocupen en una projecció horitzontal.
En el conreu pròpiament dit s’han observat tres espècies d’ocells que fan nius a terra (cruixidell, rossinyol i trist), dues amb capacitat de nidificar sobre plançons (merla i cruixidell) i onze més que fan els nius sobre les branques, encara que la mallerenga carbonera pot aprofitar els forats dels arbres més vells. Altres espècies d’ocells, com són les orenetes, l’òliba i la puput, a més, és clar, del pardal comú, poden fer nius a les construccions existents al tarongerar.
Encara és possible de precisar més les preferències de les espècies per determinades parts de l’arbre que ofereixen les condicions més adients per a l’estructura de cada niu concret. D’una manera força senzilla es pot representar en un eix la situació del niu relativa a l’alçada de l’arbre (distància vertical del niu a terra/alçada de l’arbre) i en un altre eix la posició relativa a la branca (distància del niu a la base de la branca/longitud de la branca). Així, es pot veure com el gafarró mostra preferència pels extrems de les branques (la qual cosa fa que l’afectin les esporgades) mentre que, per exemple, la merla tendeix a construir els nius sobretot a les parts intermèdies, tant de l’arbre com de la branca. Precisament en aquesta última espècie s’ha observat una certa variació estacional en la localització dels nius, ja que al mes de juny els construeix a llocs més alts de l’arbre que no pas durant els altres mesos; en canvi, no s’han pogut demostrar variacions en l’orientació dels nius en els diferents mesos, ni l’existència d’orientacions preferents.
D’altra banda, també resulta interessant l’anàlisi de la distribució dels nius sobre un pla de l’horta en el qual els tarongers es consideren aproximadament equidistants. Són diverses les qüestions que es poden plantejar: el tipus de distribució dels nius de cada espècie (que pot ser aleatòria o amb els nius més agrupats o dispersos del que es preveu en el model a l’atzar), si els nius més propers a un que s’ha pres com a referència tendeixen a ser d’una altra espècie o de la mateixa i, finalment, si un niu d’una determinada època té com a veïns més propers nius d’altres èpoques o del mateix temps de construcció. L’anàlisi estadística senzilla posa de manifest que la merla, el verdum i el gafarró tenen una distribució aleatòria, mentre que el pardal comú presenta una clara tendència a l’agrupació. Pel que fa als nius veïns més propers, el pardal comú dóna una proporció més alta de l’esperada de nius veïns que pertanyen a la seva espècie, i les distàncies mínimes mitjanes entre els nius són les més petites de les trobades al tarongerar; el gafarró, al contrari, mostra una proporció més alta que l’esperada de nius propers de merla, i les distàncies mínimes entre els seus nius i els de les altres espècies són més petites que les mesurades entre nius propis. També amb relació a l’època de construcció, hom ha pogut constatar que hi ha una certa diferència: els nius construïts en una mateixa època tenen una proporció de nius veïns superior a la deguda a l’atzar.
A l’hora d’analitzar el funcionament d’una comunitat ornítica és important de comparar les característiques de les quatre espècies d’ocells més abundants. Per exemple, l’estudi de la grossària dels ous segons l’ordre de posta ha fet veure que el gafarró tendeix a fer més gros l’ou que fa quatre, mentre que per a la merla, sol ser el tercer i per al pardal el primer (almenys en les postes més nombroses); aquestes diferències han estat interpretades com a estratègies diferents, que en el cas del gafarró potser tendeixen a compensar la asincronia de la posta produint ous cada vegada més grans, en contrast amb el pardal comú que, en fer més gros el primer ou, seguiria el que podríem anomenar «l’estratègia de l’hereu». Un altre aspecte que pot il·lustrar diferències en les estratègies de vida d’aquests quatre ocells és la comparació de corbes de creixement obtingudes pesant els polls durant la seva estada als nius (dades dels anys 1977-78): el gafarró mostra una mortalitat més forta a la fase d’ou que els altres ocells; el pardal comú experimenta un increment molt notable de la mortalitat durant els quatre primers dies de vida i, també, però amb menys intensitat, els quatre següents; pel que fa al verdum i la merla, les corbes són força paral·leles.
Densitat, biomassa i producció
En els dos anys de prospecció més intensiva a l’horta de tarongers de Sagunt, les densitats d’ocells mesurades han estat de 151,6 parelles per 10 ha el 1976 i 172,3 el 1977. Aquests són els valors més alts de densitats ornítiques trobats als Països Catalans i dels més alts dels donats a l’Estat Espanyol (altres comunitats, sobretot forestals, tenen densitats més baixes però un nombre d’espècies més gran en la biocenosi); valors semblants als trobats a Sagunt es troben als boscos caducifolis europeus. L’espècie dominant al tarongerar és el pardal comú (unes 60 parelles), seguida del gafarró i la merla, amb densitats més o menys similars, i del verdum, una mica menys abundant. La resta d’espècies representen entre una vintena i una trentena de parelles. Les densitats de parelles esmentades representen biomasses de 1,09 kg (pes fresc) per ha l’any 1976 i 1,29 l’any 1977, que vénen a ser, en pes sec, uns 218 i 259 g/ha, respectivament, que correspon aproximadament a 10 i 12 mg C/cm2.
La producció de les quatre espècies dominants s’avalua, aproximadament, en 463,3 g/ha de pes sec, prop del doble de la biomassa del total de parelles. Un càlcul fet simplement amb la suma dels pesos màxims aconseguits pels components dels nius seguits l’any 1977 va donar un valor de producció una mica inferior, 318,3 g/ha.
La comunitat de micromamífers
Al mateix conreu del tarongerar saguntí s’ha constatat la presència de quatre espècies de micromamífers: tres rosegadors, que són el ratolí mediterrani (Mus spretus), la rata negra (Rattus rattus) i la rata comuna (R. norvegicus); i un insectívor, la musaranya comuna (Crocidura russula). Només dues, però, han estat objecte d’un estudi detallat en aquest ecosistema.
Pel que fa al ratolí mediterrani, sembla que en la selecció d’hàbitat dins del tarongerar mostra preferència per zones amb estrat arbori i amb acumulacions de terra (cèrcols, cavallons) per a la construcció dels caus, que força sovint presenten més d’un forat. La distribució a l’espai presenta un patró aleatori als mesos de gener, març i juny, mentre que als de maig i novembre, coincidint amb els moments de cria, és de tipus contagiós. El sistema de captura amb trampes innòcues permet d’investigar l’activitat quotidiana i els moviments dels individus marcats, així com posar de manifest en aquest medi l’existència dels dos màxims d’activitat coneguts en aquesta espècie, l’un en clarejar i l’altre en fer-se fosc; hom pot veure, també, que l’activitat nocturna minva a mesura que avança la nit. La captura d’una proporció més gran d’individus joves en hores de poca activitat fa sospitar que aprofiten aquests moments per a accedir a territoris d’adults dominants. Les dades de desplaçament d’individus mostren una capacitat superior dels mascles a recórrer distàncies grans en períodes de temps curts. L’anàlisi de l’estat de reproducció dels ratolins capturats ha fet palesa l’existència de femelles prenyades des del mes de març fins a la darreria d’octubre. Les densitats màximes de ratolins corresponen al mes de gener: són entre 75 i 100 mascles i entre 51 i 90 femelles, traduïdes a biomasses representen entre 288,5 i 428,2 g/ha; els valors intermedis que es poden calcular per any són prop dels trobats per a tota l’ornitocenosi.
Considerarem ara el cas de la rata negra i els seus nius aeris. Malgrat el fet de ser coneguda d’antuvi la capacitat arborícola d’aquesta espècie, hi ha pocs precedents en l’estudi dels nius aeris, un dels quals es va dur a terme, precisament, al tarongerar saguntí. S’ha vist que aquest rosegador pot construir nius als arbres amb materials del mateix taronger (branquillons amb fulles, branquetes i fulles), però que també pot aprofitar els nius del pardal comú i els de la merla. Pel que fa a les preferències de la rata negra per determinades orientacions o localitzacions dins la capçada de l’arbre, s’ha pogut veure una tendència a seleccionar més l’alçada que la situació de la branca. La distribució dels nius a l’horta és en forma colonial i hom sospita una certa variació estacional en la grossària dels nius agregats. Segons algunes dades de captures tant de rata negra com de rata blanca, a més de totes les localitzacions de nius, sembla clara la preferència de la primera pels llocs de conreu menys influïts per l’home, mentre que de la segona es van trobar dues colònies importants, una en un abocador i l’altra en un aljub abandonat.
El cas dels bladars de secà

Per a la selecció i millora de les varietats de blat, la proporció del gra respecte a la palla és una de les característiques que més interessa maximitzar, sempre tenint en compte que hi ha uns límits mecànics que permeten que la planta es mantingui dreta.
Josep M. Barres
Els conreus de blat (Triticum) són poc exigents. Només volen un sòl d’una certa fondària, que no tingui sorra, ni argila, ni tampoc un excés de matèria orgànica, i un índex d’acidesa comprès entre 5,5 i 7, encara que poden admetre valors més bàsics. Els blats de tardor necessiten una integral tèrmica d’entre 1850°C i 2375°C i, en general, no convé que la temperatura sigui molt freda a l’hivern, encara que, si les plantes estan poc desenvolupades, els freds extrems poden tenir efectes positius per a la formació de les arrels. Tampoc no són convenients les temperatures molt elevades a la primavera i als moments finals de la maduració. Pel que fa a la precipitació, aquests conreus necessiten entre 500 i 600 mm/any, però amb 300 o 400 ja es poden obtenir bones collites, sempre que la distribució de pluja sigui més abundant durant la primavera que a l’hivern. El blat és una planta anual; per tant, aquest ecosistema agrícola presenta unes variacions estacionals molt més patents que els conreus d’espècies llenyoses.
Dinàmica i funcionalitat
La sembra del blat es fa a la tardor, a les zones més fredes més aviat que a les més càlides, i després d’una labor de llaurada que sol combinar-se amb un adob de fons. A les zones més temperades, hom espera per llaurar que la pluja de la tardor faci créixer les males herbes. Per a obtenir entre 300 i 400 espigues/m2, en terrenys de fertilitat mitjana, calen entre 60 i 200 kg de llavors/ha. La quantitat apropiada d’espigues depèn molt del mètode i de l’època de sembra, de la qualitat del terreny, de la precipitació, etc.
El cicle vital del blat comença amb un període de germinació que es dóna quan les llavors absorbeixen entre un 25% i un 65% del seu pes d’aigua. El moment òptim per al començament d’aquest procés correspon a una saturació del sòl que oscil·la entre el 60% i el 80% de la capacitat de camp i a un interval de temperatures de vora 30°C, que té l’òptim entre 20°C i 25°C. Quan és a punt d’aparèixer la quarta fulla té lloc la guaixada a partir d’una sèrie de borrons laterals. Aquest procés es veu limitat tèrmicament i per això sol fer-se millor a les sembres més avançades, encara que també depèn de la disponibilitat de nutrients i té un cert determinisme genètic. Les últimes fases de desenvolupament del blat són la guaixada, l’espigada i la maduració. En aquestes fases les plantes acumulen més del 75% de la seva matèria seca, que es distribueix en una proporció d’entre 1:1,5 i 1:2,5 entre gra i palla, segons una sèrie de factors, molts dels quals són genètics.
Balanços energètics i de nutrients
Encara que mantinguin un aspecte semblant, els camps de blat han estat sotmesos a canvis importants en les tècniques de conreu al llarg dels darrers decennis, amb un increment considerable de les aportacions energètiques externes. Paral·lelament, la producció primària ha passat des de valors anuals de prop dels 500 kg/ha·any als segles XIII i XIV fins als quasi 4000 kg/ha·any al secà i més de 6000 kg/ha any amb reg. En una sèrie de balanços energètics calculats en camps de blat dels Estats Units, les entrades per combustibles, electricitat, adobs, bíocides, etc. oscil·laven entre 2000 i 18 000 Mcal/ha, mentre que, d’altra banda, la producció representava entre 3000 i 18 000 Mcal/ha. El rendiment energètic (Mcal produïdes/Mcal emprades) oscil·la entre 0,43 i 4,64. L’exemple de la vall de l’Ebre se situa al rang de rendiments alts.
Hi ha força informació sobre les necessitats d’adob dels conreus de blat. Alguns elements, com el nitrogen, són especialment necessaris en certes fases del cicle vital de la planta; els continguts que es troben a les llavors i a la palla donen una informació útil per a valorar l’eficàcia de l’adob. Les corbes de resposta en experiències de camp són un dels mètodes més utilitzats per a determinar les quantitats idònies de nutrients que s’hi han d’afegir. Molt sovint, l’excés d’algun element representa una minva de la producció.
L’anàlisi dels diferents fluxos que constitueixen els cicles d’elements als conreus de blat s’ha dut a terme a Anglaterra i a Israel, i cal destacar-ne alguns aspectes. Al cicle del nitrogen, per exemple, les entrades anuals de fixació des de l’atmosfera es consideren de prop de 5 kg/ha, mentre que les aportacions d’origen atmosfèric varien entre 5 i 17 kg/ha. Les pèrdues avaluades per volatilització són, anualment, de prop de 18 kg/ha i hi ha poques mesures de les taxes de desnitrificació. En el cas del potassi les entrades atmosfèriques són més modestes (de prop dels 3 kg/ha), però queda compensat per la meteorització de la roca que n’allibera al sòl entre 30 i 35 kg/ha. Pel que fa al fòsfor, ambdós fluxos són petits: menys d’1 kg/ha any.
A continuació, la taula presenta dades de la proporció de gra i palla en tres cereals i percentatge de diferents nutrients localitzats en aquestes fraccions, segons dades compilades per l'autor. Cal destacar l’enriquiment de nitrogen i fòsfor a les llavors.
Proporció de gra i palla i percentatge de nutrients en blat, blat de moro i ordi
| % PRODUÏT PER LA PLANTA | % | ||||||
|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | |||
| Blat | |||||||
| gra | 52,2 | 60,0 | 78,8 | 18,7 | 16,2 | 37,0 | |
| palla | 47,8 | 40,0 | 21,2 | 81,3 | 83,8 | 63,0 | |
| Blat de moro | |||||||
| gra | 56,0 | 71,8 | 87,2 | 24,7 | 42,8 | 38,8 | |
| palla | 44,0 | 28,2 | 12,8 | 75,3 | 57,2 | 61,2 | |
| Ordi | |||||||
| gra | 50,7 | 58,9 | 78,7 | 22,5 | 16,5 | 53,7 | |
| Palla | 49,3 | 41,1 | 21,3 | 77,5 | 83,5 | 46,3 | |
Les comunitats

El conreu de blat ha anat minvant de superfície d’ocupació a partir del final dels anys seixanta, i ha estat substituït sobretot per l’ordi. Malgrat això, la producció no ha disminuït, ja que els rendiments han anat creixent tot aquest temps.
Jaume Orta
Als conreus de cereals es donen les comunitats arvenses dites segetals o dels sembrats (Secalion mediterraneum) formades per moltes plantes d’origen oriental; algunes produeixen granes que s’assemblen molt a la del blat, la qual cosa fa difícil de destriar-les, i d’altres les fan molt petites i contaminen fàcilment les llavors destinades a la sembra. També hi ha una sèrie d’espècies que acompleixen el seu cicle vital abans que el blat i per això dispersen les seves llavors abans de la collita. Entre les espècies arvenses més freqüents a les nostres messes cal esmentar la rosella (Papaver rhoeas), les llunetes (Biscutella auriculata), les espunyidelles o raboses (Galium spurium i G. tricorne), el jull de fulla estreta (Lolium rigidum), la rosella de flor morada (Roemeria hybrida), els morrons (Stellaria media), el matablat (Iberis amara) i els esperons de gat (Lithospermum arvense), entre d’altres. La lluita establerta durant els últims anys contra aquestes males herbes ha fet minvar de manera important la diversitat específica d’aquestes comunitats. La pèrdua del caràcter policrom, d’un innegable valor estètic, dels camps de blat, ha tingut, si més no, la compensació de l’increment dels rendiments agronòmics. Als camps de blat, s’hi fan una sèrie de fongs considerats patògens que poden afectar diverses parts de la planta, com són els rovells (Puccinia), càries (Tilletia) i carbons (Ustilago), entre d’altres.
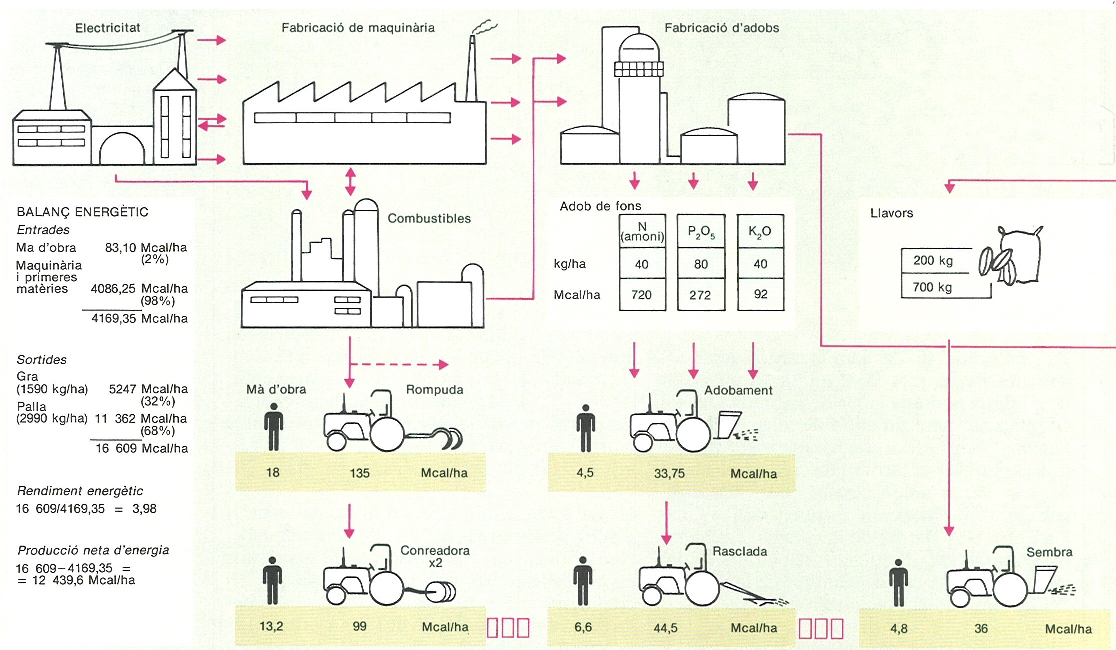
Balanç d’entrades i sortides d’energia en els bladars de secà.
Román Montull / Eduard Clavero, amb dades facilitades pels autors.
No es coneix gaire la composició faunística d’aquests conreus llevat, naturalment, d’aquelles espècies que poden considerar-se com a plagues, com ara les xinxes Aelia i Eurygaster, el mosquit del blat (Mayetiola destructor), alguns himenòpters de la família dels cèfids i certes larves de coleòpters crisomèlids. En un dels pocs estudis sobre la fauna invertebrada de les messes, fet a Anglaterra, es va trobar que els ordres d’insectes dominants a la fauna macroscòpica diürna eren els hemípters, els dípters i els coleòpters, seguits de prop pels himenòpters, amb una densitat que oscil·lava entre 250 i 820 individus/m2. Entre els hemípters dominaven els àfids, dels quals es van arribar a trobar densitats màximes de fins a 6000 individus/m2. Les famílies de dípters més ben representades als conreus són els cecidòmids, micetofílids, empídids, loncoptèrids, drosofílids i cloròpids. Les densitats apreciades, molt variables, van des de menys de 10 individus/m2, fins vora els 1000 individus/m22. L’estructura tròfica dels dípters en el sistema és dominada pels fitòfags o, alternativament, pel grup que reuneix sapròfags, escatòfags, micetòfags i necròfags, seguits pels entomòfags predadors. En l’ordre dels himenòpters, les famílies representades per un major nombre d’individus són els bracònids i calcídids, i en els coleòpters, els criptofàgids, estafilínids i latrídids. Els conreus de cereals es poden considerar com a prats modificats i, així, l’estabilitat de la seva fauna invertebrada es podria explicar per l’associació amb les poblacions del prat.
La fauna vertebrada és formada per conills i llebres, micromamífers (rates, ratolins i talpons) i, com a predadors, serps (sobretot la serp blanca), mustèlids i guineus. Entre els ocells, s’hi troben aloses, piules i especialment perdius i guatlles, de les quals hom ha citat densitats d’entre 0,02 i 0,18 individus/ha.
El cas dels arrossars

Els arrossars es poden considerar ecosistemes amb un funcionament regulat per l’acció humana per mitjà del control del cicle de creixement de l’arròs. A l’època d’inundació, a l’estiu, entre les plantes d’arròs creix una comunitat vegetal característica (sobretot, algues filamentoses flotants, a la fotografia, corresponent a un arrossar del delta de l'Ebre) i unes comunitats planctòniques formades per espècies de cicle vital curt.
Guillem Verger
L’arrós és un cereal de primera importància mundial, ja que el seu gra forma part de la meitat de la dieta de 1600 milions de persones, i d’entre una quarta part i la meitat de la dieta de 400 milions més. Els arrossars ocupen l’11% de la superfície conreada al món, situada entre 49° N i 38° S de latitud, majoritàriament al sud-est asiàtic. És conreat des de l’alta muntanya fins al nivell del mar, en terrenys de secà, inundats o bé surant a les aigües de fins 3 m de profunditat. De totes aquestes modalitats, la més generalitzada és el conreu per inundació, i és l’única que té lloc al nostre país on, com en altres països riberencs de la Mediterrània, forma part tradicional dels seus paisatges i la seva cultura.
L’arròs té uns orígens antiquíssims situats a l’hipercontinent de Gondwana. Hom creu que les espècies primitives d’arròs devien derivar cap a diferents hàbitats geogràfics en fragmentar-se aquest continent per a donar lloc a l’Àfrica, l’Antàrtida, Austràlia, Madagascar, Amèrica meridional i sud i sud-est asiàtic. Actualment se’n cultiven dues espècies amb un gran nombre de varietats cadascuna: Oryza sativa d’Àsia i O. glaberrima d’Àfrica occidental. Les subspècies cultivades d’O. sativa són tres: indica (zones tropicals), japonica (zones temperades) i javanica (zones de clima equatorial). Les primeres formes conreades eren de la primera. S’han trobat restes fòssils a l’E de la Xina i al N de l’índia. La posterior dispersió des de l’índia a l’Orient Mitjà, al N d’Àfrica i posteriorment a Europa va començar uns 1000 anys aC. De l’índia va passar a Madagascar i l’E d’Àfrica. Els EUA varen importar la llavor de Madagascar i del SE asiàtic. Els arrossos africans (Oryza glaberrima), originaris del delta del Níger, no han arribat a tenir la importància econòmica del grup asiàtic. El seu centre primari de dispersió s’estén de la zona pantanosa de la conca superior del Níger fins a dues àrees properes de la costa de Guinea.
El cultiu de l’arròs a la península Ibèrica va ser introduït pels àrabs al segle VIII i tingué importància a les terres pantanoses properes a l’albufera de València. Quan Jaume I va conquerir les terres valencianes (1233-38) l’arròs hi ocupava una gran extensió. Des de llavors fins ara s’han succeït nombroses alteracions de la superfície de conreu per raons particulars o per la implantació de lleis basades en raons sanitàries, agronòmiques i econòmiques. Al final del segle XIX es conreaven 25 000 ha a València i 3500 ha al delta de l’Ebre. A partir de l’any 1939 el conreu s’estén als maresmes del Guadalquivir i a altres llocs de Múrcia, Badajoz, Cadis, Osca, Lleida, Saragossa, Girona.
El conreu requereix unes condicions climàtiques especials. Per al seu bon desenvolupament cal una temperatura que oscil·li entre 18°C i 25°C. Cadascuna de les fases requereix un marge de temperatura adequat: la floració entre 19°C i 25°C i la fructificació entre 18°C i 20°C. A la zona de la Mediterrània, l’arròs necessita entre 130 i 180 dies per a completar el seu cicle vegetatiu. Amb les condicions esmentades, el conreu té lloc d’abril a octubre. Les terres més apropiades per al conreu de l’arròs són les al·luvials, les argil·localcàries i les argil·losilícies. Generalment, el pH del sòl oscil·la entre 4 i 6 però també es conrea arròs en sòls alcalins (Egipte, Califòrnia). El pH òptim és aproximadament 6,5. En zones com ara Nigèria, els baixos valors de pH poden donar llocs a la formació de compostos de ferro molt tòxics. Al SE asiàtic es duen a terme dos conreus per any: en un s’aprofita l’aigua procedent de les pluges de l’època humida dels monsons i, en l’altre, durant l’època seca, l’aigua necessària prové dels rius. Als països desenvolupats, la pluja pot considerar-se com un factor secundari per al conreu, ja que l’aigua és subministrada en abundància pels canals d’irrigació. Aquests canals provenen de rius i estanys de poca fondària que permeten que l’aigua tingui la temperatura adequada. L’aigua freda és perjudicial per al conreu de l’arròs per la qual cosa en cas de necessitat ha de ser escalfada en dipòsits abans de ser utilitzada.
Dinàmica i funcionalitat
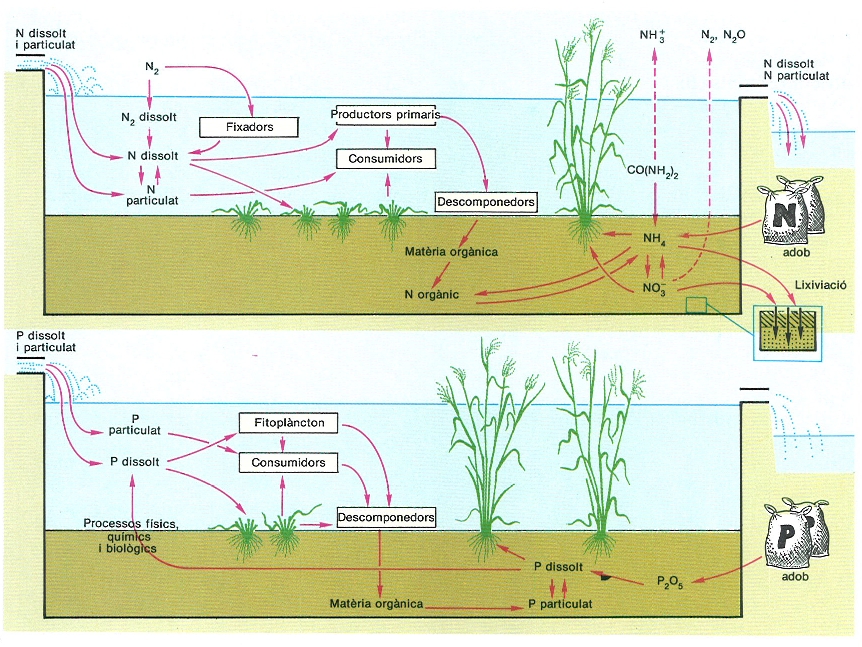
Esquemes comparatius simplificats dels cicles del nitrogen i del fòsfor en els arrossars.
Amadeu Blasco, original dels autors.
Els arrossars poden ser considerats com a ecosistemes aquàtics temporals de poca fondària, amb una fase seca durant l’hivern. Aquesta periociditat és inversa a la de la majoria dels ecosistemes aquàtics temporals naturals que, a les nostres latituds, resten secs durant l’estiu. Destaca, com en tota explotació agrícola, l’acció reguladora de l’home, que representa una aportació d’energia auxiliar, en forma d’adob, biocides i treball, per a potenciar la producció d’arròs.
L’arròs es conrea en un sòl molt planer on la capa d’aigua no ultrapassa els 15 o 20 cm de gruix. L’arrossar és dotat d’un canal d’irrigació i un altre d’evacuació que permeten la renovació continuada de l’aigua.
Un cop inundats els camps, al final d’abril o al començament de maig, segons la climatologia anual, té lloc la sembra de l’arròs. La germinació es produeix entre els 7 i 14 dies següents. Les fulles es van allargant fins a emergir a la superfície de l’aigua. Entre mitjan i final de juny apareixen una gran quantitat de tiges (de 5 a 7 per planta). La planta va creixent molt ràpidament durant el mes de juliol fins a arribar a la màxima alçada; a l’agost apareix l’espiga i es produeix la fertilització i posterior formació del gra a mesura que avança el conreu. Durant el mes de setembre, l’arròs experimenta la fase de maduració. I finalment, és el grau d’assecament del gra el que determina la data de collita.
Les anàlisis del contingut de nutrients de la planta d’arròs en diferents estadis de creixement indiquen que, com en tot agroecosistema, el nitrogen té un paper principal en la limitació de la producció. Aquest és, doncs, l’element predominant en la composició dels adobs que s’empren actualment; solament en determinats sòls àcids pot arribar el fòsfor a ser l’element limitant de la producció. Als sòls inundats, la forma assequible de nitrogen és l’amoni del sòl (el nitrogen en forma de nitrats és inaccessible per a la planta); la forma més emprada és el sulfat d’amoni, la urea o la cianamida. S’ha observat també un requeriment important de manganès. Abans de la sembra s’incorpora l’adob mineral al sòl (500 kg/ha aproximadament). Durant les primeres fases de creixement l’arròs acumula fòsfor i nitrogen a les fulles i posteriorment els trasllada a l’espiga en formació. Aquesta acumulació de nutrients i l’activitat de les poblacions biològiques acompanyants de l’arròs provoquen l’esgotament d’adob al sòl, per la qual cosa cal un suplement de nitrogen per a continuar el creixement; generalment s’empra adob foliar (urea principalment, 1,25 l/ha) per a facilitar una captació més ràpida. Durant la resta del conreu, la mineralització de la matèria orgànica present al sediment pot ser la font principal de nodriment per a l’arròs. En països del SE asiàtic o a l’índia, una font important de nitrogen són els processos de fixació duts a terme per algues cianofícies, lliures o simbionts amb el pteridòfit Azolla. Pel que fa al fòsfor, les anàlisis del contingut a la planta amb relació a la quantitat incorporada a l’adob de fons (50 kg/ha) fan pensar que aquest element hi és en excés. Als agroecosistemes conreats, una part molt important d’aquest fòsfor resta retinguda en forma d’apatites insolubles, per la qual cosa es requereix una aportació important d’aquest element cada temporada de conreu.

Les relacions ecològiques en els arrossars, i en tota mena de sistemes agrícoles en general, tendeixen a simplificar-se respecte dels sistemes sense explotació intensiva, però no deixa d’haver-hi casos d’organització típics, com aquest exemple de mutualisme que s’estableix entre diverses espècies d’ocells i l’home en el moment d’aixafar la palla i enterrar-la després de la collita de l’arròs.
Lluís Ferrés
Com a conseqüència de la seva elevada transpiració (6,5 mm/dia), l’arròs es conrea en sòls inundats dotats d’una entrada i una sortida d’aigua. Aquesta situació permet de considerar-los com a ecosistemes aquàtics on poden calcular-se balanços de nutrients com a diferències de concentració entre les sortides i les entrades. Les concentracions de fòsfor soluble i nitrogen inorgànic en forma de nitrats i nitrits al canal d’entrada sempre són superiors a les del canal de sortida; això pot indicar retenció o pèrdua d’aquests nutrients als arrossars. Ben a l’inrevés succeeix per al nitrogen en forma d’amoni, del qual s’observen concentracions més elevades als canals de sortida que als d’entrada. Aquestes diferències potser indiquen una elevada taxa de descomposició de la matèria orgànica al si dels arrossars. Durant els primers estadis de creixement de l’arròs, l’aigua dels arrossars es caracteritza pels elevats valors d’oxigen dissolt i pH, mentre que els valors d’alcalinitat són baixos. Durant els primers dies, les concentracions de nutrients dissolts (fosfats, nitrats i nitrits) són elevades, procedents de la dissolució de l’adob mineral, la qual cosa representa la pèrdua d’una fracció dels nutrients incorporats.
A partir del juliol, un cop l’arròs ha arribat a la màxima alçada (1 m) i s’ha desenvolupat un tapís important de vegetació submergida a l’arrossar, l’aigua es caracteritza pels seus elevats valors d’alcalinitat i les baixes concentracions d’oxigen dissolt i de pH; és abundant el fòsfor dissolt procedent de la redissolució dels carbonats i escàs el nitrogen inorgànic. Les condicions reductores de l’aigua afavoreixen els processos de redissolució del fòsfor i desnitrificació del nitrogen. En estudis duts a terme en arrossars japonesos s’ha estimat que el procés de desnitrificació pot representar el 38% de les pèrdues de nitrogen del sistema.
Les comunitats
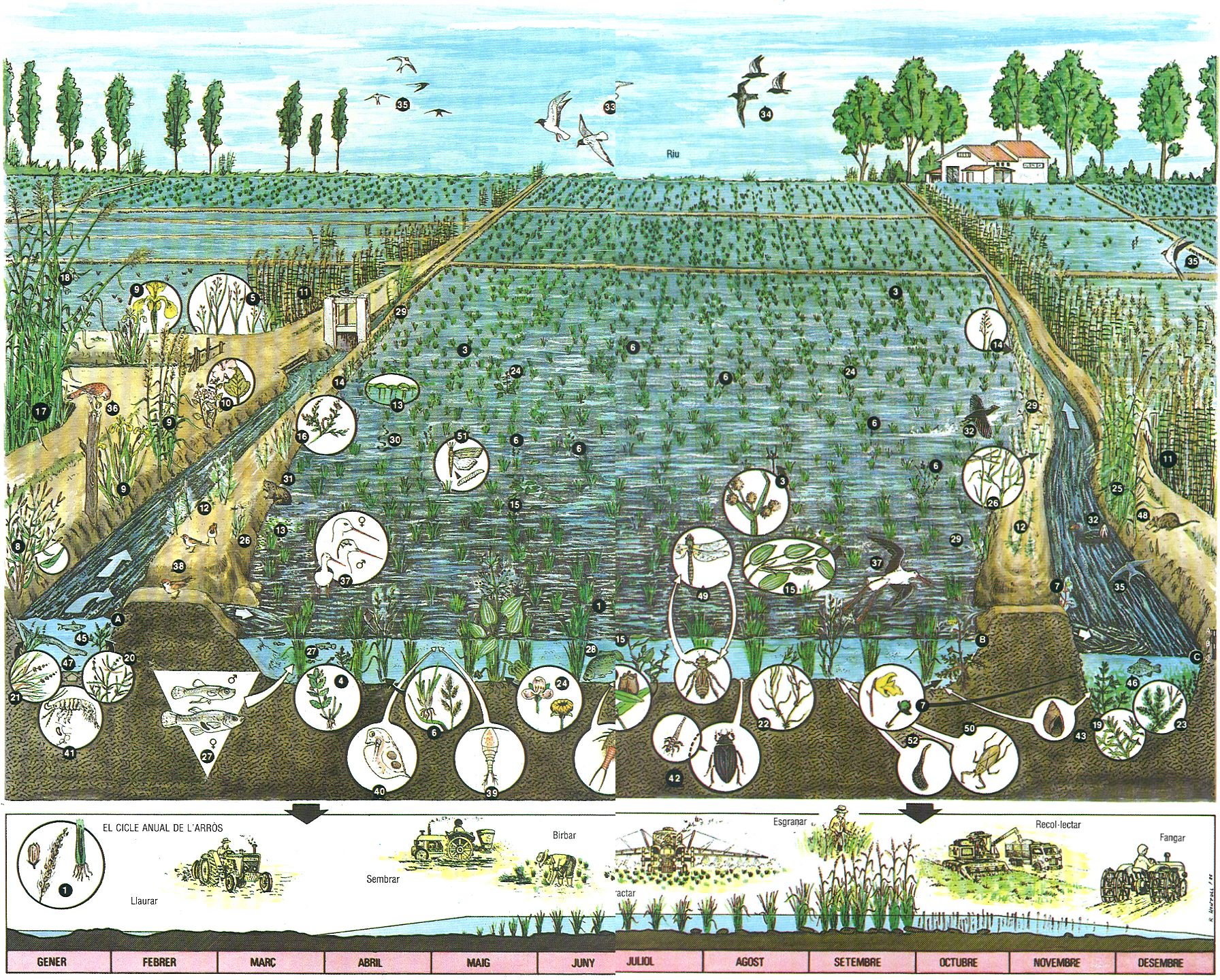
L’arrossar: estructura i espècies més representatives.1 arròs (Oryza sativa), 2 presseguera d’arrossar (Ammania coccinea), 3 jonca diforme (Cyperus difformis), 4 fàbrega (Lindernia dubia), 5 panissola repent (Panicum repens), 6 panissola (Echinochloa crus-galli), 7 utriculària (Utricularia vulgaris), 8 herba presseguera (Polygonum persicaria), 9 lliri groc (Iris pseudacorus), 10 malví (Althaea officinalis), 11 canyís (Phragmites australis), 12 cua de cavall (Equisetum ramosissimum), 13 llentia d’aigua (Lemna gibba), 14 agrostis blanca (Agrostis stolonifera), 15 potamogèton (Potamogeton natans), 16 heliotropi (Heliotropium curassavicum), 17 canya (Arundo donax), 18 corretjola gran (Calystegia sepium), 19 Najas marina, 20 i 21 potamogètons (Potamogeton pusillus, P. pectinatus), 22 Zannichellia palustris, 23 volantí (Ceratophyllum demersum), 24 plantatge d’aigua (Alisma plantago), 25 Arundo plinii, 26 agram (Cynodon dactylon), 27 gambúsia (Gambusia affinis), 28 carpa (Cyprinus carpio), 29 granota verda (Rana perezi), 30 colobra de collar (Natrix natrix), 31 rata d’aigua (Arvicola sapidus), 32 polla d’aigua (Gallinula chloropus), 33 gavina vulgar (Larus ridibundus), 34 fumarell negre (Chlidonias nigra), 35 oreneta vulgar (Hirundo rustica), 36 xoriguer (Falco tinnunculus), 37 cames-llargues (Himantopus himantopus), 38 pardal xarrec (Passer montanus), 39 Acanthocyclops vernalis, 40 Daphnia magna, 41 Gammarus aequicauda, 42 escarabat d’aigua (Hydrous piceus), 43 caragol d’aigua (Lymnaea), 44 tortugueta (Apus cancriformis), 45 madrilla (Chondrostoma toxostoma), 46 carpí ver (Carassius carassius), 47 anguila (Anguilla anguilla), 48 rata comuna (Rattus norvegicus), 49 espiadimonis (Cordulia linaenea), 50 nepa (Nepa cinerea), 51 barrinador de l’arròs (Chilo supressalis), 52 sangonera (Hirudo medicinalis).
Román Montull; Parc Natural del Delta de l’Ebre.
La presència d’aigua als camps ha afavorit l’establiment i la dispersió de nombroses espècies vegetals aquàtiques, moltes de les quals són d’origen exòtic i considerades com a males herbes per al conreu. La comunitat vegetal és molt semblant als arrossars de tot el litoral mediterrani. S’ha catalogat com a Cypero-Ammannietum coccineae, una comunitat de plantes palustres, rica en neòfits d’origen tropical, que té el desenvolupament òptim entre els mesos d’agost i octubre. Amb l’anàlisi detallada de l’estructura de la comunitat biològica hom s’adona de la seva complexitat; s’hi observen tres associacions vegetals integrades dins la mateixa: una associació d’helòfits emergents, una de plantes flotants (arrelades o no) i una altra de submergida, de macròfits i algues. L’ús d’herbicides al maig i al juny causa un empobriment important d’aquestes comunitats, però durant l’època de màxim desenvolupament (a l’agost i setembre) no és estrany d’anar trobant les espècies característiques dels arrossars.
Paral·lelament als canvis esdevinguts en les propietats físiques i químiques de l’aigua d’inundació hi ha una microsuccessió de les comunitats biològiques que s’hi desenvolupen, amb poques variants a l’àrea mediterrània.
Al cap de pocs dies de la inundació es desenvolupa un tapís de cianofícies a la superfície del sediment, que després és substituït per un altre d’algues filamentoses (Oedogonium, Mougeotia). Durant les primeres fases de creixement de l’arròs s’observa fitoplàncton en abundància, amb predomini d’algues clorofícies, volvocals i clorococals, i diatomees bèntiques (Carteria, Gonium pectorale, Pandorina morum, Ankistrodesmus falcatus, Cyclotella, i diverses espècies de Nitzschia i de Synedra). L’abundància de fitoplàncton afavoreix el desenvolupament de poblacions de zooplàncton filtrador amb predomini de cladòcers (Moina micrura, M. brachiata) i rotífers (Brachionus). L’aplicació de biocides, freqüent en aquest període, determina la desaparició dels cladòcers esmentats i dels nombrosos estadis larvals de l’ostràcode Heterocypris fretensis i del copèpode Acanthocyclops del grup vernalis, de manera que afavoreix l’increment del nombre de rotífers.
Paral·lelament al creixement de l’arròs, i amb més rapidesa, es desenvolupen diferents espècies d’helòfits (Echinochloa, Scirpus i Alisma plantago-aquatica) enèrgicament combatudes amb l’aplicació d’herbicides (molinat, propanil, entre d’altres). Durant els mesos de maig i juny, en avançar el conreu, el fitoplàncton esdevé poc abundant, cosa que coincideix amb el desenvolupament de vegetació submergida (Najas minor, Zannichellia palustris). En aquells camps on la circulació de l’aigua és més lenta es desenvolupa una cobertura vegetal neustònica, a base de llenties d’aigua (Lemna minor i L. gibba) i potamogètons (Potamogeton nodosus), que impedeix la formació d’un tapís submergit. És característica la presència de rotífers del gènere Lecane i del cladòcer Echinisca rosea, acompanyats d’alguns copèpodes (Acanthocyclops del grup vernalis) i ostràcodes (Herpetrocypris chevreuxi i Heterocypris fretensis). Durant els mesos de juny i juliol és general la presència als camps d’Hydrodictyon. Quan l’arròs ja ha assolit la màxima alçada, el fitoplàncton és poc abundant, amb presència d’euglenofícies (Euglena tuberculata, Trachelomonas volvocina i diverses espècies de Phacus), i s’observa el desenvolupament d’una densa vegetació constituïda per l’asprella Chara vulgaris i alguns helòfits d’origen tropical (Ammania coccinea, Bergia capensis, Lindernia dubbia i Utricularia vulgaris). Això determina la substitució dels organismes de tipus de vida planctònica i filtradora pels carnívors i detritívors de vida més bentònica. Són presents els copèpodes Thermocyclops dybowskii i Macrocyclops albidus i l’ostràcode Tanycypris. S’observa també un canvi d’estructura de la comunitat heleoplanctònica en el sentit d’un allargament de les cadenes tròfiques.
En conjunt es poden considerar els arrossars com a ecosistemes, on el creixement de l’arròs i, per tant, l’acció humana de control, determinen el seu funcionament. Per les seves característiques, poden servir com a model per a la investigació de processos químics i biològics d’estudi difícil en altres ecosistemes aquàtics.