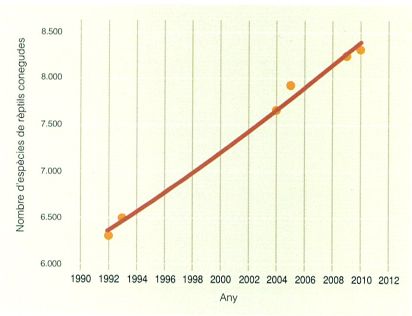
La quantitat d’espècies de rèptils conegudes ha anat augmentant de manera continuada les darreres dècades, ja que cada any se’n descriuen de noves. Això ha estat possible, entre altres raons, gràcies a la millora de les tècniques d’estudi i a l’ús d’anàlisis moleculars que permeten diferenciar espècies morfològicament molt semblants.
Dades dels autors.
El coneixement dels rèptils dels Països Catalans ha experimentat un avenç important els darrers anys. Al llarg d’aquest temps, el nombre de publicacions sobre herpetologia i el nombre d’investigadors dedicats a l’estudi de les comunitats reptilianes, del seu estat de conservació i de la taxonomia de les espècies han anat creixent de manera continuada. n’és una prova el fet que, en aquesta actualització, s’han inclòs 6 espècies noves de rèptils. Actualment es coneixen unes 8.300 espècies de rèptils arreu del món, la majoria de les quals pertanyen al grup dels escatosos. Tenint en compte que l’any 1990 se’n coneixien unes 6.000, l’increment ha estat de gairebé el 40%. Aquest augment ha estat possible no tan sols gràcies a l’increment del nombre d’investigadors sinó també al desenvolupament de tècniques d’anàlisi molecular en el camp de la genètica. Aquestes tècniques han permès dur a terme estudis taxonòmics, filogenètics i filogeogràfics que, en molts casos, han permès reconèixer espècies noves dins del que el 1987 es considerava una espècie única. D’altra banda, l’aparició d’un maquinari cada cop més potent ha permès desenvolupar els sistemes d’informació geogràfica (SIG) i altres eines de treball amb les quals és possible conèixer quines característiques ambientals determinen la distribució de les espècies i, en alguns casos, estudiar quines seran les problemàtiques ambientals que afectaran cada espècie en un futur proper.
La gran diversitat de rèptils presents als Països Catalans, i a la península Ibèrica en general, és resultat de diversos processos històrics de colonització, refugi i extinció. Totes les penínsules del sud d’Europa han constituït refugis climàtics durant els períodes glacials del darrer milió d’anys. Però, a més, la península Ibèrica ha permès que espècies africanes colonitzessin el continent europeu aprofitant els contactes que ha anat tenint periòdicament amb l’Àfrica durant els darrers deu milions d’anys. Aquesta combinació de canvis climàtics i geogràfics ha provocat fenòmens d’especiació al·lopàtrica, però també processos de colonització entre la península Ibèrica i el nord-oest d’Àfrica, tant en un sentit com en l’altre. El resultat final ha estat que la biodiversitat reptiliana dels Països Catalans és molt gran, com ho demostra el fet que hi ha representades 43 espècies de les 51 presents al conjunt de la Península i les Balears. Entre aquestes 41 espècies, les sargantanes del gènere Podarcis són les que mostren una major diversificació, no solament als Països Catalans sinó també a la península Ibèrica. Els estudis que actualment s’estan duent a terme sobre aquest gènere encara no són concloents, però semblen indicar que l’espècie que es coneix amb el nom de sargantana ibèrica (Podarcis hispanica) en realitat és un complex de diverses espècies. El gran nombre d’espècies i subespècies de rèptils presents actualment als Països Catalans demostra el gran èxit evolutiu d’aquest grup de vertebrats.
Ja s’ha comentat que la variada orografia i climatologia dels Països Catalans és un dels motius pels quals a casa nostra hi ha representades faunes reptilianes de diversos orígens. Tot i que el clima dominant als Països Catalans és de tipus mediterrani, la presència de serralades importants a la meitat nord del territori ha permès l’entrada de moltes espècies centreeuropees, com per exemple, la sargantana de paret (Podarcis muralis), i, alhora, ha afavorit l’aparició d’endemismes pirinencs. D’altra banda, el fet que als Països Catalans predomini el clima mediterrani, que es caracteritza per estius secs i calorosos i hiverns suaus, fa que les espècies termòfiles hi siguin més abundants i hi presentin àrees de distribució més àmplies.
La diversificació de les espècies i l’aparició de subespècies noves per adaptació a condicions orogràfiques i climatològiques diverses ha arribat a la màxima expressió a les Balears. En aquestes illes, la diversitat de medis i les condicions que afavoreixen l’aïllament s’han traduït en un ventall ampli de models evolutius, que van des de fenòmens de plasticitat fenotípica fins a processos de deriva, colls d’ampolla i models d’extinció i colonització. Per exemple, les nombroses subespècies de sargantanes descrites en aquest arxipèlag mostren models microevolutius i processos d’adaptació i de selecció per pressió o per deriva genètica que, per ells mateixos, mereixen ser conservats i analitzats. A més, la introducció d’altres petits saures a les Balears ha generat episodis de competència i d’exclusió competitiva.
La gran adaptabilitat i abundància dels petits lacèrtids ha fet que actualment aquests organismes s’utilitzin com a bioindicadors de l’estat dels ecosistemes, ja que, atesa la posició que ocupen en la piràmide tròfica i l’elevada taxa de renovació de les seves poblacions, reflecteixen ràpidament els canvis que es produeixen als ecosistemes de resultes de l’activitat humana.
Cal destacar, finalment, el reduït coneixement que es té de les poblacions d’ofidis dels Països Catalans, atesa la seva escassetat. Així i tot, els darrers anys la informació sobre les espècies de serps, fonamentalment les més comunes, s’ha incrementat notablement amb estudis sobre l’ecologia, la biologia, l’ús de l’espai i el domini vital d’aquests rèptils.
Els estudis de distribució i biogeografia dels rèptils
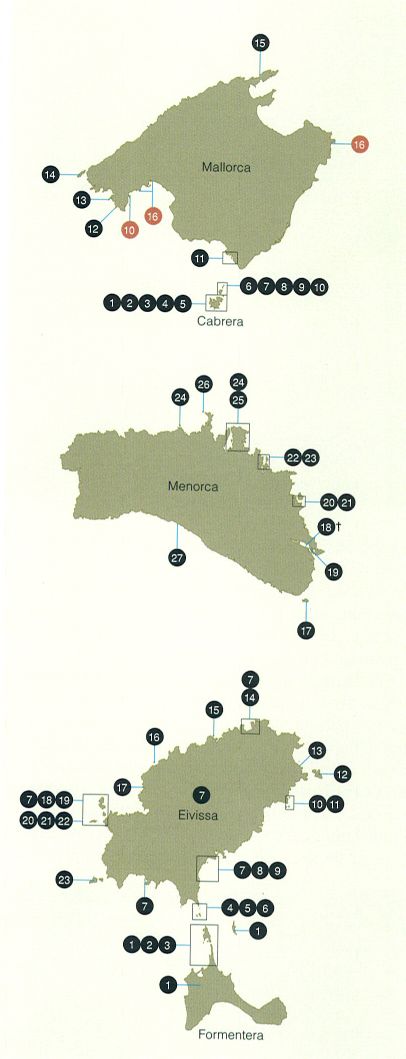
Distribució subespecífica actualitzada de les sargantanes gimnèsica i de les Pitiüses a l’arxipèlag balear. Els requadres indiquen conjunts de subespècies i illots no representables en detall per motiu d’escala. En color granat, poblacions introduïdes. Subespècies de sargantana gimnèsica o sargantana balear (Podarcis lilfordi) i poblacions introduïdes. Mallorca i Cabrera: 1 P. I. xapaticola. 2 P. I. estelicola. 3 P. I. imperialensis. 4 P. I. kuligae. 5 P. I. nigerrima. 6 P. I. conejerae. 7 P. I. espongicola. 8 P. I. planae. 9 P. I. pobrae. 10 P. I. fahrae. 11 P. I. jordansi. 12 P. I. toronis. 13 P. I. hartmanni. 14 P. I. giglioli. 15 P. I. colomi. 16 P. pityusensis pityusensis (introduïda). Menorca: 17 P. I. lilfordi. 18 P. I. rodriguezi †. 19 P. I. balearica. 20 P. I. ssp?. 21 P. I. brauni. 22 P. I. carbonerae. 23 P. I. addayae. 24 P. I. sargantanae. 25 P. I. porrosieola. 26 P. I. fenni. 27 P. I. codrellensis. Subespècies de sargantana de les Pitiüses (Podarcis pityusensis). Eivissa i Formentera: 1 P. p. formenterae. 2 P. p. torretensis. 3 P. p. gastabiensis. 4 P. p. ahorcadosi. 5 P. p. negrae. 6 P. p. caragolensis. 7 P. p. pityusensis. 8 P. p. ratae. 9 P. p. schreltmuelleri. 10 P. p. redonae. 11 P. p. canensis. 12 P. p. tagomagensis. 13 P. p. hortae. 14 P. p. canaretensis. 15 P. p. muradae. 16 P. p. hedwigkamerae. 17 P. p. calasaladae. 18 P. p. gorrae. 19 P. p. maluquerorum. 20 P. p. carlkochi. 21 P. p. kameriana. 22 P. p. frailensis. 23 P. p. vedrae.
Barreras & Creixell, a partir de Pérez-Mellado, 2009.
Pel que fa al coneixement sobre la distribució dels rèptils als Països Catalans, ha millorat força els darrers vint-i-tres anys, ja que cada vegada hi ha més herpetòlegs que duen a terme treballs de camp. Localitzar els rèptils és, en general, més difícil que localitzar els amfibis, ja que els primers no es concentren ni es fan visibles durant el període reproductor. Malgrat això, els coneixements més extensos sobre la biologia, els patrons d’activitat i els hàbitats preferits de molts rèptils han permès que, a hores d’ara, es disposi d’un coneixement molt precís sobre la distribució de la majoria d’espècies, fins i tot de les més difícils de veure.
La publicació de nous aties de distribució, tant d’abast local com general, ha permès dibuixar amb més precisió l’àrea de distribució de les espècies, primer mitjançant la projecció cartogràfica UTM (generalment en quadrícules de 10 × 10 km) i després localitzant de manera exacta els individus gràcies als aparells de GPS. Aquestes dues eines no solament han permès conèixer més bé les àrees de distribució, sinó que també han facilitat l’anàlisi de les característiques dels hàbitats, tant a nivell espacial com temporal. Cal afegir així mateix que, en molts casos, les modificacions més significatives de les àrees de distribució realitzades les dues darreres dècades són el resultat del coneixement d’aquestes distribucions a escala més petita. A més, sovint aquestes modificacions consisteixen en una fragmentació de l’àrea de distribució antiga, deguda a canvis en els usos del sòl imposats per l’activitat humana.
Actualment, l’única espècie autòctona de tortuga terrestre dels Països Catalans, la tortuga mediterrània (Testudo hermanni), té només una població natural consolidada, la del massís de l’Albera. Malauradament, les densitats de població d’aquesta espècie són cada cop més baixes i és possible que en desapareguin algunes si no s’apliquen plans de recuperació urgents. L’augment de la pressió humana a la zona de distribució, l’augment de la cobertura arbòria i la manca d’interès institucional són les causes de la reducció continuada dels seus efectius. Malgrat això, l’espècie no està en perill, ja que ha estat introduïda amb èxit en diverses localitats dels Països Catalans.

La nidificació de la tortuga babaua (Caretta caretta) als Països Catalans és un fet excepcional. Tot i que dades verbals indiquen que en altres temps aquesta tortuga havia nidificat a l’arxipèlag de Cabrera, només es té constància certa que això hagi ocorregut una vegada al País Valencià i dues vegades a Catalunya. A la foto s’observa una de les tortugues nascudes a la platja del Saler (València) l’any 2006.
Arxiu del Parc Natural de s’Albufera de Mallorca / Maties Rebassa.
A la Mediterrània hi viuen tres espècies de tortugues marines: la tortuga llaüt (Dermochelys coriacea), la tortuga verda (Chelonia mydas), i la més abundant, la tortuga babaua (Caretta caretta). Cap d’aquestes tortugues es reprodueix de manera habitual a les costes dels Països Catalans i només es té constància real de dues nidificacions. Al final del juliol del 2006 es va localitzar un niu de tortuga babaua a la platja de Puçol de València. L’11 d’agost un temporal el va deixar al descobert i es va traslladar a la platja de la Punta del Saler, on van néixer 37 tortugues. També al final d’agost del 2006 una altra tortuga babaua va niar a la platja del Pla de l’Os, de Premià de Mar, on van néixer 54 tortugues. És probable que en el passat aquesta espècie criés a les platges de les Balears, com ho denoten les descripcions verbals dels pescadors de Cabrera. Així mateix, hi ha citacions de possible cria de tortuga babaua al delta de l’Ebre, on es va trobar un nounat de tortuga babaua mort a la platja. En l’actualitat no hi ha dades de nidificadó sobre la tortuga babaua als Països Catalans, bé que aques-ta espècie és relativament abundant a les aigües de les Balears i al cap de Sant Antoni, al País Valencià, llocs que en l’època estival alberguen una elevada concentració d’exemplars de tortuga babaua i on els pescadors les capturen accidentalment, principalment entre els mesos de juny i juliol.

La sargantana àgil (Lacerta agilis) presenta una distribució molt reduïda i fragmentada, i la major part de les seves poblacions són molt petites. A Catalunya i al Vallespir-Conflent aquesta sargantana està amenaçada per les pistes d’esquí i les infraestructures que les acompanyen, que a poc a poc van eliminant els reduïts hàbitats disponibles per a l’espècie.
Albert Montori.

La sargantana cendrosa (Psammodromus hispanicus) és molt sensible a les alteracions de l’hàbitat, ja que les seves poblacions es renoven gairebé del tot d’un any a l’aitre. Aquesta espècie es distribueix per València i la franja costanera de Catalunya, bé que les poblacions litorals estan força fragmentades a causa de la pèrdua d’hàbitats.
Marc Franch.
Els saures han conquerit tots els hàbitats disponibles als Països Catalans però les espècies més grans s’han fet més rares a conseqüència de la pèrdua d’hàbitats i a la seva excessiva fragmentació, especialment a les zones periurbanes. La sargantana cendrosa (Psammodromus hispanicus), per exemple, ha reduït molt la seva presència al litoral a conseqüència de la manca d’hàbitats disponibles en aquesta àrea. En molts indrets on l’agricultura tradicional ha estat substituïda per una agricultura extensiva, la sargantana ibèrica pràcticament ha desaparegut, de vegades substituïda pel dragó comú (Tarentola mauritanica), espècie d’hàbits més nocturns que ara s’observa freqüentment de dia en substrats rocallosos, tant naturals com artificials, on abans abundava la sargantana ibèrica. L’àrea de distribució de la sargantana àgil (Lacerta agilis) també es dibuixa més petita que dues dècades enrere. En aquest cas, però, és perquè s’ha comprovat que en alguns indrets havia estat citada erròniament, confosa amb altres espècies. Actualment es considera que aquesta sargantana només es troba a Andorra, a la Cerdanya, al Ripollès i al Vallespir-Conflent. La colobreta cega (Blanus cinereus) es troba, bé que força dispersa, per tot el País Valencià. A Catalunya s’havia observat a l’extrem sud de la província de Tarragona (Ulldecona), però des del 1985 no se n’ha tornat a confirmar la presència. Actualment es considera que les citacions antigues al nord de l’Ebre s’han de descartar perquè no han pogut ser confirmades.

L’escurçó ibèric (Vipera latastei) ha anat desapareixent de forma progressiva de moltes localitats els darrers anys. Perseguit des de sempre pel fet de ser verinós, els efectius d’aquest escurçó també s’han anat reduint a mesura que s’intensificava l’ocupació humana dels seus hàbitats. Avui dia l’espècie només manté poblacions estables a les zones protegides de muntanya.
Vicent Sancho.
La distribució de la major part d’espècies d’ofidis és la mateixa, a grans trets, que es coneixia el 1987, amb una excepció important: l’escurçó ibèric (Vipera latastei). Aquest vipèrid és present des d’Alacant fins al límit entre les províncies de Barcelona i Girona. En general es tracta d’una serp molt escassa i rara de veure, que només manté poblacions d’una certa densitat en algunes localitats de la muntanya baixa o mitjana. Recentment se n’ha modificat la distribució peninsular i s’ha comprovat que l’activitat humana és la principal responsable de la desaparició de l’espècie a gran part de les zones no protegides dels Països Catalans, especialment al litoral. Per aquesta raó, la seva distribució actual està molt fragmentada.
La colobra llisa europea (Coronella austriaca) és una espècie l’àrea de distribució de la qual als Països Catalans ha estat molt ampliada i detallada. Es localitza de manera contínua des dels Pirineus fins al Montseny, però sense arribar, en cap a cas, a la franja costanera. A més, hi ha poblacions relictes a les muntanyes de Prades, als ports de Beseit, a Ares del Maestrat, al massís del Penyagolosa, al Toro i al Racó d’Ademús. Recentment, els models de distribució potencial fets per a la península Ibèrica han demostrat que, en les poblacions aïllades dels ambients més mediterranis, la colobra llisa europea es troba en condicions ambientals subòptimes, cosa que fa preveure una pèrdua progressiva de poblacions en el futur.
La taula següent presenta la llista patró dels rèptils presents als Països Catalans i estat de conservació de cadascun. Hi figuren els noms aprovats en la Lista patrón de los anfibios y reptiles de España i, entre parèntesis, els reconeguts amb anterioritat. A absent, CR en perill crític, DD dades insuficients, DE d’especial protecció, DIE d’interès especial, EN en perill, EP d’especial protecció, GE espècie amb plans de gestió específics, LC preocupació menor, NE no avaluada, NT gairebé amenaçada, P protegida, VU vulnerable. (*) indica que no estan protegides al territori corresponent. UICN Unió Internacional per a la Conservació de la Natura, CNEA Catálogo Nacional de Especies Amenazadas.
| Espècie | Nom comú | Catalunya | Illes Balears | País Valencià | La Franja | Rosselló / Vallespir | UICN (2008) | CNEA (2006) | Libro Rojo (2002) |
|---|---|---|---|---|---|---|---|---|---|
| Testudo hermanni | Tortuga mediterrània | EN | DIE | EN | A | VU | NT | DIE | EN |
| Testudo graeca | Tortuga mora | A | DIE | A | A | A | VU | DIE | EN |
| Emys orbicularis | Tortuga d’aigua europea | EN | EP | VU | VU | NT | NT | NE | VU |
| Mauremys leprosa (Mauremys caspica leprosa) | Tortuga d’aigua ibèrica | VU | NE * | P | DIE | EN | NE | NE | VU |
| Blanus cinereus | Colobreta cega | NE | A | NE | DIE | A | LC | NE | LC |
| Hemidactylus turcicus | Dragó rosat | DIE | DIE | DIE | NT | LC | DIE | LC | |
| Tarentola mauritanica | Dragó comú | DIE | DIE | DIE | LC | LC | DIE | LC | |
| Anguis fragilis | Vidriol | DIE | A | A | LC | NE | DIE | LC | |
| Acanthodactylus erythrurus | Sargantana cuarroja | VU | A | NE | A | LC | NE | LC | |
| Iberolacerta bonnali (Lacerta monticola) | Sargantana pirinenca | VU | A | A | VU | A | NT | VU | VU |
| Iberolacerta aurelioi (Lacerta monticola) | Sargantana pallaresa | EN | A | A | CR | EN | EN | EN | |
| Iberolacerta aranica (Lacerta monticola) | Sargantana aranesa | EN | A | A | EN | EN | EN | CR | |
| Psammodromus hispanicus | Sargantana cendrosa | DIE | A | DIE | NT | LC | DIE | LC | |
| Psammodromus algirus | Sargantana cuallarga | DIE | NE * | DIE | LC | LC | DIE | LC | |
| Podarcis muralis | Sargantana de paret | DIE | A | LC | LC | DIE | LC | ||
| Podarcis hispanica | Sargantana ibèrica | DIE | A | LC | NE | DIE | LC | ||
| Podareis pityusensis | Sargantana de les Pitiüses | NE * | DIE | A | DIE | NT | |||
| Podareis lilfordi | Sargantana gimmnèsica | A | DIE | A | DIE | EN | |||
| Podarcis atrata (Podarcis hispanica) | Sargantana dels Columbrets | A | A | VU | A | NE | NE | VU | |
| Podareis sicula | Sargantana siciliana (sargantana italiana) | A | EP | LC | LC | NE | LC | ||
| Scelarcis perspicillata (Lacerta perspicillata) | Sargantana marroquina | A | EP | A | LC | NE | DD | ||
| Timon lepidus (Lacerta lepida) | Llangardaix ocel·lat | VU | NE * | P | VU | NT | NE | LC | |
| Lacerta agilis | Sargantana àgil | EN | A | LC | NE | DIE | NT | ||
| Lacerta bilineata (Lacerta viridis) | Llangardaix verd o lluert | DIE | A | LC | LC | DIE | LC | ||
| Zootoca vivipara (Lacerta vivipara) | Sargantana vivípara | DIE | A | LC | LC | DIE | NT | ||
| Chalcides striatus (Chalcides chalcides) | Bívia tridàctila | DIE | A | LC | LC | DIE | LC | ||
| Chalcides bedriagai | Bívia ibèrica | NE | A | A | NT | NE | NT | ||
| Coronella girondica | Colobra llisa meridional | DIE | A | LC | LC | DIE | LC | ||
| Coronella austriaca | Colobra llisa europea | DIE | A | LC | NE | DIE | LC | ||
| Zamenis longissimus (Elaphe longissima) | Serp d’Esculapi | DIE | A | LC | LC | DIE | DD | ||
| Rhinechis scalaris (Elaphe scalaris) | Serp blanca | DIE | NE * | LC | LC | DIE | LC | ||
| Hierophis viridiflavus (Coluber viridiflavus) | Serp verda i groga | DIE | A | DIE | LC | LC | DIE | LC | |
| Hemorrhois hippocrepis (Coluber hippocrepis) | Serp de ferradura | DIE | NE * | A | LC | DIE | LC | ||
| Malpolon monspessulanus | Serp verda | NE | NE * | P | LC | LC | NE | LC | |
| Macroprotodon brevis (Macroprotodon cucullatus) | Serp de garriga (serp de cogulla) | A | A | A | NT | DIE | NT | ||
| Macroprotodon mauritanicus (Macroprotodon cucullatus) | Serp de garriga balear (serp de cogulla) | A | DIE | A | LC | DIE | NT | ||
| Natrix natrix | Colobra de collar | DIE | A | LC | LC | DIE | LC | ||
| Natrix maura | Serp d’aigua | DIE | GE | LC | LC | DIE | LC | ||
| Vipera aspis | Escurçó europeu | NE * | A | LC | LC | NE | LC | ||
| Vipera latastei (Vipera latasti) | Escurçó ibèric | VU | A | P | A | VU | NE | NT | |
| Caretta caretta | Tortuga babaua | EN | DIE | DD | EN | DIE | EN | ||
| Dermochelys coriacea | Tortuga llaüt | DIE | DIE | DD | CR | DIE | CR | ||
| Chelonia mydas | Tortuga verda | DIE | DIE | NE | EN | DIE | EN | ||
| Eretmochelys imbricata | Tortuga carei | NE * | NE * | NE | CR | DIE | DD | ||
| Lepidochleys kempii | Tortuga bastarda | NE * | NE * | NE | CR | NE | DD |
Els estudis ecològics
Els rèptils són un grup d’ecologia variada. Els saures, és a dir, les sargantanes i els llangardaixos, presenten una fisiologia i uns requeriments ecològics que no es poden comparar amb els dels quelonis o tortugues ni amb els dels ofidis o serps. És per això que es tractaran de forma diferenciada.
Els petits saures constitueixen un bon element de diagnosi de les alteracions que pot patir un ecosistema, ja que abunden a la major part d’ecosistemes dels Països Catalans, són fàcils de detectar, tenen taxes de renovació de la població altes i presenten períodes generacionals molt curts. Hi ha espècies que renoven els membres de les seves poblacions cada any, com per exemple la sargantana cendrosa, bé que la majoria d’espècies els renoven cada dos o tres anys. De manera semblant a com passa amb els amfibis, els petits saures ocupen posicions intermèdies de les cadenes tròfiques i són elements clau per al manteniment de les relacions interespecífiques dels sistemes naturals. D’altra banda, les poblacions de petits saures s’estructuren de manera fragmentada, especialment en aquelles zones on les distribucions no són contínues, com succeeix, per exemple, amb les poblacions de sargantanes del gènere Iberolacerta, que viuen en hàbitats extrems d’alta muntanya.
Una característica dels saures, i dels rèptils en general, és que necessiten termoregular-se. Com que són organismes ectoterms, han de captar calor de l’ambient exterior per a assolir una temperatura corporal òptima i poder estar actius. Aquest fet explica que la major part de rèptils mediterranis presentin un període de diapausa hivernal, durant el qual no realitzen cap tipus d’activitat. Aquest període depèn, sempre, del clima i les condicions meteorològiques. Per tant, a les zones meridionals més càlides dels Països Catalans els rèptils presenten períodes d’activitat gairebé continus al llarg de tot l’any, mentre que a les zones fredes mostren cicles d’activitat anuals més curts. Alguns cicles d’activitat poden reduir-se a tan sols quatre mesos, com els de les sargantanes del gènere Iberolacerta que viuen als Pirineus per sobre els 2.000 m d’altitud. Quan el cicle d’activitat no dura tot l’any, la major part de femelles retenen la posta a l’interior del cos, circumstància que facilita el desenvolupament de l’embrió i fa possible cercar les condicions de temperatura més adients perquè es completi el desenvolupament. La femella fa la posta quan la temperatura exterior és més alta, i aleshores el desenvolupament es finalitza ràpidament dins de l’ou. Aquest comportament és propi d’un gran nombre de lacèrtids, en els quals s’ha observat una relació inversa entre la temperatura de l’ambient i el temps que la femella reté l’ou.
Quan es parla de captació de calor, cal diferenciar la temperatura exterior de la temperatura del sòl. La major part dels rèptils petits són tigmoterms, és a dir, aprofiten l’escalfament del substrat per a captar calor. Això, juntament amb la mida corporal petita, és el que explica que es puguin trobar comunitats de saures a gran altitud, on aparentment les condicions ambientals són molt desfavorables. El fet s’entén si tenim en compte, per exemple, que mentre la temperatura de l’aire és de 6-7 °C, la temperatura d’una roca pot assolir gairebé els 20 °C. D’altra banda, el fet que hi hagi saures de mides molt diverses permet repartir els nínxols tèrmics i temporals entre les diferents espècies i mides, cosa que minimitza la competència interespecífica i intraespecífica.
Els ofidis o serps no abunden tant com els saures, ja que ocupen posicions més altes en la piràmide tròfica. Les espècies d’ofidis presents als Països Catalans tenen mides moderades, però són sempre més grans que els saures i presenten períodes d’activitat molt més curts. D’altra banda, són de costums molt més reservats i tenen densitats de població molt baixes. Normalment, després de passar una estona al descobert per a assolir la temperatura corporal òptima, passen desapercebuts, ja que realitzen les seves activitats entre la vegetació o sota el sòl. Fins i tot hi ha espècies que a l’estiu poden assolir la temperatura d’activitat sense haver d’abandonar el seu refugi. Per a la major part de colúbrids terrestres (serps dels gèneres Coronella, Rhinechis, Hierophis i Malpolon), les temperatures corporals òptimes se situen entre els 27 °C i els 32 °C; per als vipèrids, entre els 26 i els 29 °C, i per a les espècies aquàtiques del gènere Natrix, entre els 26 i 30 °C. Es tracta de temperatures força elevades que, en general, són difícils d’assolir durant els primers moments del cicle anual, especialment per a les serps més grosses.

La serp d’aigua (Natrix maura) és força resistent a la contaminació i per això se la pot trobar en gairebé qualsevol medi aquàtic, sempre que hi hagi peixos i amfibis, les seves preses principals. Llevat dels cims de les muntanyes, habita arreu dels Països Catalans.
Albert Montori.
La mida gran de la major part de serps dels Països Catalans permet instal·lar-los emissors sota la pell per a estudiar-ne l’activitat i l’ús de l’hàbitat. Fins ara s’han dut a terme estudis de radioseguiment en poblacions de serp d’aigua (Natrix maura), serp verda (Malpolon monspessulanus) i escurçó ibèric. Gràcies a aquests estudis s’han pogut analitzar els patrons d’activitat diària d’aquests ofidis, les seves àrees vitals i la selecció que fan dels microhàbitats, dades que, d’altra manera, hauria estat impossible d’aconseguir.
Pel que fa als quelonis o tortugues, es tracta, en general, d’espècies molt longeves i, en conseqüència, amb taxes de renovació poblacional molt baixes. La major part de quelonis tarden molts anys a arribar a la maduresa sexual, i compensen aquest desavantatge amb una longevitat elevada.
Les espècies d’aigües continentals, com la tortuga d’aigua ibèrica (Mauremys leprosa) i la tortuga d’aigua europea (Emys orbicularis), presenten dinàmiques de població comparables a les dels amfibis, bé que cal tenir en compte que les taxes d’extinció i colonització d’aquestes tortugues són molt més baixes que les dels amfibis.
Pel que fa a les tortugues marines, l’única espècie amb poblacions importants als Països Catalans és la tortuga babaua. Sembla que a la Mediterrània aquesta tortuga té dos nuclis de població, un oriental i l’altre occidental. La major part d’exemplars que visiten les nostres costes pertanyen al nucli occidental i són, majoritàriament, subadults o adults de mida petita. Això vol dir que la costa dels Països Catalans molt probablement no és una zona de reproducció de la tortuga babaua, tot i que en alguns casos s’hi ha reproduït. En canvi sí que és una important zona d’alimentació i creixement per a aquesta espècie.

Als Països Catalans només es reconeix una població natural de tortuga mediterrània (Testudo hermanni), la del massís de l’Albera. La resta de poblacions es consideren introduïdes.
Albert Montori.
Les poblacions de tortugues terrestres, actualment molt petites, han estat poc estudiades, de manera que no es poden generalitzar els trets bàsics de la seva ecologia. La població en millor estat de conservació és la de tortuga mediterrània que es va introduir a la punta de la Banya del delta de l’Ebre, que també és l’única que s’ha estudiat a fons. Els exemplars d’aquesta població són sedentaris, encara que no es pot saber si és a causa de la baixa densitat de població o si es tracta d’una característica pròpia de l’espècie. La poca mobilitat de la tortuga mediterrània, lligada a la gran modificació del medi i a l’elevada freqüència amb què es declaren incendis forestals als llocs on habita, fa que les poblacions naturals d’aquesta espècie als Països Catalans estiguin molt amenaçades.
Actualment ja s’ha pogut estudiar la dieta i la reproducció de moltes espècies de rèptils dels Països Catalans. En alguns casos s’han constatat diferències geogràfiques, sovint relacionades amb les característiques de l’hàbitat i, per tant, amb la disponibilitat de preses. Aquests estudis ecològics resulten molt útils per a la conservació, ja que són fonamentals a l’hora d’identificar els requeriments principals de les espècies amenaçades.
Els estudis sobre conservació
Els impactes i els riscos a què està sotmesa actualment la fauna reptiliana són d’índole molt diversa i d’una intensitat molt elevada. El 21,1% de les 151 espècies de rèptils que habiten a Europa (sobre un total de 8.300 espècies a tot el món) estan amenaçades. Les poblacions de rèptils d’Europa experimenten un declivi, menor que el dels amfibis, però molt més acusat que el dels mamífers i els ocells. Aquest declivi s’ha accentuat de manera notable les dues darreres dècades. És significatiu que l’informe realitzat per la Unió Internacional per a la Conservació de la Natura (UICN) l’any 2009 consideri que el 42% de les espècies europees estan en retrocés i només un 3% estan augmentant. Amb 65 espècies de rèptils, l’Estat espanyol és el país de la UE on hi ha més diversitat, fet que obliga a realitzar un gran esforç de conservació. Aquest esforç ha de ser especialment important als Països Catalans, on són representades les dues terceres parts de les espècies ibèriques (43 espècies), tot i ésser una de les àrees amb més densitat de població humana.
Són moltes les causes que expliquen el declivi de les poblacions reptilianes d’Europa. La fragmentació, la degradació i, especialment, la pèrdua d’hàbitats naturals en són les principals. Aquest problema afecta 22 de les 27 espècies amenaçades. Altres raons que expliquen el declivi de les poblacions de rèptils són els canvis en l’ús del sòl, la captura il·legal, els atropellaments, la contaminació i el canvi climàtic. Tanmateix, la pèrdua d’hàbitats té un impacte tres vegades més intens que el de la resta de factors. Això s’explica per la gran ocupació humana del continent europeu. Europa té actualment uns 731 milions d’habitants i és el territori més urbanitzat del món. A més, juntament amb Àsia, és el que té una major densitat de població. Dins d’Europa, els països que voregen la Mediterrània alberguen al voltant de 400 milions de persones, i d’aquestes, 135 milions viuen a la costa. Si això es té en compte, és lògic pensar que la pèrdua d’hàbitat sigui la principal causa del declivi de les poblacions de rèptils.
La pèrdua i la fragmentació dels hàbitats, considerades dues de les principals causes de pèrdua de biodiversitat, poden ser degudes a diversos motius, entre els quals cal esmentar la tala d’arbres per a l’explotació forestal, la creació d’àrees agrícoles i ramaderes, la construcció d’urbanitzacions, xarxes viàries i ferroviàries, el pas de línies elèctriques, etc. La fragmentació dels hàbitats incrementa el risc d’extinció de les poblacions de rèptils, ja que en redueix la mida. Així, doncs, la construcció d’una nova carretera pot constituir una petita reducció d’hàbitat, però, en canvi, pot disminuir molt la permeabilitat entre els hàbitats que divideix i aïllar-ne les poblacions, amb la qual cosa augmenta la probabilitat d’extinció. En una situació com aquesta es troben, per exemple, les poblacions catalanes de sargantana àgil i les poblacions de sargantanes del gènere Iberolacerta que habiten als Pirineus. També és la situació de diverses espècies de serps, com ho demostra el fet que a la franja costanera mediterrània s’hagi reduït el nombre de localitats on s’han fet observacions recents d’aquests rèptils. A més, moltes de les observacions històriques, com la de l’escurçó ibèric a la rodalia de Barcelona, no han estat actualitzades els darrers 25 anys, molt probablement a causa de l’extinció de les poblacions locals. La pèrdua d’hàbitats naturals per la urbanització del sòl és, sense cap mena de dubte, la causa d’aquesta tendència. Aquest fenomen s’està produint a tot el territori i afecta totes les espècies en major o menor grau.

Els incendis forestals fan retrocedir les comunitats vegetals a fases inicials de la successió ecològica. En aquests estadis primigenis, quan el medi s’està regenerant de manera natural, existeixen nombroses zones obertes que afavoreixen la presència de petits saures, especialment de les espècies més oportunistes i colonitzadores, com la sargantana cuallarga (Psammodromus algirus) i la sargantana ibèrica (Podarcis hispanica). A mesura que el sistema madura, la biodiversitat reptiliana augmenta.
Xavier Santos.

Els incendis forestals són la principal causa del declivi de les poblacions de tortugues terrestres. A la imatge es poden veure diversos exemplars de tortugues mediterrànies (Testudo hermanni) mortes després d’un incendi forestal a Petra-Manacor (Mallorca). Si la freqüència de focs és superior a la natural, les poblacions van disminuint fins a valors irreversibles.
Joan Mayol.
Els incendis forestals són pertorbacions que comporten un canvi molt dràstic i ràpid de l’hàbitat i que el situen en una fase molt inicial de la successió ecològica. Generalment, els estudis sobre l’impacte dels incendis forestals acostumen a incidir en la comunitat vegetal, però cal tenir en compte que l’alteració de la coberta vegetal afecta totes les comunitats d’animals i que pot canviar l’ordre o la intensitat de les interaccions faunístiques. Els efectes dels incendis sobre les poblacions de rèptils encara no es coneixen gaire bé, perquè es disposa de poques dades quantitatives. La major part d’aquestes dades fan referència a la tortuga mediterrània. Estudis realitzats a la serra de l’Albera després de l’incendi del 1994 indiquen que un 30% dels exemplars de tortuga mediterrània van morir directament per les flames. Això implica que, si la freqüència d’incendis és molt alta, la població d’aquesta tortuga s’extingirà en pocs anys. Altres treballs realitzats sobre poblacions de rèptils de fora dels Països Catalans indiquen que no existeix cap evidència que es produeixin migracions després d’un incendi forestal. No obstant això, la variabilitat interespecífica és prou gran per a no generalitzar aquesta observació. Sigui com vulgui, l’impacte directe del foc sobre les espècies menys mòbils ha de ser, certament, important. Cal comentar que la desaparició de la tortuga mediterrània al Garraf al començament del segle XX va ser deguda, molt probablement, als incendis forestals. D’altra banda, la pèrdua de cobertura vegetal a causa dels incendis afavoreix l’erosió del sòl, incrementa el risc de desertització i augmenta el risc d’avingudes. A les comarques valencianes de la Canal de Navarrés i la Vall d’Aiora el rebliment dels tolls dels barrancs i els rius d’interior ha provocat la disminució d’hàbitats disponibles per a la tortuga d’aigua ibèrica. Per a algunes espècies de rèptils, especialment de saures, els incendis forestals comporten una millora en la qualitat de l’hàbitat, que es torna tèrmicament més favorable, ja que els espais oberts permeten millors condicions de termoregulació que els boscos amb una coberta densa. Per a espècies amb requeriments tròfics o ambientals concrets, com moltes serps, els incendis poden no ser perjudicials però no comporten, tampoc, un creixement apreciable de les poblacions.

La sargantana cuallarga (Psammodromus algirus) es va introduir fa relativament poc temps a Mallorca. Avui dia presenta una distribució força estesa a la part oriental de l’illa i s’ha observat, també, en altres localitats. És probable que aquestes poblacions introduïdes s’estenguin ràpidament per tota l’illa, ja que tenen a la seva disposició un gran nombre d’hàbitats adequats. A la foto es pot veure una parella de sargantanes cuallargues durant l’acoblament.
Pere Albornà.
L’activitat agrícola tendeix a homogeneïtzar els medis i, en conseqüència, a fer disminuir la diversitat vegetal. Aquesta disminució, especialment en els casos de monocultures extensives, es tradueix, també, en una pèrdua de la riquesa específica de rèptils. A més, l’activitat agrícola porta associades tot un seguit d’actuacions perjudicials per a la fauna en general (alteració del terreny, tractaments fitosanitaris, abocaments, etc.). La transformació dels aiguamolls costaners en terres de conreu, per exemple, ha afectat especialment la tortuga d’aigua europea. Com els amfibis, els rèptils no depenen del tipus de comunitat vegetal, però sí de com s’estructura la vegetació. S’ha observat que l’increment de la superfície forestal i l’abandonament del treball tradicional al bosc ha afavorit les espècies més higròfiles en detriment de les més termòfiles. Per aquest motiu, les masses forestals que queden disperses en un mosaic de zones obertes, les franges de vegetació dels marges dels camps, les clarianes dels boscos amb matoll baix dispers, els ambients amb esquerdes i fissures, els murs de pedra seca, els tallafocs i els camins forestals, afavoreixen la intrusió de moltes espècies de rèptils en els sistemes forestals. Cal tenir en compte, però, que els tallafocs i els camins forestals també afavoreixen la penetració humana al bosc, amb la consegüent alteració del medi (atropellaments, residus, incendis, etc.). Un altre aspecte que cal tenir present és el manteniment de fusta morta als ambients forestals. Si bé un nombre excessiu de branques i troncs morts afavoreix els incendis forestals, el manteniment de troncs de gran calibre a les clarianes genera microrelleus i ambients molt propicis per als rèptils, que hi troben refugi i aliment. Els sistemes d’informació geogràfica han permès estimar la riquesa potencial de rèptils a les quadrícules UTM de 10 × 10 km de Catalunya i comparar-la amb la riquesa real calculada a partir de les observacions fetes sobre el terreny i emmagatzemades en bases de dades. La diferència entre la riquesa real i la potencial s’ha relacionat amb els diferents usos del sòl i s’ha observat que tant l’activitat agrícola com la silvicultura extensiva de coníferes suposen una pèrdua de riquesa de rèptils. En canvi, sembla que els boscos caducifolis mantenen comunitats reptilianes estables i riques.

L’atropellament és la principal causa de mort dels rèptils. Els saures i les serps de mida petita, com aquest lluert o llangardaix verd (Lacerta bilineata), perden la vida, sobretot a les carreteres, i en molt pocs casos a les autopistes i altres vies ràpides, d’on les poblacions reptilianes ja han estat eliminades pel trànsit. A les carreteres secundàries, la neteja de la vegetació dels marges redueix el nombre d’atropellaments, ja que els rèptils normalment s’assolellen a poca distància de la vegetació per poder amagar-se en cas de perill.
Marc Franch.
Pel que fa a les carreteres, en general són un factor important de mortalitat i de fragmentació dels hàbitats, no solament dels rèptils sinó de tota la fauna. Les estructures viàries són un element cada vegada més comú i abundant al nostre entorn i també més impermeable per a la fauna. Els rèptils perden la vida a les carreteres sobretot durant el dia i als mesos d’estiu, ja que moltes espècies utilitzen l’asfalt com a font de calor quan hi toca el sol. A més, la majoria del trànsit és diürn, per la qual cosa durant el dia augmenta el risc d’atropellament. En un estudi dut a terme en carreteres secundàries de la xarxa viària de Catalunya es va observar que l’espècie que patia més accidents era la serp verda. Cal tenir en compte que la incidència dels atropellaments de rèptils es tendeix a infravalorar, perquè un gran nombre són ràpidament retirats de les carreteres pels carronyaires i d’altres no es detecten perquè són molt petits o bé el mateix trànsit rodat els fa desaparèixer. Aquest és el cas dels ofidis i els saures de petites dimensions, els rèptils més freqüentment atropellats, si bé normalment no se’ls veu.
L’impacte de la contaminació amb productes químics procedents de l’activitat industrial i agrícola sobre les poblacions de rèptils encara ha estat poc avaluat. S’ha observat, però, que els rèptils, que actuen de depredadors en les xarxes trònques, acumulen nivells alts de contaminants, tant organoclorats com metalls pesants. Així, s’han trobat nivells elevats de DDT i PCB a les poblacions de serp d’aigua del delta de l’Ebre. Aquesta contaminació juntament amb el progressiu deteriorament dels ambients aquàtics dels arrossars del delta semblen els responsables de la dràstica disminució de les poblacions d’aquest ofidi, en altres temps molt abundant.

Exemplar de tortuga mossegadora (Chelydra serpentina) de l’Albufera de València. L’alliberament de tortugues exòtiques als medis aquàtics és un problema mediambiental greu, ja que competeixen amb les espècies autòctones pels llocs d’insolació i l’aliment.
Arxiu del Parc Natural de s’Albufera de Mallorca / Maties Rebassa.
Un altre factor que perjudica les poblacions de rèptils autòctons és la introducció d’espècies allòctones al medi natural. Les espècies al·lòctones alteren la dinàmica de poblacions de les comunitats naturals perquè les depreden, hi entren en competència, els prenen els recursos i els modifiquen els hàbitats. Això pot provocar la regressió i la desaparició de les espècies autòctones. n’és un bon exemple la introducció de la tortuga d’orelles vermelles (Trachemys scripta elegans) a la major part dels hàbitats aquàtics dels Països Catalans. Alguns estudis indiquen que aquesta espècie pot interactuar amb les poblacions de tortugues autòctones i desplaçar-les dels llocs on s’assolellen. Des que es va prohibir comercialitzar la tortuga d’orelles vermelles com a animal de companyia, estan apareixent altres tortugues al·lòctones, que també podrien fer la competència a les poblacions autòctones. La llista de possibles competidors és llarga: T. s. scripta, Pseudemys floridana, P. concinna, Graptemys pseudogeographica, G. kohni, Chrysemys picta, Macroclemys temminckii, Chelydra serpentina, Pelodiscus sinensis, Apalone spinifera, Pelomedusa subrufa, etc. Aquest fet demostra que resulta imprescindible regular el comerç d’espècies exòtiques, dur a terme una bona política d’educació ambiental i realitzar una gestió adequada de les espècies introduïdes per tal d’eliminar-ne les poblacions, com ja s’ha fet al Parc Nacional de Doñana i en altres espais naturals del sud de la península Ibèrica.

Els rèptils són fàcils d’observar mentre s’assolellen al descobert, com aquest llangardaix ocel·lat (Timon lepidus). Els de mida més gran necessiten més calor per a assolir una temperatura que els permeti estar actius, per això fins ben entrada la primavera no es mouen gaire.
Vicent Sancho.
El territori dels Països Catalans on s’han produït més introduccions, translocacions i naturalitzacions de rèptils els darrers anys és l’arxipèlag balear. Al llevant de Mallorca s’hi ha naturalitzat la sargantana cuallarga (Psammodromus algirus), de la qual s’han detectat diverses poblacions ben establertes. Al sud de Mallorca, la sargantana italiana o siciliana (Podarcis sicula) ha establert una petita colònia urbana, molt incipient. A la localitat mallorquina de Llucmajor hi ha una població de llangardaix ocel·lat (Timon lepidus [=Lacerta lepida]), que molt possiblement a la dècada de 1990 ja s’hi reproduïa, bé que no se’n tenen dades més recents. A Sineu, també a Mallorca, hi ha una població ben establerta de tortuga d’aigua ibèrica. La tortuga d’orelles vermelles ha estat introduïda en diverses localitats mallorquines, on ja es reprodueix; la serp de ferradura (Hemorrhois hippocrepis [=Coluber hippocrepis]) té poblacions a Mallorca i a Eivissa; la serp verda ja és present a Mallorca, i s’ha confirmat la presència de poblacions de serp blanca (Rhinechis sealaris [=Elaphe sealaris]) a Mallorca i a Eivissa. Pel que sembla, els ofidis al·lòctons han colonitzat aquest arxipèlag transportats inadvertidament a les soques d’oliveres importades com a arbres ornamentals, principalment, des d’Andalusia i Extremadura. A les illes Pitiüses, on mai no hi havia hagut serps, aquests ofidis podrien representar una amenaça per a les sargantanes insulars. Tot i alguns intents de control, no sembla factible revertir aquest procés. Malauradament les espècies citades no són les úniques que s’han detectat al medi natural. Malgrat que no es tenen indicis que es reprodueixin, cal assenyalar la captures de dos exemplars adults de tortuga mossegadora (Chelydra serpentina) al Parc de s’Albufera de Mallorca, un dels quals pesava 17 kg, com també la captura d’un exemplar de la mateixa espècie a prop de Palma. Així mateix, s’han capturat iguanes i altres espècies exòtiques. Molts d’aquests animals tropicals o subtropicals no troben a l’arxipèlag balear les condicions adients per a establir-hi poblacions fixes, però alguna espècie s’hi podria naturalitzar.
La gestió dels hàbitats i les espècies és un dels aspectes que més ha canviat els darrers anys. Alguns cops ha permès fins i tot que determinades espècies es recuperessin de manera espontània, però en la majoria de casos ha calgut reintroduir exemplars de l’espècie desapareguda o reforçar les poblacions de les espècies amenaçades per incrementar la densitat de població en la seva àrea de distribució natural, com ha succeït amb la tortuga mediterrània. Actualment aquesta espècie només té una sola població natural, la de la serra de l’Albera, la qual ha estat objecte de reforçaments successius. La resta de poblacions han estat creades de nou en indrets on l’espècie havia desaparegut, com el Garraf, el Montsant, la serra d’Irta i el desert de les Palmes, tots dins l’àrea que podria haver ocupat l’espècie antigament. També s’ha introduït a la punta de la Banya, al delta de l’Ebre, i existeixen poblacions introduïdes més antigament a les Balears. Els esforços per crear tots aquests nuclis de població han estat fructífers, i sembla que l’espècie ja no es troba en perill. Fins ara, la gestió de les diverses espècies de quelonis presents als Països Catalans s’ha planificat sota la mateixa perspectiva: reproducció en captivitat i reforçament de les poblacions naturals sense cap tipus de criteri ni base científica. De totes maneres, no sempre és així, i existeixen projectes de gestió ben estructurats. n’és un exemple el projecte Life Emys Ter, que, entre altres objectius, es proposa restablir la població de tortuga d’aigua europea del Ter mitjançant la recuperació del seu hàbitat, els aiguamolls d’aigua dolça d’inundació temporal o permanent. També al País Valencià s’està duent a terme un programa d’actuacions per a la conservació de la tortuga d’aigua europea, programa que fa el seguiment de les poblacions d’aquest queloni, restaura les marjals costaneres on viu i duu a terme reintroduccions. Aquestes actuacions són reforçades per un programa d’eradicació de tortugues exòtiques de les localitats on conviuen amb la tortuga d’aigua europea. D’altra banda, als Països Catalans s’estan duent a terme diversos projectes de l’anomenada citizen science, que es fonamenta en el voluntariat i utilitza internet per a la comunicació entre els participants. Un d’aquests projectes és el projecte SARE (Seguiment d’Amfibis i Rèptils d’Espanya), que té com a funció avaluar l’estat de conservació de les poblacions d’amfibis i rèptils de l’Estat espanyol.
Pel que fa a les lleis sobre conservació de l’Estat espanyol, tot el que s’ha comentat per als amfibis és aplicable als rèptils. Només en alguns casos l’aplicació de la legislació existent adopta algunes particularitats a cada comunitat autònoma. Per exemple, a Catalunya l’escurçó ibèric està protegit en el nou catàleg de fauna. A les Balears, la serp d’aigua ha deixat d’estar protegida perquè es tracta d’una espècie introduïda que, a més, s’ha constatat que afecta negativament les poblacions de ferreret (Alytes muletensis). Això mateix passa amb moltes altres espècies introduïdes, les quals, tot i estar protegides a nivell estatal, han de ser gestionades des d’una altra perspectiva.
L’actualització taxonòmica dels rèptils
Els estudis genètics duts a terme durant les dues darreres dècades han permès actualitzar la taxonomia dels rèptils i esbrinar la procedència d’algunes poblacions reptilianes dels Països Catalans. Els canvis taxonòmics introduïts darrerament en aquest grup de vertebrats han afectat algunes espècies dels Països Catalans, que han canviat de gènere, s’han escindit en diverses espècies o s’han subdividit en subespècies diferents.
S’ha pogut confirmar que la tortuga mora (Testudo graeca) és originaria del nord d’Àfrica i que totes les poblacions d’aquesta espècie que habiten a la península Ibèrica i les Balears són introduïdes. Actualment la taxonomia de la tortuga mora és molt controvertida i és possible que en un futur els investigadors reconeguin diversos tàxons dins d’aquesta espècie.
La situació taxonòmica actual de les tortugues del gènere Emys és força complexa. Fins ara només s’han reconegut dues espècies d’aquest gènere: la tortuga d’aigua europea (E. orbicularis) i la tortuga d’aigua siciliana (E. trinacris), endèmica de l’illa de Sicília. A l’arxipèlag balear es troba la tortuga d’aigua europea, present a gran part de Menorca i al Parc Natural de s’Albufera de Mallorca. Sembla que aquestes poblacions hi foren introduïdes en l’època dels romans. A Mallorca n’existeixen dos haplotips, mentre que a Menorca només se n’ha trobat un. L’haplotip comú és present també a les poblacions de Catalunya, del sud de França i de Còrsega. Les poblacions d’ambdues illes són, però, força semblants morfològicament. A nivell subespecífic la situació és confusa, ja que a la península Ibèrica i a les Balears s’han descrit diverses subespècies, tant des del punt de vista morfològic com des del punt de vista molecular: E. o. galloitalica, E. o. fritzjuergenobsti, E. o. orbicularis i E. o. hispanica. A la zona de la Selva (Gironès) conflueixen tres d’aquestes subespècies: E. o. fritzjuergenobsti, E. o. galloitalica i E. o. orbicularis. Pel que fa a les dues darreres, sembla que procedeixen de França i que van entrar per ambdós extrems dels Pirineus.
l 1987 es considerava que la tortuga d’aigua ibèrica (Mauremys leprosa) era una subespècie de la tortuga de rierol (M. caspica), i per tant es batejava com a M. c. leprosa. Actualment la tortuga d’aigua ibèrica i la tortuga de rierol es consideren dues espècies diferents. La primera es localitza a la península Ibèrica i al nord-oest d’Àfrica, mentre que la segona es troba al sud-est de l’antiga Unió Soviètica, a Turquia i a l’Iran. De tortuga d’aigua ibèrica se n’han descrit fins a 8 subespècies, però únicament se n’accepten dues: M.l. saharica i M. l. leprosa. La darrera és la subespècie present als Països Catalans.
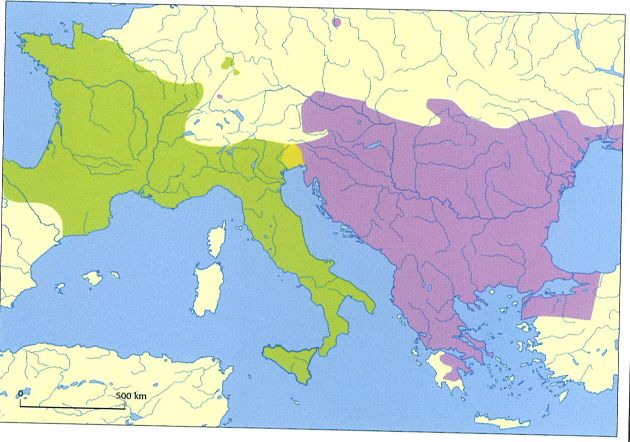
Àrea de distribució de les dues espècies de llangardaix verd o lluert: l’oriental (Lacerta viridis, en blau) i l’occidental (L. bilineata, en verd). En groc s’indica la zona d’hibridació entre les dues espècies, que es manté estable i no s’incrementa al llarg del temps, ja que la fertilitat dels híbrids és menor. L’existència d’aquesta zona d’hibridació estable referma la separació de l’antiga espècie L. viridis en dues espècies diferents. Tanmateix, el gènere Lacerta encara es troba en revisió, per la qual cosa no es descarta que properament s’hi introdueixi alguna altra novetat taxonòmica.
IDEM, a partir de dades dels autors.
Per a l’espècie Lacerta viridis, batejada amb el nom comú de llangardaix verd o lluert, es dibuixava una àrea de distribució àmplia que abraçava des de la península Ibèrica fins a Turquia. Però aquesta àrea de distribució s’ha modificat en dividir-se l’espècie en dues. Segons estudis morfològics, moleculars i d’hibridació, el llangardaix verd s’ha separat en una espècie occidental, que s’ha anomenat Lacerta bilineata, i una altra d’oriental, batejada com a Lacerta viridis. L’espècie present als Països Catalans és L. bilineata, que es distribueix també pel nord de la península Ibèrica, França, Mònaco, San Marino, Itàlia (inclosa Sicília), Alemanya occidental, la meitat meridional de Suïssa, Eslovènia i Croàcia. Per tant, el nom comú de llangardaix verd o lluert s’ha de fer correspondre, actualment, amb el nom científic de L. bilineata. Ambdues espècies presenten una zona d’hibridació estable al nord-est d’Itàlia i a les zones adjacents d’Eslovènia, on la fertilitat dels híbrids disminueix. Cal tenir en compte, però, que el gènere torna a estar en revisió, de manera que se’n podria modificar la nomenclatura.
La sargantana marroquina (Lacerta perspicillata) ha passat a anomenar-se Scelaris perspicillata, i el llangardaix ocel·lat (L. lepida) ha passat a anomenar-se Timon lepidus. A més, estudis taxonòmics sobre aquesta darrera espècie han determinat que els llangardaixos africans no pertanyen a la mateixa espècie que els europeus. Així, actualment, en aquest gènere s’inclouen quatre espècies: T. lepidus, T. pater, T. tangitanus i T. princeps, les tres primeres de les quals s’han diferenciat dins de l’anterior Lacerta lepida lato sensu. L’espècie present als Països Catalans és T. lepidus, que ocupa pràcticament tota la península Ibèrica, el sud-est i el sud-oest de França i el nord-oest d’Itàlia.
Als Països Catalans hi ha dues subespècies de llangardaix ocel·lat: Timon lepidus nevadensis i T. l. lepidus. El primer és un llangardaix de mida gran amb el cap punxegut i una coloració de fons bruna o grisenca, de vegades verdblavosa i ocre. Té un nombre més petit d’escates ventrals i un nombre més gran d’escates dorsals que T. l. lepidus, com també més porus femorals i escates al collar. Les dents són més grans. Els ocels són més petits, i en té menys. Un caràcter diagnòstic de T. l. nevadensis és l’absència d’escates de color negre a tot el cos. El segon, T. l. lepidus, és un llangardaix de mida gran amb el cap curt i el musell poc punxegut. El color de fons és verd viu i presenta quatre fileres d’ocels grans, de color blau envoltats de negre, a cada flanc.
La sargantana vivípara és un exemple de les dificultats que comporta l’establiment d’una taxonomia vàlida per als lacèrtids. Si el 1987 s’anomenava Lacerta vivipara, al final dels anys noranta va passar a anomenar-se Zootoca vivipara, i posteriorment va prendre de nou el nom original. Revisions taxonòmiques recents de la família dels lacèrtids indiquen que cal elevar el subgènere Zootoca a gènere. Per tant, tal com estan les coses actualment, la sargantana vivípara s’ha de denominar Z. vivipara. Un dels problemes d’aquesta espècie és que té una àrea de distribució molt àmplia, que va des de Galícia i Irlanda fins al Japó. Això dificulta molt l’estudi d’aquesta sargantana, tot i que sembla que les poblacions eslovenes, croates i del nord d’Itàlia podrien pertànyer a una altra espècie, Z. carniolica.
La situació taxonòmica de les sargantanes sota el nom específic de Lacerta monticola ha variat molt aquests darrers anys. Actualment aquesta espècie ha canviat de gènere i ha passat a anomenar-se Iberolacerta. A més a més, s’ha subdividit en set espècies diferents, tres de les quals habiten als Països Catalans: la sargantana aranesa (I. aranica), la sargantana pallaresa (I. aurelioi) i la sargantana pirinenca (I. bonnali, vegeu les pàgines corresponents).
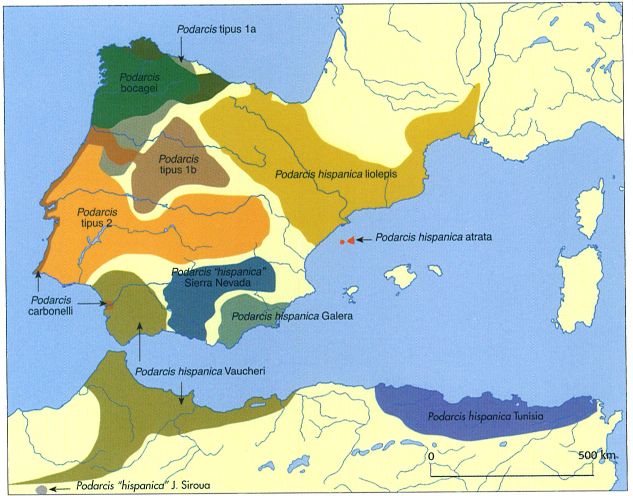
Àrea de distribució provisional del gènere Podarcis al territori continental de la península Ibèrica i al nord d’Àfrica. Aquest gènere actualment està en revisió, de manera que la situació taxonòmica de les diverses espècies i subespècies encara no es pot considerar definitiva. Al territori continental dels Països Catalans actualment hi ha reconegudes dues formes o tipus de sargantanes de l’espècie P. hispanica: P. hispanica var. liolepis, al nord, i P. hispanica var. galera, al sud.
IDEM, a partir de dades dels autors.
Una de les dificultats més grans de la dècada de 1980 va ser discriminar les dues espècies de sargantanes del gènere Podarcis presents als Països Catalans: la sargantana ibèrica (P. hispanica) i la sargantana de paret (P. muralis). Aquestes dificultats eren degudes, en part, a l’extrema variabilitat morfològica de la sargantana ibèrica, que va fer que, en el passat, la sargantana de paret fos citada erròniament, confosa amb la sargantana ibèrica, en moltes localitats de Catalunya on actualment se sap que no és present. Fins que no es van dur a terme estudis més profunds sobre la sargantana de paret no es van poder determinar els caràcters diagnòstics que van permetre diferenciar aquesta espècie de la sargantana ibèrica.
Actualment, però, la situació és molt diferent. Ara el problema rau a definir quantes espècies de sargantanes del gènere Podarcis tenen el seu hàbitat a la península Ibèrica. Les sargantanes d’aquest gènere constitueixen un grup ben caracteritzat, tant a nivell morfològic com molecular, de la família dels lacèrtids. Llevat de la sargantana de paret, la resta de sargantanes del gènere Podarcis que es documenten a la península Ibèrica constitueixen, juntament amb les formes del nord d’Àfrica, un grup monofilètic. Actualment es reconeixen sis espècies de sargantanes del gènere Podarcis a la Península (P. muralis, P. bocagei, P. carbonelli, P. hispanica, P. vaucheri i P. atrata) i dues a les Balears (P. pityusensis i P. lilfordi). Cal tenir en compte, però, que la taxonomia del gènere no es pot considerar, ni de bon tros, definitiva. Per començar, Podareis hispanica és un tàxon molt heterogeni format, aparentment, per un conjunt de diverses possibles espècies.

La sargantana de les Pitiüses (Podarcis pityusensis) és un endemisme de l’arxipèlag balear amb poblacions força estables i abundants a Eivissa i a Formentera, com també en els illots propers. La seva gran capacitat de colonització li ha permès introduir-se, a més, en diversos indrets de l’illa de Mallorca, on ha arribat a constituir poblacions estables. També hi ha una població introduïda a la plaça de les Glòries de la ciutat de Barcelona.
Marc Franch.
Als Països Catalans les espècies de Podarcis presents són P. atrata, P. pityusensis, P. lilfordi, P. muralis i, segons sembla, dues formes o tipus de P. hispanica: P. hispanica var. liolepis, al nord, i P. hispanica var. galera, al sud. El límit entre les dues formes de P. hispanica no es coneix amb precisió, i tampoc no se sap si aquestes dues formes s’hibriden entre si. Tampoc és clara la validesa de l’espècie P. atrata.
Les formes balears del gènere Podarcis, la sargantana gimnèsica (P. lilfordi) i la sargantana de les Pitiüses o pitiúsica (P. pityusensis), mostren una notable diferenciació insular, fenomen que es produeix, també, en altres espècies mediterrànies. En el cas de les Balears, el nivell de diferenciació en grups d’illes molt pròximes és molt elevat. Per exemple, només al petit arxipèlag de Cabrera s’han descrit 16 subespècies de sargantanes. En total s’han descrit 28 subespècies de sargantana gimnèsica (tres de les quals han estat invalidades) i 45 subespècies de sargantana de les Pitiüses (de les quals actualment se’n consideren vàlides 23). La sargantana gimnèsica és un dels dos únics rèptils endèmics de l’arxipèlag balear. Es localitza en nombrosos petits illots de Mallorca, Menorca i Cabrera, com també en diverses poblacions de les illes principals. La fragilitat d’aquestes petites poblacions i la seva capacitat per a colonitzar nous indrets en fan necessari un seguiment continuat. D’altra banda, la supervivència d’aquestes sargan-tanes en illots minúsculs de l’arxipèlag balear, on els recursos són molt limitats i acostumen a variar considerablement al llarg de l’any, determina l’aparició d’adaptacions i conductes poc habituals entre els rèptils: inclusió de nèctar i pol·len a la dieta, pol·linització, dispersió de llavors, fluctuacions espectaculars de la densitat de població, etc.
Tot i que en captivitat la sargantana gimnèsica i la sargantana de les Pitiüses es poden hibridar, la diferenciació genètica entre totes dues és prou evident. S’ha estimat que la separació dels dos llinatges va tenir lloc a l’època messiniana, fa uns cinc milions d’anys.
En el cas de la sargantana gimnèsica, els investigadors han identificat tres grans línies evolutives. La primera, la basal, correspon a les poblacions menorquines, la diferenciació de les quals és poc marcada, però discordant amb la proximitat geogràfica dels respectius illots, atès que sargantanes d’illes geogràficament molt properes poden presentar més diferències entre si que amb altres sargantanes d’illes més allunyades. La segona correspon a les poblacions de l’oest de Mallorca, i la tercera, a les de Cabrera. Curiosament, les sargantanes de l’illot de Colomer, a la península de Formentor, que constitueixen una població força aïllada, melànica, molt diferenciada i variable, mostren semblances genètiques amb les sargantanes del nord de Cabrera. Aquests dos llinatges es podrien haver diferenciat fa uns dos milions d’anys, ja que presenten diferències que els marcadors genètics daten d’èpoques molt antigues, d’uns 800.000 anys. Els resultats són inesperats, ja que durant les glaciacions les regressions marines permeteren la unió física de Mallorca amb Menorca fa tan sols 23.000 anys. Pel que fa a la sargantana de les Pitiüses, els estudis no estan tan avançats, i només es pot assenyalar que la separació dels llinatges d’Eivissa i Formentera sembla recular fins al milió d’anys.
L’antic complex de la bívia tridàctila (Chalcides chalcides) es va dividir al principi de la dècada de 1990 en cinc espècies: C. striatus, que es distribueix per la península Ibèrica, el sud-est de França i Ligúria; C. chalcides, amb dues subespècies: C. c. chalcides, que s’estén per la Itàlia peninsular i Sicília, i C. c. vittatus, que ocupa Sardenya i Tunísia; C. mertensi, que es distribueix pel nord de Tunísia i Algèria; C. minutus, localitzada al nord-est del Marroc, i C. pseudostriatus, que habita al nord-oest del Marroc. Les poblacions dels Països Catalans corresponen a l’espècie C. striatus.
La família dels colúbrids també ha estat objecte de revisions profundes que han afectat, sobretot, els dos gèneres més freqüents de serps europees: Coluber i Elaphe. Els estudis moleculars han demostrat que el gènere Coluber és polifilètic, i perquè la taxonomia reflecteixi aquest polifiletisme s’ha hagut de restringir el gènere al continent americà. L’anàlisi de les relacions interespecífiques i intraespecífiques, tant a nivell molecular com morfològic, unes vegades ha comportat un canvi de nomenclatura del gènere, i d’altres, la divisió d’una espècie en dues o més de diferents. Així, Coluber viridiflavus, que és la serp verda i groga, canvia el nom per Hierophis viridiflavus, mentre que Coluber hippocrepis, que és la serp de ferradura, actualment s’anomena Hemorrhois hippocrepis. També s’ha fet una revisió del gènere Elaphe, atès que tenia una àrea de distribució molt extensa i també es tractava d’un gènere polifilètic. A conseqüència d’aquesta revisió, la serp d’Esculapi (Elaphe longissima) ha canviat de nom i ara s’anomena Zamenis longissimus, mentre que la serp blanca (Elaphe scalaris) s’anomena Rhinechis scalaris.
La serp de cogulla o serp de garriga (Macroprotodon cucullatus) es troba al nord-oest d’Àfrica i a la península Ibèrica, com també en algunes illes mediterrànies, on es considera una espècie introduïda. Estudis morfològics, moleculars i biogeo-gràfics n’han aconsellat la divisió en quatre espècies diferents: M. cucullatus, que, a més de l’interior del Marroc i d’Algèria, ocupa una franja de la costa mediterrània que s’estén des de Tunísia fins a l’Orient Mitjà; M. brevis, amb una àrea de distribució que ocupa la península Ibèrica, el nord del Marroc i la costa Atlàntica d’aquest país fins al Sàhara Occidental; M. mauritanicus, pròpia del nord d’Àfrica i les Balears (que van ser colonitzades a partir d’exemplars originaris de Tunísia), i M. abukakeri, restringida a la costa occidental de Tunísia. Per tant, la serp de cogulla present als Països Catalans ha quedat dividida en dues espècies: M. brevis, al territori peninsular, i M. mauritanicus, a les Balears.

La colobra de collar (Natrix natrix), tot i estar present per tot el territori peninsular dels Països Catalans, és una espècie poc abundant. Cap al sud i l’interior es fa cada vegada més i més rara, i les seves poblacions es tornen molt aïllades i escasses. Tot i considerarse una serp aquàtica, és molt més forestal que la seva congènere, la serp d’aigua (Natrix maura), i es pot trobar molt allunyada dels punts d’aigua.
Albert Montori.
EPrenent com a base caràcters morfològics, la colobra de collar (Natrix natrix) es va dividir en nou subespècies, que més tard van quedar reduïdes a quatre. Dues d’aquestes espècies són insulars (N. n. corsa, pròpia de Còrsega, i N. n. cetti, pròpia de Sardenya), mentre que les altres dues espècies són continentals: N. n. helvetica, que s’estén per la meitat occidental del continent europeu i la franja nord-occidental del continent africà, i N. n. natrix, que ocupa la franja oriental del continent eurasiàtic. La frontera entre les dues subespècies continentals probablement se situa en una franja que va d’Hamburg a Venècia. Aquestes subespècies continentals es degueren constituir a partir de refugis faunístics durant les glaciacions plistocèniques. La subespècie de colobra de collar present als Països Catalans és, per tant, N. n. helvetica.
Pel que fa a la serp d’aigua (Natrix maura), la nova filogènia proposada recentment per al gènere Natrix la defineix com una espècie originària del nord d’Àfrica que posteriorment va envair la península Ibèrica. Tanmateix, aquest raonament filogeogràfic no concorda amb altres dades i, per tant, cal que es confirmi o es desmenteixi. En canvi, sí que és cert que dins de l’espècie N. maura s’observen tres clades ben diferenciats, un a Tunísia i Sardenya, l’altre al Marroc i un darrer a Europa. L’elevada diversitat d’haplotips presents a la península Ibèrica reforça el paper d’aquesta àrea geogràfica com a refugi plistocènic durant les glaciacions. Els darrers estudis moleculars han demostrat que les poblacions de serp d’aigua presents a Mallorca són introduccions relativament recents procedents, probablement, del sud de França.
Estudis moleculars recents han demostrat que l’escurçó europeu (Vipera aspis) és una espècie amb força variabilitat genètica que es distribueix, fonamentalment, per Itàlia, França, el nord-est de la península Ibèrica i algunes illes mediterrànies. Aquests resultats corroboren els estudis morfològics previs i confirmen la validesa de la subespècie V. a. zinnikeri, a la qual pertanyen les poblacions de la zona d’ambdós costats dels Pirineus, tant les de la Península com les del sud-oest de França.
Finalment, cal fer un darrer apunt sobre la doble nomenclatura científica de l’escurçó ibèric: Vipera latasti o V. latastei. Aquest escurçó va ser descrit l’any 1878 per Eduardo Boscá en un article publicat en el Bulletin de la Société Zoologique de France. Al final de l’article Boscá va escriure: “si de nouvelles recherches établissaient la nécessité d’élever au rang d’espèce cette forme nouvelle, je me propose de la dédier à M. Fernand Lataste, savant herpétologue de France, et je la nommerai Vipera latasti". Tot i que no va utilitzar el terme V. latastei en tot l’article, aquest nom sí que apareix en el mateix volum del Bulletin, concretament en una llegenda que acompanya un gravat amb diversos dibuixos d’escurçons firmat per J. Terrier. Un any més tard, Eduardo Boscá va fer servir en un altre article el terme V. latastei. Des de llavors, els dos noms, V. latasti i V. latastei, han estat utilitzats indistintament per diversos investigadors. Recentment, l’Asociación Herpetológica Española ha proposat acceptar el terme latastei, perquè, inequívocament, Boscá pretenia retre homenatge a la figura de l’herpetòleg francès Fernand Lataste. Malgrat tot, l’ús d’un nom o l’altre suposa un conflicte de nomenclatura que presumiblement haurà de resoldre el Comitè de Nomenclatura Zoològica d’acord amb les regles que estableix el seu codi.