Consideracions generals

Grand Prismatic Spring. Surgència termal situada al parc nacional de Yellowstone (EUA). La coloració dels marges és deguda a la presència d’organismes procariotes pigmentats i de tapissos microbians formats per organismes extremòfils, la morfologia i la fisiologia dels quals els permeten acomplir totes les funcions vitals en condicions extremes de temperatura, que pot superar els 70 °C.
Mila Zinkova.
Només es coneix un sol lloc en tot l’Univers on s’ha desenvolupat la vida, el planeta Terra, la qual cosa no implica que no s’hagi pogut desenvolupar en alguns o en molts altres indrets. El descobriment d’un nombre creixent de planetes extrasolars, alguns dels quals de constitució, mida i posició relativa respecte a la seva estrella raonablement similars als de la Terra, i el descobriment i la caracterització d’organismes extremòfils, que habiten en condicions ambientals extremes de temperatura i salinitat, ha donat un gran impuls a l’estudi de l’origen de la vida. Un origen que cal buscar en la història de la Terra.
Tots els éssers vius de la Terra comparteixen una sèrie de característiques comunes, entre les quals hi ha la manera d’organitzar la informació genètica, en el DNA o àcid desoxiribonucleic, i la manera de descodificar-la mitjançant el codi genètic, entre moltes d’altres. Les anàlisis filogenètiques han demostrat que totes les espècies actuals, i també les ja extingides, provenen d’un ésser viu primigeni ancestral, un avantpassat comú a partir del qual, per evolució –és a dir, per mecanismes de canvi preadaptatiu i per selecció natural–, s’han format totes les espècies que es coneixen. Unes espècies que la filogènia molecular ha agrupat en tres grans dominis: el dels bacteris i el dels arqueus, tots dos amb morfologia cel·lular procariota i que anteriorment s’agrupaven en un únic regne, el de les moneres; el tercer és el dels eucaris, que inclou tots els organismes amb morfologia cel·lular eucariota, i que agrupa els altres quatre regnes clàssics d’éssers vius –protists, fongs, plantes i animals.
La qüestió de l’origen de la vida és complexa, atesa la dificultat de conèixer com era exactament la Terra primitiva i la seva atmosfera, quines reaccions químiques s’hi desenvolupaven de forma espontània i quines aportacions concretes de materials rebia de l’espai, en els meteorits i cometes que hi queien. Per tenir una primera idea sobre l’origen de la vida, cal considerar com era la superfície de la Terra fa 4.000 milions d’anys (Ma), abans que s’iniciés la vida. No hi havia gens que diguessin als organismes com fabricar les proteïnes, ni enzims que catalitzessin les reaccions metabòliques, ni cap mena de metabolisme, ni fotosíntesi. La superfície de la Terra era un crosta mineral parcialment coberta per oceans salats, els quals contenien milers de compostos orgànics en dissolució. Hi havia una gran activitat volcànica i encara es produïen grans impactes amb asteroides que po dien evaporar parcialment els oceans. A causa d’aquests fenòmens violents, s’alternaven cicles geològics secs i humits. L’aigua s’evaporava constantment de la interfície entre el mar i l’atmosfera, es condensava i queia en forma de precipitacions; formava tolls amb soluts orgànics, i es tornava a evaporar. A partir d’aquest caos de terra, mar i atmosfera, d’alguna manera es va originar la vida, fa uns 4.000-3.800 Ma.
La complexitat de definir la vida
La vida no és res més que una propietat emergent de la química, un conjunt organitzat de reaccions químiques integrades i encadenades, localitzades en un espai físic concret i heretables; o, dit d’una altra manera, és un sistema químic que es manté per ell mateix i que és capaç d’evolucionar en termes darwinistes. Tanmateix, no és fàcil definir el concepte de vida, la qual cosa dificulta establir en quin moment concret aquest conjunt de reaccions químiques va esdevenir, per dret propi, un ésser viu.
Per començar, hom pot dir que la vida està vinculada a tres processos clau: reproducció, variació i selecció natural. Des del punt de vista metabòlic, es pot definir com una associació de compostos químics complexos autoorganitzats amb capacitat autoreproductora i catalítica que extreu energia de l’ambient. Una de les definicions més completes és la que diu que els organismes vius són aquells que: emmagatzemen i transmeten la informació necessària per a funcionar i per a fer organismes iguals a ells –la informació genètica–; estan capacitats per a permetre i regular les reaccions químiques gràcies a la participació d’enzims; capten energia de l’entorn i la transformen en energia pròpia, que emmagatzemen en molècules específiques per a poder-la utilitzar quan els sigui menester; tenen la capacitat de reproduir-se fent organismes iguals a ells mateixos; poden respondre de manera específica als estímuls externs i són sotmesos a selecció natural. Totes aquestes característiques han de concórrer en l’origen de la vida.
Breu història de la recerca sobre els orígens de la vida

Erasmus Darwin (1), metge, naturalista i filòsof, avi de Charles Robert Darwin, en el seu treball Zoonomia, parla d’un primer “filament vivent", origen dels éssers vius. Jean-Baptiste-Pierre-Antoine de Monet, cavaller de Lamarck (2), naturalista francès, va ser un dels primers a defensar l’evolució dels éssers vius. Charles Robert Darwin (3), en una carta a Joseph Dalton Hooker l’1 de febrer de 1871, suggereix que l’espurna original de la vida hauria començat en un “petit estany càlid, amb amoni, sals fosfòriques, llum, calor i electricitat, on s’hauria format un compost de proteïnes".
National Portrait Gallery, Londres (1); Pizzetta, J. (1893): Galerie des naturalistes, Ed. Hennuyer (2); foto de Maull i Polyblank, 1855 (3).
Després de la publicació de l’ Origen de les espècies el 1859, obra cabdal de la biologia on Charles Robert Darwin (1809 – 1882) va establir les bases de la teoria moderna de l’evolució i on va descriure la selecció natural com a procés evolutiu, molts naturalistes van adoptar la idea que els organismes vius són el resultat històric de la transformació gradual de la matèria inerta.
De manera general hom assumeix que els primers filòsofs i naturalistes apel·laven a la generació espontània per explicar l’origen de la vida, però de fet la possibilitat que la vida sorgís directament de matèria no viva era percebuda inicialment com un simple mecanisme de reproducció asexual, no pas com un mecanisme per a explicar l’origen de la vida.
Aquesta visió va canviar amb les idees transformistes desenvolupades durant els segles XVIII i XIX per diversos pensadors, com George-Louis Leclerc, comte de Buffon (1707 – 1788), que va interpretar les variacions dels éssers vius com degeneracions; Erasmus Darwin (1731 – 1802), avi de Charles R. Darwin, que en la seva obra Zoonomia (1794) va avançar algunes idees evolucionistes similars a les proposades posteriorment per Lamarck; i Jean-Baptiste-Pierre-Antoine de Monet, cavaller de Lamarck (1744 – 1829), que en la seva obra Filosofia zoològica (1809) va proposar la primera teoria coherent sobre l’evolució de les espècies, basada en l’herència dels caràcters adquirits –una teoria que tanmateix s’ha demostrat que no és correcta atès que no s’ajusta als fets observables–. Respecte a l’origen de la vida, tots ells van invocar explícitament la generació espontània com a mecanisme que va conduir a l’aparició de la vida, i no únicament com a mecanisme de reproducció asexual. En paraules de Lamarck, “La natura, mitjançant la calor, la llum, l’electricitat i la humitat, ha format per generació espontània l’organisme més simple de cada regne".
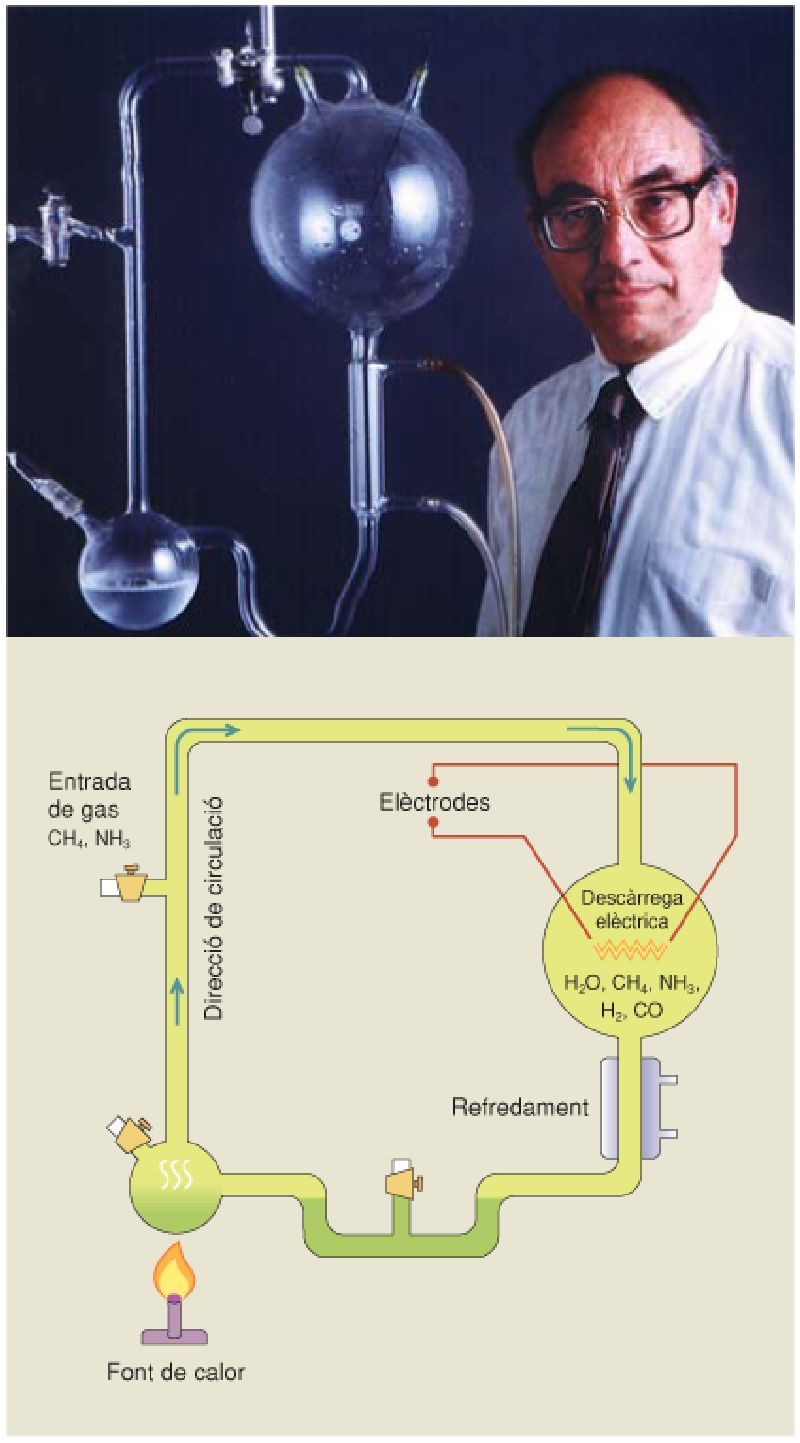
Stanley Miller (a la fotografia) i Harold Urey van dissenyar l’experiment que simulava les condicions químiques en la Terra primigènia al laboratori, per tal de demostrar que la síntesi espontània d’aquests compostos podria haver estat l’estadi previ a l’origen de la vida. Aparell de l’experiment de Miller i Urey, amb el qual volien simular les condicions de la Terra primitiva.
A partir de fonts diverses.
Com els seus predecessors, Charles Darwin va assumir que les plantes i els animals van sorgir de forma natural a partir de matèria no viva. El 1837 va escriure, en una llibreta de notes que es coneix com a Second Notebook, que “L’íntima relació entre la vida i les lleis de la combinació química, i la universalitat d’aquestes, apunta que la generació espontània no és un fet improbable". Encara més, en l’Origende les espècies hom pot llegir: “Haig d’inferir de forma anàloga que probablement tots els éssers orgànics que han viscut a la Terra descendeixen d’una forma primordial, a partir de la qual la vida va respirar per primer cop". Dotze anys després de la publicació de la primera edició de l’Origen de les espècies, Darwin va reblar el clau en una carta dirigida al seu amic Joseph Dalton Hooker (1817 – 1911): “Sovint es diu que totes les condicions per a la primera producció d’un organisme viu es troben presents en l’actualitat, i podrien haver estat presents sempre. Però si (Oh!, quin gran si) poguéssim imaginar un petit toll d’aigua càlida que contingués tots els tipus de sals amòniques i fosfòriques en presència de llum, calor i electricitat, on es pogués formar químicament un compost proteic, a punt per a experimentar canvis encara més complexos, en l’actualitat aquesta matèria seria instantàniament devorada, o adsorbida, però aquest no hauria estat el cas abans que es formessin les criatures vives".
En aquest sentit, al final del segle XIX ja s’havia demostrat que era perfectament possible la síntesi química –o abiòtica, és a dir, sense la participació d’éssers vius– de lípids i glícids, emprant diverses barreges de gasos i descàrregues elèctriques. I al principi de segle XX ja s’havien sintetitzat químicament els primers aminoàcids al laboratori, a partir de formamida humida (COH-NH2) sotmesa a descàrregues elèctriques i a llum ultraviolada.

Coberta de la primera edició en anglès de l’influent llibre d’Aleksandr I. Oparin L’origen de la vida, on va exposar la seva teoria. La hipòtesi d’Oparin explica l’origen de la vida, a partir d’uns primers aminoàcids formats de gasos com el metà, l’hidrogen i l’amoníac, i amb la intervenció d’altes temperatures, radiació ultraviolada i descàrregues elèctriques a l’atmosfera (llampecs).
Al novembre del 1923 el jove bioquímic rus Aleksandr Ivanovitx Oparin (1894 – 1980) va publicar un breu llibre intitulat L’origen de la vida, on proposava per primer cop de forma obertament explícita que abans de la vida cel·lular hi hauria d’haver hagut un llarg període de síntesi abiòtica i d’acumulació de compostos orgànics, a partir dels quals van sorgir, per agregació espontània, els primers organismes vius, en paraules d’Oparin “uns bacteris heterotròfics". El 1936 va publicar un segon llibre, Origen de la vida, molt més madur, on proposava que els primers hidrocarburs s’havien format en unes condicions atmosfèriques radicalment diferents de les actuals, en presència d’una atmosfera reductora –sense oxigen–, a partir de carburs metàl·lics d’origen geològic i vapor d’aigua. Aquests hidrocarburs haurien pogut reaccionar posteriorment amb oxigen per a formar diverses molècules orgàniques, com alcohols, cetones i aldehids, i amb amoni per a formar amines, amides i sals amòniques, uns compostos bàsics per a la formació dels aminoàcids que constitueixen les proteïnes, les quals al seu torn s’haurien agregat per a formar sistemes col·loïdals a partir dels quals haurien evolucionat aquests suposats bacteris heterotròfics.

Joan Oró i Florensa, bioquímic català, va contribuir a la recerca de l’origen de la vida mitjançant experiments de síntesi d’adenina (un component clau de l’àcid nucleic) a partir de cianur d’hidrogen.
Fototeca.cat.
La traducció a l’anglès del segon llibre d’Oparin va atreure l’atenció de nombrosos científics, que van iniciar experiments per simular la síntesi de compostos orgànics en les condicions que se suposava que tenia la Terra primitiva. De tots aquests experiments cal destacar-ne dos: el realitzat pels químics nord-americans Stanley L. Miller (1930 – 2007) i Harold C. Urey (1893 – 1981), i el realitzat pel bioquímic català Joan Oró i Florensa (1923 – 2004). Miller i Urey van dissenyar un aparell que reproduïa la interacció entre l’atmosfera i l’oceà, en el qual es podien produir descàrregues elèctriques de 60.000 V per simular llamps, i el van omplir amb diverses barreges de metà, amoni, hidrogen i aigua, que es mantenia calenta durant l’experiment. Després d’una setmana a una pressió d’1,5 bars, quasi el 50% del carboni present en la barreja havia quedat incorporat en compostos orgànics que s’havien sintetitzat químicament de forma espontània, entre els quals van identificar 13 dels 20 aminoàcids que normalment formen les proteïnes. Poc després d’aquest experiment, Oró i els seus col·laboradors van demostrar de forma similar que, si en la barreja també hi ha cianur amònic, s’obté adenina, un dels nucleòtids bàsics que formen el DNA.
Origen o orígens de la vida?
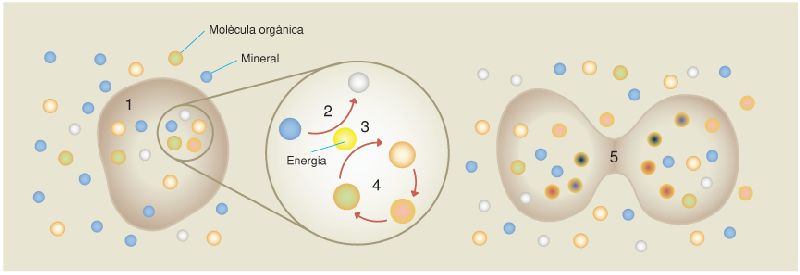
Requisits per a l’origen metabòlic de la vida. Primer s’ha de formar una barrera que separi l’entorn inert de la regió viva (1). Després fa falta una font d’energia, com ara un mineral que experimenti una reacció que alliberi calor (2), la qual ha d’anar acoblada (3) a processos metabòlics (4). Finalment, cal que el sistema incorpori més elements dels que consumeix, per poder créixer i, eventualment, reproduir-se (5).
A partir de Shapiro, R., 2008.
En el primer acte de la història de l’origen de la vida van haver d’aparèixer conjunts de molècules de carboni capaces de sintetitzar còpies de si mateixes. En aquest sentit, el carboni presenta una sèrie de característiques fisicoquímiques que el fan insubstituïble en les molècules orgàniques, com per exemple la capacitat de formar llargues cadenes i d’unir-se al mateix temps a altres àtoms o d’emmagatzemar gran quantitat d’energia en aquestes unions. Tanmateix, però, els compostos de carboni més abundants en la Terra primitiva eren gasos amb un sol àtom de carboni, com el diòxid de carboni (CO2), el monòxid de carboni (CO) i el metà (CH4). Però els components essencials dels éssers vius, com els sucres, els àcids grassos, els aminoàcids i els nucleòtids, poden tenir més d’una dotzena d’àtoms de carboni per molècula. A més, aquestes molècules s’han de combinar per a construir cadenes més llargues, com els glícids i lípids d’estructura complexa, i les proteïnes i els àcids nucleics, respectivament.
Els orígens moleculars de la vida
Inicialment, totes aquestes molècules es van haver de formar i augmentar de complexitat de forma relativament independent entre elles, per arribar un moment en què comencessin a interactuar per a formar estructures encara més complexes, com per exemple membranes protectores amb un interior aïllat però al mateix temps comunicat amb l’exterior; sistemes de replicació del material hereditari i de síntesi de proteïnes a partir de la informació continguda en aquest material, etc. Des d’aquest punt de vista, hom ha de pensar en els orígens de la vida, en plural, no pas perquè la vida que existeix a la Terra provingui de diversos organismes primigenis fundadors, sinó per l’origen químic en paral·lel de totes les molècules que, en darrer terme, van acabar sent necessàries per a constituir aquest primer organisme al qual es pugui aplicar el qualificatiu de viu.
L’origen termodinàmic de la vida
Tanmateix, és molt improbable la participació de molècules extraordinàriament complexes com l’RNA, i el DNA, les proteïnes i altres macromolècules en l’origen de la vida en la mateixa forma com les coneixem avui. En aquest sentit, hom necessita incorporar també una visió termodinàmica –o metabòlica– a l’estudi de l’origen de la vida, la qual es basa en cinc principis bàsics.
Per començar, cal una regió circumscrita on pugui augmentar l’ordre. És el que fan totes les cèl·lules quan creixen i es reprodueixen: utilitzen energia per a construir molècules orgàniques que augmenten l’ordre del sistema, i al mateix temps alliberen calor, que fa augmentar l’entropia de l’entorn. I perquè això succeeixi, cal una barrera que separi l’ordre intern de l’entropia externa, que separi allò que és viu del que no ho és. També cal que hi hagi una font d’energia que impulsi el procés d’organització. Molts organismes actuals consumeixen glícids i lípids i els combinen amb oxigen per a obtenir energia metabòlica (organismes heteròtrofs), però també n’hi ha que utilitzen altres fonts energètiques (organismes autòtrofs), com la llum solar (fotoautòtrofs) o l’energia química dels minerals (quimioautòtrofs i litòtrofs). A més, l’alliberament d’energia ha d’estar acoblat al procés d’organització que produeix i sustenta la vida, mitjançant mecanismes metabòlics. També cal que hi hagi un entramat de reaccions químiques que permeti l’adaptació i l’evolució. I, finalment, és necessari que aquest entramat de reaccions químiques pugui créixer i reproduir-se. És a dir, cal que sigui capaç de captar matèria i energia més ràpidament del que les consumeix. Des d’aquest punt de vista metabòlic, hom ha de pensar en l’origen de la vida, en singular. En el primer ésser que es pogués considerar viu, encara que s’estableixi amb fonaments teòrics, van concórrer tant els orígens moleculars com l’origen termodinàmic o metabòlic de la vida.
L’evolució prebiòtica
El carboni és un element clau en l’evolució del material orgànic prebiòtic, i esdevé biològicament interessant quan forma llargues cadenes i es combina amb nitrogen, oxigen i hidrogen, entre altres elements químics, com per exemple fòsfor i sofre. Les dades de què es disposa sobre l’aparició de la vida a la Terra se sustenten en els coneixements que es tenen sobre la formació de la Terra i sobre la síntesi química abiòtica de molècules orgàniques en les condicions de la Terra primitiva.
L’escorça i l’atmosfera de la Terra primitiva
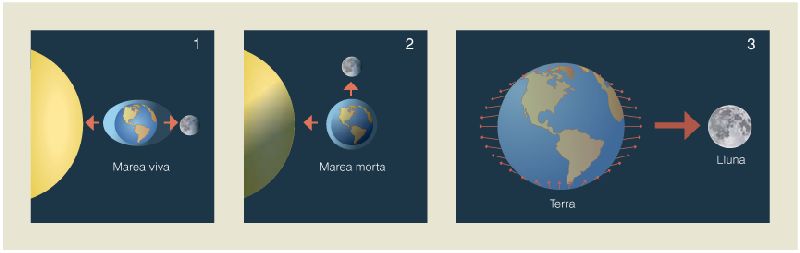
Les marees són degudes a l’atracció gravitatòria d’uns astres sobre uns altres. En el cas de la Terra, les forces de marea són causades per l’atracció del Sol i de la Lluna (1 i 2). Després de l’impacte amb el planetesimal que va originar la Lluna, la major proximitat a la Terra el va fer capaç de deformar també l’escorça terrestre (3).
A partir de fonts diverses.
La Terra es va formar fa uns 4.550 Ma per l’acreció de planetesimals la composició dels quals era similar a la de determinats meteorits anomenats condrites. El 95% de les condrites són constituïdes per materials ferruginosos (condrites ordinàries) i menys del 5%, per materials carbonosos (condrites carbonàcies). Fou un procés d’acreció a alta temperatura, durant el qual els compostos gelats es van sublimar i els materials carbonosos es van pirolitzar. En conseqüència, qualsevol molècula orgànica s’ha d’haver format després del refredament de l’escorça, o bé ha d’haver arribat juntament amb cossos celestes de petites dimensions que hi van impactar un cop ja era prou freda.

Impacte hipotètic d’un meteorit sobre la Terra primitiva.
NASA / Don Davis.
Uns 50 Ma després de la seva formació un planetesimal de la mida de Mart va col·lidir amb la Terra. Part de les restes originades es van quedar en òrbita, a uns 25.000 km de la Terra, i amb el temps es van agregar fins a formar la Lluna. Aquesta òrbita inicial tan propera –actualment la Lluna orbita a una distància mitjana de 384.400 km de la Terra–, en combinació amb la velocitat de rotació més elevada que tenia la Terra primerenca, feien que les forces de marea fossin tres-centes vegades més potents que en l’actualitat, per la qual cosa eren capaces de deformar la jove escorça terrestre.
Després d’aquesta col·lisió, la Terra va continuar rebent impactes d’asteroides i meteorits de desenes de quilòmetres de diàmetre, amb una freqüència mitjana estimada d’un projectil d’aquestes dimensions cada 1.000 anys. Hi ha qui diu que aquests impactes, juntament amb la desintegració dels elements radioactius de l’interior del planeta i la deformació de l’escorça deguda a les potents forces de marea, van fer que la Terra es trobés en estat de fusió durant uns quants milions d’anys, malgrat que l’existència d’aquest oceà de magma és un tema molt debatut. Durant aquest període, que es coneix amb el nom d’Hadeà, les temperatures eren tan altes que els elements més pesants, entre els quals cal destacar el ferro, es van anar enfonsant fins a formar el nucli, mentre que els més lleugers van formar el mantell i l’escorça.

Surgència hidrotermal situada a l’oceà Atlàntic, amb emissió de gasos reductors com ara metà, amoni i hidrogen, els quals són imprescindibles per a la síntesi abiòtica de compostos orgànics.
National Oceanic and Atmospheric Administration, NOAA.
Aquest fet també és important per a l’origen de la vida, per tres motius. D’una banda, la dinàmica del ferro defineix l’estat d’oxidació de l’escorça i, en conseqüència, la composició de l’atmosfera –i cal una atmosfera reductora per a la síntesi química abiòtica de moltes molècules orgàniques–. El ferro és extremament reactiu en presència d’oxigen, per la qual cosa l’anàlisi de l’estat d’oxidació de les roques permet inferir aspectes clau sobre l’evolució de l’atmosfera. En aquest sentit, els sediments que es van formar just després del període Hadeà, durant l’anomenat Arqueà, presenten sobretot formes no oxidades del ferro, com pirita (FeS2), la qual es dissol fàcilment de les roques en presència d’oxigen, com en l’actualitat. En segon lloc, l’enfonsament dels materials més pesants va permetre que l’escorça s’enriquís molt en carboni. El tercer motiu és que l’existència d’un nucli metàl·lic és imprescindible per a la formació d’un camp magnètic, que actua d’escut de protecció contra les radiacions còsmiques.
Els primers fòssils corresponen a uns éssers vius que són semblants a determinats cianobacteris actuals, com aquest exemplar d’Anabaena sp.
Kristian Peters
El període Hadeà va acabar fa uns 3.800 Ma i va donar pas a l’Arqueà, dins el qual s’han trobat els primers fòssils d’éssers vius, relativament semblants a determinats cianobacteris actuals. El marge per a l’origen de la vida no és gaire ampli. En aquest context, l’atmosfera que hi havia fa entre 4.400 i 3.800 Ma era una barreja de compostos volàtils portats pels cometes, entre els quals hi havia determinades molècules orgàniques que poden haver tingut un paper important en l’origen de la vida, i de gasos alliberats des de l’interior de la Terra, el component majoritari dels quals era vapor d’aigua, que no es podia condensar i formar oceans atès que la temperatura era superior al seu punt d’ebullició (100 ºC). Era dominada per gasos oxidats, com diòxid i monòxid de carboni (CO2 i CO, respectivament), i nitrogen (N2). També hi havia quantitats significatives d’hidrogen (H2), òxid de sofre (SO2) i sulfur d’hidrogen (H2S). Tanmateix, prop dels volcans i de les surgències hidrotermals hi deuria haver concentracions locals significatives de gasos reductors com metà (CH4), amoni (NH3) i també hidrogen (H2), entre d’altres, els quals són imprescindibles per a la síntesi abiòtica de compostos orgànics. Aquest fet situa en aquest ambient el possible origen de la vida.
Pel que fa al clima, aquest depenia de diversos factors, entre els quals cal destacar el camp magnètic, que molt probablement no estava plenament desenvolupat, la qual cosa feia que la termosfera fos molt més dinàmica i molt més sotmesa a l’influx solar; la lluminositat del Sol, que era el 30% menor que en l’actualitat, per la qual cosa, si l’atmosfera hagués tingut la mateixa composició que l’actual, absolutament tota la superfície terrestre hagués romàs congelada; i les propietats de difusió de la calor de l’atmosfera primitiva. Tanmateix, el rang d’energies emeses pel Sol, com la dosi de raigs X i d’energia ultraviolada, clarament superiors a les actuals, devien afectar l’estabilitat i la pervivència de moltes espècies químiques, entre les quals cal esmentar, especialment, els compostos orgànics, atès que l’energia que els confereixen desestabilitzen llurs enllaços moleculars.
Les primeres biomolècules
Les reaccions característiques de la química orgànica no són exclusives dels éssers vius, sinó que es produeixen en molts llocs de l’Univers, com per exemple en les nebuloses fosques, en asteroides i cometes i també en agregats de gel que es troben al límit exterior dels sistemes solars. Determinades molècules orgàniques produïdes fora de la Terra poden haver contribuït, i molt probablement ho han fet, a l’origen de la vida, i moltes d’altres s’han originat per síntesi química abiòtica a la Terra mateixa.
Síntesi química a la Terra
Les reaccions químiques concretes a partir de les quals, amb les molècules i condicions de la Terra primitiva, es poden formar molècules orgàniques de rellevància biològica són certament complexes, i fer-ne un catàleg exhaustiu queda fora del propòsit d’aquest suplement, per la qual cosa s’oferirà un exemple de cadascun dels grups de biomolècules constituents dels éssers vius –aminoàcids, nucleòtids, glícids i lípids–. Tots els casos provenen d’experiments realitzats al laboratori, en què, com en els experiments clàssics de Miller, Urey i Oró, es reprodueixen aspectes concrets de la Terra i de l’atmosfera primitives.
Els aminoàcids són les molècules elementals que formen les proteïnes. Són uns compostos orgànics formats per un grup carboxil (COOH), un grup amina (NH2) i una cadena lateral, la qual els dóna especificitat. En les cèl·lules actuals, els aminoàcids són ingerits amb l’aliment o bé sinteritzats amb la participació d’enzims i rutes metabòliques específiques. Tanmateix, en les condicions atmosfèriques i energètiques globals de la Terra primitiva, la reacció seqüencial de molècules orgàniques senzilles com formaldehid (CH2O), cianur (CN) i aigua (H2O) genera directament un dels aminoàcids més simples, la glicina (NH2CH2COOH). I si en comptes de formaldehid hi participen altres aldehids específics, es pot originar qualsevol dels vint aminoàcids diferents que típicament constitueixen les proteïnes.
Una via paral·lela considera que, quan el diòxid de carboni i l’aigua se sotmeten a descàrregues elèctriques, llum ultraviolada i radiacions ionitzants com raigs X, raigs gamma i radioactivitat, s’obté un elevat nombre de molècules orgàniques que formen unes mescles d’aspecte vermell marronós anomenades tolines; i les tolines, en presència d’altres molècules de localització més restringida com metà i amoni, que es trobaven en concentracions especialment elevades en surgències hidrotermals, també poden generar reaccions abiòtiques de síntesi d’aminoàcids.
Pel que fa als nucleòtids, són les molècules elementals que formen els àcids nucleics, com el DNA i l’RNA. Són formats per un glícid cíclic de cinc carbonis –una ribosa en l’RNA i una desoxiribosa en el DNA, una diferència interessant per a interpretar l’origen dels àcids nucleics en els éssers vius–; per una base nitrogenada que els confereix especificitat –citosina (C), guanina (G), adenina (A) i uracil (U) en l’RNA; i les mateixes en el DNA llevat de la U que és reemplaçada per timina (T), una diferència que també és interessant per a interpretar l’origen dels àcids nucleics en els éssers vius–, i per un o diversos grups fosfat (PO43–, un tipus de molècula inorgànica). En les condicions de la Terra primitiva, s’ha demostrat que es pot formar espontàniament adenina simplement a partir d’àcid cianhídric (HCN) i amb intervenció d’energia lumínica. I perquè es puguin formar les altres bases nitrogenades n’hi ha prou amb la participació de diversos aminoàcids i d’altres molècules orgàniques, bàsicament formaiats (CHOO−) i fosfat de carbamil (H2O3PO(CO)NH2).
Els glícids, al seu torn, són una família de compostos orgànics d’una gran diversitat estructural. Són formats per una cadena d’àtoms de carboni que pot ser lineal o bé cíclica, i es poden trobar sols, com els monosacàrids, o bé units en cadenes de longitud variable, com els oligosacàrids i els polisacàrids. Compleixen funcions energètiques, són el substrat per a la síntesi d’altres molècules biològiques i formen part de determinades estructures cel·lulars i dels àcids nucleics. Pel que fa a la seva síntesi química abiòtica, es poden generar a partir de diversos precursors, com aldehids i àcid cianhídric, en presència de fosfats. Tanmateix, però, són molt inestables en les condicions ambientals en què es poden formar espontàniament per síntesi química. Una de les explicacions més plausibles per a resoldre aquesta inestabilitat és que, un cop formats, determinats minerals com els borats els poden estabilitzar; i els minerals eren ben presents en els ambients on es va originar la vida (vegeu “Els minerals en l’origen de la vida").
Finalment, els lípids són un grup molt heterogeni de biomolècules orgàniques, insolubles en aigua però solubles en dissolvents orgànics. Són formats per una cadena lineal d’àtoms de carboni, i formen part de diverses estructures biològiques, com les membranes cel·lulars. També actuen de reserva energètica. S’ha demostrat que en condicions d’alta temperatura i ambients reductors, com els que es troben al voltant de les surgències hidrotermals, es poden sintetitzar químicament compostos lipídics de fins a 35 àtoms de carboni encadenats, a partir de determinades molècules orgàniques com àcid fòrmic o àcid oxàlic. I a partir d’aquests lípids, en presència d’hidrogen i de diòxid i monòxid de carboni, es poden formar moltes altres molècules lipídiques.
Síntesi química a l’espai
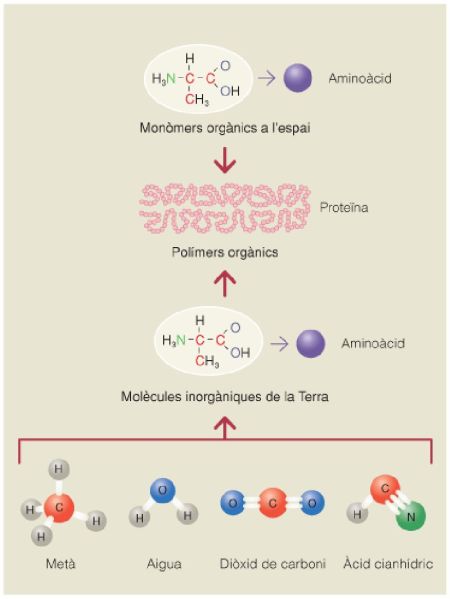
Formació d’aminoàcids i de proteïnes a partir dels compostos inorgànics i de les condicions atmosfèriques i energètiques globals presents en la Terra primitiva, amb la possible contribució de molècules orgàniques sintetitzades químicament a l’espai.
A partir de fonts diverses.
No es descarta la possibilitat que algunes molècules orgàniques s’hagin sintetitzat químicament a l’espai, i que hagin arribat a la Terra juntament amb determinats meteorits. Es coneix un tipus de meteorits, les condrites carbonàcies, que contenen carboni en forma de compostos orgànics, en proporcions que oscil·len entre l’1,5 i el 4%. S’ha detectat la caiguda de 37 d’aquests meteorits des de l’any 1806, alguns dels quals han estat molt analitzats. Per exemple, el meteorit Murchinson, de més de 100 kg de pes que va caure sobre el poble australià del mateix nom el 28 de setembre de 1969, contenia més d’un centenar de molècules orgàniques diferents, com per exemple aminoàcids, sucres i bases nitrogenades constituents dels àcids nucleics, entre d’altres.
A més, l’estudi espectroscòpic de les nebuloses fosques interestel·lars ha indicat que contenen centenars de compostos orgànics diferents. Aquestes molècules es poden formar en núvols foscos de l’espai profund gràcies a determinades característiques del gel interestel·lar, el qual, a temperatures properes al zero absolut, no es troba en forma cristal·lina sinó amorfa d’alta densitat, de manera que les seves molècules presenten una mobilitat equiparable a la de l’aigua líquida a la Terra.
Els minerals en l’origen de la vida
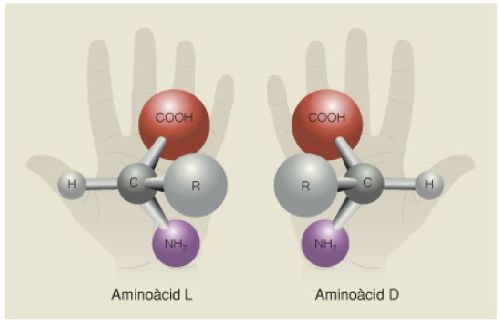
Quiralitat dels aminoàcids, amb l’estructura general d’aquestes biomolècules: el grup carboxil (COOH), el grup amino (NH2) i la cadena lateral que els distingeix. Les dues formes quirals, la L i la D, són la imatge especular l’una de l’altra.
A partir de fonts diverses.
Qualsevol model de gènesi química de molècules orgàniques a la Terra ha d’assumir que les molècules implicades s’havien de trobar en concentracions adequades, en un mateix moment i en un mateix indret, perquè poguessin interactuar entre elles. Tanmateix, hi ha poques molècules orgàniques que siguin termodinàmicament estables en aigua, per la qual cosa cal pressuposar la contribució de minerals que n’hagin incrementat l’estabilitat, com per exemple el borat, que estabilitza els glícids; o que hagin afavorit o contribuït a llur reactivitat. Hi ha diversos exemples que han estat demostrats en condicions de laboratori on s’han recreat diversos aspectes de la Terra primitiva.
Per exemple, qualsevol superfície cristal·lina submergida en aigua indueix la formació d’una capa molecular d’ions amb càrrega positiva (en forma de protons o H+) o alternativament amb càrrega negativa (en forma de grups hidroxil o OH–), la càrrega dels quals pot afavorir determinades reaccions orgàniques. També s’ha vist que es produeixen interaccions directes entre determinats minerals i molècules orgàniques. Per exemple, els òxids i els sulfits metàl·lics promouen diverses reaccions, i en presència d’òxid de coure es forma amoníac, essencial per a la síntesi química abiòtica de determinades molècules orgàniques com els aminoàcids.
Un altre cas interessant és l’efecte que té l’argila sobre moltes molècules orgàniques, atès que les atrau i concentra. Aquests minerals, presents en molts ambients, es tornen untuosos quan es mullen perquè els seus àtoms formen capes planes i llises, les quals sovint presenten una càrrega elèctrica capaç d’atreure molècules orgàniques i retenir-les. Per exemple, s’ha demostrat que quan s’evapora l’aigua que hi ha sobre les argiles, un fet no gens infreqüent en bassals i en situacions de marees, si hi ha aminoàcids aquests tendeixen a polimeritzar, un procés d’enllaç químic en cadena mitjançant el qual es formen molècules de tipus proteic. Un efecte similar s’ha observat també en argiles de gra fi i en minerals hidroxilats pel que fa a l’RNA, i s’ha vist que fan de bastida per a la construcció dels components d’aquest àcid nucleic. També s’ha demostrat que faciliten l’autoorganització de molècules lipídiques, com les que formen la membrana cel·lular de tots els éssers vius.
Finalment, la participació de minerals en l’origen de la vida permet resoldre una altra qüestió, la quiralitat dels aminoàcids. Com passa amb altres molècules orgàniques, com els glícids, tots els aminoàcids poden presentar dues variants, que contenen els mateixos àtoms però disposats en l’espai de manera especular. Aquest fenomen, que s’anomena quiralitat, fa que de cada aminoàcid existeixi una versió levogira (L) i una de dextrogira (D). En els experiments de síntesi abiòtica d’aminoàcids, com el de Miller i Urey, sempre s’obtenen mescles al 50% d’aminoàcids L i D, però en els éssers vius pràcticament el 100% dels aminoàcids són L. El motiu és que determinats minerals cristal·lins, com la calcita i molts òxids i silicats, presenten també cares quirals en els seus cristalls, D i L, les quals uneixen específicament aminoàcids D o L respectivament. Aquesta unió afavoreix que polimeritzin de forma espontània i formin cadenes proteiques, constituïdes exclusivament per aminoàcids D o alternativament L. Aquest fet permet inferir que, per atzar, el primer conjunt de molècules autoreproductores que va esdevenir precursora de totes les formes de vida de la Terra es va produir en una cara L d’un d’aquests cristalls.
Bioenergia i origen de la vida
Finalment, cal considerar també la natura i la disponibilitat de la font o fonts d’energia necessàries per a la formació química abiòtica de biomolècules, com per exemple per a la unió d’aminoàcids i nucleòtids per a formar les proteïnes i els àcids nucleics de rellevància biològica, i per al posterior manteniment dels éssers vius. En les cèl·lules actuals, la formació d’enllaços químics entre aquestes molècules requereix la participació d’enzims i d’altres molècules, entre les quals hi ha molècules donadores i acceptors d’energia metabòlica –com l’ATP, o trifosfat d’adenosina– o de poder reductor –és a dir, d’electrons, com el NADH/NADHP o dinucleòtid de nicotinamida i adenina–. Per exemple, en l’enllaç que s’estableix entre aminoàcids per a formar les proteïnes –enllaç peptídic– es requereix la participació de diverses proteïnes i molècules d’RNA. A més, es realitza en uns orgànuls cel·lulars específics, els ribosomes, formats al seu torn també per diverses molècules específiques d’RNA i per proteïnes; i es fa sota el guiatge estricte de la informació genètica, que és transportada del DNA als ribosomes mitjançant una molècula intermediària, l’RNA missatger o RNAm.
Per que fa als nucleòtids, l’enllaç que estableixen per a formar les cadenes de DNA i d’RNA –enllaç fosfodièster– és catalitzat per l’activitat d’uns enzims que genèricament s’anomenen polimerases, els quals literalment copien la informació a partir de cadenes motlle preexistents. El fet d’esmentar aquests processos no és gratuït, atès que com es veurà són clau per a interpretar l’origen de la vida.
A la Terra primitiva no hi havia cap d’aquests sistemes metabòlics, però tanmateix hi havia diverses fonts energètiques disponibles per a impulsar aquestes transformacions –és a dir, per a activar els monòmers (aminoàcids, nucleòtids, etc.), perquè es poguessin unir i formar polímers (proteïnes, àcids nucleics, etc.)–, totes elles anaeròbiques atesa la manca d’oxigen en l’atmosfera primitiva. De forma general, es poden classificar en tres grans grups: energia fotoquímica present en la llum ultraviolada, descàrregues elèctriques atmosfèriques i energia geològica electroquímica. En aquest sentit, per esmentar alguns dels molts casos analitzats, l’energia continguda en la llum ultraviolada i en les descàrregues elèctriques pot fer que el CO2, l’H2O i l’amoni (NH4) es combinin espontàniament per a formar cianida (HCN), la qual pot reaccionar amb si mateixa per a generar molècules orgàniques més complexes; i també formaldehid (CH2O), el qual, en un medi alcalí, pot formar diversos glícids. Pel que fa a l’energia geològica electroquímica, tant el metà (CH4) com molt especialment els sulfurs –sulfit d’hidrogen (H2S) i diversos sulfats de ferro– poden actuar de donadors d’electrons, els quals, per exemple, permeten que de forma espontània l’H2S i el CO reaccionin i formin molècules com tioèster (CH3-CO-SCH3), la qual permet que els aminoàcids s’uneixin aleatòriament per a formar proteïnes, sense cap intervenció enzimàtica.
Els primers éssers vius
Abans de l’aparició de la vida, molt probablement la matèria orgànica present a la Terra primitiva consistia en una àmplia varietat de compostos diferents, entre els quals hi havia aminoàcids i proteïnes, nucleòtids i àcids nucleics, lípids i glícids de diversa complexitat, etc. Tanmateix, la manera com es van encaixar aquests components per a formar els primers organismes vius és encara un tema molt debatut. Una de les millors maneres d’abordarlo és analitzar com devia ser el darrer avantpassat comú a tots els organismes actuals, un organisme hipotètic que es coneix amb el nom de LUCA (de l’anglès Last Universal Common Ancestor, ‘darrer ancestre comú universal’).
La posició filogenètica de LUCA
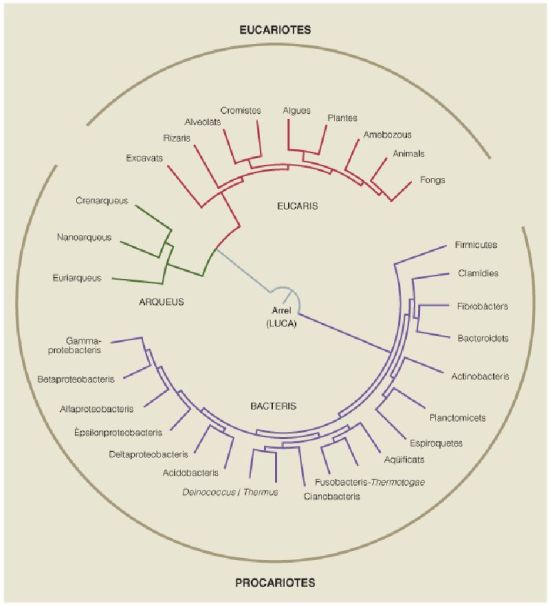
Arbre filogenètic circular on es veu la posició de LUCA a l’arrel, just abans de l’embrancament que separa el domini dels bacteris dels dominis dels arqueus i dels eucaris. S’indiquen els principals grups dins de cada domini, i si són organismes procariotes –formats per cèl·lules sense nucli– o eucariotes –formats per cèl·lules amb nucli–. Atès que el primer embrancament després de LUCA origina, d’una banda, el domini dels bacteris, que és procariota; i de l’altra el domini dels arqueus, també procariota, i el dels eucaris, que és eucariota, cal pressuposar que d’alguna manera l’estructura cel·lular de LUCA també era de tipus procariota.
A partir de dades de l’autor.
Les característiques que comparteixen tots els éssers vius, com per exemple el codi genètic, els mecanismes implicats en l’expressió gènica i les reaccions metabòliques bàsiques, entre d’altres, són tan nombroses i tan centrals en els processos vitals que és impossible assumir que han anat apareixent i evolucionant de manera independent i al mateix temps convergent en els diferents grans dominis d’éssers vius –bacteris, arqueus i eucaris–. En aquest sentit, els detalls moleculars d’aquests processos universals proporcionen una prova directa de l’origen monofilètic de la vida, és a dir, a partir d’un únic organisme ancestral –LUCA– del qual totes les formes de vida procedeixen per evolució –per canvi sotmès a l’atzar preadaptatiu i per selecció natural–. A més, les diferències que s’observen entre aquests llinatges poden ser explicades amb facilitat considerant els processos evolutius que han menat d’aquesta forma de vida ancestral fins a les actuals. Per descomptat, no hi ha cap resta geològica de l’existència concreta d’aquest antecessor universal, per la qual cosa la seva natura cel·lular, metabòlica i genètica s’ha d’inferir a partir de la comparació dels éssers vius actuals, mitjançant anàlisis filogenètiques, que el situen a la base de l’arbre de la vida.
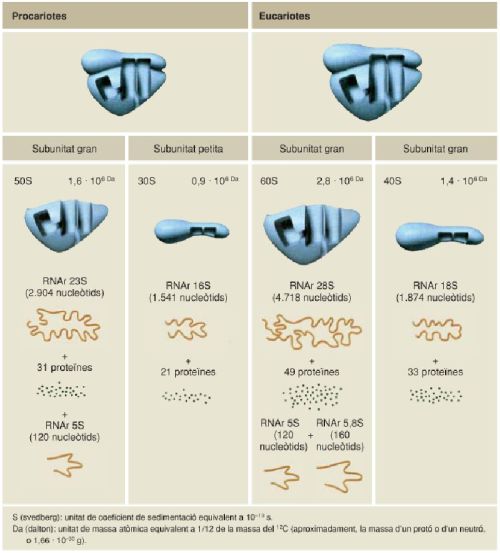
Comparació dels ribosomes típics dels organismes procariotes i dels eucariotes, amb els RNAr i les proteïnes que els formen. La comparació d’aquestes biomolècules i del mecanisme de funcionament dels ribosomes indica clarament l’origen filogenètic comú de totes elles.
A partir de fonts diverses.
Aquestes anàlisis s’han aplicat a un gran nombre d’organismes dels tres dominis d’éssers vius, i s’han comparat multitud de molècules, entre les quals les més conservades en tots els llinatges, com els RNA que formen els ribosomes –els RNA ribosòmics o RNAr–, els factors proteics que intervenen en la còpia del material genètic –com els anomenats factors d’elongació EF-G i EF-Tu–, els RNA que transporten els aminoàcids per a permetre que es vagin unint de forma específica en funció del missatge genètic –els RNA de transferència o RNAt– o els enzims que de forma específica els uneixen a aquests aminoàcids –anomenats genèricament aminoacil sintases de l’RNAt.
Els algoritmes de construcció d’arbres filogenètics han situat LUCA a la base de l’embrancament que separa, d’una banda, els bacteris, i de l’altra el grup format pels arqueus i els eucaris. Tanmateix, aquest tema encara no està del tot resolt, perquè s’han descobert molts casos de transferència gènica horitzontal entre organismes de tots tres dominis que dificulten les inferències filogenètiques, atès que tendeixen a agrupar artificialment determinats grups en funció dels gens analitzats i distorsionen els arbres filogenètics resultants. La transferència gènica horitzontal és un procés biològic mitjançant el qual un organisme incorpora material genètic d’un altre sense ser-ne el descendent. Tanmateix, com es discutirà a continuació, la transferència gènica horitzontal pot haver estat crucial en l’origen de la vida.
La natura del darrer ancestre comú universal
Són diversos els aspectes que cal analitzar quan s’aborda la natura del darrer ancestre comú universal, com per exemple el seu material genètic i les característiques de la seva replicació i expressió, incloent-hi el codi genètic, la formació de les primeres membranes cel·lulars i l’establiment de mecanismes de transport. Novament, atesa la dificultat d’analitzar la natura de LUCA des d’un punt de vista global, cal recórrer a una aproximació reduccionista i analitzar per separat els diversos components que caracteritzen la vida.
El material genètic
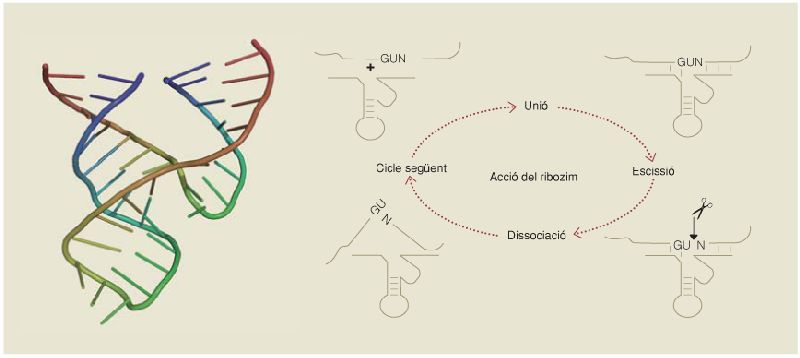
Estructura molecular i funció enzimàtica d’un ribozim model, que es coneix amb el nom de cap de martell (hammerhead ribozyme). Aquest ribozim és capaç de codificar informació genètica en la seva seqüència de nucleòtids, i al mateix temps d’actuar com un enzim, en aquest cas tallant per un punt concret molècules específiques.
A partir de fonts diverses.
Actualment, el genoma de tots els organismes procariotes i eucariotes és format per DNA, per la qual cosa la conclusió més parsimoniosa des del punt de vista filogenètic –és a dir, la que pressuposa el mínim nombre de canvis evolutius– és que aquesta molècula ja era present en LUCA, on exercia la mateixa funció. Tanmateix, l’existència d’un genoma inicial format per DNA presenta alguns problemes de difícil resolució, bàsicament pel que fa a la seva replicació –és a dir, al procés mitjançant el qual es copia–, atès que sense ell no s’hagués pogut propagar a la seva progènie. Per a replicar-se en totes les cèl·lules actuals el DNA necessita una sèrie d’enzims, entre els quals hi ha les polimerases del DNA, de natura proteica, que es troben codificades en el mateix DNA. Això genera una paradoxa. Si per a fer que el DNA es copiï calen unes proteïnes específiques, i per a fer aquestes proteïnes cal la informació contingu da al DNA, com podia la primera molècula de DNA contenir la informació necessària per a fer les proteïnes que li permetrien copiar-se?
Imatge hipotètica d’una protocèl·lula: una membrana lipídica envolta l’RNA ribosòmic.
Exploring Origins Project, National Science Foundation.
L’any 1967, Carl Woese (n. 1928), conegut per haver proposat la classificació en tres dominis dels éssers vius, Francis Crick (1916 – 2004), conegut per haver participat en el descobriment de l’estructura del DNA, i Leslie Orgel (1927 – 2007) van proposar, basant-se en el plegament que presenten algunes molècules d’RNA, que determinats RNA es poden comportar com enzims. Aquests ribozims, com se’ls anomena, l’activitat dels quals va ser demostrada per Thomas Cech (n. 1947), reuneixen en una sola molècula les característiques dels àcids nucleics pel que fa a la capacitat d’emmagatzemar informació genètica en la seva seqüència, i de les proteïnes pel que fa a tenir activitat enzimàtica.
La descoberta dels ribozims, alguns dels quals s’ha demostrat que poden copiar àcids nucleics i unir aminoàcids en cadenes proteiques, entre moltes altres funcions, va suggerir la hipòtesi que, al principi de la vida, el material genètic podia haver estat l’RNA en comptes del DNA, en forma de petites molècules que emmagatzemaven informació genètica i al mateix temps actuaven d’enzims, catalitzant les reaccions necessàries per a la vida. Una de les proves que ha deixat aquest món primigeni basat en l’RNA es troba en la maquinària de l’expressió gènica, el procés mitjançant el qual la informació continguda al DNA dirigeix la síntesi de proteïnes específiques, un procés dominat precisament per diverses molècules d’RNA, com els RNAm –que transporten la informació del DNA als ribosomes, on se sintetitzen les proteïnes–, els RNAr –que formen bona part de l’estructura dels ribosomes, i que en les cèl·lules actuals encara mostren activitat catalítica de ribozim–, i els RNAt –que incorporen un aminoàcid concret a la proteïna en creixement seguint les instruccions codificades a l’RNAm, en virtut del codi genètic.
En algun moment abans de LUCA, però, aquest món d’RNA va donar pas, per selecció natural, al món del DNA i de les proteïnes, probablement afavorit per la major especialització d’aquestes molècules pel que fa a l’emmagatzematge d’informació genètica i a l’activitat enzimàtica respectivament, una fisiologia que ja devia ser present en LUCA atès que tots els éssers vius actuals emmagatzemen la informació genètica en el DNA i utilitzen bàsicament les proteïnes com a molècules enzimàtiques. En aquest sentit, el DNA pot ser considerat com una forma modificada d’RNA en la qual el sucre ribosa és reduït en forma de desoxiribosa. Per la seva base nitrogenada uracil present en l’RNA és molecularment més senzilla que la base nitrogenada timina, l’equivalent que es troba en el DNA, la qual no és res més que un uracil al qual s’ha afegit un grup metil. Atenent a com els éssers vius actuals sintetitzen aquests àcids nucleics, sembla que el pas d’RNA a DNA es va fer en dues etapes principals: primer, es va passar de la ribosa de l’RNA a la desoxiribosa del DNA, i posteriorment de l’uracil a la timina.
Fins i tot hi ha qui ha proposat que la primera molècula capaç d’emmagatzemar informació genètica i d’actuar com a enzim no va ser l’RNA, sinó l’anomenat PNA o àcid nucleic peptídic, format per una variant acetilada de nucleòtids l’energia interna dels quals podria facilitar la seva polimerització en les condicions de la Terra primitiva. En aquest sentit, en l’experiment original de Miller i Urey es van aïllar alguns d’aquests nucleòtids acetilats, però no obstant això la seva estabilitat és clarament menor que la de l’RNA.
Sigui com sigui, la gran pregunta és per què es va acabar seleccionant el DNA en comptes de l’RNA com a material genètic. L’explicació més acceptada és que, des del punt de vista molecular, el DNA és més estable que l’RNA, i a més en cas que es produeixin alteracions en la seva estructura es pot reparar amb més facilitat i fidelitat. També hi ha qui ha proposat que el DNA va ser utilitzat inicialment per determinats virus per a evitar ser degradats, i que l’eficiència més alta d’aquesta molècula com a transportadora de la informació genètica va afavorir que fos adoptada per les protocèl·lules inicials. Sigui com sigui, es considera que a través de processos de selecció natural típicament darwinians es va passar progressivament d’un primer estadi de química abiòtica dels nucleòtids a una bioquímica catalitzada per l’RNA; d’aquí a una vida primordial basada en l’RNA; i, finalment, a una vida basada en el DNA i en les proteïnes, en la qual l’RNA es va mantenir com a molècula intermediària imprescindible del procés d’expressió gènica.
La transcripció, la traducció i els ribosomes
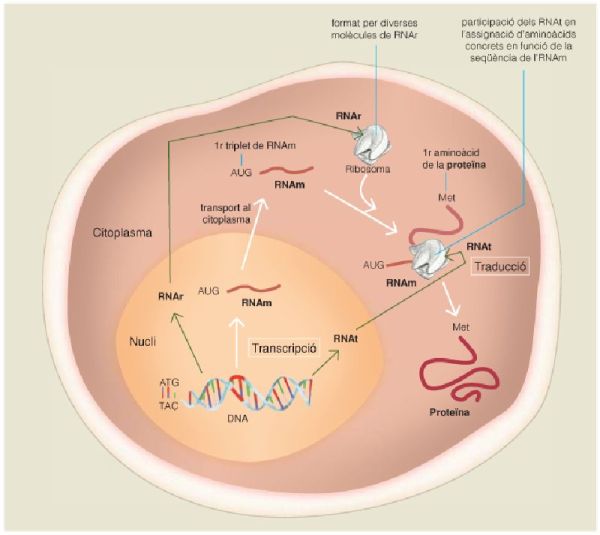
Flux d’informació gènica des del DNA a les proteïnes en una cèl·lula eucariota. La universalitat d’aquest flux i l’enorme conservació funcional i estructural de les molècules i enzims que hi intervenen, els quals han demostrat tenir un origen filogenètic comú tant en bacteris i arqueus (organismes amb estructura cel·lular procariota, sense nucli) com en eucaris (organismes amb estructura cel·lular eucariota, amb nucli), és una prova de l’origen comú de la vida i de la seva diversificació a partir de LUCA.
Bueno, 2008.
Un cop afegit el DNA com a molècula portadora de la informació genètica, cal considerar l’origen dels processos de transcripció i traducció, els quals es van fer imprescindibles. La transcripció és el pas de la informació del DNA a l’RNA, i en els organismes actuals és catalitzada per un enzim proteic anomenat polimerasa de l’RNA. A primer cop d’ull, la polimerasa de l’RNA procariota sembla prou diferent de l’eucariota per pressuposar un origen diferent, la qual cosa equivaldria a dir que aquest procés es va originar amb posterioritat a LUCA, de forma independent en aquests llinatges. Però una observació detallada de la seva seqüència revela l’existència d’un nucli catalític pràcticament idèntic en tots els éssers vius analitzats de tots tres dominis. En aquest sentit, la polimerasa de l’RNA dels arqueus s’assembla molt més a la dels eucaris que a la dels bacteris –per ser precisos, s’assembla molt a l’anomenada polimerasa II dels eucaris, atès que aquests organismes presenten diverses polimerases de l’RNA–. Tot això implica que aquesta activitat enzimàtica, realitzada per aquest nucli catalític compartit, es va originar amb anterioritat a l’existència de LUCA, i que amb posterioritat l’enzim encarregat va divergir en cadascun dels tres dominis fins a adquirir les formes actuals.
La traducció, al seu torn, és la utilització de la informació continguda a l’RNAm per a sintetitzar una proteïna específica, i es realitza en uns orgànuls cel·lulars molt concrets, els ribosomes, que són formats per diverses molècules d’RNA, els RNAr, i per proteïnes específiques. En aquest context, tant els RNAr com les proteïnes ribosòmiques de les cèl·lules actuals presenten diferències en funció del domini al qual pertanyen, però tanmateix també mostren clares homologies en tots els organismes analitzats dels tres dominis, la qual cosa indica que els seus components bàsics ja eren presents en LUCA. En aquest cas, els ribosomes dels arqueus s’assemblen molt més als dels bacteris que no pas als dels eucaris. Més encara, l’ordre i la manera com es van encaixant seqüencialment els diferents components dels ribosomes durant el seu assemblatge permet inferir-ne l’ordre d’aparició i la seva evolució prèvia a LUCA, moment en què van començar a divergir a mesura que també anaven divergint els diferents llinatges d’éssers vius.
El codi genètic
Un aspecte important de la traducció des del punt de vista evolutiu és l’establiment del codi genètic, atès que, com s’ha dit, és universal, per la qual cosa sens dubte es va establir abans de l’aparició de LUCA. El codi genètic determina la correspondència de cada aminoàcid a cada tres nucleòtids consecutius de l’RNAm, una unitat de codificació d’informació que s’anomena codó o triplet, de tal manera que l’ordre d’aminoàcids de la proteïna segueix l’ordre de codons de l’RNAm, els quals són còpia del missatge contingut al DNA –en conseqüència, és la informació continguda al DNA la que dirigeix la síntesi de proteïnes.
En aquesta relació entre codons i aminoàcids, que s’estableix físicament en els ribosomes, intervenen dues molècules que són clau: un RNAt, que reconeix el codó present en l’RNAm i transporta l’aminoàcid corresponent a aquest codó, de manera que es pugui incorporar al lloc precís que li correspon en la proteïna segons l’ordre de codons de l’RNAm; i un enzim anomenat aminoacil sintasa de l’RNAt, que uneix de forma específica cada aminoàcid a l’RNAt concret que l’ha de transportar. En aquest context, en general hi ha un aminoacil sintasa de l’RNAt específic per a cada aminoàcid i per a cada RNAt, i un RNAt específic per al reconeixement de cada codó a l’RNAm. L’anàlisi filogenètica de la seqüència dels diferents RNAt indica que tots provenen d’un únic RNAt ancestral per duplicacions successives i mutacions; i la dels aminoacil sintases de l’RNAt indica que tots provenen de dos gens ancestrals, també per duplicacions i mutacions successives, que han originat dues famílies clarament distintes d’aquests enzims.
El codi genètic, per tant, es va establir progressivament a mesura que l’RNA anava deixant pas al DNA com a suport físic de la informació genètica, i també a mesura que els enzims proteics anaven agafant el relleu als ribozims. I es va acabar d’establir abans de l’aparició de LUCA.
La replicació del DNA
Un altre aspecte que cal considerar en aquesta anàlisi reduccionista dels diversos elements que havien de concórrer en l’origen de la vida i en el darrer ancestre comú universal és la replicació del DNA. Com s’ha dit, el genoma de LUCA era format per DNA, i els mecanismes de transcripció i traducció, inclosos els ribosomes i el codi genètic, ja estaven establerts. En aquest context, també cal pressuposar que el mecanisme de còpia del DNA també havia d’estar establert; altrament LUCA no s’hagués pogut reproduir. Doncs bé, l’anàlisi dels enzims encarregats de la còpia del DNA, les anomenades genèricament polimerases del DNA, revela que tenen dos orígens filogenètics clarament diferents, atès que no hi ha cap relació estructural ni de seqüència entre la polimerasa del DNA que empren els organismes del domini dels bacteris, i la que fan servir els organismes dels dominis dels arqueus i dels eucaris, les quals sí que tenen un origen comú.
Aquest fet planteja la qüestió de com LUCA replicava el seu DNA. S’han proposat dues possibles hipòtesis, malgrat que el tema encara no està resolt. Una hipòtesi considera que LUCA tenia totes dues polimerases del DNA, les quals s’havien originat de forma independent, i que amb posterioritat a LUCA, en la branca que condueix als bacteris s’hauria perdut una de les dues polimerases, mentre que en la branca que condueix als arqueus i als eucaris s’hauria perdut l’altra. La hipòtesi alternativa considera que LUCA tenia una polimerasa del DNA diferent a aquestes dues, potser encara no proteica sinó en forma de ribozim, i que l’enzim proteic es va originar de forma independent en el llinatge dels bacteris i en el dels arqueus i dels eucaris, reemplaçant en ambdós casos l’activitat del ribozim inicial atesa l’efectivitat més alta dels enzims proteics.
De l’autoassemblatge de vesícules a les cèl·lules primitives
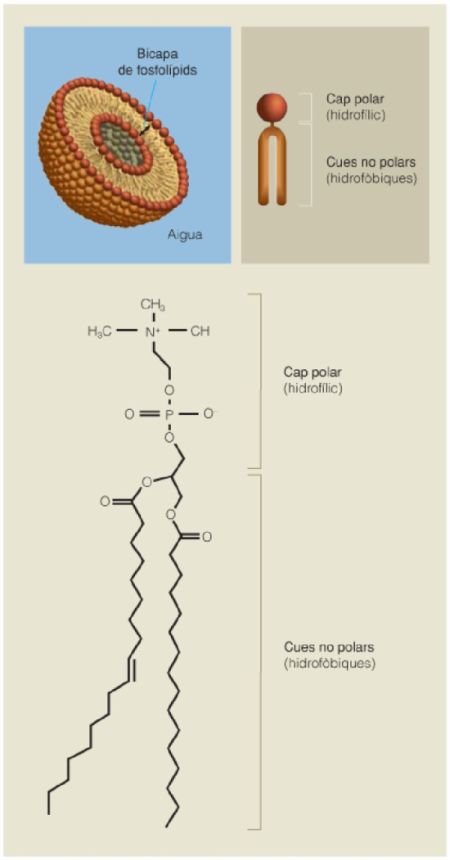
Estructura química d’un fosfolípid i vesícula delimitada per una bicapa de fosfolípids. Pot contenir un medi aquós relativament aïllat de l’exterior.
A partir de l’autor.
Com s’ha dit de forma explícita abans, una de les maneres més simples de definir la vida és com un conjunt organitzat de reaccions químiques integrades i encadenades, localitzades en un espai físic concret i heretables. Actualment, totes les cèl·lules són embolcallades per una membrana plasmàtica, que delimita l’espai físic concret on es desenvolupen les seves reaccions vitals, i contenen material genètic heretable. El principal component d’aquesta membrana plasmàtica són els fosfolípids, un tipus de lípids de natura amfifílica. Les amfifiles són molècules que tenen una part hidròfila i una part hidròfoba, per la qual cosa tendeixen a agrupar-se i a formar espontàniament vesícules: la part hidròfila s’orienta cap enfora i entra en contacte amb el medi aquós, i la part hidròfoba queda protegida de l’aigua.
La membrana dels bacteris, dels arqueus i dels eucaris és formada principalment –però no únicament– per una bicapa de fosfolípids, però cada domini presenta unes particularitats específiques, per la qual cosa es discuteix com era la membrana plasmàtica de LUCA. Cal pressuposar, però, que era formada per aquest tipus de molècules, les quals van anar augmentant de complexitat des de les protocèl·lules inicials fins a LUCA.
Tanmateix, aquest increment de complexitat en la bicapa de fosfolípids devia haver comportat també l’aparició i l’evolució de mecanismes que permetin el pas de substàncies a través de la membrana de la vesícula, atès que són poc permeables. En les cèl·lules actuals, un d’aquests mecanismes de transport es fa a través de porus estabilitzats amb proteïnes específiques. En aquest sentit, s’ha demostrat que la intercalació en la membrana d’un polipèptid format únicament per molècules d’alanina, un aminoàcid, incrementa la taxa de transport de protons i de substàncies protòniques (amb càrrega positiva) a través d’aquesta membrana.
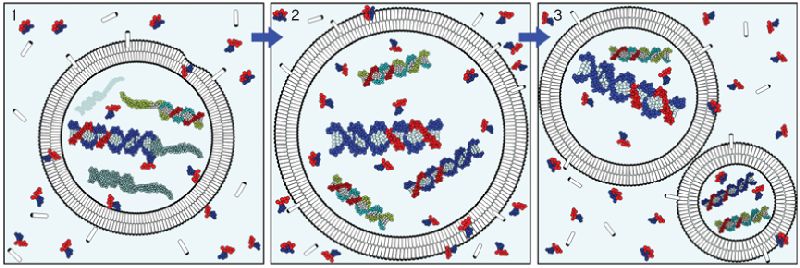
Model de replicació d’una protocèl·lula primitiva mitjançant mecanismes físics. En un ambient complex on hi ha lípids i nucleòtids es poden formar vesícules que continguin molècules d’RNA, juntament amb altres molècules orgàniques (1). Si entre les diverses molècules d’RNA hi ha ribozims, i si també hi ha alguna font d’energia externa que impulsi les reaccions, les molècules d’RNA es poden replicar, la qual cosa farà augmentar la pressió osmòtica a l’interior de la vesícula, que creixerà amb la incorporació de més amfifiles (2). Finalment, la forma de la vesícula es desestabilitzarà, per la qual cosa s’acabarà dividint i es formaran dues vesícules, que poden contenir molècules d’RNA (3).
A partir de Scrum i altres, 2010.
També és interessant destacar que aquestes vesícules poden interaccionar amb l’RNA i formar complexos liponucleics. I s’ha vist que, si es produeixen cicles de dessecament i rehidratació, aquestes vesícules acceleren la polimerització de nucleòtids en cadenes d’RNA, al mateix temps que aquests àcids nucleics contribueixen a estabilitzarles. A més, permeten l’existència d’un mecanisme senzill i directe de reproducció. Si dins una d’aquestes vesícules hi ha una molècula d’RNA i aquesta es replica, com es pressuposa que succeïa quan l’RNA era el material que emmagatzemava la informació genètica, augmenta la força iònica a l’interior de la protecèl·lula, atès que l’RNA té una lleugera càrrega positiva. Això fa que incrementi la pressió osmòtica i, en conseqüència, que s’incorporin més amfifiles a la membrana de la vesícula, per a compensar-la. La superfície membranosa creix fins que arriba a una mida crítica en la qual es distorsiona la seva forma esfèrica. Llavors, si entren en contacte dues parts diferents de la membrana, es poden fusionar i formar dues vesícules a partir de la vesícula ancestral, cadascuna amb algunes molècules d’RNA al seu interior. És l’equivalent a una reproducció cel·lular però a través de mecanismes únicament físics, que tanmateix generen un efecte en cascada en què es van generant cada cop més vesícules com aquestes, amb molècules de material genètic al seu interior.
Els ancestres de LUCA
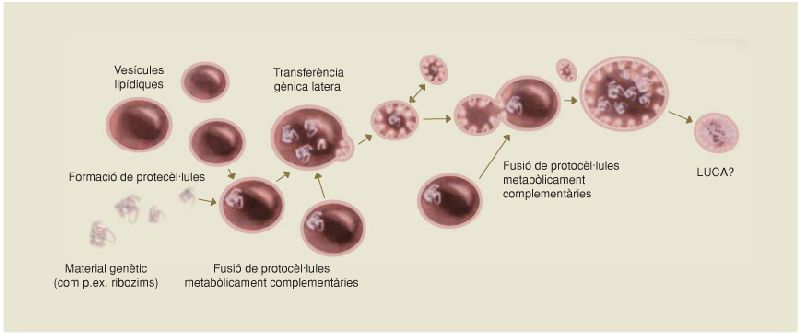
Possible origen de LUCA o d’algun ancestre del seu llinatge a partir de la fusió de diverses protocèl·lules metabòlicament complementàries i de fenòmens de transferència gènica lateral.
A partir de fonts diverses.
Totes les proves de què es disposa indiquen que tots els éssers vius de la Terra procedeixen d’un únic organisme ancestral, a partir del qual han anat divergint per processos evolutius. Això no implica, però, que LUCA fos el primer ésser viu, ni que després de LUCA no se’n formessin més de diferents, sinó que la selecció natural va afavorir únicament els descendents d’aquest. Tanmateix, hom es pot preguntar si LUCA prové d’un únic llinatge, per evolució primer abiòtica i després biològica, o bé si fou el resultat, directament o indirecta, en algun moment anterior del seu llinatge, de la fusió de diverses protocèl·lules. En aquest sentit, l’anàlisi d’un nombre creixent de gens i de genomes indica que els fenòmens de transferència gènica lateral, dels quals s’ha parlat breument, i fins i tot la fusió de genomes ancestrals sencers, foren relativament freqüents en l’origen de la vida, la qual cosa fa que moltes filogènies semblin reticulades a mesura que s’acosten a l’inici de la vida.
Aquest fet, juntament amb la discussió del punt anterior sobre la formació de les primeres vesícules i protocèl·lules a partir de membranes lipídiques amb cadenes d’àcids nucleics al seu interior, ha fet que s’hagi suggerit que LUCA, o bé una protocèl·lula anterior en el seu llinatge, s’originés a partir d’una població molt diversa de protocèl·lules metabòlicament complementàries, cadascuna de les quals amb el seu propi genoma i la seva pròpia activitat metabòlica, i entre les quals s’establien successos massius de transferència gènica lateral. És a dir, suggereixen l’existència d’una comunitat d’entitats genètiques distintes implicades totes elles en un procés de ràpida diferenciació, les quals es podrien trobar molt a prop de la base de l’arbre de la vida, i ser totes elles, d’alguna manera, antecessores de LUCA.
Sigui com sigui, en l’origen de la vida van confluir tots els elements moleculars necessaris perquè aquesta s’originés: material genètic –PNA o directament RNA, i posteriorment DNA–, activitat catalítica –ribozims i posteriorment proteïnes enzimàtiques– i vesícules membranoses en les quals es produïa una intensa i cada cop més complexa activitat metabòlica, que incrementava llur ordre intern.
L’arrel de l’arbre filogenètic de la vida permet determinar la direccionalitat dels canvis evolutius i, en conseqüència, quins dels caràcters que s’observen en l’actualitat en els tres dominis són primitius i quins són derivats. Si s’aplica aquest axioma als processos metabòlics, s’observa que el metabolisme més primitiu correspon a organismes termòfils extrems, la temperatura òptima de creixement dels quals és de 90 ºC o superior. Aquesta direccionalitat suggereix que LUCA era un organisme hipertermòfil, i en conseqüència que la vida es va originar en un ambient amb una temperatura elevada on hi havia les molècules necessàries per a la síntesi abiòtica de molècules orgàniques, com per exemple el que es troba actualment en les surgències hidrotermals oceàniques o en altres llocs en què les super-fícies minerals poden haver afavorit l’aparició de sistemes biològics quimiolitotròfics primitius, com s’ha discutit en diferents apartats d’aquestes pàgines.
Tanmateix, aquest és també un punt de debat, atès que no és fàcil reconciliar un ambient termòfil amb una primera etapa vital basada en l’RNA, com suggereixen moltes altres dades, atès que aquesta biomolècula és extremament fràgil a altes temperatures. En aquest sentit, també s’ha suggerit que potser la vida es va originar en ambients més temperats i que en conseqüència era mesòfila, però que ateses les condicions climàtiques del període arqueà ràpidament es va adaptar a ambients termòfils, i que per aquest motiu els primers organismes mesòfils van ser ràpidament desplaçats pels hipertermòfils –i aquesta és la direccionalitat que mostren els arbres filogenètics.
Novament, fos com fos LUCA, les dades de què es disposa indiquen que els organismes vius més propers a LUCA que ens permeten inferir les filogènies moleculars eren termòfils, de la mateixa manera que les dades genètiques indiquen que la primera molècula portadora d’informació genètica i al mateix temps amb capacitat enzimàtica fou l’RNA. I que totes les molècules orgàniques imprescindibles per a l’origen de la vida es van poder formar per síntesi química abiòtica i espontània en les condicions de la Terra primitiva, si bé és possible que algunes se sintetitzessin en les profunditats de l’espai i arribessin a la Terra amb meteorits i cometes. No cal que totes les molècules se sintetitzessin simultàniament, ni al mateix lloc. Només cal que en un moment determinat coincidissin les molècules mínimes imprescindibles per a iniciar la cursa vital, com RNA i vesícules d’amfifiles, les quals, gràcies a la seva activitat, van permetre que les protocèl·lules, i després les cèl·lules, anessin augmentant progressivament la seva complexitat, dins un procés evolutiu clarament darwinista –és a dir, per canvi a l’atzar i preadaptatiu i per selecció natural.
L’origen dels ecosistemes

Els estromatòlits són estructures en forma d’agregat formades per capes de microorganismes, principalment cianobacteris fotosintètics. Van constituir els primers ecosistemes, fa més de 3.500 Ma. Aquest tipus d’ecosistemes encara es troben en algunes zones d’Austràlia i del Carib.
F. Vanrenterghem.
Actualment, com probablement fou des de l’inici, els éssers vius no viuen aïllats els uns dels altres, sinó que formen comunitats complexes, els ecosistemes. Les primeres cèl·lules utilitzaven inicialment les molècules orgàniques del medi per a mantenir la seva estructura, obtenir energia i multiplicar-se. En augmentar la quantitat de cèl·lules, les molècules orgàniques disponibles es deurien haver esgotat, per la qual cosa finalment van sobreviure aquelles que, per mecanismes evolutius darwinistes, havien desenvolupat mecanismes propis de producció d’energia. Els primers mecanismes que aparegueren, a partir molt probablement d’organismes procariòtics anaeròbics termòfils, foren la fermentació i la fotosíntesi. La fermentació és un procés d’obtenció d’energia química a partir de la degradació de compostos orgànics d’alta energia, els quals es converteixen en compostos biogènics de més baixa energia que s’acumulen en el medi extracel·lular. La fotosíntesi, en canvi, és un procés d’obtenció d’energia a partir de la llum, la qual empren per a sintetitzar compostos orgànics d’alta energia a partir d’elements biogènics.

Els tapissos microbians són ecosistemes constituïts únicament per microorganismes que es disposen en capes horitzontals, d’un gruix que pot variar entre uns pocs mil·límetres i uns quants centímetres per sota de la superfície on es desenvolupen.
M. Berlanga i R. Guerrero.
En aquest context, els fòssils més antics es troben en els estromatòlits, unes estructures sedimentàries laminades constituïdes principalment per carbonat càlcic –també se n’han trobat de sílex– que es van formar per la precipitació biogènica d’aquests minerals. Els seus precursors foren comunitats constituïdes per organismes procariotes, algunes de les quals s’han conservat vives en diferents llocs de la Terra. Són els anomenats tapissos microbians. També s’han trobat alguns estromatòlits que segueixen actius, com a Shark Bay, a la costa nord-occidental d’Austràlia, i a les Bahames, a l’Atlàntic, on es pot observar el seu creixement, originat pel desenvolupament de la capa superior d’organismes, uns cianobacteris que presenten uns nivells de fotosíntesi i calcificació molt alts. Tanmateix, la seva datació és sovint controvertida. D’una banda, hi ha autors que afirmen haver identificat fòssils de bacteris en roques sedimentàries de fa 3.500 Ma, troballes molt discutides que han estat posteriorment reinterpretades com a intrusions minerals d’origen abiogènic. D’altra banda, també hi ha autors que han suggerit la presència d’inclusions carbonoses aparentment indicatives d’activitat biològica datades de fa 3.800 Ma. Sigui com sigui, el consens general és que fa 3.500 Ma la majoria de les principals vies bioquímiques que sustenten la biosfera moderna ja havia evolucionat.
Els tapissos microbians es troben principalment en zones costaneres amb entrada intermitent d’aigua marina. Als Països Catalans se’n poden trobar, per exemple, al delta de l’Ebre, que han esdevingut uns dels més estudiats a tot el món. Són formats per poblacions d’organismes procariotes disposats en capes horitzontals molt primes. La capa superior és constituïda principalment per cianobacteris fotosintètics aeròbics. Per sota d’ells acostuma a haver-hi bacteris fotosintètics anaeròbics, i per sota, bacteris reductors del sulfat. Probablement, els primers ecosistemes tenien una estructura similar, la qual afavoria l’intercanvi de substàncies i l’especialització. Una especialització que, gràcies a canvis sotmesos a l’atzar i preadaptatius, els quals inclouen mutacions en el material genètic, transferència gènica lateral, fenòmens d’endosimbiosi, duplicacions de gens i de genomes, etc., i a l’efecte de la selecció natural, han anat menant l’evolució dels organismes cap a totes les formes de vida actuals i les ja extingides, al mateix temps que han canviat la fesomia del planeta i la composició de l’atmosfera.
L’estudi de les possibles formes de vida en altres planetes
L’astrobiologia és l’estudi de l’origen, l’evolució, la distribució i el futur de la vida a l’Univers. Un dels seus objectius és la cerca de planetes amb ambients habitables, o que hagin estat habitables, malgrat que fos de forma intermitent, i analitza els planetes no només com són en l’actualitat sinó la seva història geològica, el procés dinàmic de la seva gènesi, atès que, com s’ha discutit abans, la gènesi de la Terra ha estat cabdal per al desenvolupament de la vida.
Només es coneix un sol lloc en tot l’Univers on s’hagi desenvolupat la vida, el planeta Terra, però que això no implica que no s’hagi pogut desenvolupar en alguns o en molts altres indrets. Per exemple, les observacions realitzades pel telescopi Kepler en els seus primers quatre mesos de funcionament van permetre detectar 1.235 planetes extrasolars, 54 dels quals orbiten a una distància de la seva estrella que és adequada per al desenvolupament i/o el manteniment de la vida. I es tenen dades que determinats compostos orgànics com les tolines, de les quals s’ha parlat amb relació a la síntesi abiòtica d’aminoàcids, són presents, per exemple, a l’atmosfera de Tità, un dels satèl·lits de Saturn. En aquest sentit, al sistema solar, per exemple, es considera que Mart i alguns satèl·lits gelats del sistema solar exterior, com Europa i Tità, reuneixen, o poden haver reunit, les condicions necessàries perquè s’hi hagi desenvolupat la vida, la qual cosa no implica que, necessàriament, s’hi hagi hagut de desenvolupar.
La cerca de vida a l’Univers es duu a terme a través de la identificació de biosignatures específiques. Una biosignatura és qualsevol substància, com per exemple un element, un isòtop o una molècula, o qualsevol fenomen físic o químic que proporcioni una evidència científica de l’existència, present o passada, de vida. Les biosignatures inclouen aquells minerals, gasos atmosfèrics i reserves de carboni, sulfur i qualsevol altre element que tinguin un impacte pel que fa a la utilització d’energia i a la producció de biomassa i de productes orgànics de rebuig. També es consideren biosignatures la presència de morfologies cel·lulars i extracel·lulars, de traces de metabolisme i la quiralitat de certs elements, com aminoàcids i glícids, entre d’altres, de tots els quals s’ha parlat quan s’ha discutit l’origen de la vida a la Terra. I també determinats canvis cíclics que poden afectar la superfície planetària, com per exemple la concentració de pigments complexos com clorofil·la en funció de l’estació climàtica. En general, les biosignatures representen una modificació de l’ambient observable a escala local o planetària.
Tanmateix, la troballa de biosignatures no constitueix, per si mateixa, la confirmació directa de la presència actual o en èpoques pretèrites de vida. Per exemple, l’estudi microscòpic del meteorit d’origen marcià ALH 84001 va permetre visualitzar unes estructures cristal·lines de magnetita que, com alguns autors van suggerir, podien ser rastres de formes cel·lulars de tipus bacterià, però posteriorment es va veure que també es poden produir de forma abiogènica en determinades condicions. I dues naus del projecte Viking de la NASA van recollir mostres de sòl marcià i les van sotmetre a diverses proves bioquímiques a la cerca de biosignatures, entre les quals hi havia traces d’activitat metabòlica, però els resultats no foren concloents.
Es pot especular molt sobre aquest tema, però el millor resum es troba en els set objectius primor-dials que el projecte d’astrobiologia de la NASA té plantejats: entendre la naturalesa i la distribució dels ambients habitables a l’Univers, dins i fora del sistema solar; determinar la presència de biosignatures i de química prebiòtica al sistema solar, inclosa la història dels ambients que presenten o han presentat aigua líquida, elements químics i fonts d’energia que poden haver originat i/o mantingut la vida; aprofundir en els mecanismes físics i químics a través dels quals la vida pot sorgir a partir de precursors planetaris i còsmics; entendre com la vida i l’ambient planetari han coevolucionat a la Terra a través del temps geològic, influint-se mútuament; entendre els mecanismes evolutius i els límits ambientals de la vida, inclosos els mecanismes moleculars, genètics i bioquímics que controlen i limiten l’evolució, la diversitat metabòlica i l’adaptació de la vida; analitzar els principis que modulen el futur de la vida, tant a la Terra com en qualsevol altre lloc de l’Univers, i determinar com reconèixer les biosignatures en altres planetes, tant a través de mostres de materials com a partir d’anàlisis remotes de la composició i dels canvis dinàmics de la seva atmosfera i de la superfície planetària, incloent-hi les de possibles tecnologies extraterrestres.