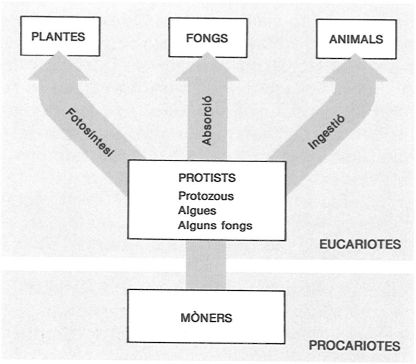
La classificació dels éssers vius en cinc regnes és un intent d’ordenar amb les tècniques modernes d’estudi els diferents grups d’organismes i imaginar-ne les possibles relacions. El concepte de mòners s’ha creat modernament per a reunir els bacteris, els cianòfits i els procloròfits.
Biopunt, original de L. Margulis i K.V. Schwartz (1985)
Històricament, hom ha agrupat els éssers vius en dos regnes, el regne animal i el regne vegetal, basant-se fonamentalment en diferències d’índole fisiològica, concretament en la capacitat (autotròfia) o incapacitat (heterotròfia) de sintetitzar matèria orgànica a partir de l’energia lluminosa i de matèria inorgànica. El grau d’organització en el conjunt dels animals i els vegetals és molt variat; hom pensava que ambdós regnes derivaven d’uns organismes més senzills, que reunien simultàniament caràcters d’índole animal i d’índole vegetal, ja que entre aquests hi ha grups heteròtrofs i grups autòtrofs. Hom considerava que aquests organismes més senzills eren formats en general per «una sola cèl·lula» —qüestió molt debatuda actualment— i que constituïen un tercer regne a part, el dels protists (Protista) o protoctists (Protoctista).
Posteriorment, en estudiar-se més a fons les característiques de la cèl·lula, es va considerar que aquest era el millor fonament per a separar els organismes. En efecte, es distingien dos grans tipus de cèl·lules, les procariotes i les eucariotes, i hom es va adonar que tant els anomenats protists com els animals i els vegetals tenen cèl·lules de tipus eucariota. La diferència fonamental entre cèl·lules procariotes i eucariotes. consisteix en el fet que les primeres no tenen membrana nuclear, per la qual cosa el material genètic no se situa al nucli, mentre que en la cèl·lula eucariota hi ha un nucli ben delimitat per una membrana, dins el qual es reuneix tot el material genètic cel·lular.
D’altra banda, tant en la seva estructura com en el seu comportament bioquímic, els procariotes i els eucariotes difereixen en d’altres aspectes. De fet, les diferències que mantenen són molt més importants que les que hi ha entre els protists, els animals i els vegetals, i actualment hom considera que els organismes procariotes constitueixen l’anomenat regne dels mòners, que reuneix els bacteris i les cianofícies o cianòfits. Els procariotes són considerats com els ancessors dels eucariotes, i és possible que l’evolució hagi tingut lloc per sèries d’associacions simbiòtiques dels ancessors procariotes, encara que, segons altres teories, l’eucariota ancestral hauria evolucionat a partir d’un ancessor procariota únic, que hom imagina de dimensions més grans que els actuals.
Pel que fa a la classificació dels eucariotes, a més dels regnes que ja hem esmentat, protist, animal i vegetal, cal afegir-hi un quart regne, el dels fongs, que abans hom havia inclòs dins el regne vegetal. Els fongs tenen paret cel·lular, com els vegetals (bé que no és feta de cel·lulosa), però són heteròtrofs, ja que no tenen plasts, com els animals. Actualment, doncs, es considera que els éssers vius poden ésser repartits en 5 regnes: els mòners (Monera), els protists (Protista), els animals (Animalia), els vegetals (Vegetalia) i els fongs (Fungi). I que l’evolució deu haver-se produït partint de procariotes, que haurien originat els protists (els eucariotes més senzills i formats en general per una sola cèl·lula) i, a partir d’aquests, per tres vies diferents, s’haurien originat els altres tres regnes (animals, vegetals i fongs), tots multicel·lulars, en els quals les diferents cèl·lules, que són especialitzades en funcions diferents, en molts casos s’organitzen fent teixits i òrgans.

Sota el concepte generalitzat d’animal s’inclouen formes d’organització molt diverses i de nivells de complexitat molt diferents, cadascun dels quals permet, però, la realització de les funcions vitals. En la fotografia, per exemple, veiem dos animals ben diferents, un opistobranqui (Flabellina affinis) i un celenterat (Eudendrium), compartint un mateix hàbitat, bé que explotant-hi recursos diferents i resolent diferentment el seu sistema d’intercanvi de gasos i substàncies amb el medi. Però no sempre és òbvia la qualitat d’animal: l’observació cada vegada més precisa de tots els organismes ha portat a replantejaments globals de conceptes abans tan indiscutibles com el d’animal i vegetal.
Xavier Safont i M. Alba Camprubí
No resulta senzill d’establir el concepte d’animal, especialment si es volen tenir en compte els protists amb tendències animals anomenats ordinàriament protozous. En el cas dels organismes multicel·lulars es pot fer una distinció clara entre animals i plantes, és a dir, entre metazous (Metazoa) i metàfits (Metaphyta). Les plantes contenen clorofil·la i són holotròfiques (autòtrofes), tenen les cèl·lules proveïdes d’una paret de natura cel·lulòsica i no tenen capacitat de moviment. Els animals, al contrari, són holozoics o sapròfits (heteròtrofs), ja que no tenen plasts, les seves cèl·lules manquen de paret cel·lular i mai no tenen cel·lulosa com a material constituent i, a més, són dotades de moviment.
Però es fa difícil d’aplicar aquestes diferències als organismes unicel·lulars, ja que, per exemple, hi ha molts flagel·lats proveïts de clorofil·la i estretament emparentats amb d’altres que no en tenen. D’altra banda, se sap que una sola mutació pot determinar la pèrdua de clorofil·la. La possessió de cel·lulosa tampoc no és un caràcter determinant en els organismes unicel·lulars, ja que hi ha grups, com els dinoflagel·lats, que tenen paret cel·lulòsica però en canvi no tenen clorofil·la, i, per tant, han d’ésser considerats heteròtrofs. Finalment, la capacitat de moviment no és un criteri vàlid ni tan sols per als organismes multicel·lulars (pensem en el cas de les esponges, que són animals immòbils); dins el conjunt dels «unicel·lulars», hi ha moltes algues que tenen fases mòbils dins el seu cicle vital i, paral·lelament, molts protozous sèssils que no tenen cap mitjà de locomoció. Això fa que sigui impossible d’establir clarament una definició d’animal que sigui vàlida tant per als multicel·lulars (metazous) com per als «unicel·lulars» (protozous).

Hom ha ordenat els diferents grups animals segons les seves afinitats en grans grups, anomenats fílums, la categoria superior de classificació dins el món animal. Els organismes pertanyents a un mateix fílum (per exemple, l’estrella Echinaster sepositus i l’eriçó Sphaerechinus granularis, en la fotografia) comparteixen un determinat nivell d’organització i unes determinades característiques comunes. Els avenços en el coneixement de les vies d’origen dels diferents grups animals (filogènia) han permès de veure que moltes vegades s’ha arribat a formes i nivells d’organització molt semblants a partir de punts d’origen diferents.
Agustí Montserrat
Si ens referim exclusivament als animals multicel·lulars o metazous, podem definir-los com a éssers heteròtrofs mancats de clorfil·la i de cel·lulosa, capaços d’efectuar moviments. A més, en general són diploides i en la seva reproducció, en general, es produeix anisogàmia (unió de dos gàmetes haploides diferents, un òvul i un espermatozoide) i el resultat de la fecundació és un zigot diploide que experimenta mitosis successives fins a donar lloc a l’anomenada blàstula. Tots els animals multicel·lulars es desenvolupen a partir d’una blàstula, i a partir d’aquesta blàstula s’origina la gàstrula. Els detalls de les fases posteriors del desenvolupament varien molt entre els diferents fílums d’animals i hom utilitza aquestes diferències per a establir les relacions que hi ha entre els diferents fílums.
L’organització unicel·lular i l’organització multicel·lular
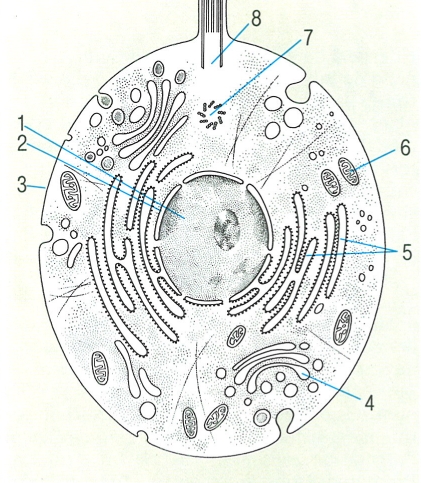
Esquema general d’una cèl·lula animal en el cas d’un organisme unicel·lular. S’hi mostren els seus elements morfològics principals: 1 nucli, 2 membrana nuclear, 3 membrana cel·lular, 4 aparell de Golgi, 5 ergastoplasma, 6 mitocondri, 7 centríol, 8 cos basal.
Biopunt, a partir de fonts diverses
Ja hem dit que els protists o protoctists deriven dels organismes procarionts (mòners) i també hem al·ludit a la dificultat de definir el concepte d’animal quan es tracta d’englobarhi també els protozous. Podem definir els protists com organismes de petites dimensions proveïts d’un complex nuclear dins un cos citoplasmàtic que generalment no està dividit en cèl·lules. En molts casos, la multiplicació per algun tipus de fissió produeix nous individus que se separen i tenen una vida independent, però de vegades aquesta separació és incompleta i llavors es formen colònies, constituïdes per individus que poden tenir diferències morfològiques i funcionals. Únicament en casos excepcionals la diferenciació entre els individus dona lloc a cèl·lules diferents al llarg del desenvolupament, com per exemple la formació d’espores en el cicle de vida dels esporozous. Tanmateix, la immensa majoria dels protists poden definir-se com acel·lulars en el sentit que equivalen a un metazou complet, és a dir, que una sola cèl·lula fa totes les funcions; en els metazous, en canvi, les cèl·lules que formen els diferents teixits i òrgans són especialitzades en funcions concretes i han perdut la totipotència, excepte en el cas de les cèl·lules sexuals (òvuls i espermatozoides), que originaran, prèvia fecundació, un nou organisme.
Els protists reuneixen diverses línies evolutives. Alguns són heteròtrofs i mostren tendències clarament animals. Altres són autòtrofs i constitueixen clarament línies evolutives relacionades amb els vegetals. Altres línies reuneixen simultàniament formes heteròtrofes i autòtrofes.
Els organismes denominats protists han estat classificats com a protozous, com a algues i com a fongs inferiors, i inclouen grups que són sempre «unicel·lulars» i d’altres que contenen formes «unicel·lulars o multicel·lulars». Pel que sembla, a partir dels protists, per via fotosintètica (autòtrofa) s’originen les plantes; per via ingestiva (hetròtrofa) s’originen els animals, i per via d’absorció (saprofítica), els fongs.
La definició original de protozous (protists de l’estirp animal) conté el terme «unicel·lular», i ací hem fet referència al terme «acel·lular». Es tracta de dos conceptes diferents, però que modernament es relacionen. Per a alguns autors, el terme «cèl·lula» es fa servir en diversos sentits i, per tal d’evitar-ho, Dobell va definir una «cèl·lula» com «una massa de protoplasma que forma part d’un organisme», anticipant que els protozous i d’altres protists són veritables organismes però no cel·lulars, amb organització diferent dels organismes multicel·lulars (metazous, metàfits i fongs). L’elecció entre tants termes implica també una opció diferent sobre l’origen dels metazous. Actualment hi ha altres definicions de «cèl·lula», com la que va donar Baker, el qual considera la cèl·lula com una massa de protoplasma àmpliament o completament envoltada d’una membrana i que conté un sol nucli, format per la transformació telofàsica d’una sèrie haploide o diploide de cromosomes anafàsics. Encara que alguns protozous s’ajusten a aquesta definició, n’hi ha molts d’altres que no s’hi ajusten, com per exemple els radiolaris, que són poliploides (el seu «nucli» és constituït per un gran nombre de nuclis), mentre que d’altres, com Opalina, són clarament multinucleats. En definitiva, Baker va arribar a suggerir que alguns protozous no són cèl·lules, i per això una definició d’aquest grup no ha de contenir el terme «unicel·lular». Baker defineix els protozous com organismes que són mòbils o holozoics o totes dues coses alhora, en els quals hom no arriba a distingir les diferents classes de cèl·lules somàtiques que són típiques dels metazous. Tot i que actualment els protozous s’inclouen dins el regne dels protists, en aquesta obra, com en la majoria dels tractats de zoologia clàssics, els considerarem com uns «animals molt especials», que es troben a la base dels metazous.
Els tipus d’organització en els animals multicel·lulars
La varietat dels organismes que viuen actualment és molt elevada. És difícil d’evaluar el nombre exacte d’espècies, però es pot considerar molt superior als dos milions (nombre hipotètic).
La visió del conjunt de les espècies existents en l’actualitat ens dona idea de la diversitat que presenta el món dels animals, però si fem un estudi més detallat de les diferents espècies ens adonem que hi ha una sèrie més aviat reduïda de formes tipus i tota una llarga sèrie de variacions d’aquestes formes que justifiquen la distinció de tantes espècies. D’altra banda, la diversitat de formes i la unitat de tipus estructurals fonamentals únicament poden explicar-se per un procés d’evolució. Els éssers vius han evolucionat a partir d’uns tipus comuns, a la recerca d’una adaptació més bona a l’ambient, que implica un canvi de l’organització però que manté les característiques fonamentals.
L’anàlisi de l’organització dels organismes ens permet de comprovar que hi ha un nombre reduït de tipus d’organització i que tots els animals que en presenten una mateixa mena constitueixen un tipus taxonòmic o morfològic (tronc o fílum), tot i que en el conjunt poden presentar canvis més o menys profunds respecte de l’esquema d’estructura o arquetip. En general, els tipus d’organització poden interpretar-se com el resultat d’un dinamisme morfològic, que modifica les estructures d’acord amb les funcions en primer lloc, i com a resultat de les adaptacions als esdeveniments que han succeït al llarg de l’evolució, en segon lloc. Per a comparar els diferents caràcters entre els tipus d’organització cal tenir en compte dos conceptes fonamentals: el d’analogia i el d’homologia. Òrgans anàlegs són els que fan la mateixa funció però tenen un origen i una estructura diferents (per exemple, l’ala d’un ocell i la d’un insecte). Són òrgans homòlegs els que tenen un mateix origen però poden desenvolupar funcions diferents (per exemple, el membre anterior dels vertebrats: aleta, pota, ala, braç). Els aspectes biològics que es consideren bàsics per a establir els tipus d’organització són els següents: simetria, nivells d’organització, desenvolupament embrionari i cavitats del cos.
Simetria
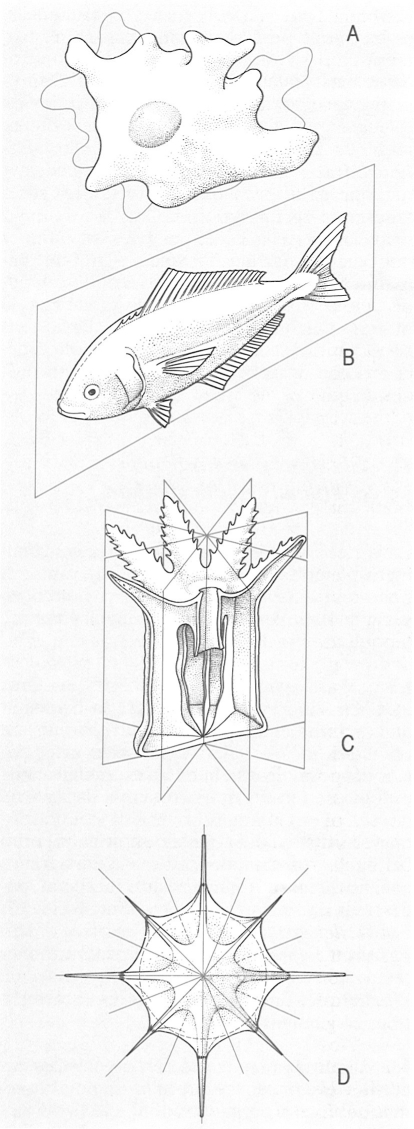
Tipus principals de simetria que hom pot trobar en els animals. A Cas especial de manca de simetria, o asimetria (ameba), B simetria bilateral (peix), C simetria radial (hidropòlip), D simetria esfèrica (heliozou).
Biopunt, a partir de fonts diverses
El criteri de simetria es pot considerar bàsic per a definir un tipus d’organització. La simetria és la divisió en parts iguals, per línies o plans, del cos dels animals. Hi ha tres grans models de simetria: esfèrica, radial i bilateral; tots tres es presenten en els protozous, mentre que en els metazous únicament trobem simetria radial i bilateral. D’altra banda, l’absència de simetria, és a dir, l’asimetria, és excepcional en el món animal i solament es troba en alguns protozous (Amoeba, per exemple), bé que sempre secundàriament pot aparèixer en d’altres grups (per exemple, els llenguados, les larves dels quals tenen una simetria clarament bilateral). En la simetria esfèrica, l’organisme és simètric respecte d’un punt. Es presenta en alguns protozous de cos totalment esfèric. La simetria és radial quan l’organisme és simètric al voltant d’un eix. Presenten aquest tipus de simetria els cnidaris i els ctenòfors, i, secundàriament, els equinoderms (les larves dels quals presenten simetria bilateral). En la simetria bilateral, l’organisme és simètric respecte d’un pla que divideix el cos en dues meitats. Aquest tipus de simetria és el més estès en el regne animal. D’altra banda, la simetria del cos es relaciona en gran part amb el tipus de vida de l’organisme. Per exemple, presenten simetria radial fonamentalment els fílums que són sèssils o planctònics, mentre que els que mostren simetria bilateral es mouen en un sentit determinant i tenen els òrgans dels sentits concentrats a la part anterior del cos; aquest procés porta a la cefalització i representa l’aparició de l’eix anteroposterior del cos. Segons la simetria, Katschel (1888-91) va dividir els metazous en dos supergrups: els radiats, que comprenien els cnidaris i els ctenòfors, i els bilaterals, que reunien tota la resta de metazous.
Nivells d’organització
Els nivells d’organització comprenen l’atòmic, el molecular, el cel·lular, el tissular, etc. Si deixem de banda els nivells inferiors, inclòs el procariota, en els animals es distingeixen diversos nivells d’organització. En el nivell cel·lular es distingeixen en primer lloc els organismes «unicel·lulars», o «acel·lulars», als quals ja ens hem referit, que són representats pels protozous, i els organismes multicel·lulars, representats pels metazous. En els metazous, segons el nombre de fulls embrionaris, distingim entre els organismes diploblàstics i els triploblàstics. Els diploblàstics presenten únicament dos fulls embrionaris (ectoderma i endoderma), que al seu torn es poden disposar com un agregat cel·lular, com passa, per exemple, a les esponges, els placozous i els mesozous, o en teixits, com és el cas dels cnidaris i els ctenòfors. Els triploblàstics, que reuneixen tota la resta dels metazous, presenten tres fulls embrionaris (ectoderma, endoderma i mesoderma) i presenten les cèl·lules organitzades en teixits, i aquests organitzats fent òrgans.
El desenvolupament embrionari i les cavitats del cos
Un altre criteri que s’utilitza per a definir els tipus d’organització dels animals és el desenvolupament embrionari i la formació de les cavitats internes del cos, tant les digestives com les celomàtiques. L’estudi comparat del desenvolupament embrionari és fonamental per a establir les relacions filogenètiques que hi ha entre els diferents grups. Des de la formació del zigot per mitjà de la fecundació fins a la formació completa de l’animal es defineixen tres etapes fonamentals: la segmentació (sèries de divisions cel·lulars de l’ou, que fan augmentar el nombre de cèl·lules); la gastrulació, que consisteix en un canvi breu i sobtat de la forma del desenvolupament de l’embrió no associat a la divisió cel·lular i que comporta la formació i la diferenciació de les dues fulles embrionàries primitives (l’ectoderma i l’endoderma); i una tercera etapa que és en realitat un complex d’un gran nombre de processos independents, i consisteix en la formació de la tercera capa embrionària (el mesoderma) i en l’organogènesi.
La segmentació
La segmentació consisteix en una sèrie de divisions cel·lulars de l’òvul fecundat, que tenen lloc abans de la gastrulació i que es relacionen amb la morfologia de l’ou (és a dir, el zigot més el vitel) i, en particular, amb la quantitat de vitel que té l’ou. De l’ou es forma la mòrula i, a partir d’aquesta, la blàstula. Quan l’ou té poc o gens de vitel, a cada divisió participa tot el zigot; aquest tipus de segmentació rep el nom d’holoblàstica. Si l’ou conté molt de vitel, únicament es divideix el pol animal i es forma un petit casquet de cèl·lules sobre el vitel; aquesta segmentació és la meroblàstica. Entre aquests dos tipus de segmentació n’hi ha molts d’altres, segons la quantitat de vitel de l’ou i la seva disposició.
El tipus de segmentació es troba en una relació molt estreta amb la morfologia de l’ou, i particularment amb la quantitat de vitel que aquest té, cosa que, al seu torn, depèn de les característiques de l’òvul (l’espermatozoide no conté substàncies de reserva). L’òvul és una cèl·lula que conté les substàncies nutritives destinades a formar i alimentar l’embrió. En casos excepcionals solament presenta la membrana plasmàtica, però generalment es troba envoltat de diverses membranes de protecció, que es formen durant l’ovogènesi, que són de naturalesa mucosa. L’òvul, i després el zigot, presenta una polaritat determinada, de manera que s’hi distingeix el pol germinatiu o pol animal, on se situa el nucli, i on es fa tota l’activitat metabòlica, i el pol vegetatiu, que és la zona on s’acumulen les substàncies de reserva o vitel. Al llarg d’un eix imaginari que aniria del pol germinatiu fins al pol vegetatiu hi ha un gradient metabòlic decreixent, dependent de la quantitat de vitel, que influeix notablement en la segmentació.
La quantitat de materials de reserva de l’ou i la seva localització permeten de classificar els ous en quatre categories, que coincideixen totalment amb els tipus d’òvuls: isolècits (o oligolècits), heterolècits, telolècits i centrolècits. Els ous isolecítics o oligolècits contenen poc vitel, i aquest és en forma de fines granulacions distribuïdes d’una manera uniforme per tot el citoplasma; pertanyen a aquesta categoria els ous de les esponges, els cnidaris, els equinoderms, els cefalocordats i els mamífers. En el cas concret dels mamífers placentats, de vegades hom parla d’òvuls alecítics o alècits, perquè són pràcticament desproveïts de vitel, ja que, gràcies al procés de desenvolupament embrionari facilitat per la placenta, no té utilitat. Els ous heterolecítics o heterolècits tenen una quantitat més abundant de vitel, que fa granulacions però d’una manera heterogènia, i es disposa desigualment: les granulacions més grosses s’acumulen a la zona més pròxima al pol vegetatiu, mentre que les més petites se situen al pol animal o germinatiu, prop del nucli. Aquests ous són característics dels anèl·lids, els mol·luscs (excepte els cefalòpodes), els amfibis i els teleostis. En el cas dels ous telolecítics o telocècits, el vitel és molt abundant i es reuneix en una massa que relega el citoplasma i el nucli al pol germinatiu, amb la qual cosa s’origina una diferenciació del disc germinatiu o cicatrula; els òvuls són voluminosos, com els ous, i són característics dels cefalòpodes, els selacis, els rèptils, els ocells i els mamífers prototeris. En els ous centrolecítics o centrolècits, el vitel és molt abundant i forma una massa central envoltada pel citoplasma, que s’estén per tota la perifèria, on també se situa el nucli; aquests ous són característics dels artròpodes. Així, doncs, els ous isolecítics i heterolecítics tenen segmentació total o holoblàstica, mentre que els ous de tipus telolecític o centrolecític la presenten de tipus parcial o meroblàstica.
D’altra banda, la segmentació total pot ésser igual o desigual. En el primer cas, tots els blastòmers són de les mateixes dimensions, cosa que és característica d’alguns ous isolecítics. Quan és desigual, que és el cas més estès (en la majoria dels ous isolecítics i en tots els heterolecítics), es distingeixen dos tipus de blastòmers: els anomenats micròmers, més petits, que se situen a la zona del pol animal, i els anomenats macròmers, més grossos, que se situen a la zona del pol vegetatiu. Quan la segmentació és parcial (meroblàstica), no afecta el pol vegetatiu, i la major part del vitel resta sense segmentar. Aquest tipus de segmentació presenta dues modalitats: segmentació parcial discoïdal, característica dels ous telolecítics, i parcial superficial, característica dels ous centrolecítics. En la segmentació parcial discoïdal, la divisió solament afecta un disc citoplasmàtic pròxim al pol animal —la part inferior (pol vegetatiu) no se segmenta. Seguidament es forma un disc de blastòmers, el blastoderma o blastodisc, a partir del qual es formarà l’embrió, que reposarà sobre la massa vitel·lina. L’ou alecític dels mamífers placentats, com que perd el vitel secundàriament, inicia la seva segmentació d’una manera holoblàstica, i prossegueix de manera meroblàstica. En la segmentació parcial superficial, típica dels ous centrolecítics, el nucli es divideix nombroses vegades, sense que apareguin límits cel·lulars definits dins la massa vitel·lina; després, els nuclis arriben al citoplasma perifèric i es disposen formant-hi una capa sincítica; finalment, sorgeixen els límits cel·lulars, que delimiten un blastoderma perifèric al voltant del vitel central no segmentat.
Una altra classificació de la segmentació es fonamenta en el destí de les cèl·lules filles o blastòmers. És indeterminada quan cada blastòmer, si se separa experimentalment, dona origen a un individu complet (principalment es troba en el deuterostomats), i és determinada quan, en separar un blastòmer experimentalment, solament origina una part determinada de l’animal, la mateixa que hauria originat en el cas de no haver estat separat (principalment es troba en els protostomats).
Una altra diferència en el tipus de segmentació consisteix en el pla segons el qual es fan les divisions dels blastòmers, i, per tant, en la disposició que aquests adopten. Els dos tipus principals que es defineixen segons aquest criteri són la segmentació radial i la segmentació espiral, i solament queda ben manifest de quin tipus es tracta en el cas de la segmentació holoblàstica (ja hem dit que quan la segmentació és meroblàstica no afecta tot l’ou). La segmentació radial és indeterminada, i té lloc principalment en els deuterostomats (per exemple els equinoderms, els hemicordats i els cordats). Durant la segmentació s’estableix una alternança regular de solcs de divisió meridians i longitudinals; els primers delimiten regions de blastòmers verticals, mentre que els segons porten a la formació de blastòmers disposats en files horitzontals. A causa que els plans perpendiculars passen per l’eix de polaritat de la blàstula, la simetria d’aquesta és radial respecte d’aquest eix. Un blastòmer determinat se situa per sota del blastòmer superior i per sobre del blastòmer inferior. La segmentació espiral és determinada i molt característica dels protostomats (per exemple els platihelmints, la majoria dels mol·luscs i els anèl·lids). Els plans de segmentació no són ni horitzontals ni verticals, sinó que són oblics en relació amb l’eix de l’ou. Els blastòmers fills es col·loquen entre els blastòmers pares i queden disposats en capes horitzontals, amb els blastòmers en posició alternada, per la qual cosa es disposen en espiral, i qualsevol blastòmer se situa entre dos blastòmers inferiors o superiors. Per anar del pol animal al pol vegetatiu cal descriure una espiral, l’eix de la qual és el de l’ou. D’altra banda, es poden localitzar fàcilment els elements derivats d’un blastòmer determinat.
Resultats de la segmentació
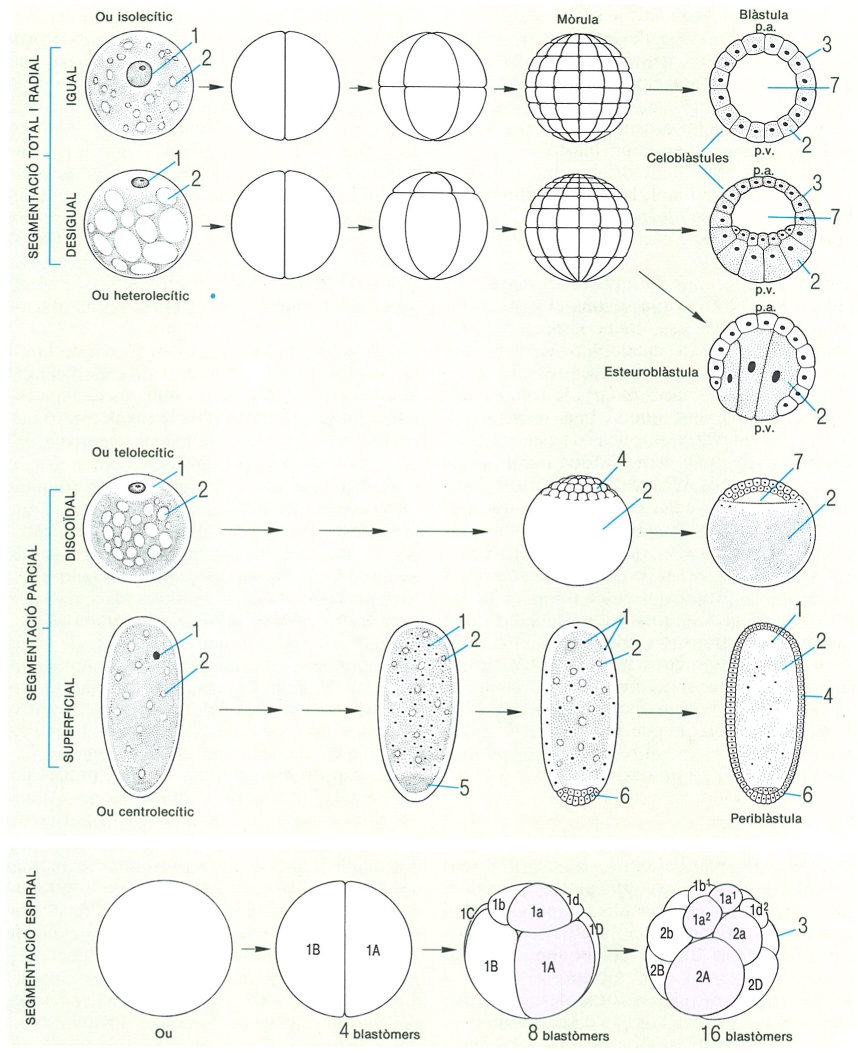
Tipus de segmentació de l’ou segons la quantitat de vitel, i formació de la mòrula i la blástula en cada cas, en els animals pluricel·lulars. 1 nucli, 2 vitel, 3 blastòmer, 4 blastoderma, 5 plasma polar, 6 cèl·lules germinals, 7 blastocel; p.a. pol animal, p.v. pol vegetatiu.
Biopunt, a partir de fonts diverses
Durant les primeres fases de la segmentació, els blastòmers es mantenen units, i això fa que l’ou adopti un aspecte semblant al d’una móra; per això, rep el nom de mòrula. No obstant això, aviat els blastòmers tendeixen a col·locar-se al voltant d’una cavitat central, l’anomenada cavitat de segmentació o blastocel, i així s’arriba a l’estadi de desenvolupament anomenat blàstula. Hi ha diversos tipus de blàstules, segons el contingut i la distribució del vitel de l’ou, que al seu torn es relaciona amb el tipus de segmentació. Quan els ous tenen poc vitel, la segmentació total igual o desigual porta a una blàstula amb el blastocel envoltat per una capa regular de cèl·lules; aquest tipus de blàstula rep el nom de celoblàstula, i en principi té tots els blastòmers de la mateixa mida; ara bé, quan els ous són heterolecítics, el blastocel es desplaça cap al pol animal, perquè els blastòmers del pol vegetatiu, més rics de vitel, són més grossos, i per això la celoblàstula és irregular. De vegades el blastocel és virtual, ja que és ocupat pels voluminosos blastòmers del pol vegetatiu; aquest tipus de blàstula, que rep el nom d’estereoblàstula, presenta un casquet de micròmers col·locats per damunt de la massa de macròmers. El resultat de la segmentació parcial discoïdal correspon a la discoblàstula dels ous telolecítics dels peixos i els rèptils. En el pol germinatiu de l’ou es forma un casquet de blastòmers que cobreix el vitel no segmentat. Finalment, el resultat de la segmentació parcial superficial dels ous centrolecítics dels artròpodes condueix a la periblàstula, formada per un blastocel virtual i els blastòmers que envolten la massa de vitel no segmentat.
La gastrulació
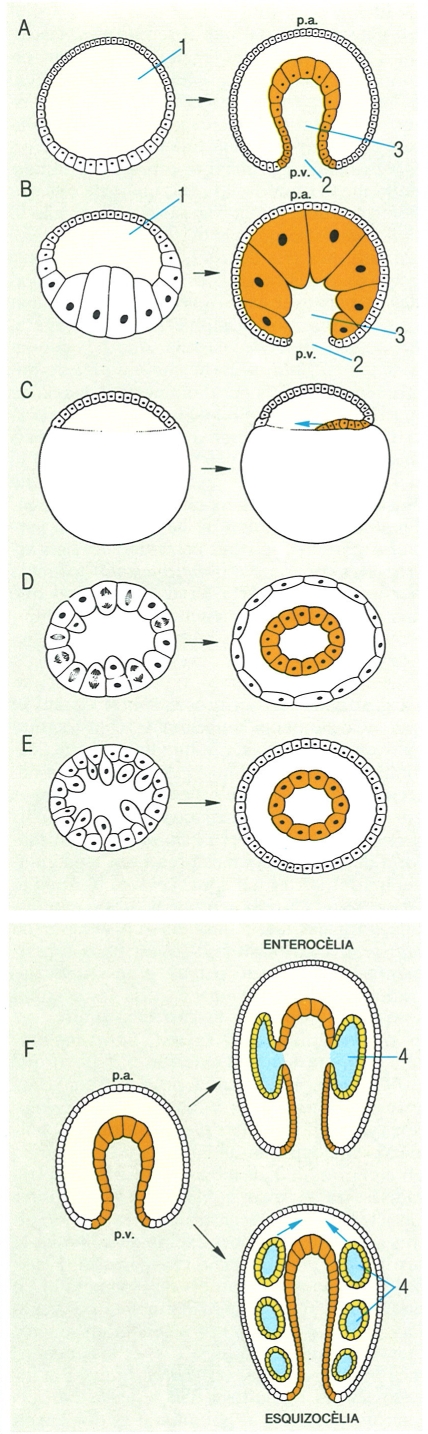
Tipus de gastrulació en el desenvolupament embrionari. En aquest procés, a partir dels diferents blastòmers que componen la blàstula, es diferencien l’ectoderma (blanc) i l’endoderma (taronja) i, més tard, es forma el mesoderma (groc), A Gastrulació per invaginació, B gastrulació per epibòlia, C gastrulació per involució, D gastrulació per delaminació, E gastrulació per ingressió, F formació de la tercera fuila embrionària, el mesoderma. 1 Blastocel, 2 blastòpor, 3 arquènteron, 4 celoma; p.a. pol animal; p.v. pol vegetatiu.
Biopunt, a partir de Barnes, Marshall, Beaumont, Cassier, Huillon
La gastrulació és un procés sobtat durant el qual els elements embrionaris (blastòmers) canvien de posició sense que hi hagi pràcticament cap divisió cel·lular. Aquest procés determina l’alineació dels elements embrionaris en dues capes, anomenades capes germinals primàries: l’ectoderma i l’endoderma. La majoria dels fílums animals presenten segmentació holoblàstica i, en aquest cas, la blàstula té l’aspecte d’una bola buida per dins (celoblàstula). La cavitat delimitada rep el nom de blastocel. Una blàstula d’aquest tipus experimenta la gastrulació per un procés d’invaginació, és a dir, que una part de les cèl·lules s’invagina i s’origina la gàstrula, alhora que es delimita una nova cavitat, l’arquènteron, que comunica amb l’exterior per un orifici o blastòpor. La gàstrula presenta una capa externa, l’ectoderma, i una d’interna, l’endoderma. Alguns animals presenten una organització semblant a la de gàstrula (els cnidaris i els ctenòfors) i d’altra banda mostren simetria radial. Ara bé, quan la blàstula presenta altres característiques, la gastrulació es fa per altres processos, com per exemple l’epibòlia, la involució, la delaminació i la ingressió. L’epibòlia es presenta quan els ous tenen vitel en una quantitat moderada i situat al pol vegetatiu, i la segmentació produeix grans macròmers vitel·lins. En aquest cas, el blastocel és més o menys virtual (estereoblàstula), i la invaginació és dificultosa. Per aquesta raó, els micròmers situats al pol animal es divideixen i es multipliquen, s’enfonsen i emboliquen els macròmers, fins a formar-se el blastòpor en el pol vegatitu. Si la quantitat de vitel és tan gran que fa que la segmentació sigui de tipus meroblàstic, es forma un petit casquet de cèl·lules o discoblàstula sobre el vitel, i llavors la gastrulació generalment té lloc per involució, un procés pel qual les cèl·lules de la perifèria del disc s’enfonsen i es dirigeixen cap enrere, sota la capa superficial, i formen un estrat doble. És el cas típic dels cefalòpodes. Hi ha altres formes de gastrulació, que no són tan freqüents. La delaminació passa quan en lloc d’una blàstula buida se’n forma una de compacta del tipus estereoblàstula; en aquest cas, la gastrulació es completa amb la separació de dues capes de cèl·lules, una d’externa i una d’interna, i no hi ha blastòpor sinó que la cavitat de l’arquènteron s’obrirà per un procés secundari. La gastrulació per ingressió es dona quan en lloc d’ésser empeses cap al blastocel sèries regulars de cèl·lules per a formar el segon full embrionari (com passa en la invaginació), el blastocel només és envaït per cèl·lules que posteriorment originen l’endoderma (en aquest cas, no es forma el blastòpor); aquest procés es presenta en els cnidaris, els ctenòfors i les esponges. Després de la gastrulació, excepte en el cas dels grups diploblàstics, té lloc la formació del mesoderma, i l’embrió va adquirint l’aspecte i la forma característica del grup al qual pertany.
La formació del mesoderma: evolució i significat del celoma
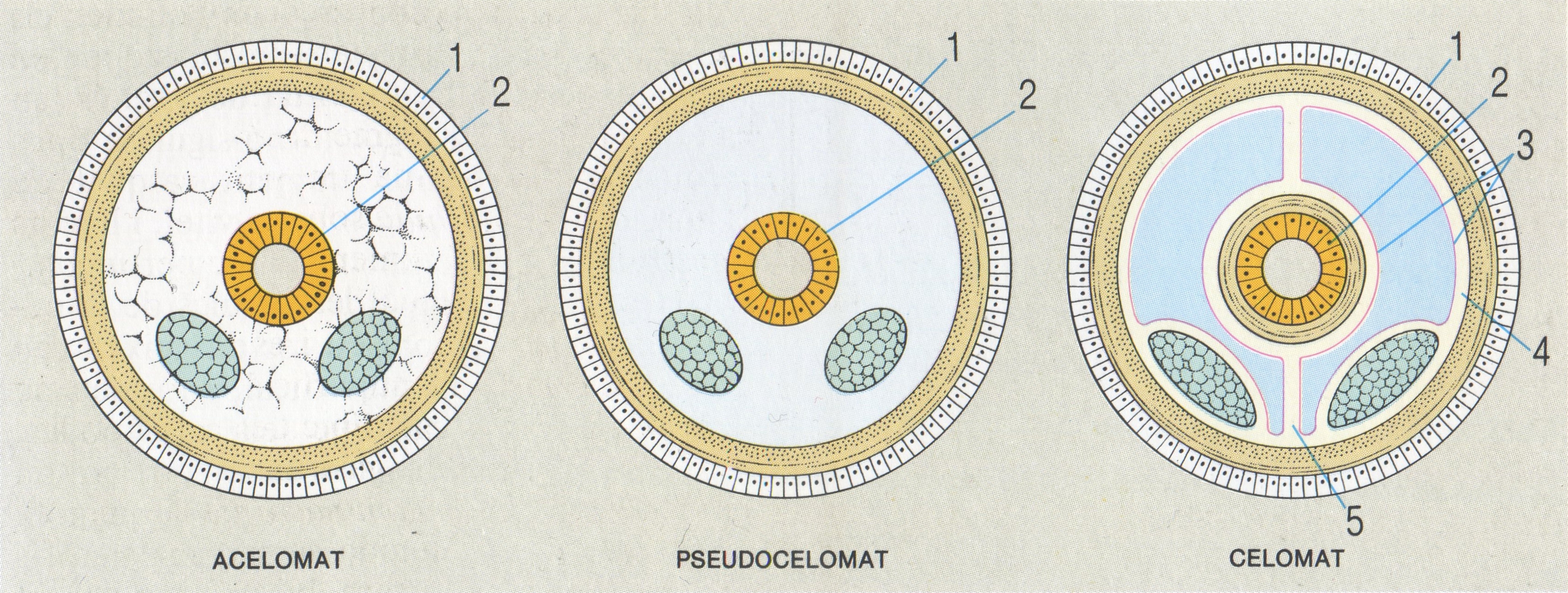
Esquema general de la secció transversal del cos en els organismes acelomats, pseudocelomats i celomats. 1 Ectoderma, 2 endoderma, 3 mesoderma, 4 peritoneu, 5 mesenteri peritoneal.
Biopunt, a partir de fonts diverses
A partir de la gàstrula s’origina una tercera capa embrionària, el mesoderma, i l’aparició del mesoderma origina al seu torn la formació d’una nova cavitat somàtica, el celoma. L’evolució d’aquesta cavitat sembla que ha tingut una importància fonamental en l’evolució dels metazous, en els quals ha permès augmentar les dimensions i la complexitat del cos. El celoma es defineix com una cavitat mesodèrmica plena de líquid, que es desenvolupa envoltada pel mesoderma embrionari. En alguns grups, en canvi, la cavitat interna del cos és representada per restes del blastocel, que persisteixen i reben el nom de pseudoceloma o fals celoma; es tracta, doncs, d’un simple espai entre el tracte digestiu i la paret del cos, que no queda delimitat pel mesoderma. En d’altres casos, no hi ha una cavitat general interna, ja que el cos és compacte. Finalment, el celoma es pot originar fonamentalment per dos processos: l’esquisocèlia i l’enterocèlia. En l’esquisocèlia el mesoderma deriva de grups de cèl·lules que se situen en el límit entre l’ectoderma i l’endoderma, als voltants del blastòpor; aquests grups de cèl·lules reben el nom d’esbossos mesodèrmics. L’esquisocèlia és característica dels protostomats. En l’enterocèlia el celoma es forma a costa de l’arquènteron, per mitjà d’una evaginació de les seves parets. És característica dels deuterostomats.
Entre els metazous triploblàstics, que sempre presenten simetria bilateral, es distingeixen tres tipus d’organització, segons el grau de desenvolupament i la natura de les seves cavitats internes: acelomats, pseudocelomats i celomats. Els acelomats presenten el cos compacte i sense cavitats. L’espai comprès entre el tub digestiu i la paret del cos és completament reblert de mesènquima i de fibres musculars, de manera que no hi ha cap espai somàtic. Són acelomats els platihelmints i els nemertins, típicament. En els metazous pseudocelomats, la cavitat somàtica és un fals celoma, ja que correspon a restes del blastocel primitiu i no és revestida d’una capa contínua de mesoderma; són pseudocelomats els rotífers, els nematodes, els acantocèfals, entre d’altres. En els celomats, la cavitat general del cos és un celoma veritable; són celomats els anèl·lids, els artròpodes, els equinoderms, els hemicordats, els cordats, entre d’altres. A més, dins dels celomats horn distingeix encara entre els esquizocelomats i els enterocelomats. En els celomats parlem d’una cavitat general, perquè no comunica en principi amb l’exterior, a diferència del que passa en el cas de l’arquènteron, que hi comunica per un blastòpor; de tota manera, indirectament hi pot comunicar pels poms celomàtics o celomípors. El líquid celomàtic, que omple la cavitat, pot tenir diverses funcions: tròfica, mecànica (esquelet hidrostàtic), excretora (eliminació de productes residuals a través dels celomípors, en aquest cas, nefròpors) i en alguns casos gonocèlica (les gònades es poden situar a les parets de la cavitat celomàtica i els gàmetes ésser expulsats a l’exterior pels gonòpors).
Una de les modificacions més importants que experimenta el celoma és la metamerització. En aquest cas, el celoma roman (almenys durant les fases embrionàries) però resta dividit en una sèrie de cavitats per mitjà d’uns envans. La metamerització del celoma comporta la metamerització del cos i el fet que estigui format per una sèrie d’unitats, els metàmers, que es van repetint al llarg del cos. A cada metàmer es repeteixen, a més de la cavitat celomàtica, els derivats mesodèrmics (nefridis, músculs, per exemple, o les gònades) i les formacions metamèriques (ganglis, apèndixs, etc.). D’altra banda, la segmentació pot ésser homònoma, la més primitiva, en la qual tots els metàmers són equivalents (per exemple, en la majoria dels anèl·lids), o heterònoma, en la qual els diferents metàmers del cos s’agrupen en regions, i mostren característiques peculiars segons la zona del cos on se situen (per exemple, en els artròpodes). En principi, la metamerització és interna i externa, però secundàriament pot romandre únicament la interna, com passa en els vertebrats. D’altra banda, alguns animals tenen pseudometameria o falsa metameria, que també pot ésser interna o externa; malgrat que l’aspecte extern en aquest últim cas és semblant al que dona la metameria, no deriva de les divisions del celoma, ja que només es repeteixen uns òrgans o formacions determinats, com passa en el cas dels cestodes. Finalment, alguns grups d’invertebrats, com els hirudinis, que són clarament metameritzats, experimenten el que s’anomena segmentació secundària, en la qual el nombre de segments aparents del cos no es correspon al nombre real, ja que cadascun és subdividit en d’altres; els òrgans interns metamèrics, però, no experimenten aquesta subdivisió.
La formació del tub digestiu i la seva evolució
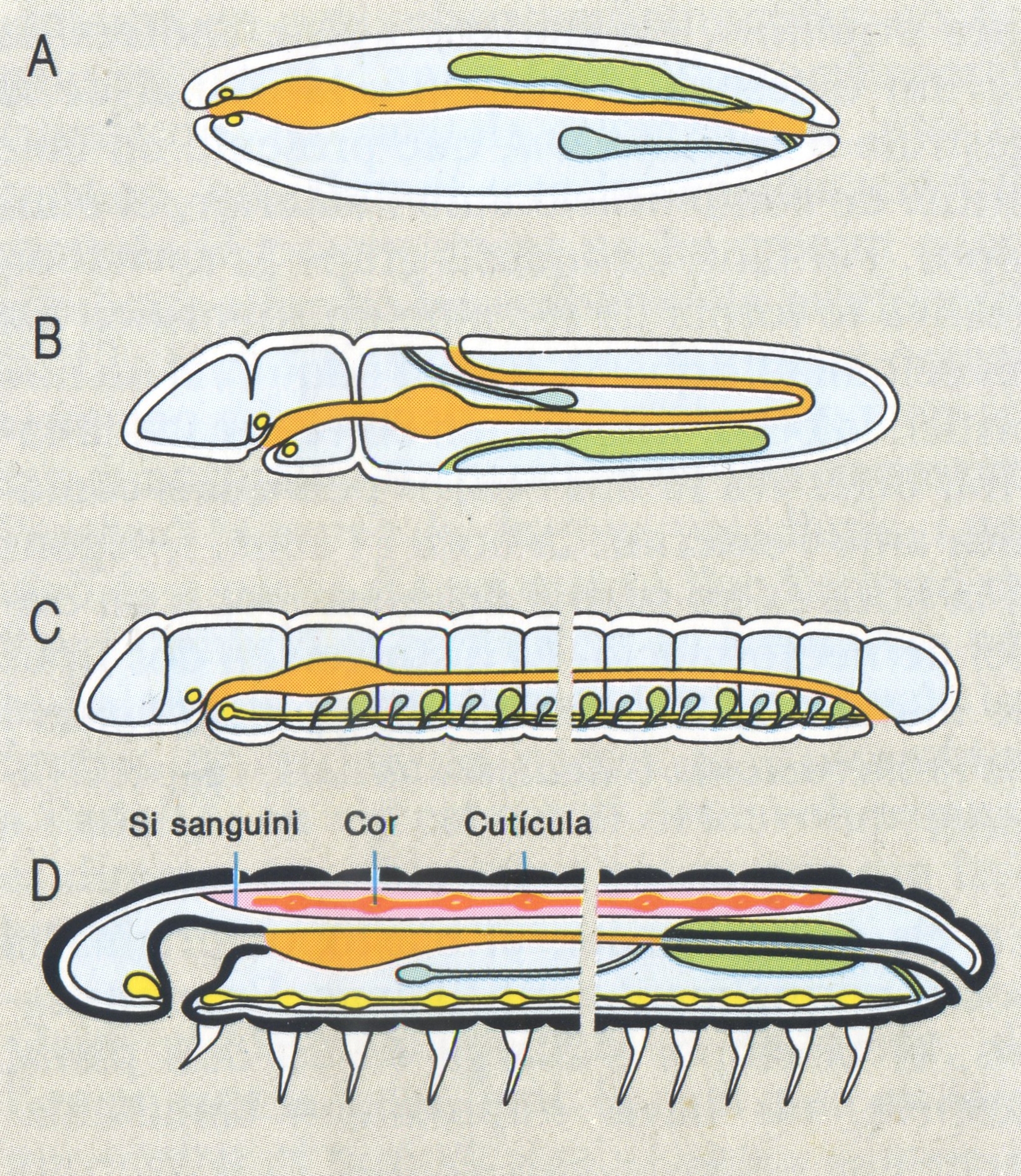
Models generals de l’organització del cos en els organismes invertebrats celomats. A Organització monomèrica. B metamerització oligomèrica, C metameritzaciò metamèrica homònoma (en el cas dels anèl·lids), D metamerització metamèrica heterònoma (en el cas dels artròpodes).
Biopunt. a partir de Barnes i altres
És un criteri molt important a l’hora d’establir els tipus d’organització. En els metazous més primitius, els cnidaris i els ctenòfors, la cavitat digestiva correspon a la cavitat de la gàstrula o arquènteron, que presenta un únic orifici de comunicació amb l’exterior, el blastòpor. Per tant, en aquests grups, la cavitat digestiva té forma de sac amb un sol orifici. En els celomats, aquesta cavitat adquireix un nou orifici, la boca. Llavors, en els celomats, la cavitat digestiva té dos orificis pels quals comunica amb l’exterior: la boca i l’anus; i, segons si la boca és un vestigi del blastòpor o un orifici de neoformació, es distingeixen dos tipus de metazous, els protòstoms o protostomats i els deuteròstoms o deuterostomats, respectivament.
En els protostomats, el blastòpor origina la boca i normalment també l’anus. En molts casos, el blastòpor s’allarga en un dels plans; després, els llavis s’aproximen simultàniament i se solda la part central de l’obertura deixant dos orificis a l’extrem, que corresponen a la boca i l’anus; la cavitat digestiva deixa d’ésser un sac per a tenir la forma d’un tub obert pels dos extrems. Un altre aspecte important és allò que succeeix durant aquest procés als elements nerviosos. Aquests se situen sobretot al voltant del blastòpor i, a causa de les modificacions que aquest experimenta, passen a ocupar una posició ventral respecte del tub digestiu, amb dos anells que envolten la part anterior i posterior del tub digestiu, l’un dels quals, el darrer, tendeix a desaparèixer. Aquest tipus de sistema nerviós rep el nom d’hiponeure i és característic de la majoria dels protostomats (anèl·lids, artròpodes, mol·luscs, entre d’altres). D’altra banda, els protostomats presenten segmentació espiral, i en molts casos el mateix tipus de formes larvals (larva trocòfora); molts són metameritzats i tenen el celoma format per esquizocèlia.
En els deuterostomats, el blastòpor es transforma en l’anus o bé marca l’emplaçament d’aquest orifici, i la boca és de neoformació. La segmentació no és mai espiral, sinó que en general és radial, i el celoma es forma per enterocèlia. El nivell de diferenciació i la posició del sistema nerviós permet de separar dins aquest conjunt d’organismes dos grans grups, d’acord amb Beaumont Cassier: els epiteloneures i els epineures. Els epiteloneures reuneixen organismes marins arcaics com els equinoderms, els estomocordats i els pogonòfors. En aquests organismes, el blastòpor manté les mateixes dimensions mentre es va desenvolupant l’embrió; durant aquest període es forma un orifici nou per mitjà de la invaginació d’una part de l’embrió fins que entra en contacte amb l’arquènteron; d’aquesta manera, el tub digestiu ja presenta dos orificis: la boca, de neoformació, i l’anus, derivat del blastòpor. El sistema nerviós en general es troba poc separat de l’ectoderma, caràcter del qual deriva el nom d’aquest grup. Paral·lelament, la cefalització és nul·la, i, almenys en la fase larval, hi ha tres parells de vesícules celomàtiques (organismes trímers). Els deuterostomats epineures (cordats) també presenten la boca de neoformació, però difereixen dels epiteloneures, entre altres caràcters, per la localització del sistema nerviós, que se situa per sobre del tub digestiu, com també per l’existència d’un eix esquelètic dorsal, la corda, situada entre el tub digestiu i el sistema nerviós, i de solcs branquials que, almenys en la fase embrionària, estableixen comunicació entre la cavitat faríngia i el medi extern.
Abans que es coneguessin tots aquests processos i les seves interrelacions, el conjunt dels celomats constituïen un grup molt divers i difícil d’interrelacionar. Durant molts anys, es va dividir el conjunt dels animals en dos grans grups: els invertebrats (animals que manquen de columna vertebral) i els vertebrats (animals que tenen columna vertebral). De fet, tots els animals excepte els del subtipus dels vertebrats pertanyen als invertebrats. Aquesta divisió entre invertebrats i vertebrats reflecteix més aviat la nostra perspectiva com a vertebrats del món animal. De fet, hi ha altres característiques diferents que, com hem vist, són d’una importància fonamental i que reflecteixen divergències evolutives molt més antigues que la formació de la columna vertebral.
La complexitat i la diversitat dels organismes multicel·lulars
L’organogènesi i el desenvolupament embrionari i postembrionari

Les formes larvals que inclouen els cicles biològics de la majoria dels grups d’animals invertebrats són variades i molt interessants des del punt de vista zoològic, ja que a partir del seu estudi hom pot imaginar possibles relacions entre diferents grups d’organismes. En alguns casos, l’aspecte i la biologia de les larves és tan diferent de la dels organismes adults que han estat considerades com a organismes adults d’una espècie diferent. Ha estat el cas de la larva dels foronis, que veiem en la fotografia, coneguda durant molt de temps pels zoòlegs com un gènere de foroni (Actinotrocha), i no com una forma larval.
Claude Carré
Amb la gastrulació no s’acaben els moviments morfogenètics, ja que encara es donen altres desplaçaments, molt variables segons el grup d’organismes. El blastòpor migra cap a una posició ventral, es formen braços i bandes ciliades en les larves aquàtiques, es destaquen blocs regulars en el meroblast de la majoria dels cordats, i es produeixen modificacions axials, com la formació del tub nerviós. En general ja es revela l’esboç de la forma embrionària. Tots aquests moviments morfogenètics, inclosa la gastrulació, constitueixen la morfocoresi.
En acabar-se la morfocoresi, grups de cèl·lules que no tenen encara cap diferenciació histológica comencen a adquirir una determinació. El germen apareixerà en aquest moment com un veritable mosaic de territoris encara no ben diferenciats, però amb un destí totalment fixat. Una vegada han aparegut, aquests territoris formadors d’òrgans es diferenciaran histològicament. Acabada la morfocoresi, cada capa embrionària originarà uns teixits, òrgans i aparells determinats: a partir de l’ectoderma s’originarà el sistema tegumentari, amb els seus derivats, i el sistema nerviós; a partir de l’endoderma s’originarà el gastroderma o teixit de revestiment del tracte digestiu; i, a partir del mesoderma es formaran, almenys en part, el sistema muscular, els òrgans excretors, l’aparell reproductor, l’aparell circulatori, les cavitats celomàtiques, etc. El germen, que durant tota la segmentació i la gastrulació pràcticament no ha crescut gens, comença a fer-ho en acabar-se la morfocoresi. Els teixits s’associen per a formar òrgans i, un cop diferenciats, els òrgans ja poden ésser funcionals. A partir d’aquest moment, l’embrió és capaç de dur una vida lliure, i per això és a partir d’aquí que té lloc l’eclosió de l’ou i l’embrió es converteix en larva. Ara bé, hi ha alguns grups d’animals en els quals les formes acabades de néixer ja s’assemblen a l’adult, i en aquest cas el desenvolupament és directe, és a dir, sense passar per cap fase larval.
Quan el desenvolupament és indirecte, de l’ou en surt la larva, amb la qual cosa es dona per acabat el període embrionari i s’inicia el desenvolupament postembrionari o període larval. La larva és capaç de viure per si mateixa i el seu aspecte en el moment de l’eclosió sovint és molt diferent del que adopta l’animal adult. En molts casos, aquesta larva ha de passar, abans no ateny l’estat adult, per moltes transformacions, conegudes en conjunt amb el nom de metamorfosi. El període que abraça des de l’eclosió de l’ou fins a l’adquisició de l’estat adult constitueix, doncs, l’anomenat període postembrionari. L’estudi de les característiques larvals i del desenvolupament postembrionari permet d’establir afinitats entre grups d’organismes. En l’organització de les larves cal distingir entre caràcters palingenètics i caràcters cenogenètics. Els caràcters palingenètics són caràcters arcaics característics de l’organització primitiva del grup taxonòmic, mentre que els caràcters cenogenètics són caràcters recents, adquirits per l’espècie o pel grup de què es tracti com a adaptació a noves condicions d’existència. Per tant, solament els caràcters palingenètics són vàlids per a establir relacions de parentiu entre grups taxonòmics distints.
Anatomia i funció

Els diferents sistemes i aparells interns són visibles per transparència en aquest organisme, una larva de llancet (Branchiostoma).
Claude Carré
Les cèl·lules que componen els organismes multicel·lulars són organitzades en teixits, òrgans, aparells i sistemes. Els teixits es defineixen com grups de cèl·lules d’estructura i funció similars. Els òrgans, formats per diversos teixits, són ben delimitats anatòmicament, i contribueixen a fer una funció més o menys específica en l’organisme. Per exemple, el cervell és format per diversos tipus de cèl·lules nervioses, i contribueix a coordinar i emmagatzemar informació a través del sistema nerviós central; el funcionament del cervell, però, depèn d’una entrada adequada de sang. Un altre exemple podria ésser el fetge, anatòmicament ben delimitat, format per diversos teixits i associat per la seva funció a l’aparell digestiu, el bon funcionament del qual també depèn d’una entrada correcta de sang.
En un sistema d’òrgans o en un aparell, els diferents components cooperen en la realització d’una funció, com ara la coordinació nerviosa o la digestió, d’acord amb els exemples emprats fa un moment. Excepte en el cas del sistema endocrí, els sistemes d’òrgans i els aparells mostren una continuïtat d’un òrgan a l’altre dins el mateix sistema. Especialment des del punt de vista anatòmic, és evident l’existència de diferències entre els teixits d’un mateix òrgan i entre els òrgans d’un sistema o aparell. Quan es comparen diferents tipus d’organismes, és a dir, diferents tipus d’organització o tipus taxonòmics, es posa de manifest que hi ha una gran varietat de sistemes i aparells, com també de models d’interrelació entre els òrgans que els componen, en el conjunt dels metazous, encara que les funcions que facin siguin les mateixes; per exemple, la funció del sistema nerviós és la coordinació de l’organisme, i la de l’aparell digestiu és la digestió. Es defineix un sistema com un conjunt d’estructures constituïdes per un sol tipus de teixit i distribuïts per tot l’organisme, és a dir, que és mal delimitat anatòmicament. Contràriament, un aparell és constituït per un conjunt d’estructures que ocupen una posició ben delimitada anatòmicament, destinades a fer una mateixa funció. A continuació ens ocuparem d’una manera breu dels diferents sistemes i aparells presents en els metazous.
El sistema tegumentari
El sistema tegumentari envolta el cos dels metazous. És constituït per un epiteli format per una o més capes de cèl·lules epitelials, a vegades recobert per cutícules dures (per exemple en els artròpodes) o per escates i estructures derivades, pèls, plomes i faneres (en els vertebrats). Associats al tegument hi ha glàndules de naturalesa diversa i altres mecanismes secretors, la funció dels quals és augmentar la naturalesa protectora de la coberta corporal exterior. El tegument és interromput bàsicament per obertures associades al tub digestiu, a l’aparell excretor, a l’aparell reproductor, a l’aparell respiratori i al sistema sensorial.
El sistema esquelètic
L’esquelet dels animals, que pot ésser extern (exosquelet) o intern (endosquelet), té una funció protectora i de suport. La naturalesa de l’exosquelet pot ésser molt variada: els coralls (cnidaris) secreten un esquelet de carbonat càlcic; diversos grups de verms presenten un tegument impermeable, recobert de cera; els mol·luscs i els braquiòpodes es protegeixen per mitjà de conquilles o valves de carbonat càlcic; els artròpodes disposen d’un exosquelet (cutícula) el component principal del qual és la quitina; el dels equinoderms és format per plaques protectores unides entre si; també les plaques òssies dels «peixos» primitius i les plaques dèrmiques de l’esquelet de la tortuga són considerades exosquelt. L’endosquelet és present en molts grups d’animals. De vegades consisteix simplement en un aparell hidrostàtic, d’altres és format per fibres de proteïna (espongina) o espícules calcàries o silícies (esponges), o bé és format per cartílag i os (vertebrats); algunes d’aquestes estructures esquelètiques són molt complexes i constitueixen valuoses fonts d’informació per als estudis de tipus evolutiu.
El sistema muscular
Aquest sistema és el responsable de la contracció de la paret del cos i de les estructures relacionades amb el moviment, tant del moviment de les estructures esquelètiques com dels moviments peristàtics del tub digestiu, dels moviments del cor, etc. La musculatura anomenada esquelètica és diferent de la musculatura visceral. El material contràctil més simple es troba en els protozous i es relaciona amb l’activitat contràctil dels cilis i els flagells i amb la formació de pseudopodis; la seva funció es relaciona amb la locomoció. En els metazous, el sistema muscular actua per mitjà de la contracció de la paret del cos, o activant el moviment dels apèndixs, per exemple. També té un paper important en el control de l’obrir i tancar de les obertures del tub digestiu i els esfínters, en el control del moviment de les brànquies, els sifons, el cor, etc. El teixit muscular és un dels més estudiats, atès l’interès que té el coneixement dels mecanismes moleculars que permeten la transformació d’energia química en mecànica, mecanismes que són molt uniformes en tot el món animal.
Els sistemes de relació i control
En els metazous més senzills el sistema nerviós pren l’aspecte d’una malla o xarxa formada per unes cèl·lules especials, les neurones. Gradualment, a mesura que els organismes presenten un grau més alt de complexitat i que el seu cos es va fent més gran, el sistema nerviós central adquireix una forma més definida i entra en relació amb els òrgans sensorials, també cada vegada més desenvolupats, la qual cosa permet una integració i coordinació més grans de les respostes als estímuls procedents del medi extern i del medi intern. Un sistema sensorial és essencial perquè els organismes duguin a terme una explotació efectiva del medi i, per tant, se situa a l’origen del comportament animal. D’altra banda, el sistema endocrí permet de regular importants funcions vitals per mitjà de les hormones. La seva existència es manifesta, per exemple, en grups d’animals tan diferents com els artròpodes i els vertebrats, i actua regulant en els primers el desenvolupament, la muda, el creixement, etc., í en els segons una gran varietat de funcions. En darrer terme, el funcionament del sistema endocrí és regulat pel sistema nerviós, la qual cosa permet una coordinació perfecta. L’existència d’hormones no és clarament provada en els organismes inferiors, tot i que hi ha proves de l’efecte que produeixen certes substàncies quan són alliberades al medi; per exemple, quan s’allibera glutanona al medi aquàtic, s’indueix el desenvolupament dels òrgans genitals en les hidres (cnidaris), i això, juntament amb altres fenòmens semblants, pot fer pensar que nombrosos invertebrats «inferiors» són capaços de segregar substàncies d’aquest tipus, o que, almenys, certes funcions són regulades per unes determinades substàncies químiques.
L’aparell digestiu
L’aparell digestiu presenta una obertura externa en relació amb una cavitat interna, o dues obertures, que posen en comunicació amb l’exterior els dos extrems de l’aparell digestiu, de tipus tubular, que presenta diverses diferenciacions en els seus diferents trams. D’aparells digestius amb una sola obertura en relació amb una cavitat gastrovascular solament se’n presenten, d’una manera primitiva, en els metazous més primitius, com els cnidaris i els ctenòfors. Les limitacions d’aquest aparell digestiu són la falta d’especialització i la dificultat de subministrar a totes les cèl·lules del cos els productes de la digestió, que en aquests grups és intracel·lular i té lloc a través de l’epiteli que envolta aquella cavitat. El transport dels productes digerits, el fan una sèrie de cèl·lules, encarregades de transportar els productes alimentaris a altres cèl·lules. En l’aparell digestiu amb dues obertures, boca i anus, gràcies a l’especialització dels diferents trams, la distribució dels productes de la digestió a totes les cèl·lules del cos és més senzilla, i a més és facilitada per l’existència d’un sistema circulatori, més o menys perfeccionat segons els grups. La digestió és fonamentalment extracel·lular i té lloc dins el tub digestiu (pot ésser afavorida per l’existència d’òrgans especials, que descarreguen els seus productes al tub digestiu i col·laboren en la digestió dels aliments), encara que en alguns casos es combina la digestió intracel·lular amb l’extracel·lular. En relació amb la part anterior del tub digestiu, poden haver-hi estructures de naturalesa i origen diversos, destinades a la trituració dels aliments (màstax dels rotífers, estómac triturador dels crustacis, mandíbules i paràgnats dels anèl·lids, ràdula dels mol·luscs, dents dels vertebrats, etc.).
L’aparell respiratori
Una altra funció vital dels éssers vius és la respiració, és a dir, la captació d’oxigen del medi extern i l’expulsió de l’anhídrid carbònic produït pel metabolisme. En els casos més senzills, l’intercanvi dels gasos respiratoris es fa a través de tota la paret del cos, que es troba profusament irrigada per vasos sanguinis. Aquest tipus de respiració rep el nom de respiració cutània, i és la que presenten en general molts organismes d’organització senzilla que viuen en ambients aquàtics o en indrets del medi terrestre amb un grau d’humitat molt elevat (si no fos així, estarien exposats a la deshidratació). A mesura que els animals augmentaren de talla corporal es va anar fent necessària l’aparició d’òrgans respiratoris—brànquies i pulmons—, localitzats en diferents parts del cos. Alguns grups, com ara els artròpodes terrestres, presenten un tipus especial d’òrgans respiratoris, les tràquees (filotràquees o pulmons dels aràcnids i dendrotràquees dels insectes i miriàpodes).
Les brànquies són formacions externes presents en nombrosos grups d’animals, en general animals aquàtics, que capten l’oxigen dissolt en l’aigua. Ara bé, les brànquies que presenten diferents fílums animals no sempre són homòlogues, ja que en molts casos són simplement òrgans anàlegs, és a dir, que fan la mateixa funció però no tenen el mateix origen. Presenten brànquies per exemple certs poliquets, els mol·luscs, els crustacis i alguns cordats, entre d’altres. Els pulmons, formacions internes que capten l’oxigen atmosfèric, són característics d’alguns grups d’animals de vida terrestre, com ara els amfibis adults, els rèptils, els ocells i els mamífers. També certs invertebrats, com els mol·luscs terrestres, tenen «pulmons», bé que són d’estructura molt diferent. Les tràquees, característiques dels artròpodes terrestres, com per exemple els insectes, consisteixen en invaginacions del tegument en forma de petits tubets pels quals circulen els gasos respiratoris, el transport dels quals queda totalment independitzat de l’aparell circulatori.
L’aparell circulatori
L’aparell circulatori és indispensable per a tots els organismes, ja que és el mecanisme de transport, és a dir, l’encarregat tant de dur els materials alimentaris des de l’aparell digestiu fins a les cèl·lules, i de recollir en aquestes els productes residuals que han de sortir a l’exterior, com del transport de gasos, l’oxigen i el diòxid de carboni (tret d’alguns casos excepcionals). L’aparell circulatori en els organismes més senzills és format per un líquid circulatori que és impulsat pels moviments del cos. En la perspectiva de l’evolució, a mesura que augmentava la talla corporal, aquest sistema va resultar insuficient i va aparèixer un sistema de vasos, primer senzill i després més complex, constituït per artèries, capil·lars i venes. Simultàniament a l’evolució d’aquest sistema de vasos va aparèixer un òrgan propulsor del líquid circulatori, el cor, el qual, per mitjà de contraccions, provoca el moviment d’aquest líquid, que, d’aquesta manera, pot arribar a totes les cèl·lules del cos. En relació amb la composició del líquid circulatori, en alguns organismes aquàtics és similar a la del medi (per exemple, en els equinoderms). No obstant, en la majoria dels organismes té una composició pròpia, i s’hi sol diferenciar un component líquid que serveix per a transportar productes metabòlics, i un componen cel·lular; aquestes cèl·lules poden contenir pigments respiratoris (com en els glòbuls rojos els vertebrats), poden formar part del sistema immunitari, etc. En la majoria dels metazous, el líquid circulatori, anomenat líquid hemàtic o hemolimfa, és l’encarregat de transportar totes les substàncies, però en els vertebrats, el líquid circulatori ha dividit les seves funcions establint dos circuits, el limfàtic i l’hemàtic.
L’aparell excretor
L’excreció és l’expulsió dels productes residuals procedents del metabolisme cel·lular. En els animals més senzills aquests productes s’eliminen per simple difusió, però, en la història evolutiva, ben aviat es van desenvolupar unes cèl·lules especials (flamígeres) i uns túbuls connectats que en conjunt constituïren els protonefridis, característics de les fases larvals de nombrosos fílums de protostomats i d’algunes formes adultes. Aquest aparell excretor va anar augmentant de complexitat. La part interna dels túbuls es va posar en relació directament amb les cavitat internes (celomàtiques) del cos, i va donar lloc a una estructura anomenada metanefridi o simplement nefridi, que caracteritza les fases adultes de diversos fílums de protostomats, o bé es va posar en relació íntima amb el sistema circulatori, amb la qual cosa va aparèixer el ronyó dels vertebrats; en aquest darrer cas, aquest sistema capta de la sang els productes que cal extreure del cos. L’activitat de l’aparell excretor també té molta relació amb l’osmoregulació de l’organisme, és a dir, amb la necessitat de conservar l’aigua del cos i de mantenir la pressió interna. Per aquesta raó, els animals que viuen en llocs molt secs excreten productes sòlids, mentre que els organismes aquàtics, que disposen d’aigua abundant, treuen els productes que cal excretar amb l’excés d’aigua. La naturalesa dels productes excretats d’origen nitrogenat també té una estreta relació amb la quantitat d’aigua disponible en l’ambient.
L’aparell reproductor
És integrat pels òrgans genitals, els testicles i els ovaris, on es produeixen els espermatozoides i els òvuls respectivament, i pels conductes que els posen en relació amb el medi extern a través dels orificis genitals. Hi ha algunes formes hermafrodites, que, per tant, presenten gònades masculines i femenines, bé que en alguns casos no coexisteixen, o no maduren al mateix temps ambdós tipus de gònades. La fecundació externa, bastant característica dels animals aquàtics, té lloc fora del cos, i els gàmetes són expulsats al medi (generalment aquàtic), i on es produeix la fecundació; en aquest cas, les estructures o glàndules accessòries, no estan, en general, gaire desenvolupades. La fecundació interna, característica dels animals terrestres, té lloc dins les vies genitals de la femella, i, per tant, va associada a la còpula (encara que de vegades la transferència és indirecta), que sovint és precedida d’un festeig nupcial més o menys complex; en aquest cas, les estructures i glàndules accessòries de l’aparell genital estan més desenvolupades i poden incloure vesícules per a l’emmagatzematge dels gàmetes, conductes per a transportar-los i òrgans copuladors. D’altra banda, en molts casos les espècies presenten caràcters sexuals secundaris, que es manifesten en un dimorfisme sexual més o menys acusat; generalment, el grau de diferència morfològica entre mascles i femelles s’acompanya d’un grau semblant de complexitat del comportament en el festeig nupcial.
Els grans grups sistemàtics
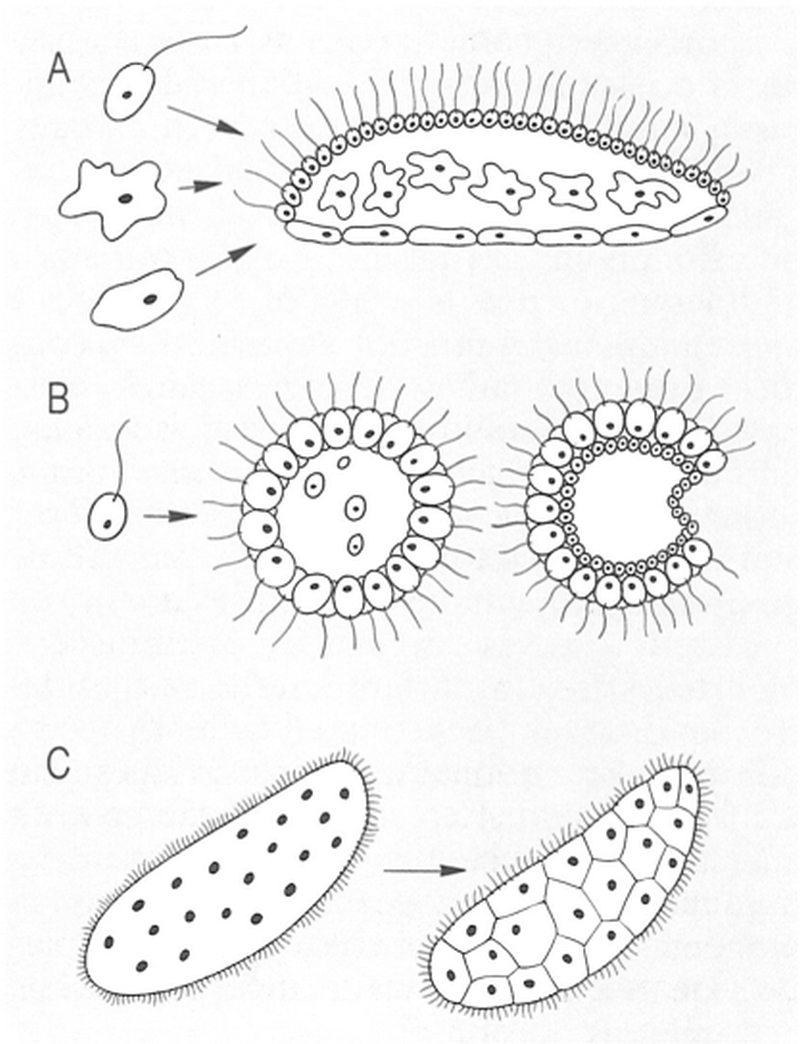
Esquema de diferents teories per a explicar les possibles vies de l’evolució dels animals multicel·lulars a partir de protists. A Via simbiòtica, B via colonial, C via de cel·lularització.
Biopunt, a partir de Barnes i altres
Els animals multicel·lulars s’han pogut originar a partir dels protozous per tres vies teòriques. En primer lloc, podria ésser que diversos protists s’haguessin associat simbiòticament i haguessin originat un organisme multicel·lular. Aquest és l’origen que es pressuposa per a les cèl·lules eucariotes a partir de les cèl·lules procariotes, i també per als líquens (metàfits), formats per la simbiosi d’una alga i un fong. En segon lloc, es podrien haver originat per via colonial, és a dir, a partir d’un protist que hagués experimentat divisions sense que els nous individus quedessin independitzats, de manera que s’haurien format colònies en les quals en alguns casos es podrien distingir diferents tipus de cèl·lules. En tercer lloc, els metazous es podrien haver originat a partir d’un protist multinucleat, mitjançant la cel·lularització dels diferents nuclis, amb la qual cosa hauria quedat compartimentat, i hauria passat així a ésser multicel·lular. D’aquests tres mecanismes possibles, el primer presenta seriosos problemes genètics, ja que no se sap com es podrien integrar diversos protists genèticament diferents en un únic organisme multicel·lular, i que fos capaç de reproduir-se; de fet, els dos o tres simbionts que formen els líquens es reprodueixen independentment i després s’associen altra vegada. Pel que fa al tercer mecanisme potencial, no hi ha indicacions sobre la compartimentació interna en els protists actuals, i no hi ha cap prova que això hagués pogut passar en el passat. Molts protists formen colònies i en alguns hi ha diferents tipus de cèl·lules, per la qual cosa aquest mecanisme sembla que és un dels més probables. De fet, és difícil de distingir entre colònies de protists i alguns organismes multicel·lulars. No tots els organismes considerats com a multicel·lulars presenten coordinació entre les seves cèl·lules, i la diferenciació cel·lular no és exclusiva dels organismes multicel·lulars. Segons quina d’aquestes hipòtesis es consideri, són uns o altres els grups de protists a partir dels quals els metazous s’han originat.
Esquema de dues de les teories que intenten explicar la radiació evolutiva dels metazous.
Carto-Tec, a partir de fonts diverses
Els organismes multicel·lulars més simples són els placozous, descrits recentment, i les esponges, que manquen de teixits organitzats en òrgans i presenten una forma poc definida. Tota la resta dels fílums, els que formen els veritables metazous, tenen teixits organitzats, en general, en òrgans, i aquests, en aparells.
Les esponges o porífers, es poden considerar per la seva organització, com una colònia de protists o com a organismes multicel·lulars. Tradicionalment s’han inclòs entre els animals, però en l’actualitat també hi ha qui els situa entre els protists. Les cèl·lules mostren una petita coordinació (no de tipus nerviós), no tenen simetria a escala individual ni a escala colonial, i els diferents tipus de cèl·lules de què disposen no formen teixits. Les esponges presenten unes cèl·lules molt característiques, els coanòcits, que són estretament emparentades amb un grup de protists, els coanoflagel·lats, que reuneix tant formes individuals com colonials. Molts zoòlegs opinen que les esponges deriven d’aquestes formes colonials. D’altra banda, els porífers constitueixen una línia evolutiva independent, a partir de la qual no s’ha originat cap altre grup.
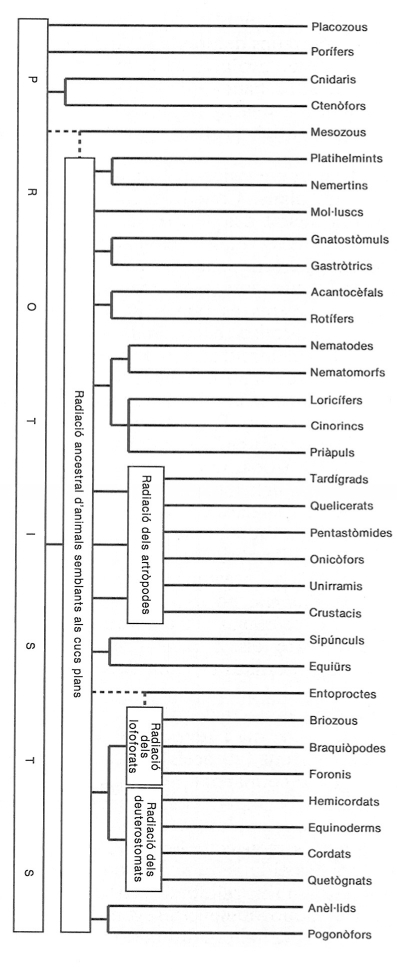
Esquema d’una de les possibles interpretacions sobre l’origen i la radiació dels animals multicel·lulars. En aquesta part de l’obra, hom ha seguit aproximadament aquest esquema, però no ha estat així pel que fa referència als artròpodes.
Carto-Tec, extret de l’obra de R.S.K. Barnes, P. Calow i P.J.K. Olive (1988), «The Invertebrates»
Com les esponges, els celenterats (cnidaris i ctenòfors) sembla que constitueixen una línia evolutiva independent, tot i que durant molt de temps es va considerar que es trobaven a la base de tota la resta dels metazous. Les peculiaritats del cos dels celenterats són la simetria radial, una organització en teixits amb coordinació entre les cèl·lules mitjançant un sistema de cèl·lules nervioses, i l’existència de dues capes cel·lulars separades per mesoglea gelatinosa i en general no cel·lular. Igualment, són molt característics orgànuls com els nematocists (cnidaris) i els col·loblasts (ctenòfors); orgànuls d’aquest tipus o semblants s’han trobat en alguns protists (mixozous, microspores i dinòfits). La forma del cos és com un sac amb una cavitat interior que comunica amb l’exterior per un orifici que fa de boca i anus simultàniament. Són diploblàstics, condició que recorda l’estat de gàstrula del desenvolupament embrionari d’altres metazous, que en ells evoluciona posteriorment per a formar la tercera capa embrionària. Els celenterats han estat considerats com un vestigi, permanentment diploblàstic, de les formes ancestrals que van originar els metazous triploblàstics, segons la teoria de la recapitulació de Haeckel. Actualment, aquesta teoria està perdent credibilitat, ja que en l’evolució les formes adultes no se succeeixen les unes a les altres sinó que cada animal és el resultat final d’un procés ontogenètic, talment que la filogènia és la successió d’ontogènies completes. De tot això es pot deduir que l’estatus dels cnidaris no està clar i que la simetria radial, l’estadi diploblàstic i el model sacciforme no són raons determinants per a situar-los a la base de l’evolució de la resta dels metazous. D’altra banda, no estan emparentats amb altres animals i és probable que derivin directament dels protists.
Un altre grup considerat com a primitiu és el dels cucs plans. Concretament, Hadzi, a la primera meitat d’aquest segle, va considerar els turbel·Iaris (verms acelomats) com els veritables metazous primitius. Aquesta teoria és potser la més acceptada actualment. Altres grups de verms es poden haver originat a partir de protists ciliats multinucleats, per mitjà de cel·lularització dels nuclis, hipòtesi que concorda amb el concepte de protozou com a organisme acel·lular. Un metazou equival a un protozou. Per tant, els ancessors dels turbel·laris podrien haver derivat de protists ciliats i, es trobarien a la base evolutiva de tota la resta dels metazous. No obstant això, hi ha molts aspectes en contra d’aquesta teoria, ja que no té en compte els criteris fonamentats en l’embriologia i dona molta més importància a l’organització de l’adult. Si bé aquesta hipòtesi no és acceptada universalment, ha promogut divergències prou profundes entre els zoòlegs i ha permès de qüestionar l’estatut primitiu dels cnidaris.
Els turbel·laris actuals presenten simetria bilateral, el cos pla i revestit de cilis, i alguns dels seus grups (els acels) no tenen un sistema digestiu permanent, tenen un sistema nerviós simple i un aparell reproductor senzill (en comparació amb els restants platihelmints, en els quals és molt complex). Pel que sembla, aquests turbel·laris primitius podrien estar relacionats amb els ciliòfors, i especialment amb els estefanopogonomorfs, segons R.S.K. Barnes. Un model alternatiu podria suposar que el protist ancestral era un flagel·lat que quan va adquirir l’esmentat estadi multicel·lular va augmentar el nombre de flagels i cada cèl·lula va adquirir la condició de ciliada.
Hi ha dos grups, els placozous i els mesozous, que sembla que deriven directament dels protists. Tots dos s’assemblen més a una colònia de protists que no a organismes multicel·lulars, ja que no tenen sistema nerviós, ni muscular, ni digestiu, ni tampoc cèl·lules esquelètiques, teixits o òrgans. Els mesozous, tot i que no tenen teixits, presenten el cos d’una forma definida i organitzada, amb simetria bilateral; són ciliats i endoparàsits d’invertebrats marins. D’altra banda, la seva simetria bilateral i l’aspecte general que tenen de verm suggereixen que podrien estar relacionats amb els cucs plans, i que la seva simplicitat és una conseqüència de la seva vida paràsita. Però les peculiaritats que presenten sobretot en relació amb els seus cicles de vida i el seu tipus de reproducció suggereixen que no són descendents d’aquells verms ancestrals.
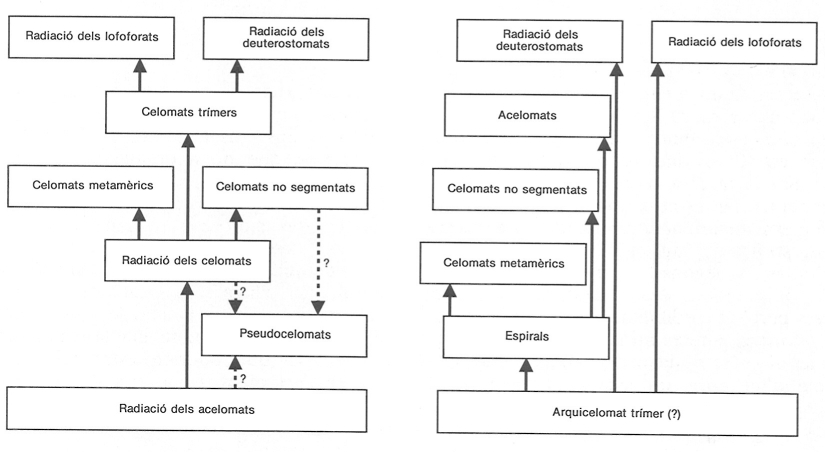
Els animals multicel·lulars poden procedir d’un stock únic de protists (serien, per tant, monofilètics), o bé podrien haver aparegut a partir de diversos grups de protists (en aquest cas, serien polifilètics). No obstant això, només una d’aquestes línies filètiques, d’animals de simetria bilateral, hauria tingut èxit i s’hauria arribat a diversificar. Malgrat que hi ha nombroses teories sobre l’origen dels metazous de simetria bilateral, en l’actualitat únicament es plantegen dues alternatives com a possibles generadores de la posterior radiació adaptativa: un cos sòlid (sense cavitats, celomat planuloide), o bé un cos amb cavitats plenes de líquid (celomats del tipus trímer). Aquesta darrera hipòtesi implica, en els metazous «inferiors», la pèrdua de la complexitat estructural, és a dir, la pèrdua de la trimeria i l’obliteració del celoma, i a més fa molt difícil d’explicar, des d’un punt de vista funcional, el desenvolupament tan precoç de les cavitats celomàtiques en el curs de l’evolució dels metazous. La primera hipòtesi, la més acceptada actualment, vincula l’increment de la complexitat estructural amb l’augment de les dimensions corporals i amb la colonització de nous hàbitats, alhora que evita les dificultats abans esmentades.
La radiació dels acelomats abasta els platihelmints i els nemertins, bàsicament. L’augment de les dimensions corporals en els animals podria haver afavorit l’explotació de nous recursos alimentaris i hauria pogut portar avantatges de tipus ecològic i evolutiu, ja que permetia de colonitzar nous hàbitats. Alhora, per tal de poder nedar, excavar i desplaçar-se, en general, es necessiten mecanismes locomotors, ja que els cilis de les espècies petites es tornen clarament insuficients; podria ésser que aquest problema locomotor s’hagués resolt en aparèixer les cavitats internes, que fan d’esquelet hidrostàtic i alhora intervenen en la locomoció. Entre els grups amb cavitats internes es distingeixen els pseudocelomats, un grup força heterogeni que podria haver sorgit per diverses vies separades dels acelomats, i els celomats. Entre els celomats s’evidencien tres tipus d’organització: els celomats unisegmentats, que són sedentaris (sipúnculs i equiürs, per exemple), els celomats metamèrics, que són mòbils (anèl·lids i artròpodes), i els celomats oligomèrics (trímers), que són tubícoles (briozous, braquiòpodes, foronis, etc.). Tots tres tipus d’organització es podrien haver desenvolupat per tres vies independents a partir dels acelomats.
L’èxit de la radiació dels metazous metamèrics està en relació amb l’aparició i l’evolució dels apèndixs locomotors, característics dels artròpodes. Malgrat l’èxit evolutiu dels artròpodes, encara no es coneix amb certesa quin és el seu punt d’origen. D’altra banda, l’organització de tipus trímer, representada pels pterobranquis (hemicordats) i els foronis, va produir dues radiacions importants, la dels deuterostomats (equinoderms i cordats) i la dels lofoforats, respectivament.