L’evolució dels vegetals al llarg dels temps geològics
De la mateixa manera que hom pot atribuir els estrats a períodes diferents segons que la fauna fòssil que contenen sigui més o menys evolucionada, també poden datar-se els estrats i assignar-los una determinada antiguitat a partir dels vegetals fòssils que hi apareixen. Per similitud amb les tradicionals eres geològiques establertes segons les restes fòssils d’animals que contenen les capes, la història del nostre planeta, tenint en compte l’evolució dels vegetals que l’han poblat, es pot dividir en les tres grans eres. El Paleofític (era dels pteridòfits o dels vegetals antics) que, en línies generals, coincideix bastant amb l’Era Paleozoica, es pot considerar que comença quan apareixen les primeres criptògames vasculars, és a dir, durant el Silurià superior, i acaba en canviar el clima i fer-se més àrid, en el Permià superior; hi predominen les equisetals, les licopodials, les filicals i les prefanerògames, i el seu moment de màxima esplendor es comprèn entre el Devonià inferior i el Carbonífer superior. El Mesofític (era de les gimnospermes o dels vegetals mitjans) que, aproximadament, coincideix amb l’Era Mesozoica, es comprèn entre el Permià superior i el Cretaci mitjà, moment en què s’accelera l’expansió de les angiospermes, mentre que fins llavors predominaven àmpliament les gimnospermes (bennettitals, pinals, taxals, etc.). El Cenofític (era de les angiospermes o dels vegetals recents), també es pot comparar amb l’Era Cenozoica i va des del Cretaci mitjà, moment en què moltes gimnospermes comencen a declinar mentre que les angiospermes assoleixen una ràpida ascensió, fins als temps actuals, en què han arribat al seu grau de desenvolupament més gran. Abans, però, de l’aparició dels pteridòfits, hi ha llargs períodes geològics durant els quals la Terra era habitada pels primers vegetals, de vida aquàtica, a partir dels quals s’originà la vida vegetal terrestre.
Les primeres restes fòssils
En estrats que s’han datat com a pertanyents al Precambrià, és a dir, amb una antiguitat superior als 600 milions d’anys, ja apareixen restes fòssils que indiquen l’activitat vital dels bacteris i de les cianofícies, éssers vius que es consideren els més primitius dins del regne vegetal. Possiblement, entre 1800 i 1500 milions d’anys enrere ja existien aquests pioners del món vegetal, els quals es desenvolupaven en aigües i en zones humides.
Hi ha citacions de Micrococcus en capes precambrianes; també es troben bacteris filamentosos ferruginosos que s’assemblen als Clamydothrix i diversos senyals de bacteris, fins i tot en els estrats més antics que existeixen al nostre planeta (Huronià).
No hi ha dubte de la presència de les cianofícies en els temps precambrians; de fet, els estromatòlits observats en sediments pertanyents a diverses divisions de l’Era Arqueozoica es consideren productes de la seva activitat. Les escitonemàcies es coneixen també des del Precambrià. Des dels primers moments de l’activitat sedimentària al nostre planeta, apareixen impressions fòssils de l’activitat vital dels vegetals més primitius, com ara les cianofícies i els bacteris; i de llavors ençà hom aprecia senyals de la seva existència en estrats corresponents a tots els períodes geològics.
Els diversos grups d’algues constitueixen vegetals ja més evolucionats, pel fet que presenten cèl·lules eucariotes, cosa que els diferencia dels bacteris i les cianofícies, els quals presenten cèl·lules procariotes (sense un veritable nucli); aquestes plantes aquàtiques (rodofícies, crisofícies, carofícies, feofícies, clorofícies, etc.) poblen tant les aigües salades com les aigües dolces de la Terra, ja des de temps anteriors al Cambrià; es van fent freqüents des de les primeries dels temps cambrians, i apareixen des d’aleshores en les capes de les diverses eres, amb formes similars a les existents en l’actualitat.
Les algues necessiten, per a desenvolupar-se, l’existència d’aigua, i per aquest motiu viuen sempre en medis aquosos. Presenten un tipus d’organització vegetal anomenat tal·lus i que consisteix en el fet que tenen un veritable nucli cel·lular, manquen de vasos conductors, arrels i arquegonis; llurs esporangis i gametangis no són gairebé mai envoltats d’un estrat de cèl·lules estèrils.
Fins al Silurià superior, és a dir, fa uns 320 milions d’anys, tots els vegetals que poblaven la Terra vivien a les aigües, salades de la mar o dolces dels llacs, els rius i les masses palustres, i també podien desenvolupar-se en llocs molt humits de la superfície terrestre.
Per tal que les plantes poguessin alliberarse d’aquesta dependència total de l’aigua i començar a colonitzar la terra ferma dels continents, fins llavors desproveïda de vegetals, van haver de desenvolupar les estructures vitals adequades que els van fer possible de viure en el medi aeri. Les plantes que han aconseguit evolucionar fins a assolir el nivell de perfecció suficient procedeixen de les algues clorofícies.
El Paleofític
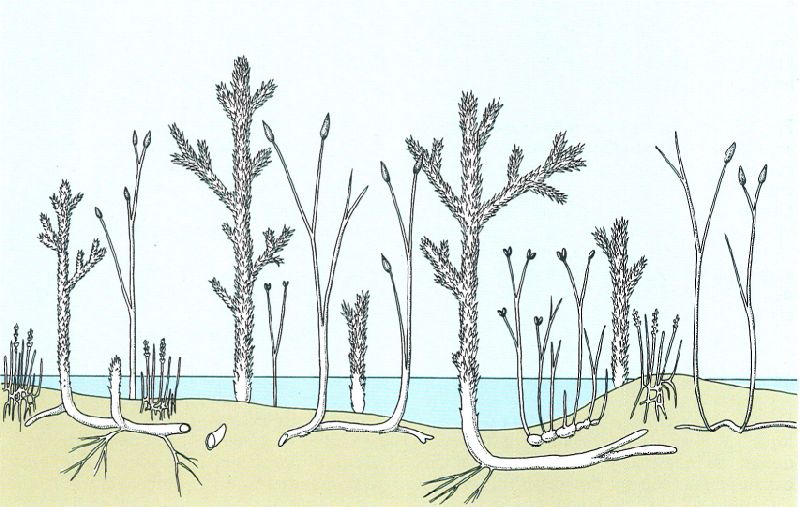
Reconstrucció teòrica d’un paisatge del Silurià superior que pot caracteritzar els ambients vegetals del que hom coneix com a Paleofític, en els temps d’aparició de les plantes vasculars terrestres (Silurià superior-Devonià mitjà). Hom hi ha representat Rhynia, Asteroxylon. Hornea i Zosterophyllum, entre d’altres.
Jordi Corbera, a partir de fonts diverses
Durant el Devonià i el Carbonífer es desenvolupen les diverses línies evolutives dels pteridòfits, els quals arriben a la seva màxima expansió i al seu màxim desenvolupament, s’adapten als distints medis terrestres, però necessiten, per a reproduir-se, un ambient humit en determinats moments del seu cicle vital. Presenten un sistema vascular que forma els feixos conductors ben diferenciats, constituïts per una part cribrosa i una altra de llenyosa; com a elements conductors d’aigua, posseeixen les traqueides lignificades i molt rarament existeixen també tràquees. Mitjançant les traqueides lignificades, augmenta considerablement la resistència, fet que permet als pteridòfits de desenvolupar-se com a plantes terrestres —la qual cosa no han pogut dur a terme les algues ni els briòfits— i arribar a assolir port arbori de vegades molt ramificat, tal com ho demostren les restes de grans plantes trobades en estat fòssil en estrats pertanyents al Carbonífer.
En el Carbonífer, fa uns 250 milions d’anys, el nostre planeta era cobert per una luxuriant vegetació constituïda per nombroses famílies de les diverses classes de pteridòfits. El clima devia afavorir el desenvolupament d’aquests vegetals en àmplies zones, atès que era més aviat càlid i humit, i, per això, moltes espècies van arribar a assolir dimensions enormes.
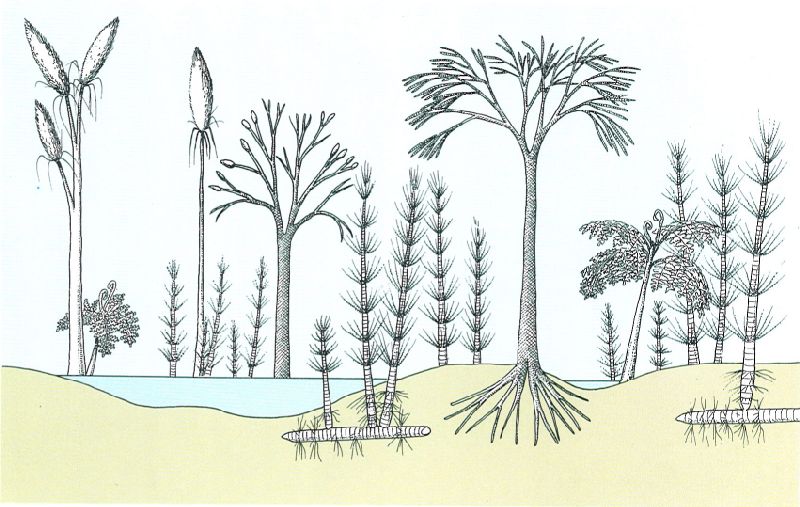
Reconstrucció teòrica d’un paisatge del Carbonífer, al Paleozoic. Hom hi ha representat diverses espècies arbòries que caracteritzaven aquestes formacions vegetals que són l’origen de les conques hulleres, amb Lepidodendron, Calamites, Pecopteris i Sphenophyllum, entre d’altres.
Jordi Corbera, a partir de fonts diverses
Les principals classes en què es divideixen els pteridòfits són: les licopodials, les equisetals i les filicals. Les licopodials assoleixen la seva màxima esplendor durant el Carbonífer mitjà, i a partir del Permià inicien la davallada; en destaquen els gèneres Lepidodendron, Sigillaria, Bothrodendron, Ulodendron, Lepidocarpon, Pleuromeia, etc., representats per nombroses formes arborescents que han arribat a atènyer dimensions molt grans. Les lepidospermals presentaven formacions semblants a les llavors. La poca perfecció en el sistema conductor de l’aigua en aquests vegetals i en la seva absorció degué conduir a la desaparició de les formes arbòries al final del Paleozoic, en iniciar-se un període d’aridesa que eliminà les espècies que no pogueren adaptar-se a aquestes noves condicions, i féu que s’expandissin altres tipus amb un sistema conductor més perfecte, i, així, ha arribat als nostres dies una mínima relíquia d’aquesta classe tan important durant el Paleozoic.
Les equisetals, que actualment són representades per un únic gènere, assoliren el seu moment de més gran expansió al llarg de l’Era Primària, i començaren a minvar al començament del Mesozoic. Entre els gèneres més representatius trobats en estat fòssil podem assenyalar: Calamites, Annularia, Asterophyllites, Pinnularia, etc. Les formes heterospòriques que apareixen en determinades famílies són les més recents (no es coneixen plantes amb aquesta característica anteriors al Carbonífer), fet que les converteix en menys evolucionades que les licopodials, que ja presenten formes heterospòriques des del Devonià. Pel que fa a les seves estructures anatòmiques, són, en conjunt, més evolucionades que no pas la classe tractada anteriorment, ja que mostren una elevada diferenciació funcional.
Entre les criptògames vasculars, les filicals constitueixen la classe que ha arribat a un major desenvolupament i, actualment, n’existeixen unes 10 000 espècies vives, de les quals unes tres quartes parts pertanyen a la família de les polipodiàcies. Aquest important grup de pteridòfits aparegué durant el Devonià mitjà, i els seus avantpassats cal cercar-los, igual que els de les altres dues classes, entre les psilofitals; les primofilicines constitueixen el nexe d’unió entre els dos grups. Presenten un cert grau d’afinitat amb aquestes primitives plantes vasculars, però tenen caràcters propis d’un nivell evolutiu superior, el trànsit entre les psilofitals i les primofilicines és tan insensible que és impossible de precisar si formes com Protopteridium han de col·locar-se ja entre les falgueres.
Totes les filicals tenen megafil·les, anomenats frondes, proveïts de nervació abundant, enrotllats en l’àpex quan són joves, excepte en les ofioglossals, i amb nombrosos esporangis a l’anvers.
Les falgueres actuals comprenen tres subclasses: les eusporangiades, les leptosporangiades i les hidroptèrides. Les eusporangiades, les més primitives, assoliren un gran desenvolupament durant el Carbonífer i el Permià, i començaren a davallar a partir del Mesozoic. Els ordres més importants són, en primer lloc, les ofioglossals, falgueres recents, i les marattials, que ja havien aparegut al Carbonífer, procedents de primofilicines, que eren molt abundants al Carbonífer i al Permià, i de les quals destaquen els gèneres Megaphyton, Hagiophyton, Caulopteris, etc., que assoleixen mides superiors als deu metres d’alçada.
Les leptosporangiades constitueixen el grup de falgueres vivent més nombrós, amb unes 9000 espècies, que viuen principalment a les zones intertropicals, tenen dimensions molt variables, des de pocs mil·límetres de longitud fins a 15 m d’alçada, i entre ells es compten les formes arborescents dels gèneres Cyathea, Alsophila, Dicksonia, Cibotium, etc. Les falgueres leptosporangiades apareixen ja al Carbonífer i anaren progressant durant el Mesozoic i el Cenozoic; actualment es distribueixen en uns 250 gèneres. La subclasse de les hidropteridals comprèn dues famílies amb uns quants gèneres que viuen en aigües i sòls palustres. Es coneixen restes fòssils pertanyents a la família de les salviniàcies: el gènere Azolla s’ha citat ja en capes del Cretaci inferior i Salvinia en el Paleocè.
Una gran línia filogenètica dins del regne vegetal que es pot situar entre les criptògames vasculars i les gimnospermes és la que formen les denominades prefanerògames, que comprenen les pteridospermes i les cordaïtes. Les prefanerògames han estat incloses durant molt de temps dins de les gimnospermes, però aquestes plantes contenen òvuls, i no llavors, característica principal de les espermafites (gimnospermes i angiospermes); constitueixen un estadi superior en l’evolució dels pteridòfits, en el qual diversos grups de criptògames vasculars posseeixen òvuls, és a dir, macrosporangis més o menys embolcallats per un tegument, mentre que, en els pteridòfits, es mantenen nus. Aquests vegetals són pròxims a les fanerògames i, en un nou pas en l’evolució, podran assolir aquest nivell; per aquest motiu hom les anomena prefanerògames per tal de diferenciar-les alhora de les fanerògames, que són més evolucionades, i de les criptògames vasculars, més primitives que elles. Es tracta d’un esglaó filogenètic situat entre aquests dos grans grups sistemàtics, els quals han ocupat una posició important entre totes les flores d’anteriors períodes geològics, sobretot durant el Paleozoic, i presenten actualment restes molt escasses que pertanyen al grup de les cicadals i al de les ginkgoals.
Les prefanerògames comprenen les pteridospermes, dins de les quals s’inclouen les pteridospermals (del Devonià al Cretaci), les glossopteridals (del Permocarbonífer al Retià), les caitonials (del Juràssic al Cretaci), les cicadals (del Triàsic fins a l’actualitat) i les nilsonials (del Keuper al Cretaci superior); i les cordaïtes, dins les quals trobem les cordaïtals (del Devonià al Permià, com a mínim) i les ginkgoals (del Permià inferior fins als nostres dies). D’entre les prefanerògames podem destacar els gèneres següents: Lyginopteris (Carbonífer), Heterangium (del Carbonífer inferior al Permià inferior), Sphenopteris (Carbonífer), Medullosa (del Carbonífer mitjà al Permià inferior), Rhexoxylon (del Permocarbonífer al Retià), Colpoxylon (Permià), Megaloxylon (Carbonífer), Cycadoxylon (del Carbonífer superior al Permià), Calamopitys (del Devonià superior al Carbonífer inferior), Mariopteris (Carbonífer mitjà), Callipteris (del Carbonífer superior al Permià), Glossopteris (del Permocarbonífer al Retià), Mesoxylon (Carbonífer), Ginkgo (del Juràssic mitjà als nostres dies), etc.
El Mesofític
Els espermatòfits són els vegetals que han assolit el màxim grau d’evolució i presenten alternança de generacions heteromorfa i de fases nuclears, amb haplofase i diplofase. Tal com succeeix amb els pteridòfits heterosporis i les prefanerògames, els espermatòfits originen, després de la meiosi, micròspores i megàspores. La reducció dels gametòfits o protal·lus masculins i femenins arriba a la màxima expressió: no són visibles externament i han de nodrir-se a partir de l’esporòfit. La divisió dels espermatòfits comprèn les gimnospermes, que tenen les llavors nues sobre les bràctees carpel·lars, i les angiospermes, amb les llavors protegides per les fulles carpel·lars, que formen l’ovari, el qual, posteriorment, originarà el fruit.
Les gimnospermes comprenen les coníferes i les bennettitines. Les coníferes s’han originat a partir de les cordaïtals i són les gimnospermes més antigues, atès que hom comença a tenir-ne senyals en el Carbonífer superior; no obstant això, les bennettitals podrien tenir una antigor similar que encara no s’ha confirmat. Les coníferes ocupen encara avui un lloc important en la vegetació del planeta, però des del Cretaci mitjà es troben en una davallada progressiva i són substituïdes per les angiospermes, més adequades a expandir-se en tots els ambients terrestres; així, les coníferes queden relegades a les zones on els espermatòfits més evolucionats tenen més dificultat a accedir.
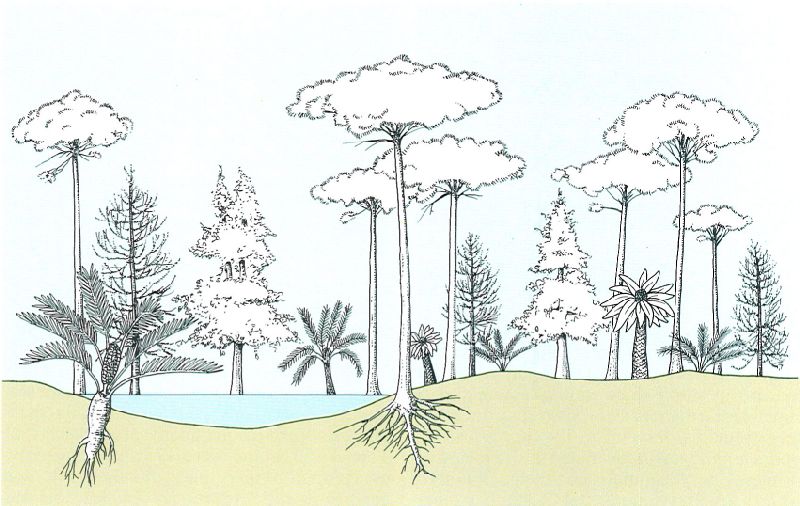
Reconstrucció teòrica d’un paisatge durant el Mesozoic, en què abundaven les falgueres i les gimnospermes prefanerògames, com Gynkgo, diverses cicadals, Araucarites, filicals, etc.
Jordi Corbera, a partir de fonts diverses
La línia evolutiva de les coníferes comprèn vegetals llenyosos que arribaren a la seva màxima expansió al final del període juràssic, en què dominaren a les flores mesofítiques; n’existien uns 55 gèneres, que reunien unes 580 espècies vives i s’adaptaven millor que els pteridòfits als medis terrestres. Podien viure en ambients més freds i secs que no pas aquests últims, per la qual cosa, a partir del Permià mitjà, moment en què iniciaren un cert desenvolupament, començaren a eliminarlos en les zones on no podien competir-hi avantatjosament. Les coníferes han mantingut una elevada força de competència, sobretot en els biòtops extrems per a les plantes arbòries. La classificació sistemàtica de les coníferes inclou, ultra els gèneres aïllats fòssils que visqueren en el Paleofític i el Mesofític i que no han pogut situar-se en unitats sistemàtiques superiors de manera clara, sis grups principals: les lebaquials, les pinals, les cupressals, les podocarpals, les araucarials i les taxals. Les primeres, les lebaquials, que inclouen només formes fòssils, visqueren del Carbonífer superior al Permià inferior, i comprenen la família de les lebaquiàcies, amb diversos gèneres, entre els quals podem destacar Lebachia, que només apareix a l’hemisferi N (de l’Estefanià mitjà al Permià inferior), i Ernestiodendron (de l’Estefanià al Permià); entre els gèneres afins trobem Voltzia (Triàsic inferior), Pseudovoltzia (Permià superior) i Podozamites (del Retià al Cretaci superior). L’ordre de les pinals inclou la família de les pinàcies, la més important de les gimnospermes actuals, que compta amb uns 10 o 11 gèneres, alguns dels quals tenen nombroses espècies: Pinus (unes 150 espècies), Abies (60 espècies), Picea (50 espècies). Entre les formes fòssils trobem diverses espècies d’aquests gèneres. Les cupressals, amb les famílies de les cupressàcies i les taxodiàcies, que comprenen 22 i 10 gèneres vius respectivament, tenen una gran quantitat de representants fòssils, sobretot al Cretaci i al Terciari, dels quals podem destacar: Taxodium distichum, Sequoia langsdorfii, Metasequoia glyptostroboides, Tetraclinis articulata, etc., freqüents a les flores terciàries de l’hemisferi N. Les podocarpals inclouen 7 gèneres vivents i aproximadament 150 espècies, principalment de l’hemisferi austral, difoses pels boscos montans dels països tropicals i subtropicals; sis d’aquests gèneres tenen representants fòssils, entre els quals cal remarcar Podocarpus, abundant al Cretaci i al Terciari. El cinquè ordre, el de les araucarials, es compon d’una única família amb dos gèneres vivents: Araucaria i Agathis, tots dos de l’hemisferi S. Les restes més antigues fossilitzades d’Araucaria, o de gèneres molt propers, apareixen al Triàsic superior i són freqüents al Juràssic i al Cretaci. Finalment, l’ordre de les taxals inclou dues famílies vivents, les cefalotaxàcies, petita família amb un gènere i sis espècies a l’Àsia oriental, i les taxàcies, amb cinc gèneres encara vius, entre els quals destaquen Taxus i Torreya, coneguts tots dos des del Juràssic mitjà.
Les bennettitals constituïren un món durant el Mesofític. Formaven un grup molt nombrós que hom ha estimat en 30 000-40 000 espècies. Apareixen en el Permocarbonífer i troben el període de màxima esplendor durant el Cretaci inferior. Són descendents de les pteridospermes, juntament amb les cicadals; s’extingiren durant el Cenomanià (Cretaci). Les bennettitals tenen molts trets comuns amb les cicadals, tants que sembla que no hi ha cap mena de dubte sobre les seves estretes relacions filogenètiques; tanmateix, el fet que les bennettitals formin llavors autèntiques assenyala el grau de diferenciació entre dues línies tan afins i un major desenvolupament evolutiu d’aquesta última, que l’ha portada a assolir el nivell de planta espermatòfita. No es coneixen formes intermèdies entre ambdós grups, per la qual cosa es desconeix, de moment, l’origen precís de les bennettitals; sembla que tenen un origen cicadal, però les dues línies devien separar-se molt aviat, com a màxim al començament del Triàsic, per a evolucionar paral·lelament. Els principals gèneres de les bennettitals, tots fòssils, són: Cycadeoidea, que comprèn un gran nombre d’espècies, del Keuper al Cretaci inferior, i és particularment freqüent a les fàcies wealdianes; Monathesia, del Cretaci superior; Williamsonia, del Triàsic al Wealdià; Williamsoniella, del Retià al Juràssic mitjà; i Wielandiella, del Retià al Juràssic.
Les clamidospermes són un fílum de vegetals molt aïllats en el món actual, amb afinitats molt discutides; presenten característiques a la vegada de gimnospermes i d’angiospermes. Les restes fòssils que s’han trobat i que poden situar-se en aquest grup són molt escasses i dubtoses. L’origen de les clamidospermes sembla bastant antic, ja que se n’esmenten possibles fòssils al Carbonífer, se cita la seva presència en capes datades com a pertanyents al Permià, al Triàsic, al Juràssic i al Cretaci, i també en capes del Terciari. Actualment comprenen tres ordres amb una família cadascun: efedrals, welwitsquials i gnetals, que presenten molt poques espècies i que tenen una incidència mínima en la flora actual.
El Cenofític
A partir del Cretaci mitjà comencen a davallar les gimnospermes i el seu lloc privilegiat en les flores del planeta comença a ser ocupat per les angiospermes, que són els espermatòfits més evolucionats. En aquest moment les pteridospermes han desaparegut pràcticament del tot, les ginkgoals han disminuït molt i les cicadals, abundants al començament del Cretaci, entren en fase d’extinció a la fi del Mesozoic i constitueixen grups merament supervivents ja a l’inici de l’Era Terciària. Les bennettitals tenen el seu màxim apogeu al principi del Cretaci, davallen ràpidament i s’extingeixen al Cretaci superior. Les coníferes, tan nombroses a l’inici del Cretaci, comencen a desaparèixer lentament al final de l’Era Secundària i arriben als nostres dies només amb una petita mostra del que havien estat en el passat.
L’aparició de les angiospermes
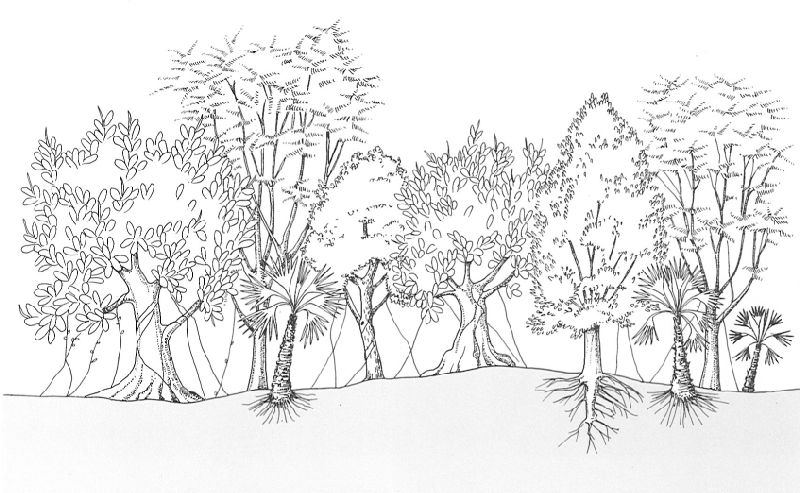
Reconstrucció teòrica d’un paisatge característic de l’Oligocè, a l’Era Terciària, on predominen les fanerògames, entre les quals Ficus, Sabal, Sapindus, Persea, etc.
Jordi Corbera, a partir de fonts diverses
El Cenofític o era dels vegetals recents es considera que comença a partir del Cretaci mitjà i arriba fins als temps actuals, ja que fou a partir d’aquest moment que les angiospermes començaren a substituir les gimnospermes com a nucli fonamental de les flores del planeta; aquest relleu es produí en un període curt de temps. El paper primordial que agafen a partir de llavors aquests vegetals més versàtils, més adaptats a la vida terrestre, amb una reproducció més eficaç i un cicle vital més adequat, és el que els ha permès de suplantar les gimnospermes, menys perfeccionades, en el domini dels principals ambients de la superfície de la Terra.
Les angiospermes ja existien molt abans del Cretaci mitjà; hom ha assenyalat la presència de restes fòssils d’aquest grup en el Triàsic, i possiblement pogueren, fins i tot, haver sorgit anteriorment. Durant el Mesofític devien haver viscut entre la gran massa de gimnospermes i passar totalment desapercebudes a causa del seu desenvolupament escàs; no és fins al Cretaci mitjà que adquireixen la força suficient, o no es donen les condicions oportunes, i, d’aleshores ençà, es produeix una ràpida expansió de les angiospermes, que, al cap de poc temps, predominen en les flores dels continents. Segons Axelrod, les angiospermes tenen el seu origen en regions del planeta de latitud baixa i, des d’allí, anaren progressant fins arribar a zones d’una latitud més elevada. Durant el Cretaci inferior les angiospermes no sobrepassaven els 40-43 graus de latitud N, mentre que, en latituds superiors, la flora continuava essent de tipus precretaci, sense presència d’angiospermes. Al Cretaci superior ja havien sobrepassat els 70 graus de latitud N. Les angiospermes constituïen el 10% de la flora del jaciment de la Val de Lobos (Portugal) durant el Cretaci inferior i, en canvi, les flores contemporànies d’Anglaterra, França i Bèlgica no contenen vegetals d’aquesta mena. A l’Aptià (Cretaci mitjà) la flora de Cercal (Portugal) era formada per un 25% d’angiospermes, mentre que a Greensand (Anglaterra) tot just comencen a aparèixer. En acabar l’Albià constitueixen el 85% de les espècies presents al jaciment de Nazaré (Portugal) i, en canvi, només representen el 10% del de Kome (Grenlàndia).
Les fanerògames mostren una diferenciació més gran en els seus teixits que no pas els vegetals que les han precedides, amb formació de tràquees i de tubs crivellats amb cèl·lules annexes. Tenen més plasticitat vegetativa, amb l’existència de formes anuals i perennes i presenten, també, vegetals llenyosos i herbacis. Totes aquestes possibilitats permeten que aquests vegetals s’adaptin amb facilitat a tota mena d’hàbitats.
L’origen filètic de les angiospermes
L’origen filètic de les angiospermes, ara com ara, no s’ha pogut determinar ben bé; hi ha raons per a considerar que el seu origen és bastant antic i que es van poder formar a partir de grups afins a les pteridospermes (liginopteridàcies). Un problema que encara no s’ha resolt és el de precisar quina de les dues classes en què hom les ha dividides, dicotiledònies i monocotiledònies, donà origen a l’altra. Avui en dia s’admet que la diferenciació entre aquestes dues classes és artificial i que totes dues es relacionen a través de les magnòlides. Les troballes fòssils de fulles que recorden les de les palmeres (Sanmiguelia), al començament de l’Era Secundària (Triàsic), sembla assenyalar que les monocotiledònies són més antigues, però existeixen restes, també del Triàsic, que se suposa que pertanyen a dicotiledònies (Furcula i Homoxylon). Si això fos així, aquestes dues línies serien bastant antigues i d’una edat bastant similar.
El gran polimorfisme que hom troba en les angiospermes, fins i tot en les seves branques més antigues, permet d’arribar a la hipòtesi que no procedeixen d’un únic grup de gimnospermes, sinó que el seu origen és múltiple, i que la complexa estructura d’aquests vegetals es va assolir mitjançant l’evolució de diverses formes. Aquest origen múltiple explicaria l’aparició simultània d’angiospermes a diferents punts del globus molt distants entre ells. No obstant això, hi ha nombrosos aspectes dubtosos que dificulten la confirmació que aquests vegetals tinguin un origen monofilètic o polifilètic, malgrat la gran quantitat de dades que afavoreixen aquesta segona hipòtesi.
Per tal de tenir una idea sobre l’origen filogenètic de les angiospermes, cal analitzar el món vegetal del qual han sorgit, ja que és a partir de l’evolució d’aquest que s’han originat. Així, si considerem les flores existents abans del Cretaci, remarquem els fets següents: d’una banda, entre les criptògames vasculars, les grans línies del Paleofític han desaparegut gairebé per complet, tret de les falgueres leptosporangiades, que continuen diversificant-se durant el Mesofític; en segon lloc, d’entre les prefanerògames, les pteridospermes i les cordaïtes típiques ja s’han extingit i han estat reemplaçades pels seus descendents (peltaspermàcies, caitonials, cicadals i ginkgoals), els dos últims grups dels quals arriben, en aquest moment, a la seva màxima esplendor; i, en tercer lloc, en aquests moments predominen les gimnospermes (bennettitals i coníferes), que es troben en ple apogeu i desenvolupament i posseeixen la diversitat de formes i prou força genètica per a originar aquest nou fílum vegetal.
Sembla clar que únicament les gimnospermes han pogut donar lloc a les angiospermes; tanmateix, hi ha una sèrie de problemes sense resposta adequada la qual ens permeti de precisar a partir de quin grup s’han originat o bé si ho han fet a partir de formes diferenciades anteriorment. Les clamidospermes poden tenir-se en compte a l’hora de determinar els avantpassats de les angiospermes, ja que l’organització floral d’aquests vegetals podria haver donat lloc a les flors angiospermes per simplificació de les seves inflorescències; lamentablement, no s’han trobat gaires restes fòssils d’aquest grup, i això fa que no disposem de prou dades sobre el seu passat.
Hi ha tres hipòtesis principals sobre l’origen de les angiospermes. La primera, defensada per Scott, Bower, etc., proposa que es relacionen amb les bennettitals per mitjà del grup primitiu de les policàrpiques, a causa de les seves flors hermafrodites; tanmateix, no sembla que sigui així, ja que, entre altres diferències, els suports dels primordis seminals són completament diferents. La segona hipòtesi es basa en el que Parkins i Arber anomenen hemiangiospermes: possibles grups de vegetals amb caràcters intermedis entre les bennettitals i les angiospermes que les enllacessin filogenèticament; no se n’ha trobat, però, cap resta fòssil. D’altres botànics, com Navaschin, Wettstein, etc., consideren que és per mitjà de les monoclamídies, les quals conceben com el grup més primitiu de totes les angiospermes, que s’estableix l’enllaç entre aquestes últimes i les gimnospermes, i troben en les clamidospermes l’explicació de molts dels caràcters intermedis existents entre les gimnospermes i les angiospermes.
Les característiques que han proporcionat una clara superioritat a les angiospermes sobre les gimnospermes i que els ha permès de vèncerles en la difícil competència pel domini en els medis terrestres, han pogut ser, principalment, sis: en primer lloc, l’hermafroditisme, que possibilita una reproducció més eficaç; en segon lloc, la protecció dels primordis seminals a l’interior dels carpels; en tercer lloc, l’existència d’una pol·linització zoòfila (principalment entomòfila), més econòmica i més ben dirigida que l’anemòfila; en quart lloc, la reducció considerable dels gametòfits i dels òrgans reproductors, amb la qual cosa la reproducció és més ràpida; en cinquè lloc, la plasticitat del seu cos vegetatiu, que no ha assolit cap més grup; i finalment, un sistema de xilema i floema molt més eficaç que en les altres plantes vasculars.
Les flores més recents
La flora existent al nostre planeta durant els últims 60 milions d’anys, és a dir, al llarg del Cenozoic, no ha canviat substancialment; es constitueix d’un elevat percentatge d’angiospermes, amb reajustaments florístics a causa dels canvis climàtics que han anat succeint-se i als quals ha hagut d’adaptar-se la vegetació. Les formes fòssils trobades en estrats que pertanyen a l’Eocè i a l’Oligocè no presenten diferències notables amb les espècies actuals del mateix gènere o de grups pròxims filèticament; això ens permet d’establir comparacions entre les espècies fòssils i les recents que ens proporcionen dades sobre les condicions ecològiques i climatològiques en què havien pogut desenvolupar-se.
L’àrea de distribució de molts gèneres i moltes famílies ha sofert modificacions a causa de les translacions de les plaques de l’escorça terrestre que han fet que els continents hagin anat canviant de posició amb el decurs del temps. Aquestes translacions originen canvis lents en el clima de la zona que fan que la flora hagi d’adaptar-s’hi i que les formes que no en són capaces siguin progressivament eliminades i altres espècies més adequades ocupin el seu lloc per desenvolupar-se en la nova situació.
Els canvis esdevinguts en la composició florística a les nostres latituds posen de manifest que durant el Terciari es produí un refredament lent però progressiu. Les flores de l’Eocè (fa uns 40 o 50 milions d’anys) tenien components clarament tropicals, com si es trobessin poblant zones intertropicals, cosa que s’aprecia a les flores extretes dels jaciments eocens europeus (Hordle i illa de Mall, a les illes Britàniques; Menat, Sézanne i Gelinden, a França; etc.). Durant l’Oligocè (23-35 milions d’anys), la vegetació al S d’Europa era de tipus tropical atenuada o subtropical, amb temperatures mitjanes anuals superiors en gairebé 10°C a les actuals (jaciments de Bonneville, Ais de Provença i Cèlas, a França; Monte-Promina, a Iugoslàvia; Cervera, etc.). Al Miocè (5-22,5 milions d’anys) el clima de les nostres terres era de tipus subtropical: uns 20°C de temperatura mitjana, amb un ambient sec durant el Miocè mitjà que esdevingué un clima temperat càlid a la fi del període, tal com ho indiquen flores dels jaciments de Bilin i Radoboj, a Iugoslàvia; Senegaglia, a Itàlia; Ralligen i Oeningen, a Suïssa; la Cerdanya, etc. Al Pliocè, darrer període de l’Era Terciària (2-5 milions d’anys enrere), existia un ambient temperat una mica més càlid i humit que l’actual; segons es desprèn de l’estudi de les flores dels jaciments del Papiol, la vall del Roine, Podgumer, a Bulgària; Meximieux, Val de l’Arno, etc., la vegetació durant el Pliocè era semblant a l’actual, però s’hi aprecia una mescla d’elements diversos: al costat de formes típicament temperades n’apareixen unes altres de tipus temperat càlid, que són representants subtropicals i de la vegetació de tipus mediterrani.
Les flores del Terciari del nostre continent eren molt més riques que les actuals. Nombroses espècies que vivien a Europa han desaparegut del tot. La vegetació a l’hemisferi N era molt més homogènia, amb formes que tenien una àrea de distribució àmplia, que abastava extenses zones d’Euràsia i de l’Amèrica del Nord.
En començar les glaciacions quaternàries, i a causa de la considerable disminució de les temperatures, els vegetals que no pogueren suportar aquells freds desaparegueren i moltes espècies s’aixoplugaren a les regions més càlides del S o en zones arrecerades, en les quals les temperatures els permetien de viure.
Moltes espècies desaparegueren definitivament d’Europa. Durant els principals períodes glacials, gran part de l’hemisferi N romangué cobert pels gels, bé que entre dues glaciacions existia una etapa interglacial amb temperatures més moderades.
L’especial configuració del continent europeu no afavorí el desplaçament de les espècies vers el S en començar els freds glacials, i aquestes tornaren a ocupar la seva àrea de distribució quan augmentaren les temperatures i retrocediren els gels. En efecte, les cadenes muntanyoses europees més importants es troben orientades d’E a W i, més o menys escalonadament, ocupen gran part del continent i deixen massa poc espai perquè sigui possible el desplaçament dels vegetals. D’altra banda, la mar Mediterrània tanca pel S la comunicació entre Europa i Àfrica i impedeix qualsevol mena de comunicació entre els dos continents; en una latitud en què els freds de l’època glacial ja havien eliminat les formes de tipus subtropical i temperat càlid, aquestes no tingueren cap més oportunitat de tornar a formar part de la flora europea. Consegüentment, la vegetació continental experimentà un gran empobriment: la major part de les formes càlides s’extingiren, desaparegueren gairebé totes les lauràcies i altres plantes de caràcter subtropical, mentre que d’altres sobrevisqueren tot refugiant-se a l’extrem S o a les regions caucàsiques entre la mar Negra i la mar Càspia.
En altres continents de l’hemisferi N, les glaciacions es feren sentir amb una intensitat similar, però no afectaren tant les flores: les espècies pogueren, a mesura que disminuïen les temperatures, concentrar-se a les regions del S, molt menys fredes i, un cop les condicions ambientals els foren favorables, pogueren avançar novament cap al N; això permeté que la riquesa florística es conservés molt millor que pas no al continent europeu. A l’Amèrica del Nord, les condicions són molt diferents de les que es donen a Europa: les principals serralades s’orienten de N a S i existeixen amplis passadissos que feren possible que els vegetals avancessin i retrocedissin durant els períodes glacials; a més, les dimensions d’aquesta part del continent americà són més grans, la qual cosa possibilità que les plantes poguessin assolir latituds més baixes on les temperatures eren molt més suaus. Aquesta especial configuració va permetre la preservació de la flora de l’Amèrica del Nord, molt més rica que l’europea. A l’Àsia, l’orientació de les principals cadenes muntanyoses és d’E a W, però com que el continent és molt més gran que Europa i hi ha més extensió de zones no ocupades per les serralades, els vegetals gaudiren d’un major nombre de possibilitats d’emigrar quan les condicions ambientals els foren hostils.
L’extinció d’un nombre tan elevat d’espècies a Europa donà lloc a la formació d’importants disjuncions en formes que poblaven grans extensions de l’hemisferi boreal: en desaparèixer totalment d’aquest continent, quedaren poblacions de la mateixa espècie o d’espècies molt afins separades per milers de quilòmetres. Així, per exemple, formes molt properes del gènere Platanus que poden hibridar-se (cosa que posa de manifest les seves semblances genètiques) viuen en llocs molt distants entre ells: l’Amèrica del Nord, la península balcànica i alguns punts d’Àsia. Altres gèneres presenten característiques similars, com ara Liquidambar, Magnolia, Liriodendron, Rhus, etc., les quals tenen espècies pròximes entre elles però separades per distàncies considerables.
A mesura que s’acabaren les glaciacions s’anà constituint la flora existent als nostres dies, amb un clar predomini de les angiospermes, distribuïdes en unes 300 famílies, amb més de 10 000 gèneres i un total de més de 226 000 espècies, que s’han adaptat a les condicions ecològiques i als climes del planeta, i han poblat la majoria de les regions de la Terra.
L’evolució de les flores a partir del Paleogen als Països Catalans
Hom pot començar a estudiar amb més coneixement de causa l’evolució de les flores a les nostres terres a partir del Cenozoic, ja que fins que no apareixen les angiospermes i s’expandeixen fins arribar a ser les plantes dominants en tots els ambients i els llocs de terra ferma, no es poden establir comparacions amb les flores actuals i la seva distribució al planeta. Aquestes comparacions ens donen referències de la naturalesa de les flores pretèrites, les condicions ambientals sota les quals es podien desenvolupar i el clima aproximat en què devien viure. L’estudi de les flores anteriors al Cenozoic presenta moltes dificultats d’interpretació; no hi ha garantia suficient per a afirmar que existeixen afinitats filètiques entre les flores actuals i les passades i això fa que la seva comparació no sigui del tot vàlida.
Al llarg del Cenozoic, la flora de la nostra àrea va patir una profunda transformació: passà de ser de tipus tropical, durant l’Eocè, a ser de tipus temperat a principi del Quaternari, i existiren flores de caràcter intermedi a mesura que ens acostem a les èpoques més recents. Sembla que és constant el lent però progressiu refredament; s’aprecia una tendència al seu descens lent i no es dóna un augment cap a condicions més càlides fins que no s’arriba als temps quaternaris. No sembla que passi el mateix amb la humitat ambiental, ja que ha pogut sofrir diverses alteracions, tot i que l’estudi de les flores assenyala l’existència d’un mínim d’humitat a la darreria del Miocè mitjà o bé durant els nivells inferiors del Miocè superior.
Els vegetals fòssils més antics del Cenozoic es troben a les capes eocenes que apareixen pels voltants de Manresa (Bages) (Sant Vicenç de Castellet, Vacarisses, Castellgalí, etc.). Aquesta flora presenta característiques tropicals per la presència d’espècies de Nipadites burtini i d’Acrostichum lanzaeanum, formes afins de les quals formen actualment els manglars de regions intertropicals. Les mitjanes tèrmiques anuals devien ser a la ratlla dels 25°C.
Durant l’Oligocè predominen les formes de clima càlid, amb elements tropicals i subtropicals. Abunden els vegetals perennifolis capaços de viure en ambients amb humitat i temperatura altes i amb escasses variacions estacionals. Apareixen falgueres tropicals i palmeres (Acrostichum lanzaeanum, Goniopteris stiriaca, Flabellaria haeringiana i Sabal major), i hi són freqüents les lauràcies, que atenyen un desenvolupament considerable (apareixen unes 10 espècies diferents, de les quals n’hi ha nombroses restes). S’aprecia ja una tendència a la sequera, la qual cosa sembla posar en evidència l’existència d’estacions seques. En la flora oligocena començen a adquirir una certa importància les lleguminoses, que ja tenen diverses espècies. En aquestes flores es barregen elements asiàtics amb elements americans (els seus descendents filètics viuen preferentment en aquests dos continents), i són especialment freqüents els primers.
Durant el Miocè inferior encara són freqüents les espècies tropicals i subtropicals, però disminueixen considerablement les lauràcies (s’han trobat algunes impressions foliars d’arbres d’aquesta família). Augmenten considerablement les lleguminoses, especialment les mimosàcies i les cesalpinàcies de port arbori o arbustiu; això permet d’interpretar que la flora s’adaptava a l’existència de períodes sense pluges, més o menys prolongats, seguits d’altres de pluges torrencials. També cal destacar la presència d’espècies de la família de les proteàcies, tant en el Burdigalià inferior de Catalunya com en el jaciment de Mallorca del mateix període. Aquesta flora presenta una barreja de formes que actualment tenen representants a Àsia, Àfrica i Amèrica, principalment a les regions tropicals i subtropicals.
El Miocè mitjà es caracteritza a les nostres terres per un desenvolupament extraordinari de les lleguminoses arbòries i arbustives, que atenyen la seva expansió màxima a la darreria d’aquest període (són molt freqüents les impressions de petits folíols d’aquest grup). Continua encara la preponderància dels tipus tropicals i subtropicals, però ja s’inicia la lenta penetració d’elements de caràcter temperat que, en acabar el període, predominaran en les flores europees, i queden com a relictes algunes formes de clima càlid, que desapareixeran definitivament en arribar els freds quaternaris.
Les flores de les diferents èpoques que hom ha descobert a les nostres terres són comparables a les seves contemporànies trobades en altres llocs del S d’Europa, principalment les del migdia francès i les d’algunes conques d’Itàlia. S’aprecia clarament que, en canviar les condicions ambientals, la desaparició de les formes inadaptades s’inicià pel N i la seva extinció s’anà estenent cap al S.
En el transcurs del Miocè superior, la flora sofreix un canvi força considerable, i passa a ser composta per una majoria d’elements de tipus temperat en detriment de les formes càlides, que van essent eliminades progressivament. Per exemple, les flores del Vallesià de la Cerdanya, la Seu d’Urgell (Alt Urgell) i la Bisbal (Baix Empordà), contenen un percentatge elevat d’espècies de tipus temperat, conseqüència de l’adaptació de la vegetació a les noves condicions existents a les nostres latituds (la disminució de les temperatures i l’aparició d’un ambient semblant al de les zones temperades). El percentatge de restes d’espècies de tipus temperat és més gran en les conques de la Cerdanya i la Seu d’Urgell, on constitueixen una majoria important, possiblement pel fet de tenir les condicions climatològiques més rigoroses.
La flora del Pliocè presenta característiques semblants a la del Miocè superior, ja que hi ha també una majoria d’espècies de climes temperats que, posteriorment, s’imposen en aquestes latituds. Cal destacar un nou moment d’esplendor de les lauràcies, que, difuminades i poc representatives durant gran part del Miocè, trobaren condicions més apropiades per al seu desenvolupament en el Pliocè, probablement perquè hi hagué una humitat adequada al seu desenvolupament, ja que no hi ha bases reals per a suposar que les mitjanes tèrmiques foren més elevades que en el Miocè superior; també cal tenir en compte que aquesta família no és gaire exigent pel que fa a les temperatures.
En iniciar-se el Quaternari, durant el Pretiglià, disminuïren les temperatures mitjanes i desaparegueren les lauràcies i moltes formes de tipus temperat càlid, de manera que la flora s’empobrí considerablement. L’augment de les temperatures durant el Tiglià afavorí el desenvolupament dels roures i la tornada d’espècies de climes temperats càlids, com ara Laurus canariensis, L. nobilis, Parrotia persica, etc. Els freds glacials que continuaren al llarg del Quaternari eliminaren definitivament de la nostra flora la majoria de les espècies de climes temperats càlids.