


Llevats domesticats per fer vi
-
-
- 3 of 10

Els humans hem domesticat animals i plantes, però també hem anat “domesticant” alguns microbis al llarg de la història en anar desenvolupant els aliments fermentats. Els grups principals d’aquests microorganismes són tres: els bacteris làctics de molts productes lactis i de vegetals fermentats, els llevats de les begudes alcohòliques i les floridures d’alguns formatges i derivats de la soja.
En aquest article em limitaré a parlar dels llevats de la fermentació alcohòlica del vi, o sigui Saccharomyces cerevisiae. Cal recordar que, a més dels llevats, uns altres microbis importants relacionats amb el vi són els bacteris làctics de la fermentació malolàctica, com Oenococcus oeni (Bartowsky 2008, Liu 2002).
Domesticació
Quan el flux genètic entre poblacions de la mateixa espècie es redueix, usualment per aïllament, l’evolució pot donar lloc a subpoblacions diferents, que eventualment poden esdevenir espècies diferents. Els humans poden interferir en aquest procés natural limitant el flux genètic deliberadament, seleccionant determinats trets desitjats, és a dir, fent una selecció artificial. D’aquesta manera, algunes espècies acaben essent dependents d’ambients antropogènics i adquireixen trets evolutius que interessen als humans. Això és la domesticació.
Els trets o característiques buscats amb la selecció artificial són molt diversos, des del rendiment (ramaderia bovina o ovina, agricultura intensiva, etc.), passant per la tolerància a l’estrès (collites resistents a la sequera, etc.), fins arribar a l’estètica (flors, etc.) o a la companyia (gossos, etc.).
Amb l’excepció dels gossos ja domesticats fa uns 30.000 anys, la majoria d’espècies foren domesticades durant la revolució neolítica, fa uns 12.000 anys, amb el canvi del nomadisme de caçar i recol·lectar al sedentarisme lligat als conreus i al bestiar. En seleccionar els descendents d’animals i plantes que millor cobrien les necessitats específiques, aquests humans prehistòrics efectivament van començar a controlar els processos de selecció i el flux genètic (Steensels et al. 2019).
Domesticació de microorganismes
En comparació amb la de plantes i animals, la domesticació de microbis ha estat ignorada molt de temps. Recentment, el gran augment de dades genòmiques disponibles ha permès conèixer amb precisió les relacions genètiques entre diferents microorganismes, amb la qual cosa s’ha vist que els microbis industrials sovint representen branques genètiques diferenciades i aïllades als arbres filogenètics. Tanmateix, les circumstàncies ambientals que la domesticació microbiana requereix i les característiques genètiques concretes no s’han estudiat amb detall.
Curiosament, mentre que la domesticació d’animals i plantes són sobretot el resultat de l’esforç humà voluntari, la domesticació dels microbis ha succeït sobretot sense intenció. Cal recordar que el coneixement sobre els microorganismes començà a finals del segle XVII amb Antonie van Leeuwenhoek; que el seu paper a les fermentacions no fou desvelat fins el segle XIX per Louis Pasteur i altres; i que, sobretot, els primers cultius purs —en concret, de llevats cervesers— no foren obtinguts fins a finals del segle XIX per Emil C. Hansen, fisiòleg i micòleg danès de l’empresa cervesera Carlsberg.
Però és clar, les fermentacions eren conegudes des del neolític o abans, encara que no se sabés res dels microbis com a tals. Un dels primers passos per controlar una mica el procés de fermentació fou probablement la retroinoculació (backslopping en anglès), en què s’utilitza material —i per tant microbis— d’una fermentació prèvia per començar-ne una de nova. Amb això, aquells primerencs artesans prehistòrics van promoure sense pretendre-ho l’adaptació dels microbis a l’ambient fermentatiu antropogènic, duent a terme una selecció “inconscient”.
Al llarg del procés de domesticació dels microbis, hi ha una competència contínua entre diferents variants, ja siguin espècies, soques o mutants, a cada ronda de fermentació. O sigui, també hi ha una supervivència dels més adaptats, com a la selecció natural, encara que aquí és en un ambient controlat pels humans. En canvi, això ha canviat aquests darrers decennis des que s’utilitza la inoculació de cultius purs, seleccionats prèviament d’acord amb les característiques volgudes. Aquesta selecció més “artificial” s’assembla més a la domesticació tradicional d’animals i plantes. A més, la introducció de les noves eines biotecnològiques aquests darrers anys ha permès dissenyar encara millor el microorganisme amb les propietats desitjades per a la fermentació.
Domesticació dels llevats
Els llevats són fongs unicel·lulars, i per tant microscòpics. Tenen unes 4 µm de diàmetre, encara que alguns poden arribar fins a 40 µm, i ocasionalment alguns fan prolongacions o cadenes de cèl·lules conegudes com pseudohifes. Es reprodueixen sobretot asexualment per mitosi amb gemmació, una divisió asimètrica en què el llevat fill més petit és la gemma (Figura 1). En condicions d’estrès poden esporular i després de la meiosi es produeixen espores haploides, que poden conjugar. La majoria són ascomicets però també n’hi ha de basidiomicets, i encara que hi ha unes 1.500 espècies de llevats, el llevat per antonomàsia és Saccharomyces cerevisiae.

Figura 1. Micrografia electrònica de cèl·lules de Saccharomyces cerevisiae en gemmació - Imatge de lliure ús de Wikimedia Commons, llicència de Creative Commons
Encara que no són exclusius del vi, els llevats S. cerevisiae són el model perfecte de domesticació microbiana perquè, malgrat ser eucariotes, tenen un genoma petit (12 Mb i 6.500 gens, enfront de la mitjana als ascomicets que és de 36 Mb i 11.000 gens) i ben estudiat, temps de generació curts i es disposa de moltes eines per fer-hi recerca genètica i fenotípica.
S. cerevisiae només és abundant a les fermentacions de fruita recollida artificialment, com als raïms veremats un cop són al celler. En canvi, de forma natural s’ha trobat a molts hàbitats però en nombres escassos, i en particular a les fruites, al raïm mateix, i a l’escorça dels roures, que semblen ser els seus nínxols ecològics (Steensels et al. 2019).
Per això s’ha proposat que S. cerevisiae no és que estigui adaptat a cap nínxol específic, sinó que es pot dir que és una espècie nòmada, capaç de sobreviure com a generalista poc abundant en tot un ventall d’ambients, i encara que el seu genoma és petit, és força complex (Goddard & Greig 2015).
Però malgrat que no es té clar quin és el seu hàbitat natural, està clar que el seu segon hàbitat el constitueixen diversos ambients artificials de fermentacions, com les del pa, la cervesa i el vi. S. cerevisiae és conegut per les seves excel·lents característiques fermentatives, fins i tot en presència d’oxigen. Aquest caràcter sorgí com un procés evolutiu, adquirint la capacitat de créixer en medis amb molts sucres però amb pocs dels altres nutrients, i de convertir aquests diversos sucres en etanol, un compost antimicrobià per al qual aquest llevat és molt tolerant. Com que l’etanol és un bon preservatiu natural enfront de microbis no desitjats, i addicionalment té la capacitat de ser una substància addictiva, la fermentació d’aliments per llevats va esdevenir una pràctica ben estesa, i va sorgir independentment en diverses civilitzacions arreu del món.
El vi es considera la primera beguda fermentada alcohòlica, i es calcula que té una antiguitat d’uns 9.000 anys (Fay et al. 2019). Amb tot, S. cerevisiae no és l’única espècie per produir begudes alcohòliques, vins inclosos, ja que també en produeixen altres Saccharomyces relacionats: S. uvarum, S. eubayanus, S. kudriavzevii i altres derivats híbrids (González et al. 2006).
Per estudiar el probable origen de S. cerevisiae i la relació entre les seves diverses soques, en un estudi recent de seqüenciació i anàlisi dels SNP (polimorfismes d’un sol nucleòtid) de més de 600 soques d’aquesta espècie (Duan et al. 2018), s’ha vist que les soques silvestres, de diversos continents però sobretot de la Xina, se separen quasi totes de les domesticades, excepte les aïllades en roures i altres Quercus, que són més properes a les soques pròpies de vins i de cerveses (Figura 2). Com veiem, les soques domesticades es distribueixen en dues branques evolutives. Una branca agrupa soques de fermentacions en estat sòlid, sobretot de l’Àsia i en particular de la Xina. I l’altra inclou les pròpies de fermentacions líquides com els vins i les cerveses i altres, a més de les esmentades dels Quercus. A partir de tot plegat, els autors (Duan et al. 2018) postulen un hipotètic origen de l’espècie, i sobretot de la seva domesticació, a la Xina o a l’est asiàtic en general.
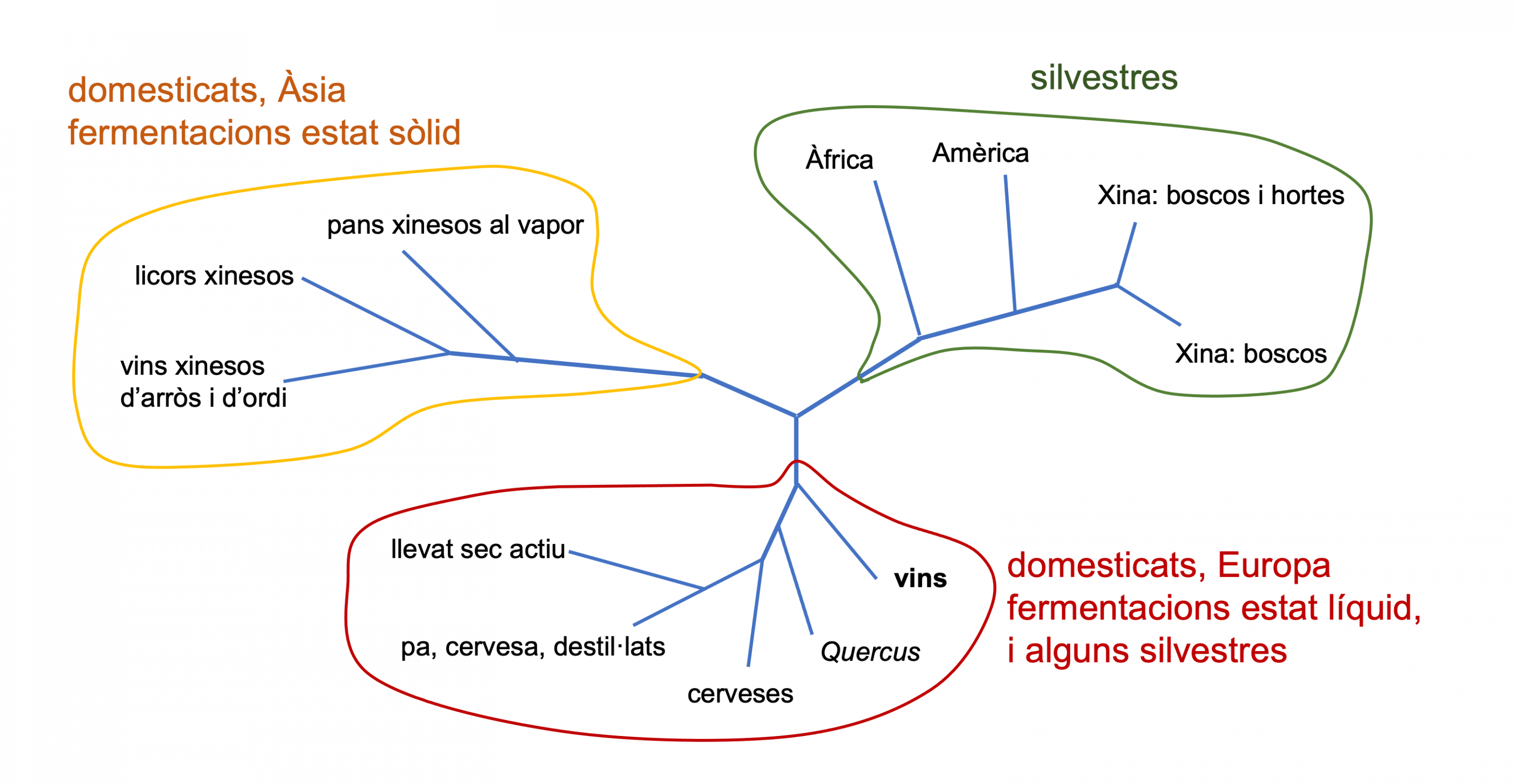
Figura 2. Esquema de l’arbre filogenètic de soques de S. cerevisiae - Gràfic adaptat de Steensels et al. 2019 i Duan et al. 2018
Aquesta proximitat evolutiva de les soques de S. cerevisiae del vi amb les dels Quercus podria ser perquè aquests arbres fossin l’origen silvestre de les víniques o també perquè hi hagués hagut un retorn a la natura d’alguna soca vínica (Steensels et al. 2019).
Les soques de llevats domesticades dels diversos processos en estat líquid, o industrials “europees”, a més de ser diferents les silvestres, s’agrupen filogenèticament força amb l’aplicació industrial, ja sigui en l’elaboració de cervesa o de vi. Les soques domesticades de S. cerevisiae tenen reforçats uns trets que les fan idònies per a l’ambient fermentatiu, com l’adaptació a concentració alta de sucres, ambients amb poc O2 i tolerància a l’etanol, però també alguns trets específics del producte.
Per exemple, els llevats cervesers poden metabolitzar maltotriosa, sucre específic del malt, mentre que els llevats vínics tenen una bona resistència als agents antimicrobians emprats, el sulfat de coure a la vinya i el sulfurós al celler. Els S. cerevisiae del vi tenen una domesticació més limitada que els cervesers. A més, els de la cervesa han perdut la capacitat de supervivència fora de l’entorn fermentatiu, mentre que els vínics poden esporular i sobreviuen a l’entorn extern. Això segurament és degut al fet que la fermentació del vi només passa un cop cada any, després de la verema, i la resta de l’any els llevats han de sobreviure al celler o a l’entorn en un ambient més natural (Steensels et al. 2019).
Cal afegir a aquestes diferències entre els llevats vínics i els cervesers, la diversitat gènica molt més gran dels llevats cervesers. De fet, molts d’aquests són S. cerevisiae, que inclouen els anomenats d’alta fermentació com els de les cerveses ale i stout i moltes més de tot el món, però justament les cerveses de baixa fermentació lager, que són el 94% del mercat mundial, són dutes a terme per S. pastorianus (sinònim S. carlsbergensis), per tant una altra espècie. Aquest S. pastorianus és un híbrid al·lotetraploide de S. cerevisiae amb el llevat criotolerant S. eubayanus, i s’originà domèsticament cap al segle XV a Baviera sotmetent les cerveses a les baixes temperatures de les coves dels Alps. Per això aquest llevat actua entre els 7 i 13ºC, i al fons del fermentador (Libkind et al. 2011).
Totes les altres cerveses d’alta fermentació són elaborades amb molt diverses S. cerevisiae i constitueixen un grup polifilètic, amb moltes branques evolutives, algunes de les quals són filogenèticament properes a les del sake, del vi i del pa. La diversitat de nucleòtids dels S. cerevisiae cervesers és més del doble que la dels vínics. Aquesta major diversitat segurament és deguda a la suara comentada utilització estacional dels llevats vínics a diferència dels cervesers que s’utilitzen tot l’any, i també als règims de selecció més intensos i diferents associats a la seva elaboració (Gonçalves et al. 2016).
Amb tot això, tenim l’aparent paradoxa que les cerveses, com que la majoria de les consumides són lager, i moltes d’elles produïdes a gran escala industrial, semblen ser més estandarditzades comercialment i per tant més uniformes, i en canvi els vins semblen ser més diversos, en contra del que acabem de veure sobre la molt major diversitat de soques de llevats cerveseres que víniques.
Canvis genètics cap a la domesticació de Saccharomyces cerevisiae del vi
Si ens centrem en el vi, cal recordar que en començar la fermentació espontània del most de raïm les soques de S. cerevisiae no són predominants i hi ha també tot un ventall d’altres llevats, coneguts com no-Saccharomyces: Hanseniaspora, Pichia, Lachancea, Metschnikowia o Torulaspora i altres gèneres (Fleet et al. 1984). Per efecte de la producció d’etanol, S. cerevisiae acaba predominant i desplaçant en nombre als no-Saccharomyces. Encara que cada cop més es van veient els beneficis d’aquests altres llevats a nivell d’aromes i es fan servir com a estàrters addicionals a S. cerevisiae (Jolly et al. 2014), no es pot parlar de la domesticació d’aquests no-Saccharomyces perquè no tenen fins ara un paper primordial en la vinificació, ja que no es pot elaborar cap vi només amb aquests altres llevats. Per tant, aquí ens limitarem a S. cerevisiae.
Molts dels mecanismes de canvis genètics que han portat a la domesticació de S. cerevisiae en la vinificació són presents també en altres espècies domesticades, tant microbianes com animals i plantes. El mecanisme més fàcil d’establir com a canvi genètic són els polimorfismes d’un sol nucleòtid o simples (SNP, sigla en anglès) deguts a mutacions puntuals d’un sol nucleòtid, però s’ha vist que aquests SNP representen tan sols una petita fracció dels mecanismes relacionats amb la domesticació de S. cerevisiae, i en canvi la majoria són rearranjaments estructurals del genoma, com els que s’explicaran a continuació.
Les variacions en el nombre de còpies (CNV, sigla en anglès) són mutacions molt freqüents en molts organismes diversos, en què fragments de DNA de >1kb són repetits, de manera que tenen diverses còpies, en nombre variable. El cas més conegut en els llevats vínics és el gen CUP1, que codifica per a una metal·lotioneïna, proteïna captadora de coure. Algunes soques poden tenir fins a 18 còpies d’aquest gen, amb el qual aquests S. cerevisiae vínics produeixen més quantitat de la proteïna i, per tant, són més tolerants al coure de la “barreja de Bordeus” usada com a fungicida a les vinyes (Steensels et al. 2019).
Els rearranjaments cromosòmics, amb delecions, insercions o translocacions de grans fragments de cromosomes, sovint causades per transposons, són relativament freqüents en l’evolució dels llevats vínics i de Saccharomyces en general. De fet, aquests rearranjaments que poden afectar un nombre important de gens són una causa important d’especiació. Un exemple n’és la tolerància al sulfit —l’antibacterià més usat en vinificació— degut a l’al·lel SSU1-R, que és el producte sobreexpressat d’una translocació entre els cromosomes 8 i 16, només present als llevats vínics (Pérez-Ortín et al. 2002).
La hibridació interespecífica és un altre mecanisme de canvis que ja hem vist com a origen dels llevats domesticats a les cerveses lager. Les espècies de Saccharomyces poden hibridar molt fàcilment entre elles ja que aquests híbrids poden propagar-se per mitosi tan eficientment com els seus progenitors, encara que rarament poden fer la meiosi i, per tant, quasi mai produeixen espores viables (Sipiczki 2008). Malgrat això, alguns dels llinatges híbrids poden acabar sent reconeguts com a espècies. Els casos més coneguts són l’esmentat S. pastorianus (sinònim de S. carlsbergensis, encreuament de S. cerevisiae amb S. eubayanus) de les lager, i els S. bayanus (S. cerevisiae x S. eubayanus x S. uvarum) del vi. Tanmateix, S. bayanus és controvertit per les seves similituds amb S. cerevisiae i genera confusió de noms d’espècies amb alguna de les soques comercials més utilitzades (Sipiczki 2008).
Els híbrids també han estat clau en l’aparició de força llevats vínics, sobretot amb relació a la possibilitat de fermentar a baixes temperatures. Per exemple, algunes de les soques comercials d’estàrters que fermenten vi a 15ºC són híbrids “naturals” de S. cerevisiae i S. kudriavzevii amb poliploïdia parcial, aïllats el 1979, que tenen la bona producció d’etanol de S. cerevisiae i la tolerància al fred de S. kudriavzevii (Erny et al. 2012).
De fet, recentment la hibridació interespecífica de diversos Saccharomyces vínics —per exemple S. cerevisiae x S. uvarum— s’està duent a terme als laboratoris per tal d’aconseguir soques amb millors característiques relacionades amb caràcters poligènics. La hibridació com a mètode té l’avantatge de no ser considerada una tècnica GMO ( sigla en anglès d’organisme modificat genèticament) i, per tant, les soques poden ser transferides ràpidament a la indústria (García-Ríos et al. 2019).
La transferència genètica horitzontal (TGH) és un altre mecanisme molt important per a l’evolució en molts organismes, i en els llevats vínics també se’n coneixen alguns casos en els quals la TGH ha contribuït a la seva domesticació. Un cas és el gen FSY1, codificant del transportador de fructosa, que permet a S. cerevisiae la utilització de la fructosa al final de fermentació, després d’haver consumit la glucosa. És molt probable que aquest gen li fou transferit del similar de S. pastorianus, amb el qual presenta molta homologia (Galeote et al. 2010). Un altre cas demostrat de TGH és el del gen FOT originari d’un altre llevat, Torulaspora microellipsoides, que li permet a S. cerevisiae una millor incorporació d’oligopèptids, que li donen avantatge competitiu quan les fonts de nitrogen són escasses (Marsit et al. 2015).
Conclusió
El llevat Saccharomyces cerevisiae és l’espècie principal implicada en la fermentació alcohòlica del vi. En canvi, abans de la fermentació quasi no hi és. Si no s’inoculen estàrters, el nombre de S. cerevisiae són molt baixos al most. Per tant, es pot dir que si els humans no haguessin començat a fer vi —i cervesa— fa alguns milers d’anys, aquesta espècie segurament no existiria. És clarament un microbi domesticat en el sentit que ha aparegut evolutivament en un temps relativament breu com a derivat de microbis previs que no tenien les característiques específiques d’aquest llevat.
Com hem vist, l’excepcionalitat de Saccharomyces (“el fong del sucre”), i en particular de S. cerevisiae, és sobretot la capacitat de fermentar medis amb molts sucres, convertint-los en quantitats importants en etanol, per al qual aquest llevat és molt tolerant. Aquesta producció d’etanol és bioquímicament excepcional i única entre els fongs i els altres organismes. Vegeu pel que fa a això el treball de Dashko et al. (2014) sobre l’evolució de la fermentació alcohòlica als llevats.
Bibliografia
- Bartowsky, EJ. (2008): “Oenococcus oeni and malolactic fermentation —moving into the molecular arena”. Aust J Grape Wine Res 11: 174-187
- Dashko, S.; Zhou, N.; Compagno, C.; Piškur, J. (2014): “Why, when, and how did yeast evolve alcoholic fermentation?”. FEMS Yeast Res, 14, 826-832
- Duan, S.F.; Han, P.J.; Wang, Q.M.; Liu, W.Q.; Shi, J.Y. et al. (2018): “The origin and adaptive evolution of domesticated populations of yeast from Far East Asia”. Nat. Commun. 9, 2690
- Erny, C.; Raoult, P.; Alais, A.; Butterlin, G.; Delobel, P. et al. (2012): “Ecological Success of a Group of Saccharomyces cerevisiae/Saccharomyces kudriavzevii Hybrids in the Northern European Wine-Making Environment”. Appl Environ Microbiol 78, 9
- Fay, J.C.; Liu, P.; Ong, G.T.; Dunham, M.J.; Cromie, G.A. et al. (2019): “A polyploid admixed origin of beer yeasts derived from European and Asian wine populations”. PLoS Biol 17(3): e3000147
- Fleet, G.H.; Lafon-Lafourcade, S.; Ribereau-Gayon, P. (1984): “Evolution of yeasts and lactic acid bacteria during fermentation and storage of Bordeaux wines”. Appl Environ Microbiol 48, 1034-1038
- Galeote, V.; Novo, M.; Salema-Oom, M.; Brion, C.; Valerio, E. et al. (2010): “FSY1, a horizontally transferred gene in the Saccharomyces cerevisiae EC1118 wine yeast strain, encodes a high-affinity fructose/H+ symporter”. Microbiology 156, 3754-3761
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V. et al. (2016): “Domestication and divergence of Saccharomyces cerevisiae beer yeasts”. Cell 166, 1397-1410.e1316
- García-Ríos, E.; Guillén, A.; de la Cerda, R.; Pérez-Través, L.; Querol, A.; Guillamón, J.M. (2019): “Improving the Cryotolerance of Wine Yeast by Interspecific Hybridization in the Genus Saccharomyces”. Front Microbiol 9, 3232
- Goddard, M.R.; Greig, D. (2015): “Saccharomyces cerevisiae: a nomadic yeast with no niche?”. FEMS Yeast Res. 15, 1–6
- Gonçalves, M.; Pontes, A.; Almeida, P.; Barbosa, R.; Serra, M. et al. (2016): “Distinct Domestication Trajectories in Top-Fermenting Beer Yeasts and Wine Yeasts”. Current Biology 26, 2750-2761
- González, S.S.; Barrio, E.; Gafner, J.; Querol, A. (2006): “Natural hybrids from Saccharomyces cerevisiae, S. bayanus and S. kudriavzevii in wine fermentations”. FEMS Yeast Res 6, 1221-1234
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J. et al. (2011): “Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast”. Proceedings of the National Academy of Sciences, 108(35), 14539-14544
- Liu, S.Q. (2002): “A review: Malolactic fermentation in wine — Beyond deacidification”. J Appl Microbiol 92, 589-601
- Jolly, N.P.; Varela, C.; Pretorius, I.S. (2014): “Not your ordinary yeast: non-Saccharomyce yeasts in wine production uncovered”. FEMS Yeast Res, 14, 215-237
- Marsit, S.; Mena, A.; Bigey, F.; Sauvage, F.; Couloux, A. et al. (2015): “Evolutionary advantage conferred by an eukaryote-to-eukaryote gene transfer event in wine yeasts”. Mol Biol Evol 32, 1695-1707
- Padilla, B.; Gil, J.V.; Manzanares, P. (2016): “Past and future of non-Saccharomyces yeasts: from spoilage microorganisms to biotechnological tools for improving wine aroma complexity”. Front. Microbiol. 7, 411
- Pérez-Ortı́n, J.E.; Querol, A.; Puig, S.; Barrio, E. (2002): “Molecular Characterization of a Chromosomal Rearrangement Involved in the Adaptive Evolution of Yeast Strains”. Genome Res 12, 1533-1539
- Steensels, J.; Gallone, B.; Voordeckers, K.; Verstrepen, K.J. (2019): “Domestication of industrial microbes”. Curr Biol 29 R381 (Review)
- Sipiczki, M. (2008): “Interspecies hybridization and recombination in Saccharomyces wine yeasts”. FEMS Yeast Res 8, 996-1007
NOTA:
Adaptació d’una part de l’article “Microbis domesticats per fer vi”, publicat el 24 d’agost de 2022 al blog “Bios i altres” d’Albert Bordons
