Els acantocèfals són animals vermiformes paràsits que, en estat adult, viuen sempre en l’intestí de vertebrats. El seu estudi és d’un gran interès pel fet que són agents causants de malalties d’animals tant silvestres com domèstics, i també, més rarament, de l’home. La seva morfologia reflecteix una adaptació profunda a la vida parasitària i al seu microhàbitat entèric; destaca, sobretot, la poca extensió dels sistemes muscular, nerviós, circulatori i excretor, com també la pèrdua completa del sistema digestiu. L’organisme no és, doncs, sinó un sac pseudocelomàtic d’òrgans reproductors proveït d’una estructura espinosa d’anclatge en un extrem. Generalment de color blanc o crem, o lleugerament groc o ataronjat, i de cos allargat, entre cilíndric i lleugerament aplatat, amb nombrosos solcs transversals, són verms de dimensions compreses entre 1 mm escàs i més d’1 m, segons les espècies. Són organismes dioics amb un cert dimorfisme sexual: en general les femelles són una mica més grans que els mascles. Dins els anomenats helmints o verms paràsits, que comprenen una part dels platihelmints (trematodes, monogenis i cestodes), els nematodes i els acantocèfals, aquests són, de molt, el grup amb un nombre més petit d’espècies (aproximadament unes 1000) com també el grup menys abundant.
L’interès principal dels acantocèfals es deu al fet que constitueixen agents causals de malalties (desencadenades essencialment pel mal que produeix la seva trompa en penetrar en la mucosa intestinal) en diversos animals domèstics i en l’home. En l’home han estat trobades cinc espècies paràsites: Macracanthorhynchus hirudinaceus (pròpia del porc), Moniliformis moniliformis (de rosegadors), Acanthocephalus rauschi (de peixos), Corynosoma strumosum (de foques) i Acanthocephalus bufonis (de gripaus). Cal no oblidar, a més, la seva transcendència com a paràsits d’animals silvestres o en captivitat, ja que s’han descrit epizoòties fulminants capaces de matar un gran nombre d’animals en un espai curt de temps.
Les escasses citacions d’espècies d’acantocèfals als Països Catalans són, sens dubte, conseqüència dels escassos estudis que hom hi ha dedicat. Aquestes citacions sempre s’han fet en ocasió d’estudis helmintofaunístics de grups o d’espècies d’hostes determinats. Així, al nostre país se’n coneixen fins ara únicament sis espècies.
Morfologia
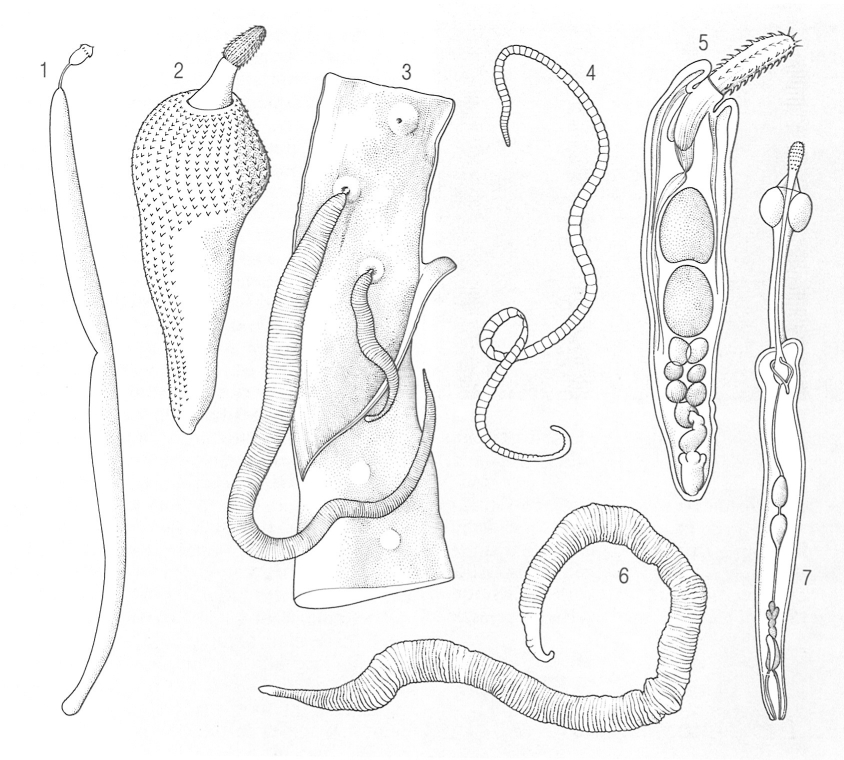
Diferents tipus morfològics dels acantocèfals: 1 Bolbosoma, 2 Corynosoma, 3 Macracanthorhynchus hirudinaceus (una femella, més gran, i un mascle, adherits a la paret de l’intestí d’un porc), 4 Moniliformis, 5 Acanthocephalus, 6 Hamanniella microcephala, 7 Pomphorhynchus bulbocolli.
Biopunt, a partir de fonts diverses
Externament, el cos d’un acantocèfal consta de dues regions diferents: la regió anterior o prosoma, que comprèn la probóscide o trompa i el coll, i que és responsable de la fixació del paràsit a la mucosa intestinal; i la regió posterior o metasoma, que comprèn el tronc i és la responsable fonamentalment de les funcions reproductores.
El tronc és la part més llarga del cos, i en moltes espècies també presenta espines externes simples, però només en aquelles parts que mantenen contacte amb la mucosa intestinal de l’hoste. El tegument que el recobreix, com en el cas de la trompa i el coll, comprèn diverses capes musculars internes. Internament, inclou el pseudoceloma o cavitat corporal dins la qual hi ha els sacs lligamentaris, els òrgans sexuals, els del sistema excretor (quan n’hi ha) i, en els mascles, els ganglis genitals. Pel tronc es fa, a més, l’absorció i la distribució de nutrients del contingut intestinal de l’hoste.

Detall de la probòscide de l’acantocèfal Centrorhynchus appendiculatus, paràsit de la musaranya comuna (Crocidura russula). L’exemplar fotografiat (mitjançant un microscopi d’interferència), en estat larval, fou extret dels budells d’una musaranya estudiada a Eivissa.
Santiago Mas-Coma.

Diferents tipus de probòscide en els acantocèfals. Noteu l’estructura espinosa, especial per a anclar l’animal a les parets entèriques de l’hoste.
Biopunt
La probóscide o trompa, que adopta formes diferents segons les espècies, des d’esfèriques a cilíndriques, és recoberta per un tegument i una paret muscular prima, en la qual s’implanten les rels d’uns ganxos esclerotitzats i corbs. La forma, les dimensions i el nombre de ganxos constitueixen dades d’interès taxonòmic. Per dins, aquesta trompa és plena de líquid. Inserits al seu àpex intern hi ha un parell de músculs (músculs inversors de la probòscide) que s’estenen al llarg de la trompa i del coll i van a fixar-se a la paret d’un sac muscular (receptacle de la probòscide) que és fixat a la paret interna de la trompa. La forma d’aquest sac també varia molt segons la família, però en general consisteix en una o dues capes de fibres musculars. Quan els músculs inversors de la trompa es contreuen, la probòscide s’invagina dins un receptacle i els ganxos queden a dins; quan el receptacle es contreu la trompa es veu obligada a evaginar-se per un sistema hidràulic. En el receptacle hi ha un gangli nerviós (gangli cerebral o cervell).
El coll és una zona llisa, sense espines, situada entre els ganxos posteriors de la trompa i un replec de la paret del cos; uns músculs retractors fixen aquest replec a la superfície interna del coll. En algunes espècies hi ha, a més, uns músculs protrusors que es fixen al receptacle de la trompa. Quan els músculs retractors de la trompa i els del coll es contreuen, tota la part anterior de l’animal es retreu dins el tronc. En algunes espècies hi ha una fosseta sensorial a cada costat del coll, mentre que en moltes d’altres hi ha dues fossetes similars a la punta de la trompa.
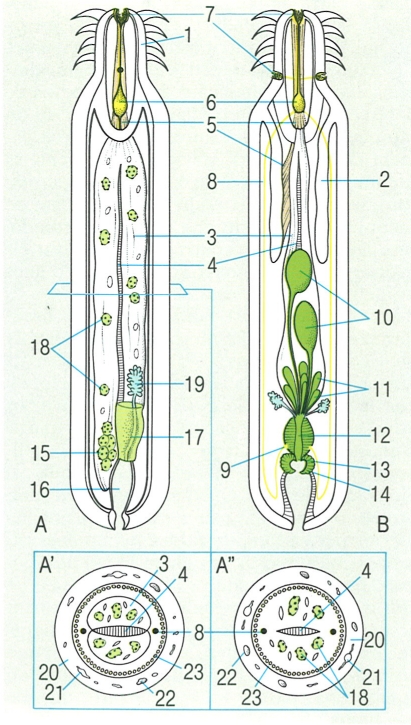
Morfologia interna dels acantocèfals: A femella, en visió lateral, i detalls del tall transversal d’una femella d’arquiacantocèfal (A′) i de paleoacantocèfal (A″). B Esquema de l’organització interna d’un mascle de paleoacantocèfal, vist per la cara dorsal. Hom ha indicat en el dibuix: 1 trompa o probòscide armada de ganxos, 2 lemnisc, 3 sac lligamentari, 4 cordó lligamentari, 5 múscul retractor, 6 encèfal, 7 papil·les sensorials, 8 nervi lateral, 9 gangli, 10 testicles, 11 glàndules del ciment, 12 conducte ejaculador, 13 penis, 14 bossa copuladora evaginable. amb orifici genital extern, 15 ovari, 16 úter, 17 campana de l’úter, 18 ous. 19 protonefridis, 20 epidermis, 21 llacunes del sinciti epidèrmic, 22 nucli cel·lular, 23 capa muscular.
Biopunt, a partir de fonts diverses
La paret del cos és un síncit complex que conté elements nuclears i una sèrie de canals interconnectats i reblerts de líquid, que constitueix l’anomenat sistema lacunar. En algunes espècies, aquests nuclis són molt escassos però gegantins, mentre que en d’altres, els nuclis es fragmenten durant el desenvolupament larval i es distribueixen en gran nombre per tota la paret del tronc. Quan els nuclis són sencers, mantenen un nombre constant en cada espècie. El tegument no té veritables capes diferenciades, però hom hi pot distingir, de fora a dins, diferents regions: una capa superficial, una zona ratllada, una zona vesicular, una zona de feltre, una zona de fibres radials i una làmina basal. Internament, després del tegument, hi ha una capa de teixit connectiu irregular, seguida de diverses capes de músculs circulars i longitudinals. El sistema lacunar, de funció encara no ben coneguda (hom creu que pot actuar com a sistema circulatori), consta de dues parts, una de presomàtica i una altra de metasomàtica, aparentment sense connectar. El sistema lacunar presomàtic és constituït per canals que desemboquen en dues estructures de funció desconeguda, els lemniscs, que creixen de la base del coll vers el pseudoceloma; cada lemnisc presenta un canal central que és continuació del sistema lacunar presomàtic. El sistema lacunar metasomàtic consisteix en una complicada xarxa de canals interconnectats, bé que en la majoria d’espècies se’n distingeixen dos de longitudinals principals (dorsal i ventral o bé laterals), connectats amb nombrosos canalets transversals irregulars; d’altres tenen, a més, un parell de canals longitudinals de posició mitjana. Els músculs de la paret del cos formen una capa longitudinal envoltada externament per una altra capa de músculs circulars, d’estructura i activitat peculiars: l’obertura que deixen aquests músculs és connectada amb el sistema lacunar, de tal manera que la circulació del fluid lacunar hi porta nutrients i elimina els materials residuals dels músculs. Malgrat que no hi ha ni cor ni cap altre òrgan circulatori, la contracció dels músculs circulars fa anar el fluid cap als components longitudinals, i viceversa.
L’excreció, en la majoria de les espècies, sembla que s’efectua per difusió a través de la paret corporal. De tota manera, els membres d’una família (oligacantorínquids) presenten dos òrgans excretors protonefridials, cadascun dels quals comprèn diversos bulbs anucleats, en forma de flama, portadors de flocs de cilis, que poden ésser capsulats o no segons cada espècie; en els mascles, aquests òrgans es fixen al vas deferent i s’hi buiden, mentre que en les femelles són fixats a la campana uterina i es buiden a l’úter. En general, els acantocèfals tenen poca capacitat per a l’osmoregulació: s’inflen fins a fer-se cilíndrics i turgents en solucions hipotòniques, salines o de sacarosa, i es tornen flàccids, plans i arrugats en solucions hipertòniques. La tonicitat del seu líquid pseudocelomàtic és semblant o lleugerament superior a la del contingut intestinal de l’hoste.
El sistema nerviós és simple. En les espècies estudiades, el gangli cerebral, situat en el receptacle de la trompa, és constituït tan sols per de 50 a 94 cèl·lules. D’aquest gangli surten relativament pocs nervis, els més grans dels quals són el nervi anterior de la trompa i els nervis posteriors laterals. Els nervis innerven els dos òrgans sensorials laterals i l’òrgan sensorial apical, quan n’hi ha. Ventralment i una mica per davant del gangli cerebral hi ha una gran cèl·lula multinucleada (cèl·lula de suport), de la qual surten cap als òrgans sensorials laterals i apicals unes formacions no nervioses, de funció desconeguda, potser secretora.
Pel que fa al sistema reproductor, tant els mascles com les femelles tenen un o dos sacs lligamentaris prims, fixats a l’extrem posterior del receptacle de la trompa, que s’estenen fins a arribar prop del porus genital distal. Dins aquests sacs hi ha les gònades i alguns òrgans accessoris dels sistemes reproductors. En algunes espècies, els sacs lligamentaris són permanents, mentre que en d’altres es destrueixen a mesura que el verm esdevé madur. Els mascles presenten, en general, dos testicles, si bé s’ha descrit l’existència d’individus monòrquids (amb un sol testicle) en algunes espècies. La localització i les dimensions dels testicles són força constants per a cada espècie. Cada testicle té un conducte eferent a través del qual els espermatozoides madurs, en forma de fils prims i sense cap, migren fins al conducte deferent comú i/o fins a un petit penis. Hi ha també diversos òrgans accessoris, entre els quals cal destacar les glàndules del cement (d’un a vuit òrgans sincítics que contenen un o més nuclis gegants o diversos fragments nuclears), que en moltes espècies són unides per ponts prims i que secreten un cement copulatori de naturalesa proteica (proteïna adobada) que de vegades queda emmagatzemat en un receptacle especial fins al moment de la còpula. Un altre òrgan sexual accessori del mascle és la bossa copuladora, una part especialitzada de la paret distal del cos, proveïda de papil·les sensorials, que roman invaginada dins l’extrem posterior del celoma, excepte durant la còpula. Enganxat a la base d’aquesta bossa hi ha un sac muscular anomenat bossa de Saefftigen, que, quan es contreu, força el fluid cap al sistema llacunar de la bossa copuladora i, per pressió hidrostàtica, fa que s’evagini. Les femelles presenten un ovari molt peculiar perquè es fragmenta en boles ovàriques molt aviat, generalment quan el verm no és més que un estadi jove dins l’hoste intermediari. Aquestes boles d’oogonis floten lliurement dins el sac lligamentari i creixen lleugerament de volum abans de la inseminació. L’extrem posterior del sac lligamentari es troba fixat a una campana uterina muscular, que, d’una banda, permet que els ous madurs passin a l’úter i la vagina i arribin a l’exterior a través gonòpor, i, de l’altra, evita que hi passin els ous immaturs, els quals retorna al sac lligamentari.
Biologia
Reproducció i desenvolupament
La reproducció dels acantocèfals parteix d’una còpula: quan l’extrem posterior del mascle entra en contacte amb el de la femella, la bossa copuladora s’evagina i l’abraça per mitjà de contraccions musculars, mentre s’efectua la transferència de l’esperma; després, el cement copulador obtura la vagina de la femella i s’endureix ràpidament fent-hi un tap que hi romandrà durant el desenvolupament dels embrions, fins que, finalment, es desintegri. Després de la còpula, els espermatozoides emigren des de la vagina, a través de l’úter i la campana uterina, fins a l’interior del sac lligamentari, on comencen a fertilitzar els ovòcits de les boles ovàriques; després d’unes primeres divisions, escasses, els embrions es desprenen de la bola ovàrica i queden surant lliurement en el líquid pseudocelomàtic, la qual cosa permet que els ovòcits subjacents quedin exposats a la fertilització. Per això, en una mateixa femella podem trobar diversos estadis de l’embriogènesi inicial i, a partir d’una sola còpula, arriben a produir-se i a alliberar-se uns quants milers, fins i tot milions, d’ous embrionats. Quan els embrions amb protecció són empesos endins de la campana uterina per l’acció peristàltica, tenen dues rutes possibles: passar novament al pseudoceloma a través de fenedures de la campana o bé entrar en l’úter. Els embrions completament desenvolupats són lleugerament més llargs que els immaturs i, en conseqüència, com que no poden passar a través de les fenedures de la campana, passen a l’úter, mentre que els ous immaturs són retinguts. Els ous embrionats madurs surten pel porus genital, cauen al lumen de l’intestí i surten al medi extern amb els excrements de l’hoste definitiu. Hi ha dos tipus d’ous, que es corresponen a un cicle biològic aquàtic o a un cicle terrestre; en aquest darrer cas, la coberta de l’ou és més gruixuda.
La larva completament embrionada, o acàntor, que infestarà l’hoste intermediari (un artròpode), és un organisme allargat i usualment armat a l’extrem anterior amb de 6 a 8 ganxos en forma de fulles de navalla o bé amb petites espines (òrgan aclide o rostel); l’acàntor, tancat dins l’ou, constitueix un estat latent de resistència que només continuarà el seu desenvolupament quan arribi a l’hoste intermediari. Efectivament, quan un artròpode ingereix l’ou, el verm es converteix en un paràsit, que creix i absorbeix nutrients, i inicia així l’estadi de desenvolupament conegut amb el nom d’acantel·la: en algunes espècies, l’extrem anterior de l’acàntor (que porta l’òrgan aclide) es transforma aparentment a l’extrem anterior de l’adult, mentre que en d’altres sembla que hi ha un curiós canvi de polaritat de 90 graus, de manera que l’extrem anterior de l’adult prové de la part lateral de l’acàntor. Durant l’estat d’acantel·la, els sistemes orgànics es desenvolupen a partir de la massa nuclear central i dels nuclis hipodèrmics de l’acàntor. Al final d’aquest període de desenvolupament s’arriba a l’estadi jove, en què, amb totes les estructures de l’adult, ja és infestant i rep el nom de cistacant. En la majoria de les espècies, els extrems anterior i posterior s’invaginen i tot el cistacant queda englobat dins un embolcall hialí molt prim. En aquest moment, el paràsit ha d’accedir al seu hoste definitiu per via oral (depredació de l’hoste intermediari) per poder seguir el seu desenvolupament i donar lloc als mascles i femelles adults, un cop a l’intestí, procés pel qual no hi ha cap mena de migració intraorgànica complexa.
Cicle biològic
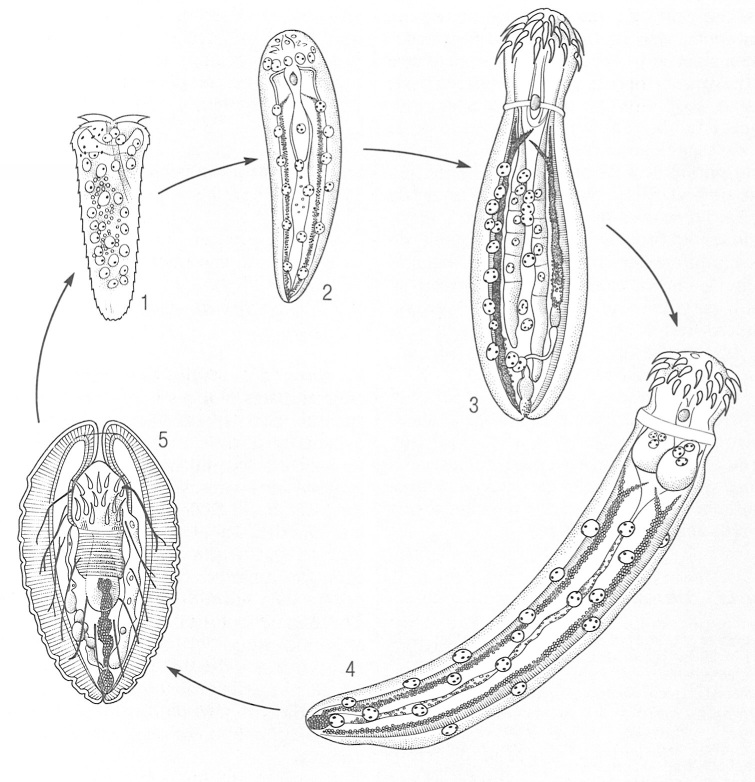
Cicle biològic de l’acantocèfal Macracanthorhynchus hirudinaceus. 1 Larva acantor, 2, 3 estadis intermedis de desenvolupament, dins l’hoste intermediari, 4 acantel·la (tambe dins l’hoste intermediari), 5 cistacant infestant, amb la probóscide invaginada.
Biopunt, a partir de fonts diverses
La vida dels acantocèfals adults dura entre unes setmanes i uns mesos, segons cada espècie. El seu cicle biològic és diheteroxè, és a dir, hi intervenen solament dos hostes: un artròpode, que fa d’intermediari, i un vertebrat, que actua d’hoste definitiu. Els adults viuen en l’intestí de peixos, ambifis, rèptils (rarament), ocells i mamífers, i, en general, les espècies mostren poca especificitat d’hoste (per exemple, una espècie pot infestar i desenvolupar-se en exemplars de peixos de diferents famílies, però no en ocells o mamífers, i viceversa), però, contràriament, sí que mostren una gran especificitat pel microhàbitat: sempre es troben en el mateix tram del tracte digestiu, i només amplien el seu camp d’infecció quan formen una població molt densa.
Els acantocèfals són polígams, i el nombre de còpules depèn de la densitat poblacional. La femella deixa anar els ous embrionats madurs, que contenen l’acàntor (que ja és infestant per a l’hoste intermediari) dins l’intestí del seu hoste vertebrat, de manera que surten a l’exterior amb els excrements. El període prepotent (temps entre el moment de la infestació i l’eliminació dels primers ous per l’hoste) oscil·la entre 22 i 70 dies en les espècies estudiades fins ara. La supervivència i el manteniment del poder d’infestació de l’acàntor depèn de factors abiòtics (temperatura, humitat i dessecació), però són més resistents els ous d’espècies de cicle terrestre que no les de cicle aquàtic. En condicions ambientals normals, l’acàntor pot arribar a romandre viable en el medi extern (sòl, aigua) durant setmanes, mesos o anys, segons cada espècie.
L’hoste intermediari s’infesta en ingerir l’ou que conté l’acàntor viable. En acantocèfals de cicle terrestre sempre es tracta d’un artròpode, i gairebé sempre d’un insecte (generalment un coleòpter o un ortòpter), rarament un miriàpode, i si són de cicle aquàtic es tracta d’un microcrustaci (generalment amfípodes, copèpodes, isòpodes o ostràcodes). L’especificitat per l’hoste intermediari és molt marcada, però s’ha observat que hi ha un factor geogràfic important, de tal manera que l’acantocèfal tria sempre la mateixa espècie d’hoste intermediari dins un àmbit geogràfic reduït, encara que al llarg de tota la seva àrea de dispersió n’utilitzi diverses. D’altra banda, una mateixa espècie d’acantocèfal pot tenir diverses soques o races específiques per a espècies determinades d’hostes intermediaris. Una vegada dins l’hoste intermediari, l’acantor eclosiona en el canal alimentari i, ajudantse dels ganxos de l’òrgan aclide penetra en la paret intestinal. En algunes espècies arriba a travesar completament l’intestí i a instal·lar-se a l’hemocel de l’hoste, però en d’altres es para justament sota la serosa; en tots dos casos, s’inicia a continuació la fase d’acantel·la, que es desenvolupa fins a atènyer l’estadi infestant de cistacant. El cicle es tanca quan l’hoste intermediari que allotja el cistacant és depredat per un vertebrat, que actua d’hoste definitiu, ja que en ell es desenvoluparà l’adult.
En el cicle biològic d’algunes espècies, tant de cicle aquàtic com terrestre, pot entrar-hi un altre hoste, entre l’intermediari i el definitiu: I’hoste d’espera o paratènic. En general és un vertebrat (molt rarament un mol·lusc o un anèl·lid), i el cistacant travessa llur paret intestinal com si fos l’hoste definitiu; aleshores el cistacant s’instal·la en els mesenteris o òrgans viscerals, on procedeix a reencistar-se, de manera que el desenvolupament del verm es deté. Aquest estadi de latència en l’hoste paratènic sol anomenar-se juvenil.
Des del punt de vista biològic, doncs, l’hoste paratènic és innecessari per a tancar el cicle, però augmenta la capacitat de l’espècie tant pel que fa a la seva disseminació en l’espai com a la seva dispersió per hostes de nivells tròfics diferents o d’espècies diferents d’un mateix nivell tròfic. I hom pensa que, en molts casos, la intervenció de l’hoste paratènic s’ha de considerar imprescindible per a assegurar la infestació de l’hoste definitiu, especialment quan aquest pertany a grups de vertebrats que ocupen un nivell molt alt en la cadena trófica (cetacis, pinnípedes, carnívors, ocells marins ictiòfags, rapinyaires), i que rarament consumeixen artròpodes. D’altra banda, cal tenir present que els hostes paratènics poden actuar de vegades alhora d’hostes definitius: s’ha vist que una mateixa espècie d’hoste, fins i tot un mateix individu, pot arribar a acollir cistacants encistats extraintestinals i també adults intraintestinals de la mateixa espècie, de manera que fa alhora d’hoste paratènic i d’hoste definitiu per a un mateix paràsit. Els cistacants procedeixen de la ingestió d’acantel·les no infestants, mentre que els adults procedeixen de l’adquisició de cistacants infestants. En el cas de la ingestió d’acantel·les no infestants es podria parlar en realitat més d’hoste intermediari que no d’hoste paratènic, ja que sí que hi ha un cert desenvolupament fins a arribar a atènyer l’estadi infestant de cistacant.
La distribució geogràfica dels acantocèfals es condicionada per la dels seus hostes, i depèn del grau d’especificitat per l’intermediari i pel definitiu que mostra cada espècie. Hi ha representants d’aquest grup de paràsits a tots els continents, oceans i mars.
Filogènia i sistemàtica
Les primeres citacions d’acantocèfals daten del segle XVII, i la 13ena edició del «Systema Naturae» (1788 i 1793) ja en recull gèneres i espècies. El seu nom, Acanthocephala (del grec «akantho», espinós, «kephala», cap), fou proposat per Rudolphi al començament del segle XIX (1802, 1809) com un nou ordre. Més tard (1931-33) Meyer els proposà com una classe dins els asquihelmints i ja hi distingí els ordres dels paleacantocèfals (Palaeacanthocephala) i els arquiacantocèfals (Archiacanthocephala). Finalment, van Cleave (1941-1948) elevà el grup a la categoria de fílum. La base del sistema de Meyer i van Cleave és reconeguda unànimement avui en dia, i hom hi accepta tres classes diferents: els arquiacantocèfals, els paleacantocèfals i els eoacantocèfals. Fins a l’actualitat no s’ha trobat cap representant d’aquesta darrera classe al nostre país.
La classe dels eoacantocèfals (Eoacanthocephala) comprèn paràsits de peixos, amfibis i rèptils. Es caracteritzen pels canals lacunars longitudinals; nuclis hipodèrmics escassos, gegants i de vegades ameboides; el receptacle de la trompa, de paret simple; el múscul retractor de la trompa, que travessa l’extrem posterior del receptacle; el cervell, prop de la part anterior o mitjana del receptacle; els nuclis dels lemniscs, escassos i gegants; dos sacs lligamentaris persistents en la femella; la glàndula cementària única, sincitial, amb diversos nuclis i receptacle de cement annex, i ous de forma variable. Inclou els ordres dels giracantocefàlides (Gyracanthocephalida) i els neoequinorínquides (Neoechinorhynchida).
Hom sol considerar els acantocèfals relacionats amb un o més grups d’asquihelmints, especialment amb els rotífers i els priàpuls. Descobriments fòssils recents, datats del Cambrià (d’uns 530 milions d’anys d’antiguitat), en una fauna marina que vivia en sediments situats a 100 m de fondària han permès de descriure Ancalagon minor, un protoacantocèfal de vida lliure aparentment excavador i amb afinitats amb els priàpuls. Tot sembla indicar que cal cercar, doncs, l’origen dels acantocèfals paràsits molt enrere en el temps, en aquelles formes ancestrals de vida lliure que constituïen part de la dieta d’artròpodes sedimentaris, probablement crustacis primitius que a la llarga es convertirien en els seus primers i únics hostes. L’aparició de vertebrats depredadors d’aquests artròpodes pot haver marcat una possible colonització ulterior d’aquests vertebrats i, amb això, el pas d’un cicle monoxè a un cicle heteroxè, com en l’actualitat. La filogènia dels acantocèfals encara és força especulativa, bé que tot sembla indicar que de les tres classes que comprenen, els paleacantocèfals, majoritàriament paràsits de crustacis aquàtics, peixos teleostis i llamprees, són els més primitius.
Els grups d’acantocèfals
La classe dels arquiacantocèfals inclou espècies paràsites d’ocells i de mamífers, caracteritzades pels canals lacunars longitudinals, principals dorsals i ventrals, o simplement dorsals, els pocs nuclis hipodèrmics, els nuclis gegants en lemniscs i les glàndules cementàries, els dos sacs lligamentaris persistents en les femelles, les glàndules cementàries separades i piriformes i els ous ovalats i amb coberta gruixuda. Aquesta classe reuneix els ordres dels monilifòrmides (Moniliformida), els gigantorínquides (Gigantorhynchida), els oligacantorínquides (Oligacanthorhynchida) i els apororínquides (Apororhynchida). Al nostre país ha estat trobat solament, al Principat i a les Balears, Moniliformis moniliformis, un arquiacantocèfal paràsit de rosegadors i eriçons.
La classe dels paleacantocèfals inclou paràsits de peixos, amfibis, rèptils, ocells i mamífers. Es caracteritzen pel fet de tenir canals lacunars longitudinals principals laterals, nuclis hipodèrmics fragmentats i nombrosos, sac lligamentari únic (no persistent en la femella), glàndules cementàries separades, de forma des de tubular fins a esferoïdal, i ous ovalats o allargats, de vegades amb engruiximents polars. Inclou els ordres dels equinorínquides (Echinorhynchida) i els polimòrfides (Polymorphida). Al nostre país fins ara s’han trobat cinc espècies de paleacantocèfals: Centrorhynchus aluconis (=C. appendiculatum), trobat com a paràsit de musaranyes al Principat i a Eivissa, i de rapinyaires a les Balears; C. buteonis, trobat a les Balears com a paràsit de rapinyaires, llangardaixos i colobres; Acanthocephalus anthuris, trobat a la Catalunya del Nord i al Principat en granotes i tritons; i, finalment, A. clavula (=Echinorhynchus clavula) i Echinorhynchus truttae, tots dos trobats a la mar valenciana com a paràsits d’anguiles.