Ens hem referit sovint, en les pàgines precedents, a les gimnospermes i a les angiospermes, els dos grups que clàssicament hom distingeix dins les plantes superiors. Les diferències entre les unes i les altres són molt nombroses i transcendentals, de manera que la separació és prou justificada. Recordem, entre les característiques de les angiospermes, els primordis seminals tancats dins d’un recipient, el pistil, format per les fulles carpel·lars; l’ovari, de vegades acompanyat d’altres parts de la flor, que es converteix després de la fecundació en un fruit que guarda les llavors; la «doble» fecundació (de les dues cèl·lules espermàtiques, l’una es fusiona amb l’ovocèl·lula i origina el zigot, i l’altra ho fa amb el nucli secundari i dona lloc al nucli endospèrmic).
En el decurs dels darrers decennis els estudis paleobotànics han aportat noves dades que permeten de dibuixar amb més precisió la trajectòria evolutiva dels espermatòfits. Els nous coneixements han posat de manifest, per exemple, que dins de les gimnospermes hi ha dos grans grups independents des de la segona meitat del Paleozoic, i això obliga a reconsiderar-ne la sistemàtica.
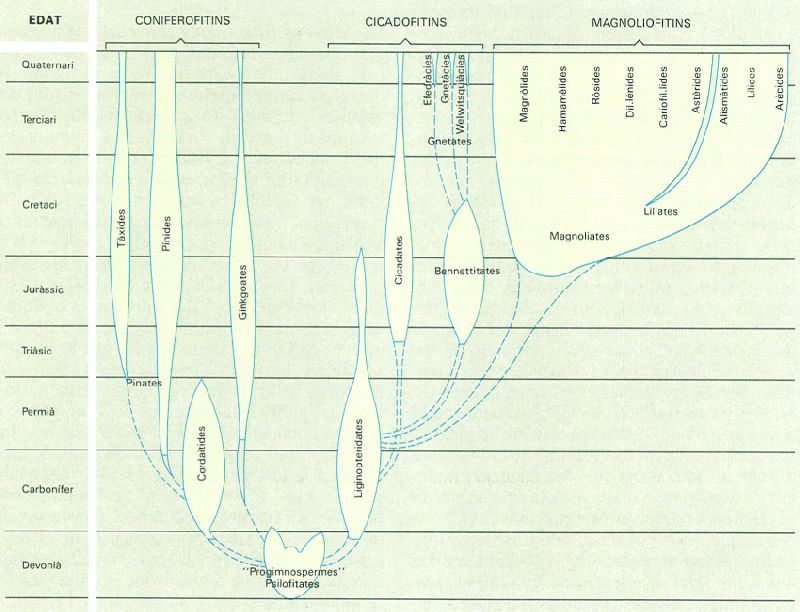
Diagrama evolutiu dels espermatòfits al llarg dels diversos períodes geològics. Les línies discontínues representen relacions insegures; l’amplada de cada rama figura la diversitat i la riquesa quantitativa dels grups respectius.
Carto-Tec, a partir d’Ehrendorfer (in Strasburger, 1986), simplificat
L’estudi de les restes fòssils actualment conegudes permet de suposar que els espermatòfits provenen de les progimnospermes, un grup de pteridòfits emparentat amb les psilofitates que va desaparèixer a la darreria del Devonià. Precisament les primeres llavors fòssils provenen d’aquest període, època en què els pteridòfits eren, encara, el grup dominant al medi terrestre. Amb l’inici del Carbonífer, les plantes amb llavor se separaren en dues línies evolutives que corresponen a dos grans grups d’organització gimnospèrmica (amb els primordis seminals oberts, fecundació senzilla, etc.), coneguts actualment amb els noms de coniferofitins i cicadofitins. La derivació paral·lela d’aquests dos grups a partir de les progimnospermes significa que tenen un origen comú, però que han evolucionat independentment. Les gimnospermes aconseguiren de senyorejar la terra, en detriment dels pteridòfits, a l’acabament del Paleozoic. És molt probable que les angiospermes s’hagin originat de cicadofitins primitius, coneguts actualment només en estat fòssil, però és particularment difícil de saber en quin moment s’inicià la desviació perquè hi ha molt poques restes fòssils antigues del grup. Les angiospermes dominen la vegetació terrestre des del final del Mesozoic.
Les tendències evolutives dels espermatòfits
L’estudi de les estructures actuals i la comparació amb les restes fòssils conegudes permeten de constatar l’existència d’unes pautes generals (discutibles i, certament, discutides) en el procés evolutiu dels espermatòfits, i fan possible, a més, la reconstrucció de les grans línies de diversificació del grup.

La flor del nelumbe o flor de l’Índia (Nelumbo nucifera) reuneix molts caràcters considerats primitius, entre els quals hi ha l’eix floral allargat i el caràcter cíclic de la flor, el periant vistent i amb moltes peces i el gineceu coricàrpic format per nombrosos carpels lliures. El nelumbe és una nelumbonàcia aquàtica cultivada com a ornamental i tan propera a les nimfeàcies que alguns autors la inclouen dins aquesta darrera família.
Jordi Vidal
Pel que fa al port, les primeres gimnospermes eren segurament plantes llenyoses i arborescents de les quals han derivat les formes arbustives, lianoides i herbàcies a través de vies diverses però paral·leles. Dins de les formes arbòries, els sistemes amb moltes rames primes i amb fulla caduca deriven d’arbres perennifolis més primitius, amb branques gruixudes i poc nombroses. La ramificació simpòdica prové de la monopòdica.
Quant a les fulles, sembla que de les ramificades en tres dimensions s’ha passat a fulles planes, encara profundament dividides i, finalment, a fulles simples, cada vegada més petites. De la disposició esparsa han derivat les fulles oposades i les verticil·lades. A les angiospermes, la nervadura més primitiva és la pinnada.
Les primeres flors es formaren per segregació de les fulles fèrtils, portadores d’esporangis, que s’agruparen helicoïdalment a l’extrem de rames curtes. Una vegada aïllades les peces florals, s’inicià un procés de reducció i fixació del seu nombre (oligomerització) coincident amb el pas cap a una disposició verticil·lada. En relació amb l’hermafroditisme floral, aparegué un periant que envoltava habitualment les peces fèrtils. L’adquisició de pètals és evolutivament més tardana que no pas la de sèpals (que són, simplement, bràctees modificades), però secundàriament aparegueren novament flors apètales. Les peces soldades es consideren més evolucionades que no les lliures. Entre les actuals angiospermes, els estams més primitius són òrgans laminars, amples, amb els sacs pol·línics en una de les cares; provenen, segurament, d’esporofil·les que portaven els microsporangis al marge.

La pol·linització entomòfila mobilitza diverses menes d’insectes amb resultats ben desiguals. Molts dels coleòpters, per exemple, visiten les flors perquè se n’alimenten, sobretot del pol·len; a causa del fregament amb els estams (com a les flors en estadi masculí de la composta de la fotografia superior), el pol·len s’adhereix a la superfície del cos i pot ser dipositat més tard, pel mateix procediment, sobre els estigmes de les flors en estadi femení del mateix capítol o d’un capítol veí. L’abella de la mel (fotografia inferior) és un dels pol·linitzadors més eficients i habituals del romaní (Rosmarinus officinalis); de manera general, visita preferentment flors zigomorfes de coloracions groguenques o blavoses, amb els senyals de nèctar ben marcats i amb zona d’aterratge.
Enric Curto
Hom ha discutit molt si les primitives angiospermes tenien les flors grosses o petites, i actualment es pot suposar que eren mitjanes; les flors grosses que trobem a les magnoliàcies i a d’altres famílies properes podrien tenir origen secundari, com a resposta a la pressió de selecció dels mecanismes de pol·linització. Pel que fa a la simetria, el camí seguit a partir de les flors acícliques, primàriament asimètriques, passa per les flors actinomorfes, les bilaterals i les zigomorfes, i pot arribar novament a una situació d’asimetria (però en aquest cas en flors cícliques). Les flors més primitives són unisexuals i anemògames tot i que, a les angiospermes, l’anemofília s’ha de considerar secundària. Els primers animals pol·linitzadors, encara poc especialitzats, foren segurament coleòpters pol·linívors. La necessitat d’estalviar pol·len reconduí el camí evolutiu cap a flors amb nèctar, visitades inicialment per altres grups d’insectes, i posteriorment per ocells petits, rats-penats i d’altres animals. Els primordis seminals de les gimnospermes són directament accessibles als grans de pol·len. A les angiospermes primitives, els carpels no s’han soldat del tot per les vores i els grans de pol·len germinen als marges de la fulla carpel·lar, que representen l’antecedent immediat de l’estigma. Ben aviat, però, els carpels es tanquen, de primer separadament els uns dels altres (gineceu apocàrpic) i, més tard, fusionats tots junts donant lloc a gineceus sincàrpics o cenocàrpics.
Pel que fa a les inflorescències, hi ha raons per a suposar que les racemoses deriven de les cimoses, les més primitives de les quals són els monocasis. Del pleocasi pot provenir el raïm, i d’aquest s’originen la resta de les inflorescències racemoses.
Els gametòfits manifesten una tendència molt clara cap a la reducció i la simplificació progressives. Aquest fet comporta avantatges adaptatius considerables, perquè la maduració és més precoç i la reproducció més ràpida i efectiva. Per aquest procés es passa de zoidiogàmia (fecundació mitjançant espermatozoides) a sifonogàmia (fecundació mitjançant cèl·lules espermàtiques que són transportades al llarg del tub pol·línic).
La testa de les llavors més primitives consta sovint d’un estrat carnós, exterior, i un altre d’interior lignificat i més dur. A la resta (la gran majoria) dels espermatòfits, la coberta de la llavor és típicament seca i dura, però el seu gruix es va reduint gradualment perquè el fruit col·labora també en la protecció de la llavor. L’albumen és constituït inicialment per les restes del megaprotal·lus (endosperma primari de les gimnospermes); a les angiospermes, l’endosperma secundari només es forma després de la fecundació. El nombre de cotilèdons disminueix: són molts a les gimnospermes, dos a les dicotiledònies i un de sol a les monocotiledònies. A les angiospermes, les llavors amb endosperma abundant i amb l’embrió petit i poc diferenciat són pròpies dels representants més primitius; les llavors dels grups més evolucionats tenen poc endosperma (o gens) i un embrió gros i ben diferenciat. La coberta parcialment carnosa de les llavors dels primers espermatòfits fa pensar en una disseminació endozoòcora, potser duta a terme per rèptils (saurocòria); la diversificació de la zoocòria i l’aparició de l’anemocòria i de la hidrocòria són posteriors; i segurament la mirmecocòria és un dels sistemes evolutivament més avançats.
La sistemàtica
Els espermatòfits (Spermatophyta) es reparteixen en tres subdivisions, la dels coniferofitins, la dels cicadofitins i la dels magnoliofitins. Les dues primeres corresponen a les gimnospermes, considerades avui, simplement, com un nivell de desenvolupament primitiu dins dels espermatòfits i no pas un grup sistemàtic natural; els magnoliofitins corresponen precisament a les angiospermes. La subdivisió dels coniferofitins (Coniferophytina) inclou poques espècies —unes 600—, però de notable importància en el paisatge mundial; són les gimnospermes de fulla acicular o de nervadura dicòtoma. La subdivisió dels cicadofitins (Cycadophytina) agrupa les gimnospermes de fulla pinnada, unes 200 arreu del món; en provenen, molt probablement, els magnoliofitins, originats segons tots els indicis del grup actualment extingit de les pteridospermes. La subdivisió dels magnoliofitins (Magnoliophytina) és la més nombrosa, ja que té aproximadament un quart de milió d’espècies que es reparteixen entre dues classes perfectament delimitades, la de les magnoliates (Magnoliatae) o dicotiledònies, i la de les liliates (Liliatae) o monocotiledònies; la distinció entre unes i altres arrenca de la darreria del segle XVII, a proposta de J. Ray.