L'arbre de la vida al principi del segle XXI

Arbre de la vida és el títol de la instal·lació al·legòrica del lligam entre el DNA i les relacions filogenètiques, amb què el CosmoCaixa de Barcelona rep els visitants. Es tracta d’un tronc de 300 anys d’antiguitat d’un arbre amazònic (Minquartia guianensis), anomenat acariquara, que és famós per les qualitats de la seva fusta.
CosmoCaixa Barcelona - Obra Social Fundació "La Caixa"/ Ronald Stallard
Des de mitjan segle XX, amb la incorporació progressiva de les tècniques i eines de la biologia molecular a la filogènia, és a dir, amb el desenvolupament de la filogènia molecular, la classificació dels éssers vius ha experimentat diverses transformacions que han afectat molts grups taxonòmics, inclosos els grans clades basals, uns canvis que han proporcionat una visió diferent i molt més real de l’arbre de la vida i de les relacions filogenètiques entre els diversos grups d’éssers vius. Tanmateix, no ha estat només una qüestió tècnica. Hi ha un altre motiu, molt més profund, que fa referència a la manera com ha anat evolucionant la visió que hom té de la jerarquització de la biodiversitat, la qual es basa en els processos algorítmics que s’empren per assegurar que l’anàlisi i la comparació de les estructures dels éssers vius condueixen a una classificació completament filogenètica.
En biologia, com en molts altres aspectes de la vida, sovint hom acostuma a distingir entre els objectes i els processos. En filosofia es discuteix fins a quin punt aquesta distinció té fonaments merament ontològics, però el cert és que en biologia tots els éssers vius estan sotmesos a un procés de canvi continu, que els dóna les seves característiques en cada moment donat i, per tant, en conseqüència, els defineix, la qual cosa fa que sovint sigui difícil de destriar l’objecte del seu procés. Aquesta visió de canvi impregna l’estudi de pràcticament totes les disciplines biològiques, de la biologia evolutiva a l’ecologia i de la biologia del desenvolupament a la filogènia. Hi ha canvis ràpids, com els que s’esdevenen durant el desenvolupament embrionari, i d’altres de molt més lents, com els de l’evolució; però en biologia, l’atenció sobre els objectes que hom estudia implica necessàriament una atenció sobre els processos que els han generat.
Llegir i representar gràficament la història de la vida

Ramificació dels llinatges dels diàpsids en un arbre filogenètic simplificat. Aquest arbre indica que tots dos grups d’animals comparteixen un avantpassat comú més recent entre ells (representat per un punt vermell) que no pas amb els llangardaixos (node blau), però no que en el moment de la ramificació ja sorgissin cocodrils i ocells amb les característiques que actualment tenen. Cal fer el mateix raonament per a tots els nodes i les ramificacions dels llinatges de l’arbre de la vida.
Original de l’autor / iStockphoto / Studio Araminta, Corel i Stock.xchng.
Charles Darwin (1809 - 1882) va incloure la paraula origen en el títol de la seva obra més coneguda de totes, L’origen de les espècies (1859), i fou precisament aquesta paraula, el problema dels orígens, el que va motivar la major part de les objeccions que es van fer a la seva teoria, especialment quant a l’origen de l’espècie humana. La cerca dels orígens és la cerca dels avantpassats i, més concretament, dels ancessors més antics d’un determinat llinatge. La filogènia, amb els atractius i elegants nodes i branques dels arbres que produeix, es percep sovint com la ciència dels avantpassats i, en conseqüència, com una manera d’acostar-nos científicament als orígens dels llinatges. Tanmateix, però, els cladogrames i els arbres filogenètics no representen de forma estricta les relacions ancessor-descendent, com fan les genealogies, malgrat que a primera vista s’hi puguin assemblar.
Com s’ha dit abans, els nodes (els punts de ramificació) indiquen la presència de sinapomorfies (caràcters derivats que són comuns als llinatges que es ramifiquen a partir d’aquell node) i les branques encarnen la continuïtat històrica d’un conjunt de caràcters sotmesos a modificacions successives i, per tant, també a l’origen de nous caràcters, que en conjunt condueixen al següent node. Més encara, en els cladogrames, o arbres sense arrel (la representació gràfica més utilitzada en l’actualitat per a representar les relacions filogenètiques), ni tan sols es pot assegurar quin caràcter dels que s’utilitzen per a separar dos nodes veïns és el més antic (el plesiomòrfic) i quin és el derivat (l’apomòrfic). La direcció del canvi només es fa evident quan l’arbre es dota d’arrel, és a dir, quan es determina quina és la bifurcació més antiga, la basal, dels grups que s’analitzen. De fet, quan es fixa l’arrel és quan el cladograma esdevé un arbre filogenètic. És llavors quan és possible establir un marc sobre el qual reconstruir un fragment de la història evolutiva, una seqüència temporal de transformació dels caràcters.
Per analitzar dos grups d’animals el llinatge dels quals sorgeix d’un node comú en un arbre filogenètic es pot considerar, per exemple, el cas dels cocodrils i els ocells, dos grups germans segons les dades aportades per la filogènia molecular entre els quals hi ha registrades nombroses diferències morfològiques i genètiques, però que tanmateix llurs llinatges respectius es bifurquen a partir d’un mateix node. Si només es consideren els representants vius d’aquests dos grups, hom podria arribar a pensar que les diferències que els separen es van originar en un instant, representat pel node en qüestió, però és ben segur que no va ser pas així. És molt més probable que, per exemple, les tres innovacions morfològiques més òbvies dels ocells respecte als cocodrils (i la resta de rèptils), és a dir, les plomes, les ales i el reemplaçament de les vèrtebres posteriors per una cua, evolucionessin de forma seqüencial, probablement en aquest ordre. No obstant això, en construir l’arbre cal situar la gènesi teòrica de totes aquestes diferències i de les moltes altres que els distingeixen en el node a partir del qual van divergir aquests grups, a l’era mesozoica.
Tanmateix, en termes filogenètics, aquest node indica que el grup dels cocodrils comparteix un avantpassat comú més recent amb el grup dels ocells que no pas amb qualsevol altre grup d’éssers vius, inclosos els llangardaixos i la resta de rèptils, la qual cosa no s’observava en cap de les filogènies clàssiques, però en cap cas implica que totes aquestes diferències sorgissin en un instant. A més a més, aquest node suggereix una llista de caràcters que probablement eren presents en el darrer ancessor comú a ocells i cocodrils, però tampoc no dibuixa pas una imatge concreta literal d’aquest avantpassat, ni indica, per exemple, quin va ser ni com era el que es pot considerar com el primer grup d’ocells. És així com cal interpretar els nodes i les ramificacions dels cladogrames i dels arbres filogenètics, inclosos els nodes i les ramificacions més basals, els que es troben a l’origen dels llinatges actuals, dels quals hom parlarà tot seguit.
A més, el fet que el grup dels cocodrils comparteixi un avantpassat comú més recent amb els ocells que amb qualsevol altre grup de rèptils indica que, des del punt de vista de la cladística, els ocells i els rèptils formen part del mateix grup. Aquest és un dels molts canvis de visió sobre la filogènia dels éssers vius que la filogènia molecular ha aportat a l’arbre de la vida.
Regnes, imperis i dominis
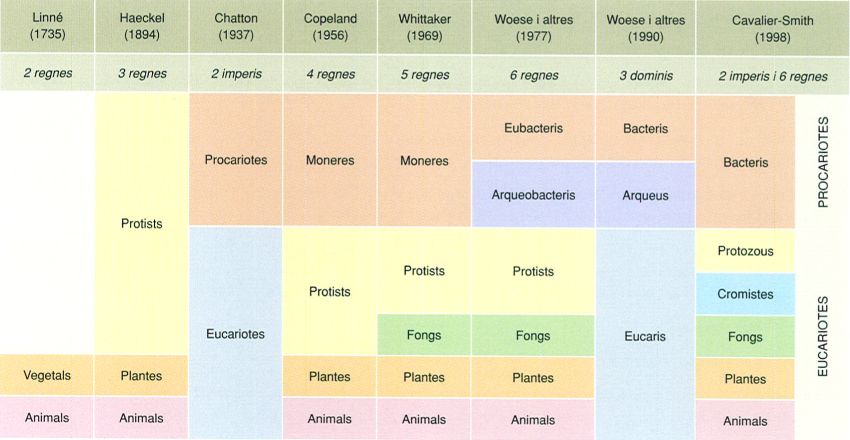
Resum de les diverses classificacions dels grans grups basals en el decurs de la història de les classificacions filogenètiques. En color, els grups aproximadament equivalents segons els diversos sistemes de classificació. També s’indica el nom de qui ho va proposar i l’any.
A partir de fonts diverses
Com s’ha explicat més extensament abans, la inferència filogenòmica implica dos passos crucials. Primer s’identifiquen els caràcters homòlegs entre els tàxons a relacionar, aquells que procedeixen d’un avantpassat comú. I després es reconstrueixen les relacions de parentiu i, en conseqüència, la història evolutiva d’aquests tàxons mitjançant la utilització de mètodes de construcció d’arbres, amb els quals es comparen aquests caràcters. Així, per a inferir filogènies hom pot utilitzar gairebé qualsevol caràcter, amb la condició que sigui homòleg, com per exemple estructures morfològiques, característiques ultraestructurals de les cèl·lules, rutes bioquímiques, presència i disposició de gens en el genoma i seqüències d’aminoàcids o de nucleòtids. Quan es comparen seqüències, l’homologia es determina mitjançant processos informàtics de cerca de similitud. Llavors, amb tots aquests caràcters es construeix una matriu, amb la qual es quantifica cada caràcter concret en les espècies que es comparen. Finalment, a partir d’aquesta matriu s’infereix l’arbre o el cladograma, mitjançant un dels mètodes de reconstrucció filogenètica: els d’unió de veïnatge o d’evolució mínima, que converteixen la matriu de caràcters en una matriu de distàncies evolutives; el de màxima parsimònia, que selecciona l’arbre filogenètic que requereix el mínim nombre de canvis per a explicar les dades observades respecte als caràcters usats en la matriu; o el de màxima versemblança, que selecciona l’arbre que atorga la màxima probabilitat que es puguin observar les dades que es comparen en funció d’un model determinat entre d’altres.
L’aplicació de la revolució conceptual i metodològica que fonamenta l’anàlisi cladística ha canviat alguns dels grups clàssics que es fonamentaven en estudis morfològics, inclosos els grans grups basals. D’aquesta manera, s’han succeït progressivament la concepció clàssica de Linné del 1735 de dos regnes: el dels vegetals i el dels animals; la posterior proposta d’Ernst Haeckel (1834 - 1919), que el 1894 va canviar el nom del regne dels vegetals pel de regne de les plantes i va incorporar el regne dels protists per a incloure tots els éssers vius que Linné no havia considerat i, per tant, no havia incorporat, com els bacteris, els protozous i els fongs; la simplificació d’Édouard Chatton (1883 - 1947), que en distingir entre els organismes eucariotes (organismes unicel·lulars o pluricel·lulars conformats per cèl·lules amb nucli) i procariotes (organismes unicel·lulars sense nucli) va proposar el 1937 l’existència de sengles dos imperis; la posterior ampliació de Herbert Faulkner Copeland (1902 - 1968), que el 1938, i amb més detall l’any 1956, va proposar l’existència d’un nou regne, el de les moneres, per a agrupar els organismes procariotes, segregant-los així del regne dels protists proposat per Haeckel; la nova ampliació de Robert Whittaker (1920 - 1980), que el 1969 va definir un nou regne, el dels fongs, i establí la classificació basal encara utilitzada actualment (però filogenèticament inexacta) basada en l’existència de cinc regnes: el de les moneres, el dels protists, el dels fongs, el de les plantes i el dels animals; una altra ampliació, proposada per Carl Richard Woese (1928-) el 1977, el qual, utilitzant per primer cop la filogènia molecular d’acord amb la seqüència de l’RNA ribosòmic 16S, va segregar el regne de les moneres en els regnes dels eubacteris i dels arqueobacteris; la simplificació proposada pel mateix Woese el 1990, que també amb dades de filogènia molecular va proposar agrupar tots els éssers vius en tres dominis: els eucaris (on resten inclosos tots els éssers vius constituïts per cèl·lules eucariotes), els bacteris i els arqueus (on hi ha tots els éssers vius de base procariota), que és la classificació basal actualment acceptada i utilitzada de forma general; i finalment, la proposta de Thomas Cavalier-Smith (1942-), l’any 1998, d’una classificació basal jeràrquica en dos imperis, el dels procariotes (que englobaria novament els bacteris o eubacteris i els arqueus) i el dels eucariotes; i en sis regnes, de categoria inferior als imperis: el dels bacteris (l’únic regne de l’imperi dels procariotes) i els dels protozous, els cromistes, els fongs, les plantes i els animals, tots dins l’imperi dels eucariotes. Cal destacar que aquesta darrera proposta incorpora un nou regne, el dels cromistes (Chromista), que inclouria totes les algues amb cloroplasts que contenen clorofil·la de tipus a o c, com també diverses espècies sense acolorir íntimament relacionades amb elles; aquesta proposta, però, no ha tingut mai un suport ampli i actualment està en desús.
L’accés a les seqüències de DNA i de proteïnes, facilitat per tècniques com la reacció en cadena de la polimerasa i la seqüenciació automàtica, també ha permès establir moltes altres relacions filogenètiques insospitades per comparació amb les filogènies basades en caràcters morfològics.
A continuació es comenten les característiques dels grups basals en funció dels dos sistemes de classificació actualment utilitzats: el dels cinc regnes de Whittaker (1969), que, basat en caràcters mofològics i malgrat que algunes de les relacions que proposa no són confirmades per la filogènia molecular, encara és emprat en diversos àmbits com el de l’ensenyament; i el dels tres dominis de Woese (1990), que actualment és acceptat i que recolza en diversos conjunts de dades procedents de la filogènia molecular.
La llista següent resumeix els principals canvis introduïts en les relacions filogenètiques a partir de l’aplicació dels estudis de filogènia molecular.
- El parentiu de llinatge entre els eumlcets (un grup de fongs sense valor taxonòmic que comprèn les classes dels ficomicets, els ascomicots i els basidiomicots, que s’ha emprat tradicionalment per a distingir-los dels mixomicets) i els microsporidis (un ordre de protozous de la classe dels esporozous).
- La construcció del clade dels opistoconts, que comprèn els eumicets i els microsporidis esmentats en el punt anterior, els coanoflagel·lats (protozous monoflagel·lats que comparteixen un avantpassat comú amb els animals) i els metazous (grup filogenètic que comprèn els animals).
- El parentiu de llinatge entre els clorobionts (un grup taxonòmic que reuneix les algues verdes i les plantes terrestres) i els rodòfits (algues pluricel·lulars de tal·lus filamentós o laminar, sovint d’estructura complexa i generalment de color vermellós).
- La construcció del clade dels alveolobionts, que comprèn els ciliats (protozous en general relativament grossos i d’estructura complexa, amb el cos recobert de cilis disposats formant sèries característiques de cada espècie), els dinòfits (un grup d’algues unicel·lulars nedadores que tenen dos flagels desiguals, un en posició longitudinal i un altre típicament transversal, que els permeten moure’s), els esporozous i els hematozous (dos grups de protozous paràsits).
- La construcció del clade dels estramenòfils, que comprèn les algues brunes, les diatomees (algues unicel·lulars desproveïdes de flagels i envoltades per una membrana pèctica impregnada de sílice), els crisòfits (algues unicel·lulars biflagel·lades típiques de les aigües dolces), els xantòfits i els eustigmatòfits (dos grups d’algues groc verdoses), els oomicets (subclasse de ficomicets integrada per fongs paràsits o sapròfags, de miceli ben desenvolupat i de parets cel·lulars cel·lulòsiques) i els hipoquitridiomicets (un grup de fongs).
- La parafílla de les algues verdes.
- La parafília dels briòfits (plantes arquegoniades que es caracteritzen per tenir la generació esporofítica unida a la gametofítica i normalment paràsita d’aquesta darrera, com les molses).
- La parafília de les gimnospermes (plantes llenyoses de ramificació monopòdica i generalment perennifòlies que es caracteritzen perquè els primordis seminals no són inclosos en un ovari, com les coníferes).
- La parafília dels diblàstics (metazous el desenvolupament embrionari dels quals es caracteritza per presentar dos fulls embrionaris, l’ectoderma i l’endoderma, com les esponges, els cnidaris i els cnetòfors).
- La divisió dels protostomats en lofotrocozous (grup taxonòmic, en el qual el desenvolupament de les espècies que inclou presenta lofòfor o larva trocòfora, com els mol·luscs i els platihelmints, entre molts d’altres) i ecdisozous (grup taxonòmic que inclou els nematodes i els panartròpodes), que anul·la l’antiga divisió dels triblàstics en acelomats, pseudocelomats i celomats.
- La parafília dels crustacis.
- La parafília dels peixos i els rèptils.
- El parentiu de llinatge dels hipopotàmids i els cetacis, el qual implica la parafília dels artiodàctils (mamífers amb extremitats acabades en unglots).
Els cinc regnes
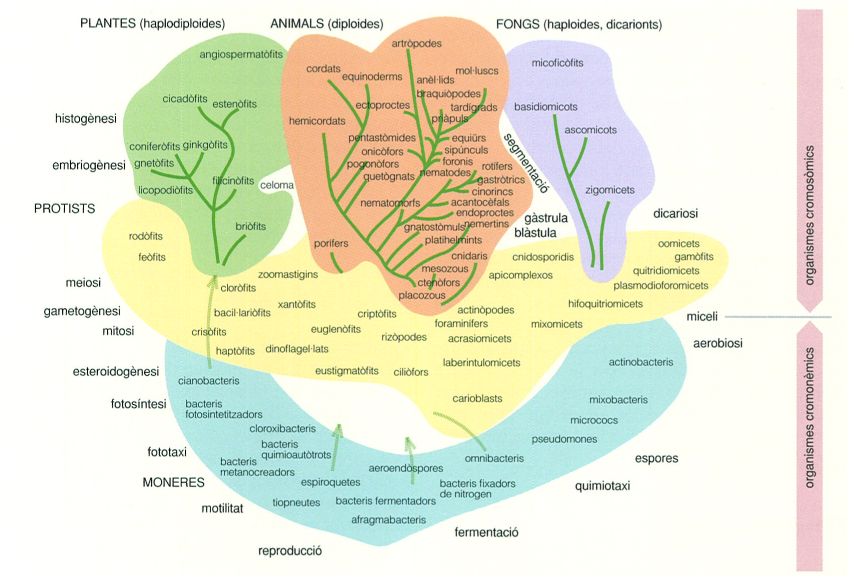
Classificació dels organismes en cinc regnes de R.H. Whittaker del 1969, en què el regne de les moneres conté tots els procariotes, i la resta són eucariotes. La proposta recollia la nova visió dels organismes gràcies al coneixement de la microestructura cel·lular i les diverses hipòtesis emeses sobre l’origen de la vida i de la cèl·lula eucariota.
Barreras & Creixell, a partir de fonts diverses.
S’anomena regne cadascuna de les cinc categories taxonòmiques més àmplies en què són agrupats els éssers vius d’acord amb característiques morfològiques segons la proposta de Whittaker de l’any 1969. Aquestes categories han estat reconegudes durant diverses dècades pels codis internacionals de nomenclatura zoològica, botànica i bacteriològica, i que malgrat que no es corresponen exactament amb allò que aconsellen els estudis filogenètics continuen essent emprats en diversos contextos, per la seva utilitat pràctica i intuïtiva.
La taula següent resumeix les principals característiques dels cinc regnes d’éssers vius proposats per Whittaker l’any 1969: moneres, protists (o protoctists), fongs, metàfits (plantes) i metazous (animals).
| Tipus de cèl·lules | Tipus d’organització cel·lular | Tipus de nutrició | Tipus de digestió | ||
|---|---|---|---|---|---|
| 1. Regne de les moneres | Procariotes | Unicel·lular | Autòtrofs o heteròtrofs | Externa | |
| 2. Regne dels protists o protoctists | Protozous | Eucariotes | Unicel·lular | Heteròtrofs | Interna |
| Algues | Eucariotes | Unicel·lular o pluricel·lulartal·lofític | Autòtrofs fotosintetitzadors | ||
| 3. Regne dels fongs | Eucariotes | Unicel·lular o pluricel·lular tal·lofític | Heteròtrofs | Externa | |
| 4. Regne dels metàfits (plantes) | Eucariotes | Pluricel·lular tissular | Autòtrofs fotosintetitzadors | ||
| 5. Regne dels metazous (animals) | Eucariotes | Pluricel·lular tissular | Heteròtrofs | Interna |
El regne de les moneres
El regne de les moneres, nom actualment en desús i que és substituït pel de procariotes, inclou tots els organismes unicel·lulars les cèl·lules dels quals són mancades d’un nucli envoltat de membrana per a contenir el material genètic. La seva morfologia és molt simple, atès que també són mancats de la major part d’orgànuls cel·lulars, com mitocondris i cloroplasts, i de tota la resta d’estructures membranoses típiques de les cèl·lules dels protozous, les algues, els fongs, les plantes i els animals, com per exemple el reticle endoplasmàtic, els lisosomes, els vacúols i l’aparell de Golgi. Una altra de les característiques pròpies d’aquest grup és que en la major part d’organismes el material genètic és compost per una única molècula de DNA de forma circular. La seva alimentació pot ser autòtrofa, com els bacteris fotosintetitzadors i quimiosintetitzadors, o heteròtrofa. La filogènia molecular ha dividit aquest regne en dos dels tres dominis actualment reconeguts: el dels bacteris (o eubacteris) i el dels arqueus.
El regne dels protists
Els protists, anomenats també protoctists, són un grup divers de microorganismes eucariotes (les seves cèl·lules tenen el material genètic contingut dins un nucli envoltat d’una membrana nuclear) que no tenen gaire cosa en comú entre ells, llevat d’una organització relativament senzilla. Viuen en gairebé qualsevol medi que contingui aigua líquida i hi ha més de 150.000 espècies descrites. N’hi ha tant d’unicel·lulars, com els protozous i les algues unicel·lulars, com de pluricel·lulars tallofítics, és a dir, que llurs cèl·lules no estan organitzades en teixits especialitzats, com la resta d’algues. Molts d’aquests microorganismes, com ara les algues unicel·lulars, són fotosintetitzadors, productors primaris essencials dels ecosistemes, especialment a l’oceà, on formen part del plàncton. D’altres són heteròtrofs. Els uns són sèssils i uns altres estan dotats de sistemes de locomoció, com per exemple cilis o flagells, o bé es mouen mitjançant moviments ameboides.
Tanmateix, la taxonomia molecular tendeix a evitar els noms clàssics d’aquest grup, com per exemple el de protozou per referir-se a les espècies heteròtrofes de protists no filamentosos. Un dels motius principals d’aquests canvis és que els protists com a grup clàssic no és monofilètic, com exigeix la cladística, sinó polifilètic. És a dir, que no inclou l’avantpassat comú més recent de tots els seus membres o, dit d’una altra manera, que és constituït per la unió artificial de branques disperses de l’arbre de la vida. Per exemple, la filogènia molecular situa dins un mateix grup, el dels Archaeplastida, les algues verdes i les plantes terrestres, i en un grup completament diferent, el dels excavats, les euglenes, uns organismes unicel·lulars eucariotes proveïts de flagells presents en molts cursos d’aigua que tradicionalment també han estat considerades dins el regne dels protists.
Els protists han estat tradicionalment subdividits en diversos grups, sovint per les seves semblances amb els altres regnes eucariotes, segons un punt de vista clarament antropocèntric. Així, s’han reconegut els protozous per la seva semblança amb els animals quant a mobilitat; els protòfits, majoritàriament algues unicel·lulars, per la seva semblança amb les plantes quant a la presència de pigments fotosintetitzadors; i els mixomicets i oomicets també anomenats pseudofongs per la seva semblança amb els fongs. Malgrat que aquests grups sovint s’encavalquin, a vegades encara es fan servir com a noms informals per a descriure la seva morfologia i ecologia.
El regne dels fongs
Els fongs són un regne d’organismes eucariotes heteròtrofs, que poden ser sapròfags o paràsits. N’hi ha tant d’unicel·lulars, com els llevats, com de pluricel·lulars, com els ficomicets i els basidiomicots, que formen filaments multicel·lulars anomenats hifes, les quals constitueixen el miceli del fong. Tots els fongs es reprodueixen per espores i no formen teixits veritables (és a dir, els fongs pluricel·lulars tenen estructura tal·lofítica). Els basidiomicots, a més, formen una estructura especial, el bolet, un cos fructífer especialitzat en la producció i la disseminació de les espores. També es poden reproduir de forma vegetativa, per trencament del miceli. Hi ha més de 100.000 espècies de fongs descrites.
Tradicionalment, els fongs s’han estudiat paral·lelament a les plantes, com una disciplina més de la biologia vegetal, però les dades aportades per la filogènia molecular indiquen clarament que són el grup germà dels animals, no pas de les plantes.
El regne de les plantes
El regne de les plantes o regne dels metàfits inclou els organismes pluricel·lulars eucariotes les cèl·lules dels quals tenen paret cel·lular (cèl·lules vegetals) i s’organitzen en teixits veritables (estructura tissular), la qual cosa exclou les algues. Les plantes són autòtrofes fotosintetitzadores i capten l’energia solar a través de la clorofil·la dels cloroplasts. Hi ha més de 350.000 espècies descrites.
Tradicionalment aquest regne s’ha dividit en funció de si les plantes eren vasculars (traqueòfits) o no vasculars (briòfits, com les molses). Els traqueòfits, al seu torn, tenint en compte si fan llavors (espermatòfits) o no en fan (pteridòfits, com les falgueres); i els espermatòfits, segons si les llavors són contingudes en un fruit (angiospermes) o no formen fruit (gimnospermes, com els pins i els avets). En aquest cas, la filogènia molecular no ha alterat substancialment la classificació interna d’aquest grup, llevat de la parafília de les monocotiledònies i les dicotiledònies, però ha permès relacionar directament les plantes amb les algues i explicar la divergència del grup de les angiospermes amb la resta de grups de plantes, a través de cinc grups basals d’angiospermes, coneguts col·lectivament com el clade ANITA per la inicial dels cinc grups: el format per l’única espècie del gènere Amborella, i els grups de les nimfeals (Nymphaeales), les il·licials (Illiciales), les trimeniàcies (Trimeniaceae) i les austrobaileials (Austrobaileyales).
El regne dels animals
Finalment, el regne dels animals, o regne dels metazous, constitueix un grup d’éssers vius eucariotes, heteròtrofs i pluricel·lulars organitzats en teixits veritables. També es caracteritzen per la seva capacitat de locomoció, per l’absència de clorofil·la i de paret en les seves cèl·lules i pel seu desenvolupament embrionari, que inclou, a diferència de les plantes i els fongs, una fase de blàstula que determina un pla corporal fix, malgrat que moltes espècies poden patir una metamorfosi posterior.
Com en els regnes anteriorment esmentats, la filogènia molecular ha donat una perspectiva diferent de la classificació interna dels animals i de la seva relació filogenètica amb els altres grups, per exemple, situant-los com a grup germà dels fongs i reordenant la seva filogènia interna, amb la incorporació de diversos tàxons, subdividint els protostomats (grup d’animals la boca dels quals s’origina a partir del blastòpor, i l’anus, a partir del pol embrionari oposat, i que comprèn els platihelmints, els nemertins, els asquelmints, els anèl·lids, els mol·luscs i els artròpodes, entre d’altres) en lofotrocozous (animals protostomats que presenten un estadi de larva trocòfora, com els braquiòpodes, els ectoproctes o briozous, els cicliòfors, els entoproctes, els gnatostomúlids, els mol·luscs, els nemertins, els platihelmints, els forònids, els rotífers i els sipúnculs) i en ecdisozous (grup que comprèn els cefalorincs o escalidòfors, els nematodes i els panartròpodes).
Els tres dominis
S’anomena domini cadascun dels tres nivells superiors de la classificació dels éssers vius tal com els va definir Carl Woese el 1990 utilitzant eines i dades de filogènia molecular. Aquests tres dominis, actualment acceptats, són el domini dels bacteris, el domini dels arqueus i el domini dels eucaris. Tots tres es consideren monofilètics, és a dir, que les seves espècies comparteixen un ancessor comú i alhora engloben totes les espècies que en descendeixen. Actualment és acceptat que constitueixen els grups basals de l’arbre de la vida.
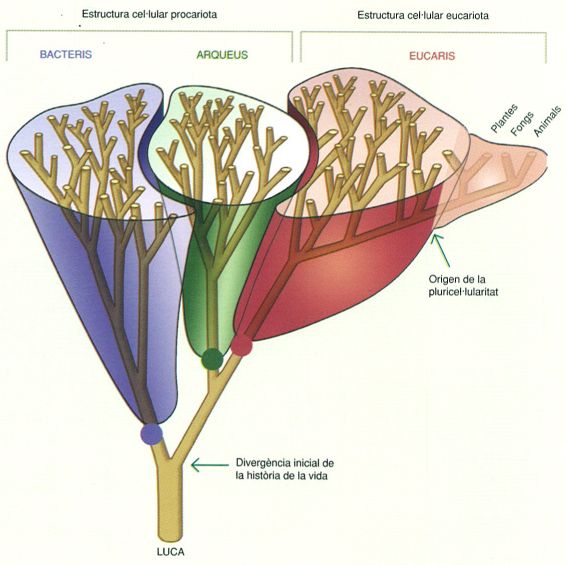
Arbre de la vida simplificat on es mostren els tres dominis d’éssers vius tal com els va definir Carl Woese el 1990, una classificació actualment acceptada.
A partir de Woese, Kandler i Wheelis, 1990.
Arbre filogenètic (circular, a dalt) que mostra la major proximitat filogenètica del domini dels arqueus amb el domini dels eucaris, en comparació amb el domini dels bacteris. A baix, esquema hipotètic en forma d’arbre que mostra la transferència horitzontal de gens entre organismes no relacionats per llinatge.
Un dels problemes principals per a entendre la natura de la monofília i les relacions entre aquests grups d’organismes fa referència al lloc concret on cal situar l’arrel de l’arbre, és a dir, a la posició teòrica de l’últim ancessor comú universal a tots els éssers vius actuals, conegut com LUCA (acrònim de Last Universal Common Ancestor), del qual tots procedeixen per divergències successives del seu llinatge. Les dades moleculars de què es disposa, molt especialment quant a la presència de gens duplicats, situen aquesta arrel a l’inici de la branca que hi ha entre el domini dels bacteris i els dominis dels arqueus i els eucaris.

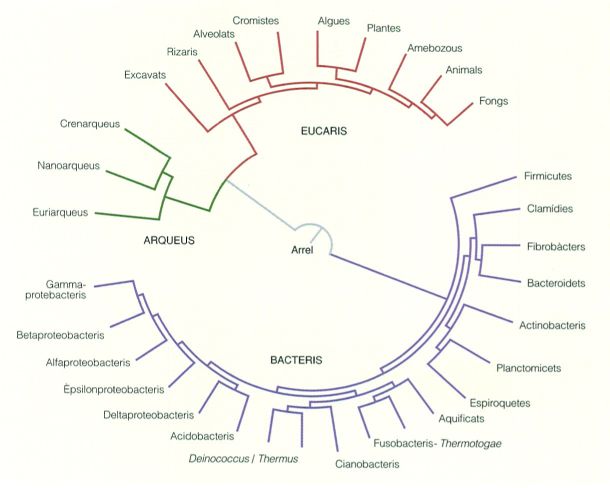
Arbre de la vida on s’indiquen els tres dominis i els principals grups dins de cada domini. Els fongs, les plantes i els animals es mostren emmarcats per a situar-los en el conjunt de l’arbre de la vida.
Un dels problemes concrets que presenta l’assignació de la posició de l’arrel de l’arbre de la vida és el de la transferència horitzontal de gens entre grups d’organismes relativament distants, un fenomen biològic que pot emmascarar les relacions filogenètiques reals. La transferència gènica horitzontal, anomenada també transferència gènica lateral, és un procés biològic mitjançant el qual un organisme incorpora material genètic d’un altre sense ser-ne el descendent. Aquesta transferència de gens no té res a veure amb la transferència vertical, la que es dóna quan un organisme rep el material genètic del seu progenitor per mitjà de la reproducció. La transferència gènica horitzontal fou descrita per primer cop el 1959 per explicar el traspàs de gens que confereixen resistència a determinats antibiòtics entre bacteris d’espècies diferents. És un fenomen biològic relativament comú entre organismes procariotes, fins i tot entre espècies distants filogenèticament. En els eucariotes, en canvi, la transferència horitzontal de gens és un fenomen molt poc freqüent, però, malgrat tot, l’anàlisi de determinades seqüències de DNA ha demostrat que s’ha produït un cert nombre de vegades. Així, per exemple, els mitocondris i els cloroplasts, que segons la teoria actualment acceptada de l’endosimbiosi com a origen de les cèl·lules eucariotes provenen de bacteris endosimbionts, van traspassar bona part del seu material genètic al genoma nuclear, un cas clar de transferència horitzontal de gens. També s’ha demostrat la transferència horitzontal d’alguns gens entre bacteris i llevats de l’espècie Saccharomyces cerevisiae; entre bacteris endosimbionts del gènere Wolbachia i els seus hostes, com per exemple escarabats asiàtics de l’espècie Callosobruchus chinensis, alguns artròpodes del gènere Drosophila i nematodes de l’espècie Brugia malayi; entre el genoma mitocondrial i plantes paràsites de la família de les rafflesiàcies (Rafflesiaceae), pròpies del sud-est asiàtic; entre el genoma dels cloroplasts d’una planta encara no identificada i els mitocondris de la pesolera (Phaseolus sp.); i entre diatomees i l’opistobranqui Elysia chlorotica, que se n’alimenta.
En aquest sentit, la transferència horitzontal de gens pot alterar la inferència filogenètica. Per exemple, si es consideren dues espècies de bacteris filogenèticament distants entre els quals s’ha produït un episodi de transferència gènica horitzontal, la inferència filogenètica tendirà a apropar-les, atès que comparteixen un mateix gen, malgrat que els altres siguin diferents. Per aquest motiu les filogènies robustes es construeixen amb el major nombre de caràcters possibles, inclòs un ampli ventall de gens diferents. El gen més utilitzat en la filogènia de procariotes és el corresponent a l’RNA ribosòmic 16S, atès que acostuma a estar molt conservat en les espècies més emparentades i al mateix temps presenta prou diferències per a poder agrupar-les convenientment. Tanmateix, en alguns casos també ha estat transferit horitzontalment, i això obliga a replantejar alguns nodes i embrancaments de la filogènia d’aquests grups.
La taula següent recull les característiques distintives principals dels tres dominis (bacteris, arqueus i eucaris): el nombre d’espècies identificades, els fòssils més antics coneguts i la distribució actual de sengles membres.
Els tres dominis| Bacteris | Arqueus | Eucaris | |
|---|---|---|---|
| Característiques específiques distintives | La paret cel·lular conté àcid muràmic. L’RNA de transferència que inicia la traducció porta una N-formilmetionina. |
La membrana cel·lular conté lípids exclusius d’aquest domini, amb unions èter en lloc d’èster, que li donen més rigidesa. Presenten canvis exclusius al braç TψC de l’RNA de transferència. Tenen ribosomes específics. |
El DNA és contingut dins un nucli limitat per una membrana nuclear. Tenen citosquelet format per microtúbuls de tubulina i mitocondris. El DNA es compacta en cromosomes abans de la divisiò cel·lular. Les cèl·lules es reprodueixen per mitosi, amb participaciò de centríols i microtúbuls. Tenen sexualitat veritable. |
| Nombre d’espècies | 9.021 classes identificades mitjançant l’RNA ribosòmic 16S | 259 classes identificades mitjançant l’RNA ribosòmic 16S | Més d’1,7 milions |
| Fòssils més antics coneguts | Estromatòlits de fa 3,5 milions d’anys (roques sedimentàries formades per l’activitat metabòlica de cianobacteris filamentosos fotosintetitzadors) | No identificats | Més d’1 milió d’anys, en forma d’espores del Precambrià |
| Distribució actual | Mundial | Mundial | Mundial |
El domini dels bacteris

Espiroquetes acumulades al costat d’una Beggiatoa per quimiotaxi (microscòpia electrònica) i, en els requadres a l’esquerra i a la dreta, flagell modificat que recorre tot el cos del microbi i li permet fer el moviment flexuós que el caracteritza.
Francesc Torrella.
Els organismes que pertanyen al domini dels bacteris (Bacteria), coneguts també com eubacteris o bacteris veritables, constitueixen un grup d’éssers vius amb estructura cel·lular procariota, és a dir, desproveïda d’un nucli envoltat de membrana, però molt divers quant a morfologia i molt especialment també quant a fisiologia. Aquest domini inclou la major part de bacteris que han estat objecte d’estudi de la microbiologia clàssica (microbis del sòl, agroalimentaris, amb implicacions mèdiques, etc.). Presenten una gamma de formes i mides molt diversa. Els més petits són els micoplasmes i els nanobacteris, amb una mida que va de 0,3 µm a 0,05 µm respectivament, mentre que el bacteri més gran conegut és Epulopiscium fishelsoni, un bacteri simbiont que viu a l’intestí de peixos del gènere Acanthurus, propis de l’oceà Índic, i que fa 600 µm de llarg i 80 µm d’ample.
Poden presentar una coloració grampositiva o gramnegativa. La tinció de Gram, dita així pel bacteriòleg danès Christian Gram (1853 - 1938), que la desenvolupà el 1884, és un tipus de tinció que permet diferenciar els bacteris en funció de determinats components de la seva paret cel·lular. Els bacteris dits grampositius es visualitzen de color violeta i els gramnegatius de color rosa. També poden tenir diverses morfologies externes, com per exemple morfologia esfèrica (cocs), en bastonet (bacils), en espiral (espirils), lobulada, etc., i es poden presentar de forma aïllada o bé en agregats i en filaments.

Filaments de cianobacteris del gènere Spirulina.
Francesc Torrella.

Beggiatoa (4-5 µm d’ample; microscòpia de contrast de fase i objectiu 100x d’immersió) de fonts d’aigües sulfuroses.
Francesc Torrella.
Actualment, la utilització de tècniques d’anàlisi i seqüenciació de DNA a gran escala de mostres de l’ambient permet la identificació de molts nous grups de bacteris tant aquàtics com del sòl. Aquesta tècnica, anomenada metagenòmica, treballa amb material genètic obtingut directament de mostres, que inclouen fragments dels bacteris que hi viuen i que poden ser identificats. Per exemple, l’anàlisi metagenòmica de mostres del mar dels Sargassos, al mig de l’Atlàntic nord, va permetre identificar la presència d’unes 2.000 espècies bacterianes, 148 de les quals mai no havien estat descrites. Al principi del 2009 es va iniciar l’anàlisi metagenòmica de la Mediterrània, de diverses mostres de sòl i també de dos llacs dels Països Catalans (l’estany de Banyoles i el lac Redon, a la Vall d’Aran).
La diversitat de fonts energètiques que poden utilitzar els eubacteris i de les seves vies metabòliques és molt gran, per la qual cosa ocupen molts nínxols ecològics diferents. N’hi ha d’autòtrofs fotosintetitzadors i quimiosintetitzadors, d’heteròtrofs, d’aeròbics estrictes i facultatius i d’anaeròbics estrictes i facultatius. Els bacteris són molt importants en el manteniment dels cicles del fòsfor, el sofre i el nitrogen a la biosfera, atès que converteixen el gas i els compostos minerals no utilitzables per la majoria d’organismes en compostos que sí poden fer servir. A més, ocupen un lloc predominant en les piràmides ecològiques. Intervenen en les primeres etapes de formació d’un sòl, són font d’aliment per a molts eucariotes unicel·lulars i per a petits metazous, es troben en simbiosi al tub digestiu dels vertebrats i constitueixen també una important font de patògens per a les plantes i els animals. En un gram de sòl, especialment si és d’horta, es poden trobar fins a centenars de milions de bacteris.
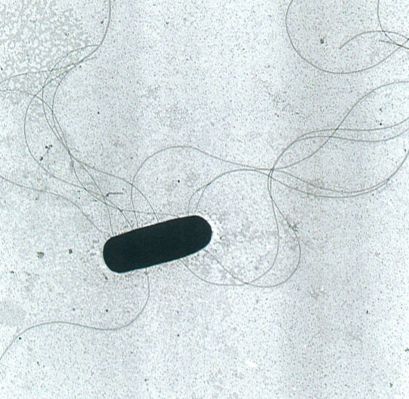
Escherichia coli (microscòpia electrònica de transmissió, forma de cocobacil, amb llargs flagells perítrics que s’emboliquen entre ells. El cos del bacteri fa 1,5-2 µm de llarg).
Francesc Torrella.
Als Països Catalans s’han identificat espècies pertanyents a diversos grups d’aquest domini. En són alguns exemples els cianobacteris (o oxifotobacteris), bacteris fototròfs que alliberen oxigen i que es distribueixen tant en hàbitats terrestres com aquàtics; els anoxifotobacteris, bacteris fototròfs que no alliberen oxigen, típicament bentònics o plantònics, malgrat que algunes espècies poden formar desenvolupaments massius en hàbitats rics en sulfur d’hidrogen, com ara en llacs meromíctics o holomíctics, tapissos microbians, fangs reductors, basses de maresmes i fonts sulfuroses i termals; els beggiatoals, que oxiden compostos de sofre reduït i l’acumulen en forma de grànuls refringents, i que es poden trobar a la superfície de fangs negres, als aiguamolls litorals, a les salines, a les maresmes i a les fonts sulfuroses; els bacteris reductors de sulfats, que habiten en sediments d’aigües dolces i marines, en tapissos microbians, en tubs intestinals i en instal·lacions de depuració d’aigües residuals; les espiroquetes, de morfologia helicoïdal, àmpliament distribuïdes en ambients aquàtics i en el cos d’animals; els bacteris depredadors d’altres bacteris; els bacils i cocs gramnegatius aerobis, que habiten en aigües dolces i marines, en la superfície de peixos i al sòl, i que poden ser patògens d’animals i de plantes; els bacils gramnegatius anaerobis facultatius, molts dels quals són indicadors de contaminació fecal; els cocs grampositius, un gran nombre paràsits o patògens, i d’altres utilitzats en la indústria alimentària; els bacils i cocs grampositius formadors d’endòspores, presents al sòl, en medis aquàtics i al tracte intestinal de mamífers; els bacils irregulars grampositius no formadors d’endòspores, sapròfags del sòl o patògens d’animals i plantes, i els nocardiformes, que són molt freqüents al sòl.
El domini dels arqueus
Els arqueus (Archaea) constitueixen, com els bacteris, un domini d’organismes d’estructura cel·lular procariota, és a dir, desproveïda de nucli envoltat de membrana, molt divers quant a morfologia i fisiologia. També poden presentar coloració grampositiva i gramnegativa a la tinció de Gram, i diverses morfologies externes, com per exemple esfèrica, lobulada, en bastonet, espiral, etc. Es poden trobar aïllats o bé en agregats o en filaments. El seu diàmetre varia entre 0,1 µm i 15 µm, en funció de l’espècie. Fisiològicament poden tenir metabolisme aeròbic, anaeròbic facultatiu o anaeròbic estricte. Aquests darrers inclouen organismes metanogènics, que produeixen gas; halobacteris, que viuen en ambients salins, i termoacidòfils, que viuen en ambients termals o molt àcids.
Malgrat que els arqueus hagin estat tradicionalment considerats com un tipus de bacteri, les dades moleculars els situen filogenèticament més a prop dels eucaris que dels bacteris, per exemple pel que fa a la constitució dels ribosomes, els orgànuls cel·lulars encarregats de descodificar el missatge contingut als gens i sintetitzar les proteïnes corresponents. A més, el DNA s’empaqueta de forma similar a la dels eucariotes, en nucleosomes estructurats al voltant de proteïnes semblants a les histones eucariotes (les proteïnes encarregades d’empaquetar el DNA en tots els organismes eucariotes), i alguns dels seus gens posseeixen introns (zones no codificants intercalades entre les zones codificants), com passa també en els gens dels eucariotes. A més, com a característica distintiva, la major part d’espècies (llevat d’algunes del gènere Thermoplasma) presenten una paret cel·lular que, a diferència de la paret cel·lular dels eubacteris, no conté el peptidoglicà mureïna (àcid muràmic). Tanmateix, com en els eubacteris, el DNA generalment és organitzat en forma d’un únic cromosoma circular.
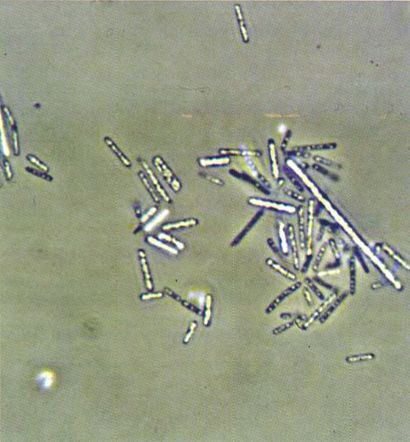
Micrografia de contrast d’una soca pura d’Halobacterium halobium en forma de bacil (5-10 µm de llarg) i amb les cèl·lules plenes de vacúols de gas, que es veuen com a punts refringents. Són halòfils extrems que viuen a les salines.
Francesc Torrella.
Els arqueus van ser identificats originalment en ambients extrems, però des de llavors se n’han trobat en tota mena d’hàbitats; són molt freqüents en les aigües oceàniques, on poden arribar a constituir fins a un 20% de la biomassa microbiana. Respecte als ambients extrems que ocupen, hom sol distingir, sense validesa taxonòmica i sense que coincideixin estrictament amb els grups que genera la filogènia molecular, entre halòfils, termoacidòfils, metanogènics i psicrofílics. Els halòfils només viuen en ambients extremament salins; Halococcus i Halobacterium, per exemple, que només viuen en medis aquosos amb més d’un 12% de sal i poden sobreviure a concentracions de fins el 32% de sal. Els termoacidòfils viuen en ambients aquosos i a temperatures superiors als 60 °C, o bé en ambients molt àcids, amb un grau d’acidesa de pH 1 a 3; així, n’hi ha que es troben en fonts termals, d’on obtenen energia mitjançant l’oxidació del sofre, mentre que d’altres viuen en el compost en combustió; l’arqueu més termòfil que es coneix és Pyrolobus fumarii, que creix a 113 ºC, mentre que el més acidòfil és el gènere Picrophilus, que pot créixer en un pH de -0,06. Els metanogènics viuen en ambients anaeròbics, com per exemple en determinats tipus de sediments i al tracte digestiu d’alguns animals, com els rumiants, i produeixen metà; s’han trobat arqueus metanogènics vius en mostres de gel extretes a tres quilòmetres de fondària en glaceres de Grenlàndia. Finalment, els psicrofílics viuen en ambients extremament freds; per exemple, es calcula que els arqueus constitueixen el 34% de la biomassa de la superfície de les costes antàrtiques.
En l’àmbit dels Països Catalans s’han detectat espècies pertanyents a diversos grups d’arqueus, una de les quals, del gènere Haloarcula, és halòfila, és a dir, viu en ambients salins extrems, com ara en salines i estanys naturals salats. La resta, que pertanyen als gèneres Methanobacterium, Methanobrevibacter, Methanococcus, Methanogenium, Methanoplanus, Methanosarcina, Methanospirillum i Methanothrix són metanogènics i habiten en ambients anòxics, com ara embassaments, zones entollades i dins l’aparell digestiu de remugants. També se’n poden trobar als sòls de boscos, prats i camps d’arròs, en ambients aquàtics i termofílics com les fonts termals, en ambients hipersalins com els tapissos microbians, a l’intestí gros dels humans, a l’intestí d’insectes que mengen fusta, com els tèrmits, i a la cavitat bucal, associats a la placa dental.
El domini dels eucaris

Arbre filogenètic simplificat del domini dels eucaris. Les línies ratllades assenyalen grups la monofília dels quals encara resta dubtosa.
A partir de fonts diverses.
Els organismes del domini dels eucaris (Eukarya) es caracteritzen per ser formats per cèl·lules eucariotes, és a dir, amb un nucli envoltat per una membrana específica que conté el material genètic. A més, el citoplasma és molt estructurat, i presenta diversos orgànuls que compartimenten les activitats metabòliques de la cèl·lula: reticle endoplasmàtic, aparell de Golgi, lisosomes, vacúols, mitocondris, cloroplasts, etc. Tenen una gran diversitat morfològica, des d’organismes unicel·lulars a pluricel·lulars amb teixits veritables. La mida de la cèl·lula eucariota és compresa entre 10 i 100 µm, mentre que la talla dels organismes varia des d’uns pocs µm en els organismes unicel·lulars fins a més de 100 m (algues marines) en alguns pluricel·lulars. Comprenen els organismes més vistosos i per aquest motiu són els que omplen més pàgines en qualsevol història natural, malgrat ser quantitativament els que es troben en menor nombre a qualsevol ecosistema.
Els organismes del domini dels eucaris són presents a tots els hàbitats terrestres, des de les altures a les profunditats marines, i a totes les latituds. Són fonamentalment aerobis i representen una biomassa considerable en tots els ecosistemes. N’hi ha d’autòtrofs fotosintetitzadors, com les plantes, les algues i alguns protists, i d’heteròtrofs, com els fongs i els animals.
En general, els éssers vius que són més familiars per als humans són els eucariotes, atès que són els únics éssers vius que es distingeixen a ull nu de forma individual, especialment els pluricel·lulars. Certament, la major part d’organismes eucariotes que hom coneix sense necessitat de ser un especialista són els pluricel·lulars macroscòpics, formats per milers, milions i àdhuc milers de milions de cèl·lules, però tanmateix també n’hi ha molts, tant d’unicel·lulars com de pluricel·lulars, que passen desapercebuts, atesa la seva mida. No obstant això, si es fan servir els òrgans dels sentits sense cap ajut tècnic, hom ja s’adona que els eucariotes macroscòpics representen una enorme diversitat. Però l’autèntica diversitat dels eucariotes és molt més gran del que a simple vista es pot apreciar. De fet, bona part del 1.200.000 espècies d’eucariotes, aproximadament, que es coneixen resten amagades de la nostra vista, ja que la majoria són organismes microscòpics.
Com en molts altres aspectes de l’arbre de la vida, la utilització de la filogènia molecular ha alterat la concepció clàssica de la filogènia i la taxonomia dels eucariotes, per exemple en demostrar que els fongs i els animals són grups germans amb relació a tots els altres éssers vius. Un altre dels canvis conceptuals respecte a l’arbre de la vida tal com era concebut tradicionalment és la constatació que els protists, considerats per Whittaker un regne, són clarament un grup polifilètic, amb representants a gairebé totes les subdivisions que es fan del domini dels eucaris. Per aquests motius, en la discussió dels principals dades i grups del domini dels eucaris que es farà a continuació, no se seguirà la classificació tradicional en regnes (protists, plantes, fongs i animals), sinó en els grups que actualment es consideren combinant dades morfològiques i sobretot moleculars, seguint un ordre decreixent de magnitud taxonòmica. Tanmateix, i per a una millor situació dels grups que s’esmentaran, s’anirà indicant en quin dels regnes clàssics quedarien englobats.
També cal assenyalar que les filogènies de què es parla aquí no tenen vocació d’exhaustivitat, sinó que es transcriuen únicament per presentar els grans grups d’organismes del domini dels eucaris, amb definicions breus i concises, especialment en aquells grups que ja tenen una clara representació en els anteriors volums d’aquesta obra. A més, per tractar-se precisament d’una història natural, es farà més èmfasi en aquells grups que tenen una major representació en una obra d’aquestes característiques.
Respecte als criteris filogenètics de les agrupacions, cal dir que en molts casos són diversos, principalment moleculars amb participació de diverses molècules però també morfològics. En alguns grups es comentaran explícitament, però en la majoria, atesa la diversitat de criteris, no se’n farà esment. Finalment, un darrer advertiment: constantment es van acumulant dades filogenètiques noves tant sobre l’arbre de la vida com sobre les relacions filogenètiques de grups concrets d’organismes, la qual cosa fa que tots els arbres filogenètics i cladogrames que hom pugui presentar tinguin un cert component intrínsec de provisionalitat, en el sentit de coneixements en creixement i refinament constants. Aquest fet es posarà de manifest en alguns grups en què la relació filogenètica amb els seus possibles grups germans és encara incerta, cosa que es farà clarament explícita quan es discuteixi el grup en qüestió.
Cladograma simplificat on es mostra la ramificació dels uniconts i els biconts dins el llinatge dels eucaris, i la seva relació amb els altres dos dominis, arqueus i bacteris. També hi figuren les principals ramificacions dins els grups dels uniconts i dels biconts. Les línies vermelles indiquen la incorporació de cloroplasts específics per endosimbiosi en determinats llinatges dels biconts i les fletxes vermelles indiquen la procedència d’aquests cloroplasts segons dades moleculars.
Barreras & Creixell, a partir de fonts diverses.
Dins el domini dels eucaris, el primer node que hom ha establert ramifica els eucariotes en dos grans grups, els uniconts (Unikonta) i els biconts (Bikonta), els quals van ser proposats inicialment per Thomas Cavalier-Smith. A nivell morfològic, els uniconts es caracteritzen per tenir, en la seva forma ancestral, un únic flagel, que després molts dels seus descendents van perdre de forma secundària. Dins el grup dels uniconts hom troba els animals, els fongs i les amebes, entre d’altres. Els biconts, d’altra banda, es caracteritzen per tenir dos flagels, si més no en la seva forma ancestral, que molts grups descendents també han perdut de forma secundària. Dins els biconts hom inclou les plantes, les algues i tota la resta de protists. Tanmateix alguns uniconts presenten dos flagels, però aquest és un caràcter secundari d’alguns llinatges concrets.
Des del punt de vista genètic, la distinció entre uniconts i biconts és molt més clara. Així, els uniconts presenten la fusió de tres gens concrets que es troben separats en els biconts i en els organismes procariotes, una característica compartida per tots els uniconts que dóna consistència a aquest grup filogenètic. Els gens que es troben fusionats codifiquen tres enzims implicats en la síntesi de pirimidines (citosina, timina i uracil), uns nucleòtids presents als àcids nucleics (DNA i/o RNA): la carbamoïl fosfat-sintasa, la dihidroorotasa i l’aspartat carbamiltransferasa. Respecte als biconts, a nivell genètic presenten la fusió distintiva d’uns altres dos gens, la timidilat-sintasa (implicada en la síntesi de timidina monofosfat, després de noves fosforilacions en la síntesi de DNA) i la dihidrofolatreductasa (implicada en el metabolisme de l’àcid fòlic), que es troben separats en tots els uniconts i que en conseqüència donen consistència filogenètica al grup dels biconts. Tanmateix, les relacions filogenètiques dins els biconts encara no han estat del tot clarificades.
Malgrat que el grup dels uniconts és acceptat en la confecció general de l’arbre de la vida, no tothom inclou el biconts com a tals, atès que les relacions filogenètiques dins aquest grup encara no han estat del tot clarificades. En els arbres filogenètics en els quals no es considera el grup dels biconts, hom considera llavors uns altres tres grups germans dels uniconts: els arqueplasts, els cromalveolats i els excavats.
Els arqueplasts
Els arqueplasts (Archaeplastida) constitueixen un dels clades principals del domini dels eucaris. Comprenen les plantes terrestres i les algues verdes o clorobionts, les vermelles o rodòfits i un grup petit d’algues microscòpiques d’aigua dolça anomenades glaucòfits. És un grup clarament monofilètic els membres del qual es caracteritzen, entre altres factors, per tenir cloroplasts envoltats de dues membranes, la qual cosa suggereix que procedeixen directament de cianobacteris endosimbionts (els cianobacteris actuals constitueixen un fílum d’eubacteris que es caracteritza per fer fotosíntesi oxigènica). En tots els altres grups d’organismes les cèl·lules dels quals estan proveïdes de cloroplasts, com els cromalveolats, els rizaris i els excavats, es tracta de cloroplasts envoltats de tres o quatre membranes, la qual cosa suggereix que els devien adquirir secundàriament a partir d’algues verdes o roges.
Altres característiques morfològiques distintives del grup dels arqueplasts és que a la major part els manquen els centríols i que els mitocondris tenen una estructura molt característica, amb les crestes mitocondrials planes. A més, la majoria d’arqueplasts també tenen les cèl·lules protegides per una paret de cel·lulosa i emmagatzemen energia en forma de midó, uns caràcters que tanmateix són compartits amb altres grups del domini dels eucaris.
Dins els arqueplasts hom pot distingir dues línies filogenètiques principals. D’una banda, les algues verdes i les plantes terrestres (clorobionts), que es caracteritzen per tenir clorofil·la dels tipus a i b en els cloroplasts. De l’altra, les algues roges (rodòfits), els cloroplasts de les quals contenen clorofil·la a però no clorofil·la b, i que també contenen un altre tipus de pigments, les ficobiliproteïnes, entre els quals cal destacar les ficoeritrines i les ficocianines, que els donen el seu color característic. Els pigments fotosintetitzadors de les algues roges són presents també en la majoria de cianobacteris. Dins aquest esquema filogenètic, la posició dels glaucòfits no és encara prou clara, atès que també presenten els pigments fotosintetitzadors típics de les algues roges (i dels cianobacteris), però en canvi tenen una inusual paret cel·lular dins els cloroplasts, semblant també a la dels cianobacteris.
Les plantes verdes
El grup de les plantes verdes, més pròpiament dit dels clorobionts (Chlorobionta) o viridòfits (Viridophytae), inclou un ampli ventall d’organismes fotosintetitzadors que es caracteritzen per tenir clorofil·les a i b com a pigments fotosintetitzadors, emmagatzemar els productes de la fotosíntesi en forma de midó dins de cloroplasts envoltats d’una doble membrana i tenir cel·lulosa a les parets cel·lulars. Dins d’aquest grup hom ha identificat diversos milers d’espècies d’algues verdes i diversos centenars de milers de plantes terrestres.

Les falgueres pertanyen al clade dels embriòfits, que reuneix totes les plantes terrestres.
Fototeca / MC.

Arbre filogenètic de les plantes verdes o clorobionts i, a baix, característiques principals dels diferents grups.
A partir de fonts diverses / Miquel Alonso.
Així, hom distingeix tres llinatges essencials de plantes verdes d’acord amb la taxonomia molecular: el dels prasinòfits, format íntegrament per algues verdes; el que inclou les clorofícies, les trebouxiofícies i les ulvofícies, també format íntegrament per algues verdes, i el dels estreptòfits, que inclou algues verdes fortament emparentades amb les plantes terrestres i les mateixes plantes terrestres o embriòfits (plantes vasculars).

Arbre filogenètic dels embriòfits (plantes terrestres) i característiques principals dels diferents grups. Els noms assignats als clades encara no són noms científics establerts formalment.
a partir de fonts diverses.
Dins els clorobionts, els organismes tradicionalment més representats en una història natural són els embriòfits (Embryophyta), per la qual cosa s’aprofundirà una mica més en la filogènia concreta d’aquest grup. Els embriòfits són el clade format pels descendents de certes algues verdes, i es caracteritzen per posseir una sèrie d’adaptacions per a la vida fora de l’aigua. Aquest clade comprèn totes les plantes terrestres: les hepàtiques (marcantiòfits), les antocerotes, les molses, els licòfits (licopodis, isòets i selaginel·les), les falgueres i les plantes amb llavor, entre les quals hi ha les gimnospermes i les angiospermes. La monofília dels embriòfits és àmpliament demostrada tant per dades morfològiques com moleculars.
Tradicionalment, els embriòfits han estat classificats en dos grans grups: els briòfits en sentit ampli, que inclourien les hepàtiques, les antocerotes i les molses; i les plantes vasculars, que inclourien les falgueres, les gimnospermes i les angiospermes. Tanmateix, les anàlisis de filogènia molecular han demostrat que les molses són un grup parafilètic pel que fa a les plantes vasculars, si bé les seves relacions filogenètiques precises encara estan en discussió.
Els rodòfits

Les algues roges o rodòfits reuneixen prop de 6.000 espècies a tot el món. Tot i que s’admet la seva condició d’arqueplasts, es continua debatent el seu grau de distanciament dels clorobionts.
Josep M. Barres.
Els rodòfits (Rhodophyta), o algues roges, constitueixen un llinatge específic dins els arqueoplasts que es caracteritza per la presència de pigments fotosintetitzadors accessoris, concretament ficoeritrina, ficocianina i al·loncocianines, els quals s’agrupen en complexos proteics anomenats ficobilisomes que s’ancoren a la membrana til·lacoidal dels cloroplasts; l’absència de flagels i centríols, i la presència de midó com a biomolècula de reserva. Habiten totes les mars i els oceans, preferentment en àrees costaneres i de plataforma continental, incloses les costes dels Països Catalans. Són uns importants productors primaris, i proporcionen l’hàbitat estructural per a moltes altres espècies marines.
Els glaucòfits
Finalment, els glaucòfits (Glaucophyta) comprenen un grup d’algues d’aigua dolça que es caracteritzen per la presència de cloroplasts que mantenen moltes de les característiques típiques dels cianobacteris, dels quals procedeixen per endosimbiosi, com per exemple, una paret residual amb peptidoglicans i la presència de carboxisomes, uns compartiments típicament bacterians que contenen enzims implicats en la fixació de carboni, la qual cosa permet fer la hipòtesi que els glaucòfits representen el grup d’organismes més semblant a l’ancestral que originàriament va establir una endosimbiosi amb un cianobacteri.
Una altra característica pròpia d’aquest clade és la presència de betacarotens, zeaxantines i B-criptoxantina com a pigments fotosintetitzadors accessoris, a més a més de la clorofil·la a. Tots els glaucòfits són unicel·lulars; tanmateix n’hi ha de mòbils flagel·lats, els quals són mancats de paret cel·lular; d’immòbils que conserven flagels vestigials molt curts i que tenen una paret cel·lular de cel·lulosa; i d’altres que durant el cicle vital passen per fases mòbils i immòbils i que presenten una paret cel·lular no cel·lulòsica.
Els uniconts
Els uniconts (Unikonta) constitueixen un altre dels quatre grans llinatges en què es divideix el domini dels eucaris. Es caracteritzen pel fet que presenten tres gens específics fusionats (els corresponents a tres enzims implicats en la síntesi de nucleòtids pirimidínics: la carbamoïl fosfat-sintasa, la dihidroorotasa i l’aspartat carbamiltransferasa). Dins els grup dels uniconts resten inclosos els metazous (animals), els coanoflagel·lats, els fongs i els amebozous, entre d’altres. La monofília dels uniconts encara no està completament demostrada.
Es reconeixen dos grans grups dins els uniconts: els amebozous (Amoebozoa) i els opistoconts (Opisthokonta), uns grups clarament monofilètics el darrer dels quals inclou els animals i els fongs, juntament amb alguns protists filogenèticament molt emparentats amb els animals. La principal característica dels opistoconts és que presenten, com a mínim en algun moment del cicle biològic, cèl·lules flagel·lades, com per exemple, els espermatozous dels animals o les espores dels quitridiomicots o quítrids (Chytridiomycota), un tipus de fongs, o durant tota la seva vida, com en els coanoflagel·lats.
Els metazous
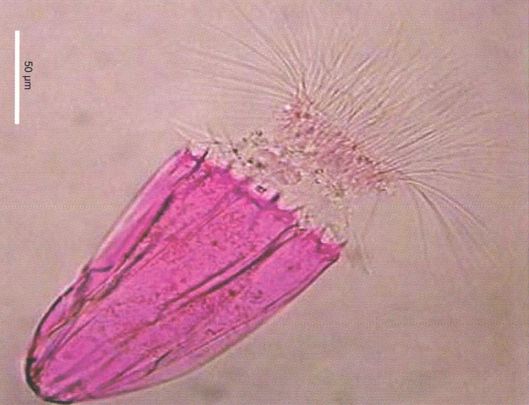
Els loricífers de les cubetes anòxiques hiperhalines de les profunditats de la Mediterrània (a la fotografia, Spinoloricus, amb tinció) són els primers metazous coneguts que viuen en condicions del tot anòxiques. Estudis de traçadors radioactius, anàlisis bioquímiques, microanàlisis de raig X i espectroscòpia d’infrarojos, microscòpia electrònica de transmissió i d’escombratge en ultraseccions, han permès demostrar adaptacions específiques, com la falta de mitocondris i un gran nombre d’orgànuls associats a procariotes endosimbiòtics.
Danovaro i altres, 2010.
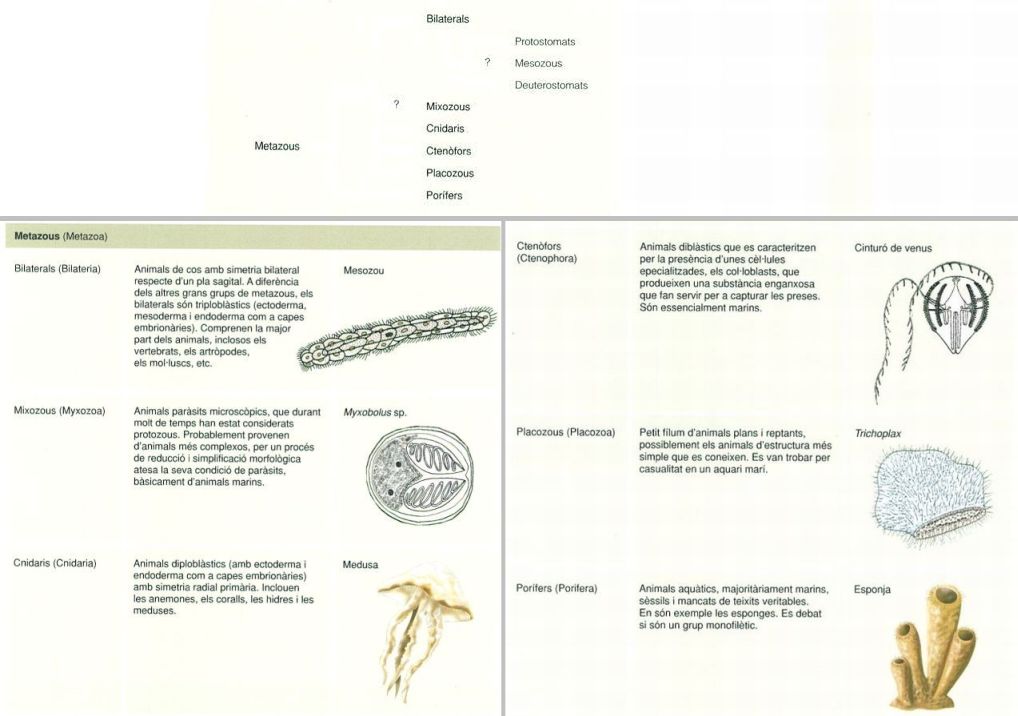
Els metazous (Metazoa) constitueixen un grup monofilètic d’organismes pluricel·lulars les cèl·lules dels quals es troben diferenciades en teixits i òrgans amb funcions específiques, i que generalment tenen una cavitat digestiva i un sistema nerviós. Les filogènies moleculars permeten dividir aquest clade en diversos grups, la major part dels quals són també, sense cap mena de dubte, monofilètics.
Els animals bilaterals (Bilateria) es classifiquen, segons dades combinades de filogènia clàssica i filogènia molecular, en dos grans grups: protostomats i deuterostomats.
Els metazous protostomats
Els protostomats (Protostomata) es caracteritzen pel fet que la boca de l’adult deriva del blastòpor embrionari (el blastòpor és l’obertura que s’origina durant la fase embrionària de gastrulació, en la qual es forma un cavitat interna que prefigura la futura llum del sistema digestiu), i inclouen tots els invertebrats bilaterals, llevat dels equinoderms, els hemicordats, els urocordats i els cordats. Els animals deuterostomats (Deuterostomata), en canvi, es caracteritzen per tenir una boca de nova formació, que no es correspon amb el blastòpor embrionari, el qual genera l’anus de l’adult. Es reconeix un tercer grup de bilaterals, els mesozous (Mesozoa), uns animals petits, de forma de vida paràsita (parasiten animals marins), d’assignació i classificació encara incerta. Així, es discuteix si els mesozous són bilaterals molt primitius o, en canvi, platihelmints morfològicament reduïts i simplificats. Tanmateix sí que és clar, atenent a la filogènia molecular, que són polifilètics, formats per dos grups filogenèticament no emparentats.
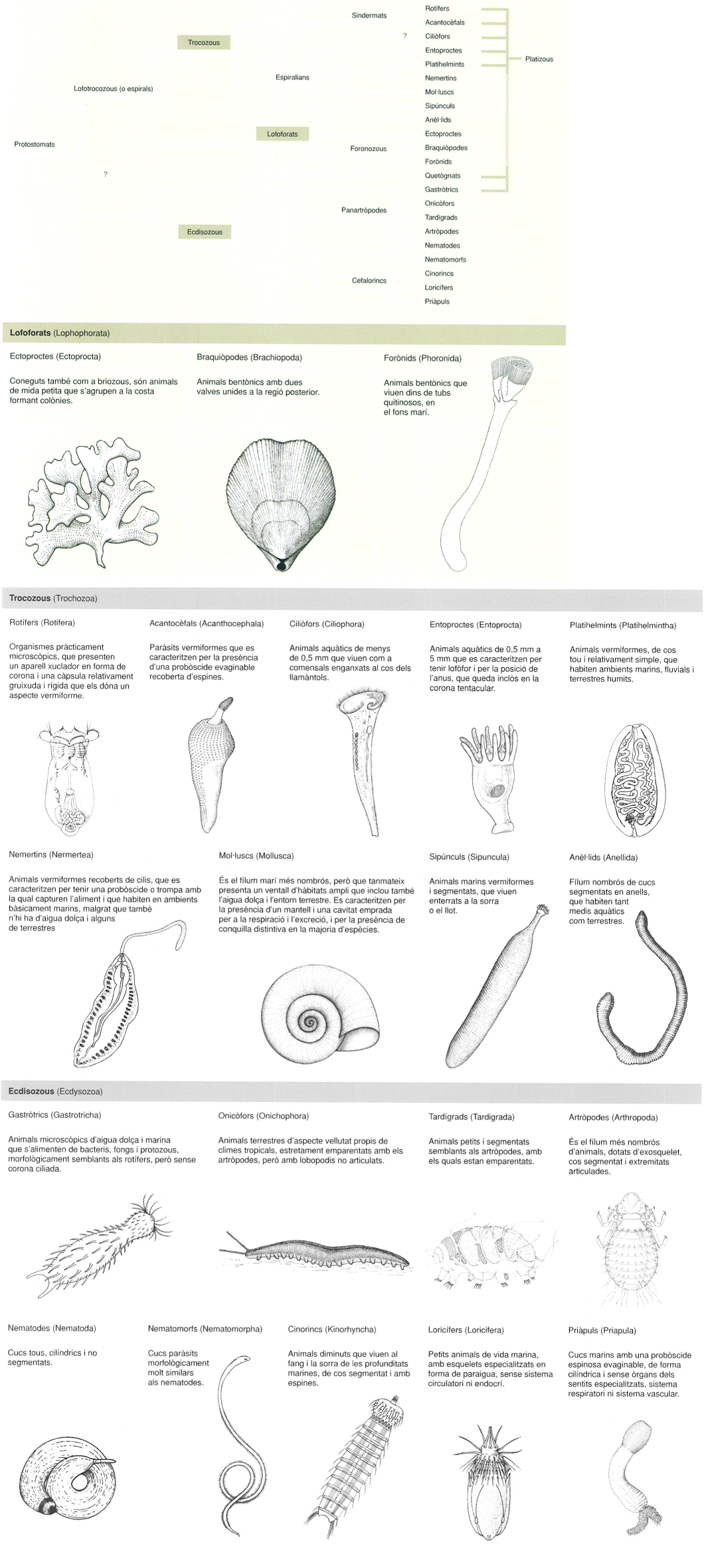
Arbre filogenètic dels protostomats i les característiques principals dels clades destacats amb un requadre. Els dibuixos no són a escala.
A partir de fonts diverses / Jordi Corbera, Lluís Centelles, Biopunt, Amadeu Blasco, Ray Gibson i Sedgwick.
Arbre filogenètic dels metazous i característiques principals dels diferents grups.
A partir de fonts diverses / Josep L. Ferrer, Jordi Corbera i Biopunt.
Els protostomats, al seu torn, es classifiquen, segons les dades de la filogènia molecular, en dos grans grups, els lofotrocozous (Lophotrocozoa) i els ecdisozous (Ecdysozoa), dins els quals queden agrupats tots els protostomats, llevat dels quetògnats, cucs marins depredadors que són un dels principals components del plàncton en totes les mars i oceans, de classificació encara incerta. Els lofotrocozous constitueixen un tàxon monofilètic introduït el 1995 a partir de dades moleculars, i comprèn dos grans grups: els lofoforats (Lophophorata), que es caracteritzen per la presència de lofòfor, un òrgan anular format per tentacles ciliats que envolten la boca; i els trocozous (Trochozoa), que es caracteritzen per presentar una larva trocòfora durant el desenvolupament embrionari, una larva marina plantònica nedadora. Els ecdisozous són també un grup monofilètic, definit el 1997 basant-se en dades de filogènia molecular. Aquest grup se sosté igualment per caràcters morfològics, i es pot considerar que inclou tots els animals que muden l’exosquelet.
Tanmateix, el 1998 Thomas Cavalier-Smith (1942-) va proposar un nou grup de protostomats, els platizous (Platyzoa), que inclou alguns dels grups prèviament classificats com a lofotrocozous, un grup concret d’ecdisozous, i els quetògnats, un grup que les dades de filogènia molecular també situen com a monofilètic, la qual cosa genera una aparent incongruència filogenètica que encara s’està debatent.
Els metazous deuterostomats
Finalment, l’altre gran grup d’animals bilaterals és el dels deuterostomats (Deuterostomata), els quals tenen com a característica morfològica comuna el fet de tenir una boca de nova formació, i que el blastòpor esdevé l’anus. Tanmateix, la filogènia molecular ha establert també el seu origen monofilètic, la qual cosa dóna consistència a aquest grup des del punt de vista de llinatges evolutius. Els deuterostomats representen també els animals morfològicament més complexos.
Els coanoflagel·lats

Arbre filogenètic dels deuterostomats i característiques principals dels diferents grups.
A partir de fonts diverses.
Els coanoflagel·lats (Choanoflagellata) són organismes de vida lliure, unicel·lulars o colonials, que es troben en tots els ambients aquàtics, tant marins com d’aigua dolça. Tenen forma ovoide o esfèrica, amb un únic flagel apical envoltat per una corona de 30-40 microvil·lis (prolongacions microscòpiques de la membrana cel·lular). El moviment del flagel els permet desplaçar-se i, alhora, atrapar aliment amb la corona de microvil·li, bàsicament bacteris i detritus.

Filera de coanoflagel·lats sèssils subjectes a una colònia d’algues planctòniques. S’hi aprecien les coanes, on acumulen els bacteris de què s’alimenten, i el gros flagell central que en sobresurt. El punt blanc que s’hi distingeix en cada un és el petit vacúol pulsàtil que expulsa les restes de la digestió.
Francesc Torrella.
Una altra característica pròpia d’aquest grup és que la distribució dels orgànuls cel·lulars al seu interior és constant, i sovint la seva membrana cel·lular és envoltada per una matriu cel·lular o periplast.
N’hi ha de nedadors i de sèssils, els quals s’uneixen al substrat pel periplast o bé per una prolongació específica anomenada pedicel. La filogènia basada en l’RNA 18S i la comparació de gens nuclears i mitocondrials situa els coanoflagel·lats com a grup germà dels metazous.
Els filasteris
Els filasteris (Filasterea) són un grup d’eucariotes unicel·lulars de difícil assignació filogenètica. Comprèn dos grups molt concrets d’organismes opistoconts emparentats, uns que s’alimenten de bacteris i els altres que són paràsits i viuen a l’hemolimfa del cargol Biomphalaria glabrata. Les anàlisis filogenètiques els emparenten amb els animals i els coanoflagel·lats, però no els situen pas dins de cap d’aquests grups, per la qual cosa se situen dins un tàxon propi.
Els ictiosporis
Els ictiosporis (Ichthyosporia), anomenats també mesomicetozous, són un petit clade monofilètic de protists majoritàriament paràsits de peixos i altres animals. Clàssicament eren agrupats dins el grup de les algues, els fongs o els protozous, però les anàlisis filogenètiques han demostrat la seva monofília i el seu parentesc amb els animals i els coanoflagel·lats, dels quals serien un grup germà, i també dels fongs.
Els fongs

Llevat del pa (una soca de Saccharomyces cerevisiae), vist al microscopi òptic (objectiu de 100x d’immersió), tenyit amb el colorant blau de metilè. Les cèl·lules tenen una mida variable, entre 5 i 10 µm.
Francesc Torrella.
Els fongs (Fungi), que formen un llinatge monofilètic d’organismes heteròtrofs amb representantsunicel·lulars i pluricel·lulars, han estat situats per la filogènia molecular (utilitzant entre altres molècules la subunitat petita de l’RNA ribosòmic) com a grup germà dels animals i dels coanoflagel·lats. Comparteixen amb els animals la facultat d’exportar enzims hidrolítics des de l’interior de les cèllules per a degradar els biopolímers, els quals són absorbits com a elements nutritius. Tanmateix, a diferència dels animals, els fongs no degraden l’aliment al seu interior, sinó que en fan una digestió externa.
També comparteixen amb els animals el fet de tenir estructures quitinoses, emmagatzemar l’energia en forma de glicogen i tenir un triplet especial al seu RNA missatger per codificar el triptòfan mitocondrial (UGA). Tanmateix, algunes de les relacions filogenètiques concretes dins el clade dels fongs encara estan en discussió, atesa la gran diversitat morfològica que presenten, des dels anomenats fongs superiors (o més pròpiament anomenats dicaris, Dikarya) fins a les floridures i els diversos tipus de fongs unicel·lulars, com els llevats o els quítrids, entre d’altres.
Els amebozous

Ameba de morfologia típica, pertanyent al grup dels maiorèl·lids (Mayorella), que viu als fangs actius de depuradora. La cèl·lula fa uns 80-90 µm d’envergadura, però molt més (fins a 200 µm) quan s’estira per viatjar. A la imatge (contrast de fase), s’hi distingeixen bé els lobopodis, el nucli i els vacúols digestius.
Francesc Torrella.
Els amebozous (Amoebozoa) són el principal grup de protozous ameboides, la major part dels quals presenten mobilitat deguda a fluxos interns de citoplasma. Els pseudopodis (o lobopodis) són roms i digitats. La majoria són unicel·lulars, i habiten principalment al sòl i als hàbitats aquàtics, malgrat que se’n coneixen també de paràsits. També n’hi ha de pluricel·lulars i de sincicials (una sola massa citoplasmàtica amb diversos nuclis), els quals tenen l’aspecte d’una massa mucilaginosa.
Els cromalveolats
Els cromalveolats (Chromalveolata) formen un grup d’organismes la posició filogenètica dels quals ha canviat diverses vegades en el decurs de la història de les filogènies, a mesura que s’anaven refinant i ampliant els sistemes taxonòmics i filogenètics. Així, històricament, quan els éssers vius es classificaven únicament de forma binària en animals o plantes, els cromalveolats coneguts eren considerats plantes, atesa la paret cel·lular que tenen les seves cèl·lules i els sistemes fotosintètics dels seus cloroplasts.
Posteriorment, amb la proposta de cinc regnes de Robert Whittaker, molts cromalveolats van ser englobats dins el regne dels protists, mentre que d’altres van ser inclosos dins el regne dels fongs, com els oomicets (Oomycetes) (organismes semblants als fongs, entre els quals s’inclou el míldiu) i els labirintulomicets (Labyrinthulomycetes) (organismes majoritàriament marins paràsits o descomponedors que formen xarxes de tubs filamentosos que fan servir de pistes per les quals les cèl·lules es desplacen per absorbir els nutrients). Més recentment, Cavalier-Smith va proposar la creació d’un nou regne per a aquests organismes, que s’hauria d’afegir als cinc regnes clàssics de Whittaker.
Tanmateix, encara es debat avui dia si els cromalveolats són monofilètics o polifilètics, una distinció important per a les anàlisis cladístiques. El motiu d’aquest debat és que els diversos grups de cromalveolats no tenen gaires característiques morfològiques comunes entre ells, llevat que els seus cloroplasts comparteixen el mateix origen i que la majoria de les cèl·lules tenen parets cel·lulars amb cel·lulosa. No obstant això, com ja s’ha dit, cada subgrup essencial de cromalveolats té certs trets únics. Hom en distingeix cinc llinatges: els cercozous, els foraminífers, els radiolaris, els alveolats i els heteroconts o estraminòpils.
Els cercozous
Els cercozous (Cercozoa) són un grup monofilètic de protozous principalment heteròtrofs, especialment abundants al sòl de terra ferma, on són els eucariotes més nombrosos, però que també es troben en ambients aquàtics d’aigua dolça i marina. També n’hi ha de paràsits, sobretot de plantes, d’animals invertebrats i d’altres protists. Els cercozous de vida lliure s’alimenten de bacteris, de fongs, d’algues, d’altres protozous i fins i tot d’animals microscòpics. Probablement es tracta dels depredadors més nombrosos de terra ferma. Morfològicament n’hi ha d’ameboides, de flagel·lats i d’ameboflagel·lats.
Els foraminífers

El foraminífer Globigerina, probablement el més comú dels protozous marins proveïts de closca calcària. L’exemplar fotografiat és viu, i mostra els filopodis estesos. S’aprecien petites partícules d’aliment que són transportades cap a la cèl·lula.
Francesc Torrella.
Els foraminífers (Foraminifera) són organismes gairebé exclusivament marins i generalment ben tònics, bé que també hi ha formes planctòniques. Són proveïts d’una closca calcària, que pot ésser formada per una o per diverses cambres, i pot presentar una sola obertura o nombrosos foradets, pels quals surten uns rizopodis molt delicats (estructures filamentoses que s’encreuen formant una xarxa).
Els radiolaris
Els radiolaris (Radiolaria) constitueixen un grup de protozous ameboides que es caracteritzen per estar proveïts d’una intricada càpsula central perforada, que divideix la cèl·lula en seccions externes i internes, anomenades ectoplasma i endoplasma, al centre del qual hi ha el nucli cel·lular. Llevat de casos excepcionals, tenen un exosquelet, generalment de sílice, el qual s’acumula en les malles de la xarxa ectoplasmàtica que els envolta. Entre els forats de l’exosquelet surten pseudopodis rectes, anomenats reticulopodis. Viuen aïllats o en colònies i constitueixen un important element del plàncton marí. La seva alimentació consisteix en flagel·lats, diatomees i petits crustacis vius.
Els alveolats

Dins el mateix clade dels alveolats se situen organismes antigament considerats ben diferents, com ara els dinoflagel·lats, que es classificaven entre les algues, i els ciliats, que es trobaven entre els protozous. Els dinoflagel·lats (Ceratium, a dalt) són abundants al plàncton marí (encara ara considerats dins el fitoplàncton). Els paramecis (a baix) viuen en aigues dolces eutrofitzades de llacunes i tolls, i també en basses de llacunatges, on amb la seva alimentació contribueixen a l’eliminació de petits microorganismes (bacteris, algues unicel·lulars).
Francesc Torrella.
Els alveolats (Alveolata) són un grup monofilètic d’organismes eucariotes unicel·lulars que han adoptat tipus extremament diversos de nutrició, tant autòtrofa per fotosíntesi com heteròtrofa per depredació i parasitisme intracel·lular. Les principals sinapomorfies dels alveolats són la presència de sacs membranosos anomenats alvèols, situats sota la membrana plasmàtica, i la presència de microporus a la superfície cel·lular, amb funció pinocítica (incorporació de líquids orgànics de l’exterior com a font d’aliment). Els alveolats es classifiquen en apicomplexos, dinoflagel·lats i ciliats.
Els apicomplexos (Apicomplexa) són paràsits obligats, i presenten un orgànul específic que els facilita l’acoblament al seu hoste. Aquest orgànul, una mena de complex apical, és format per un conus de tubulina (una proteïna) anomenat conoide, que serveix de bastida a un orgànul evaginable anomenat roptria. Els apicomplexos infecten principalment cèl·lules animals, des de cèl·lules intestinals d’insectes i invertebrats marins fins als glòbuls rojos dels primats, incloent-hi els humans. Un exemple molt conegut és el del plasmodi, l’agent que causa la malària.
La majoria dels dinoflagel·lats (Dinoflagellata) són depredadors de vida lliure o organismes fotosintetitzadors propis d’ambients aquàtics. Molts dels fotosintetitzadors són també consumidors de bacteris i d’altres petits organismes eucariotes, un tipus de nutrició combinada que s’anomena mixotròfia. Les espècies fotosintetitzadores i mixotròfiques fan un paper molt important en els cicles oceànics del carboni, i si són presents en grans quantitats produeixen les anomenades marees roges. Dues característiques típiques dels dinoflagellats són que tenen el citosquelet diferent del de qualsevol altre eucariota i que presenten els cromosomes sempre condensats.
Els ciliats (Ciliata) són predadors que s’alimenten bàsicament de bacteris. Molts són de vida lliure, però també n’hi ha al tracte intestinal dels animals i als músculs d’alguns peixos. La principal sinapomorfia dels ciliats és la presència de dos nuclis heteromòrfics i d’un còrtex cel·lular que conté molts cilis, alineats en configuracions específiques.
Els heteroconts
Els heteroconts (Heterocontha), anomenats també estraminòpils (Straminopila), són un dels llinatges principals dels eucariotes, amb més de 100.000 espècies conegudes. Formen un grup morfològicament heterogeni, per bé que filogenèticament relacionat. Inclouen les algues brunes (feòfits), pluricel·lulars i fotosintetitzadores; les diatomees, algues unicel·lulars encapsulades en una coberta de sílice i que són un dels principals components del plàncton marí; i els oomicets, uns organismes filamentosos superficialment semblants als fongs entre els quals s’inclou el que causa el míldiu, una malaltia que afecta algunes plantes i que els causa lesions característiques a les fulles. Una de les característiques comunes de la major part d’estraminòpils és que presenten flagels, generalment dos de desiguals, o bé produeixen cèl·lules fiagellades en algun moment del cicle biològic, com per exemple, gàmetes o zoospores (espores asexuals mòbils).
Els hacrobis
Finalment, els hacrobis (Hacrobia) constitueixen un grup que comprèn una gran diversitat d’eucariotes microscòpics, incloent-hi tant organismes autòtrofs com heteròtrofs, com per exemple, microalgues, organismes flagel·lats i ameboides. Malgrat que no tenen característiques morfològiques comunes, la filogènia molecular els situa com a grup monofilètic, probablement descendent d’un avantpassat comú fotosintetitzador que va passar per una endosimbiosi secundària amb una alga vermella.
Els excavats

Fragment de l’arbre filogenètic dels metazous que recull la branca dels excavats.
A partir de fonts diverses.
Els excavats (Excavata) constitueixen el darrer dels quatre grans grups del domini dels eucaris, un clade format bàsicament per protists. Inclou organismes de vida lliure i simbionts, i també alguns paràsits, que poden afectar, entre altres organismes, els humans. A molts excavats els manquen els mitocondris típics, però la majoria tenen un orgànul mitocondrial modificat. També n’hi ha que presenten mitocondris amb crestes tubulars, discoïdals i, de vegades, planes. La majoria tenen diversos flagels (típicament dos, quatre o més de quatre), i molts presenten un solc ventral digestiu ciliat que utilitzen per a capturar i fagocitar partícules d’aliment. Tots són unicel·lulars, però n’hi ha alguns que es poden agrupar en petites comunitats, les quals tanmateix no esdevenen un organisme pluricel·lular en sentit estricte.
El grup dels excavats també inclou organismes que no presenten cap d’aquestes característiques, però que tanmateix queden dins el mateix grup segons les filogènies moleculars. Aquest és un dels principals motius pels quals encara es debat si els excavats representen un clade monofilètic o un grup polifilètic o parafilètic. En aquest sentit, els arbres filogenètics situen alguns excavats com els eucariotes més primitius, la qual cosa permetria fer pensar que podrien ser un grup parafilètic. Tanmateix, les anàlisis multigèniques de filogènia molecular assenyalen que la major part de grups dels excavats tenen un origen monofilètic.
Els excavats es classifiquen en set grups: els malawimonàdids, els euglenozous, els heterolobosis, els jacòbids, els parabasàlids, els fornicats i els preaxostils o anaeromònades.
Els malawimonàdids
Els malawimonàdids (Malawimonadidae) són un grup de petits organismes unicel·lulars flagel·lats que comprèn un únic gènere amb només dues espècies conegudes, que viuen en sediments d’aigua dolça i al sòl. Desconeguts fins el 1990, són un grup filogenèticament significatiu atès que els arbres filogenètics els situen a la base dels excavats, per la qual cosa ocupen una posició clau per a entendre’n l’origen. Són organismes heteròtrofs de vida lliure, de no més de 5 µm de longitud. La seva forma recorda una mongeta, amb dos flagels i un solc ventral que utilitzen, com altres excavats, per a recollir i fagocitar petites partícules d’aliment, que són transportades gràcies al corrent generat pel flagel posterior.
Els euglenozous
Euglenes vistes amb microscopi de contrast de fase. La de la fotografia (Euglena polymorpha) s’estira i s’arronsa en nedar. La majoria de les euglenes no fan més d’uns 40 µm de llarg, però algunes poden arribar a mig mil·límetre.
Francesc Torrella.
Els euglenozous (Euglenozoa) són un grup monofilètic format per organismes unicel·lulars flagel·lats que presenten diversos tipus de nutrició, incloent-hi la depredació, el parasitisme i el comensalisme, l’autotròfia per fotosíntesi i l’alimentació per osmosi. Les principals sinapomorfies d’aquest grup són la presència de filaments paraxonèmics heteromòrfics (flagels relativament gruixuts) i extrosomes tubulars (estructures embolcallades de membrana que, en determinades condicions, descarreguen el seu contingut a l’exterior de la cèl·lula).
Els euglenozous depredadors es troben filogenèticament molt distribuïts dins aquest grup, i mostren una àmplia diversitat d’estructures quant a l’aparell i les estratègies d’alimentació i les preferències de preses. Per exemple, algunes espècies depredadores s’alimenten de bacteris, mentre que altres consumeixen preses més grosses, com cèl·lules eucariotes, les quals fagociten senceres o bé foraden per aspirar-ne el contingut.
Els euglenozous depredadors són un element important de la biota microbial en molts sediments de superfície. Els euglenozous fotosintetitzadors, al seu torn, representen un clade específic d’aquest grup, i es van originar a partir d’una endosimbiosi secundària entre una euglena primitiva i una cèl·lula d’una alga verda. En canvi, els euglenozous paràsits i comensals s’han originat diverses vegades dins aquest grup, com a adaptacions alimentàries secundàries.
Els tripanosomes formen un grup monofilètic considerat actualment dins dels euglenozous.
Cristina Riera i Roser Fisa.
Tanmateix, els euglenozous paràsits i els fotosintetitzadors comparteixen uns quants trets característics del seu citosquelet, associats a l’aparell flagel·lar i d’alimentació. Dins aquest grup s’inclouen, per exemple, els tripanosomes, que provoquen malalties importants, com la malaltia de la son, la leishmaniosi i la malaltia de Chagas.
Finalment, els euglenozous que s’alimenten per osmosi són mancats d’aparell d’alimentació i absorbeixen els nutrients directament del medi on es troben.
Els heterolobosis
El grup dels heterolobosis (Heterolobosea), o percolozous, es caracteritza per la forma ameboide dels organismes que el formen, els quals es mouen formant protuberàncies eruptives; per realitzar mitosis intranuclears sense divisió del citoplasma que incrementen les dotacions cromosòmiques; per presentar, en molt casos, una fase temporal flagellar, i per tenir les crestes mitocondrials planes, sovint agregades en disc, i un aparell de Golgi poc convencional no organitzat en piles membranoses. Viuen principalment en aigües dolces i marines, i cada gènere és específic d’un d’aquests dos tipus d’hàbitat, llevat d’unes poques excepcions més generalistes. També s’han descrit quatre espècies d’heterolobosis extremòfils, que habiten en zones termals àcides o bé en ambients hipersalins.
Els jacòbids
Els jacòbids (Jakobida) són un petit grup de protists flagel·lats heteròtrofs de vida lliure. La seva principal característica és la presència de dos flagels que s’insereixen a prop d’un dels extrems de l’organisme, un dels quals s’orienta en sentit posterior i va associat a un solc conspicu amb funcions alimentàries i que ocupa la major part d’una de les superfícies cel·lulars (per conveni es diu que és la ventral); aquest flagel proporciona un corrent d’aigua que impulsa les partícules d’aliment cap al solc.
Una altra característica distintiva d’aquest grup és la presència d’un cromosoma mitocondrial molt semblant als cromosomes bacterians. Així, doncs, en la gran majoria d’eucariotes el cromosoma mitocondrial es troba reduït aproximadament a una vintena de gens, un reducte evolutiu del procés d’endosimbiosi que va generar aquest orgànul cel·lular. En canvi, el cromosoma mitocondrial dels mitocondris dels jacòbids conté gairebé un centenar de gens, organitzats de forma molt semblant als cromosomes proteobacterians.
Els parabasàlids
Els parabasàlids (Parabasalia) són un grup de protozous flagel·lats la majoria dels quals simbiòtics amb animals. Se n’han trobat diversos que viuen a l’intestí dels tèrmits i dels escarabats, els quals ajuden a digerir la fusta, i alguns presenten bacteris simbionts dins seu. També se n’han descrit de patògens, per exemple, en humans. Els organismes d’aquest grup tenen els flagels ordenats en un o més grups que es localitzen a prop de l’extrem anterior de la cèl·lula. Són mancats de mitocondris, per la qual cosa tenen un metabolisme anaerobi, i tampoc no tenen el solc típic dels excavats, però totes dues pèrdues són secundàries, i comparteixen avantpassats comuns amb la resta d’excavats, els quals tenien tant mitocondris com el solc ventral ciliat amb funcions digestives.
Els fornicats
Els fornicats (Fornicata) constitueixen un grup de protists unicel·lulars heteròtrofs i flagel·lats amb un o dos cariomastigonts per cèl·lula (estructures cel·lulars de suport a flagels), cadascun dels quals suporta entre un i quatre flagels. Un dels flagels s’orienta enrere, cap al solc ventral típic dels excavats. Els fornicats exhibeixen unes quantes característiques inusuals, com l’absència de mitocondris clàssics. N’hi ha que són mancats completament d’aquests orgànuls, mentre que d’altres presenten mitocondris molt reduïts, anomenats mitosomes. Els fornicats que no tenen mitocondris tenen un metabolisme anaerobi, mentre que els que tenen mitosomes són microaerobis. Aquesta característica fa que durant un temps es pensés que eren els organismes basals del domini dels eucaris, però tanmateix les dades moleculars assenyalen que aquest caràcter és derivat, per simplificació d’organismes ancestrals. N’hi ha de vida lliure i de paràsits, entre els quals cal destacar Giardia intestinalis, un important paràsit de l’intestí humà. El seu origen monofilètic és clar, d’acord amb dades estrictament moleculars.
Els preaxostils
Finalment, el darrer gran clade dels excavats són els preaxostils (Preaxostyla) o anaeromònades (Anaeromonada), un grup de protists anaerobis o microaerobis que, com els fornicats, són mancats de mitocondris o bé presenten mitocondris molt reduïts (mitosomes). La sinapomorfia morfològica característica d’aquest grup és la presència de preaxostil, una banda de microtúbuls que van des de la base del flagel fins a l’altre extrem de la cèl·lula. Les anàlisis de filogènia molecular indiquen que és un grup monofilètic.