Introducció a la taxonomia, la sistemàtica i la filogènia
Les espècies

En la desena edició de Systema Naturae (‘Sistema natural’), del 1758, Carl von Linné va incloure la classificació de més de 4.000 espècies d’animals i més de 7.000 de plantes, en les quals, a més d’utilitzar els noms emprats en aquell moment, va afegir els noms binomials, que a poc a poc es van anar imposant entre la comunitat científica i que avui són tan familiars.
Per a poder establir relacions entre les diferents espècies i entre els grups taxonòmics d’ordre superior, com els gèneres, les famílies, els ordres, etc., primer cal agrupar i classificar els éssers vius. La unitat bàsica de classificació dels éssers vius és l’espècie. De manera general, una espècie és un grup d’individus que tenen atributs comuns i que són designats pel mateix nom. A cada espècie se li dóna un nom científic, el qual és format per dues paraules llatines, d’acord amb una nomenclatura binària proposada pel naturalista suec Carl von Linné (1707 – 1778) i acceptada universalment. En aquesta nomenclatura, el primer nom s’escriu en majúscula i correspon al gènere, una unitat taxonòmica de nivell superior a l’espècie que inclou aquelles espècies emparentades que mostren un nombre suficient de característiques comunes.
Aquest nom genèric és qualificat per un adjectiu en minúscula, de vegades un substantiu en aposició. Per exemple, el nom científic de la nostra espècie és Homo sapiens, la qual cosa indica que pertanyem al gènere Homo, juntament amb altres espècies actualment extingides, com H. erectus o H. neanderthalensis, entre d’altres (de fet, les persones som els únics representants vius del gènere Homo). Els dos noms junts constitueixen el nom científic de l’espècie, i s’escriuen en un tipus de lletra diferent de la resta del text on es troben, generalment en cursiva o bé subratllats. Sovint, un cop s’ha anomenat en un text una espècie pel nom complet, la resta de cops que se’n fa esment s’escriu només la primera lletra del gènere seguida del qualificatiu d’espècie, per exemple, H. sapiens per Homo sapiens. Tanmateix, si l’espècie no és ben definida però hom està segur del gènere al qual pertany, s’acostuma a posar el nom del gènere seguit de les lletres sp., com per exemple, Drosera sp. per referir-se a una planta d’aquest gènere, propi dels aiguamolls dels Pirineus, sense especificar-ne l’espècie.
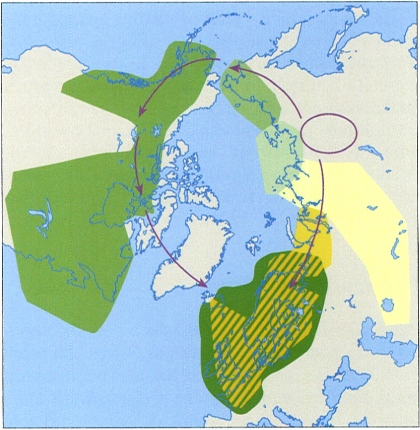
Corona circumpolar de distribució de diverses subespècies de Larus argentatus i L. fuscus. Les àrees marcades indiquen les distribucions de les espècies i subespècies del grup. S’indica la zona geogràfica d’origen d’aquestes espècies i subespècies, i la direcció de les rutes de migració que les va fer coincidir a la zona que comprèn les illes Britàniques i els països escandinaus.
IDEM, a partir de Mayr, 1999.
En algunes ocasions, a més, per a reflectir la complexa realitat natural, cal distingir entre diverses subespècies d’una mateixa espècie. Llavors s’afegeix un altre qualificatiu al nom de l’espècie. Per exemple, el gavià argentat de potes roses (Larus argentatus), del qual parlarem a continuació, presenta diverses subespècies, com el gavià argentat britànic (L. argentatus argenteus, en què Larus designa el gènere, argentatus el nom específic i argenteus la subespècie) i el gavià argentat europeu (L. argentatus argentatus), entre d’altres. Els noms vàlids són triats d’acord amb la regla de prioritat de publicació de la descripció, però tanmateix la nomenclatura és regulada per una normativa internacional l’objectiu de la qual és eliminar al màxim l’ambigüitat.
Tanmateix, el concepte biològic d’espècie no és senzill de delimitar teòricament, com posa de manifest el fet d’haver de reconèixer, en algunes ocasions, subespècies d’una mateixa espècie. El naturalista francès Georges Léopold Cuvier (1769 – 1863) el definí com el conjunt d’individus nascuts els uns dels altres o de pares comuns i que s’hi assemblen tant com entre ells mateixos. Aquest concepte i altres de semblants, bé que útils, no acaben de ser completament satisfactoris, atès que no concreten el grau de dissimilitud mínima que permet de diferenciar una espècie d’una altra de morfològicament propera. En aquest sentit, el criteri més objectiu per a distingir espècies pròximes és el genètic, elaborat pel biòleg evolucionista Ernst W. Mayr (1904 – 2005), el qual es basa en l’esterilitat entre espècies pròximes o en l’esterilitat dels híbrids resultants de llur encreuament. Així, per a considerar que dos individus pertanyen a una mateixa espècie, cal que en encreuar-se generin descendents que s’hi assemblin tant com entre ells mateixos, i que aquests descendents també siguin fèrtils. Dit d’una altra manera: si dues poblacions d’individus no generen mai descendents o bé els seus descendents no són fèrtils, hom els ha de considerar espècies diferents.
No obstant això, i malgrat que aquest criteri sigui objectiu, a la pràctica no sempre és fàcil d’aplicar, especialment en organismes sense reproducció sexual, com els bacteris i molts eucariotes unicel·lulars, o en què és difícil encreuar-los sota control. També hi ha altres situacions complexes, com, per exemple, les que es donen en alguns casos de distribució geogràfica contínua d’espècies molt emparentades. Per exemple, el gavià argentat de potes roses (Larus argentatus), un gavià atlàntic present ocasionalment a les costes dels Països Catalans, no es pot encreuar amb el gavià fosc (L. fuscus), un altre gavià atlàntic comú a tots els Països Catalans, a la tardor i a l’hivern. L’àrea de distribució d’aquestes dues espècies es sobreposa a la zona de les illes Britàniques i dels països escandinaus, a la mar del Nord i en altres ports d’Alemanya, França, Països Baixos, Dinamarca i Polònia, entre d’altres, però com que no es poden encreuar, cal considerar-les espècies diferents a tots els efectes.
La distribució geogràfica actual d’aquests gavians forma una corona circumpolar contínua que ressegueix les costes europees, americanes i asiàtiques. En aquesta corona de distribució es poden reconèixer diverses espècies i subespècies amb característiques morfològiques distintives i amb àrees d’ocupació que limiten les unes amb les altres.
Totes aquestes formes biològiques de gavià es poden encreuar amb les formes veïnes, tant amb la que es localitza a l’est de la seva àrea de distribució com amb la que està a l’oest. Pràcticament, l’única excepció són Larus argentatus i L. fuscus, que no es poden encreuar entre si, però sí que ho poden fer amb les formes adjacents cap a l’oest i cap a l’est respectivament, malgrat que la seva àrea de distribució se sobreposi àmpliament a les illes Britàniques i als països escandinaus, entre altres indrets del nord d’Europa. Tot això fa que aquestes dues formes biològiques hagin de ser classificades com a espècies diferents l’una de l’altra i explica la necessitat de definir subespècies transitòries, atès que aquestes sí que es poden encreuar.
L’explicació d’aquest fet cal buscar-la en la història evolutiva d’aquests gavians, probablement a partir d’una única població ancestral de la qual van anar divergint.

Morfologia i cariotip de dues espècies de planària d’aigua dolça, Polycelis felina (a l’esquerra) i P. nigra (a la dreta). En moltes espècies de planària es fa servir el cariotip com a criteri de classificació taxonòmica: 9 parells de cromosomes (n = 9, 2n = 18) en el cas de P. felina i 8 parells (n = 8, 2n = 16) en el cas de P. nigra. Totes dues espècies són 2n, és a dir, presenten dues dotacions de cromosomes homòlegs o iguals (n representa el nombre bàsic, o haploide, de cromosomes de l’espècie).
Maria Ribes / Biopix i R. Romero / M. Vila.
Tanmateix, malgrat les limitacions inherents a qualsevol definició, no hi ha dubte que les espècies són una categoria natural. Actualment, a part de la similitud morfològica, per a establir la distinció entre espècies també s’empren criteris de similitud genètica, com, per exemple, mitjançant la comparació de seqüències concretes de DNA o d’estructura cromosòmica, els quals permeten resoldre moltes de les ambigüitats i les incerteses que la taxonomia té plantejades. Els cromosomes són les estructures cel·lulars que contenen la molècula portadora de l’herència biològica, el DNA (més endavant parlarem àmpliament de les anàlisis genètiques i de les eines i molècules implicades). Per exemple, una de les característiques importants per a descriure i identificar algunes espècies de planària d’aigua dolça, uns platihelmints presents en molts rius dels Països Catalans, és el cariotip. El cariotip és la descripció ordenada i completa de la dotació cromosòmica d’un organisme, atenent al nombre, la mida i la forma dels cromosomes de les seves cèl·lules.
La taxonomia i els tàxons

Comparació dels principals tàxons a què pertanyen una espècie animal, l’espècie humana (Homo sapiens), i una de vegetal, l’alzina (Quercus ilex). L’esquema de classificació taxonòmica no inclou totes les categories de la jerarquia taxonòmica, sinó només les que són essencials per a la classificació.
A partir de fonts diverses.
La taxonomia és la branca de la biologia que classifica els éssers vius i n’estableix els grups, que s’anomenen tàxons. Un tàxon és cada entitat de classificació dels éssers vius reconeguda pels codis internacionals de nomenclatura botànica, zoològica i bacteriològica. Els tàxons s’ordenen segons una jerarquia estricta. Com hem dit anteriorment, el tàxon bàsic de classificació dels éssers vius és l’espècie. Les unitats taxonòmiques clàssiques de rang superior a l’espècie són, en ordre creixent de magnitud i segons les va establir Linné, el gènere, la família, l’ordre, la classe, el fílum (en animals) o la divisió (en plantes) i el regne. Així, el gènere agrupa una o diverses espècies filogenèticament relacionades, la família, un o diversos gèneres emparentats, etc. Tanmateix, la utilització d’eines i dades genètiques i moleculars ha revelat l’existència d’una nova unitat taxonòmica de grau superior al regne: el domini.
Tot sovint també s’utilitzen tàxons de rang inferior a l’espècie per a ajudar a distingir agrupacions dins una mateixa espècie, com la subespècie, la varietat i la forma. I també es fan servir categories addicionals entre les diverses unitats taxonòmiques tradicionals proposades per Linné, per exemple, amb l’ús dels prefixos super-, sub-, infra- davant del nom del tàxon, i algunes agrupacions a les quals no s’ha donat encara una categoria taxonòmica concreta.
La taxonomia clàssica
Per a classificar els organismes en els diferents tàxons es poden utilitzar diversos criteris. La taxonomia clàssica agrupa els organismes en funció de les afinitats existents amb relació als caràcters morfològics, la qual cosa obliga a definir si un determinat caràcter és primari o derivat, atesa la diferent utilització que se’n pot fer de cara a les classificacions taxonòmiques. Els caràcters primaris són aquells que ja eren presents en els ancessors comuns a les espècies que es volen agrupar taxonòmicament. Els caràcters derivats, en canvi, són el resultat d’adaptacions puntuals d’una espècie concreta o d’un grup restringit d’espècies.
Per exemple, a les illes Canàries hi ha diverses espècies d’aràcnids pertanyents al gènere Dysdera. Una d’aquestes espècies, D. gollumi, identificada a la cova de Los Roques, a Las Cañadas del Teide, és troglòbia (habita en les parts profundes de les coves) i es caracteritza per una absència total d’ulls. En canvi, D. levipes, identificada a tot el vessant nord de l’illa de Tenerife i a la Gomera, és epigea (viu sobre la superfície del sòl) i presenta desenvolupament ocular, com la resta d’aranyes epigees del seu gènere. En aquest cas, el caràcter morfològic d’absència d’ulls s’ha de considerar derivat, atès que tots els representants d’aquest gènere i d’altres gèneres emparentats amb els quals comparteixen ancestres comuns i que viuen a la superfície del sòl presenten ulls, la qual cosa implica que els ancessors comuns també en presentaven.
Un cas diferent és el de les aranyes del gènere Nesticus que habiten la serralada pirinenca i el conjunt dels Països Catalans. N. obcecatus, per exemple, identificada en algunes cavitats dels Pirineus d’Osca, també és troglòbia i presenta una reducció molt significativa quant al nombre d’ulls respecte d’altres aranyes del mateix gènere. N. cellulanus, en canvi, present a bona part dels Països Catalans, és epigea i té un nombre d’ulls molt superior al de N. obcecatus. En aquest cas, però, els ancessors de N. obcecatus que van colonitzar els Pirineus d’Osca ja presentaven una reducció del nombre d’ulls, per la qual cosa aquest caràcter ha de ser considerat primari.
A continuació la taula presenta la classificació taxonòmica completa de l’espècie humana. S’hi indiquen tots els tàxons establerts, en ordre creixent de rang taxonòmic, alguns que encara no tenen una categoria taxonòmica definida i d’altres que estan en discussió. Els que coincideixen amb els set proposats inicialment per Linné apareixen en negreta. Algunes agrupacions taxonòmiques s’han establert partint de criteris de similitud molecular (entre seqüències de DNA). Quan es basen en comparacions amb organismes fòssils, s’indica l’era geològica en què van viure (A partir de fonts diverses).
| CATEGORIA TAXONÒMICA | NOM | CARACTERÍSTIQUES |
|---|---|---|
| Espècie | Homo sapiens | |
| Gènere | Homo | Desenvolupament específic i especialitzat de la memòria i l’aprenentatge. Els humans en són els únics representants vius. |
| Subtribu | Hominina | Bipedisme. Els humans en són els únics representants vius. |
| Tribu | Hominínids (Hominini) | Inclou també els ximpanzés i els bonobos, però no els goril·les. |
| Subfamília | Homininis (Homininae) | En caminar utilitzen els artells de la mà. Inclou també els goril·les, però no els orangutans. |
| Família | Homínids (Hominidae) | Grans primats. Inclou també els orangutans. |
| Superfamília | Hominoïdeus (Hominoidea) | Primats sense cua i amb la locomoció arbòria restringida als braços. Inclou també els gibons. |
| Parvordre | Catarins (Catarrhini) | Locomoció terrestre. Inclou també la resta de primats del Vell Món. |
| Infraordre | Simiformes (Simiiformes) | Utilització d’elements naturals no modificats com a eines. Inclou també els primats del Nou Món. |
| Subordre | Haplorins (Haplorrhini) | No tenen membrana als orificis nasals externs ni vibrisses al morro. Inclou també els prosimis del Nou Món. |
| Ordre | Primats (Primates) | Locomoció arbòria prènsil. |
| Superordre | Euarconts (Euarchonta) | Basat en la similitud de seqüències de DNA. Inclou també els lèmurs. |
| Magnordre | Euarcontoglirs (Euarchontoglires) | Basat en la similitud de seqüències de DNA. Inclou també els rosegadors. |
| No definida | Boreoeuteris (Boreoeutheria) | Testicles externs. Inclou els ratpenats, els eriçons, els ungulats i els carnívors. Es defineix també d’acord amb la similitud de seqüències de DNA. |
| Cohort | Euteris (Eutheria) | Formació de placenta durant el desenvolupament embrionari. Inclou tots els mamífers placentarís i mamífers extingits més semblants als mamífers placentaris actuals que als marsupials. |
| Supercohort | Teris (Theria) | Mamífers que pareixen les cries sense que estiguin envoltades d’una closca d’ou. Inclou també els marsupials. |
| Infralegió | Tribosfènids (Tribosphenída) | Inclou també alguns mamífers extingits dels grups taxonòmics Hypomylos, Necrolestidae i Aegialodontia, que van viure a l’inici del Cretaci. |
| Sublegió | Zateris (Zatheria) | Inclou també alguns mamífers extingits dels grups taxonòmics Arguitheriidae, Arguimuridae, Vincelestidae i Peramuridae, que van viure al final del Juràssic i l’inici del Cretaci. |
| Legió | Cladoteris (Cladotheria) | Inclou també alguns mamífers extingits que van viure al final del Juràssic i a l’inici del Cretaci del grup taxonòmic Dryolestoídea. |
| Superlegió | Trecnoteris (Trechnotheria) | Inclou també alguns mamífers extingits dels grups taxonòmics simetrodonts i espalacotèrids, que van viure al Mesozoic. |
| Infraclasse | Holoteris (Holotheria) | Inclou també alguns mamífers extingits del grup taxonòmic Chronoperotidae, que van viure al Mesozoic. |
| Subclasse | Teriformes (Theriiformes) | Mamífers que pareixen les cries sense que estiguin envoltades d’una closca d’ou. Inclou tots els mamífers placentaris i els marsupials, vius i fòssils. |
| Classe | Mamífers (Mammalia) | Animals les femelles dels quals posseeixen glàndules mamàries per a alimentar les cries. Inclou també els monotremes, com els ornitorincs, que pareixen les cries envoltades d’una closca. |
| No definida | Mamaliaformes (Mammaliaformes) | Inclou també protomamífers extingits del Triàsic tardà, com els del gènere Sinoconodon. |
| No definida | Probainògnats (Probainognathia) | Inclou també alguns rèptils mamiferians extingits del Triàsic i el Juràssic, com els del gènere Lumkuia. |
| No definida | Cinodonts (Cynodontia) | Inclou també rèptils extingits semblants als mamífers de l’inici del Triàsic, com els del gènere Cynosaurus. |
| No definida | Teriodonts (Theriodontia) | Inclou també alguns rèptils mamiferians extingits del Permià i el Cretaci mitjà, com el gènere Cynognathus. |
| No definida | Teràpsids (Therapsida) | Presenten mandíbules complexes, amb incisives frontals i canines laterals. Inclou també alguns rèptils mamiferians extingits de l’inici del Cretaci. |
| No definida | Sinàpsids (Synapsida) | Inclou protomamífers extingits (o rèptils mamiferians) del Carbonífer tardà i també els rèptils i els ocells. |
| (Sèrie) | Amniotes (Amniota) | Vertebrats tetràpodes els ous dels quals estan adaptats a terra ferma, bé perquè estan envoltats d’una closca (com les aus i els rèptils) o de membranes amniòtiques (com els mamífers). Durant el desenvolupament no presenten estadis larvaris. |
| Superclasse | Tetràpodes (Tetrapoda) | Vertebrats amb quatre extremitats. Inclou també els amfibis. |
| No definida | Sarcopterigis (Sarcopterygii) | Inclou també els peixos amb aletes carnoses, com els celacants i els peixos pulmonats. |
| Infrafílum | Gnatostomats (Gnathostomata) | Vertebrats amb mandíbules. Inclou també els peixos ossis. |
| Subfílum | Vertebrats (Vertebrata) | Cordats craniats amb espina dorsal o columna vertebral. Inclou tota la resta de peixos. |
| No definida | Craniats (Craniata) | Cordats amb crani. Inclou també els anomenats "peixos bruixa". |
| Fílum | Cordats (Chordata) | Animals que presenten, com a mínim en algun període del cicle vital, notocordi o cordó nerviós dorsal, fenedures branquials, endostil i cua postanal. Inclou també els urocordats i els cefalocordats, com l’amfiox (Amphioxus). |
| Superfílum | Deuterostomats (Deuterostomata) | Animals en què la primera obertura embrionària, el blastòpor, esdevé l’anus en l’adult. Inclou també els equinoderms, els hemicordats i els xenoturbèl·lids. |
| No definida | Celomats (Coelomata) | Animals triploblàstics (amb tres capes embrionàries) que presenten celoma entre les capes ectodèrmica i endodèrmica. Inclou també els anèl·lids, els mol·luscs, els artròpodes i els lofòfors. |
| Subregne | Bilaterals (Bilateria) | Animals amb simetria bilateral. |
| No definida | Eumetazous (Eumetazoa) | Animals els embrions dels quals passen per un estadi de gàstrula. Inclou tots els animals excepte les esponges i els placozous. |
| Regne | Animals o metazous (Animalia) | Organismes eucariotes pluricel·lulars de nutrició heteròtrofa les cèl·lules dels quals no presenten paret cel·lular. A més, tots passen com a mínim per una fase mòbil durant el cicle vital. |
| Superregne | Opistoconts (Opisthokonta) | Organismes eucariotes pluricel·lulars de nutrició heteròtrofa. Inclou també els fongs. |
| Subdomini | Uniconts (Unikonta) | Inclou també les amebes. |
| Domini | Eucaris (Eukarya) | Organismes eucariotes (formats per cèl·lules amb nucli cel·lular). Inclou també les plantes, les algues i tota la resta de protists. |
| No definida | Ciots (Cyota) | Tots els éssers vius amb estructura cel·lular. Inclou també els arqueus i els bacteris (cèl·lules procariotes, sense nucli cel·lular). |
| No definida | Biots (Biota) | Grup teòric proposat per a incloure tota la vida, tant terrestre com extraterrestre, en cas d’identificar-se. |
La taxonomia numèrica
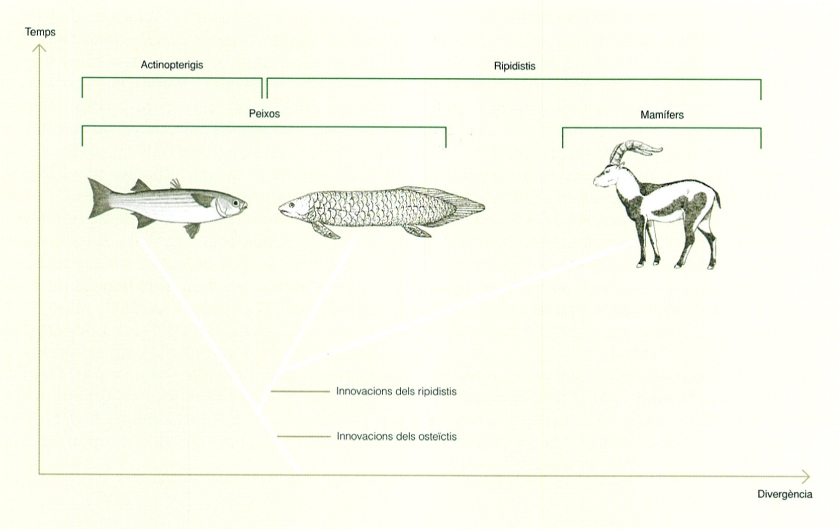
El criteri de l’anomenada fenètica, que incorpora trets adaptatius i ecològics al costat dels morfològics, pot portar a agrupacions errònies, com la que l’arbre dels actinopterigis i els ripidistis mostra entre els actinopterigis i els dipnous (peixos pulmonats). L’esquema indica també la incorporació de les característiques pròpies dels ripidistis, que inclouen les principals innovacions dels tetràpodes, i dels osteïctis, que engloben els actinopterigis i els ripidistis i que inclouen la calcificació de l’esquelet intern.
Dades de l’autor / Agnès Perelló i Gustavo Hormiga.
Una altra aproximació a la classificació taxonòmica és l’anomenada taxonomia numèrica o fenètica. La taxonomia numèrica es basa en la descripció de tantes característiques com sigui possible dels grups que s’han de comparar, preferiblement més d’un centenar, a les quals s’assignen valors numèrics. Si la característica és qualitativa, és a dir, excloent, com per exemple, presència o absència d’ulls, hi haurà dos valors numèrics possibles. En canvi, si és quantitativa, com per exemple, nombre d’ulls dins un interval determinat, llavors a cada estat se li assigna un valor. Un cop atorgats els valors, es tracten matemàticament per establir el grau de similitud fenètica entre els grups d’organismes que es comparen.
La fenètica s’utilitza molt per a classificar microorganismes, en els quals no es reconeixen gaires caràcters morfològics distintius però sí fisiològics, que són quantificats d’aquesta manera. Per exemple, es pot establir experimentalment si un tipus de bacteri determinat és capaç de fermentar o no un hidrat de carboni concret, o si pot créixer en presència d’altes concentracions de clorur de sodi. Les unitats taxonòmiques que estableix la taxonomia numèrica s’anomenen UTO (en anglès OTU, acrònim d’Operational Taxonomic Units, ‘unitats taxonòmiques operatives’) i entre elles es poden establir coeficients de similitud i de distància taxonòmica. També s’utilitza en la classificació dels organismes superiors, generalment per a establir categories per sota del rang taxonòmic de gènere.
La taxonomia molecular
Finalment, la taxonomia molecular estableix el grau de relació entre espècies o tàxons de rang superior mitjançant l’anàlisi i la comparació de seqüències de DNA i/o de determinades proteïnes, atenent al principi que les semblances moleculars conjecturen l’existència d’ancessors comuns, i que el grau de similitud és un reflex de la distància filogenètica (o evolutiva) i del temps que fa que llurs llinatges van divergir. A l’apartat 4 d’aquesta secció parlarem a bastament de filogènia i taxonomia molecular.
La sistemàtica i la filogènia
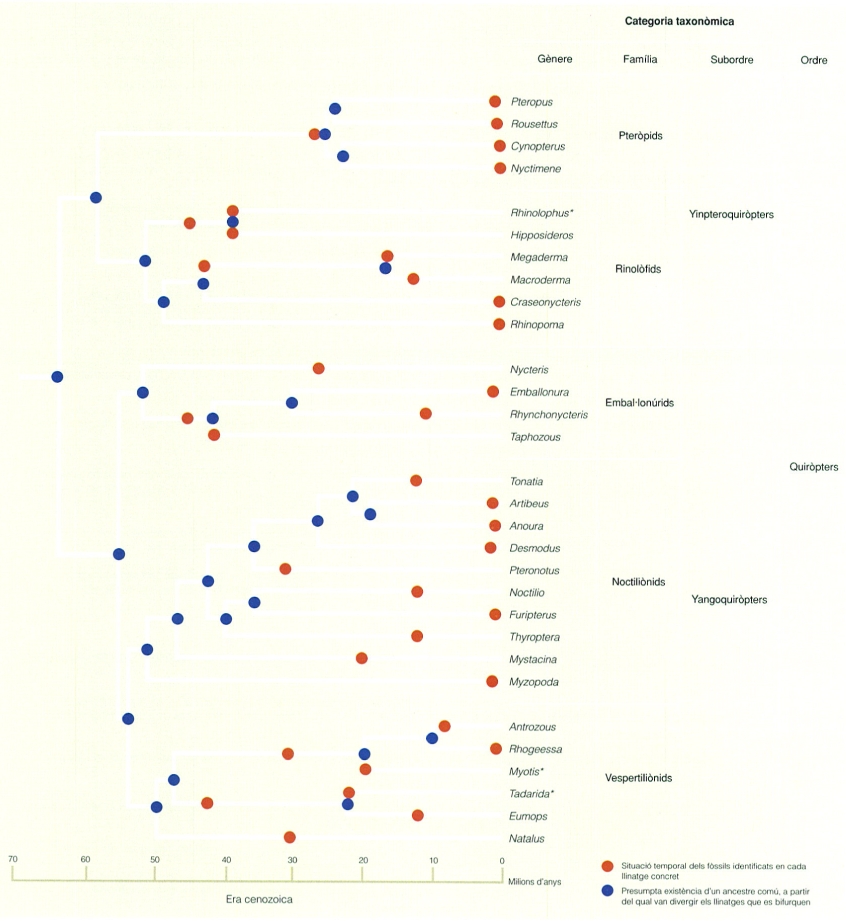
Exemple d’arbre filogenètic, en aquest cas concret, dels quiròpters, que mostra quatre categories taxonòmiques diferents: gènere, família, subordre i ordre (per simplificar-ne la interpretació, no s’indiquen les espècies, ni tampoc conté tots els gèneres). L’ordre dels quiròpters inclou unes 1.100 espècies identificades de ratpenats, les quals representen aproximadament el 20% del total d’espècies de mamífers. Amb (*) s’indiquen els gèneres presents als Països Catalans.
A partir de Teeling i altres, 2005.
En aquest context, la sistemàtica és la branca de la biologia que estudia la relació de parentiu i les relacions evolutives entre grups d’organismes. La filogènesi, en canvi, és el conjunt d’aquestes relacions de parentiu d’acord amb la història evolutiva. Així, la filogènia és la branca de la biologia que estudia el procés de la filogènesi. El terme filogènesi fou introduït pel biòleg alemany Ernest Haeckel (1834 - 1919) en la seva obra Generelle Morphologie der Organismen (‘Morfologia general dels organismes’, 1866), per analogia amb el terme ontogènesi. L’ontogènesi és la descripció del procés de desenvolupament d’un organisme des de la fecundació fins que ateny l’estat madur. Així, la filogènesi és la descripció del procés evolutiu d’un grup d’organismes, des del seu origen fins a l’actualitat.
L’estudi de la filogènesi dels organismes ha fet canviar l’enfocament de la sistemàtica, la qual ja no es basa solament en la morfologia i la fisiologia dels organismes sinó també en el seu parentiu evolutiu. És a dir, que ha incorporat una visió dinàmica (la de l’evolució) a les classificacions, la qual s’afegeix a la visió estàtica que tradicionalment li han conferit els estudis morfològics. Per aquest motiu, la zoologia, la botànica i la microbiologia, vistes des de la filogènia, poden ser definides com l’estudi comparatiu de la diversitat d’animals, de plantes i de microorganismes respectivament.
Els estudis filogenètics han experimentat un gran auge amb l’adveniment de les tècniques i les eines d’anàlisi molecular. La filogènia molecular és la branca de la biologia que estudia les relacions evolutives entre grups d’organismes en funció del parentiu avantpassat-descendent sobre la base de dades o caràcters moleculars, com per exemple, comparant seqüències d’àcids nucleics (com DNA o RNA) o proteïnes. Els estudis filogenètics són indispensables per a conèixer el grau de parentiu entre els diversos grups d’organismes i el temps que fa que es va produir la divergència evolutiva.
En aquest context, un arbre filogenètic és un diagrama que representa les relacions de parentiu evolutiu entre diferents grups d’organismes, els quals es van ramificant a partir d’un tronc comú en branques que condueixen cap a les espècies, agrupades progressivament en tàxons de nivell superior. Així, cada enforcadura o punt de ramificació pressuposa l’existència d’un avantpassat comú als llinatges que es bifurquen. I l’arbre de la vida és un arbre filogenètic que pretén recollir totes les espècies d’éssers vius.
La importància social dels arbres filogenètics
Hi pot haver algú que pensi que la construcció d’arbres filogenètics només resulta útil com a ciència bàsica, per a entendre les relacions evolutives i comprendre les relacions de parentiu entre els éssers vius actuals, però el cert és que les dades que aporten proporcionen beneficis tangibles a la societat, en aspectes diversos. N’esmentarem uns quants, a tall d’exemple.
Un dels camps en què la construcció d’arbres filogenètics cada cop més acurats reporta enormes beneficis és el de la medicina. Per exemple, l’anàlisi de les relacions filogenètiques entre diferents virus que poden afectar els humans permet establirne l’origen evolutiu, geogràfic i epidemiològic, la qual cosa contribueix a trobar la millor forma d’eradicar-los. Un cas concret és el del virus SNV (acrònim de sin nombre virus), que entre els mesos d’abril i juny del 1993 va provocar la mort de deu persones al sud-oest dels Estats Units a conseqüència d’una malaltia llavors desconeguda, amb una taxa de mortalitat del 70%. L’agent infecciós va resultar ser un virus de la família dels hantavirus, i es va constatar que provenia d’un parell d’espècies de ratolí de camp (Peromyscus maniculatus i P. leucopus) típiques de la zona i llunyanament emparentades amb els ratolins de camp europeus.

Superposició dels arbres filogenètics d’alguns hantavirus (en gris) i d’algunes espècies de rosegador (en blanc) del Nou Món, per a mostrar l’evolució paral·lela d’ambdós grups. A baix, imatges de rata comuna (Rattus norvegicus), de ratolí de camp nord-americà (Peromyscus maniculatus) i de l’hantavirus que causa una síndrome pulmonar en els rosegadors.
A partir de Yates i altres, 2002 / MDG CN, National Park Service / John Good i Centers for Disease Control and Prevention (EUA).
L’anàlisi filogenètica d’aquest virus, comparada amb la d’altres hantavirus, va revelar que havia evolucionat paral·lelament a diferents espècies de rosegadors, un fenomen biològic que es coneix com a coevolució, la qual cosa va permetre als investigadors predir que moltes espècies de rosegadors a les quals encara no se’ls havien trobat virus d’aquesta família molt probablement també en serien portadores, amb les consegüents conseqüències per a la salut humana. Això va fer possible cercar-los de manera específica i dirigida, identificar-ne uns quants i explicar l’etiologia d’algunes malalties pulmonars de les quals fins llavors es desconeixia l’agent infecciós. I comprendre la causa d’una malaltia és un pas previ necessari per a diagnosticar-la de forma inequívoca i cercar-hi remei mèdic i farmacèutic.
La biologia de la conservació és sens dubte l’àrea que rep una influència més directa del coneixement cada cop més acurat que es té de l’arbre de la vida. El motiu és ben simple: aquest increment de coneixement porta implícit un augment del nombre d’espècies identificades. I no es pot conservar allò que no es coneix. Actualment, un dels temes més importants que intenta resoldre la biologia de la conservació és quantes espècies d’éssers vius hi ha. S’estima que són entre 12,5 i 13 milions d’espècies, però un percentatge significatiu encara no ha estat científicament identificat. I tan sols el 0,4% ha estat incorporat a l’arbre general de la vida, que ha d’acabar incloent-les totes.

Arbre filogenètic dels principals grups de plantes terrestres, el qual ha permès deduir que els gens que confereixen tolerància a la dessecació ja eren presents en les plantes primitives, es van perdre abans de l’embrancament del llinatge dels traqueòfits i van reaparèixer posteriorment, almenys, en tres llinatges de manera independent. A més, aquests gens van tornar a evolucionar de forma també independent, almenys, en vuit grups d’anglospermes (no es mostra en aquest arbre) En blau, s’indiquen els grups que contenen espècies tolerants a la dessecació.
Adaptat a partir d’Oliver i altres, 2000.
A més, l’arbre de la vida, o els arbres filogenètics parcials en el seu defecte, contribueixen a aclarir les dues qüestions primordials que es planteja la biologia de la conservació: com es distribueix la diversitat en el paisatge i com s’ha assolit aquesta diversitat. Per exemple, fa dècades que s’invoca l’aparent estabilitat dels boscos tropicals com a paradigma per a explicar la gran diversitat d’espècies que hi ha en aquestes àrees. Tanmateix, diversos estudis filogenètics i filogeogràfics han demostrat que la biodiversitat es forma progressivament i és més acusada en zones heterogènies i inestables que no pas en àrees amb aparentment poques pertorbacions. En definitiva, la informació que proporcionen els arbres filogenètics és extremament útil per a fer una bona planificació de la conservació i per a establir les àrees més efectives en les quals reintroduir poblacions d’espècies concretes.
El valor potencial dels arbres filogenètics en agricultura també és enorme. D’una banda, l’existència d’una filogènia acurada ajuda els investigadors a buscar gens concrets en els ancessors silvestres de les plantes de conreu actuals, la qual cosa permet conèixer com ha anat canviant llur funció durant el procés de domesticació i contribueix a la modificació selectiva de les noves generacions de plantes de manera molt més precisa, en funció de les necessitats. Per exemple, la utilització de l’arbre filogenètic de les plantes ha permès identificar i traçar els gens responsables de la tolerància a la sequera, des de plantes morfològicament senzilles com les hepàtiques i els marcantiòfits fins a les modernes angiospermes. Atesa la taxa actual de desertització en alguns indrets del planeta, dades com aquesta poden resultar de gran importància a l’hora de desenvolupar una agricultura global més sostenible.

Relació filogenètica entre el blat de moro conreat i el teosinte silvestre actual. Aquesta relació ha ajudat a explicar les variacions morfològiques entre aquestes plantes.
Adaptat a partir de Wang i altres, 2001.
Un altre exemple fa referència al blat de moro, la planta conreada que té més importància econòmica. El blat de moro és una planta herbàcia anual amb un únic cos fructífer, la panotxa, una condició que normalment no es troba en les plantes herbàcies silvestres. Fent servir anàlisis filogenètiques s’ha demostrat que les espècies silvestres actuals més emparentades amb el blat de moro conreat són les de teosinte, pròpies de Mèxic i Guatemala, precisament els llocs on es va domesticar i es va començar a seleccionar el blat de moro fa 9.000 anys. Així, doncs, també s’ha vist que el blat de moro conreat es pot encreuar amb algunes variants de teosinte, la qual cosa permet incorporar característiques d’aquesta planta que es considerin útils per a les varietats conreades.
Les espècies invasores s’han convertit en un gran problema a tot el món que provoca danys irreparables en moltes espècies natives de molts ecosistemes. Les anàlisis filogenètiques són una eina important per a identificar les espècies invasores i determinar-ne l’origen geogràfic. A més, permeten abordar la qüestió de les associacions evolutives d’aquestes espècies amb altres que en siguin preses o bé depredadores, la qual cosa fa possible predir l’abast de la invasió. Per exemple, s’ha fet una anàlisi filogenètica d’espècies invasores de coleòpters en relació amb diversos hàbitats, el qual ha mostrat quines espècies són susceptibles de provocar plagues i ens quins hàbitats. Aquest estudi teòric ha estat demostrat pel cas de Tetropium fuscum, una espècie invasora d’escarabat que a Europa no provoca plagues però en canvi sí que ho fa a l’Amèrica del Nord. En aquest sentit, s’ha proposat que aquestes anàlisis d’associacions poden servir també per a la lluita biològica contra aquestes plagues, per exemple, afavorint espècies autòctones que desplacin les espècies invasores.

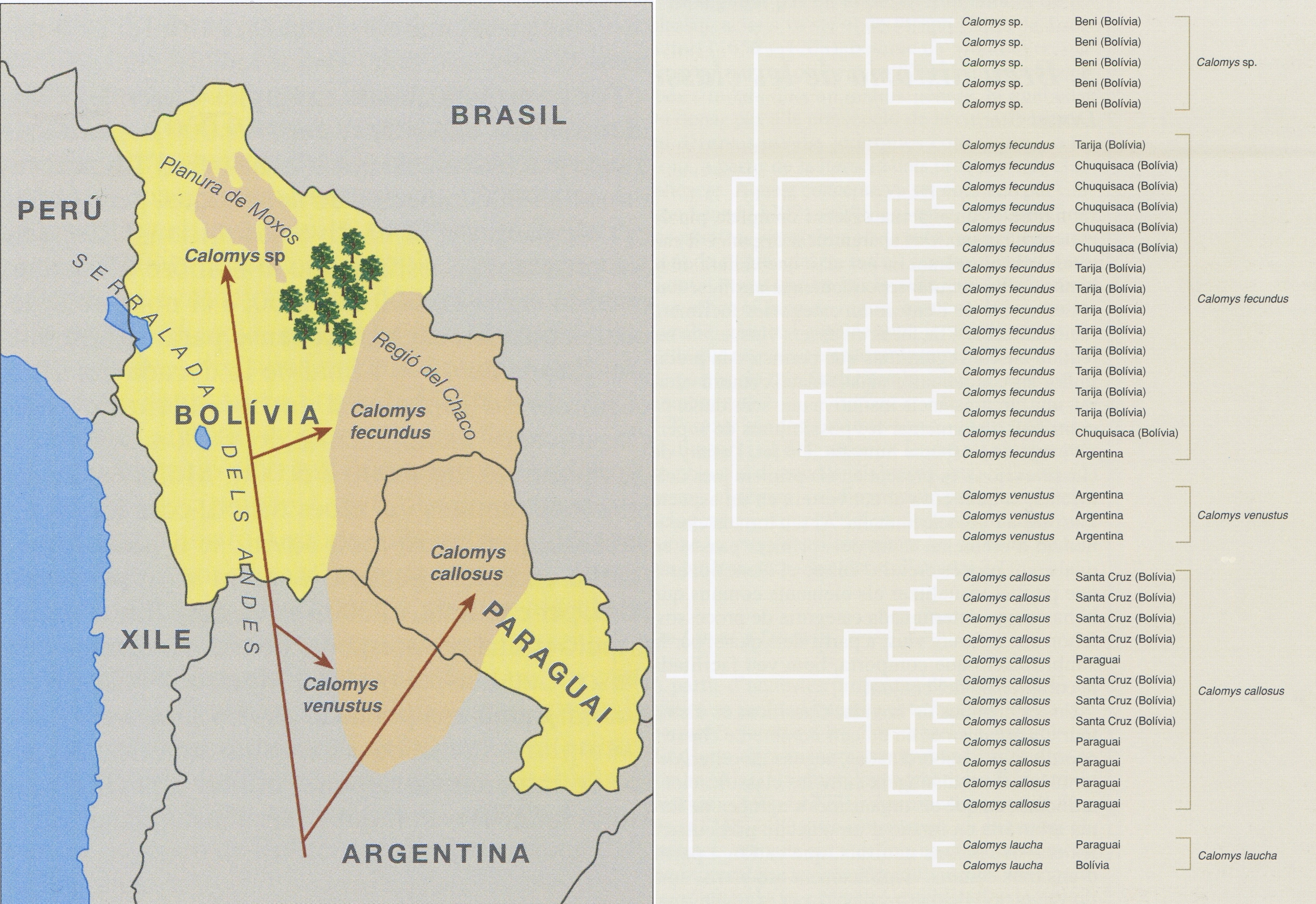
Àrea de distribució de diverses espècies de ratolí del gènere Calomys a la regió del Gran Chaco: una espècie no identificada (Calomys sp.) és portadora del virus Machupo (MACV), causant de la febre hemorràgica boliviana, i C. fecundus, C. venustus i C. callosus ho són del virus Latino. Les fletxes indiquen la colonització de la regió segons les dades de l’arbre filogenètic (a baix). L’arbre també inclou una espècie no esmentada en el mapa: C. laucha, com a grup extern que permet situar l’arrel de l’arbre. S’indica el departament de Bolívia o el país d’on ha estat obtinguda cada població.
IDEM, a partir de Salazar-Bravo i altres, 2002.
Els arbres filogenètics també tenen implicacions que contribueixen a una bona gestió del sòl, especialment si s’integren en altres disciplines. Per exemple, s’han emprat anàlisis filogenètiques per a determinar l’existència i l’origen geogràfic de diverses espècies molt emparentades de ratolins a l’est de Bolívia, pertanyents al gènere Calomys, cadascuna de les quals és portadora d’un tipus específic d’arenavirus. Els individus que habiten la planura de Moxos (departament de Beni, Bolívia) pertanyen a una espècie no prou ben definida d’aquest gènere (Calomys sp.) i poden transmetre el virus Machupo (MACV), que causa la febre hemorràgica boliviana. En canvi, les espècies que hi ha a la regió del Chaco (departament de Santa Cruz, Bolívia), concretament C. fecundus, C. venustus i C. callosus, poden transmetre el virus Latino, que no és patogènic. Tanmateix, si els individus de la planura de Moxos es troben amb individus de les espècies de la regió del Chaco poden generar descendència híbrida, la qual podria escampar el virus patogènic en districtes actualment lliures d’aquesta malaltia. La frontera natural entre aquestes dues poblacions és el sòl que les separa, cobert de bosc, però una gestió inadequada permetria obrir corredors a través dels quals aquestes espècies podrien franquejar-la.
La biodiversitat i l’arbre de la vida
Per a establir una filogènia, el primer punt és haver identificat totes les espècies que hom vol incloure-hi. I per a abordar la construcció de l’arbre de la vida, cal haver-les identificat i classificat totes. Tanmateix, actualment no coneixem ni tan sols l’ordre de magnitud del nombre d’espècies d’éssers vius, atès que moltes no han estat mai descrites. Els càlculs oscil·len àmpliament en funció del mètode utilitzat, des de 3,6 milions fins a més de 100 milions d’espècies, malgrat que sovint s’acota a un interval d’entre 12,5 i 13 milions d’espècies. Unes dades que contrasten amb el nombre d’espècies descrites, que al començament de l’any 2010 oscil·laven, segons els còmputs, entre 1,5 i 1,8 milions.
Per posar uns quants exemples concrets, s’han identificat i classificat unes 69.000 espècies de fongs, però es calcula que n’hi ha més d’ 1,6 milions, i unes 15.000 espècies de nematodes, però també es calcula que n’hi ha diversos milions. Respecte als vertebrats, s’estima que el nombre d’espècies oscil·la entre 15.000 i 40.000, i el nombre d’espècies identificades creix sense parar. Entre mitjan dècada de 1990 i final de la dècada del 2000 el nombre d’espècies d’amfibis identificades i classificades ha augmentat de 4.000 a 5.300, i el de mamífers, de 4.000 a 5.000. Molts es troben en zones remotes i d’accés difícil, com les selves amazòniques, però molt sovint se n’identifiquen també en zones geogràficament molt properes. Així, per exemple, el 2005 es va fer pública la descripció d’una nova espècie de tritó endèmica dels Països Catalans, el tritó del Montseny (Calotriton arnoldi), amb una població que no supera els 1.500 exemplars, i el 2009 es va citar una nova espècie de ratpenat, el ratpenat orellut alpí (Plecotus macrobullaris), en l’àmbit dels Països Catalans, en una cova situada al terme municipal de Queralbs (Ripollès).
Respecte a les plantes amb flors, són el grup d’éssers vius que ha estat més estudiat a nivell sistemàtic, atesa la major facilitat d’observació i classificació que presenta. S’han identificat i classificat unes 272.000 espècies diferents, però es calcula que el nombre total superarà les 300.000. Cada any s’identifiquen unes 2.000 espècies noves, que són afegides a l’Index Kewensis del Jardí Botànic Reial de Kew, al Regne Unit, que recull el nom i la classificació de totes les espècies de plantes i de fongs identificades.
Totes aquestes dades indiquen que només s’ha començat a explorar la vida a la Terra. I malgrat que el màxim desconeixement ateny el grup taxonòmic dels bacteris i els arqueus, el cert és que en major o menor grau afecta tots els grups taxonòmics. L’arbre de la vida és a l’inici del camí, però encara ha de fer un llarg recorregut. Tanmateix, els arbres filogenètics, com hem dit, aporten informació molt valuosa sobre els éssers vius presents i passats, i sobre el mateix fenomen vital, des de la perspectiva dinàmica que confereix l’evolució.
Breu història de les classificacions dels éssers vius
La natura és un sistema divers i complex, ple de materials i éssers vius aparentment dispars i de naturalesa heterogènia. És per això que al llarg de la història s’ha intentat repetidament classificar tots els elements de l’entorn en categories fàcilment racionalitzables, inclosos els éssers vius, en un intent de buscar regularitats que permetessin predir millor les condicions d’aquest mateix entorn natural, una circumstància instintiva que sens dubte ha contribuït a augmentar les possibilitats de supervivència de l’espècie humana. De fet, l’afany de classificació ja és present en els primers jocs dels infants, la qual cosa el converteix en un aspecte intrínsec a l’espècie humana. Al seu torn, la classificació d’elements és un dels primers passos en qualsevol procés creatiu, àdhuc el científic, atès que permet visualitzar els elements comuns que hi ha en una determinada categoria de processos, fenòmens o éssers vius, a partir dels quals, per la simplificació que comporta, hom veu facilitada la comprensió del conjunt.
En les ciències de la vida, classificar és distribuir els organismes en un cert nombre de classes, coordinades o subordinades, segons un criteri determinat. Hom distingeix entre les classificacions artificials, en què els organismes s’agrupen de forma arbitrària en funció d’un objectiu particular, i les classificacions naturals, que es fan a partir d’uns trets comuns científicament establerts, amb l’observació i l’experimentació, per exemple, analitzant la morfologia específica d’un grup de plantes, com el tipus d’arrel i de tija, la forma del limbe de les fulles, les especificitats dels òrgans florals, etc.
El mot taxonomia, que com s’ha dit designa la ciència de la classificació dels éssers vius, va ser proposat el 1813 pel botànic suís Augustin-Pyrame de Candolle (1778 – 1841). Tanmateix, l’interès a classificar els éssers vius no va néixer al segle XIX amb el mot utilitzat per a designar aquesta disciplina científica, sinó que ve de molt més antic, i va ser desenvolupat inicialment, de manera especial, en el context de la botànica. El motiu pel qual els botànics han estat els primers interessats a disposar d’una classificació dels subjectes del seu estudi, les plantes, és molt simple: si bé és relativament senzill per a qualsevol persona reconèixer i recordar el nom dels animals més importants o característics de la seva contrada, les plantes, i molt especialment les plantes medicinals, les primeres plantes no conreades que sens dubte han estat d’interès científic, exigeixen, per a reconèixer-les i per a no confondre-les amb altres espècies semblants, un grau d’aprenentatge més elevat, que tradicionalment ha estat reservat a determinats professionals, els quals han hagut de disposar d’elements precisos d’identificació (i, per tant, de classificació). Com tants altres aspectes de la ciència i el pensament occidental, la taxonomia va néixer a la Grècia clàssica.
D’Aristòtil al Renaixement
Una de les primeres persones de qui hi ha constancia escrita del seu afany per a classificar la natura fou Aristòtil (385 – 322 aC), el qual va establir una primera classificació dels animals en Anaima (‘sense sang’) i Enaima (‘amb sang’), atenent a aquesta característica biològica. Així, els Anaima incloïen, segons Aristòtil, els animals de cos tou, amb closca o sense o teguments durs, com per exemple, els insectes i els cucs, mentre que els Enaima incloïen bàsicament els vertebrats, com per exemple, els peixos, les serps, els ocells i els quadrúpedes vivípars i ovípars (mamífers i rèptils respectivament).
Teofrast (372 – 288 aC), successor d’Aristòtil al capdavant del Liceu (l’escola peripatètica), va escriure Perí phytón historía (‘Història de les plantes’), possiblement l’obra fundacional de la botànica com a disciplina científica. Posteriorment, Dioscòrides (s. I dC) va escriure un nou tractat de botànica en sis volums, en què va classificar les plantes en aromàtiques, alimentàries, medicinals, vinícoles i metzinoses (va dedicar un volum a cadascun d’aquests tipus de plantes llevat de les medicinals, a les quals en va destinar dos). Finalment, en el món llatí hom pot destacar la Historia Naturalis (‘Història natural’) de Plini el Vell (24 – 79 dC), una obra en 37 volums en què proposà una classificació basada també en la utilitat de les plantes, i la classificació dels animals feta per sant Agustí (354 – 430), que va agrupar segons fossin útils, damnosos o superflus.
A partir d’aquest moment, en els més de mil anys compresos entre la realització d’aquests treballs i el segle XVI, la major part de tractats es van dedicar a discutir sobre els llibres d’aquests autors, als quals es van afegir figures i nous textos per ajudar al reconeixement de les plantes, i a parlar dels diversos noms que en el decurs de la història s’havien donat a una mateixa planta. Tanmateix, al segle XVI es va començar a veure la necessitat de disposar de regles i principis de classificació més rigorosos. Però la dificultat de proposar una classificació nova és molt superior a la que comporta discutir els errors de classificacions ja proposades. A partir d’aquell segle diversos botànics i naturalistes, entre els quals figuren l’alemany Leonhard Fuchs (1501 – 1566) el 1531, el suís Conrad Gesner (1516 – 1565) el 1541, i el també alemany Rudolf Jacob Camerarius (1665 – 1721), el 1686, van proposar un nou sistema de classificació basat en criteris purament alfabètics, un intent a priori ben poc útil si hom no coneixia el nom de la planta, però que tanmateix té el mèrit d’haver trencat amb la creença generalitzada que els treballs dels clàssics eren, per definició, intocables.
Del Renaixement a Linné
Un cop superada aquesta etapa, entre els segles XVI i XVII es van anar succeint tot un seguit de propostes de mètodes de classificació basats, en el cas majoritari de les plantes, en les característiques morfològiques de les seves parts, com la tija, les fulles, les flors i els fruits. En general es van emprar dues grans estratègies lògiques. D’una banda, l’heretada d’Aristòtil, que consisteix a començar la classificació pels grups més grans d’organismes i anar-los dividint subsegüentment en grups més petits, tot aplicant criteris predefinits, fins a arribar a les espècies. Aquest procediment lògic, conegut com a divisori, va evolucionar cap a les claus dicotòmiques de classificació, en què en pràcticament totes les successives divisions un dels termes es defineix de manera negativa respecte de l’altre (per exemple, plantes amb flors / plantes sense flors).
L’aproximació alternativa, en canvi, consisteix a procedir per agrupació, és a dir, considerar primer les espècies (els grups més petits) i després anar-les agrupant de manera progressiva en funció de criteris de similitud, fins a arribar als grans grups d’organismes. Hom pot pensar que ambdós procediments lògics són equivalents i que, en conseqüència, al final han de conduir cap a un mateix resultat, però no és pas així, atès que en el primer cas els criteris de divisió seqüencial s’estableixen a priori, mentre que en el segon els criteris d’agrupament depenen de l’observació experimental dels organismes.
El següent pas en el desenvolupament de les classificacions va ser el de Joseph Pitton de Tournefort (1656 – 1708), botànic de Montpeller, que va proposar per primer cop d’agrupar les espècies més similars dins un mateix gènere, la qual cosa va donar pas a la concepció de les classificacions en diversos nivells jeràrquics consecutius, o tàxons, a partir de la iteració dels processos de classificació. Aquest concepte va ser codificat per Carl von Linné (1707 – 1778), el qual, per classificar les plantes, va establir els tàxons de nivell decreixent de regne, classe, ordre, gènere, espècie i varietat, tàxons que en la classificació dels animals van rebre el nom de regne, branca, classe, ordre, família, gènere i espècie (una jerarquia en set nivells que respectava escrupolosament la concepció de perfecció que tradicionalment s’ha atribuït al nombre set).
Tornant als dos sistemes lògics de classificació, el divisori i el d’agrupació, Linné va emprar una combinació d’ambdós: el divisori per als tàxons superiors al gènere, i el d’agrupació per a les espècies i els gèneres. Aquest és el motiu conceptual pel qual Linné va proposar d’anomenar les espècies de forma binària, com s’ha vist en l’apartat 2.1.
De Linné a Darwin
Dos arbres filogenètics clàsics. A l’esquerra, la primera mostra gràfica de pensament genealògic global de l’arbre de la vida: l’arbre que Charles R. Darwin va utilitzar en l’obra L’origen de les espècies (1859) per a explicar el concepte de selecció natural. A la dreta, arbre filogenètic elaborat per Ernst Haeckel poc temps després, el 1866, que situava els diversos grups de vertebrats amb relació als cordats no vertebrats (amfiox, Leptocardia) i els llinatges que englobava cadascun segons els coneixements de l’època.
A partir d’aquestes propostes de classificació va néixer un profund interès per trobar una lògica que permetés accedir a les classificacions naturals; o, dit d’una altra manera, per trobar l’ordre dins la natura. Així, els naturalistes francesos Bernard de Jussieu (1699 – 1777) i Antoine-Laurent de Jussieu (1748 – 1836) es van adonar, en examinar els caràcters emprats per a definir un determinat nivell taxonòmic, que no tots són equivalents, una observació que els va permetre establir el principi de subordinació dels caràcters, una autèntica revolució taxonòmica. Aquest principi postula que, per a definir un tàxon, és millor fer servir pocs caràcters però que siguin presents en totes les espècies que aquest engloba que no pas utilitzar molts caràcters però que no es trobin en totes les espècies analitzades. Aquest principi fou emprat per a construir la primera classificació botànica que pot ser considerada com a natural, atesos els coneixements de l’època. També fou aplicat pel naturalista i zoòleg francès Georges Cuvier (1769 – 1832) per a classificar els animals en quatre grans grups atenent al seu pla d’organització corporal: els vertebrats, els artròpodes, els mol·luscs i els radiats. Totes aquestes classificacions, tanmateix, consideraven l’ordre natural com a immutable, en el sentit d’estàtic i en contraposició a dinàmic, un fet que es reflectia en l’essència de les classificacions.
Poc després, però, amb la publicació de les primeres teories evolutives, les classificacions van donar un nou tomb conceptual. Així, amb la teoria evolutiva del naturalista francès Jean-Baptiste Lamarck (1744 – 1829), segons la qual el motor de l’evolució és l’herència dels caràcters adquirits (una explicació que ha demostrat no ser correcta), i la dels naturalistes anglesos Charles Darwin (1809 – 1882) i Alfred Russel Wallace (1823 – 1913), que van proposar la selecció natural com a mecanisme per a explicar el fet de l’evolució (un mecanisme àmpliament demostrat), es va començar a considerar que totes les característiques dels organismes, incloses les utilitzades en les classificacions taxonòmiques, procedeixen d’un procés d’evolució. Això permet agrupar els organismes, ja no tan sols en funció de les seves similituds morfològiques, sinó també atenent al seu llinatge o, alternativament, al seu grau de parentiu, els quals han d’explicar aquestes similituds (i al mateix temps també poden ser interpretats a partir d’elles).
De fet, com s’ha comentat en un apartat precedent, l’única figura que consta en L’origen de les espècies de Darwin és un arbre, amb el qual pretenia il·lustrar el seu concepte de selecció natural, entesa com la supervivència d’aquelles formes que, en un moment concret, entre tota la variabilitat existent entre els organismes d’una mateixa espècie, resultaven més ben adaptades al medi. Una supervivència diferencial que, convenientment plasmada en una taxa característica de reproducció, es troba a la base de tots els llinatges d’éssers vius i dels nodes a partir dels quals es bifurquen les diverses ramificacions. Arribats a aquest punt, ja només falta incorporar les dades i les tècniques genètiques i moleculars per a arribar als sistemes de classificació i als arbres filogenètics actuals, un procés de desenvolupament de la sistemàtica i la taxonomia que va durar encara pràcticament un segle més.
De Darwin a Hennig
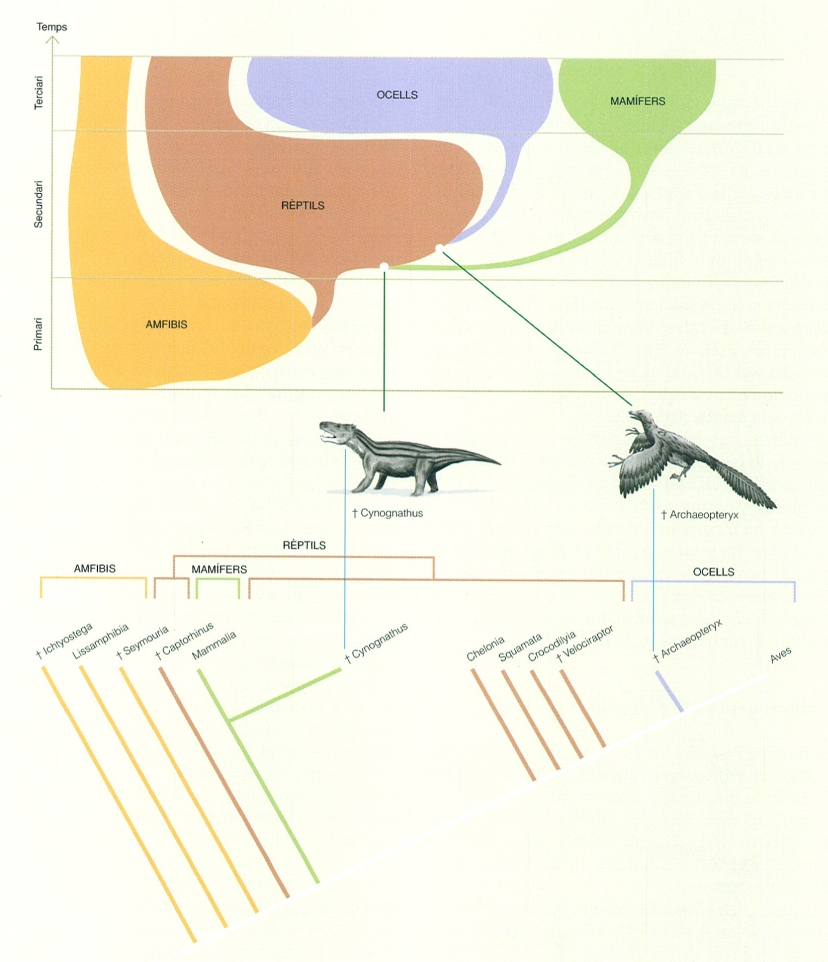
Comparació de la interpretació que feia Alfred S. Romer (a dalt) i l’obtinguda per l’anàlisi cladística (a baix) de les relacions entre diversos tàxons de vertebrats. Com a exemple, s’hi pot veure la posició dels fòssils Cynognathus i Archaeopteryx, que Romer situa al node dels embrancaments. També es pot comprovar que en l’anàlisi cladística actual els rèptils resten separats en dos grups pels mamífers, que són al mig. (†) indica els grups extingits.
A partir de Romer i Parsons, 1986 i fonts diverses Nobumichi Tamura i Biopunt.
A partir dels treballs de Darwin, i durant un centenar d’anys, diversos naturalistes es van dedicar a classificar els éssers vius amb l’afany de fer coincidir les classificacions establertes amb llurs idees sobre la genealogia i la filogènia de la vida (és a dir, sobre qui descendeix de qui i sobre qui està més emparentat amb qui, respectivament), però sense un mètode de treball veritablement explícit. Cal destacar, per exemple, el treball realitzat pel biòleg, naturalista, filòsof i físic alemany Ernst Haeckel (1834 – 1919), que a més d’encunyar diversos termes de la biologia, com el mot ecologia, també en va crear d’altres relatius a les anàlisis filogenètiques, com fílum, filogènia i el nom del regne Protista, i va proposar un arbre filogenètic dels vertebrats fent servir encertadament l’amfiox, un cordat no vertebrat, com a grup extern per a determinar l’arrel de l’arbre.
En aquesta mateixa línia, cal destacar també les arborescències realitzades pel paleontòleg americà Alfred S. Romer (1884 – 1973), en què barrejava les relacions entre avantpassats i descendents (relacions genealògiques) amb les relacions de parentiu (relacions filogenètiques), i en les quals situava, en els nodes d’embrancament, com a ancessors comuns, els fòssils coneguts. A més, en la construcció dels seus arbres també incorporava consideracions adaptatives i ecològiques i criteris de complexitat relativa i de similitud global, una aproximació anomenada fenètica, de la qual es parlarà més endavant, a més de les estrictament morfològiques.
Ara bé, la introducció de consideracions adaptatives i ecològiques en les anàlisis filogenètiques, com feia Romer, pot emmascarar les relacions reals. Per exemple, si hom intenta establir la filogènia dels peixos en relació amb els altres vertebrats atenent a criteris ecològics i de similitud global, situarà dins un mateix grup els peixos actinopterigis, com la truita Salmo trutta, l’espècie oriünda dels Països Catalans, i els dipnous o peixos pulmonats, com Neoceratodus fosteri, propi d’Austràlia, atès que ambdós ocupen hàbitats aquàtics d’aigua dolça, tenen el cos afusat hidrodinàmic i presenten aletes amb una aparença externa relativament similar. Les característiques principals dels peixos actinopterigis, però, són posseir un esquelet d’espines òssies en les aletes, tenir el crani cartilaginós i en part calcificat, recobert per fusos dèrmics, i presentar un sol parell d’obertures branquials cobertes per un opercle. En canvi, els dipnous presenten, entre altres característiques distintives, pulmons i un esquelet ossi ben desenvolupat dins les extremitats en forma d’aletes. Així mateix, les anàlisis filogenètiques situen els dipnous dins el tàxon dels ripidistis, el mateix grup que inclou tots els altres vertebrats llevat, precisament, dels peixos actinopterigis.
Finalment, el darrer gran avenç conceptual en la història de les classificacions va ser el de la sistemàtica filogenètica (o cladística), fundada el 1950 pel biòleg alemany Willi Hennig (1913 – 1976), a la qual s’ha incorporat més recentment la filogènia molecular. La seva base, que es desgranarà en els apartats posteriors, és agrupar els organismes en tàxons que incloguin un ancessor comú i la totalitat de la seva descendència, una tipologia de grups que fa que rebin el nom de monofilètics.
L’adveniment de la filogènia molecular
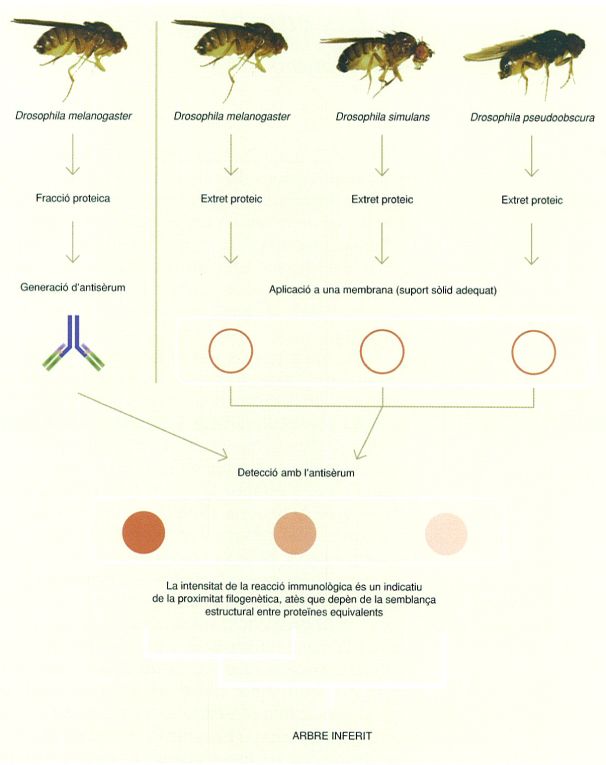
Esquema de l’experiment per a inferir un arbre filogenètic a partir de la relació serològica o immunològica de tres espècies de mosca de la fruita (Drosophila). En l’esquema es mostren només els passos principals: s’obté un extret proteic de cadascuna de les tres espècies, que es tracta amb l’antisèrum obtingut de la fracció proteica de D. melanogaster; la intensitat de la reacció immunològica indica la proximitat filogenètica, atès que depèn de la semblança estructural entre proteïnes equivalents. L’experiment és del mateix tipus que el que va fer George H.F. Nuttall per a inferir quins eren els organismes evolutivament més propers als humans.
Original de l’autor/ David Bueno.
La filogènia molecular és l’estudi de les relacions evolutives entre organismes utilitzant tècniques i eines de la biologia molecular, una incorporació no tan sols metodològica sinó també conceptual que a partir de la darrera dècada del segle XX ha canviat el panorama de les classificacions i de la mateixa filogènia.
Els inicis conceptuals de la filogènia molecular, però, cal situar-los al final del segle XIX, abans del redescobriment de les anomenades lleis de Mendel l’any 1900. Per exemple, l’any 1904 el biòleg d’origen nord-americà George H.F. Nuttall (1862 – 1937) va utilitzar anticossos en reaccions serològiques (o immunològiques) per a inferir les relacions filogenètiques entre diversos grups d’organismes, atès que la intensitat d’aquestes reaccions es correlaciona amb la seva proximitat evolutiva. Així, va determinar que els parents vius més propers als humans són els grans simis, seguits dels micos del Vell Món, els micos del Nou Món i els prosimis, respectivament.
Tanmateix, fins que aquestes tècniques no es van desenvolupar sistemàticament, no es van començar a aplicar de forma consistent en estudis filogenètics. Així, durant les dècades de 1960 i 1970, els avenços en separació de proteïnes i en hibridació d’àcids nucleics (DNA i RNA) i el desenvolupament dels mètodes immunològics van permetre avenços significatius en l’establiment de relacions filogenètiques entre espècies molt emparentades, malgrat que aquests mètodes no fossin tan acurats com els de seqüenciació de DNA o de proteïnes (però en canvi eren molt més ràpids i econòmics). A més, l’aplicació d’aquests mètodes va estimular el desenvolupament de les tècniques de mesura de distància genètica i els mètodes de construcció d’arbres filogenètics.
A partir del 1970, l’acumulació d’un nombre creixent de seqüències de DNA va tenir un gran impacte en la filogènia molecular, atès que aquestes seqüències proporcionen una gran quantitat de dades fàcilment analitzables i comparables de manera molt més objectiva. Finalment, l’aparició de les tècniques automàtiques de seqüenciació de DNA i el desenvolupament d’eines bioinformàtiques cada cop més potents han permès la seqüenciació d’un gran nombre de gens i de multitud de genomes complets (el genoma és el conjunt de totes les seqüències de DNA d’un organisme, les quals inclouen tots els seus gens), i també l’anàlisi comparativa de totes aquestes dades, la qual cosa ha incrementat exponencialment la robustesa dels arbres i el nombre d’espècies utilitzades.
Els fonaments filosòfics de les classificacions
Hom no pot acabar una història de les classificacions sense esmentar el concepte de Scala Naturae o ‘escala de la natura’, àmpliament desenvolupat pel filòsof i científic alemany Gottfried W. Leibniz (1646 – 1716), les bases del qual es remunten als filòsofs grecs Plató i Aristòtil, ateses les implicacions que ha tingut (i a vegades, en certs aspectes, encara té) en les classificacions i en les filogènies. Leibniz va proposar el principi de gradació lineal de la classificació dels éssers vius, que condueix, en cada graó, a un grau superior de perfecció, i al final de la qual va situar l’espècie humana. Aquest concepte de Scala Naturae es troba fortament arrelat a la cultura occidental i és l’origen d’alguns errors conceptuals comunament acceptats i, per tant, difícils de corregir. Entre aquests errors, els més destacables són l’antropocentrisme, el finalisme i l’essencialisme.
De l’antropocentrisme n’hem tret la visió que els humans es troben al final de l’escala de la natura, al graó de més perfecció. Des d’una òptica evolutiva, aquesta idea es tradueix en la consideració de l’espècie humana com la més evolucionada de totes, quan en realitat totes les espècies estan igualment evolucionades, però cadascuna respecte a unes característiques concretes, les que han estat afavorides per la selecció natural en cada contingència concreta entre tota la varietat morfològica i fisiològica existent en aquella població. Totes provenen d’un mateix ancestre comú, a partir del qual han anat evolucionant fins a la seva forma actual.
L’antropocentrisme és un factor que comporta el manteniment d’unes classificacions de determinats grups que no tenen cap parentiu filogenètic proper. N’hi ha uns quants exemples: el dels procariotes, que són un grup sense cap significat filogenètic però que tanmateix encara són presents en moltes classificacions, formats per cèl·lules sense nucli, que actualment representen dos dels tres grans dominis que constitueixen els éssers vius; el dels invertebrats (o metazous sense vèrtebres), representats en diversos grups que no comparteixen cap ancestre comú immediat; el dels peixos en sentit ampli, que són els craniats sense extremitats i que inclouen els peixos pulmonats, malgrat que aquests estiguin molt més emparentats amb els tetràpodes que amb la resta de peixos; els rèptils, que són els amniotes sense pèls, etc. En tots aquests casos, el factor antropocèntric queda reflectit en la comparació d’aquestes característiques amb les pròpies de l’espècie humana (organismes eucariotes per oposició a procariotes, vertebrats per oposició a invertebrats, tetràpodes per oposició a vertebrats sense extremitats evidents, i amniotes amb pèl per oposició a amniotes sense pèl, respectivament).
Respecte al finalisme, cal emfasitzar que la forma com es representen moltes classificacions, amb els mamífers i dins seu els humans al capdamunt, indueix a pensar que l’evolució ha conduït a l’aparició dels humans per se, però la selecció natural és l’únic element que afavoreix unes variants concretes entre tota la variabilitat present en una població d’organismes en funció de les contingències de cada moment. Dit d’una altra manera, com en la història, si tot tornés a començar, la probabilitat que tot passés de la mateixa manera és pràcticament nul·la.
Finalment, l’essencialisme porta a veure la natura des d’un punt de vista filosòfic, és a dir, que hom es pregunti sobre el perquè de l’existència, però la filogènia, com a disciplina científica, respon al com i al quan dels processos que han portat els organismes a les seves formes actuals, però en cap cas al perquè. En altres paraules, l’essencialisme construeix les entitats a priori i després les fa encaixar dins la realitat, una aproximació completament oposada a la de la filogènia, en què les entitats (els tàxons) es formulen i es reformulen tantes vegades com faci falta en funció de la realitat (o, més ben dit, de les anàlisis filogenètiques d’aquesta realitat).
Cal tenir en compte totes aquestes consideracions filosòfiques per a entendre com la filogènia actual ha fet variar la classificació dels éssers vius i, en definitiva, l’arbre de la vida, i per què continuarà canviant fins a arribar a l’arbre de la vida definitiu.
Els caràcters en taxonomia i filogènia
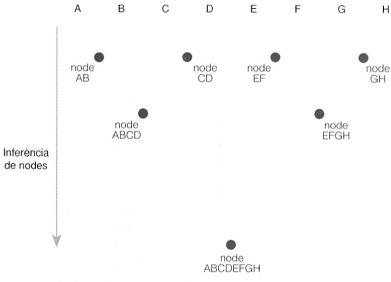
L’anàlisi dels caràcters dels organismes en el seu estat actual o fòssil i la seva comparació filogenètica permeten inferir els nodes teòrics a partir dels quals van sorgir els llinatges respectius. Així, per exemple, en l’arbre que relaciona els organismes A, B, C, D, E, F G i H, el node AB reuneix els llinatges A i B basant-se en la comparació de les característiques compartides actuals, les quals haurien de trobar-se en el seu darrer avantpassat comú; el node ABCD reuneix els llinatges procedents del node AB i del node CD (el qual, al seu torn, prové de la reunió dels llinatges C i D), i així successivament.
Original de l’autor.
En ciències naturals, la sistemàtica és la ciència de les classificacions. El seu primer objectiu és identificar i descriure els éssers vius presents i passats, per fer-ne un inventari tan complet com sigui possible quant a diversitat i característiques. Després ve la classificació pròpiament dita, que consisteix a fer una ordenació lògica que permeti entendre la diversitat dels organismes i les seves relacions de parentiu.
Totes les espècies són el resultat d’una llarga història evolutiva. Si hom pogués retrocedir en el temps, veuria com els diferents llinatges es van reunint consecutivament amb altres llinatges, en nodes que van agrupant més i més organismes com més a la base de l’arbre de la vida es troben. Tanmateix, per a establir aquestes relacions, hom no disposa de la història evolutiva completa dels organismes, sinó dels resultats visibles en el seu estat actual o fòssil, de les característiques presents en els organismes que es troben a la punta de les branques, en forma de caràcters morfològics, fisiològics i també moleculars. L’anàlisi d’aquests caràcters i la seva comparació filogenètica permeten inferir els nodes a partir dels quals van sorgir llurs respectius llinatges, uns teòrics avantpassats comuns que devien presentar els caràcters compartits que són presents en els grups actuals, en un retrocés teòric que permet construir arbres fïlogenètics i reconstruir l’arbre de la vida.
Els caràcters morfològics i fisiològics
Un caràcter és un atribut observable d’un organisme. Les classificacions clàssiques generalment es fonamenten en caràcters morfològics dels organismes i també, de vegades, en aspectes fisiològies, molt especialment en els organismes procariotes. La immensa majoria de les classificacions i dels arbres elaborats abans de la dècada de 1980 es van fer emprant aquest tipus de caràcters.
Els caràcters en taxonomia
Tots els organismes presenten una gran quantitat de caràcters observables i analitzables, però no tots són igualment útils o vàlids per a fer una bona classificació. Cal que els caràcters siguin comparables i que, a més, es puguin comparar. És a dir, cal que es trobin en els organismes que hom vol estudiar i que siguin similars, però també cal que no siguin idèntics, de tal manera que hom en pugui reconèixer com a mínim dos estats diferents pertanyents a un patró general comú.
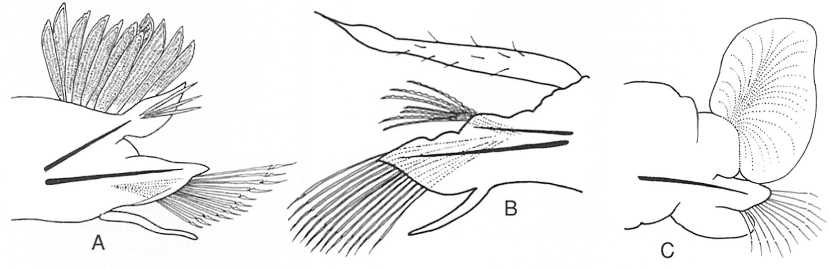
Els parapodis dels poliquets, exemple de caràcter vàlid per a analitzar en taxonomia: són comparables però no idèntics, i n’hi ha més de dos estats diferents: A birrami (Chrysopetalum debile), B subbirrami (Harmothoe areolata) i C unirrami (Anaitides maculata).
Jordi Corbera.
Com s’ha dit, un caràcter és la designació d’allò que hom observa. Per exemple, els parapodis dels poliquets, uns apèndixs laterals parells situats als metàmers somàtics d’aquests cucs marins. L’estat, en canvi, és la forma concreta que té aquell caràcter en l’organisme analitzat. En l’exemple, el caràcter parapodis pot presentar diversos estats, que són específics de cada espècie, per la qual cosa poden ser utilitzats en la taxonomia dels poliquets. Així, dins el caràcter parapodis hom pot trobar l’estat parapodis birramis (formats per dues branques similars), parapodis subbirramis (formats per dues branques de diferent morfologia), parapodis unirramis (formats per una sola branca), etc.
Pel que fa als caràcters fisiològics o metabòlics, són emprats sovint en la taxonomia de procariotes, atesa la dificultat d’observació de caràcters morfològics en aquests organismes i la limitació a uns pocs estats diferents. Per exemple, per a classificar bacteris del tàxon dels enterobacteris, hom utilitza un test anomenat IMVIC que consisteix a analitzar, mitjançant assaigs microbiològics i bioquímics, quatre aspectes del seu metabolisme: la capacitat de convertir l’aminoàcid triptòfan en indol (I), un altre compost orgànic; la capacitat de produir àcids, que es detecta amb el roig de metil (M), un indicador de pH; la capacitat de fermentar glucosa per la via butanodiòlica, que s’efectua mitjançant la detecció d’acetil metil carbinol amb la prova bioquímica de Voges-Proskauer (V), i la capacitat de créixer fent servir citrat com a única font de carboni (C). L’acrònim IMVIC prové de les inicials dels productes que s’empren o es detecten en aquestes quatre proves consecutives. Segons aquest test, els bacteris del gènere Salmonella (uns bacils gramnegatius de distribució mundial, anaerobis facultatius, envoltats de flagels perítrics) es caracteritzen per donar negatiu a la primera prova (I-), positiu a la segona (M+), negatiu a la tercera (V-) i positiu a la quarta (C+), i per això es diu que el seu IMVIC característic és - + - +.
La relació entre els caràcters morfològics i fisiològics i els caràcters moleculars
L’adveniment i la posterior simplificació i difusió de les tècniques d’anàlisi molecular han permès incorporar una nova gamma de caràcters a les anàlisis taxonòmiques i filogèniques, procedents de les seqüències de determinades molècules, concretament dels àcids nucleics (DNA i RNA) i de les proteïnes, els quals mereixen un tractament específic. Tanmateix, els caràcters morfològics i fisiològics i els caràcters moleculars no són independents entre ells, sinó que estan íntimament relacionats, perquè la morfologia i la fisiologia depenen del contingut genètic específic de cada organisme, que els determina. O, dit d’una altra manera, els caràcters morfològics i fisiològics proporcionen el context global als caràcters moleculars, atès que són la conseqüència integrada de les seves funcions.
A més, els estudis taxonòmics i filogenètics també han impulsat altres disciplines científiques, de les quals, alhora, s’han nodrit, com per exemple, de l’anàlisi genètica del desenvolupament embrionari, atès que durant aquest procés la maquinària molecular dels organismes (els gens i les proteïnes) afaiçona i dirigeix l’adquisició de llurs característiques morfològiques i fisiològiques. També cal fer esment de la bioinformàtica, la disciplina científica que aplica els principis de la informàtica a l’estudi de fenòmens biològics, sense la qual no seria possible examinar les ingents quantitats de dades que proporcionen les anàlisis moleculars.
Caràcters homòlegs i caràcters anàlegs en taxonomia clàssica
Des d’una òptica comparativa, un caràcter és una estructura identificada com a similar en dos o més organismes, entenent per similar que pugui ser idèntica o bé presentar diversos estats. Tanmateix els caràcters que s’usen en taxonomia i filogènia han de tenir un mateix origen evolutiu en tots els organismes que es comparen. Això comporta que, en aquest context, la noció de caràcter sigui indissociable del concepte d’homologia.
Hom diu que un determinat caràcter és homòleg en dos o més organismes (o que presenta homologia) si ha evolucionat en tots ells a partir d’un mateix avantpassat comú que ja el presentava, malgrat que pugui aparèixer en els organismes actuals sota qualsevol variació de forma o mida. A escala molecular, aquestes variacions es plasmen en canvis de seqüència en les proteïnes i/o els àcids nucleics, dels quals es parlarà a continuació.
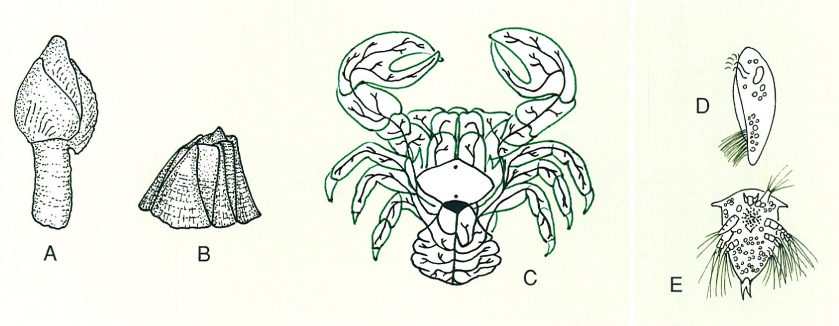
Comparació morfològica entre diversos tipus de cirrípedes. A percebe comú (Pollicipes pollicipes); B gla de mar (Semibalanus balanoides); C Briarosaccus callosus, paràsit, els teixits del qual (en negre) s’adapten completament a l’hoste (en verd, la forma del crustaci infectat); D i E vista lateral i frontal de la larva naupli corresponent, que és pràcticament idèntica en tots ells.
Agnès Perelló, a partir de Skelton i Gilmour, 1994.
Les homologies tenen una importància fonamental en les inferències filogenètiques, ja que només la comparació de caràcters homòlegs dóna informació consistent sobre les relacions de parentiu entre organismes. Per aquest motiu, el primer pas per a avaluar les relacions evolutives o de parentiu filogenètic és determinar quins dels molts caràcters que a priori hom podria emprar són homòlegs, tot analitzant llur correspondència topològica, embriològica i/o de composició. Així, atenent llur morfologia externa, hom difícilment podria determinar la presència de caràcters homòlegs entre els diversos membres adults de la infraclasse dels cirrípedes, un tàxon comprès dins el grup dels crustacis i que inclou, entre altres organismes, el percebe comú (Pollicipes pollicipes), els glans de mar (Semibalanus balanoides) i els briarosacs, uns cirrípedes paràsits que en l’etapa adulta no tenen cos amb forma pròpia perquè l’adapten completament a la forma del crustaci que parasiten (com per exemple Briarosaccus callosus). Tanmateix, però, tots passen per un mateix estadi larvari, el de naupli, durant el qual són morfològicament molt semblants. A partir d’aquestes semblances larvals hom pot establir les corresponents homologies topològiques en els adults. No obstant això, per a fer una classificació útil en la construcció d’arbres filogenètics, cal distingir entre dos tipus diferents d’homologia (la primària i la secundària) i cal també contraposar homologia a analogia.
Relacions d’homologia i analogia possibles entre extremitats anteriors i ales. Dins els tetràpodes, el cas de la geneta és homòleg quant al fet de ser extremitat anterior, però no amb relació a les ales.
Agnès Perelló.
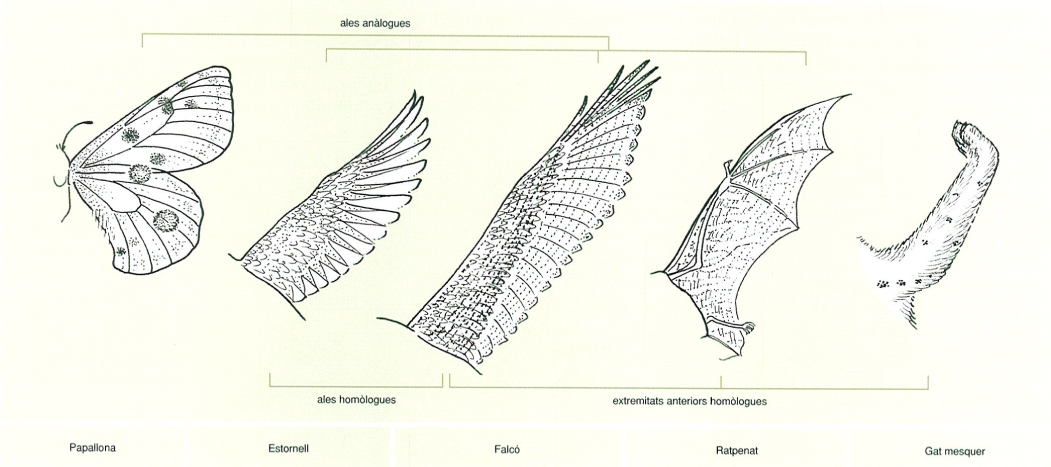
Un caràcter presenta homologia secundària en dos o més organismes si prové, en tots, d’un mateix ancestre comú, per divergència evolutiva. Dit d’una altra manera, l’homologia secundària és l’homologia per descendència. Aquesta homologia, però, no és operacionalment bona en la construcció d’arbres filogenètics, ja que només té sentit dins un marc taxonòmic concret. Sense un marc taxonòmic de referència, dins l’arbre general de la vida tot caràcter pot acabar sent homòleg de qualsevol altre caràcter, atès que tots els organismes procedeixen d’un mateix avantpassat comú, a l’origen de la vida, a partir del qual han anat divergint els diversos llinatges actuals de manera progressiva. Per exemple, dins el marc taxonòmic dels ocells, les ales dels estornells són homòlogues a les dels falcons, atès que ambdues procedeixen d’una ala ancestral per descendència. Però si hom considera el marc taxonòmic general dels tetràpodes, les ales dels estornells també són homòlogues de les extremitats anteriors de les genetes, atès que ambdues procedeixen d’una extremitat anterior ancestral per descendència. De la mateixa manera, les ales dels estornells i les ales dels ratpenats també són homòlogues dins el marc taxonòmic dels tetràpodes quant a extremitats anteriors, pero tanmateix, i aquí és on radica el problema de les homologies secundàries, no són pas homòlogues quant al fet de ser ales, atès que no provenen d’una mateixa ala ancestral, sinó de dos processos independents de divergència evolutiva a partir d’extremitats anteriors de tetràpodes ancestrals, malgrat que hagin conduït a unes estructures funcionalment similars. Per aquest motiu, l’homologia secundària o per descendència no és operacionalment bona en la construcció de filogènies.
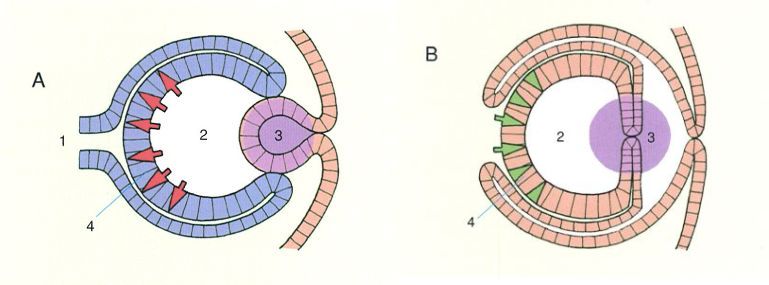
Analogia entre els ulls dels cefalòpodes i els dels vertebrats. L’esquema mostra el diferent origen embrionari dels ulls dels vertebrats (A) i dels cefalòpodes (B), que és un reflex del diferent origen filogenètic. 1 nervi òptic, 2 cos vitri, 3 lent, 4 retina.
Agnès Perelló, modificat a partir de Fernald, 2000.
En aquest context, els caràcters que tenen una estructura i un origen filogenètic diferents però que tanmateix presenten una semblança funcional, es diu que són anàlegs (o que presenten analogia). Per exemple, les ales dels ocells són anàlogues a les ales de les papallones, atès que tant l’estructura com l’origen embrionari i evolutiu són diferents, i que presenten una semblança funcional (adaptar aquests organismes al vol). Passa el mateix, per exemple, amb els ulls dels cefalòpodes i dels vertebrats, o amb les espines de l’argelaga i del grèvol. Així, doncs, els ulls dels cefalòpodes i dels vertebrats obeeixen a una mateixa funció i en aquest cas, a més a més, tenen una estructura anatòmica global molt semblant, tanmateix els dels cefalòpodes es formen a partir d’una invaginació de l’ectoderma que entra en contacte amb la neuroectoderma, i per tant són constituïts majoritàriament per cèl·lules ectodèrmiques, mentre que els dels vertebrats es formen a partir d’una projecció de la neuroectoderma que entra en contacte amb l’ectoderma, i per tant són constituïts majoritàriament per cèl·lules neuroectodèrmiques. Aquestes diferències quant a processos de desenvolupament i quant a origen embrionari de llurs cèl·lules indiquen un origen filogenètic diferent i, en conseqüència, que es tracta de caràcters anàlegs. En el cas de les espines de l’argelaga (Genista scorpius) i del grèvol (Ilex aquifolium), les de l’argelaga tenen un origen caulinar, a partir de cèllules modificades de la tija, mentre que les del grèvol són foliars. Per aquest motiu han de ser considerats també caràcters anàlegs.
Homologia primària entre el radi (en groc) de sis espècies diferents de tetràpodes. Malgrat que la funció (contribuir a volar, caminar, saltar i nedar) i la forma del radi són diferents en cada cas, sempre es troba en un mateix pla d’organització (l’extremitat anterior) i manté la mateixa relació amb les estructures veïnes, atès que se situa entre l’húmer i els ossos carpians.
Agnès Perelló.
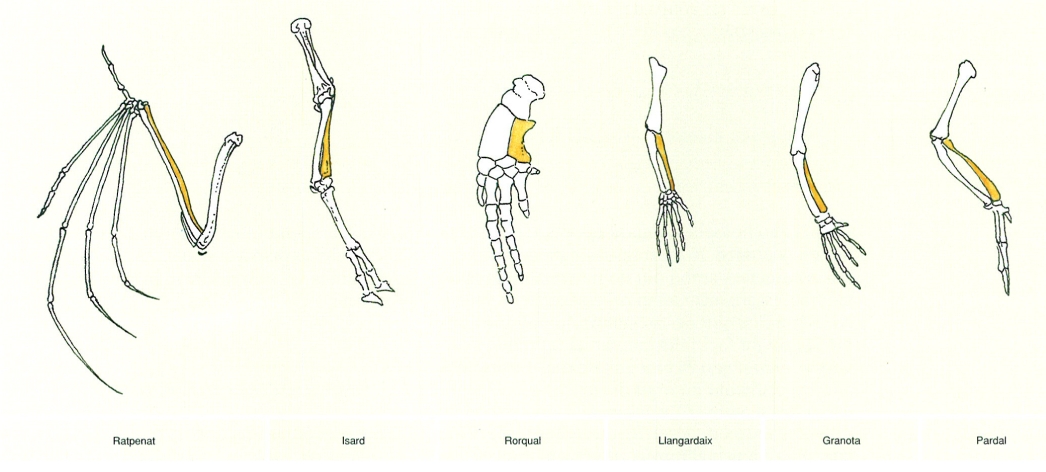
Finalment, hom diu que un caràcter present en dos o més tipus d’organismes presenta homologia primària si la seva similitud es basa en la comparació d’estructures o característiques morfològiques o moleculars que comparteixen un mateix pla d’organització, és a dir, que mantenen les mateixes connexions o relacions amb les estructures veïnes, amb independència de la seva forma o funció. Per exemple, el radi dels tetràpodes, l’os ubicat a la part externa de les extremitats superiors, presenta homologia primària en tots els organismes del grup, atès que es troba circumscrit en un mateix pla d’organització, el de l’extremitat anterior, i manté les mateixes relacions amb els ossos veïns. Així, per exemple, els radis d’un rorqual (Balaenoptera physalus), una balena relativament comuna davant les costes dels Països Catalans, d’un isard (Rupicapra rupicapra), d’un ratpenat com el de ferradura gros (Rhinolophus ferrumequinum), d’un llangardaix ocel·lat (Timon lepidus), de la granota roja (Rana temporaria) i del pardal comú (Passer domesticus) presenten homologia primària atès que, malgrat que la forma no sigui la mateixa ni tampoc sigui exactament idèntica la funció global de l’estructura on es troba, en tots els casos es localitza entre l’húmer i els ossos carpians, circumscrit dins un mateix pla d’organització. És a dir, per a formular una hipòtesi d’homología hom s’ha de circumscriure a un pla d’organització concret, una mena de lectura abstracta de les similituds entre estructures. L’homologia primària és operacionalment útil en la confecció d’arbres filogenètics.
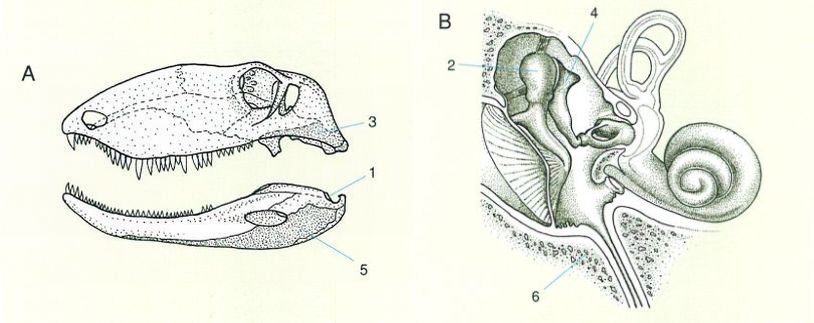
Homologia primària entre els ossos mandibulars d’un mamífer marsupial típic (A) i els de l’oïda mitjana d’un mamífer placentari (B). Els estudis embriològics (ontogenètics) han demostrat que l’os articular (1) és homòleg del martell (2); el quadrat (3) ho és de l’enclusa (4), i l’angular (5), de l’os timpànic (6), entre d’altres.
Agnès Perelló i Amadeu Blasco.
Els caràcters amb homologia primària han de tenir necessàriament un mateix origen embrionari. Això implica que per a establir homologies primàries sovint faci falta emprar dades ontogenètiques (l’ontogènesi és el seguit de transformacions que experimenta un organisme des de la fecundació fins a atènyer l’estat madur). Per exemple, diversos ossos de l’estructura mandibular dels marsupials tenen el mateix origen embrionari que determinats ossets interns de l’orella mitjana dels mamífers placentaris, per la qual cosa, malgrat que la seva estructura i funció siguin diferents, han de ser considerats com a homòlegs primaris, i en conseqüència la seva comparació pot ser emprada en filogènesi.
Els caràcters moleculars

Representació dels estats que pot presentar un caràcter molecular en l’exemple concret dels aminoàcids de l’hèlix E de la flavohemoglobina de tres espècies de fongs aspergils. Cada aminoàcid (representat per una lletra) és un caràcter diferent. L’alineació de les seves seqüències parcials permet quantificar-ne la similitud de manera objectiva, indicada per asteriscs: un de sol indica la presència de diferències entre la primera i la segona respecte de la tercera; dos, diferències entre la primera i la tercera respecte de la segona; i tres, diferències entre totes tres.
A partir de Biesebeke i altres, 2009.
De la mateixa manera que en taxonomia hom pot considerar els caràcters morfològics i fisiològics, també es poden utilitzar caràcters moleculars. Un caràcter molecular designa una posició específica dins d’una molècula determinada (no pas tota la molècula), com per exemple "l’aminoàcid que es troba en la posició número 59 de la flavohemoglobina" (la flavohemoglobina és una proteïna fúngica de la família de les globines que té una funció relacionada amb la respiració mitocondrial, l’estrès oxidatiu i l’estrès per compostos nitrogenats), o una posició determinada d’un nucleòtid en una seqüència de DNA. En aquest context molecular, l’estat és la forma concreta que té aquell caràcter en els organismes analitzats. Per exemple, en la posició 59 de la flavohemoglobina dels aspergils, un grup de floridures de coloració diversa, hom pot trobar glicina en organismes del gènere Aspergillus o alternativament arginina en els del gènere Ralstonia. Aquests aminoàcids representen dos estats diferents per al caràcter "posició aminoacídica 59 de la flavohemoglobina".

Seqüència de nucleòtids i aminoàcids dels segments codificants del gen i de la proteïna WOX4 de la planta de la mostassa (Arabidopsis thaliana), tal com apareixen al Gen-Bank del National Center for Biotechnology Information (NCBI) dels Estats Units, un dels principals bancs de dades moleculars. Aquest gen està implicat en la diferenciació de les cèl·lules del càmbium vascular en les plantes i presenta certes diferències filètiques.
GenBank de l’NCBI.
El fet que cada posició dins una molècula pugui ser considerada com un caràcter concret implica que en l’anàlisi d’una sola molècula hom pugui identificar molts caràcters diferents i en diferents estats, la qual cosa genera una quantitat de dades molt superior a les que normalment s’obtenen de la morfologia i la fisiologia. Tanmateix, com en el cas dels caràcters morfològics i fisiològics, no tots els caràcters d’una molècula ni totes les molècules són igualment útils o vàlids en taxonomia i filogènia. També cal que les molècules i els caràcters siguin comparables i que, a més, es puguin comparar. És a dir, cal que les molècules que hom compara siguin similars però no exactament idèntiques, de tal manera que es puguin reconèixer com a mínim dos estats diferents pertanyents a un patró general comú. I, per descomptat, cal que les molècules siguin homòlogues (d’homologia molecular se’n parlarà més endavant, en aquest mateix apartat).
En taxonomia i filogènia molecular s’empren dos tipus de molècules diferents: els àcids nucleics, que inclouen el DNA i l’RNA, i les proteïnes. En tots dos casos, les inferències filètiques es basen en la comparació de la seqüència de molècules homòlogues, és a dir, en l’ordre concret de les molècules bàsiques constituents que formen els àcids nucleics i les proteïnes.
Els àcids nucleics i les proteïnes
Les molècules que s’empren en filogènia molecular són els àcids nucleics i les proteïnes, atès que la seva seqüència és un reflex de la història evolutiva del seu llinatge; de fet, en sentit ampli, en són alhora causa i conseqüència. Els àcids nucleics constitueixen una família de biomolècules formades per la unió lineal d’unes molècules més senzilles, els nucleòtids, que es van repetint al llarg d’una cadena no ramificada de longitud variable. Hi ha dos tipus d’àcids nucleics: l’àcid desoxiribonucleic, o DNA, i l’àcid ribonucleic, o RNA. El DNA és format per quatre nucleòtids diferents: la citosina (C), la guanina (G), l’adenina (A) i la timina (T). L’RNA, al seu torn, també és constituït per quatre nucleòtids, tres dels quals són comuns amb el DNA, però conté uracil (U) en comptes de timina (T).
Les proteïnes, en canvi, són formades per la unió també lineal i no ramificada d’unes altres molècules constituents: els aminoàcids. Cal parlar d’aquestes molècules atès que totes s’empren en anàlisis filogenètiques.
El DNA
Esquema del cromosoma d’una cèl·lula procariota (A) i d’una cèl·lula eucariota (B) amb el seu nucli(5). S’hi observen els nucleòtids (1) que s’aparellen en la doble hèlix del DNA (2), el plegament del DNA (3) fins a constituir el cromosoma (4) i la diferent morfologia del cromosoma, que és circular en la immensa majoria de les cèl·lules procariotes i lineal en tots els eucariotes.
Adaptat a partir de Bueno, 2008.
El DNA és la molècula encarregada d’emmagatzemar la informació genètica i de transmetre-la d’una generació a la següent. Les molècules de DNA són formades per dues cadenes d’àcids nucleics que es disposen en forma de doble hèlix, amb els nucleòtids aparellats seguint una complementarietat específica: una T en una de les cadenes sempre s’aparella amb una A en l’altra cadena, i una C sempre ho fa amb una G. Aquesta doble hèlix, que constitueix la fibra de DNA, s’empaqueta amb l’ajut de proteïnes específiques per formar els cromosomes.
Cada molècula de DNA conté molts gens. Els gens són les unitats bàsiques de l’herència, les unitats funcionals que determinen les característiques biològiques dels organismes. Des d’un punt de vista molecular, un gen és un segment de DNA que fa una funció concreta, la qual és codificada en la seqüència dels seus nucleòtids. El conjunt de totes les seqüències de DNA d’un organisme és el seu genoma. El genoma humà, per exemple, és format per uns 3.200 milions de nucleòtids, i conté uns 24.000 gens.
Les proteïnes
Com s’ha dit, les proteïnes, també anomenades polipèptids, són formades per la unió lineal i no ramificada d’unes molècules més senzilles, els aminoàcids. De forma estàndard són formades per una combinació de vint aminoàcids diferents, malgrat que en alguns organismes es poden trobar altres aminoàcids, generalment modificacions específiques d’alguns dels vint aminoàcids estàndards, com la selenocisteïna i, en certs arqueus, la pirrolisina. La funcionalitat de les proteïnes depèn de la seqüència concreta dels aminoàcids que les formen, la qual depèn, al seu torn, de la seqüència de nucleòtids del gen corresponent.
Del DNA a les proteïnes amb la contribució de diversos tipus d’RNA
Com s’acaba de dir, la seqüència d’aminoàcids d’una proteïna és determinada per la seqüència de nucleòtids del gen corresponent, en una correspondència unívoca entre ambdues seqüències.
Del DNA a l’RNAm: la transcripció del missatge genètic
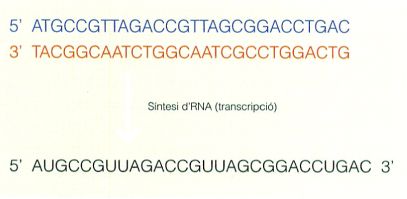
Esquema de la síntesi de l’RNA per mitjà de la transcripció de la informació continguda al DNA. El DNA és format per dues cadenes de nucleòtids aparellats. La cadena que es transcriu (en color vermell) serveix de motlle. L’RNA resultant (en color negre) té la mateixa seqüència de nucleòtids que la cadena acompanyant (en color blau) canviant la T per la U, i és complementària de la cadena que es transcriu.
A partir de fonts diverses.
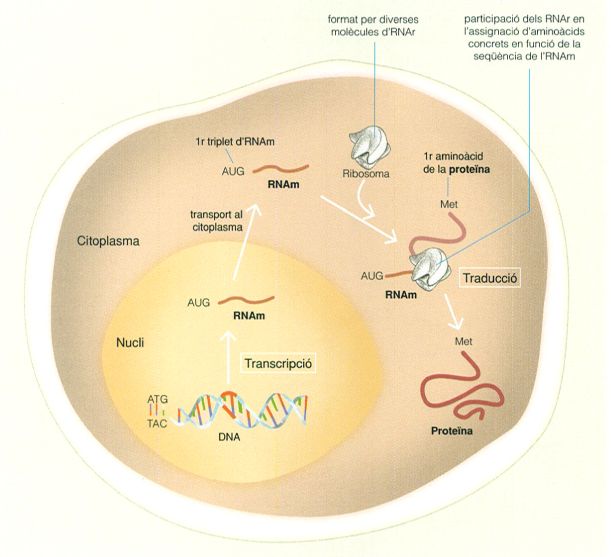
Flux d’informació gènica, que relaciona el DNA amb els diversos tipus d’RNA i porta a la síntesi de proteïnes. El procés de la traducció relaciona la seqüència de nucleòtids de l’RNAm (que al seu torn prové de la seqüència de nucleòtids del DNA) amb la seqüència d’aminoàcids de les proteïnes.
Barreras & Creixell, a partir de Bueno, 2008.
Per tal de sintetitzar una proteïna, el missatge contingut en la seqüència d’un gen es transcriu primer en una molècula intermediària d’RNA, anomenada RNA missatger, o RNAm. L’RNAm és l’encarregat de transportar la informació continguda als gens des del DNA fins als ribosomes, els orgànuls cel·lulars on es fabriquen les proteïnes. Els nucleòtids que formen l’RNAm són una còpia dels nucleòtids que formen una de les dues cadenes del DNA del gen corresponent, també en virtut de la complementarietat específica que s’estableix entre ells. Aquest procés de còpia s’anomena transcripció. En la transcripció intervenen diversos enzims; i, abans de ser funcional, l’RNAm experimenta un procés de maduració, que inclou l’addició de nucleòtids als seus extrems que li confereixen estabilitat i l’eliminació dirigida i controlada d’algunes seqüències internes no codificants (és a dir, que no contenen informació per a sintetitzar la proteïna corresponent), anomenades introns (vegeu més endavant).
De l’RNAm a la proteïna, amb la contribució de l’RNAr i l’RNAt: la traducció
Llavors, l’RNAm s’acobla als ribosomes, els quals descodifiquen el missatge i sintetitzen la proteïna corresponent, en un procés anomenat traducció. Els ribosomes, al seu torn, són formats també per unes altres molècules d’RNA, anomenades RNA ribosòmic, o RNAr. Els RNAr també estan codificats en gens concrets del genoma. Els organismes procariotes presenten tres tipus d’RNAr, que s’anomenen, segons el grau de sedimentació en unes condicions fisicoquímiques determinades: RNAr 5S, RNAr 23S i RNAr 16S. Els dos primers constitueixen l’anomenada subunitat gran del ribosoma, i l’altre, la subunitat petita.
Els organismes eucariotes, en canvi, presenten quatre tipus de RNAr: l’RNAr 5S, l’RNA 5,8S i l’RNA 28S, que constitueixen la subunitat gran del ribosoma; i l’RNAr 18S, que constitueix la subunitat petita del ribosoma. Atesa la funció crucial dels ribosomes, les seqüències dels RNAr s’han mantingut en organismes diferents, i són unes molècules molt emprades per a establir relacions filètiques. Així, per exemple, l’RNA 18S s’ha utilitzat per a construir la filogènia molecular dels fongs, els platihelmints, els tetràpodes i les angiospermes; l’RNA 28S també s’ha emprat per a construir la filogènia molecular dels tetràpodes, etc.
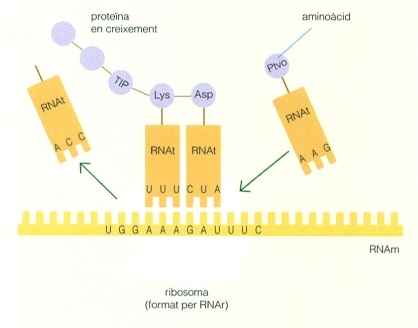
Procés de traducció, que relaciona la seqüència nucleotídica de l’RNAm (que al seu torn prové de la seqüència nucleotídica del DNA) amb la seqüència aminoacídica de les proteïnes. L’RNAt incorpora els aminoàcids a la proteïna en funció del missatge de l’RNAm.
A partir de fonts diverses.
Representació tridimensional d’un ribosoma eucariota, amb les seves subunitats gran (1) i petita (2), i disposició entrellaçada de les diverses molècules d’RNAr que les formen.
Agnès Perelló.
Finalment, un cop l’RNAm s’ha acoblat als ribosomes, intervé un altre tipus d’RNA: els RNA de transferència o RNAt, els quals també són codificats en gens concrets del DNA. La funció de l’RNAt és fer correspondre aminoàcids concrets a la seqüència específica de l’RNAm, una relació que s’anomena codi genètic. En filogènia, les dades procedents de la comparació de la seqüència nucleotídica dels RNAt s’han utilitzat, entre molts altres exemples, per a determinar l’origen d’orgànuls cel·lulars com els mitocondris i els cloroplasts a partir de bacteris ancestrals endosimbionts (teoria de l’endosimbiosi, àmpliament acceptada gràcies a aquestes i altres dades moleculars). En resum, totes les molècules que s’empren en filogènia es relacionen entre elles a través del flux d’informació gènica, nom genèric amb què es coneix el pas de dades moleculars del DNA a les proteïnes.
El codi genètic
El codi genètic, que estableix la correspondència específica entre els codons de l’RNAm (UUU, UUC, etc.) i aminoàcids concrets (Phe, Leu, etc). Hi ha tres codons que determinen el final de la síntesi de la proteïna corresponent, designats amb el nom de codons de terminació. A la dreta, nom dels aminoàcids i abreviatura en el codi de tres lletres i en el d’una lletra. En la taula del codi genètic es pot veure l’abast de la degeneració: diversos codons codifiquen un mateix aminoàcid.
A partir de fonts diverses.
Com s’ha dit, la relació que s’estableix entre el missatge contingut a l’RNAm i els aminoàcids que s’incorporen a les proteïnes constitueix el codi genètic. El codi genètic, que és universal en el sentit que és igual per a tots els organismes (llevat d’algunes excepcions menors, producte d’adaptacions particulars), fa correspondre un aminoàcid concret a cada tres nucleòtids consecutius de l’RNAm, sense excepció, una unitat de codificació d’informació que s’anomena codó o triplet.
Atès que hi ha quatre nucleòtids diferents, i que aquests es combinen de tres en tres per a formar els codons, hom pot trobar 64 codons diferents (és a dir, 64 combinacions diferents de tres nucleòtids, o 43 codons), però en la síntesi de proteïnes només s’empren vint aminoàcids diferents. Això fa que un mateix aminoàcid pugui ser codificat per diversos codons diferents, una característica del codi genètic que s’anomena degeneració. La degeneració del codi genètic és de gran importància en els estudis moleculars aplicats a la filogènia, atès que afecta les possibilitats que es fixin mutacions concretes i, en conseqüència, que es puguin observar diversos estats per a un caràcter molecular determinat (vegeu més endavant).
Per exemple, els codons UUU i UUC codifiquen tots dos el mateix aminoàcid, la fenilalanina, mentre que els codons UCU, UCC, UCA i UCG codifiquen, tots quatre, l’aminoàcid serina. També hi ha tres codons específics que en lloc de promoure la incorporació d’un aminoàcid a la proteïna determinen la fi de la traducció, el final de la síntesi de la proteïna corresponent, anomenats codons de terminació (del terme anglès stop codons).
Els tipus de DNA
Segons la seva localització cel·lular, hom pot distingir diversos tipus de DNA. Aquesta distinció és important perquè en funció del grup d’organismes i del nivell taxonòmic que es vulgui analitzar és més informatiu emprar dades d’un tipus de DNA o d’un altre. Així, hom distingeix el DNA nuclear, el DNA mitocondrial i el DNA del cloroplast.
El DNA nuclear és el que hi ha al nucli de les cèl·lules eucariotes o, per analogia, al nucleoide de les procariotes (en les procariotes el DNA no és segregat de la resta de components cel·lulars, però tanmateix la zona on es troba condensat s’anomena nucleoide). Aquest DNA conté la major part de la informació genètica dels organismes.
El DNA mitocondrial o DNAmt és a l’interior dels mitocondris de les cèl·lules eucariotes. Els mitocondris són els orgànuls cel·lulars encarregats de transvasar l’energia química continguda en les molècules d’aliment a altres biomolècules, des d’on la cèl·lula la pot utilitzar per a dur a terme les seves funcions vitals. El DNAmt conté un nombre molt reduït de gens en comparació amb el DNA nuclear, unes poques dotzenes, en funció de cada grup d’organismes. Els fongs i els animals, per exemple, tenen els mateixos gens en el seu DNAmt, però es troben ordenats de manera diferent, una informació que dóna consistència a la posició filètica d’aquests dos tàxons com a grups germans. Els gens presents al DNAmt mitocondrial exerceixen la seva funció en els mateixos mitocondris, els quals també reben moltes aportacions dels gens continguts al genoma nuclear. Tanmateix, les diferències d’ordre entre els gens del DNAmt i les diferències de seqüència són sovint emprades en filogènia molecular. A més, la gran similitud entre els gens i l’estructura morfològica del DNAmt i del cromosoma de determinats bacteris ha estat emprada per a bastir i donar consistència a la teoria endosimbiòtica d’aquests orgànuls.
De manera similar al DNAmt, els cloroplasts de les cèl·lules eucariotes també tenen les seves pròpies molècules de DNA, conegudes com a DNA del cloroplast o DNAcp. El DNAcp conté més de 100 gens, en funció de cada espècie, un nombre més elevat que en el DNAmt però també clarament inferior al que es troba en els genomes nuclears. Els cloroplasts són els orgànuls cel·lulars encarregats de convertir l’energia lluminosa en energia química, i de transvasar-la a determinades biomolècules des d’on les cèl·lules la poden utilitzar. També en aquest cas els gens presents en el DNAcp exerceixen la seva funció en els mateixos cloroplasts, els quals també reben moltes aportacions dels gens continguts al genoma nuclear. Els gens del DNAcp són emprats també en inferències filogenètiques en aquells organismes que contenen aquests orgànuls. Així mateix, la similitud entre els gens i l’estructura morfològica del DNAcp i del cromosoma de determinats bacteris fotosintetitzadors ha estat emprada per a bastir i donar consistència a la teoria endosimbiòtica respecte a aquests orgànuls.
Les mutacions en filogènia
La seqüència de nucleòtids que conforma qualsevol molècula de DNA no és constant en el temps, sinó que canvia. Aquests canvis són les mutacions, les quals constitueixen una font de variabilitat imprescindible per a l’evolució dels organismes. A més a més, aquests canvis proporcionen estats diferents als caràcters moleculars que hom compara en taxonomia i filogènia. La idea bàsica és que una major similitud entre dues seqüències de dos organismes diferents implica una major proximitat de parentiu entre aquests organismes. Les mutacions són errors que es produeixen durant el procés de còpia del DNA, un procés que esdevé abans de la reproducció cel·lular per assegurar que les dues cèl·lules filles heretin una collecció completa de tots els gens específics d’aquella espècie.
Els tipus de mutacions
Les mutacions es classifiquen segons el nombre de nucleòtids que afecten i la manera com afecten la seqüència de DNA. Per començar, poden afectar un únic nucleòtid (mutacions puntuals) o diversos nucleòtids adjacents. Respecte a com afecten la seqüència de DNA, hom distingeix les substitucions, que consisteixen en el bescanvi d’un nucleòtid per un altre; les delecions, que són la pèrdua d’un o més nucleòtids; les insercions, que consisteixen en l’addició d’un o més nucleòtids a la seqüència; les inversions, en què un segment de DNA queda invertit respecte a la seva polaritat inicial; les translocacions, en què un segment de DNA canvia de lloc i se situa en el mateix o en un altre cromosoma; i les duplicacions, en què un segment de DNA es duplica i queda incorporat en un altre lloc més o menys proper del mateix cromosoma o bé en un altre cromosoma. També es poden produir altres canvis més extensos, com per exemple, la duplicació de cromosomes i de genomes complets.
La distribució de les mutacions
Les mutacions són sempre fortuïtes, en el sentit que no van adreçades a cap objectiu morfològic ni fisiològic concret predeterminat, però tanmateix no es reparteixen aleatòriament en la fibra de DNA. Hi ha determinades zones, anomenades regions calentes, que són més propenses a incorporar mutacions que d’altres. El motiu obeeix a les característiques químiques dels mateixos nucleòtids, i també a determinats aspectes del procés de còpia. Per exemple, determinades combinacions de nucleòtids, com CG, es poden alterar químicament amb certa facilitat i esdevenir TG (substitució d’una C per una T).
Respecte al procés de còpia del DNA, els enzims que la dirigeixen poden "relliscar" amb una certa facilitat quan troben seqüències curtes que es repeteixen un cert nombre de vegades, com per exemple, TCTCTCTC..., la qual cosa provoca que afegeixin més nucleòtids del compte (una inserció) o bé que en copiïn menys dels que hi ha (una delectó).
A aquest efecte no aleatori de la distribució de mutacions, cal afegir també l’efecte de la pervivència de les mutacions, atès que quan hom compara seqüències de nucleòtids o d’aminoàcids no observa tots els canvis que s’han produït sinó només aquells que han perdurat en el temps, en virtut de la influència de la selecció natural sobre ells. Així, per exemple, les mutacions que afecten zones no crítiques de la fibra de DNA, és a dir, que no pertanyen a cap gen o a la seva zona reguladora, tindran més probabilitats de perdurar que els que afecten segments gènics implicats a codificar dominis proteics o d’RNA essencials, atès que amb molta més probabilitat aquestes darreres seran eliminades per la selecció natural si alteren la funció del gen en qüestió. De la mateixa manera, si la mutació afecta un nucleòtid que no fa canviar l’aminoàcid codificat, en virtut de la degeneració del codi genètic, molt probablement no es veurà sotmès a selecció, per la qual cosa podrà quedar fixat en aquell llinatge. En canvi, si la mutació en el DNA afecta l’aminoàcid codificat, llavors la funcionalitat de la proteïna pot quedar afectada i la selecció natural actuarà. Inevitablement això provoca un biaix en els canvis que hom observa cap a posicions en què no s’altera el missatge original, la qual cosa ha de ser tinguda en compte a l’hora de triar les molècules més informatives per a fer inferències filogenètiques.
Així, doncs, per exemple, si el darrer nucleòtid del codó GGG, que codifica l’aminoàcid glicina, és substituït per una A (GGA), continua codificant el mateix aminoàcid i, per tant, la seqüència de la proteïna no en quedarà afectada, ni tampoc la funcionalitat. En canvi, si en el mateix codó la primera G és substituïda per una A (AGG), llavors ja no codificarà l’aminoàcid glicina sinó que codificarà alanina, la qual cosa farà canviar la seqüència de la proteïna i probablement alterarà la funcionalitat global de la proteïna en qüestió. Per això s’aprofundirà una mica més a continuació en l’estructura del genoma i de les proteïnes.
L’estructura del genoma i la seva relació amb la pervivència de les mutacions
Com s’ha dit, les fibres de DNA són formades per una llarga seqüència de nucleòtids encadenats, en els quals es troben els gens. Així, els gens són segments concrets d’aquesta fibra de DNA que duen codificada una informació genètica determinada. Tanmateix, no tota la fibra de DNA és plena de gens, ni tots els gens duen codificat un mateix tipus general d’informació. Aquestes distincions són necessàries atès que la probabilitat de trobar variacions en un segment o altre de les cadenes de DNA depèn de la funció concreta de cada segment de DNA, com per exemple, formar part d’un gen o d’una zona intergènica.
L’estructura del genoma i els dominis de les proteïnes
Per començar, tots els gens tenen determinades seqüències molt conservades que els flanquegen, les quals contribueixen a la seva funcionalitat. Per exemple, els gens eucariotes que codifiquen proteïnes tenen sempre, davant de l’origen de transcripció, una seqüència formada bàsicament per adenines (A) i timines (T), anomenada caixa TATA; unes seqüències riques en citosines (C) i guanines (G), anomenades caixes GC; i sovint també una seqüència específica anomenada caixa CAAT (pels nucleòtids concrets que les formen).
Dominis funcionals de la proteïna PTK2 humana, implicada en l’adhesió entre cèl·lules (en tots els organismes pluricel·lulars). Cada domini és marcat en un color diferent. S’indiquen (en taronja) les proteïnes amb les quals interactua cada domini, i la funció global d’aquestes interaccions. Si es produeix un canvi que afecta un d’aquests dominis, probablement en quedarà afectada la funcionalitat. Però, si el canvi es produeix fora dels dominis, la probabilitat que s’alteri la funció serà molt menor, per la qual cosa tindrà més probabilitats de fixar-se i de poder ser observat.
Original de l’autor, a partir de fonts diverses.
Dins un gen, no totes les posicions nucleotídiques acumulen canvis amb igual probabilitat. El motiu és ben simple: per a realitzar les seves funcions, les proteïnes han d’interactuar amb altres molècules, incloses altres proteïnes. (Cal recordar que la seqüència aminoacídica de les proteïnes és conseqüència directa de la seqüència nucleotídica del gen corresponent.) Generalment, en qualsevol proteïna hom pot reconèixer diversos dominis, que són segments de la molècula la funció dels quals és interactuar de forma específica amb altres molècules, com per exemple, el centre catalític dels enzims, la zona que afavoreix una determinada reacció bioquímica.
En aquest context, els dominis d’una proteïna que interactuen específicament amb altres molècules acostumen a estar molt conservats, atès que pràcticament qualsevol canvi afecta llur funció, mentre que els segments d’enllaç poden acumular més canvis. A nivell del DNA, això implica que els nucleòtids implicats en la codificació de determinats dominis proteics (en virtut del codi genètic) tenen molt més restringida la possibilitat de canviar, mentre que els altres poden acumular un nombre més gran de canvis. És a dir que, segons el nivell taxonòmic que hom analitza, és més informatiu utilitzar un segment o un altre d’un determinat gen, per poder comparar un nombre adequat de canvis informatius.
D’altra banda, en els organismes eucariotes la informació continguda als gens no és contínua. Així, la majoria dels gens són formats per diverses seqüències que contenen informació per a sintetitzar la proteïna, anomenades exons, separades entre si per unes altres seqüències que no porten informació per a sintetitzar la proteïna corresponent, anomenades introns. Durant la formació de l’RNAm, els introns són eliminats com a part de processament d’aquesta molècula, per la qual cosa no són traduïts a proteïna. Això no exclou que alguns introns es puguin comportar, a vegades, com a exons, un fenomen genètic anomenat processament alternatiu de l’RNAm.
En general, qualsevol canvi que afecta un exó té més conseqüències fisiològiques que un que afecta un intró i, per tant, s’observen més canvis en els introns que en els exons. Tanmateix, la distribució d’introns dins un gen en diversos organismes o la seva utilització alternativa com a exó o com a intró també s’utilitza per a fer inferències filogenètiques, atès que les seves reorganitzacions tenen també un origen evolutiu, sovint restringit a llinatges concrets.
El DNA no codificant
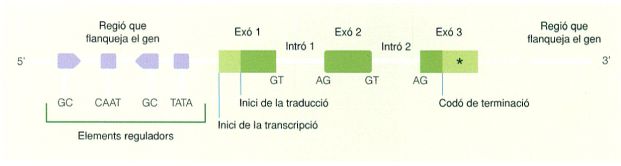
Estructura interna general d’un gen eucariota, estructurat en exons i introns, i flanquejat per elements reguladors. S’hi indica el lloc d’inici de la traducció i de la transcripció.
A partir de fonts diverses.
Hom distingeix quatre tipus de DNA no codificant: les zones reguladores, que controlen el funcionament de gens concrets i determinen on, en quin moment i en quina intensitat han de funcionar, i que per tant són importants per al funcionament genètic; les regions intergèniques, que serveixen d’enllaç entre gens per tal de mantenir la unitat de les fibres de DNA; els introns, que són seqüències no codificants que es troben intercalades dins els gens, i que són eliminades durant la formació de l’RNAm (només tenen introns els organismes eucariotes i alguns arqueus); i els anomenats pseudogens, que són restes de gens que han perdut la funcionalitat i per tant poden acumular canvis (mutacions) sense que interfereixin en el funcionament cel·lular i genètic. Generalment els pseudogens provenen de gens que s’han duplicat i en què una de les còpies, el pseudogèn, ha perdut la funcionalitat per haver acumulat un nombre significatiu de mutacions en llocs clau per al seu funcionament. Les conseqüències de les mutacions en aquestes zones són diverses, la qual cosa condiciona la seva pervivència i, en conseqüència, que puguin ser observades.
Els canvis en les seqüències
Per a visualitzar els diversos estats d’aquests caràcters moleculars cal conèixer llurs seqüències. Dur a terme els procediments per conèixer la seqüència d’aquestes molècules s’anomena seqüenciar. Es tracta d’un conjunt de processos que es fan mitjançant tècniques específiques, en bona part automatitzades, la descripció de les quals va més enllà dels objectius d’aquesta obra.
Nombre de substitucions entre dues seqüències de DNA
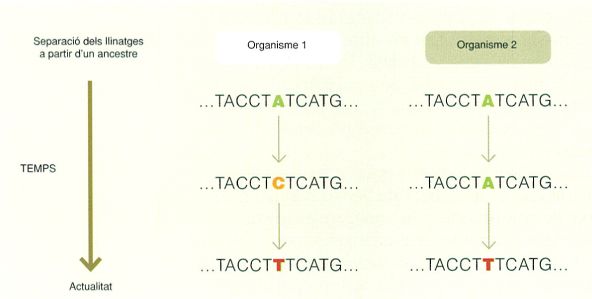
Els canvis que s’han produït des del moment de la separació de diferents llinatges a partir d’un organisme ancestral fins a l’actualitat no són tots visibles en els organismes que avui es poden analitzar. L’esquema mostra com en la seqüència de DNA de dos organismes que parteixen d’un mateix ancestre es poden produir diversos canvis que no són visibles en la seqüència analitzable en l’organisme actual. El nombre de canvis és sempre igual o inferior als que s’han produït realment, per la qual cosa cal emprar algoritmes correctors. Observant els dos organismes, avui no es pot saber que en l’organisme 1 es va produir un canvi de més que en el 2.
A partir de fonts diverses.
Quan dos llinatges divergeixen, les seves seqüències de DNA acumulen canvis per mutació, atès que en divergir la selecció natural actua sobre elles de manera diferent. En general, la variable que es fa servir més sovint en anàlisis filètiques és el nombre de canvis que s’han produït des de l’inici d’aquesta divergència. Quan el grau de divergència entre dues seqüències és petit, la probabilitat que en un mateix lloc s’hagi produït més d’un canvi consecutiu és molt petita, i el nombre de diferències que hom observa és molt similar a les que realment s’han produït. En canvi, si el grau de divergència és substancial, és possible que alguna mateixa posició nucleotídica o aminoacídica hagi canviat diverses vegades, però només s’observa el darrer canvi, i dels altres no en queda cap registre. En aquests casos, el nombre de canvis observats és inferior als que realment s’han produït.
Per exemple, si un nucleòtid determinat canvia de A a C en l’evolució d’un llinatge i després experimenta un nou canvi de C a T, l’observador només és conscient d’un canvi, de la A inicial a la T final, quan en realitat n’hi ha hagut dos. I si per casualitat en l’altre llinatge que s’analitza s’ha produït un canvi independent en el mateix nucleòtid de la A original a una T, llavors l’observació no revela cap diferència entre els organismes que es comparen, atès que les dues seqüències finals tenen una T en aquesta posició. Però en realitat s’han produït tres canvis, dos en una de les seqüències analitzades i un en l’altra.
Per a corregir aquestes distorsions, s’empren diversos algoritmes que permeten calcular estadísticament el nombre de substitucions. Aquests algoritmes tenen en compte també la regió genòmica que hom analitza, atès que aquest és uns dels factors que influeixen en la probabilitat que es produeixin més o menys canvis. Això facilita la comparació del grau de divergència entre dues parelles de seqüències analitzades.
Els rellotges moleculars
A mitjan dècada de 1960 es va proposar que la taxa de canvi (o d’evolució) d’una proteïna deter-minada devia ser aproximadament constant en el decurs del temps en tots els llinatges que la tinguessin, una hipòtesi que implicava l’existència d’una mena de rellotge molecular. Aquesta proposta va desvetllar l’interès per la utilització de macromolècules en els estudis evolutius i filogenètics, i va obrir definitivament la porta a la concepció molecular de la filogènesi actual. A més, si existís una mena de rellotge molecular reflectit en la taxa de canvi de les proteïnes (i per extensió, també del DNA, atès que conté la informació per a sintetitzar-les), hom podria utilitzar aquestes dades per a determinar quan van divergir dues espècies concretes i per a construir relacions filogenètiques.
Tanmateix, aquesta hipòtesi també va estimular diverses controvèrsies. Per exemple, des del punt de vista de la filogènia i l’evolucionisme clàssics, es va dir que no encaixava bé amb les dades morfològiques i fisiològiques, ja que si hom considera les formes fòssils, els tempos d’evolució no són constants, sinó sovint erràtics. També des del punt de vista de la filogènia molecular s’han posat objeccions, per exemple, pel fet que generalment la taxa de canvis observats s’accelera després de la duplicació d’un gen, ja que llavors una de les còpies queda alliberada de la pressió selectiva de la selecció natural i pot acumular molts més canvis. No obstant això, sovint s’utilitza aquesta hipòtesi per a estimar el temps de divergència entre llinatges d’organismes i per a reconstruir arbres filogenètics, tenint sempre present que es tracta d’una estimació. Per això, en la construcció d’arbres filogenètics amb dades moleculars, hom ha de considerar sempre la taxa de canvi.
Taxa de canvi en funció de la regió genòmica
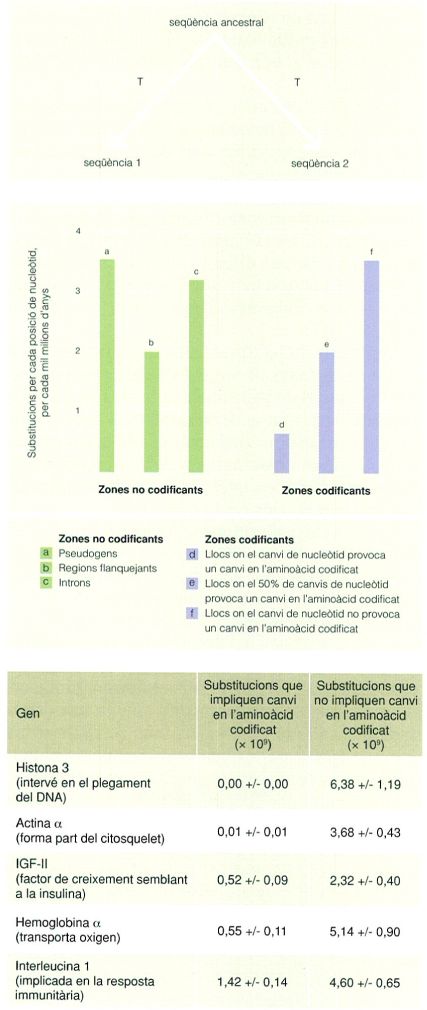
Taxa de canvi o de substitució de nucleòtids en una proteïna i temps de divergència. A dalt, esquema del temps de divergència de dues seqüències homòlogues a partir d’una seqüència comuna ancestral, de fa T anys. El temps de divergència permet calcular la taxa de substitució de nucleòtids. Al mig, taxa de substitució mitjana en diferents regions del genoma. A baix, taxa de substitució de nucleòtids en funció del gen i de si les substitucions afecten posicions que impliquen canvi d’aminoàcid o no, a partir de la degeneració del codi genètic. La taula mostra cinc exemples de gens molt conservats (historia), mitjanament conservats (actina) i relativament poc conservats (IGF-II, hemoglobina i interleucina 1), i també els intervals de variació sobre les taxes (error estàndard).
A partir de Graur i Li, 2000.
La taxa de substitució nucleotídica es defineix com el nombre de substitucions que ha experimentat cada lloc ocupat per un nucleòtid cada any, i es calcula dividint el nombre de substitucions detectades entre dues seqüències homòlogues pel doble del temps que fa que van divergir (és a dir, taxa de substitució = nombre de substitucions / 2 × temps de divergència). S’assumeix que aquest temps és el mateix que el temps de divergència entre les dues espècies d’on s’han obtingut les molècules homòlogues, inferit a partir de dades paleontològiques.
Aquesta taxa de canvi, però, no és constant, sinó que depèn de la regió de la molècula que s’analitzi, la qual cosa ha de ser curosament valorada quan hom utilitza aquestes dades per a fer inferències filogenètiques. Així, per exemple, s’ha vist que les zones de DNA on es detecta una taxa més alta de substitucions són els pseudogens, els introns i els segments que flanquegen els gens però que no en formen part (és a dir, les zones no codificants), ja que en no portar informació per a sintetitzar cap RNA ni per a regular la funció gènica, en general els canvis que s’hi produeixen no es veuen tan sotmesos als efectes de la selecció natural i es fixen amb molta més facilitat. També presenten una elevada taxa de substitució aquelles posicions dins el gen que, sigui quin sigui el nucleòtid que portin, en virtut de la degeneració del codi genètic no fan canviar l’aminoàcid codificat, per la qual cosa la proteïna final no es veu afectada. En canvi, les zones del genoma que presenten una menor taxa de substitucions són les que contenen informació per a sintetitzar proteïnes, i concretament aquelles en què un canvi de nucleòtid implica un canvi de l’aminoàcid codificat (i en conseqüència un canvi en la proteïna sintetitzada).
A més, la taxa de substitució també depèn de cada gen concret, en funció de la conservació de la seva funció en els diversos llinatges, i de cada regió no codificant específica. Així, per exemple, la taxa de substitució en els gens de les histones, les proteïnes encarregades d’empaquetar el DNA, que són presents en tots els eucariotes i en els quals realitzen exactament la mateixa funció, és pràcticament nul·la. Això no vol dir que no es produeixin mutacions en aquests gens, sinó que les mutacions que es produeixen són eliminades per la selecció natural i, per tant, no queden fixades i no s’observen.
Altres gens, en canvi, presenten taxes de substitució baixes però detectables, com els de l’actina, una proteïna que forma part del citoesquelet de totes les cèl·lules eucariotes. Aquesta proteïna realitza la mateixa funció en totes les cèl·lules, però ateses les diferències morfològiques i fisiològiques que presenten les cèl·lules dels diversos organismes, toleren petites diferències en la proteïna sintetitzada per aquest gen; diferències que al seu torn són a la base d’algunes diferències de morfologia cel·lular.
Finalment, hi ha altres gens que presenten, en comparació, taxes de substitució molt més elevades, com ara el de la insulina, la hemoglobina o les interleucines, i en general qualsevol gen la funció del qual estigui implicada en les diferències morfològiques i fisiològiques dels diferents llinatges; unes diferències que, precisament, es basen en aquestes diferències genètiques.
A més, la taxa de substitució tampoc no és igual en tota la seqüència codificant d’un gen. Hi ha segments que presenten una taxa de substitució molt més elevada que d’altres, i segments en què aquesta taxa és pràcticament nul·la, en funció de la conservació fisiològica del domini proteic que codifiquen.
En definitiva, en funció dels organismes o llinatges que s’han de comparar per a establir una determinada filogènia molecular, hom ha de triar les dades moleculars que li resultin més informatives, és a dir, aquelles que presentin un nombre adequat de variacions per a fer les comparacions pertinents; ni massa per a no introduir heterogeneïtats ni poques per a no limitar el nombre de caràcters que cal comparar.
Taxa de substitució en el DNA mitocondrial i dels cloroplasts
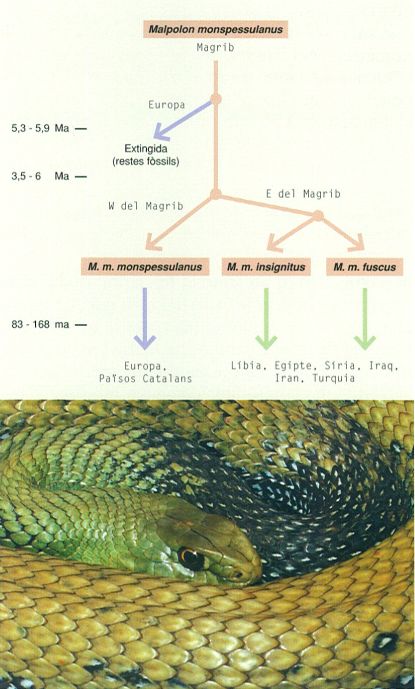
Separació filogenètica i dispersió geogràfica de diverses subespècies de serp verda (Malpolon monspessulanus), originària del Magrib, a partir d’inferències filètiques fetes amb dades moleculars de dos segments concrets del seu DNAmt. Després d’una primera dispersió per Europa, es va produir una divergència de subespècies al mateix Magrib. Un grup es va instal·lar cap a l’oest, i d’allà va passar a efectuar la segona dispersió per Europa (present ara als Països Catalans); i un altre grup, distribuït per l’est, va donar lloc a dues altres subespècies, que van colonitzar el sud-est asiàtic. La línia del temps no és a escala. Ma milions d’anys; ma milers d’anys.
A partir de Carranza, Arnold i Pleguezuelos, 2006 / Vicent Sancho.
En comparació amb els genomes nuclears, el DNA dels mitocondris i dels cloroplasts és molt més senzill d’investigar experimentalment, ja que és petit. A més, en general la taxa de substitucions nucleotídiques és superior en el DNA d’aquests orgànuls cel·lulars en comparació amb el genoma nuclear, i presenta moltes més reorganitzacions gèniques, és a dir, canvis en l’ordre i la posició relatius dels gens entre llinatges diferents. Per aquests motius, l’anàlisi del genoma d’aquests orgànuls constitueix un element clau en la construcció de l’arbre de la vida.
Per exemple, l’anàlisi de la variació detectada en dos fragments de DNAmt, un de 300 nucleòtids corresponent al gen del citocrom b i un altre de 395 a 515 nucleòtids corresponent al gen de l’RNAr 12S d’aquest orgànul, ha permès establir l’origen africà i les relacions filogenètiques de diverses subespècies de serp verda (Malpolon monspessulanus), així com la història concreta de la seva distribució per Europa i el Pròxim Orient a partir d’un origen nord-africà. La serp verda, present en alguns indrets dels Països Catalans, és una colobra de coloració uniforme i tons verdosos, característica d’ambients secs i pedregosos, que pot atènyer els 2 m de longitud. Segons aquestes dades moleculars, M. monspessulanus és originària del Magrib, des d’on va colonitzar el sud d’Europa fa uns 5,3-5,9 milions d’anys, aprofitant el pont de terra que unia els dos extrems de l’estret de Gibraltar. Tanmateix, els exemplars d’aquesta primera colonització, que ha deixat restes fòssils, es van extingir. Mentrestant, fa uns 3,5-6 milions d’anys, M. monspessulanus del Magrib es va separar en tres subespècies: M. m. monspessulanus a l’oest, i M. m. insignitus i M. m. fuscus a l’est. Aquestes dues darreres subespècies han anat colonitzant progressivament i de forma ininterrompuda el sud-est asiàtic (Líbia, Egipte, Síria, Iraq, Iran i Turquia), mentre que M. m. monspessulanus va tornar a colonitzar Europa fa entre 83.000 i 168.000 anys.
L’alineació de seqüències
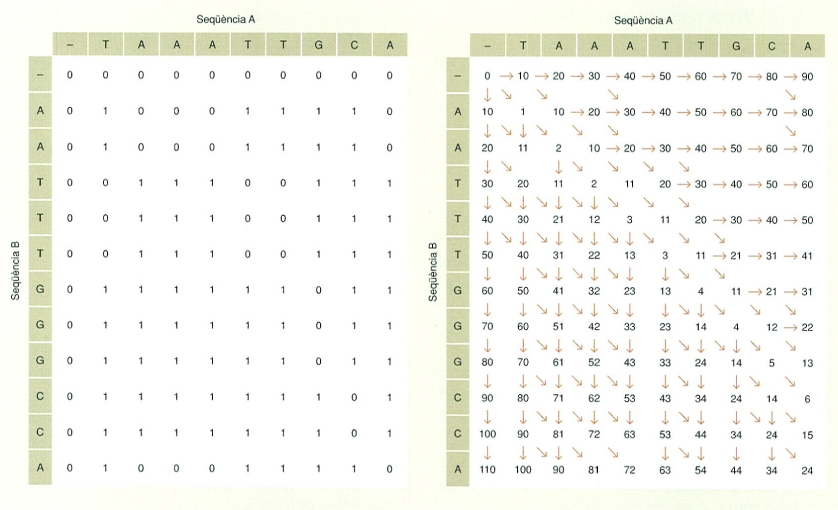
Matriu inicial d’aparellament de seqüències de nucleòtids, on 0 indica que no hi ha diferència, i 1 que hi ha una substitució. A la dreta, matriu completa de distàncies. Basant-se en matrius com aquestes, fetes amb algoritmes determinats, els programes d’alineació de seqüències estableixen les alineacions òptimes entre les diverses seqüències que es comparen.
A partir de Phillips i altres, 2000.
Comparar seqüències homòlogues implica identificar les diferències de nucleòtids o d’aminoàcids que hi ha entre elles, degudes tant a canvis puntuals de nucleòtids o aminoàcids com a delecions i insercions de més extensió, que han ocorregut en qualsevol dels llinatges que s’analitzen des de la seva divergència a partir d’un ancessor comú. Aquest procés es realitza mitjançant l’alineació de les seqüències que s’han de comparar. L’alineació de seqüències és l’ordenació de les seqüències de DNA, RNA o proteïnes que hom vol comparar per identificar regions de similitud que puguin ser conseqüència de les seves relacions funcionals, estructurals o evolutives. Normalment es representen en fileres paral·leles aparellades que permeten observar i quantificar objectivament el grau de similitud. Tanmateix, aquestes alineacions no indiquen si una seqüència determinada ha experimentat una deleció o si és l’altra la que ha tingut una inserció, o bé quina és la que ha tingut una determinada substitució, però permeten detectar i quantificar de forma objectiva els diversos estats d’aquests caràcters.
Alineació de seqüències de nucleòtids d’un fragment del gen de l’RNAr 18S de diverses espècies de mol·luscs. La major part de posicions són compartides, però tanmateix hi ha estats diferents en alguns caràcters. (*) indica els estats que són diferents en algunes de les seqüències. (−) omple els buits que deixen les insercions o les delecions. A partir d’aquestes diferències, els algoritmes estableixen les corresponents matrius de similitud.
A partir de Giribet i altres, 2006.
Si entre les seqüències que es comparen hi ha poques molècules o pocs segments que no s’aparellen bé, es pot obtenir una alineació global raonable pràcticament a simple vista. Tanmateix, si hi ha un nombre significatiu de diferències, cal utilitzar algoritmes matemàtics i procediments estadístics per a predir quines de les molècules de la seqüència són les que realment s’aparellen bé –i per tant són caràcters conservats– i quines han divergit –o presenten estats diferents d’aquell caràcter-. El criteri general en l’alineació de seqüències és reduir el nombre de molècules no aparellades al mínim.
Normalment, per a fer les alineacions, hom utilitza eines bioinformàtiques, les quals permeten fer dos tipus generals d’alineacions, les anomenades alineacions globals i les locals. L’alineació global abasta tota la longitud de les molècules analitzades, mentre que l’alineació local identifica regions de similitud dins de seqüències més llargues i les compara, amb la qual cosa evita segments molts divergents de difícil alineació (cal recordar que la taxa de canvi depèn de cada regió gènica concreta i que pot ser molt variable). En general, les alineacions locals són preferibles a les globals.
Es poden utilitzar diversos mètodes matemàtics per a generar les alineacions, generalment matrius de distàncies en què es representen les substitucions i els camins estadístics més probables que les justifiquen, els quals es basen en els seus propis algoritmes i que resulten més o menys informatius en funció de les seqüències concretes que hom està analitzant. Quan es comparen més de dues seqüències, els procediments matemàtics inclouen totes les alineacions possibles entre totes les seqüències considerades de dues en dues, a la recerca de l’alineació que atorga una màxima similitud al conjunt de totes elles.
A més, si s’utilitzen algoritmes, hom pot establir numèricament l’índex de similitud entre dues seqüències concretes en funció de cada possible alineació, la qual cosa permet triar com a òptima l’alineació que mostra un índex de similitud màxim (és a dir, aquella alineació en què hi ha més nucleòtids o aminoàcids aparellats). També s’utilitzen altres algoritmes que calculen l’índex de distància entre dues seqüències en funció de cada possible alineació. En aquest cas, hom tria com a òptima l’alineació que mostra un índex de distància mínim (és a dir, aquella alineació en què hi ha menys nucleòtids o aminoàcids no aparellats).
L’homologia molecular
Com en el cas dels caràcters morfològics, per a poder inferir relacions filogenètiques utilitzant dades moleculars, cal comparar seqüències nucleotídiques o aminoacídiques que siguin homòlogues. Dos o més segments d’àcids nucleics o dues o més proteïnes són homòlegs si presenten similituds quant a la seva seqüència, és a dir, si presenten intervals de seqüència que són parcialment iguals entre ells, amb la condició indispensable que procedeixin d’una seqüència ancestral comuna a partir de la qual han divergit per evolució. L’homologia molecular s’estableix mitjançant la comparació bioinformàtica de les seqüències analitzades, és a dir, a partir de l’alineació de les corresponents seqüències d’aminoàcids o nucleòtids per a quantificar-ne de manera objectiva la similitud.
No obstant això, en algunes ocasions la semblança entre seqüències pot haver sorgit de manera independent (per exemple, per evolució convergent), sense que comparteixin una seqüència ancestral comuna, com es va esmentar també en el cas dels caràcters morfològics. Aquest fet es pot donar especialment si hom analitza seqüències massa curtes, les quals poden haver esdevingut semblants simplement per atzar, per exemple, perquè la selecció natural les ha afavorit de forma independent pel fet de realitzar una funció també semblant, com determinar un domini d’unió a una determinada proteïna.
Així, moltes proteïnes la funció de les quals és unir-se al DNA per a regular la funció gènica comparteixen un domini comú, una seqüència de nucleòtids anomenada domini homeòtic que té l’estructura tridimensional adequada per a unir-se al DNA. En aquest cas concret, moltes de les proteïnes que contenen aquest domini no són globalment homòlogues, però el domini homeòtic específic prové, molt probablement, d’una única seqüència ancestral amb aquesta funció, la qual ha estat aprofitada i reciclada en múltiples ocasions. La filogènia molecular es nodreix de l’homologia molecular, de les semblances i les diferències en la seqüència entre molècules homòlogues.
L’ortologia i la paralogia
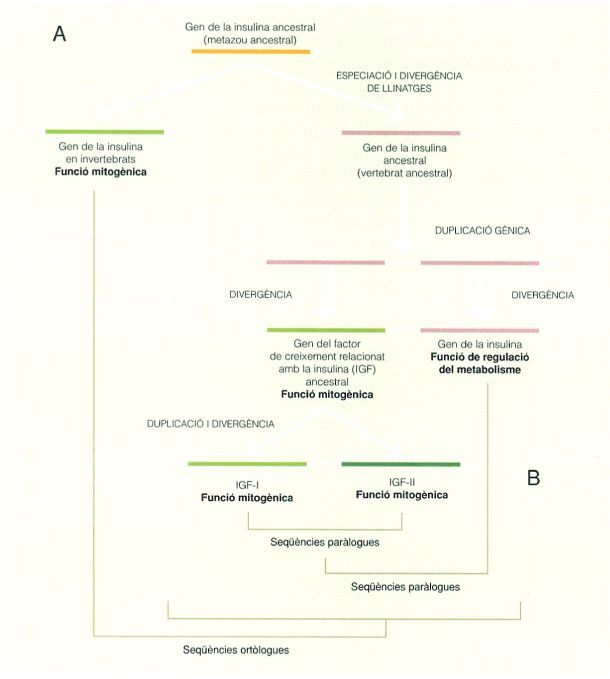
Ortologia i paralogia del gen de la insulina i d’altres relacionats. El gen de la insulina té l’origen en un metazou ancestral (A) i únic, del qual va derivar per processos iteratius d’especiació i divergència de llinatges. En aquest cas d’ortologia, la funció es conserva parcialment. B indica paralogia del gen de la insulina i del factor de creixement relacionat amb la insulina en vertebrats, els quals van adquirir funcions diverses a partir del gen ancestral, i paralogia entre els dos gens del factor de creixement relacionat amb la insulina (IGF-I i IGF-II) dels vertebrats.
Original de l’autor, a partir de fonts diverses.
Quan es comparen seqüències d’àcids nucleics o de proteïnes per a fer inferències taxonòmiques o filogenètiques, hom ha de distingir entre diversos tipus d’homologia molecular, atès que les implicacions i la utilitat són diferents: les seqüències ortòlogues i les seqüències paràlogues.
Es diu que dues seqüències homòlogues són seqüències ortòlogues (o que presenten ortologia) si ambdues procedeixen d’una seqüència ancestral a partir de la qual han divergit en paral·lel al procés d’especiació que ha afectat llur llinatge. Dit d’una altra manera: quan dues poblacions d’una mateixa espècie divergeixen evolutivament fins a esdevenir espècies diferents, les còpies divergents d’una mateixa seqüència ancestral present en aquestes espècies són ortòlogues entre si, atès que es van originar a partir d’un avantpassat comú. Els gens ortòlegs acostumen a conservar la mateixa funció, però tanmateix hi ha algunes excepcions. L’evidència més consistent que dos gens homòlegs són ortòlegs prové de les anàlisis filogenètiques del llinatge d’aquelles seqüències. Per exemple, l’anàlisi molecular i filogenètica del gen de la insulina, una hormona proteica, ha demostrat que el seu origen tant en vertebrats com en invertebrats es remunta a un únic gen ancestral, malgrat que la conservació de la funció d’aquesta hormona entre aquests dos grups d’organismes només sigui parcial.
Les seqüències ortòlogues proporcionen una informació molt útil per als estudis taxonòmics i filogenètics, atès que el seu patró de divergència permet traçar les relacions filogenètiques entre organismes, com s’ha dit per al cas de l’homologia primària. En general, una major semblança quant a gens ortòlegs implica una major proximitat de parentiu.
En canvi, es diu que dues seqüències són paràlogues (o que presenten paralogia) si han divergit dins un mateix llinatge d’organismes després d’haver experimentat un procés de duplicació gènica en l’organisme ancestral d’aquell llinatge. Dit d’una altra manera: si un gen d’un organisme es duplica, les dues còpies esdevenen paràlogues. Generalment, els gens paràlegs tenen la mateixa funció o bé una funció molt relacionada, però també hi ha algunes excepcions, atès que la manca de pressió selectiva sobre una de les còpies permet que muti més lliurement i, en conseqüència, que pugui adoptar noves funcions (la manca de pressió selectiva sobre una de les còpies és deguda al fet que l’altra manté la funció original). Per exemple, retornant al cas de la insulina, la seva anàlisi molecular i filogenètica ha demostrat que, en els vertebrats, va patir com a mínim una duplicació, la qual cosa va generar dos gens paràlegs: el de la insulina pròpiament dit, que va adquirir la funció de regulació del metabolisme, i l’anomenat factor de creixement relacionat amb la insulina (o IGF, de l’anglès Insulin Growth Factor), que va mantenir la funció mitogènica ancestral. Tanmateix, en el llinatge dels vertebrats, l’IGF es va tornar a duplicar amb posterioritat i va generar dos nous gens paràlegs, anomenats IGF-I i IGF-II, amb funcions similars.
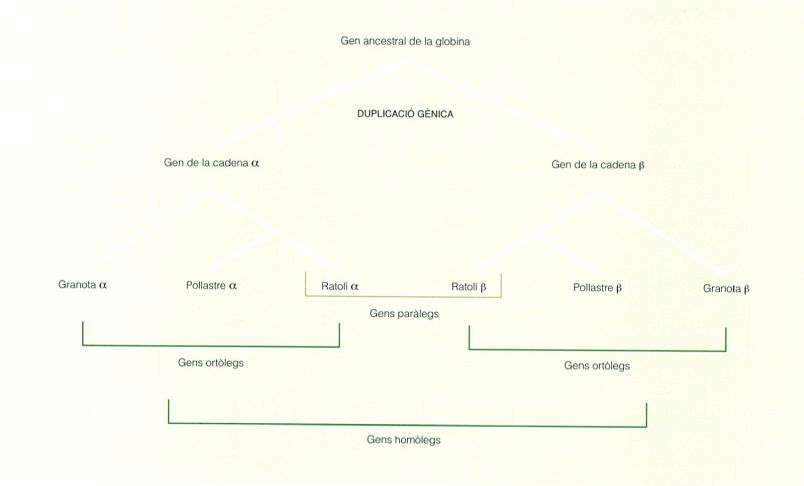
Esquema simplificat de la relació d’ortologia i paralogia entre diversos gens de la família de les globines dels vertebrats. Els gens α i β són paràlegs entre ells, mentre que els diversos gens α presenten ortologia entre ells, igual que els gens β.
Original de l’autor, a partir de fonts diverses.
Tanmateix, no sempre és tan senzill establir les relacions d’ortologia i paralogia. Per exemple, el gen de l’hemoglobina dels humans és ortòleg respecte al gen de l’hemoglobina dels ximpanzés, però al mateix temps és paràleg del gen de la mioglobina tant dels mateixos humans com del dels ximpanzés, atès que provenen de processos de divergència paral·lela a la dels llinatges o de duplicació interna en un llinatge i posterior divergència, respectivament. L’hemoglobina és la proteïna encarregada de transportar l’oxigen per la sang, mentre que la mioglobina emmagatzema oxigen de reserva als músculs. A més, el gen de l’hemoglobina ha experimentat diverses duplicacions, la qual cosa ha generat també diverses hemoglobines amb funcions concretes que també són paràlogues entre elles.
La comparació de gens paràlegs quant a nombre i seqüència entre diversos organismes o llinatges (no pas en un mateix organisme) també proporciona informació útil per als estudis taxonòmics i filogenètics. En general, una major semblança de nombre i seqüència entre gens paràlegs implica una major proximitat de parentiu entre els llinatges analitzats.
L’homologia estructural i l’homologia de processos

Homologia dels ulls de vertebrats i d’insectes a partir de la comparació de les seqüències de nucleòtids d’un segment de DNA corresponent als gens Pax-6 del ratolí i eyeless de Drosophila. Els nucleòtids idèntics estan marcats en verd. La taxa de similitud en la seqüència de nucleòtids d’aquest segment és del 76,66%, i pel que fa als aminoàcids, del 100%. Això indica que el segment de proteïna corresponent a aquesta seqüència és idèntica en insectes i vertebrats. Tanmateix, la morfologia dels ulls del ratolí i la mosca és ben diferent.
A partir de fonts diverses.
El descobriment de sorprenents homologies moleculars en el camp de la biologia evolutiva del desenvolupament ha posat en qüestió la distinció clàssica entre homologia i analogia. Un cas paradigmàtic és el de la relació entre els ulls i els cors dels vertebrats i dels insectes. Els ulls dels vertebrats i dels insectes han estat considerats, tradicionalment, com a estructures funcionalment anàlogues, ateses les seves diferències morfològiques i d’origen embrionari. Al final de la dècada de 1990, però, es va demostrar que tant la formació dels ulls dels insectes com la dels vertebrats, i també la dels cefalòpodes, depèn de l’expressió de gens ortòlegs, que provenen d’un ancestre comú, anomenats eyeless en insectes i Pax-6 en vertebrats.
A partir d’aquest fet, s’ha deduït que els ulls dels vertebrats, dels insectes i dels cefalòpodes provenen molt probablement d’una cèl·lula bàsica fotoreceptora, que ja des de l’inici era regulada pel mateix gen. Aquest gen hauria mantingut la funció de dirigir la formació d’òrgans fotoreceptors, amb independència de la morfologia final d’aquests òrgans, que depèn d’altres gens específics.
De la mateixa manera, des d’una perspectiva morfològica i anatòmica, el cor dels vertebrats i dels insectes només té en comú el fet de servir per a bombar fluids, motiu pel qual clàssicament han estat considerats un altre exemple d’analogia. No obstant això, s’ha demostrat que ambdós tipus de cor depenen també de l’expressió de gens ortòlegs, anomenats Csx en vertebrats i tinman en insectes.
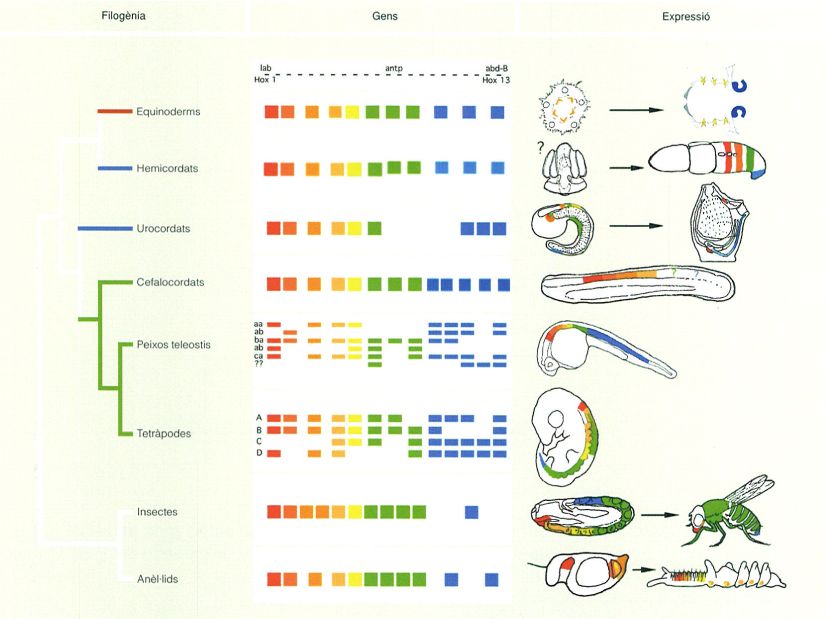
Esquema simplificat de la família gènica Hox en diversos grups de metazous, en què s’indica la relació filogenètica d’aquest grup, el nombre de gens i de famílies paràlogues dins de cada grup i l’expressió d’aquests gens respecte de l’eix anteroposterior, seguint un codi de colors. Cada caixa acolorida representa un gen. Observeu que els peixos teleostis i els tetràpodes tenen diverses famílies paràlogues amb relació a aquests gens, mentre que els altres metazous presenten una única família, i que el nombre de gens per família també és superior en aquests dos llinatges. La presència de diverses famílies en els vertebrats és deguda a dues duplicacions successives a l’inici del seu llinatge. La correspondència de colors entre els diferents llinatges indica els gens que són ortòlegs.
Adaptat a partir de Swalla, 2006.
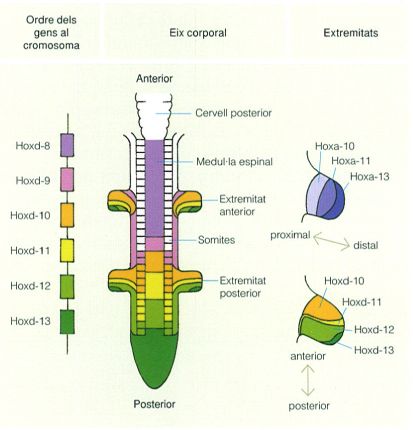
Expressió d’alguns membres de la família gènica Hox en la determinació del patró anteroposterior i de les extremitats de vertebrats. Alguns dels gens d’aquesta família s’expressen en tots dos eixos corporals.
A partir d’Alberts i altres, 2002.
D’altra banda, també s’ha demostrat que un nombre significatiu de gens homòlegs poden expressar-se en estructures que no ho són. Un cas paradigmàtic és el dels gens Hox. Els gens Hox constitueixen una família gènica que té la funció de coordinar i establir el patró d’estructures durant el desenvolupament embrionari. Aquesta família presenta gens ortòlegs en tots els metazous, i gens paràlegs dins de cada organisme concret. Els organismes morfològicament més simples tenen menys paràlegs que els complexos. Els gens Hox dels insectes determinen llur patró anteroposterior d’estructures, però no el de les seves extremitats, mentre que en els vertebrats determinen ambdós patrons, a banda d’expressar-se en moltes altres estructures anatòmiques en determinats moments del desenvolupament embrionari, com el sistema nerviós central.
Tanmateix, durant el desenvolupament de les extremitats tant d’insectes com de vertebrats intervé un altre gen que és ortòleg en aquests dos grups d’organismes, anomenat distal-less en Drosophila, el qual també intervé en el desenvolupament dels parapodis dels poliquets, dels lobopodis, dels onicòfors, de les butllofes dels ascidis i de l’aparell ambulacral dels equinoderms, la qual cosa situa aquestes estructures anàlogues dins el camp de 1’homología molecular.
Això implica que hom hagi de definir en cada cas a quin nivell d’homologia es refereix. Els ulls dels vertebrats i dels cefalòpodes, per exemple, serien anàlegs quant a estructures fotoreceptores complexes i organitzades, però els gens que regulen llur formació embrionària serien ortòlegs (i per tant homòlegs). Per aquest motiu, per a designar patrons d’expressió gènica homòlegs, hom parla d’homologia de processos. I per a les homologies clàssiques, hom empra l’expressió homologia estructural.
Els caràcters primitius i els caràcters derivats
Finalment, tenint en compte totes les consideracions fetes quant a homologia dels caràcters, hom ha de distingir si l’estat d’un caràcter determinat és ancestral, primitiu o derivat, atès que les conseqüències a l’hora de fer inferències filogenètiques i taxonòmiques són diferents.

Els halteris dels dípters (a l'esquerra, típula) provenen d’una modificació del patró bàsic d’ales dels insectes, concretament del parell d’ales posteriors. Constitueixen una sinapomorfia (apomorfia compartida per dos o més tàxons) dels dípters, atès que no es troben en cap altre grup d’insectes (a la dreta, ales de papallona).
Marta Goula.
Si un caràcter, ja sigui morfològic o molecular, presenta el mateix estat o la mateixa seqüència bàsica que l’ancestre comú, llavors es considera un caràcter primitiu, i en terminologia filogenètica s’anomena caràcter plesiomòrfic o plesiomorfia. Així, es diu que una forma és plesiomòrfica si representa el caràcter ancestral, és a dir, el caràcter homòleg primitiu. Les plesiomorfies compartides entre diversos grups d’organismes s’anomenen simplesiomorfies, i pel fet de ser presents en tota una sèrie de grups no permeten discriminar els seus clades. Per exemple, la presència de cinc dits en les extremitats anteriors dels rosegadors i dels primats és una simplesiomorfia, atès que aquest caràcter es va originar molt abans de la separació filogenètica d’aquests dos grups respecte de la resta de vertebrats en l’evolució dels tetràpodes, i també és present en altres grups, com els llangardaixos. És a dir, la presència de cinc dits en les extremitats anteriors no pot ser utilitzada com a caràcter d’exclusió d’altres grups respecte als rosegadors i els primats. Per aquest mateix motiu, un caràcter simplesiomòrfic no és una prova que diversos tàxons que el continguin estiguin directament emparentats.
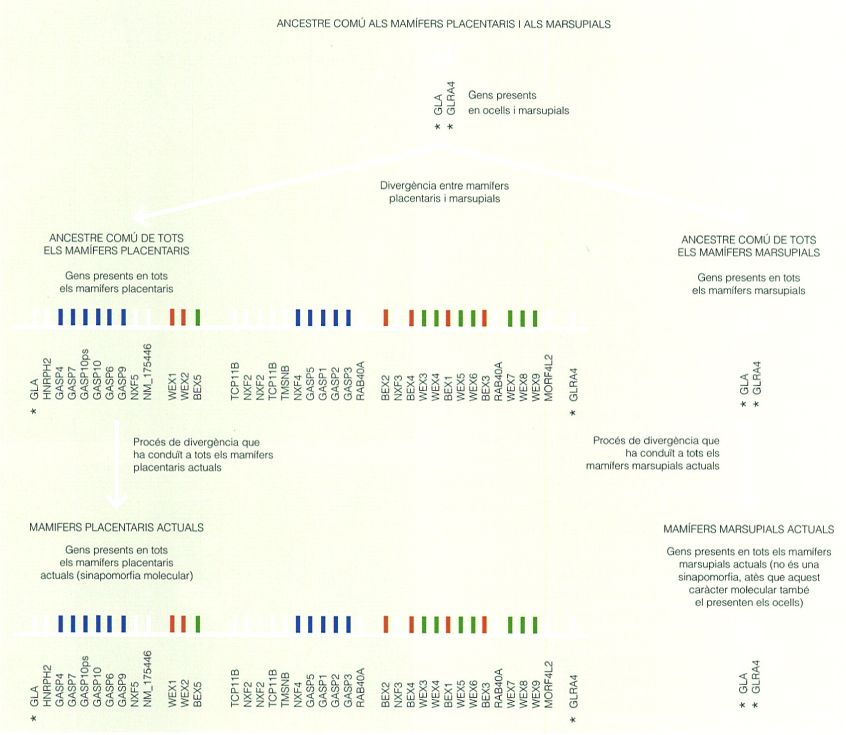
Sinapomorfia molecular dels mamífers placentaris quant a la regió de DNA que comprèn les famílies gèniques BEX, WEX i GASP. (*) indica els gens de l’extrem d’aquesta regió que són presents en l’ancestre comú als mamífers marsupials i als mamífers placentaris i que flanquegen aquesta sinapomorfia.
Adaptat a partir de Winter i Ponting, 2005.
En canvi, els caràcters derivats que distingeixen una espècie ancestral i tots els seus descendents, i que, per tant, no són compartits per cap altre tàxon, s’anomenen caràcters apomòrfics o apomorfies. Els caràcters apomòrfics representen novetats evolutives associades a llinatges concrets i són una prova suficient de la consistència del parentiu del grup que els manifesta. Per exemple, la presència de plomes és exclusiva dels ocells, per la qual cosa aquest caràcter es pot utilitzar per a definir i agrupar de forma consistent tots els membres de la classe de les aus. També hi ha apomorfies que es troben restringides a una sola espècie. Llavors s’anomenen autoapomorfies, i es poden emprar per a definir l’espècie en qüestió. Les autoapomorfies per si mateixes no proporcionen informació sobre les relacions filogenètiques d’aquella espècie, però no obstant això en poden indicar el grau de divergència respecte als parents més propers. Finalment, una apomorfia que sigui compartida per dos o més tàxons s’anomena sinapomorfia. Els caràcters sinapomòrfics defineixen de forma estricta els grups monofilètics, és a dir, aquells grups que inclouen un ancessor comú i absolutament tots els seus descendents. En el proper apartat es parlarà a bastament de grups monofilètics, atès que són la base dels sistemes cladístics de classificació utilitzats en filogènia. Així, doncs, una sinapomorfia és un caràcter compartit per dos o més tàxons i per llur ancessor comú, però que no és present en l’ancessor d’aquest. És, per tant, un caràcter homòleg derivat compartit per un grup d’organismes. Un exemple de sinapomorfia són els halteris dels dípters. Els halteris són unes estructures en forma de mall que provenen d’ales modificades i que actuen de balancí durant el vol dels insectes de l’ordre dels dípters. No hi ha cap altre grup d’insectes que tingui aquestes estructures, per la qual cosa són un caràcter homòleg compartit per tots els dípters actuals i, en conseqüència, per l’ancessor comú d’aquest ordre, però que no tenia llur ancessor respectiu, ja que que no són presents en cap altre grup d’insectes.
El mateix concepte es pot aplicar a caràcters moleculars. Per exemple, hi ha una regió concreta al genoma dels mamífers placentaris que conté diversos gens de les famílies gèniques BEX, WEX i GASP, que estan implicades en diverses funcions cel·lulars, algunes de les quals d’àmbit cerebral. Aquesta regió del DNA, que en humans es localitza al cromosoma X, no es troba ni en ocells ni en mamífers marsupials, per la qual cosa representa una sinapomorfia molecular dels mamífers placentaris. És a dir, que prové de l’ancestre comú a tots els mamífers placentaris, però que no es troba a l’ancessor comú que comparteixen aquest ancestre i el dels marsupials. Tanmateix, l’origen filètic dels gens GASP es remunta a l’origen dels vertebrats, però els gens BEX i WEX també són exclusius dels mamífers placentaris.

Fenestra anterorbital (1) de dinosaure (A), pterosaure (B), cocodril (C) i ocell (D). La seva presència en dinosaures i pterosaures és plesiomòrfica, ja que també es troba en els avantpassats dels cocodrils (en els cocodrils actuals, la fenestra està tapada). Tanmateix, és sinapomòrfica si es considera el grup format pels cocodrils, els dinosaures i els pterosaures.
A J.M. Luijt; C Mark Pellegrini; D Didier Descouens / B Kyle Mc-Quilkin i Ryan Ridgely;.
Així mateix, cal tenir present que una apomorfia pot ser considerada sinapomorfia o alternativament plesiomorfia en funció del nivell taxonòmic que hom estigui resolent. Per exemple, la presència de fenestra antorbital en dinosaures i pterosaures és plesiomòrfica, ja que també es troba en grups més inclusius i no és un caràcter que permeti separar els pterosaures i els dinosaures d’altres animals, com els cocodrils. Però si es considera el grup format pels cocodrils, els dinosaures i els pterosaures, llavors el mateix caràcter esdevé exclusiu per a aquest grup i esdevé una sinapomorfia.
Un altre exemple molt clar que la distinció entre sinapomorfia i plesiomorfia no és absoluta sinó que depèn del nivell taxonòmic en què hom se centri és el de la columna vertebral. Així, la columna vertebral és un caràcter sinapomòrfic que distingeix els vertebrats de la resta de metazous, atès que és exclusiu d’aquest grup i compartit per tots els seus membres, però al mateix temps és plesiomòrfic si hom el considera quant a la presència en els ocells respecte a tota la resta de vertebrats, atès que aquests també en presenten.
L’homoplàsia
A vegades la similitud entre caràcters en organismes de diferents tàxons pot ser deguda també a una convergència independent, el que en termes evolutius s’anomena convergència evolutiva o alternativament evolució paral·lela. La convergència evolutiva és el fenomen biològic pel qual diferents organismes tendeixen, sota pressions ambientals selectives equivalents, a desenvolupar en la seva evolució, a partir d’estructures preexistents, característiques morfològiques, fisiològiques o etològiques similars a partir de condicions ancestrals diferents, les quals esdevenen estructures anàlogues. Tanmateix si aquesta convergència s’ha produït a partir de condicions ancestrals similars es parla d’evolució paral·lela. En aquest context, un caràcter present en dos o més tàxons però que no es trobi en l’ancessor comú, és a dir, que hagi sorgit de forma independent, s’anomena caràcter homoplàsic o homoplàsia.

Procés de pèrdua secundària de les extremitats en les serps, com a exemple d’homoplàsia. Es mostren imatges d’organismes actuals en representació dels ancestres d’aquests llinatges.
Josep L. Ferrer.
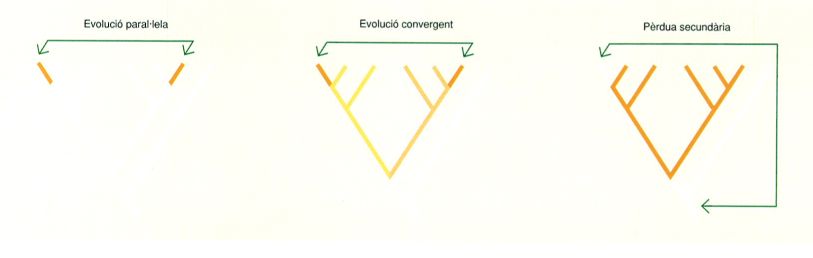
Diferents formes d’homoplàsia: evolució paral·lela (evolució independent de la mateixa característica, des d’una mateixa condició ancestral), convergent (des d’una condició ancestral diferent) i pèrdua secundària (retorn a la condició ancestral). En groc, divergència progressiva dels caràcters.
Original de l’autor.
Un exemple d’homoplàsia és l’homeotèrmia en mamífers i ocells, és a dir, la capacitat de mantenir la temperatura del cos constant amb independència de la temperatura externa. L’homeotèrmia és un caràcter derivat respecte a l’estat ancestral, la pecilotèrmia (en què la temperatura interna depèn completament de la del medi ambient), que ha evolucionat de forma independent en els avantpassats d’aquests dos grups, atès que ha es coneixen avantpassats comuns que la manifestessin.
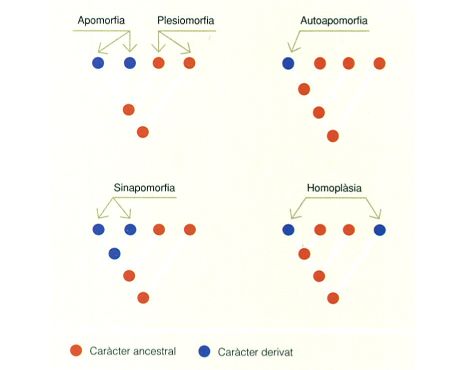
Arbres filogenètics esquematitzats que il·lustren els conceptes emprats per a descriure els diversos patrons dels caràcters ancestrals i derivats: apomorfia, plesiomorfia, autoapomorfia, sinapomorfia i homoplàsia.
Original de l’autor, a partir de fonts diverses.
Finalment, també és possible la presència d’homoplàsia deguda a la pèrdua d’una característica derivada, fet que provoca que, en aparença, un determinat llinatge retorni a una determinada condició ancestral, un fenomen evolutiu que es coneix com a pèrdua secundària. Un exemple d’homoplàsia per pèrdua secundària és la pèrdua de les extremitats en les serps i en alguns amfibis, com les cecílies pròpies de les regions tropicals humides —les anàlisis filogenètiques situen les cecílies com el grup germà dels amfibis, amb els quals constitueixen un clade de grau superior conegut com a lissamfibis—. Els caràcters homoplàsics no poden ser emprats per a fer inferències taxonòmiques i filogenètiques.