
La percepció del concepte de comunitat vegetal és haguda intuïtivament per la saviesa popular, sobretot quan els agrupaments vegetals són fisiognòmicament distints: en aquest cas de la vall de Boí, tothom diferencia l’avetosa (bosc), l’avellanosa (matoll alt) o el prat dallador (prat).
J. Nuet i Badia
La distribució de les plantes sobre la superfície de la Terra no obeeix a un atzar capriciós. Tampoc, per a una contrada determinada, llur distribució sobre un relleu concret: el cim, el raiguer de la muntanya, la cinglera rocallosa, el fons suau de la vall. Una rigorosa estratègia competitiva ha afaiçonat, al llarg dels segles, l’harmoniós "patch work" del mantell vegetal, la rigorosa estratègia competitiva que, implacable, ha fet evolucionar el món de la vida des del començament de les coses. La intervenció dels animals, ells també fills del mateix procés evolutiu, ha introduït tensions en aquest teixit de vida vegetal, però no pas distorsions dissolvents: no hauria estat esperable de dues concepcions vitals nascudes de la mateixa lògica, la de la vida. Però amb l’home, el més singular representant del món animal —tant, que li costa o es nega a admetre l’evidència de fer-ne part—, la tensió ha esdevingut molt forta, sense precedents, i potser per això mateix qualificable provisionalment d’alarmant. Nosaltres, que mirem d’interpretar les coses, tenim la paradoxal tendència, al mateix temps, d’interferir-les, fet que dificulta la nostra pròpia desitjada comprensió. Per això, perquè interpretem, ens hem adonat que les plantes no es distribueixen capriciosament, i també per això altre, perquè interferim, no sempre copsem prou clarament aquest fenomen. Es tracta ara de comprendre’l i d’analitzar-lo.
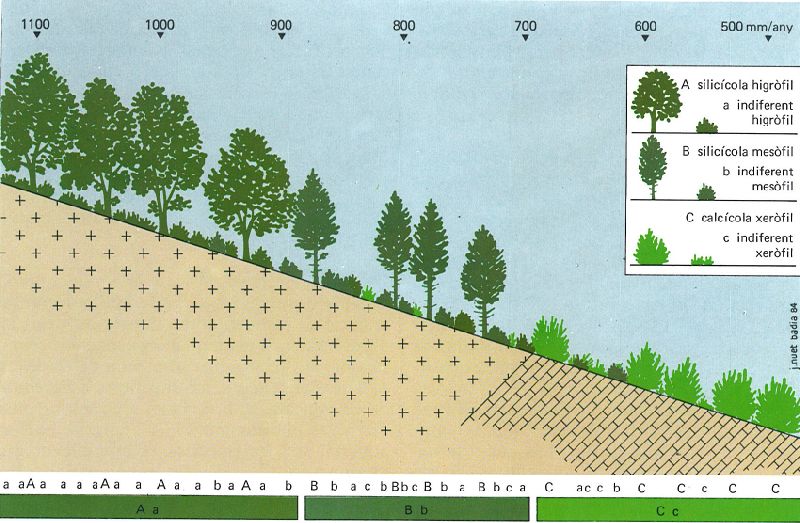
Agrupament de les espècies en comunitats. Disposició exemplar de sis espècies hipotètiques, en funció del gradient de precipitacions lligat a l’altitud i de la natura del substrat (silícic a l’esquerra, calcari a la dreta). La lectura de la seqüència dona una aparent barreja sense sentit, però una visió de conjunt permet de diferenciar tres agrupaments dominants, amb sentit ecològic, o sigui tres "comunitats": la comunitat d’Aa, la de Bb, i la de Cc.
J. Nuet i Badia: original dels autors
Hi ha, en efecte, una tensió entre les plantes i els animals. Però també entre les plantes i els factors inanimats de l’ambient: la temperatura, la humitat, la insolació, els vents, tants d’altres. D’un manera bàsica aquests factors queden compendiats en dos grans blocs: el clima i la natura del sòl. El clima es defineix sobretot en funció de la temperatura i de les precipitacions, tant en llurs valors absoluts, com en llurs oscil·lacions i combinacions al llarg del dia i de l’any, cosa que configura un conjunt extensíssim de situacions. La natura del sòl, a son torn, comporta aspectes molt diversos, entre els quals la textura física, la composició química i la riquesa en elements nutritius, fets que determinaran un ample ventall de possibilitats i limitacions. Cada espècie vegetal té les seves exigències i toleràncies respecte a aquest conjunt de factors i, particularment, respecte a la manera com es combinen. El resultat és que, tal serà la distribució dels factors, tal serà el marc compatible o incompatible per a una espècie determinada. Per això la distribució no és capriciosa, bé que a vegades pot resultar desconcertant: tant com n’és la variació del clima i del sòl.
Davant dels factors ambientals, les plantes reaccionen d’una manera sintetitzadora, no pas analítica. En unes condicions determinades, una espècie concreta pot viure o no pot viure, sense mitges tintes. La impossibilitat de vida ve donada per la inadequació d’un sol factor ambiental o de diversos alhora, o també per la combinació nociva de dos factors que, aïlladament, no són intrínsecament incompatibles (per exemple: temperatura suportable amb humitat alta, o sequedat tolerable amb temperatura baixa, però combinació inadmissible d’aquestes mateixes sequedat elevada i temperatura alta). La compatibilitat d’una espècie amb un indret, doncs, pot determinar-se mitjançant una funció matemàtica força complexa, en la qual intervenen moltes variables, cadascuna de les quals pot invalidar el resultat, si ultrapassa uns límits màxims, o pot mantenir-se en uns marges tolerables que, combinats amb les altres variables, acabin donant un resultat realment vàlid o paradoxalment invàlid malgrat la validesa de cada variable considerada aïlladament. Seduït pel factor ambiental més vistent, l’observador superficial pot arribar a concloure que la distribució és atzarosa, car no troba una correlació entre aquest factor i el comportament de les espècies. La conclusió, aleshores, serà errònia perquè, cal insistir-hi, les plantes reaccionen davant la globalitat dels factors ambientals d’un indret determinat, no només davant d’un de sol, per espectacular que a nosaltres pugui semblar-nos.
Que una espècie pugui viure en un lloc, però, no vol dir que hi pugui viure òptimament. A la noció de compatibilitat, doncs, cal afegir la d’optimació. Si per a un indret concret podem establir una relació relativament reduïda de les espècies que hi resulten compatibles, més reduïda serà encara la llista de les espècies que poden viure-hi òptimament. A l’hora de poblar aquest indret, les espècies incompatibles no arribaran a fer-s’hi, i d’entre les compatibles, les òptimament adequades desplaçaran les que no en són tant. Això porta a la configuració de lots d’espècies que tendeixen a retrobar-se juntes cada vegada que es repeteixen unes determinades condicions ecològiques. D’aquests lots d’espècies, més o menys "fidels" entre elles, hom en diu comunitats vegetals. I les comunitats vegetals, com una mena de supraorganisme, o d’organisme d’organismes, reprodueixen al seu nivell d’integració els mateixos mecanismes de compatibilitat o incompatibilitat, d’optimació o simple tolerància, exposats per a les espècies individuals.
El concepte de comunitat vegetal, per la seva mateixa natura, exigeix una gran flexibilitat de definició. Una comunitat vegetal no és un ens d’estructura fixa i preexistent, sinó un conjunt d’ens resultat d’un procés. Una comunitat vegetal és aquell conjunt d’espècies que solen tendir a aparèixer cada vegada que es donen unes determinades condicions ecològiques. Però les condicions ecològiques no són mai absolutament idèntiques en dos llocs determinats; però l’accés a aquests llocs no és igualment fàcil per a totes les espècies de la comunitat (hi pot haver hagut barreres: un gran riu, per exemple, superable per espècies que confien la disseminació de llurs granes als ocells i insuperable per a d’altres espècies de la comunitat que les confien als insectes); però una maltempsada imprevista (una allau, un incendi, una esllavissada) pot haver afectat diversament les distintes espècies. Això fa que el concepte de comunitat no sigui monolític i calgui entendre’l més aviat en termes estadístics. Un excés de rigidesa en la concepció duria a haver de definir una comunitat diferent per a cada indret, i un excés de laxitud portaria a trobar que totes les comunitats s’assemblen les unes a les altres, o si més no a les veïnes.
El terme comunitat té òbvies connotacions socials. La mateixa ciència que estudia les comunitats vegetals rep el nom de Fitosociologia o Fitocenologia, fet prou revelador. No cal pensar, però, que les relacions entre les plantes que integren una comunitat siguin comparables a les que s’estableixen en una comunitat humana, per exemple. Hi ha més una coincidència en l’espai i en el temps, que no pas una interacció entre els individus. Per això la idea d’organisme d’organismes, més amunt apuntada, ha d’ésser entesa com a recurs retòric més que com a cap altra cosa. Això no obstant, les plantes introdueixen variacions en les condicions ecològiques del lloc on viuen (ombra, retenció, consum d’aigua, etc.), i en aquest sentit actuen indirectament sobre les seves acompanyants. Aquesta actuació consolida la coherència de la comunitat, car restringeix encara una mica més el lot d’espècies (d’entre les òptimes a l’indret, les òptimes a les noves condicions que elles mateixes creen en l’indret) i crea veritables dependències interespecífiques, fet aquest que dona al terme "comunitat" —o a la locució "organisme d’organismes"— un valor més ajustat al sentit semàntic habitual en aquestes expressions. I encara caldria afegir, en aquesta línia, el cas de determinades espècies que, de manera directa, actuen sobre les altres del lot segregant substàncies químiques que en dificulten la presència: són els fenòmens dits al·lopàtics, encara no prou coneguts.
El mateix raonament, doncs, que mena a sostenir que les espècies no es disbribueixen a l’atzar, porta a concloure que s’agrupen en comunitats i que aquestes, a son torn, tampoc no es distribueixen atzarosament. Fets, per altra banda, que la saviesa popular ja havia establert empíricament: és per això que hom sap des de molt antic que existeixen fagedes, bardisses, alzinars o gespets, i que hom no pot esperar de trobarlos, a més, en qualsevol lloc, sinó en aquells que els resulten adequats. Aquest mateix home que ha copsat intuïtivament i empírica el fenomen, però, ha incidit tantíssim sobre la vegetació, ha transformat talment l’entorn, ha urbanitzat de tal manera la seva forma de vida, que en assajar de fer una interpretació científica d’aquesta realitat ha hagut de recórrer a una certa metodologia arqueològica per a reconstruir lògicament allò que, potser, li resultaria evident si pogués contemplar-ho en estat virginal (i no precisament còmode per a l’home, per cert): la distribució no capriciosa de les plantes i llur agrupament, igualment previsible, en comunitat. És a dir, l’harmonia funcional de la vegetació, de la qual la Fitosociologia, aleshores, no fóra més que una descripció quasi pictòrica. Però això ens retorna al començament d’aquest apartat, on ja s’ha comentat el fenomen.
Clímax i comunitats vegetals permanents

Diversitat paisatgística i concepte de clímax. Paisatge nòrdic uniforme i poc humanitzat (vall de l’Athabasca, Canadà) i paisatge mediterrani, divers i molt transformat per l’home (vall de la Tordera, Catalunya): les actituds interpretatives són espontàniament diferents davant dels dos casos.
AISA i J. Nuet i Badia
Hom parla de bioclima per a referir-se al conjunt de factors climàtics que incideixen més immediatament sobre els éssers vius (humitat, temperatura, llum, etc.). Els factors climàtics són, sens dubte, els principals responsables de la distribució de la vegetació. Per això, en cada zona de clima més o menys homogeni (domini climàtic o bioclimàtic)*, és esperable una vegetació també més o menys homogènia (domini climàcic). Sorgeix així el concepte de clímax, que s’aplica a la comunitat normalment esperable en un domini climàtic determinat. Això no obstant, el substrat juga també un paper destacable en la distribució de la vegetació, fins al punt que pot introduir variacions ecològiques de molta consideració al si d’un domini climàtic determinat. Aquestes variacions del substrat, traduïdes en modificacions dràstiques del relleu, en àrees mancades o quasi de sòl, en superfícies on determinats elements químics són inexistents, molt escassos o excepcionalment abundants, etc., poden fer inviable la presència de la clímax en zones que corresponen al seu domini. El lloc de la clímax, aleshores, serà ocupat per d’altres comunitats compatibles amb la situació climàtica i capaces, a més, de suportar les singularitats del substrat en qüestió. Aquestes comunitats, integrades parcialment o no per espècies que també fan part de la clímax i per espècies singulars que normalment no hi apareixen, reben el nom de comunitats permanents, locució que fa referència al seu caràcter de vegetació permanentment instal·lada al si d’una àrea on, per raons climàtiques, més aviat hom n’esperaria una altra. La clímax i les diverses comunitats permanents, doncs, constitueixen la dotació fitocenològica d’un domini climàcic determinat, les anomenades comunitats primàries de l’indret.

Variacions de la vegetació per raó de l’exposició o del substrat. Alternança de vegetació caducifòlia (obacs i fondals) i perennifòlia (solells i dorsals), a Rocacorba (a dalt). Seqüenciació de la vegetació arrelada en substrat submergit (comunitat d’espargani muntanyenc), amarat (comunitat de càrex inflat) i emergit (prat de pèl caní), a la Balmeta, estany del Capcir (a baix).
Ernest Costa, J. Nuet i Badia
No tots els autors coincideixen en aquesta manera de veure les coses. A part els qui ni la tipificació clara de les comunitats no admeten, hi ha els qui donen primacia als caràcters del substrat sobre les característiques del clima, en el qual cas i des del seu punt de vista, tant les clímaxs com les comunitats permanents foren comunitats permanents en els llocs on es troben i prou. En aquestes diferents actituds influeixen molts factors extracientífics d’adscripció a una escola o a una altra —com en política, la manca de millors motius invita a fer bandera irrenunciable de foteses secundàries— i també diferents realitats ambientals dels llocs on operen aquestes escoles.
En efecte, les escoles nòrdiques observen sovint una realitat climàtica prou uniforme i moltes vegades poc transformada per l’home. Tendeixen aleshores a exaltar el paper del substrat davant del clima o els costa de veure la diversitat de les comunitats perquè, efectivament, s’enfronten amb un paisatge vegetal prou uniforme i monòton. Les escoles d’extracció latemediterrània, per contra, viuen en un entorn secularment modificat per l’home, sotmès a contrastos climàtics molt forts i d’una diversitat paisatgística enorme. Es comprèn que aquests diferents punts de partida influeixin fortament en la metodologia d’estudi i que, durant anys, hi hagi hagut serioses discrepàncies, accentuades, com ha estat ja apuntat, per personalismes i altres febleses humanes. Però actualment hom s’adona que aquestes discrepàncies, o són extracientífiques, o afecten més que res la terminologia, raó per la qual tothom tendeix a posar-se conceptualment d’acord, per sobre de divergències de llenguatge. Sobretot perquè les raons autèntiques dels uns completen i matisen les raons autèntiques dels altres: tant el clima com el substrat són importants, i en zones ecològicament molt homogènies els límits entre les comunitats —si hi són— es desdibuixen.

Variacions de la vegetació per efecte del vent. El vent esdevé un factor ecològic tan important com els climàtics els derivats del substrat a les àrees on bufa de forma enèrgica i sostinguda: conformació eòlica de la vegetació als ullastrars de Scandola (Còrsega).
Teresa Franquesa
En qualsevol cas, una cosa és certa: els constrastos deguts al substrat són més forts que els deguts al clima, i això perquè el clima varia gradualment d’una zona a una altra, mentre que les variacions del substrat poden ésser dràstiques en intensitat i sobtades en l’espai. Altrament dit, les variacions significatives del substrat solen traduir-se en canvis molt aparents en la vegetació. És per això que al costat de conceptes genèrics i amplament divulgats —bé que popularment potser no prou precisats— com els de vegetació mediterrània o de vegetació tropical (condicionaments climàtics), coexisteixen concepcions no pas menys divulgades —i popularment prou més precises— com vegetació de roques, de platges o de ribera fluvial (condicionaments del substrat). En aquests darrers ambients és on trobem comunitats integrades per espècies sens dubte compatibles amb el clima general de l’àrea, però versemblantment poc competitives amb les "normals" del domini climàtic, bé que capaces de suportar alguna condició extremosa: quan aquesta condició apareix (empobriment del sòl, presència de certs elements químics, amarament edàfic, etc.), les espècies en conjunt poc competitives, però sofertes davant d’aquest nou factor limitant, desplacen les altres i constitueixen la corresponent comunitat permanent. Entès així, no és difícil comprendre per què en els ambients extremosos és on es concentren la majoria de les espècies "rares": si poguessin viure i competir també en els llocs normals, ja s’hi haurien instal·lat i, esdevingudes comunes, deixarien d’ésser rares (bé que continuarien essent notables). Aquest és el cas de moltes "vulgaritats": es tracta d’espècies remarcables, precisament perquè en tenir una capacitat de viure i competir admirablement àmplia poden aparèixer pertot.
Vegetació primitiva o potencial i vegetació actual: dinàmica i successió
Aquesta suara al·ludida diversitat dels paisatges mediterranis —que, tanmateix, no els és exclusiva, ni de bon tros— és en bona part deguda a la tradicional acció transformadora de l’home. I és que, en efecte, el panorama teòric d’extensos dominis climàcics coberts per les corresponents clímaxs i comunitats permanents no coincideix amb la nostra realitat quotidiana. Enfront d’una més aviat deduïda vegetació primitiva teòrica tenim la realitat més complexa de la nostra vegetació actual, fruit de moltes transformacions i ferides cicatritzades o en cicatrització.
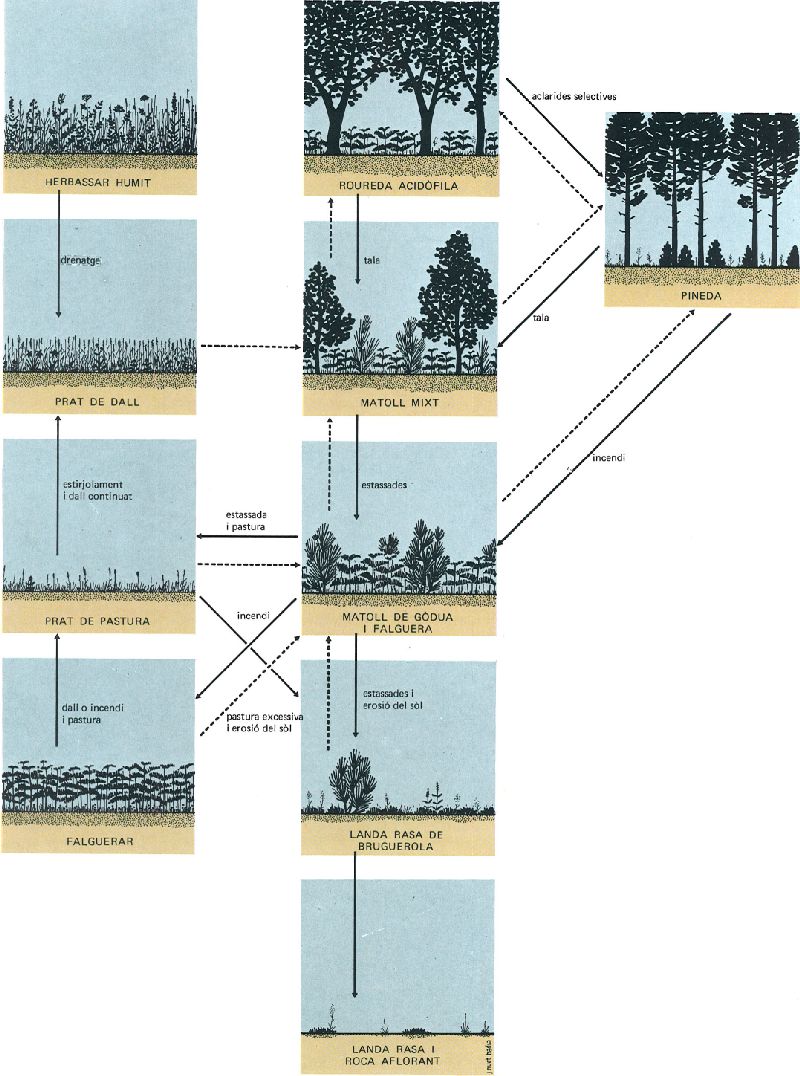
Model de successió de degradació i reconstitució. Esquema de la marxa teòrica dels processos de successió de degradació (línies contínues) i de successió secundària de reconstitució (línies discontínues) d’una roureda humida acidòfila. Noti’s la irreversibilitat del procés quan el sòl és destruït.
J. Nuet i Badia; original de J. Vigo, modificat
En efecte, la destrucció d’una comunitat climàtica o permanent no comporta pas l’anorreament definitiu de la vegetació a l’indret. Ben al contrari, un procés de restauració, comparable com hem dit a la cicatrització d’una ferida, es desencadena immediatament. Granes vingudes d’arreu (el vent, els ocells, etc.) i rebrots de les plantes malmeses reprenen immediatament la recolonització del lloc afectat. En una primera fase, totes les espècies compatibles, posat que arribin a l’indret, tindran l’oportunitat de fers’hi presents. No pas necessàriament les més ben adaptades —les òptimament compatibles—, sinó les més rústegues i diligents en la creixença seran les que, inicialment, dominaran l’espai, tal com es comprèn fàcilment. Això comportarà la instal·lació d’unes comunitats més o menys efímeres, d’espècies sofertes i ràpides, però no òptimament eficaces, les anomenades comunitats secundàries o transitòries. Amb el temps, i fins i tot a l’empara de les condicions ambientals més propícies que les mateixes comunitats transitòries hauran creat, les espècies òptimes acabaran reprenen llur domini i restaurant, doncs, la comunitat climàcica o la comunitat permanent que hom havia destruït. Haurà tingut lloc, així, un procés de recuperació mitjançant la successió de diverses comunitats transitòries, cada vegada més pròximes i assemblades a la definitiva, l’anomenada per això mateix successió reconstitutiva.


Successió secundària en camps abandonats. Ocupació espontània de feixes d’antics vinyets per herbassars d’albellatge i pi pinyer, a la península del cap de Creus (a dalt). Cups vinícoles del s. XIX i vinyes abandonades engolides per la pineda de pi blanc, al Pont de Vilomara i Rocafort (Bages) (a baix).
Teresa Franquesa
La successió reconstitutiva pot exigir molts d’anys, algun segle fins i tot. Les etapes intermèdies en què domina una determinada comunitat transitòria representen sempre graus creixents de complexitat organitzativa, més difícil d’incrementar com més avança el procés. Les primeres comunitats transitòries, normalment herbàcies, s’implanten en mesos, i són desplaçades aviat per d’altres d’igualment transitòries, però habitualment arbustives i d’estructura més complexa. Les espècies llenyoses de gran port —posat que la clímax o comunitat permanent destruïda fos de caràcter forestal— apareixen més tardanament, o bé es mantenen en una lògica fase de plançó durant els primers anys. Es tracta, per tant, d’un procés que sempre avança en el sentit d’incrementar la complexitat i que dura anys. És per això que no resulta evident als nostres ulls, perquè és lent i aparentment insensible, com el moviment de les busques d’un rellotge, tan cert com imperceptible a primera vista.
Les comunitats transitòries, bé que passatgeres, són autèntiques comunitats, no pas mescles aleatòries d’espècies oportunistes. Llur composició florística, això sí, és més laxa que la de les comunitats madures climàciques, sobretot la de les comunitats transitòries corresponents als primers estadis de la successió reconstitutiva. Es tracta de comunitats poc exigents, capaces de repetir-se en successions que menen a clímaxs distintes, si més no en les primeres fases; després, com més avançada està la successió, més específiques en resulten les comunitats transitòries. A vegades, aquestes comunitats transitòries són idèntiques a les que resulten permanents en certs indrets especials (llocs amb poc sòl, per exemple): hom dedueix que en aquests llocs ingrats la successió ja no pot anar més endavant i acaba esdevenint-hi permanent allò que en punts més dotats no és sinó una fase transitòria vers la complexitat climàcica.

Colonització primària. Sobre roques nues aflorants tenen lloc processos primaris de colonització (successió primària), edificadors de sòl, com els protagonitzats per aquests pins negres i nerets sobre granitoides dels Pirineus.
Adolf Sostoa
Una reconstitució com la descrita correspon a una successió secundària, car resulta subsegüent a una destrucció prèvia. Hom anomena, per contra, successió primària el procés que duu a la colonització per part de la vegetació d’una àrea de sempre desproveïda. És com a culminació d’una interminable successió primària que la Terra primigènia es cobrí gradualment de vegetació, i són encara actualment en fase de successió primària els estadis de colonització de roques nues, poblades primer de líquens i molses, després de petites herbetes i més tard de vegetació llenyosa. En aquest sentit, les comunitats permanents dels llocs especials esmentades més amunt hi fan també el paper de comunitats transitòries en el llarguíssim procés de la successió primària. En conjunt pot dir-se que la successió primària segueix el ritme pausadíssim de la formació del sòl, a la qual formació contribueix de manera decisiva. Per això, si la destrucció de la vegetació d’un indret —allò que hom anomena la degradació— comporta la destrucció del sòl, no pot desencadenar-se la successió secundària de reconstitució (que dura decennis), i en el seu lloc s’inicia una lentíssima successió primària (que necessita mil·lenis).
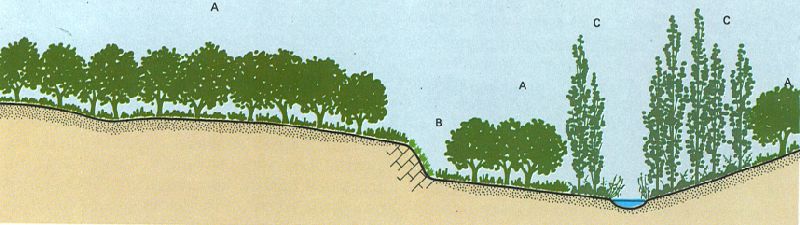
Vegetació primitiva, actual i potencial. Seqüència hipotètica de la vegetació primitiva (a dalt), actual (al mig) i potencial (a baix) d’un indret determinat. Els boscos esclerofil·les (A) o de ribera (C) i la vegetació rupicola (B) primitius, romanen més o menys ben constituïts en l’actualitat —parcialment desplaçats per camps i construccions, o per brolles (A’) i bardisses (C’)— i podran refer-se en el futur, excepte en les àrees en què el sòl hagi desaparegut o esdevingut inaccessible.
J. Nuet i Badia; original dels autors
Aquesta dialèctica entre la degradació i la reconstitució constitueix la dinàmica de la vegetació, fatalment dirigida sempre vers la consecució de la vegetació estabilitzada final d’un indret o vegetació potencial. Aquesta vegetació potencial —la vegetació que s’hi pot fer— és precisament la vegetació primitiva, si res no l’ha destruïda, o coincideix teòricament amb ella, si el sòl primitiu no està malmès. Si en degradar la vegetació primitiva hom ha malmès també el sòl, la vegetació potencial esperable ja no serà lògicament com la primitiva. La vegetació actual d’un indret humanitzat, doncs, sol ésser un complex mosaic de comunitats transitòries i de restes de vegetació primitiva (fragments de comunitats climàciques i permanents), mosaic que tendeix vers una vegetació estabilitzada final tant més semblant a l’antiga vegetació primitiva com menys destructora per al sòl hagi resultat la degradació (les àrees sense o amb poc sòl, encara en fase de successió primària i les variacions que mentrestant pugui anar experimentant el clima, completaran i complicaran el panorama, tanmateix, de la suposable vegetació estabilitzada final). En resum podem dir que la successió reconstitutiva secundària tendeix a transformar la vegetació actual (on solen predominar les comunitats secundàries o transitòries) en una vegetació estabilitzada final, és a dir a dotar cada domini climàcic de la vegetació potencial (les comunitats estables o primàries, o sigui les climàciques i les permanents) que hi és esperable, vegetació potencial que correspondrà en força casos a la vegetació primitiva que hi havia hagut com a fruit primigeni de la successió constitutiva primària.
Tipus de vegetació
Termes com prat, brolla o bosc, o d’altres de més precisos com gespet, boixeda o carrascar, pertanyen immemorialment al llenguatge comú. Serveixen per a designar realitats conegudes popularment, fàcilment identificables per al camperol o per qualsevol persona interessada. Corresponen a casos concrets de tipus de vegetació, de les agrupacions que hom pot fer amb les comunitats d’aspecte semblant.
L’aspecte o fisiognomía d’una comunitat és, en efecte, el primer tret de què hom s’adona. Una segona aproximació permet d’identificar l’espècie o espècies dominants. I un estudi més detallat fa possible la determinació de totes o de la majoria de les espècies presents, així com de llur importància quantitativa. Per a reconèixer inequívocament una determinada comunitat cal atènyer aquest tercer estadi, precisament l’objecte primordial de la fitocenologia. La fisiognomia, doncs, suposa una primera aproximació fitocenològica i permet una primera designació dels tipus de vegetació, prèvia a 1’apel·lació de les comunitats. Fisiognòmicament hom diferencia, per a la nostra vegetació autòctona, tres grans tipus de vegetació: els boscos, les bosquines i els prats.
Els boscos

Tipus fisiognòmics de formacions forestals. La majoria de les nostres comunitats o formacions forestals són referibles a algun o alguns d’aquests cinc grans tipus fisiognòmics: A, bosc pluriestratificat (en aquest cas, alzinar); B, bosc monolbiestratificat o boscana (en aquest cas, avetosa); C, arbreda artificial (en aquest cas, pollancreda); D, bosc gros o de llavor (en aquest cas, pineda molsosa de pi roig); E, bosc menut o de rebrot (en aquest cas, perxades de castanyeda). Aquests cinc grups no són excloents, de manera que l’alzinar d’A, és un bosc gros com la pineda de D, per exemple.
J. Nuet i Badia; original dels autors
Apliquem el terme de bosc a un conjunt vegetal on un o diversos estrats d’arbres dominen sobre un o diversos estrats herbacis i/o arbustius, l’anomenat sotabosc. Bé que el sotabosc és part integrant i inseparable del bosc, són els arbres qui dona al bosc el seu caràcter distintiu i característic. En alguns casos, com en els alzinars, el sotabosc és dens i riquíssim en espècies llenyoses; en d’altres, com algunes rouredes i les fagedes, és esclarissat o quasi només integrat per espècies herbàcies (d’aquests boscos se’n diuen boscanes, prou diferents a les simples arbredes o plantacions artificials d’arbres que no són de cap manera un bosc). Reben el nom de bosquetons els boscos de port discret, integrats per arbres baixos o d’aspecte arbustiu, i hom denomina boscanys les restes d’un antic bosc que han quedat isolades al si d’una àrea conreada o d’un jardí. La denominació de bosc en galeria és aplicada a la comunitat forestal que ressegueix, allargassada, un accident geogràfic determinat, com és el cas dels boscos de ribera, que són els que es fan a les ribes fluvials o a vegades lacustres.
Els boscos, siguin comunitats permanents o transitòries, han sofert intensament l’acció de l’home i se’ns presenten transformats molt sovint. A l’àrea mediterrània, on els boscos naturals solen dur sotaboscos atapeïts, hom s’ha esmerçat a destruir els estrats no arboris mitjançant estassades més o menys enèrgiques. L’obtenció de llenya en alguns casos, o l’impossible desig de convertir aquests boscos en boscanes transitables com les medioeuropees, han estat els mòbils usuals. Això ha popularitzat expressions com la de "bosc brut", aplicada a boscos amb sotabosc dens, i "bosc net" destinada a les boscanes o als boscos estassats. És cert que una autèntica boscana, d’arbres alterosos i potents, resulta molt bella i còmoda de transitar, però no per això els boscos de sotabosc dens són abominables, ni sempre prou bons els resultats d’una estassada: un "bosc net" pot massa sovint fer l’efecte d’un paraigua sense tela, privat de cohesió i solidesa (no podem eliminar la majoria de les espècies d’una comunitat sense que aquesta perdi sentit i funcionalitat). També per intervenció humana directa s’estableix la situació de bosc baix o menut, en els quals els arbres procedeixen de soques rebrotades, subsegüents a tales prèvies, que s’oposa a la de bosc alt o gros, la normal, en la qual els arbres són tots nascuts de llavor.
L’arbre dominant acostuma a donar nom propi als boscos, designats d’aquesta manera rouredes o rovires, freixenedes, alzinars, salzedes, avetoses, pinedes, etc. Aquestes denominacions, que afegeixen precisions florístiques al simple terme de bosc, ultrapassen la mera aproximació físiognòmica i s’acosten molt —a vegades del tot— al concepte de comunitat vegetal. Tindrem ocasió de comprovar-ho més endavant.
Les bosquines

Tipus fisiognòmics de formacions embosquinades. La majoria de les nostres comunitats o formacions embosquinades són referibles a algun d’aquests sis grans tipus fisiognòmics: A, màquia; B, brolla o timoneda; C, brolla arbrada o pineda secundària; D, matoll; E, landa; F, bardissa.
J. Nuet i Badia; original dels autors
Els conjunts vegetals desprovists d’arbres i integrats majoritàriament per plantes arbustives són les denominades bosquines. Hom parla més concretament de màquia, de brolla o de timoneda, de garriga, de matoll, de landa i fins de bardissa segons el port i densitat dels arbusts dominants. Com el cas dels boscs, també hi ha bosquines amb nom propi, derivat de l’arbust preponderant: boixedes, brugueres, arboçars, etc.
La màquia és la més imposant de les bosquines. El terme, que és d’origen italià, designa formacions molt atapeïdes i altes de fins 1,5-2 m, normalment de caràcter mediterrani; actualment les màquies resulten ben escasses al nostre territori. La brolla, per contra, constitueix la bosquina més difosa; hi dominen els arbusts baixos o subarbusts (0,5-1 m), com els romanins i les estepes, i si tan menuts i esclarissats són, amb predomini de les farigoles o timons, la brolla pren el nom de timoneda (que fóra un nom propi, si no fos que designa totes les brolles d’escassa entitat quantitativa). Aquest darrer és també el cas de la garriga, denominació d’origen català i occità, aplicat a les brolles rases, denses i punxoses, hi domini o no el garric o coscoll (alguns autors, sobretot francesos, arriben a utilitzar el terme garriga com a sinònim i substitutiu del de brolla). L’espinar, encara, fóra una mena de garriga esclarissada, de brolla poc densa i punxent, si es vol. I, finalment, hom parla de brolla arbrada per a referir-se al conjunt de vegetals, corrents al nostre país, on una simple brolla es ve completada per un estrat arbori lax superposat i més o menys independent; moltes de les pinedes de terra baixa es troben en aquest cas, bé que popularment hom les qualifiqui sempre de boscos (se’n diferencien perquè la brolla no és un sotabosc que depengui de la presència dels arbres, sinó un conjunt que viu prou independentment d’aquests).
Les bosquines extramediterrànies tenen un aire ben diferent de les màquies, les brolles o les garrigues. Poden estar integrades per arbusts alts de 2 m i més, de fulla blana o no, i aleshores constitueixen els matolls, dels quals foren casos concrets les avellanoses, els baleguers o fins les boixedes. O bé poden prendre l’aspecte d’un tapís arranat, o si més no baix, fet que s’esdevé sobretot sobre sòls àcids, i en aquest cas prenen el nom genèric de landa. A les landes predominen els brucs, la bruguerola i espècies de fulla menuda afins, mentre que els matolls més típics estan presidits per les genísties (per això les boixedes o les avellanoses suara esmentades no són considerades matolls per alguns autors).
Un últim cas de bosquina, jà molt extrem, és el de la bardissa, conjunt vegetal impenetrable, propi sobretot d’indrets humits, en el qual predominen diverses lianes punxents i arbusts espinosos: rosers, arítjols, esbarzers, etc.; contràriament a les altres bosquines, no solen recobrir grans extensions, car es limiten als enclavaments particularment humits d’on són pròpies.
Els prats
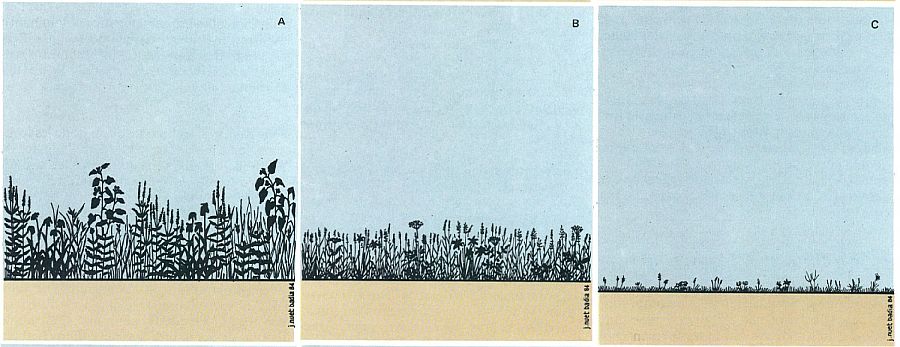
Tipus fisiognòmics de formacions herbàcies. La majoria de les nostres comunitats o formacions herbàcies són referibles a alguns d’aquests tres grans tipus fisiognòmics: A, herbassar alt o fenen B, prats de dall; C, prat ras.
J. Nuet i Badia; original dels autors
Als prats, la dominància, i fins la presència exclusiva, correspon a les plantes herbàcies. No hi ha mai arbres ni grans arbusts, només a vegades alguna planta de port Ilenyós. El trànsit hi és, òbviament, molt fàcil, bé que pot veure’s dificultat en alguns casos per l’elevadíssima densitat de l’herbei.
Els prats constituïts per herbes altes i de fulla més aviat grossa reben el nom genèric d’herbassars, tot i que el terme, per a molts, és simple sinònim de comunitat herbàcia. Bons herbassars foren les comunitats megafòrbiques o megafòrbies, conjunts de grandíssimes herbes d’alta muntanya, altes com un home, o bé els feners, comunitats també muntanyenques, riques i humides. En canvi, els conjunts integrats per herbes menudes, d’aspecte graminoide, no són qualificats d’herbassars, i més aviat són ells qui s’emporten la denominació de prat per antonomàsia.
És en el conjunt dels prats pròpiament dits que hom distingeix els pradells (prat d’extensió reduïda, integrat per herbetes menudes) dels gramenets (el cas més genuí, car en són tots els prats dominats per gramínies o plantes graminoides perennes). Un cas especial de gramenet el constitueixen les gespes, formades per plantes altes de pocs centímetres, però molt atapeïdes, prats aquests ben típics de l’alta muntanya, el paisatge de la qual caracteritzen en constituir-se en dilatats rasos que cobreixen grans extensions. Aquestes gespes rases, justament perque suporten una pastura extensiva tradicional, reben el nom de prats de pastura, enfront dels anomenats prats de dall, gramenets alts o herbassars que hom dalla regularment. A l’altre extrem de la gamma, hom parla d’erm per a referir-se a un prat eixut d’estructura molt laxa, amb espais sense vegetació enmig de llurs herbetes esparses, propi d’indrets particularment àrids. I, finalment, de gramenet embosquinat per a designar conjunts a cavall dels prats i de les brolles, mixtura de vegetació herbàcia i arbustiva baixa, com és el cas de les joncedes.
Altres tipus de vegetació
La majoria de les nostres comunitats són assimilables a un bosc, a un prat o a una bosquina, però n’hi ha que no s’ajusten a aquesta tipologia. Les mateixes bardisses foren un mal exemple de bosquina, i cal admetre que resulta una mica excessiu considerar les joncedes com un prat. Però les discrepàncies més grans es produeixen en el cas de les comunitats pobladores d’ambients especials, majorment roques i zones aigualoses. Així, els canyissars, els poblaments de llentilles d’aigua, els creixenars, les jonqueres, les comunitats rupestres, etc., no són pròpiament ni un prat, ni una bosquina. Constitueixen tipus de vegetació singulars, no assimilables a cap dels tres bàsics i habituals al nostre país.
Formacions vegetals
Els tipus de vegetació no esgoten totes les possibilitats d’una descripció fisiognòmica, ni molt menys. Així, hi ha conjunts ben coneguts, com els canyissars, les jonqueres, els saladars, etc., que no són ni un bosc, ni ben bé una bosquina, ni tampoc un prat. Altrament, la gamma de realitats de vegetació que s’ofereix als nostres ulls va molt més enllà de la vintena de tipus suara enumerats. Una aproximació fisiognòmica més precisa ens permet de matisar els tipus de vegetació amb qualificatius fenològics o morfològics, és a dir amb qualificatius referits a l’aspecte de les plantes observades (morfologia) o a Ilur capteniment al llarg de l’any (fenologia). És així com neix el concepte de formació vegetal.
En termes de formació vegetal, hom parla de boscos planocaducifolis, de boscos aciculoperennifolis o de boscos esclerofil·les, per exemple, per a referir-se respectivament a boscos dominats per arbres de fulla plana i caduca, en forma d’agulla (acicular) i perenne, i de fulla petita i dura i també perenne. (Són caducifòlies les plantes que, estacionalment, perden (i renoven) totes llurs fulles de cop. Per contra, hom anomena perennifòlies aquelles que renoven rotativament llur fullam, sense quedar-ne mai del tot desproveïdes. Segons que les fulles siguin amples i planes o bé estretes i en forma d’agulla, hom parla de plantes planocaducifòlies (els roures, el faig) o aciculocaducifòlies (els làrixs), o bé de plantes planoperennifòlies (l’alzina, el llorer) o aciculoperennifòlies (els pins, l’avet). Són plantes esclerofil·les les de fulla petita, plana i dura.) Així, les rouredes o les fagedes serien boscos planocaducifolis, mentre que les pinedes o les avetoses foren aciculoperennifolis, i els carrascars o les suredes resultarien boscos esclerofil·les. Aquesta metodologia és extensible a d’altres tipus de bosc i, naturalment, a les bosquines i als prats.
Per a referir-se a grans extensions de territori, l’ús de les formacions vegetals és útil i interessant. Ofereix, en efecte, un nivell de precisió prou bo, i té l’avantatge que resulta fàcilment comprensible per a una persona mínimament iniciada. És per això que l’emprarem nosaltres mateixos en sumaritzar els dominis mundials de vegetació. Per contra, resulta encara molt insuficient quan hom es refereix, amb un cert detall, a la vegetació d’una àrea més restringida, el nostre país per exemple. Tant és així que fins la terminologia popular, quan parla d’alzinar o de freixeneda, ateny una precisió més elevada, bé que menys intuïtivament gràfica, que quan hom diu bosc esclerofil·le o bosc planocaducifoli: les primeres designen comunitats concretes, les segones només famílies fisiognòmiques de comunitats.
Denominació i sistemàtica de les comunitats vegetals
El progrés en el coneixement de la vegetació ha portat a la identificació i descripció gradual de noves comunitats, que han hagut d’ésser denominades de manera precisa per a poder-s’hi referir inequívocament. Per això les diverses escoles de fitocenòlegs han desenvolupat sistemes apel·latius diversos per a designar les comunitats identificades, però aquests sistemes sovint resulten complexos i quasi mai comprensibles sense una iniciació prèvia. Els estudis fitocenològics exigeixen un cert domini d’aquests sistemes, alguns dels quals han estat adaptats a les exigències del llenguatge comú. En aquesta obra, concretament, hom adopta la metodologia i llenguatge dits sigmatistes (de S.I.G.M.A.: Station International de Géobotanique Méditerranéenne et Alpine), creats pel botànic grisó establert a Montpeller Josias Braun-Blanquet (1884 - 1980); alhora, hom empra una adaptació al llenguatge català (els termes sigmatistes conserven la forma llatina usual en la taxonomia biològica) en la línia seguida per diversos botànics catalans, entre els quals Oriol de Bolòs, Josep Vigo i nosaltres mateixos. Aquesta doble apel·lació permet a l’especialista de guiar-se per la nomenclatura internacional, i tant a l’especialista com a l’home del carrer d’emprar també una terminologia catalana vulgaritzada, bé que culta.
En la confecció d’aquesta terminologia, hom ha recuperat mots d’ús popular, tot fixant-ne l’abast, hom ha creat termes vulgars nous, i sobretot hom ha generat locucions de nova planta mitjançant la combinació de tots aquests elements. És per això que en l’apartat destinat als tipus de vegetació hem precisat el sentit de tots aquests termes elementals: perquè tenen un valor històric en la llengua vulgar i perquè ens permeten ara de redimensionar la llengua culta mitjançant recursos genuïns. D’aquesta manera han nascut locucions com màquia litoral, roureda de roure pènol, bardissa amb boix, herbassar de vorada de bosc, timoneda gipsícola, gespa de pèl caní. Els uns, normalment els que duen el nom d’una espècie concreta, solen designar comunitats precises (aquelles en què l’espècie esmentada és característica o dominant); els altres, usualment els que porten qualificatius geogràfics, topogràfics, físiognòmics o ecològics, acostumen a designar conjunts o sèries de comunitats relacionades entre elles.
La sistemàtica sigmatista mereix un comentari concret, a fi de comprendre l’abast de les locucions de forma llatina emprades, i la lògica de la terminologia catalana que les tradueix. L’associació és la unitat fonamental del sistema, i equival a comunitat vegetal elemental. Les diverses associacions són designades afegint el sufix -etum a una locució formada amb els noms científics d’una o de dues espècies típiques de la comunitat (a vegades hom empra també noms llatinitzats de caràcter geogràfic, circumstància que ha donat lloc a alguns abusos localistes, raó per la qual hom ha acordat de no formar més noms amb aquest criteri, i fins de revisar els antics que els duien). Per aquest sistema apareixen denominacions com Saniculo-Taxetum (de Sanícula i Taxus), Brachypodietum retusi (de Brachypodium retusum), Quercetum ilicis galloprovinciale (de Quercus ilex i galloprovincialis, o sigui provençal), etc. El guió separa els noms de dues espècies diferents i per això quan no hi és sabem que hom ha designat la comunitat mitjançant una sola de les seves espècies. Actualment hom tendeix a emparar el nom científic complet de la o de les espècies escollides (com en Brachypodietum retusi) i a desestimar formes simplificades com Saniculo-Taxetum (que esdevindria Saniculo europaeae-Taxetum baccatae o almenys Saniculo-Taxetum baccatae, de Sanícula europaea i Taxus baccata).
Les associacions semblants són convencionalment agrupades en subaliances i aliances, aquestes en ordres, els ordres en classes, i les classes en divisions. Els noms d’aquests distints rangs jeràrquics s’obtenen de la mateixa manera que els de les associacions, però emprant sufixos diferents: -enion i -ion per les subaliances i aliances, -etalia per als ordres, -etea per a les classes i -ea per a les divisions. D’aquesta forma apareixen noms com Seslerio-Pinenion, Onopordion arabici o Rosmarino-Ericion, com Prunetalia spinosae o Pino-Juniperetalia, com Quercetea ilicis o Calluno-Ulicetea, o com Galeopsiea o Vulpio-Brachypodiea retusi. Quan hom vol matisar petites diferències al si d’una mateixa associació, és possible recórrer a la subassociació, que serà designada afegint al nom complet de l’associació un nou terme afectat del sufix -etosum; això dona noms prou complexos com és ara Buxo-Quercetum pubescentis fagetosum o bé Buxo-Quercetum pubescentis hylocomio-pinetosum sylvestris, que es corresponen (són els casos de màxima complicació, cal aclarir-ho) a les denominacions catalanes de roureda de roure martinenc amb faig i de pineda rovirosa de pi roig amb molses.