
La situació de cada punt del riu dins la seva conca condiciona en gran part les seves característiques. Hom ha ideat, per això, un sistema per a jerarquitzar-los definint diferents graus: per exemple, els rius de grau 2 es formen amb la unió de dos rius de grau 1 (que no tenen cap afluent); els de grau 2 no salten de nivell jeràrquic fins que no es troben amb un altre del seu mateix grau, i així successivament.
Román Montull.
Els rius formen una xarxa que reuneix i drena cap a la mar les aigües que recull de les diferents conques hidrogràfiques. La quantitat i la qualitat d’aquestes aigües és en relació amb els processos que s’hi han desenvolupat, tant els d’origen natural com els que són resultat de l’acció de l’home. Margalef ha comparat la conca hidrogràfica i els rius que la drenen amb un ronyó: així com una bona anàlisi d’orina dóna la informació necessària per saber què té un malalt, una bona anàlisi de la qualitat de l’aigua ens indica què passa a la conca, ens informa dels processos que hi ocorren. Certs processos que passen a les conques hidrogràfiques o els símptomes de la contaminació requereixen anàlisis més o menys complexes i repetides. En el cas dels rius, els organismes que hi viuen es comporten com a metabòlits complexos: pel fet que no són arrosegats avall directament, acumulen informació del que s’ha esdevingut al riu, i es transformen en veritables indicadors del seu estat de salut. Podríem comparar-ho a les pedres del ronyó, que segons la grandària, forma o composició ens informen de la gravetat i de la duració del procés. No és estrany, doncs, que l’estudi dels rius s’hagi transformat en l’estudi de les conques hidrogràfiques els aspectes generals de les quals ja han estat tractats en les pàgines anteriors. Ens referirem ací, doncs, als aspectes relacionats especialment amb les comunitats d’organismes que viuen als rius.
La grandària, la forma i la complexitat de la xarxa de drenatge dels rius no són factors independents. La relació ha estat ben estudiada emprant un ordre jeràrquic, de manera que hi ha rius de grau 1 (els que no tenen afluents), rius de grau 2, que són la unió de dos rius de grau 1, rius de grau 3, que resulten de la fusió de dos rius de grau 2, i així succesivament. En aquest sistema, un riu de grau inferior no modifica el grau d’un riu d’ordre superior: un riu de grau 1 no modifica l’ordre d’un de grau 2, etc. L’escala en què es realitza el mapa de la jerarquia dels rius és molt important, ja que determina el grau màxim a què pot arribar un riu. En el cas de l’ecologia, s’empren aquells mapes on es considera que el grau 1 correspon a cursos d’aigua permanent. L’escala és normalment el mapa 1:200 000. Als Països Catalans, amb aquesta escala s’arriba a un grau màxim de 6-7. La majoria de les conques dels Països Catalans són petites o mitjanes. L’única conca gran, la de l’Ebre, hi és compresa només parcialment. Això explica, en conjunció amb el clima, el règim mediterrani típic dels nostres rius, amb aiguats i inundacions freqüents alhora que amb secades importants. Els gràfics que hom pot elaborar del cabal mitjà mensual de diferents rius són prou eloqüents en aquest sentit, com també la variació que hi ha, en una mateixa conca, d’anys amb molta aportació d’aigua i d’anys molt secs; això genera la imprevisibilitat pròpia dels nostres rius.
La majoria dels rius mediterranis són fortament influïts per les activitats de l’home. Amb els anys, l’home ha anat incidint cada vegada més en el règim dels rius com a resultat de les demandes cada vegada més grans d’aigua. Avui, gairebé cap dels nostres rius és lliure d’aquesta influència i, per tant, el flux d’aigua que s’observa a les parts més baixes és un reflex de la importància d’aquestes activitats de regulació. Fins i tot un riu com l’Ebre és fortament regulat, i l’aigua que porta és relacionada amb l’activitat de les grans preses que hi ha al mateix riu o les conques pirinenques dels seus afluents: poden arribar a ser més importants les fluctuacions que hi ha entre dia i nit que no pas entre dos mesos successius. Evidentment, les comunitats d’organismes es troben sotmeses a aquestes intervencions. Un bon diagnòstic de tots els rius ha de basar-se en els processos que ocorren en la seva conca. Els processos de regulació dels cabals d’aigua són, sens dubte, els canvis més importants que els rius, com a sistemes naturals, han patit.
Dinàmica i funcionalitat dels ecosistemes fluvials

Vistes de les parts alta i baixa del Ter, per a il·lustrar la manera com la posició d’un tram determinat del riu pot influir en les seves característiques globals. A la part alta del riu (a dalt), de grau 2, el fort pendent (predomini dels processos d’erosió) i el cabal reduït (conca pròpia encara petita) fan que prengui l’aspecte dels cursos d’alta muntanya: llit pedregós, amb palets relativament grans que ofereixen nombroses oportunitats de colonització als animals, vegetació de ribera que s’endinsa força en la llera, aportació quantitativament important de fulles dels arbres caducifolis, aigües fredes i corrent fort. A la part baixa del riu (a baix), l’amplada i el cabal ja són considerables i el poc pendent i l’augment de la fondària creen condicions de vida molt diferents per als organismes. L’acumulació de sals i nutrients arrossegats des de les parts més altes i el flux més laminar i lent poden, fins i tot, afavorir la presència de plàncton; en aquest tram han desaparegut els còdols i els fons són fangosos o sorrencs, i tot i l’exhuberància de la vegetació litoral, la seva aportació de materials al riu és relativament petita en relació amb els que el riu transporta. Així, si el riu a les parts altes mostra una gran dependència de la conca, a les parts baixes les seves característiques depenen més d’ell mateix i del transport de materials provinents d’altres parts de la conca.
Narcís Prat i Josep M. Barres
Del que hem dit fins ara deduïm que no podem pas separar el riu de les altres parts que formen la seva conca, ja que tot el que passa dins del riu en depèn. Així, tots els processos ecològics que hi puguem identificar manifesten aquesta dependència, des de les característiques morfològiques de la llera en un punt determinat fins al flux d’energia que s’hi estableix. L’amplada del riu, el tipus de substrat, la quantitat d’aigua que porta depenen de la situació del riu dins de la conca (capçalera o tram baix), però també del que s’ha esdevingut en el tram immediatament anterior. Tot plegat ho trobarem reflectit en el funcionament del riu en cada tram, en el tipus i la complexitat de les comunitats d’organismes, i també en les relacions que hi ha entre els organismes i el seu medi. Tot i que els estudis ecològics dels nostres rius han tingut un caire descriptiu de les comunitats presents o s’han centrat en alguns temes de composició química, és molt important conèixer com s’origina el flux d’energia en el riu, per entendre la composició i dinàmica de les seves comunitats. Aquest tipus d’aproximació és la més corrent avui dia en els estudis sobre ecologia de rius. Els estudis dels darrers anys ens han permès també posar dins d’aquest marc general el que avui coneixem dels nostres rius com a entitats ecològiques.
Organització de les comunitats segons el flux d’energia
La xarxa tròfica amb relació als materials al·lòctons
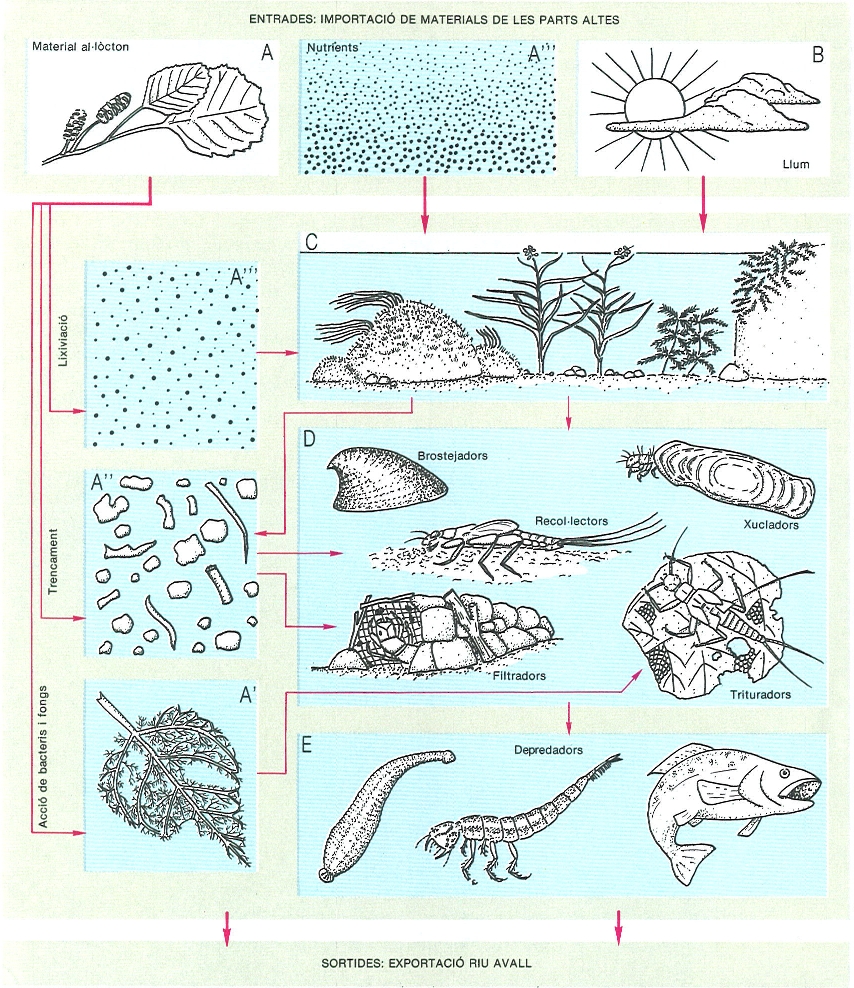
Organització de la comunitat en els rius segons el flux d’energia. La presència d’un grup determinat d’organismes en un punt del riu depèn de la font d’energia externa. Si el que s’utilitza és el material format a la conca (fulles, si el riu és cobert pel bosc, com a A), dominen els materials particulats grollers, colonitzats per bacteris i fongs (A’); com a conseqüència, la comunitat animal és formada especialment per trituradors (plecòpters). Si, en canvi, el riu és obert a l’entrada de llum (B) i porta nutrients abundants (A"’) la producció primària és alta, tant d’algues (epilíton) com de macròfits o molses (C); en aquest cas, els animals dominants són brostejadors (gasteròpodes) o xucladors (tricòpters). En tots dos casos, els materials de gra fi que resulten (A") poden quedar acumulats en zones de poc corrent, on són recollits pels recol·lectors (efemeròpters) o bé són arrossegats avall i capturats en les xarxes dels filtradors (tricòpters). Tots poden ser menjats pels depredadors (E). El grau de complexitat de la comunitat és amb relació a la importància del flux d’energia i a la categoria jeràrquica del riu.
Maria Rieradevall.
Sovint hom considera el riu com un sistema heterotròfic, que depèn de l’entrada d’energia externa originada a la seva conca; és a dir, dels materials orgànics que provenen dels ecosistemes veïns, com ara les fulles o els branquillons dels arbres propers (matèria orgànica al·lòctona). En aquest cas se suposa que la producció primària dins del riu (derivada dels organismes autòctons autòtrofs, com molses o algues) és poc important. Això és especialment veritat a les parts altes de la conca, on els rius són estrets i amb poc cabal.
En un riu situat en una conca ben forestada, l’aportació de materials orgànics que provenen d’aquesta cobertura, com les fulles, és molt alta i és especialment important, si el riu no és gaire gran (d’ordre 1 a 3). Això originarà, dins del riu, d’una banda, una gran abundància de materials que hauran de ser descompostos, i de l’altra, la presència de barreres físiques (branques caigudes al riu, per exemple) que retenen el pas de l’aigua. Alhora, la densa capçada dels arbres limita l’arribada de la llum i això fa que el creixement dels vegetals aquàtics sigui limitat; en canvi, hi ha molt de material al·lòcton per descompondre, per la qual cosa trobarem molts bacteris i fongs fent aquesta funció. La majoria dels nostres torrents que corren per conques forestades tenen aquestes característiques, sigui al Montseny, als Pirineus o a les muntanyes de Prades. Els vegetals de vida aquàtica que trobem en aquests casos són, preferentment, molses adaptades a ambients ombrívols i frescals.
En aquests mateixos rius, i associats a aquests materials en descomposició, hi ha una munió de petits animals que, aparentment, s’alimenten de les fulles caigudes al riu, però que en realitat les trenquen a trossets i es mengen preferentment els fongs i bacteris que hi han crescut al damunt. Un experiment al laboratori mostra que les fulles esterilitzades (és a dir, de les quals hom ha eliminat la població bacteriana i fúngica) no són menjades per aquests animals; només les fulles que han estat un cert temps a l’aigua (o sigui, que han tingut temps de ser colonitzades per microorganismes) permeten que els animals hi creixin. Així, doncs, el material de dimensions mitjanes o grans que ha caigut dins del riu, que hom anomena matèria particulada orgànica grollera (MOPG), afavoreix la vida d’una munió de petits invertebrats, especialment larves d’insecte, que, mastegant i triturant el material, n’acceleren la descomposició. Per això, genèricament els coneixem com a animals trituradors. Uns dels organismes més característics dels nostres rius que podem incloure dins d’aquesta categoria són els tricòpters, que fan les seves casetes amb petits trossets de pedres o restes de troncs. Com un exemple característic, podem citar Potamophylax, abundant a les muntanyes de Prades o a la riera de Balà, o Allogamus auricollis, molt característic dels rius pirinencs. Aquests tricòpters, que en alguns indrets del Principat reben el nom de terrendosos, són molt apreciats pels pescadors com a esquer. La seva abundància és condicionada a l’entrada i la presència d’abundant matèria orgànica grollera al riu.
Una observació més acurada del nostre riu a la seva capçalera en llocs de poc corrent ens permet veure gran quantitat de material acumulat en el fons. Si filtrem aquest material amb un sedàs d’1 mm de porus, ens queda retingut el material groller que aprofiten els trituradors; però no sols s’ha filtrat aigua, sinó també moltes petites partícules orgàniques (restes de fulles, en aquest cas). Si fem passar l’aigua resultant de la filtració primera per un filtre més fi, de 0,45 µm, aquest es tenyeix de color fosc, perquè reté el material fi i només deixa passar el matèria orgànica dissolta (MOD). Tenim, doncs, una fracció de matèria particulada orgànica fina (MOPF) que ha estat generada per l’actuació dels animals trituradors. Alguns animals es poden alimentar d’aquests materials que queden acumulats en el fons dels rius. Per fer-ho hauran de tenir algun sistema per a recollir-lo, com una mena de raspall, per exemple. Els animals que presenten aquesta estratègia alimentària els anomenem recol·lectors. Com més abundant sigui el material fi, com més zones de poc corrent hi hagi, més possibilitats hi ha que els animals recol·lectors hi siguin abundants. Entre els organismes recol·lectors que són més característics, podem citar els ostràcodes i els insectes plecòpters, que recullen entre els pèls de les seves potes els materials del fons del riu; el plecòpter Protonemura en seria un exemple.
Si augmenta la quantitat d’aigua en el riu, per la pluja o pel desglaç, per exemple, en augmentar la velocitat de baixada de l’aigua, una part del material orgànic acumulat pot ser arrossegada, especialment aquella en què el material és més fi. Aquest material, que queda ara en suspensió, o bé pot sedimentar-se més avall o bé pot ser retingut per filtres en el riu, físics (barreres de branquillons o fulles) o deguts a l’acció dels organismes. Efectivament, alguns animals poden realitzar aquesta filtració retenint la matèria orgànica particulada fina amb finalitats tròfiques. Per això en certs organismes trobem algunes parts del cos modificades per a la filtració, com les mandíbules de Fefemeròpter Ephoron virgo, abundant a la part baixa del riu Ebre. D’altres animals són capaços de fabricar els seus propis filtres mitjançant diferents tipus de secrecions sedoses. Tots aquests reben el nom de filtradors. Les larves dels dípters simúlids o les del tricòpter Hydropsyche, també en són exemples típics. Aquests darrers són relativament ben coneguts en l’àmbit dels Països Catalans i fins i tot sabem que els filtres que fan tenen una llum de 230 × 360 µm i presenten una notable precisió i regularitat. Naturalment, l’abundància d’animals filtradors és en relació amb la quantitat de material fi que hi ha en suspensió a l’aigua, sigui quin sigui el seu origen.
La xarxa tròfica originada a partir de la producció primària

Fragment de l’epilíton que recobreix una pedra del riu Ges al seu pas per Vidrà (juliol de 1983), vist al microscopi electrònic d’escandallatge. Hi ha materials orgànics i inorgànics, i algunes espècies d’algues: la cianofícia Schizothrix affinis (fílaments prims que travessen la imatge), la clorofícia Spirogyra (a baix a la dreta) i diverses diatomees (Navicula, al mig a l’esquerra, i Gomphonema, al mig a la dreta). Aquesta comunitat constitueix l’aliment dels animals brostejadors del riu.
Sergi Sabater/ SME Univ. Barcelona
Si el riu és desproveït de vegetació a les vores o la conca és desforestada, la penetració de la llum pot originar la presència més o menys abundant de plantes dins del riu. Això depèn també de la quantitat de nutrients de l’aigua, una característica que es relaciona amb la conca o el punt del riu on ens trobem. Les petites diatomees, les algues filamentoses, les molses o les fanerògames aquàtiques poden ser abundants segons les característiques del substrat del riu (pedres, grava o sorra) i dels nutrients que conté l’aigua.
Moltes vegades les algues es presenten com una capa fina d’uns pocs milímetres que recobreix les pedres, sobretot a les zones de més corrent, on han d’enganxar-se fortament al substrat per no ser arrossegades. En realitat, aquesta capa és un complex on són abundants alhora bacteris i algues, inclosos tots en una matriu de polisacàrids complexa anomenada epilíton. Aquesta cobertura funciona com una màquina d’absorbir no solament nutrients inorgànics (com nitrats o fosfats), sinó també matèria orgànica dissolta (MOD), i pot suportar una biomassa considerable d’organismes que pasturen pel seu damunt. Precisament de la capacitat de pasturar d’aquests organismes, n’ha derivat el nom de brostejadors. Els seus aparells bucals presenten adaptacions per raspar l’epilíton del damunt de les pedres o els epífits de les plantes aquàtiques. Un representant genuí en seria el mol·lusc pulmonat Ancylus fluviatilis, ben conegut per la seva forma de barret frigi característica, però també molts insectes aquàtics, com els dípters blefarocèrids i alguns efemeròpters (Ecdyonurus, Baetis) o tricòpters (Synagapetus). La presència abundant de brostejadors indica la importància del procés de fixació d’energia solar mitjançant l’epilíton. Una gran abundància de brostejadors pot, fins i tot, arribar a limitar el creixement de les algues, i, de fet, el controlen. Els briòfits i les fanerògames aquàtiques també poden ser rosegats per aquests organismes.
En aquests ambients on predominen els vegetals aquàtics també hem de considerar la importància del corrent. Així, una part de les diatomees o de l’epilíton pot ser arrossegada fàcilment i aquest és l’origen també d’una part de la matèria orgànica particulada fina, que pot mantenir-se en suspensió i afavorir la presència d’animals filtradors o bé pot acumularse en llocs de poc corrent, on l’aprofiten els recol·lectors.
A les parts més baixes dels rius trobem una quantitat considerable de potamoplàncton, perquè l’amplada, el cabal del riu i el corrent en permeten la proliferació. Aquests organismes, pel fet d’estar en suspensió a l’aigua, passen a formar part del material orgànic particulat i ofereixen encara més possibilitats de desenvolupament als filtradors. La gran proliferació de potamoplàncton a la part baixa de l’Ebre és l’origen de la gran abundància de les palometes (Ephoron virgo) en aquesta part del riu.
És també a les parts mitjanes i baixes dels rius on poden desenvolupar-se millor els macròfits, és a dir plantes aquàtiques que viuen totalment o parcialment submergides dins de l’aigua. Les condicions perquè es desenvolupin són la presència d’un substrat de sorres o fang, un corrent moderat o fort però no turbulent i una certa quantitat de nutrients a l’aigua. La majoria dels macròfits són fanerògames (Myriophyllum, Ceratophyllum, Potamogeton), però algunes algues (com per exemple Cladophora) també poden trobars’hi formant grans masses, en aquestes condicions. Els nutrients que afavoreixen el creixement dels macròfits poden ser derivats del rentatge de la conca. Per tant, com més gran sigui la conca, més forta serà la tendència a acumular-ne, cap a les parts baixes, de manera que hi haurà un creixement més gran dels macròfits a les parts baixes dels rius, gràcies a les característiques del substrat, molt fi, i a les concentracions de nutrients.
L’acció de l’home sobre els ecosistemes fluvials es caracteritza per la seva aportació de nutrients a l’aigua (eutrofització). Per això és fàcil que a les parts baixes dels nostres rius hi hagi grans proliferacions de macròfits, com Potamogeton pectinatus i d’altres. A les parts mitjanes del Llobregat i a la part baixa del Ter se’n poden trobar masses considerables. Els macròfits poden suportar una variada fauna a base de brostejadors, o de vegades encara més especialitzada, de tipus xuclador. Aquests darrers són animals amb les parts bucals convenientment modificades per xuclar els líquids de les plantes. Malgrat tot, no és una adaptació que es pugui trobar fàcilment a l’aigua, on la majoria de macròfits són explotats per brostejadors o herbívors que els masteguen.
Els depredadors
Els depredadors són el conjunt d’animals que es mengen els altres animals. Hem vist fins ara que hi havia a bastament representats en els rius els dos grups d’organismes que obtenen el seu aliment o bé de la producció primària (la xarxa dels herbívors) o bé dels materials al·lòctons detrítics (que podríem anomenar xarxa dels descomponedors). Tota aquesta biomassa animal és a disposició d’aquells altres animals que siguin capaços d’aprofitar-la, que són els depredadors.
Els depredadors que podem trobar als rius són de tots els grups zoològics, des de petits unicel·lulars que es mengen altres unicel·lulars, fins als més grans, com la llúdria, avui escassa als nostres rius. Una gran part dels petits depredadors són insectes de tots els ordres. Les grans perles (Perla, Dinocras) o, entre els tricòpters, els del gènere Rhyacophila, en són els més característics. Però segurament els depredadors més típics dels rius són els peixos, la majoria dels quals s’alimenten d’aquests petits animals, encara que alguns, com la carpa, són més aviat omnívors. Cal esmentar els grans depredadors, com el llop de riu (Esox lucius), i d’altres, ben apreciats pels pescadors. Els depredadors en els rius han despertat molt d’interès en aquest darrers anys. De ser considerats gairebé com un element poc important dins l’ecosistema fluvial, han passat a ser considerats, en molts casos, com uns veritables controladors de la dinàmica de les poblacions de certes espècies. A més dels peixos i alguns insectes aquàtics, cal tenir en compte el paper dels crancs de riu.
Variació de l’estructura de la comunitat en l’espai

Cap a les parts central i final del riu els substrats sorrencs i fangosos, la velocitat moderada del corrent i l’elevat contingut de nutrients de l’aigua i/o els sediments poden donar lloc al desenvolupament d’una biomassa considerable de macròfits. Els macròfits són el suport de molts organismes animals que els brostegen o masteguen, o de molts filtradors que els utilitzen només com a substrat de fixació. En la fotografia, feta al riu Cardener després del pas per Manresa al novembre de 1979, hom pot intuir que el riu, en aquest tram d’1 m de fondària, és reblert, en tota la seva amplada, per una fanerògama aquàtica: Potamogeton pectinatus; aquest creixement tan abundant es deu a la seva capacitat de resistir la salinitat que el riu té en aquest tram (conseqüència de l’aportació de sals de les mines de Súria i Cardona) i a la seva gran velocitat de creixement en aigües ben proveïdes de nutrients (en aquest cas, aportats per la ciutat de Manresa). P. pectinatus és una planta poc valorada pels animals i pels epífits, per la qual cosa pràcticament només s’instal·len en les seves branquetes petits brostejadors (que mengen més aviat els materials que s’hi acumulen i els epífits), com el gasteròpode Potamopyrgus jenkinsii, o filtradors, com Hydropsyche exocellata, un tricòpter que teixeix unes xarxes de filtració finíssimes.
Narcís Prat
A cada segment del riu podem observar l’organització de la comunitat en relació amb la manera com es produeix el flux d’energia. Aquest depèn de les característiques de la conca i les seves modificacions; és per això que podem emprar la comunitat com a indicadora del que passa a la conca. Però en molts dels nostres rius el que trobem és un mosaic considerable de condicions en sectors relativament petits i, per tant, una comunitat molt diversa d’organismes. En un espai reduït pot haver-hi totes les estratègies que hem definit fins ara.
L’estudi de la comunitat que vivia al riu Llobregat, fet a l’alçada de Navars (Bages) l’agost de 1982, pot ser en aquest sentit molt aclaridor. Si, en aquesta zona, i de banda a banda del riu, en un transsecte perpendicular, agafem una mostra (recol·lecció de tots els organismes de més de 250 µm que hi ha en un quadrat de 20 × 20 cm) cada 2 m aproximadament, ens trobem que cada mostra és un xic diferent de les altres: en unes zones dominen certs substrats, i en d’altres, certs organismes. Les condicions particulars de substrat i de corrent, a més de les derivades de possibles interaccions entre els organismes (depredació, herbívors, competència, etc.), fan que hi hagi fins 100 espècies diferents en un espai reduït, de no gaire més d’un metre quadrat. En realitat, més que una sola comunitat, tenim un mosaic de comunitats. En uns llocs se sedimenten materials i per tant tenim diferents tipus de recol·lectors; en d’altres llocs hi ha moltes pedres recobertes d’abundant epilíton i al seu damunt la presència abundant dels brostejadors corresponents; hi ha també una zona amb macròfits, on la composició de la fauna és influïda pel substrat fi sobre del qual es troben; en tots els llocs amb corrent hi ha abundants filtradors. En un espai relativament reduït respecte a la dimensió longitudinal del riu se’ns manifesta tota la diversitat de la vida en aquests ambients. En diferents llocs, però que es troben ben pròxims, una estratègia d’alimentació pot dominar sobre les altres, segons quines siguin les condicions del riu.
Aquesta organització a petita escala té un reflex a una escala més gran: tota la conca. En efecte, hem dit que els rius s’organitzen jeràrquicament, de les parts altes a les parts baixes. Els materials grollers de les capçaleres es transformen en materials fins o nutrients dissolts. Si examinem a grans trets les característiques del riu al llarg del seu eix, veurem que és possible definir en diferents trams algun tipus de comunitat, precisament per una dominància més forta de certs processos d’organització. A la part alta d’un riu és molt més probable que la conca hi sigui forestada i que en el riu, els processos derivats de l’entrada de material al·lòcton hi siguin més importants que no pas la producció primària; per tant, sembla que l’epilíton hi ha de ser poc important, i entre els animals han de dominar els trituradors. A les parts baixes en canvi, els materials fins i els nutrients en dissolució fan que l’organització de la comunitat sigui diferent, amb un domini dels animals filtradors i del potamoplàncton. Aquest fet i les diferències de règim tèrmic i la vivor de les aigües fan que l’estudi i l’observació del riu en la seva organització tot al llarg de l’eix principal sigui un bon model ecològic de referència.
El riu com a sistema ecològic
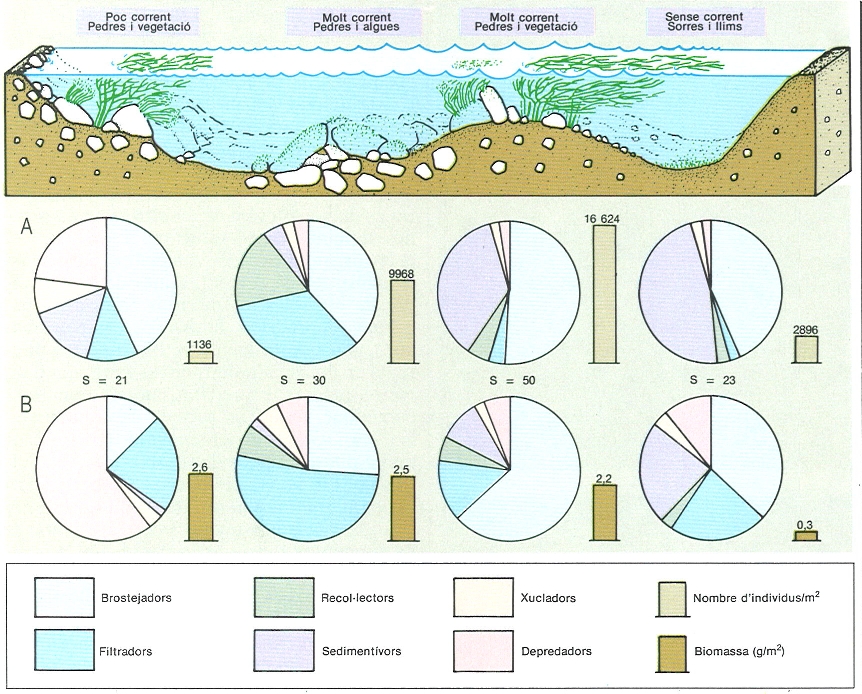
Heterogeneïtat espacial dels rius, en un exemple pres d’un tram del riu Llobregat, en el qual es va efectuar un transsecte de banda a banda, amb 4 punts de mostreig, a l’agost del 1982. Hom hi ha representat la densitat d’organismes (A) relativa (sectors) i absoluta (barres), la biomassa (B), també relativa i absoluta i el nombre d’espècies (S). A la banda esquerra, en una zona de poc corrent i de substrat dur, hi ha poca densitat, poques espècies i les sangoneres predadores hi són dominants, per la qual cosa la biomassa hi és elevada. Més cap al centre, amb més corrent i amb un substrat dur cobert d’algues, la dominancia es reparteix entre brostejadors (Baetis) i filtradors (Hydropsyche), tant pel que fa al nombre com a la biomassa; els tricòpters són més grans que els efemeròpters brostejadors i per això la seva biomassa és més important. En el tram de substrat fi, hi arrelen abundants potamogètons i algues, i la dominancia en nombre i en biomassa és dels brostejadors i dels sedimentívors (cucs, de biomassa individual petita). A la part dreta, el riu és encalmat i el substrat llimós, cobert d’algues; hi dominen els sedimentívors (cucs, quironòmids) i els brostejadors (quironòmids petits), de biomassa individual molt petita (els pocs filtradors, tricòpters, que hi ha en conjunt sumen una biomassa semblant); els baixos valors de densitat, biomassa i diversitat d’espècies és en relació amb el poc corrent.
Maria Rieradevall, original de l’autor
L’eix horitzontal dels rius no és pas igual a l’eix vertical dels llacs i els embassaments. El model més senzill és considerar el riu com un continu funcional que va des de les parts altes fins a les més baixes, transformant els materials que hi entren o s’hi fabriquen. Com que l’aigua no pot anar cap amunt, tota la informació generada a les parts altes pot anar cap a les parts baixes, però no a l’inrevés. Només els animals capaços de remuntar el corrent (peixos, fase adulta dels insectes) poden retornar aigües amunt. En aquest sentit són els únics que poden «explotar» uns ambients on és relativament fàcil trobar aliment.
Aquest model prediu que al llarg del riu canvien les condicions: des de la prevalença dels materials particulats grollers i un flux d’energia generat pels materials externs, a les parts altes, es passa per la dominància dels processos lligats a la producció primària de l’epilíton, a les parts mitjanes, fins que s’arriba a una gran importància dels materials fins en suspensió en el riu i un potamoplàncton abundant, a les parts més baixes. Això origina, també, un canvi en les comunitats: hi ha dominància d’animals trituradors a les parts altes; una importància més gran dels brostejadors a les parts mitjanes; i una dominància total dels filtradors a les parts més baixes. Entre els productors primaris, els briòfits són propis de les capçaleres de grau menor i a les parts més baixes hi ha les fanerògames aquàtiques, mentre que a les parts mitjanes són importants les diatomees, i totes les altres algues capaçes d’enganxar-se a les pedres. Això es pot expressar segons el grau del riu i en relació amb la temperatura, ja que aquests paràmetres també canvien gradualment de dalt a baix del riu.

L’heterogeneïtat en l’espai i la complexitat biològica que hi havia al riu Llobregat a l’altura de Navars (Bages) a l’agost del 1982 (a dalt) contrasta fortament amb l’aspecte que prengué uns anys més tard, després d’una riuada (a baix). Els rius són, doncs, uns sistemes extremament actius i canviants. El creixement de masses de macròfits que s’inicia després d’una riuada portarà a augmentar l’heterogeneïtat del medi i la diversitat de les comunitats, si la magnitud i la durada de la pròxima riuada és petita. Però si la pertorbació és molt forta, es trencarà de nou l’organització del riu. Les pertorbacions fortes són comunes als rius mediterranis. Moltes espècies hi estan adaptades i la comunitat es regenera força ràpidament, a diferència del que passa en els processos de successió biològica en altres sistemes, com per exemple els boscos.
Narcís Prat / Maria Rieradevall
La realitat mostra que en els rius situats en zones amb poca alteració humana aquest model és variable, tot i que es manté una certa tendència al que prediu la teoria. D’altra banda, cal tenir en compte que les intervencions humanes poden canviar molt les coses. Aquest aspecte és cabdal en la interpretació de l’ecologia dels rius dels Països Catalans, on gairebé no n’hi ha cap que es pugui considerar que té un estat proper al que seria un riu natural. En realitat, cada riu és un mosaic de zones, de vegades ben properes, on predominen uns o altres processos, o fins i tot de zones dins les quals hi ha fragments amb característiques ben diferents, com hem vist en el cas del Llobregat. El que trobem al llarg de l’eix principal del riu, en realitat, és un mosaic de mosaics. Aquest punt de vista sembla el més adequat per a un sistema tan extraordinàriament dinàmic com és el riu. Els rius dels Països Catalans s’acosten molt més al model del mosaic que no al del continu, tant des del punt de vista natural (una bona part dels nostres rius són temporals i això imposa una restricció important en el procés d’organització de la comunitat) com des del punt de vista de la importància de la intervenció humana, que provoca discontinuïtats molt grans, com la presència d’embassaments o la contaminació.
Per a comprendre aquest mosaic complex hem de preguntar-nos quins són els factors que hi intervenen. A més d’estudiar com es genera el flux d’energia, hi ha altres factors que cal tenir en compte: diferències de pendent, de substrat, de velocitat de l’aigua, de cabal o de les característiques físico-químiques de les aigües, entre rius que s’assemblen per altres característiques de la conca (vegetació, insolació) poden donar una estructuració i composició de les comunitats ben diferents. Aquests aspectes diferencials semblen accentuats en les conques mediterrànies i hom esperaria, fins i tot, un gradient N-S en els Països Catalans, bé que fins ara no podem demostrar-ho. Finalment, un altre aspecte influeix en aquest immens mosaic de mosaics. Hi ha una variació temporal (superposada a tota aquesta variació en l’espai) que és doble: d’una banda, el cicle anual mediterrani amb el fort contrast estiu-hivern més o menys matisat per l’altitud; d’altra banda, les variacions interanuals, a les quals ja ens hem referit quan hem parlat de cabals. Així, moltes de les nostres comunitats es poden interpretar d’acord amb una succesió ecològica doble, una a curt termini (anual) i una altra a un termini mitjà, com per exemple el temps que hi ha entre dues avingudes catastròfiques.
Les comunitats fluvials

Comunitats característiques dels cursos alt, mitjà i baix dels principals rius catalans. Hi dominen gairebé sempre els animals filtradors (groc) i els brostejadors (taronja), mentre que els trituradors són poc importants. Pel que fa als productors primaris (verd), en el curs alt hem indicat les comunitats de substrat silici (1-5) i calcari (6-7); només en el curs baix dels rius més grans es pot parlar d’un potamoplàncton (36-40). Pel que fa als predadors (rosa), els més importants són els peixos, sobretot a les parts baixes, on, a més d’omnívors o fitòfags (49-51), poden haver-n’hi d’ictiòfags (52-53). 1 Gomphonema angustata, 2 Achnanthes minutissima. 3 Cymbella ventricosa, 4 Pleurocapsa, 5 Cratoneuron, 6 Hygroamblystegium, 7 Rhynchostegium riparoides, 8 Hydropsyche instabilis, 9 Simulium, 10 Rheotanytarsus, 11 Epeorus, 12 Ecdyonurus, 13 Paraorthocladius, 14 Oligoneurella, 15 Perla, 16 Rhyacophila, 17 truita de riu (Salmo trutta), 18 Amphora pediculum, 19 Gomphonema variegatum, 20 Navicula, 21 Diatoma elontatum, 22 Cladophora, 23 Myriophyllum, 24 Potamogeton crispus, 25 P. nodosus, 26 Hydropsyche pellucidula, 27 Cheumatopsyche, 28 Psychomyia pusilla, 29 Ancylus, 30 Orthocladius, 31 Baetis, 32 Elmis, 33 Dina lineata, 34 Rhyacophila, 35 Barbus barbus, 36 Biddulphia laevis, 37 Cyclotella, 38 Scenedesmus, 39 Thalassiosira, 40 Pediastrum simplex, 41 Cladophora, 42 Ceratophyllum, 43 Potamogeton pectinatus, 44 Hydropsyche exocellata, 45 Ephoron virgo, 46 Theodoxus fluviatilis, 47 Melanopsis, 48 Cardiocladius, 49 barb caní (Blennius fluviatilis), 50 gobi (Gobius gobius, 51 madrilla (Chondrostoma toxostoma), 52 lluç de riu (Esox lucius), 53 perca americana (Micropterus salmoides) MOPF matèria orgànica particulada fina. La gruixària de les fletxes indica la importància del flux entre compartiments.
Maria Rieradevall, original de l’autor.
Ja hem comentat que les comunitats d’organismes que habiten un tram determinat del riu depenen essencialment dels processos que es donen a la seva conca juntament amb les característiques del mateix riu: substrat, velocitat del corrent i dinàmica temporal. Tot això fa que el riu sigui més un mosaic d’ambients diversos que no pas un espai uniforme.
L’estudi de la composició, la dinàmica temporal i la importància relativa de les comunitats d’organismes en els rius dels Països Catalans és relativament recent. Per això, no és estrany que el coneixement de les comunitats sigui encara fragmentari: els estudis funcionals (producció primària i secundària), com també les relacions entre els components de la comunitat, tant des del punt de vista tròfic (depredació) com d’altres interaccions relacionades (competència, cicles de vida), són gairebé inexistents. D’altra banda, l’estudi descriptiu de les comunitats d’organismes ha perdut la transcendència que tenia fa uns anys, potser perquè les agrupacions d’espècies que es poden fer són degudes a moltes causes i la interpretació en resulta difícil. De tota manera, la composició i estructura de la comunitat té l’interès de proporcionar-nos una visió sintètica de la conca molt millor que la de les anàlisis químiques.
Com podem definir i caracteritzar una comunitat determinada? La definició més senzilla és acceptar que un conjunt d’organismes que coincideixen en el mateix espai formen una comunitat on el grau d’interacció entre els seus membres és molt variable. Aquesta és una definició pràctica, ja que ens permet utilitzar les recol·leccions d’organismes que fem en una zona determinada com a representatius de la comunitat que hi viu.
D’acord amb les característiques de la seva conca i la posició en la xarxa hidrogràfica podem trobar diferents «estils» de comunitats en el conjunt dels Països Catalans, tot i que les espècies variïn. Amb les dades de què fins ara disposem hem fet alguns grups que podrien representar les comunitats tipus dins de les nostres contrades. Primer distingirem les comunitats pròpies dels rius de capçalera de conca, siguin pirinencs o no; d’altra banda, les comunitats pròpies del tram mitjà dels rius més grans (Llobregat, Ter) i les de les parts més baixes (Ebre). Al llarg de l’eix del riu haurem de distingir aquelles comunitats pròpies dels afluents amb molt menys cabal que el riu principal al qual van a parar. Finalment considerarem el cas dels rius mediterranis de recorregut curt i poc cabal, com el Foix o el Besòs. En tots els casos intentarem donar una guia de la funcionalitat de la comunitat.
Les comunitats dels rius de capçalera de conca

En els rius de capçalera (rius petits, de grau 1, 2 o, com a màxim, 3), on el bosc cobreix fins i tot el llit del riu, la comunitat que hi viu pot dependre en gran part o totalment del flux de materials al·lòctons grollers que hi arriben, especialment les fulles dels arbres caducifolis. A la riera de Balà (Osona), que reprodueix la fotografia, l’abundància de faigs fa que l’aigua corri pel mig de grans masses de fulles, que són arrossegades, triturades i descompostes per l’acció física i química de les aigües i pels animals trituradors, que hi són molt abundants. Ací, per exemple, dominen els tricòpters del gènere Potamophylax, que fabriquen casetes amb petits grans de matèria mineral (bé que, en llocs de menys corrent, fins i tot a la mateixa conca, les fan amb trossets de fulles).
Narcís Prat / Maria Rieradevall
Aquestes comunitats corresponen a la part més alta dels nostres rius, on podríem considerar que neixen tant els rius principals com els seus afluents. Són, per tant, rius de grau 1 o 2. Solen ser de fort pendent (generalment més del 10%) i, per tant, les aigües són ràpides. Les comunitats de productors primaris, com també les de macroinvertebrats i peixos, són molt similars quant a adaptacions i a composició, ja que els organismes estan adaptats especialment al corrent fort.
Els factors de variabilitat que poden influir en la seva composició són el cabal, la composició química o l’altitud en què s’origina el riu. Així hi ha certes diferències entre un riu que neix als Pirineus i un que neix a la Serralada Litoral o Prelitoral; o, també, n’hi ha entre el Cardener o el Llobregat, que neixen amb un doll d’aigua important. D’altra banda, sabem que hi ha una dependència de la manera com s’estructura la comunitat respecte del flux d’energia segons que sigui molt dependent de l’entrada de materials de la conca o hi domini la producció primària. Així, podem tenir en llocs d’altitud, clima, pendent i característiques físico-químiques similars comunitats molt diferents. Si la conca és forestada, dominen les comunitats que depenen del material al·lòcton, amb pocs productors primaris (majoritàriament molses) i animals trituradors. Si la conca no és forestada, les comunitats s’organitzen entorn del flux d’energia originat en la producció primària. Per això, dins de la uniformitat hi ha una variació important entre els nostres rius de muntanya, tot i que des del punt de vista morfomètric i físico-químic poden ser similars.
D’una banda tenim els rius pirinencs, que a les parts més altes es caracteritzen per la temperatura baixa de l’aigua tot l’any (mitjanes de 7-8°C o menys), el fort pendent (15-30%), la feble mineralització (20-100 µS/cm) i la bona oxigenació. Dins d’aquesta aparent uniformitat podem trobar comunitats ben diferents, com les dominades pels simúlids (diverses espècies de Prosimulium) i pel quironòmid Pseudodiamesa, que són propis de les sortides dels llacs de muntanya, com el Redó, situat a més de 2000 m. L’abundància d’aquests organismes es deu als seus hàbits filtradors, que aprofiten els materials produïts en el llac (fitoplàncton) que aquest exporta. En certa manera són comunitats molt similars a les que es poden trobar a la sortida d’alguns embassaments, encara que amb una composició específica molt diferent.
En molts llocs els rius freds i ben oxigenats circulen a través de boscos més o menys ben formats. En aquests casos els animals estenotèrmics són abundants (plecòpters, especialment), però numèricament la comunitat és dominada per animals de tipus triturador. A Andorra, i a les capçaleres del Ter i del Freser, aquests són representats especialment pels tricòpters Allogamus auricollis i Drusus discolor, ben característics i identificables pels seus estoigs fets per mitjà de petits grans de sorra o pedres.

Els materials calcaris, molt abundants al nostre país, faciliten la formació de fonts i surgències naturals d’aigua, fins al punt que aquestes són un element comú del nostre paisatge. Les aigües amb abundància de calci sorgeixen amb cabals i temperatures relativament constants. La seva riquesa relativa de nutrients permet el creixement de comunitats de l’epilíton i molses característiques (Cratoneuron commutatum, per exemple), al damunt de les quals es desenvolupa una comunitat que, gràcies a l’estabilitat de les condicions físiques i químiques, és molt similar al llarg de l’any i té elements característics, bé que difereix entre una font i una altra. A la fotografia veiem les fonts del Cardener, al vessant oriental de la serra del Port de Compte, que en són un exemple característic, com podrien ser-ho també les fonts del Llobregat (Berguedà) o les deus de Sant Quintí de Mediona (Alt Penedès).
Joaquim Reberté
Curiosament, en altres rius de capçalera de conca, sempre en llocs de considerable altitud i d’aigües fredes, hi és present una comunitat similar, però on l’espècie dominant és una altra. En efecte, a les capçaleres dels torrents de les Guilleries (riera de Balà) o de les muntanyes de Prades les fulles són també l’element bàsic per a fer funcionar les comunitats, però els elements dominants, que també són tricòpters amb estoig, són del gènere Potamophylax. El mateix es presenta a les rieres de la part alta de la serra de Tramuntana de Mallorca. En tots aquests casos, els acompanyen abundants plecòpters, especialment dels gèneres Nemura i Protonemura que també eren importants als Pirineus i són presents de forma abundant a la capçalera de les rieres del Montseny. Aquests plecòpters també estan lligats a la presència de materials en descomposició, encara que es podrien caracteritzar millor com a recol·lectors de material que obtenen pel fet de viure enmig de les fulles en descomposició.
En canvi resulten molt diferents els rius de capçaleres calcàries, especialment quan s’originen en surgències importants. Tot i que la temperatura mitjana hi pot ser igualment baixa, hi ha una quantitat de sals més gran en dissolució que permet la presència de més nutrients en solució i el creixement de més productors primaris. Així, dominen els brostejadors, especialment els blefarocèrids (Liponeura cinerarescens), alguns efemeròpters, especialment Baetis alpinus, tricòpters del gènere Agapetus, quironòmids (Diamesa i Breoheptagia) i mol·luscs (Ancylastrum fluviatile, Bythinella).
Les comunitats de productors primaris (de les quals s’alimenten els brostejadors esmentats) en aquestes aigües fredes de muntanya són molt similars, independentment de les característiques físiques i químiques de les aigües. Són comunitats d’epilíton, gairebé exclusivament sense macròfits o, en tot cas, molses i hepàtiques aquàtiques. Així, tant al Ter com al Llobregat, les comunitats són dominades per diatomees (Ceratoneis arcus, Meridion circulare, Diatoma hiemale), amb dominància a la primavera d’altres algues que poden formar grans masses, com Hydrurus vulgaris, que recobreix totes les capçaleres dels rius pirinencs en aquella època. Les quantitats de clorofil·la que poden desenvolupar-se en aquestes aigües netes són més aviat minses, amb una mitjana propera als 65 mg/m2.
Les aigües calcàries del Llobregat permeten que hi siguin presents algunes diatomees (Diatoma elongatum o Gomphonema olivaceum) que no trobem en els rius granítics. Aquestes diferències també són clares entre les molses. En els rius poc mineralitzats dominen Hygrohypnum i Philonotis entre les molses, i també són comunes algunes hepàtiques exclusivament aquàtiques, entre les quals assenyalarem Scapania undulata i Chiloscyphus polyanthus, mentre que als rius calcaris hi ha Cratoneuron commutatum i Barbula ehrenbergii.
Els depredadors també són abundants en aquestes aigües i molts atenyen dimensions considerables. En molts rius mancats de poblacions de peixos, els depredadors poden ser exclusivament invertebrats (insectes sobretot) o petits vertebrats, com els tritons. La fracció de depredadors és formada en els rius de muntanya principalment per grans plecòpters (Perla, Dinocras, Isoperla) o tricòpters (Rhyacophila), encara que també hi podem trobar alguns no artròpodes ben característics, com els cucs plans del gènere Polycelis (P. felina, P. nigra). En el cas que hi hagi peixos, exclusivament hi trobem truites.
Les comunitats dels rius de muntanya

Comunitats característiques dels rius de muntanya. Els rius de muntanya proporcionen una gran varietat d’ambients, segons les característiques físiques i químiques de l’aigua i la seva dependència de la conca adjacent. En el cas de l’aigua de sortida de l’estany Redó, la riquesa d’elements planctònics (1 Fraguaría crotonensis, 2 Cyclotella comta, 3 Cryptomonas, 4 Chromulina, 5 Mallomonas) afavoreix la dominància dels filtradors (17 Simulium). A la capçalera del Ter, en aigües poc mineralitzades de zones poc forestades (per damunt del límit superior altitudinal de la vegetació arbòria), les pedres es recobreixen d’algues abundants (6 Meridion circulare, 7 Diatoma vulgare, 8 Ceratoneis arcus, 9 Hydrurus foetidus), que afavoreixen la prèsencia de diferents brostejadors (18 Diamesa, 19 Baetis alpinus, 20 Boreoheptagia). L’exemple del riu d’Andorra és un cas típic de riu de zona de caducifolis, les fulles dels quals es descomponen per l’acció de fongs i bacteris (10); a la comunitat dominen els trituradors (21 Amphinemura, 22 Potamophylax, 23 Drusus, 24 Allogamus). L’aigua de les Fonts del Llobregat és característica pel seu contingut alt de calci, que dóna lloc a l’abundància de molses i algues característiques (11 Ceratoneis arcus, 12 Gomphonema acuminatum, 13 Nitzschia pungens, 14 Cocconeis placentula, 15 Barbula unguiculata, 16 Cratoneuron commutatum) i a una gran varietat de brostejadors (25 Micrasema, 26 Liponeura, 27 Baetis, 28 Synagapetus). Els depredadors són gairebé els mateixos en tots els casos (29 Polycelis, 30 Perla, 31 Isoperla, 32 Rhyacophila, 33 Salmo trutta), bé que varia la dominància relativa d’un sobre els altres en cada cas.
Maria Rieradevall, original de l’autor
Els rius més grans dels Països Catalans (Llobregat, Ter, Segre, Noguera Pallaresa i Ribagorçana), pel fet d’originar-se als Pirineus o als Prepirineus, ja tenen una certa importància quan encara travessen zones de condicions de muntanya. És a dir, les aigües continuen descendint per un pendent fort, fredes la major part de l’any; però ja no són petits rierols sinó que transporten un cabal elevat provinent de la unió de diferents torrents o rierols. Això fa que la dependència de la conca immediata ja no sigui tan gran i que l’aigua tingui més nutrients i, també, arrossegui una considerable quantitat de materials en suspensió. En aquestes aigües desapareixen gairebé totalment els trituradors, i es fan més abundants els brostejadors i els filtradors.
A les nostres aigües de muntanya destaca la presència del tricòpter Hydropsyche instabilis, que és un dels organismes més comuns quan l’aigua transporta una quantitat considerable de materials en suspensió. També són comuns entre els filtradors els simúlids, especialment a la primavera (Simulium monticola, S. cryophilum), com també alguns quironòmids del gènere Rheotanytarsus, que formen tubs característics. Entre els brostejadors d’aquests rius cal esmentar l’abundància de certs efemeròpters en els rius de conques calcàries, cosa que no es dóna en els de conca silícia, com Oligoneurella rhenana; però també és el lloc on es troben en gran abundància els efemeròpters més grans i buscats per la seva qualitat d’esquer de pesca, com els dels gèneres Epeorus, Ecdyonurus o Rhithrogena, i molts quironòmids que proliferen al damunt de l’abundant epilíton en totes les èpoques (Parorthocladius nudipennis, diverses espècies d’Eukiefferiella).
Les comunitats d’algues que formen l’epilíton són dominades per diatomees, especialment Achnanthes minutissima, Cymbella ventricosa i Gomphonema angustatum, i també d’altres algues incrustants, com Pleurocapsa; també és important la cobertura de Cladophora, especialment a l’estiu o quan els nutrients són abundants. Aquestes comunitats poden formar recobriments relativament importants a les pedres, amb uns valors mitjans al voltant dels 112 mg/m2 de clorofil·la. Els macròfits continuen representats gairebé exclusivament per molses, que en els rius de conca silícia acostumen a ser Rynchostegium riparioides i Hygroamblystegium, i en els calcaris, espècies del gènere Cratoneuron.
Depredadors importants en aquestes zones continuen essent els plecòpters com Dinocras cephalotes o Perla, i els tricòpters (diferents espècies de Rhyacophila). Però en aquests ambients el depredador que domina i de vegades controla la comunitat és la truita, que és justament en aquests trams on millor es desenvolupa.
Les comunitats del tram mitjà
El tipus de funcionament, més independent dels ecosistemes propers i amb més dependència de la producció pròpia, derivada dels materials que provenen de la conca transformats aigües amunt, és el tret més característic de la part mitjana dels rius. És important, doncs, el paper dels vegetals (productors primaris) i dels animals que hi viuen associats (brostejadors), i també el dels filtradors, ja que molts dels materials provinents de la part alta del riu són en forma particulada. En aquest tram de riu, la temperatura de l’aigua és molt menys variable i no arriba als valors hivernals que caracteritzen els rius de muntanya; per això, deixa de ser un factor clau. El desenvolupament de certes espècies es pot realitzar gairebé al llarg de tot l’any. Són zones amb més sals, més conductivitat i més nutrients, i també molt alterades per l’acció humana, que hi intervé de diverses formes (aportació de nutrients, contaminació per aigües negres, canalitzacions, canvis en els cabals per la presència d’embassaments, contaminació salina, extraccions d’aigua o d’àrids etc.). Les comunitats que avui tenim als Països Catalans en aquests trams de riu depenen la majoria d’aquests canvis.
Els productors primaris s’hi caracteritzen per l’abundosa cobertura algal (primavera i estiu sobretot), feta de multitud de diatomees, que gairebé sempre viuen a l’epilíton o com a epífites de Cladophora glomerata. En les zones de substrat no rocós, hi trobem abundants macròfits; encara que les molses hi poden ser presents (Cinclidotus i Fissidens, sobretot), hi dominen algunes fanerògames (Potamogetón nodosus, P. crispus, Myriophyllum verticillatum). Les quantitats de clorofil·la poden arribar fins a 300 mg/m2.
Entre aquesta abundant cobertura vegetal hi trobem molts brostejadors, dels quals destaquem la variadíssima comunitat de quironòmids (de vegades, 20 espècies en una sola pedra) i efemeròpters (Baetis rhodani i B. vardarensis, com els més representatius), i els mol·luscs (Ancylastrum fluviatile). Barrejats amb aquests, a les zones de més corrent, hi trobem una munió de filtradors: simúlids quan hi ha a la pedra espais lliures o sobre els macròfits, o tricòpters (Hydropsyche pellucidula, H. exocellata, Psychomyia pusilla o Chematopsyche lepida). Els depredadors són també alguns insectes (Rhyacophila evoluta, Rh. dorsalis) o sangoneres, especialment Dina lineata. També els peixos poden ser importants (barbs, carpes, gòbids, bagres). En aquest darrer cas una de les comunitats més interessant és la que hom ha estudiat al Matarranya.
La contaminació canvia d’una manera característica aquesta comunitat. Per exemple, al Llobregat, la contaminació salina fa que el macròfit dominant absolut sigui Potamogeton pectinatus, mentre que les diatomees més abundoses també canvien, a més d’alguns insectes. Per exemple, les dues espècies de Baetis que hem citat més amunt són substituïdes per B. meridionalis i B. nigrescens, i entre els quironòmids hi ha certes espècies que dominen, com Cardiocladius fuscus. Canvis similars es donen a les parts mitjanes del Ter, en aquest cas per causa del funcionament dels embassaments de Sau i Susqueda.
Les comunitats del tram inferior

Els peixos solen ocupar la part superior de la xarxa tròfica als rius. La truita, a muntanya, i la carpa, a les parts baixes, potser en són els elements més populars. Però el nombre d’espècies és molt més elevat, com mostren estudis recents fets al Matarranya, per exemple (on és feta la fotografia), o a les parts baixes de l’Ebre. Mengen tant els organismes que viuen a la superfície de les pedres com els materials que el riu arrossega (deriva), i també, en el cas dels superdepredadors, altres peixos.
Adolf de Sostoa
La part baixa dels rius més grans dels Països Catalans es troba molt degradada per causa de la contaminació, o bé perquè és gairebé seca per l’ús que hom fa de les seves aigües més amunt. En aquestes condicions, les comunitats estan molt empobrides. Si hi resta una mica d’oxigen i hi ha transparència, podem trobar-hi algun macròfit (Potamogeton pectinatus) i petites cobertures de diatomees gairebé monospecífiques (com la diatomea Nitzschia ganderheimensis), entre les quals poden viure alguns quironòmids, especialment del gènere Cricotopus. Si hi falta l’oxigen, els macroinvertebrats gairebé desapareixen o es redueixen a espècies resistents a la pol·lució (cucs tubifícids i larves d’alguns dípters, com Eristalis o Chironomus).
La part baixa de l’Ebre conserva, però, una qualitat acceptable de les aigües, alterada només per la regulació que els embassaments fan del riu. La gran quantitat de nutrients fa que en aquestes aigües hi pugui créixer una notable quantitat d’algues en suspensió formant part del potamoplàncton (valors de clorofil·la de fins 30 µg/l). D’altra banda, a les pedres, hi pot proliferar un epilíton abundant, amb grans masses de l’alga Cladophora. En aquestes condicions apareix una comunitat molt típica, amb molts filtradors i brostejadors. A les zones amb pedres dominen com a filtradors els tricòpters (Hydropsyche exocellata) i a les zones amb sorres, l’efemeròpter Ephoron virgo, ben conegut a Tortosa i a tota la zona baixa de l’Ebre per les seves eclosions en massa. Com a brostejadors, destaca la presència abundant dels mol·luscs, sobretot espècies de Melanopsis i Theodoxus fluviatilis. És una comunitat particular i interessant, que ens fa veure el que podia haver estat la part baixa dels nostres rius fa uns quants anys. La comunitat de peixos en els rius grans és molt variada i complexa, amb espècies molt característiques. A l’Ebre, n’hi ha fins a 37 espècies diferents, onze de les quals són sedentàries, mentre que les altres són marines que poden invadir-lo temporalment.
Quan el riu s’acosta a la mar pot reduir la velocitat i alhora rebre la influència salina. En aquests casos pot desenvolupar-se un veritable potamoplàncton, que manté en suspensió espècies pròpies de llocs salats (Thalassiosira weissflogii). Al Ter, aquestes comunitats assoleixen valors elevats de biomassa, amb concentracions de clorofil·la de fins 247 mg/m2. En aquestes zones són comuns els macròfits. En el cas de l’Ebre, pot desenvolupar-se una ploma salina fins a 25 km de la seva desembocadura, quan el cabal del riu es redueix per sota dels 50 m3/s (especialment a l’estiu). Llavors la part baixa del riu es transforma en un veritable llac on l’aigúa dolça s’escola lentament per sobre l’aigua salada. La poca transparència de l’aigua i la gran quantitat de nutrients fan que la producció primària sigui elevada. Al fons d’aquesta zona (6-7 m) la anòxia pot ser total.
Les comunitats dels afluents de les parts mitjana i baixa

Els afluents de molts dels rius catalans corren per terrenys calcaris. L’aigua hi té un fort contingut de calci, per la qual cosa els materials submergits sovint es recobreixen d’una capa d’incrustació de carbonats. Això fa que les comunitats algals i animals hi siguin peculiars. El torrent de la fotografia és un exemple d’aquest tipus d’ambients, alguns dels quals poden arribar a assecar-se a l’estiu o convertir-se en una successió de basses. També són medis extraordinàriament diversos, que mantenen sovint una biomassa considerable.
Narcís Prat / Maria Rieradevall
A la part mitjana i baixa, els rius principals dels Països Catalans (de la Tet, al nord, fins al Xúquer, al sud) reben una sèrie de cursos d’aigua de cabals i característiques molt diferents. En tots aquests afluents, les condicions del medi hi són menys extremes que a les capçaleres de conca, especialment pel que fa a la temperatura. El règim de cabals hi és típicament mediterrani, per la qual cosa, la dinàmica temporal és molt important. La longitud d’aquests rius és també molt variable. Tot i que és difícil de fer comparacions, se’n pot fer un resum atenent les característiques relacionades amb la mineralització.

Molts dels afluents dels nostres rius situats a la zona de clima mediterrani prenen, en algunes èpoques de l’any, les característiques d’una successió de basses més que no pas d’un curs d’aigua. En aquestes condicions, que a la fotografia es veuen representades pel curs alt del Ripoll, afluent del Besòs, el riu es colonitza d’algues (Spirogyra, Zygnema) al damunt de les quals es desenvolupa una comunitat semblant a la de les basses temporànies. Moltes d’aquestes basses actuen com un refugi per a les espècies sense capacitat migratòria, que són pròpiament del riu, o per als alevins de peixos.
Narcís Prat
Els afluents del Ter i del Besòs que provenen del Montseny tenen una mineralització relativament feble comparada amb la dels rius on van a parar. Aquesta característica fa que els puguem diferenciar de la majoria dels rius catalans que neixen a la zona prelitoral i que normalment van molt carregats de carbonats o de sulfats. Aquests afluents de conca silícica i de grau 2, 3 o 4 presenten una cobertura algal feble, normalment amb valors relativament baixos de clorofil·la (al voltant dels 80 mg/m2), composta de diatomees i Cladophora fracta, que es fa més abundant com més nutrients porta l’aigua. L’estructura de la comunitat de macroinvertebrats està molt relacionada amb l’estructura de la conca (més o menys forestada); però s’assembla molt a les dels rius pirinencs de grau 2-4, amb els grans efemeròpters (Epeorus, Ecdyonurus, Rithrogena) com a elements característics.
En canvi, els afluents calcaris presenten característiques notables, especialment si hi ha una certa quantitat de sulfat, que els apropa als coneguts «chalk-streams» anglesos. La disponibilitat de nutrients és més gran en aquestes comunitats, per la qual cosa és fàcil trobar valors de concentració de clorofil·la elevats (una mitjana de 200 mg/m2). La major part d’aquesta biomassa és formada per algues incrustants, com Gongrosira, Calothrix, Rivularia, Schizothrix o Phormidium, sobre de les quals es desenvolupa una gran quantitat de diatomees epífites. També s’hi pot trobar la comunitat de briòfits dominada per Cratoneuron. Una gran munió de macroinvertebrats viu entre aquests organismes, amb alguns elements característics i exclusius d’aquest tipus de rius. Entre els efemeròpters és de destacar la presència de Thraulus bellus i Choroterpes picteti, exclusius d’aquests ambients. Tot i això, la comunitat és extraordinàriament diversa, amb animals brostejadors com a dominants (quatre espècies de Baetis, Echinogammarus del grup berilloni, Ancylastrum fluviatile i una munió de quironòmids). Dels filtradors cal esmentar Hydropsyche pellucidula i H. siltalai i alguns simúlids.
Les comunitats dels rius mediterranis

Comunitats i dels rius de zona calcària, dels rius típicament mediterranis que poden assecar-se fent basses a l’estiu, i dels rius contaminats i amb molta matèria orgànica. En el primer cas, les comunitats són riques tant en productors primaris (verd) i brostejadors (taronja) com en filtradors (groc) i depredadors (rosa). La biota dels rius mediterranis és canviant: a l’hivern i la primavera és típicament rivícola, i a l’estiu, limnícola. La comunitat rivícola és formada per espècies oportunistes de brostejadors i filtradors que s’alimenten dels productors primaris; la limnícola, per algues zignemals o carofícies i macroinvertebrats, sobretot brostejadors o recol·lectors; entre els depredadors, també poden haver-hi amfibis, insectes i peixos. Els rius contaminats tenen una flora i una fauna simplificada i adaptada a resistir els tòxics, l’abundant matèria orgànica i les baixes concentracions d’oxigen. 1 Achnanthes minutissima, 2 Diatoma elongatum, 3 Calothrix (a aspecte general de la colònia, b detall dels tricomes), 4 Rivularia, 5 Phormidium autumnale, 6 Gongrosira, 7 Hydropsyche siltalai, 8 Simulium, 9 Micropsectra atrofasciata. 10 Baetis. 11 Caenis, 12 Choroterpes, 13 Habrophlebia, 14 Thraulus, 15 Gammarus, 16 Eukiefferiella, 17 Ancylus, 18 góbit (Gobius gobius), 19 madrilla (Chondrostoma toxostoma), 20 barb (Barbus barbus), 21 Cymbella helvetica, 22 Achnanthes minutissima, 23 Gomphonema, 24 Amphora, 25 Diatoma elongatum, 26 Navicula, 27 Cladophora, 28 Hydropsyche siltalai, 29 Agapetus, 30 Ephemerella, 31 Elmis, 32 Simulium, 33 Baetis, 34 Spirogyra, 35 Zygnema, 36 Chara, 37 Psectrocladius, 38 ostràcodes 39 àcars, 40 Caenis, 41 Cloeon dipterum, 42 zigòpter, 43 carpa (Cyprinus carpio) 44 Sphaerotilus, 45 Nitzschia, 46 tubifícid, 47 Chironomus, 48 Physa, 49 Eristalis. MOPF matèria orgànica particulada fina.
Maria Rieradevall, original de l’autor
La majoria dels nostres rius tenen un règim típicament mediterrani. (Molts dels considerats en l’apartat anterior es podrien incloure aquí.) Però també alguns dels nostres rius s’originen a la Serralada Prelitoral i van directament a la mar. Tenen un recorregut curt i de vegades assoleixen només graus de 2-3. Les condicions ambientals poden ser molt extremes, i la probabilitat que la llera quedi reduïda a una bassa és molt gran; el Foix o el Besòs en serien uns exemples ben característics. Això fa que la comunitat d’organismes que s’hi troba sigui poc especialitzada i que la majoria d’espècies siguin elements ja coneguts de comunitats anteriors. L’organització de la comunitat és interrompuda gairebé cada any per les riuades o les secades, per la qual cosa s’hi poden trobar espècies amb mecanismes resistents a l’arrossegament o a l’assecament. D’altra banda han de ser, també, espècies de creixement ràpid, que puguin aprofitar els moments favorables, que de vegades són només dos o tres mesos.
En aquestes condicions les comunitats d’algues són formades gairebé sempre per una munió de diatomees, i en els casos d’una certa riquesa de nutrients, per Cladophora. A la zona de les capçaleres la netedat de l’aigua, les temperatures més baixes i la sequera estiuenca fan que certs organismes coneguts per les seves adaptacions a aquests factors en siguin elements característics, com els plecòpters Capnioneura mitis o Eulectra geniculata o l’efemeròpter Baetis muticus intermedius. Però la comunitat és formada sobretot per elements d’àmplia dispersió en tots els rius que són afluents dels més grans, sobretot quironòmids i simúlids, amb la presència sempiterna de Baetis rhodani i Ancylastrum fluviatile. No sembla que hi sigui gaire important l’estratègia alimentària, però hi trobem abundants brostejadors, com els citats juntament amb Agapetus i èlmids, i, en certs casos, filtradors (simúlids i, com a tricòpter característic, Hydropsyche siltalai). Si el riu és molt calcari, s’hi poden trobar, també, Thraulus bellus i Choroterpes picteti.
A l’estiu, és fàcil seguir la successió biològica en aquests rius, ja que la llera sol convertirse en un seguit de basses on la conductivitat i la temperatura solen incrementar-se i les comunitats, tant d’algues com de macroinvertebrats, s’adapten a les noves condicions. Sempre s’hi troben efemeròpters (Cloeon inscriptum i Caenis luctuosa), i hi apareixen larves d’odonat i de ditíscids i altres escarbats. Les comunitats evolucionen cap a les característiques de les basses.