Característiques adaptatives al vol
Els ocells són vertebrats homeoterms (és a dir, que la seva temperatura interna no varia amb la del medi) de temperatura elevada i de reproducció ovípara, proveïts de plomes i amb els membres anteriors transformats en ales, cosa que porta a un bipedisme obligat.
Aquests caràcters i molts d’altres poden explicar-se a la llum d’un objectiu: adquirir la facultat de volar. Altrament, un primer cop d’ull general al món dels ocells, ens el mostra àmpliament diversificat, tant pel que fa als colors i les dimensions com per la forma de les potes, el bec o les ales.
Podem examinar els trets que caracteritzen els ocells sota el mateix prisma amb què ho farien els enginyers aeronàutics. Quins són els materials, els paràmetres i les característiques que utilitza l’home quan dissenya avions? Hi ha tres coses primordials i obligades: lleugeresa, potència i perfil aerodinàmic. Els ocells les han aconseguides primordialment mitjançant els trets evolutius de les plomes i les ales, la pneumatització dels ossos, la sang calenta, un sistema remarcable de respiració, uns músculs pectorals molt potents i un cor molt gran i fort que batega molt ràpid.
Les adaptacions anatòmiques
Per aconseguir la lleugeresa, al llarg de l’evolució ha tingut lloc una reducció dels òrgans i s’han suprimit particularitats anatòmiques i fisiològiques que podrien afegir un pes superflu.
La reducció dels òrgans reproductors n’és un bon exemple. Llevat del kiwi i d’algunes rapinyaires, en què són funcionals ambdós ovaris, a la resta d’ocells en funciona un de sol (l’altre és vestigial) i els òrgans sexuals s’atrofien durant l’època no reproductora. El que això pot representar per a l’animal ho pot expressar, per exemple, el fet que, a l’estornell, durant la reproducció, aquests òrgans pesen 150 vegades més que a la resta de l’any. Paral·lelament, en els ocells s’han reduït quasi del tot les glàndules de la pell, de les quals solament resten les uropigials (i no en tots els ocells), situades a la base de la cua.
L’oviparisme o reproducció per ous també és una bona adaptació al vol: la reducció de pes és evident, si ho comparem amb el viviparisme dels mamífers. Una constatació de la lleugeresa dels ocells ens ve donada per la seva densitat específica, que en el cas d’un ànec és de 0,6, mentre que en la major part dels mamífers volta l’1.
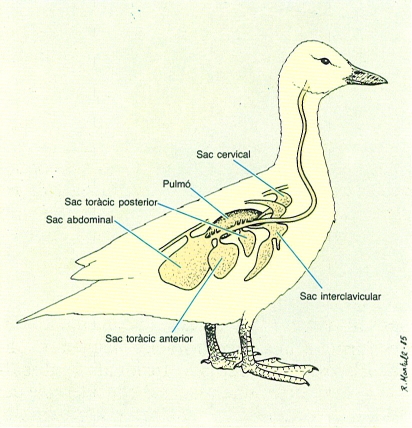
Els sacs aeris constitueixen un sistema de ramificacions múltiples i independents originades als bronquis, que es fiquen per entre les vísceres i a dins dels ossos de l’esquelet. Participen en la reducció de la densitat del cos, en el manteniment de la temperatura corporal i en l’aireig dels pulmons.
S.F.F.B. / Román Montull.
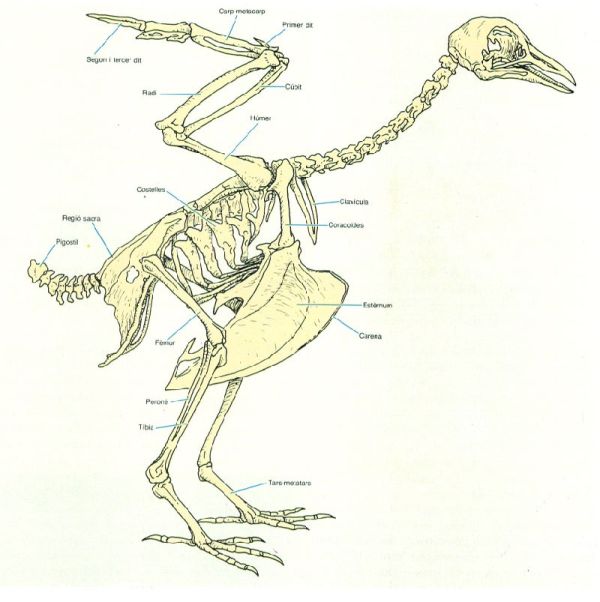
L’esquelet dels ocells correspon al model general a tots els vertebrats, però amb algunes particularitats en relació amb l’adequació al vol, que afecten principalment les cintures pectoral i pelviana i les extremitats; altrament, la forma de les costelles confereix una especial solidesa a la caixa toràcica; la clavícula i el procoracoides mantenen el pit eixamplat durant el vol; l’estèrnum presenta una carena on s’insereixen els potents músculs que accionen el vol. A la cintura pelviana, la regió sacra constitueix el punt d’inserció dels músculs que mantenen el cos dret sobre les potes. A les ales, l’húmer rep la inserció dels músculs del vol; el radi i el cúbit formen la part extensible i més visible de l’ala; el primer dit suporta l’àlula; el segon i el tercer porten les rèmiges primàries.
S.F.F.B. / Román Montull.
L’esquelet dels ocells és molt fort, elàstic i lleuger, en una bona part gràcies a la seva pneumatització, que consisteix en la presència de cambres d’aire en lloc del moll d’alguns ossos. Per reforçar l’estructura buida, algunes espècies presenten trabècules internes i obliqües en forma de V. El grau de pneumatització varia segons els grups; els cabussadors en tenen molt poca i excepcionalment, algun grup molt volador com el de les gavines, no en presenta. La forma dels ossos pectorals s’entén perquè, en el vol, la pressió s’exerceix sobre les ales i aquestes descansen sobre el pit; per això, aquests ossos són grans, particularment l’estèrnum, que presenta una forma carenada molt acusada típica dels ocells. Com més voladora és l’espècie, més gran i carenat és l’estèrnum, damunt del qual s’implanten els poderosos músculs pectorals, impulsors de les ales; la seva forma, alhora, ajuda a l’aerodinamisme del cos. La llarga cua dels rèptils (els avantpassats dels ocells) queda reduïda al mínim: la formen unes poques vèrtebres caudals, que en la major part de les espècies es troben soldades formant el pigostil. Llevat de les vèrtebres del coll, que permeten una gran mobilitat al cap, la resta de vèrtebres són anquilosades o semianquilosades, i constitueixen un conjunt gairebé indivís. Les costelles són primes i aplanades, i s’acoblen a les veïnes formant una caixa toràcica lleugera i sòlida. Els ossos del cap són fusionats i, globalment, comporten una reducció considerable del pes. Entre dos animals de talla comparable, com són el colom i la rata, el cap de l’ocell pesa una sisena part del del rosegador. L’absència de dents, de la barra mandibular que els sosté i dels músculs trituradors, juga un paper important en l’estalvi de pes. Aquestes peces queden substituïdes pel bec que, majorment, és buit per dins. La funció trituradora, que demana músculs poderosos i, per tant, pesants, davalla al pedrer o estómac triturador (element de l’aparell digestiu típic dels ocells), que se situa prop del centre de gravetat de l’animal. Per ajudar a triturar els aliments, els ocells s’empassen sovint petites pedretes que actuen com les rodes de molí. A l’esquelet del cap també hi ha dues grans concavitats que allotgen els ulls, els més desenvolupats i perfectes de tots els animals. La visió primordialment, i en un grau menor, l’audició, són els sentits més desenvolupats dels ocells.
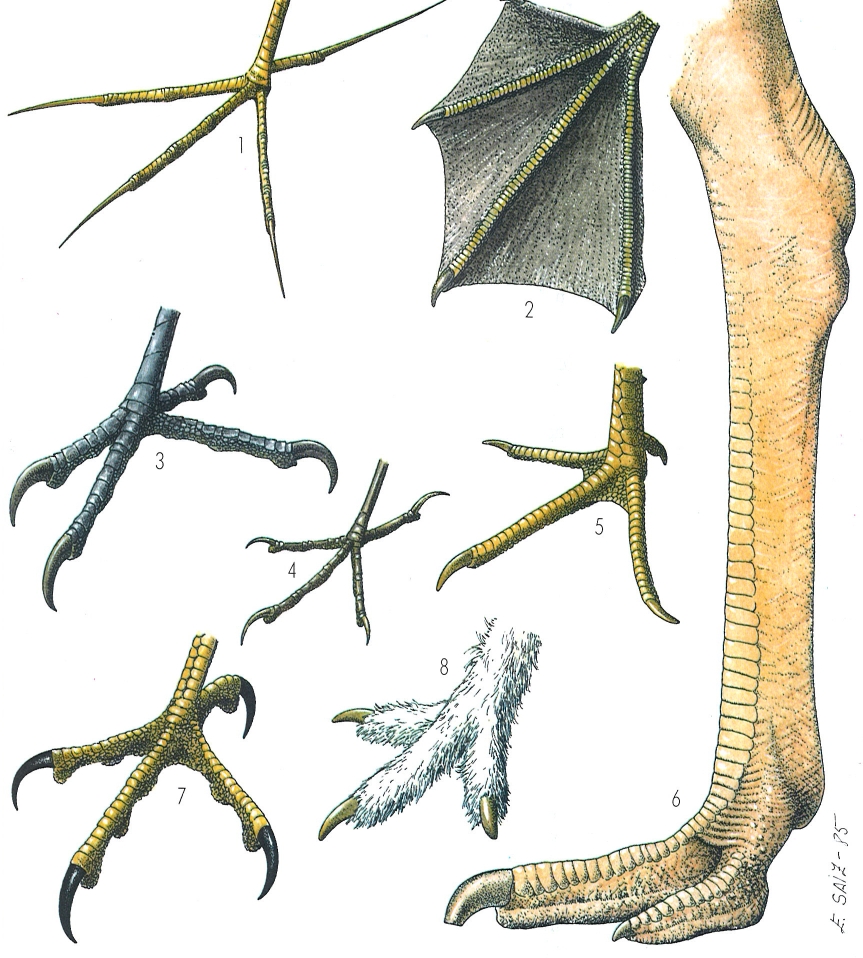
La morfologia de les potes dels ocells reflecteix les característiques dels seus hàbitats i els seus costums. Les potes de dits llargs, prims i ben oberts permeten a la jacana (1 Jacana jacana) caminar pel damunt de les plantes surants a l’aigua. Les del mascarell (2 Sula bassana) reflecteixen la seva capacitat nedadora. En el picot verd (3 Picus viridis), la posició dels dits, permet que l’ocell es subjecti als troncs dels arbres per un mecanisme semblant al dels moixons com el pit-roig (4 Erithacus rubecula), que té els dits disposats fent com una mena de pinça amb la qual es pengen dels branquillons. Les potes del faisà (5 Phasianus colchicus) permeten de caminar per terra i també parar-se als troncs dels arbres. Les robustes potes de l’estruç (6 Struthio camelus) són un exemple del tipus de potes dels ocells caminadors de les estepes. Les potes dels ocells de presa com les del xoriguer (7 Falco tinnunculus), tenen una morfologia distintiva ben clara; són fortes i amb les ungles esmolades i corbades per a subjectar les preses. Les potes de la perdiu blanca (8 Lagopus mutus), amb els tarsos emplomallats, donen idea de l’aïllament tèrmic que necessiten aquests ocells pera viure a l’estatge nival de les muntanyes.
Eduardo Saiz.
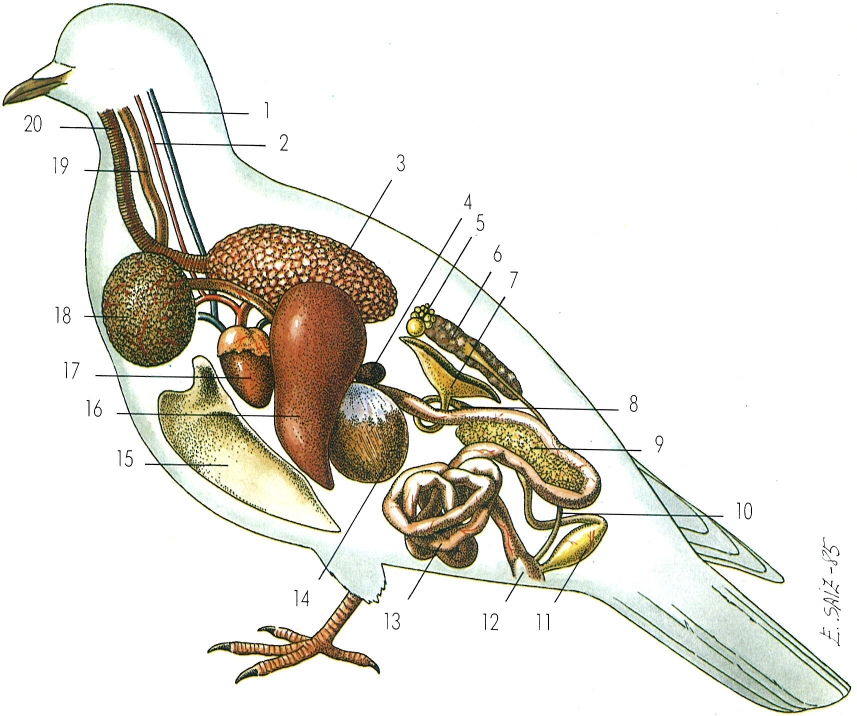
Anatomia interna dels ocells. 1 lugular, 2 caròtida, 3 pulmó, 4 melsa, 5 ovari, 6 ronyó, 7 infuldíbul, 8 oviducte, 9 pàncrees, 10 urèter, 11 glàndula de la closca, 12 cloaca, 13 budells, 14 pedrer, 15 estèrnum, 16 fetge, 17 cor, 18 pap, 19 esòfag, 20 tràquea.
Eduardo Saiz.
L’organització de la musculatura tendeix a concentrar al sector central i ventral la major part d’elements (entre els paquets musculars del pit i de les cuixes hi ha el centre de gravetat de l’animal) i a desplaçar cap a les parts perifèriques (ales, potes) tendons prims i forts. Així, els músculs pectorals, molt poderosos, són els agents del vol batut, i en les espècies molt voladores —colibrís i alguns colúmbids— poden gairebé arribar al 40% del pes de l’animal. Els albatros i altres planejadors tenen menys musculatura central i més longitud de lligaments i tendons prims. La simple presència d’abundant múscul pectoral no indica necessàriament una bona aptitud voladora, ja que per a volar sostingudament cal que el múscul sigui molt ben irrigat; això se’ns manifesta pel color de la carn, que és més vermella com més irrigació té. El pollastre o el gall dindi són una bona prova domèstica de la relació entre la carn blanca i la ineptitud per al vol: com a màxim, fan voladetes ràpides i curtes.
Una característica important dels ocells és la presència de sacs aeris. Connectats als ossos pneumatitzats i als pulmons, que són relativament petits, els ocells disposen, en general, de nou o deu bosses limitades per membranes i distribuïdes per l’abdòmen, el tòrax i les zones interclavicular i cervical. Aquests sacs contribueixen a donar lleugeresa a l’animal, alhora que fan més eficient la funció dels pulmons. Però, principalment, tenen funció refrigerant, cosa que és fonamental en un organisme d’alta temperatura que, a més, desprèn una quantitat important de calor com a conseqüència del vol; per exemple, es calcula que, de l’aire emmagatzemat en els sacs del colom, més de la meitat és emprat per a refrigerar i la resta és destinat estrictament a la respiració. Als ocells aquàtics també ajuden a la flotabilidad, mentre als que es cabussen des d’una certa altura, com els xatracs o els mascarells, els protegeix el cos de l’impacte. Globalment, el sistema de respiració-ventilació d’un ocell ocupa un volum considerablement més important que el d’un mamífer: el 20% del volum total en el cas de l’ànec contra el 5% en el cas de l’home.
Les plomes
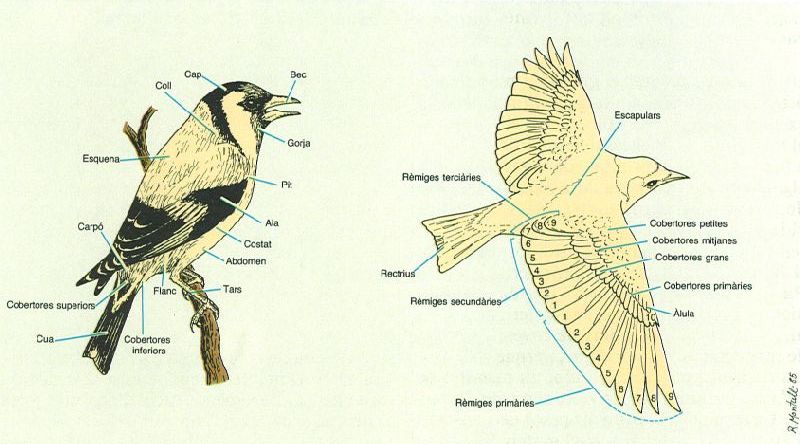
Morfologia externa i nom de les parts principals del cos dels ocells. L’enumeració de les plomes que s’indica a l’esquema de la dreta correspon a l’ordre de renovació durant la muda i també al de les modificacions hagudes en el curs evolutiu. En canvi, els grups d’anellament i alguns manuals és corrent que utilitzin l’enumeració de les rèmiges en sentit contrari (ascendent), perquè això evita la dificultat d’identificar la primera rèmige primària.
S.F.F.B./Román Montull.
La presència de plomes és, potser, el tret més típic dels ocells. Les plomes compleixen diverses funcions, de les quals les més importants són les següents: conformar la superfície de vol de les ales i la cua, crear una capa aïllant al voltant del cos, ajudar eficaçment a l’aerodinamisme del cos, mantenir el cos impermeable i, finalment, aprofitar-ne el color i la forma per a camuflar-se o per a d’altres funcions, com les del reconeixement específic, el del company sexual, per a la cohesió del grup (per exemple, mitjançant l’efecte de "flash" de taques blanques a les ales o a la cua), etc. En un animal homeoterm, són fonamentals els mecanismes que tendeixen a mantenir la temperatura del cos. Les plomes retenen prop de la pell una sèrie de capes d’aire immobilitzades, que actuen d’aïllant tèrmic. Aquestes capes es formen per la superposició de plomes, a tocar de la pell el plomissol i més amunt les cobertores. Quan fa molt fred, l’ocell s’estufa, és a dir, alça les plomes exteriors per augmentar l’espessor de la capa aïllant. L’aïllament tèrmic que comporta el plomatge és molt eficaç, tal com ho demostra el fet que els ocells siguin els vertebrats més capacitats per a aguantar temperatures extremes. Les plomes de contorn, que a grans trets són totes, llevat de les de vol (rèmiges i timoneres) i les del plomissol, són molt importants per a l’aerodinàmica de l’ocell, ja que poden fer variar el contorn de l’animal segons les circumstàncies, i evitar amb això les turbulències. Un altre element regulador de les turbulències és l’ala bastarda, petit apèndix alar que s’articula al polze i se situa i funciona igual que els alerons dels avions. Un altre fet de la morfologia externa dels ocells, relacionat amb Paerodinamisme, és l’absència d’apèndixs sobresortints de la línia general del cos: no hi ha orelles externes i la major part de les espècies, mentre volen, recullen les potes sota el plomatge ventral d’una manera semblant a com els avions recullen el tren d’aterratge. Els avantatges que representen les plomes per a la funció de volar són múltiples. N’esmentarem la duresa i la flexibilitat (no es trenquen contra un corrent d’aire fort), la lleugeresa, i el fet que siguin elements morts i proveïts d’un sistema especial d’unió entre els elements que conformen la superfície de la ploma. En efecte, el fet d’ésser inerts evita a l’ocell d’instal·lar un sistema d’irrigació sanguínia que gravaria el pes de la cua i de les ales. D’altra banda, el sistema d’unió tipus "velcro" dels barbicels que conformen la superfície de la ploma, permet que aquesta torni a la forma original ràpidament, sense esforç ni despesa, simplement amb un toc del bec, després d’alguna topada o accident. La importància de les plomes es palesa pel fet que en molts ocells el plomatge pot pesar més del doble de l’esquelet. Per exemple, l’àguila marina americana, amb 4 kg de pes, té un esquelet de 272 g i un plomatge de 677 g.
Les adaptacions fisiològiques
Sembla evident que una activitat energèticament tan alta com és la de volar impliqui necessàriament un metabolisme alt, ja que, com és sabut, l’increment de temperatura comporta una acceleració de les reaccions químiques. En els ocells, les temperatures internes oscil·len entre 38° C i quasi 45°C, les concentracions de sucre a la sang doblen aproximadament les dels mamífers, mentre que la pressió arterial és també considerablement més alta. La circulació és doble, tancada i completa i el cor és gran (proporcionalment, més que en els mamífers), poderós i amb una freqüència alta de batecs, més alta com més petit és l’ocell. Per exemple, si el cor de l’home batega 72 cops per minut, el del colom ho fa 135 vegades, el del canari 514 i el del colibrí 615. Tot plegat, un motor potent i eficient per a poder volar.
Però un motor d’aquest tipus demana un combustible que sigui altament energètic i que deixi pocs residus. Per tant, no és casualitat que no hi hagi gaires espècies d’ocells que s’alimentin de dietes de baixes calories, com per exemple de fulles i herbes, que demanen a més a més un període llarg de permanència en el tub digestiu. Tot i ser molt ampli el mostrari d’aliments que consumeixen els ocells, podem dir que, globalment, es tracta d’aliments calòrics: llavors, fruits, insectes, cucs, petits vertebrats, etc. En comparació amb els mamífers, aprofiten una fracció més gran de l’aliment, i aquesta és consumida d’una manera molt eficient i ràpida.
Un caràcter no relacionat amb el vol, forçat per la reproducció ovípara, és l’excreció d’àcid úric en lloc d’urea, cosa que suposa una despesa d’aigua molt minsa, amb les consegüents implicacions sobre l’adaptació a medis terrestres pobres en aigua.
Podríem dir, per acabar, que igual que els motors sobrerevolucionats, els ocells tenen una vida molt intensa, però curta; això ho demostra el fet que l’esperança de vida de les espècies de mida petita és inferior a dos anys, i la de les més grans, encara que alguns individus poden arribar als seixanta anys, usualment no arriba als cinc anys, perquè les mortalitats al primer any i al segon són molt fortes.
La uniformitat del pla estructural

L’adequació anatòmica dels ocells al vol és ben clara en aquest exemplar de fragata (Fregata magnificens), en el qual s’aprecia el perfil aerodinàmic del cos.
Carme Borràs.
En el món dels ocells, hi ha una relativa uniformitat externa, que esdevé rigorosament implacable en estudiar-ne l’estructura interna, car el model estructural que els permet volar té unes exigències molt estrictes, que permeten poques variacions.
Una comparació amb el grup dels mamífers és ben il·lustrativa d’aquesta uniformitat. Per exemple, si bé és cert que hi ha ocells adaptats al medi marí, com el cas dels pingüins, aquesta adaptació mai no és total, de manera que, en algun moment (a l’època de la reproducció) han de tornar a terra; per contra, els mamífers de vida marina, com els dofins i les balenes, passen tota la vida a la mar, i fins i tot, per a ells, el fet de tornar a terra els representa la mort. Passa el mateix en el cas de la vida subterrània: alguns ocells, com el sud-americà Steatornis, viuen tot l’any a dins de coves, però han de sortir a la nit per a menjar, perquè s’alimenten de fruits; en canvi, el talp, un mamífer de vida subterrània, no pot viure si no és sota terra. La comparació entre els pesos dels animals d’ambdós grups reflecteix el mateix fenomen.
Tot i que hi ha molta diferència entre el pes de l’ocell més gros (l’estruç, de 150 kg) i el del més menut (un colibrí cubà, de 2,5 g), la diferència entre els extrems corresponents al grup dels mamífers (la balena blava i la musaranya nana) és mil vegades més gran. Pel que fa a la forma i les característiques externes del cos, els mamífers són clarament més diversificats: en trobem de molt peluts, com el visó, i de glabres, com la rata del desert, alguns d’allargassats, com la mostela, i d’altres d’arrodonits, com l’elefant, etc.
Examinem, ara, el cas de la talla. Els ocells més grans són els no voladors, els quals, en tot el món, probablement no ultrapassen la cinquantena d’espècies. Dins dels voladors, l’energia necessària per a volar imposa unes dimensions límit: si doblem les dimensions lineals del cos d’un ocell, el pes o el volum s’incrementa al cub, mentre que la superfície ho fa al quadrat. Per tant, si tenim dos ocells amb les mateixes proporcions relatives, i l’un és el doble de llarg que l’altre, el gran tindrà quatre vegades més superfície alar, però, alhora, pesarà vuit vegades més que el petit. Amb aquesta regla s’entén que s’arribi relativament ràpidament a un límit superior, ja que l’ocell gran, per a volar amb la mateixa facilitat que el menut, ha d’impulsar-se més, és a dir, ha d’incrementar el seu volum muscular, la qual cosa implica alhora una sobrecàrrega, etc. A la pràctica s’observa que aquest límit superior és similar en diversos grups d’ocells voladors i es troba al voltant dels 13,5-20 kg. Això no obstant, coincideix a diverses famílies que els exemplars més grans s’escapen una mica de la tirania d’aquesta regla geomètrica, gràcies al vol a vela, que els permet aprofitar una energia suplementària (tèrmiques, crestes de les onades, etc.); en són exemples les cigonyes, les àguiles, els voltors, els albatros, les grues, etc., molts dels quals migren pels estrets, defugint tant com poden la travessa de les masses d’aigües obertes, on és obligat l’ús de l’energèticament costós vol batut. L’exemple de les migracions en V de grues, oques o pelicans és un altre recurs dels ocells grans per a estalviar energia de vol. La presència d’un límit superior en la mida dels ocells és paral·lela al límit que s’imposa en els avions que funcionen amb motor d’explosió, i que l’home ha superat canviant totalment la concepció del motor, és a dir, amb el motor a reacció, invent que encara no han posat a punt els ocells.
En els ocells menuts observarem que com més petit és l’animal, més superfície té en relació al seu volum i més fàcilment perd la calor del seu cos. En conseqüència, ocells diminuts com els colibrís, desprenen una gran part de l’energia solament en el manteniment de la seva temperatura interna. La supervivència d’aquestes joies es deu a l’explotació d’un aliment altament calòric (el nèctar de les flors), la possibilitat d’esdevenir letàrgics durant la nit i en els períodes meteorològicament desfavorables (amb la qual cosa estalvien molta energia), i al fet que, majoritàriament, viuen a les zones més càlides del món, o en el cas de viure a les regions fredes, les exploten solament durant l’estiu.
Les estratègies alimentàries i la morfologia

El bec dels ocells és de forma variada en relació als hàbits alimentaris de cada espècie. En els insectívors pot ser llarg i punxegut, com en l’abellerol (1 Merops apiaster) i el raspinell comú (2 Certhia brachydactyla) o bé rabassut i ample, com en l’oreneta (3 Hirundo rustica). El dels granívors és curt i robust, com en el durbec (4 Coccothraustes coccothraustes), el trencapinyes (5 Loxia curvirostra) i la perdiu roja (6 Alectoris rufa) Els piscívors solen tenir-lo llarg i punxegut, com el blauet (7 Alcedo athis) o el cabussó emplomallat (8 Podiceps cristatus) serrat o acabat en ganxo, com en el corb marí gros (9 Phalacrocorax carbo) i el gavot (10 Alca torda). En els frugívors pot adoptar aspectes ben diferents, com en el tudó (11 Columba palumbus), el turac (12 Turaco livingstonii) o el tucà (13 Ramphastos toco). Els ocells depredadors tenen becs potents i corbats, com el falcó pelegrí (14 Falco peregrinus), l’òliba (15 Tyto alba) o el capsigrany (16 Lanius senator). Els filtradors tenen un sistema de làmines que actuen com un filtre, com el cas de l’ànec cullerot (17 Anas clypeata) i el flamenc (18 Phoenicopterus ruber), encara que poden no tenir-ne, com el bec d’alena (19 Recurvirostra avosetta). El bec dels ocells omnívors és de formes variades, en relació amb la poca especificitat del seu règim; és il·lustratiu comparar el bec de la garsa (20 Pica pica) amb el del pardal (21 Passer domesticus) i el de l’estornell vulgar (22 Sturnus vulgaris) Hi ha altres tipus de bec, encara més especialitzats com el del colibrí (23 Amazilia sp.), amb el qual extreu el nèctar de les flors, el de la garsa de mar (24 Haematopus ostralegus), amb el qual obre hàbilment els mol·luscs bivalbs i el del picot negre (25 Dryocopus martius) —complementat amb una llarguíssima llengua que s’envolta dins del crani—, adequat per a foradar les soques de pins vells.
Eduardo Saiz.
Una de les característiques més evidents dels ocells és la seva gran diversitat pel que fa als colors del plomatge i al tipus d’alimentació; entre els ocells hi ha tota una gamma de règims alimentaris, des dels que mengen de tot (omnívors) fins als especialistes més estrictes. Les adaptacions alimentàries bàsiques van lligades a la velocitat i al mètode de locomoció, al grau d’agudesa dels òrgans sensorials, a inclinacions fisiològiques, a la morfologia del bec i de la llengua, a la forma i a la potència dels peus, al tipus de plomatge, al període d’activitat (diürna o nocturna), a la tolerància fisiològica a paràmetres ambientals (temperatura, humitat, etc.), i al llindar de tolerància a certs ingredients del menjar, com toxines, àcids, sals o partícules indigeribles. De totes aquestes, les adaptacions externes més importants —el bec, els peus i, en un grau menor, les ales— ajuden a imaginar com i de què s’alimenta un ocell. Però, encara que sovint aquests signes externs poden indicar dietes selectives (podríem endevinar el règim alimentari d’una espècie simplement observant-ne el bec), la realitat és que a les zones temperades hi ha moltes excepcions, i els especialistes estrictes són escassos.
Els ocells de les regions temperades tenen, en general, una dieta variable al llarg de l’any. En aquestes regions se solen produir més granes que fruits (pensem simplement en les grans extensions dels cultius cerealistes). Això fa que hi hagi més ocells que s’alimenten de gra (granívors) que no pas de fruits (frugívors), encara que dins dels moixons (ordre dels passeriformes) hi hagi tantes espècies d’una mena com de l’altra. Són relativament pocs els granívors que, com el trencapinyes, s’alimenten de granes tot l’any (aquesta espècie en concret, de pinyons); per exemple, els fringíl·lids (pinçà, cadernera, gafarró, etc.), els emberízids (sits, cruixidell, etc.), els passèrids (pardals), els remízids (teixidors) i els estríldids (vídues, becs de corall, etc.), durant la primavera, i parcialment a l’estiu, pugen els polls amb tants o més insectes que granes. També tenen tendència a alimentar els pollets de les niades més tardaneres amb una proporció més alta de granes que no les primeres llocades. Les raons per les quals donen insectes a les cries no són encara prou ben conegudes, però probablement es relacionan amb la seva riquesa en determinades proteïnes necessàries per al creixement animal; d’altra banda, els insectes, al contrari de les granes, contenen grans quantitats d’aigua, element imprescindible per al creixement del poll.
Els ocells granívors exploten i consumeixen la futura descendència de les plantes. A causa d’això, les plantes, al llarg de la seva evolució, han desenvolupat mecanismes diversos dedicats a dificultar l’extracció de les granes per part dels animals; d’una banda, les llavors van protegides amb closques dures; de l’altra, els arbres i els arbusts produeixen les granes en un temps curt i d’una manera sincronitzada, amb la qual cosa els animals disposen d’un període curt de sobreabundància d’aliment i d’un període llarg (la resta de l’any) d’escassetat. Les adaptacions dels animals, al seu torn, giren entorn del principi d’emmagatzemar la collita per poder passar el mal temps hivernal. Així, les llavors es conserven bé durant períodes de temps llargs, i permeten al gaig, al picot garser gros o al trencanous, explotar durant l’hivern el seu rebost de grans, pinyons, etc. Altres ocells més petits, com les mallerengues o el picasoques blau, guarden dins les fenedures de l’escorça dels arbres petites llavors, que es menjaran posteriorment. Aquesta activitat d’emmagatzematge de llavors ocupa una bona part de la tardor i l’estiu de moltes espècies granívores.
El cas de les espècies frugívores és ben diferent. Per a les plantes que fan fruits sucosos, els ocells constitueixen un dels principals agents dispersadors, ja que encara que el fruit és comestible, les llavors són extremament resistents als sucs digestius dels ocells i usualment passen el tracte digestiu sense danys; d’altra banda, l’animal ha desplaçat la llavor i l’ha llençada amb el fertilitzant inclòs (l’excrement). Les plantes que fan aquests tipus de fruits han desenvolupat estratègies evolutives ben diferents, dedicades a atreure l’ocell, que en aquest cas és beneficiós i necessari per a la planta.
Els colors vius càlids, molt conspicus, preferentment vermells i taronges, són un primer senyal visual que assenyala a l’ocell la seva font d’aliment. Per tal d’estimular l’ocell a menjar i, conseqüentment, a dispersar les llavors, la planta ofereix a l’ocell una recompensa: el fruit. Si la planta fa els fruits molt nutritius, certament atrau bé els ocells, però li resta poca energia per a produir llavors; si els fruits són poc nutritius, podrà fer moltes llavors, però no compensarà energèticament els ocells de menjar-ne els fruits. Per tant, l’estratègia evolutiva gira entorn d’aconseguir fruits nutritius, però només suficientment nutritius per a atraure els ocells. D’això es desprèn que la dieta dels ocells frugívors és sovint nutritivament pobra i, consegüentment, dediquen una bona part del seu temps a les activitats d’alimentació. Just al contrari del cas explicat més amunt, les plantes que disposen de fruits sucosos, sovint els ofereixen durant llargs períodes de l’any, per tal d’augmentar la probabilitat que tots els seus fruits siguin menjats pels ocells.
A les zones temperades, contràriament a les tropicals, no hi ha fruits durant tot l’any. Això significa que moltes de les espècies que mengen fruits una determinada època de l’any, han de servir-se d’una altra dieta a les altres èpoques, que en molts casos és de tipus insectívor. Per tant, els ocells frugívors de les regions temperades no estan especialment adaptats a menjar fruits. Dins aquestes regions, gairebé totes les espècies que aprofiten fruits d’estiu i de tardor formen part de dues categories: la dels residents que mengen fruits estacionals, però amb un espectre alimentari que canvia quan aquests desapareixen, i la dels migradors que romanen estacionats mentre duren els fruits.
A Europa, la maduració de molts fruits s’allarga durant tot el temps que els petits migradors són més abundants a la zona. Mentre que la floració segueix un gradient latitudinal i és dona primerament a la conca mediterrània i posteriorment a les àrees més nòrdiques europees, la fructificació segueix un altre pla. En efecte, la maduració dels fruits a la Mediterrània sempre és posterior a la de les zones més nòrdiques i coincideix amb el fort del pas migratori dels frugívors, cosa que constitueix un bonic exemple de coevolució entre els ocells, que dispersen les llavors, i les plantes. La importància dels fruits a la conca mediterrània no es restringeix al pas migratori, sinó que també juga un paper preponderant per als hivernants. En el matollar esclerofil·le mediterrani, la qualitat nutritiva dels fruits de tardor és baixa, però la dels que maduren a l’hivern és suficientment elevada com per a permetre la supervivència dels consumidors de petita mida, de requeriments energètics molt grans: el pit-roig, els tallarols capnegre i de casquet (típicament insectívors durant la primavera-estiu), la merla, els tords o la griva, que en molts casos, a l’hivern, s’alimenten quasi exclusivament d’aquests fruits. Dintre de les espècies vegetals del matollar esclerofil·le mediterrani, són especialment importants, pel que fa als ocells hivernants, el llentiscle o mata, l’olivera o ullastre i el marfull, els fruits dels quals, en alguns casos, contenen a la polpa una quantitat de lípids que voreja el 60% del seu pes. Hi ha, per tant, un ajust estacional entre la qualitat dels fruits i els requeriments dels ocells frugívors, i de fet, en aquest cas sembla que també s’ha produït una coevolució difusa entre les espècies del matollar esclerofil·le mediterrani i la comunitat ornítica hivernant.
Tot plegat ens fa entendre que a les zones temperades és difícil d’ésser insectívor total. Les espècies que ho són, o bé migren quan vé el mal temps, o bé estan especialment adaptades a alimentar-se d’insectes i larves en hivernació, tal com alguns picots. També veiem que no és possible d’ésser un frugívor tot l’any, i que la major part de les espècies, encara que presenten certes adaptacions, tenen principalment becs adequats per a molts usos i fan servir dietes mixtes, especialment en relació amb l’estacionalitat.
El cicle anual
Els canvis estacionals
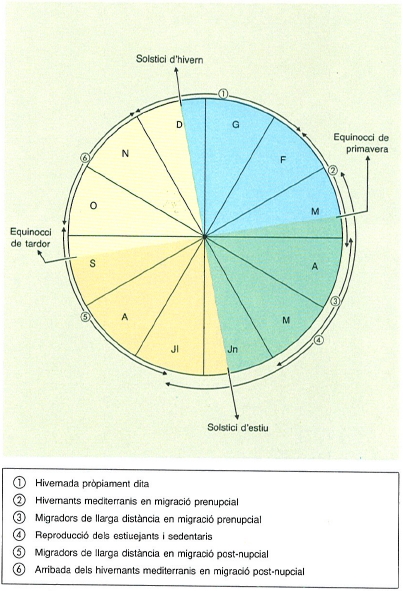
Cicle anual de l’avifauna als Països Catalans, amb indicació de les diferents categories. S’assenyalen els equinoccis i els solsticis per fer referència a la importància fonamental del fotoperíode.
Maber, original d’Enric Carrera.
En tots els ocells, migradors i sedentaris, se succeeixen una sèrie de canvis de periodicitat estacional. La successió ordenada i completa d’aquests canvis és l’anomenat cicle anual de l’ocell. Els principals canvis estacionals es relacionen amb la reproducció i la muda, i amb la migració, en el cas particular de les espècies migradores. Pel que fa a l’ocell migrador, hi ha dos fenòmens peculiars. D’una banda, el "Zugunruhe", expressió alemanya que indica un estat especial d’inquietud, que coincideix aproximadament amb l’època de la migració; i de l’altra, una acumulació de greix (subcutani i peritonial) abans i en el transcurs dels períodes migratoris. El greix, utilitzat com a reserva, els predisposa i faculta a realitzar llargs viatges sense escala. Un exemple ben clar és la boscarla dels joncs (Acrocephalus schoenobaenus), que en condicions normals pesa 11 gr, i just abans de la migració arriba a pesar de 20 a 22 gr; aquestes reserves la capaciten per a volar de 100 a 115 hores seguides sense parar. Un altre cas es el territ de tres dits (Calidris alba), que amb un pes habitual de 50 gr, arriba a pesar fins a 110 gr just abans de començar el seu viatge migratori. En aquest cas, les reserves extres de greix li permeten volar distàncies de l’ordre de les 2000 milles.
Actualment, quasi tots els científics coincideixen a creure que l’únic factor extern d’efecte sincronitzador general del cicle anual, es la variació del fotoperíode natural, és a dir, el canvi gradual de la durada del dia (llum) i de la nit, per raons còsmiques conegudes per tothom. Dins d’aquest cicle, els equinoccis marquen el punt de viratge, i l’allargament successiu dels dies a l’hemisferi nord correspon a l’escurçament de les nits a l’hemisferi sud i a l’inrevés.
La reproducció
La reproducció dels ocells és àmpliament coneguda i, per això, n’esmentarem solament algunes característiques generals, que tenen incidència a l’hora d’examinar la situació de la nostra avifauna, i que en bona part són relacionades amb la fenologia de cria. Així, hom pot preguntar-se si els ocells es reprodueixen tot l’any, quants cops ho fan, si el període de la cria és igual arreu, etc.

Els nius dels ocells continentals, com els d’hàbits aquàtics, són de nivells de complexitat molt variable segons cada espècie, des dels més rústecs, gairebé inexistents, com el del siboc (Caprimulgus ruficollis, a dalt, a l’esquerra), passant per les simples perforacions als marges terrosos, típiques dels nius de l’abellerol (Merops apiaster, a la dreta), o l’acumulació de branquillons en forats de penya-segats, com el cas de la gralla (Corvus monedula, al centre superior, a l’esquerra), o els teixits en forma de cassoleta oberta, de vegades situats a terra, cas típic dels alàudids, com el cotoliu (Lullula arborea, a la dreta) o penjats amb fang a l’enforcadura de les branques dels arbres, com el de la merla (Turdus merula, al centre inferior, a l’esquerra), fins als més complexos, tancats i entreteixits, com el característic de les mallerengues (com la cuallarga, Aegithalos caudatus, a la dreta) i el de la garsa (pica pica, a baix, a l’esquerra); un cas particular és el dels picots, com el picot negre (Dryocopus martius, a la dreta), que consisteix en un forat fet al tronc d’un arbre.
Jordi Muntaner, Xavier Ferrer, Josep M. Barres, Oriol Alamany i Anna Motis.

Els nius dels ocells aquàtics poden ser d’estructura molt variada, des d’una simplicitat absoluta fins a una gran complexitat. Efectivament, hi ha ocells que gairebé no fan niu, sinó que deixen els ous en petites concavitats del terreny, a la sorra, com el xatrac (Sterna albifrons, a dalt, a l’esquerra), o a les pedres, com el corriol petit (Charadrius dubius, a la dreta), o bé els dipositen sobre plataformes de penya-segats, com el corb marí emplomallat (Phalacrocorax aristotelis, al centre superior, a l’esquerra), o amagats entre la vegetació del litoral, com el cas de l’ànec coll-verd (Anas plathyrhynchos, a la dreta); d’altres construeixen lleugeres plataformes totalment flotants, com el fumarell carablanc (Chidonias hybrida, al centre inferior, a l’esquerra) o sobre el canyís, com el martinet ros (Ardeola ralloides, a la dreta); els nius més complexos són teixits en forma d’embut, com el de la boscarla de canyar (Acrocephalus scirpaceus, a baix, a l’esquerra), o completament tancats, com el del teixidor (Remiz pendulinus, a la dreta),
Marcel·la Chinchilla, Jordi Muntaner, Anna Motis i Xavier Ferrer.
Els ocells es reprodueixen en un període de l’any molt determinat, que varia segons les espècies i les contrades. En tots els ocells la criança dels polls coincideix amb el període de l’any de màxima abundància de recursos alimentaris; per exemple, mentre que la majoria dels falcònids dels Països Catalans tenen els pollets al final de la primavera o al començament de l’estiu, el falcó de la reina, que s’alimenta d’ocells petits (ornitòfag), puja els polls durant el mes de setembre, que és quan a les Illes es produeix el pas fort de moixons migradors. En el proper capítol ja s’especifiquen els períodes de cria de les espècies indígenes i s’observen les variacions que hi ha des del trencapinyes o la tórtora turca, que poden nidificar al desembre o al gener, fins al mateix falcó de la reina, que ho fa al setembre. Globalment, però, els mesos típics de cria al nostre país són l’abril, el maig i el juny, i amb menys intensitat el març i el juliol. En els tròpics, els ocells poden niar a qualsevol època de l’any.

Els nius dels ocells antropófils són construïts aprofitant raconades i enfonys d’edificacions, en indrets suficientment tranquils i arrecerats. És el cas de l’oreneta (Hirundo rustica, a dalt), que fa el niu amb fang, en forma de cassoleta, o el del niu completament rodó, anomenat ull de bou, del cargolet (Troglodytes troglodytes, a sota, a l’esquerra); l’òliba (Tyto alba, a la dreta) aprofita forats o relleixos, on diposita els ous, gairebé sense cap protecció, mentre que les teules són el punt escollit pels estornells (Sturnus unicolor, a baix, a l’esquerra) i els pardals; els nius de la cigonya blanca (Ciconia ciconia, a la dreta), molt vistents, són construïts al capdamunt de campanars i edificacions altes.
Oriol Alamany, Lluís Solé, Jordi Muntaner, Anna Motis i Xavier Ferrer.
A part les variacions específiques del període reproductor, lligades al règim alimentari, hi ha variacions geogràfiques produïdes per condicions particulars hidrològiques, de temperatura o microclima i també per oscil·lacions fenològiques interanuals a l’alta muntanya, que depenen del desglaç. Dues d’aquestes variacions geogràfiques són molt evidents i generals, referents a la latitud i a l’altitud. A l’hemisferi nord, el període de la reproducció s’endarrereix a mesura que s’avança cap al N, procés força general, i que també es dona a d’altres grups biològics. Paral·lelament al fenomen latitudinal hi ha l’altitudinal, que fa que la nidificació s’endarrereixi a mesura que es puja en altitud. A les nostres contrades, aquest desfasament entre la terra baixa i l’alta muntanya, tot i que és variable, gira entorn dels quinze dies o un mes.
Quan es parla del període de la reproducció, generalment s’entén el que va des de la posta del primer ou fins que els pollets ja volen. Des del punt de vista de l’ocell, aquest període és més llarg, ja que comprèn l’establiment del territori (si en té), l’establiment de la parella, la construcció del niu i, en molts casos, un temps de seguiment del jove, especialment a les espècies carnívores-piscívores, a les quals les tècniques de caça que ha d’aprendre el jove són complexes i demanen temps. Veurem, tot seguit, els dos períodes més importants de l’època de la cria: la incubació i l’alimentació dels polls.
El període d’incubació va des de la posta de l’últim ou fins a la seva desclosa, i bé que hi ha variacions considerables entre les espècies, a grans trets és més llarg com més gran és l’espècie. En el cas dels picots i els petits moixons, els ous desclouen abans de la fi de la segona setmana d’incubació; en els ocells més grossos, com cigonyes, falcons, flamencs, oques, ducs i pingüins, aquest període és de cinc o sis setmanes; a la major part de l’avifauna mediterrània oscil·la entre aquests límits. Períodes més llargs els tenen els voltors, les grans àguiles, els petrells i casuaris, que coven entre set i vuit setmanes; el rècord d’incubació de tots els ocells del món l’ostenta l’albatros reial, amb 81 dies.

El desenvolupament dels polls i la seva permanència als nius s’estén durant períodes variables segons si es tracta d’espècies nidícoles, com és el cas de l’escorxador (Lanius collurio, a dalt), a les quals els polls neixen desvalguts i absolutament incapaços de volar i de procurar-se aliment, o d’espècies nidífuges, com és el cas dels ànecs, que neixen proveïts de ploma i són capaços de picar menjar. En un estadi de desenvolupament posterior, els polls volanders, com el de l’oreneta (Hirundo rustica, a baix) es caracteritzen, entre altres coses, pel fet de tenir la cua curta i boqueres grogues.
Ricard Gutiérrez i Xavier Ferrer.
El període que va des de la desclosa fins que el poll vola, varia també entre les espècies; el megapòdid d’Australàsia és capaç de volar pràcticament des del seu naixement, però per a la major part dels passeriformes, aquest període és inferior a tres setmanes, mentre que, a l’altre extrem, corbs marins, cigonyes, calaus i grans rapinyaires nocturns, triguen vuit setmanes a volar. Són rares les famílies amb un llarg període pre-volador, com el de dotze setmanes, característic dels grans voltors del món antic i els casos extrems que representen l’albatros reial (de 36 setmanes) i l’albatros vagabund (més de 40). Hi ha unes certes precisions arran de l’aplicació dels termes poll, poll volander i jove. Es considera poll o pollet l’exemplar sortit de l’ou que encara no vola; des que vola fins que, a la tardor, canvia el plomatge i passa a vestir-se d’immatur o d’adult (segons l’espècie) pren el nom de jove. El terme, no tant precís, de poll volander s’aplica als pollets que viuen a les rodalies del niu i que, sovint, hi van a passar la nit; fan petites volades, però encara els creix el plomatge (cosa que s’observa externament en la cua, que és curta); mantenen les taques externes de color groguenc de les comissures del bec i no saben alimentar-se sols, de manera que depenen totalment o en una bona part dels pares. Aquesta condició de poll volander dura poc temps.
Coneguent les variacions en els períodes de cria, s’entén que certes espècies, preferentment les de mida petita i reproducció ràpida, puguin pujar, a les nostres latituds, dues i fins i tot tres llocades per temporada. Aquest és un fenomen molt freqüent a la conca mediterrània, on el temps temperat i càlid permet un període de reproducció més dilatat i una alta possibilitat de postes de reposició en el cas de pèrdues per pluja, predadors, etc.
La migració

El vol en formació és habitual en moltes espècies migradores, com aquests flamencs (Phoenicopterus ruber) arribats a les salines de Santa Pola.
Jordi Vidal / TAVISA.
Des d’un punt de vista ecològic, els consumidors capaços de desplaçar-se ràpidament grans distàncies, són els més ben adaptats per a explotar els recursos alimentaris dels medis fluctuants i utilitzar-ne els excedents, que es presenten en abundància durant un període limitat de l’any. Aquestes facultats de desplaçament segons un ritme regular, expliquen el quasi cosmopolitisme dels ocells i la seva presència, en densitats relativament importants, en regions quasi desproveïdes d’altres vertebrats, atès el curt període en el qual són habitables.
La migració constitueix, en definitiva, una resposta de l’organisme a les condicions fluctuants del medi, i per tant té una considerable importància ecològica. Cal fer una distinció inicial entre la migració pròpiament dita i altres moviments (irrupcions, erratismes, etc.). El terme migració només s’aplica quan es tracta d’un fenomen biològic de natura instintiva, que consisteix en un desplaçament d’una certa envergadura i durada, intencional o autònom, regular en el temps (periòdic) i l’espai (retorn a l’origen), de caràcter popular (poblacional) i que imposa un antagonisme entre les residències alternatives. La migració implica, doncs, dos viatges, el pre-nupcial o de primavera, que porta els ocells a l’àrea de cria, i el postnupcial o de tardor, que els duu a les àrees de repòs hivernal anomenades quarters d’hivern.
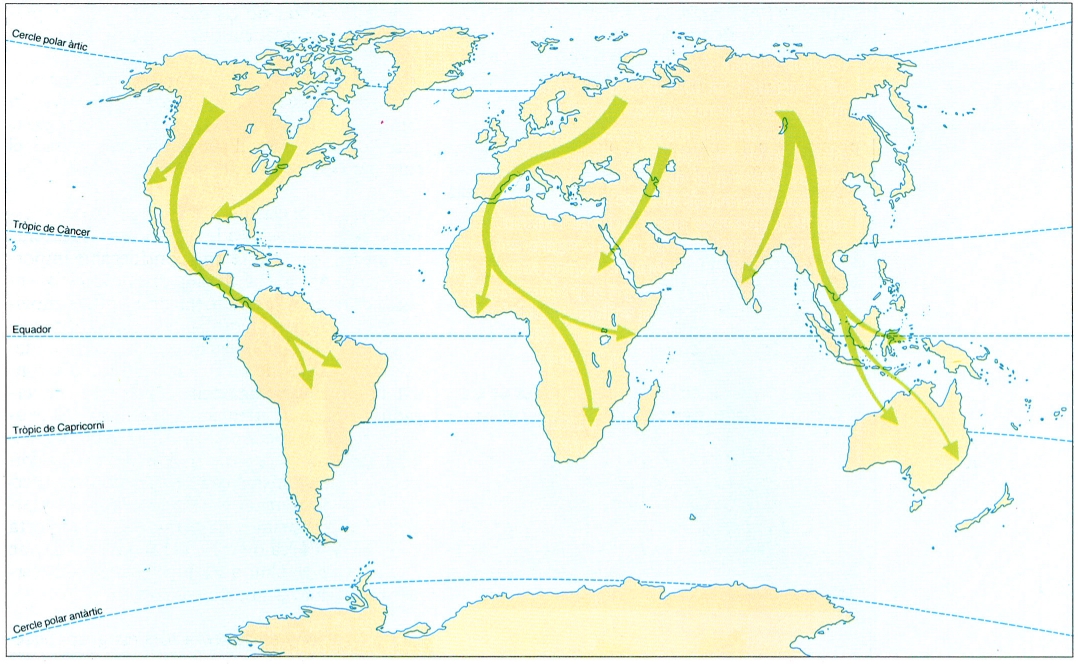
Principals rutes migratòries del món. El vessant occidental de la Mediterrània, on se situen els Països Catalans, cau dins de les vies principals de migració.
Maber, a partir de fonts diverses.
Les migracions es donen a tots els grups sistemàtics, excepte en aquells que reuneixen espècies no voladores. De fet, més del 60% de les famílies d’ocells disposa de representants migradors. Les migracions no són pròpies de regions determinades, encara que les zones fredes de tots dos hemisferis són l’escenari dels desplaçaments més espectaculars, atesa la gran amplitud de les fluctuacions climàtiques anuals. Els moviments estacionals afecten també les regions intertropicals, sotmeses a fluctuacions cícliques de l’ambient. Solament els ocells de les selves tropicals no manifesten cap tendència a desplaçar-se, gràcies a la uniformitat de l’ambient al llarg de l’any. Per exemple, al Canadà, el 75% dels passeriformes abandonen el país durant l’hivern, i al paleàrtic ho fan el 40%. La proporció d’individus migradors d’una mateixa espècie varia també amb la latitud; el pitroig, posem per cas, és totalment migratori a Escandinàvia, mentre que a Centreuropa, el 50% dels exemplars són migradors i a l’Europa meridional són tots sedentaris.
Sovint la migració s’efectua en un front ample, però en general hi ha vies predilectes de migració, que ressegueixen les costes, els colls de les muntanyes, les carenes batudes per vents regulars que afavoreixen el vol, els deserts, on els corrents tèrmics permeten mantenir-se a bastant altura amb poca despesa econòmica, etc. La major part de les vies migratòries van de l’hemisferi nord a l’hemisferi sud. La raó principal per la qual hi ha tan pocs ocells terrestres que, reproduint-se al S emigrin vers el N, és l’espai, ja que llevat de l’Antàrtida (que la cobertora de gel fa especialment inhòspita), hi ha una superfície cinc vegades més gran a l’hemisferi nord que al sud. Es coneixen poques espècies terrestres de l’hemisferi sud que passen l’Equador per hivernar més cap al N.
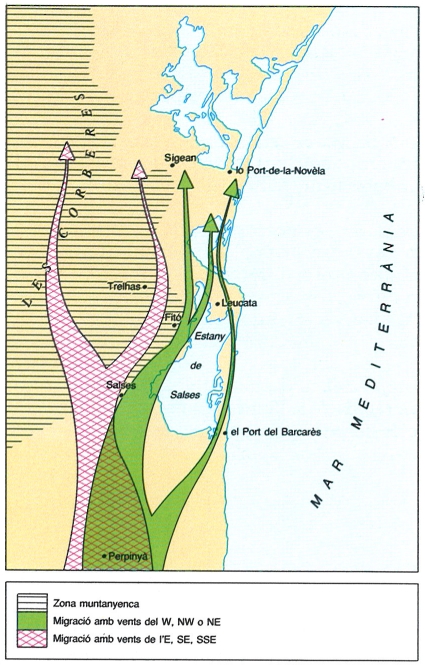
Vies migratòries del pas pre-nupcial a Leucata (Rosselló), que il·lustren la importància de la direcció dels vents en la migració.
Maber, segons dades de Sagot i Seriot.
La migració pre-nupcial o primaveral, és a dir, el viatge entre l’àrea de repòs hivernal i la zona de cria, té una durada més curta que la postnupcial i, per tant, implica una concentració més gran de migradors. Els ocells tenen una pressa especial per a arribar a les zones de nidificació, atès que aquestes romanen disponibles un període curt de temps i que de la seva rapidesa depèn que puguin triar els millors territoris de cria. Els ocells que arriben a les nostres terres al començament de març acaben de travessar el gran desert del Sàhara i una part de la Mediterrània si no tota. Malgrat tot, només fan una petita parada per tal de recuperar l’energia perduda en el llarg viatge. Un exemple n’és el cas de l’exemplar de mosquiter groc gros (Phylloscopus trochilus) anellat al delta de l’Ebre en pas primaveral i recuperat al cap de sis dies en una localitat alemanya situada 1500 km al NE en línia recta; aquest ocell va volar, doncs, un mínim de 250 km diaris. Al litoral continental català, el període de fort pas migratori pre-nupcial abraça des de mitjan març fins a mitjan maig.
La migració postnupcial o pas de tardor és formada per les mateixes poblacions del pas primaveral, a excepció de les espècies que migren en llaç, és a dir, que tenen un itinerari tardorenc diferent del primaveral. Als Països Catalans correspon als casos de l’escorxador (Lanius collurio), el mosquiter xiulaire (Phylloscopus sibilatrix) i d’altres, representats als gràfics adjunts. Les nostres terres són recorregudes, doncs, pe’ s mateixos individus que ja hi havien passat a la primavera, més les seves llocades (llevat, òbviament, de la fracció corresponent a la mortalitat d’estiu). El pas postnupcial, per tant, és més massiu i dilatat que el pre-nupcial. Els ocells no es veuen sotmesos a presses per arribar al lloc de destí, sinó que viatgen més tranquil·lament, tot preparant-se fisiològicament per a les llargues travesses sense intervals que els esperen. Al continent europeu, la major part dels petits migradors travessen d’una tirada la Mediterrània i el Sàhara. Aquest temps, a la regió mediterrània és característica una gran producció de baies que, com ja hem explicat, són explotades per un gran nombre de passeriformes (especialment sílvids i túrdids), que mostren en aquesta època una considerable hiperfàgia. Tot això els permet, d’una banda, recuperar-se del gran desgast que ha suposat tant el viatge prenupcial com la reproducció, i de l’altra emmagatzemar suficients energies pera acabar la muda i continuar el viatge cap a les àrees d’hivernada amb reserves suficients. Els primers ocells que arriben al nostre país durant el pas postnupcial són els limícoles, que ho fan al final de juliol; del començament d’agost al final d’octubre passen els migradors de llarga distància, és a dir, els que hivernen més enllà del Sàhara i a la vora dels tròpics (mosquiter groc gros, còlits, oriol, abellarol, hortolà, oreneta de ribera, cotxa cua-roja, etc.); i quan encara no han acabat de passar aquests, arriben els ocells que hivernen a l’àrea mediterrània i, en definitiva, a les nostres terres.
La muda

La muda o renovació del plomatge és un fenomen obligat per a tots els ocells. Sol ser un procés lent i progressiu, de manera que, en determinats períodes de l’any, el plomatge mostra la intercalació de plomes noves, de les quals apareixen primer els canons blavosos, com en aquesta foto d’una ala de valona (Tringa glareola).
Isabel Martínez.
L’estructura i la ubicació de les plomes impliquen un desgast de tot el plomatge que obliga l’ocell a reemplaçar-lo periòdicament per un de nou. Aquest conjunt de canvis, complets o parcials, del plomatge s’anomena muda. No és muda, per tant, el canvi de plomes no periòdic degut à un accident o bé a alguna altra causa. Durant la muda hi ha una alteració important del metabolisme i, en conseqüència, és un període de gran "stress" per a l’animal. D’altra banda, es requereix un esforç energètic important per a canviar el plomatge i fer-lo de nou (cal tenir en compte, a més, la necessitat d’aconseguir una gran quantitat d’aminoàcids, especialment arginina i cisteïna, per tal de poder fer les queratines), per a regular la temperatura (l’homeotèrmia queda alterada per la manca de plomes, la qual cosa implica un esforç suplementari per tal de compensar l’alteració de l’aïllament tèrmic) i per a volar (en el cas que mudin les plomes de l’ala i la cua, l’esforç per poder volar és més gran).
La mida de l’espècie juga un paper important en el tipus de muda. La majoria dels ocells grans muden irregularment durant un període llarg, que pot ser de més d’un any. Els altres muden, com a mínim, un cop l’any o cada dos anys. En el cas particular dels anseriformes (cignes, oques i ànecs), la majoria passen uns dies sense volar, perquè muden simultàniament les plomes de les ales i de la majoria del cos; com que en aquesta situació resten indefensos, els anseriformes trien indrets especialment segurs i productius per passar-hi aquest temps. Això comporta l’existència de veritables "migracions de muda" a aquests indrets (per exemple, el delta del Volga, diversos punts de la mar del Nord, etc.). Aquestes migracions particulars són fonamentals en el cicle anual de l’ocell i afecten no solament els anseriformes, sinó també d’altres grups aquàtics (gavines, limícoles, etc.), i probablement, també, no aquàtics.
En general, el procés de la muda inclou una muda postnupcial completa i una muda primaveral incompleta (o rarament completa), però hi ha diferències segons l’edat dels individus. Per exemple, els joves de l’any tornen a canviar totalment el plomatge cap a la tardor o l’hivern, fenomen que es coneix amb el nom de muda postjuvenil. L’explicació d’aquesta renovació tan ràpida de la lliurea és que el primer plomatge és de pobra qualitat; aquesta muda es pot fer servir, en alguns casos, per a datar l’animal.
La muda va íntimament lligada als ambients on es desenvolupen els ocells i a les seves fluctuacions. Els ocells muden quan les condicions els són més favorables; l’època de la muda els ha de proporcionar una gran quantitat de menjar, i d’altra banda, l’ajusten a d’altres fenòmens del seu cicle biològic, com la migració i la nidificació, que també demanen molta energia, i amb els quals gairebé mai no coincideixen en el temps. Els països del N d’Europa, de clima més rigorós, obliguen els ocells a mudar ràpidament per tal de poder migrar; en alguns casos, surten sense mudar i ho fan als quarters d’hivernada. Els països mediterranis, en canvi, permeten una certa laxitud, i això fa que hi hagi una gran varietat d’estratègies de muda dins d’una mateixa espècie, a causa de la coincidència temporal de diverses poblacions amb orígens geogràfics i ambientals diferents (i per tant, amb diversos estats de muda) i potser també per la coexistència de diverses estratègies dins d’una mateixa població. En parlar d’aquestes estratègies ens referirem solament a la muda completa anual que tenen la majoria d’adults (que és la més important al llarg del cicle anual de l’ocell) i a la fenologia dels Països Catalans.
Les espècies sedentàries, com per exemple el pardal comú, no tenen necessitat de mudar ràpidament perquè no han de migrar; comencen la muda cap a mitjan juny i l’acaben el novembre. Les poblacions estivals migradores (per exemple, el rossinyol) han de mudar més de pressa per tal de tenir a punt plomatge nou abans de migrar. La muda té lloc, en la gran majoria dels casos, després de la nidificado i abans de la migració. En alguns casos aïllats, per exemple el de la tallareta vulgar, la muda comença després de la primera posta, s’atura durant la segona posta i continua després, fins a finalitzar-la. Un cas en el qual la muda comença després de la nidificació, se suspèn i continua en els quarters d’hivernada és el de la tórtora. Moltes espècies no muden fins que no arriben a Àfrica (principal àrea d’hivern dels nostres ocells estivals), com per exemple l’oreneta i la major part dels transsaharians. Hi ha alguns ocells, com la boscarla de canyar, el gruix poblacional dels quals muda a Àfrica i una petita part de la població ho fa a la península Ibèrica. La gran majoria dels ocells que hivernen als Països Catalans tenen mudes parcials, que acostumen a afectar tan sols les plomes del cos i les secundàries més internes o terciàries; aquesta muda pre-nupcial normalment comença cap al gener i acaba a la primavera, i és important per a l’adquisició del plomatge nupcial.