
L’observació microscòpica dels cianòfits pot oferir imatges molt espectaculars al microscopista afeccionat, ja que el contrast entre les beines i el cos cel·lular pot ésser tan patent com en aquests dos casos: a baix, filament de Petalonema alatum, d’una superfície higropètrica de les muntanyes de Prades; a dalt, Chroococcus turgidus procedent del mateix ambient, amb beines concèntriques.
Francesc Torrella
A l'hora d'introduir els cianòfits o cianobacteris (les antigues cianofícies, o algues procariòtiques), cal recordar que el concepte d’alga correspon no pas a un grup homogeni d’organismes amb una història evolutiva comuna, sinó a un conjunt polifilètic d'éssers vius que solament tenen en comú la fotosíntesi de tipus superior i l’adaptació a viure en medis aquàtics o humits, on moltes espècies de grups diferents viuen juntes i d’on les recullen i les estudien els mateixos especialistes, els algòlegs (o ficòlegs). Per començar, i malgrat que a moltes obres d’algologia, de limnologia (estudi de rius, llacs, etc.) i d’oceanografia, hom estudia tot el conjunt de les algues, hi ha una primera diferenciació òbvia, que ningú no s’atreviria a negar, i és que les algues d’organització procariòtica tenen poc a veure amb les d’organització eucariòtica. La vella denominació d’algues blaves va essent desaconsellada cada cop amb més força, i es prefereix la denominació de cianòfits o la de cianofícies (l’única classe de què consta el fílum cianòfits). Darrerament, els microbiòlegs han dedicat gran atenció a alguns representants d’aquest grup, a l’estudi del qual apliquen les tècniques clàssiques de cultiu pur o axènic (és a dir, amb una sola espècie, sense cap altra de forastera), amb resultats molt interessants referits sobretot a llur fisiologia i a llur plasticitat morfològica. Per tal de posar de manifest llurs indubtables afinitats amb els bacteris, els microbiòlegs han proposat per als cianòfits el nom de cianobacteris, que ha estat bastant acceptat, però que desplau a molts algòlegs. Hom ha expressat aquesta disjuntiva bizantina de forma una mica humorística, tot dient que el que són cianofícies quan les estudiem tal com apareixen al camp, es converteix en cianobacteris quan els isolem i els estudiem en cultiu axènic. Com que l’enfocament d’aquesta obra és fonamentalment de camp, parlarem, doncs, de cianofícies.
Característiques del grup
Les cianofícies o cianobacteris són, dins del grup dels procariotes fotosintetitzadors, el grup que presenta la més gran diversificació, pel que fa al nombre d’espècies i a la seva varietat. Podríem dir que les cianofícies inclouen gairebé tots els procariotes fotosintetitzadors capaços de trencar les molècules d’aigua i, després d’utilitzar-ne l’hidrogen com a reductor (i no dependre, així, d’una substància reductora ja existent en el medi, com els bacteris fotosintetitzadors), desprendre oxigen com a subproducte (fotosíntesi de tipus superior, o oxigènica). Aquest tipus de fotosíntesi, que confereix a qui la sap fer una eficàcia i independència més grans respecte del medi, només es troba en un altre grup d’organismes procariotes, els procloròfits, descoberts fa poc, i molt rars.

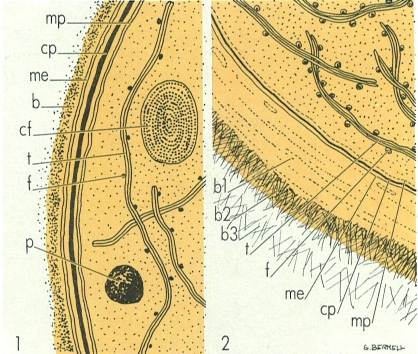
Estructures dels embolcalls cel·lulars i del citoplasma, esquematitzades a partir de seccions fines vistes al microscopi electrònic: 1 Gloeocapsa alpicola (croococcal) i 2 Dermocarpa (pleurocapsal) (b1/b2/b3 diferents capes de la beina, me membrana exterior, cp capa de peptidoglicà, mp membrana plasmàtica, t tilacoides, f ficobilisomes, cf grànul de cianoficina, p grànul de polifosfat o volutina).
Maber, a partir de dades facilitades per l’autor
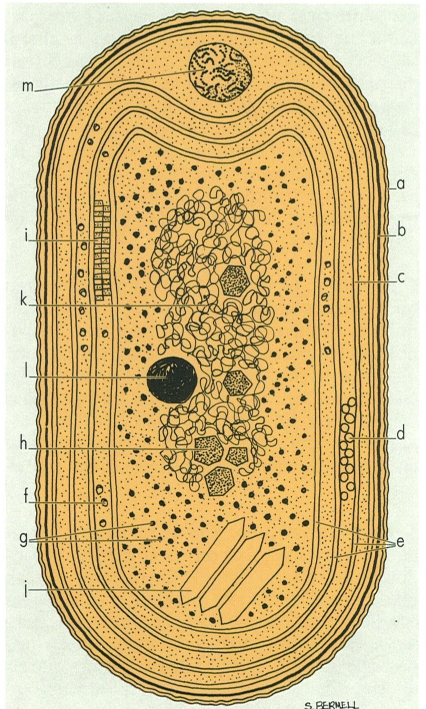
Organització cel·lular dels cianòfits, en el cas d’una cianofícia ideal estàndard: a membrana exterior; b capa de peptidoglicà; c membrana plasmàtica; d/i ficobilisomes (en visions frontal i lateral); e tilacoides; f grànul de glucogen; g ribosoma; h carboxisoma; j vesícula gasífera; k nucleoide; I grànul de polifosfat; m grànul de cianoficina.
Maber, a partir de dades de l’autor
Totes les cianofícies poden sintetitzar ficobiliproteïnes, pigments compostos per un grup cromòfor, la ficobilina, i una proteïna. La ficobilina principal, la ficocianina, és blava. Sovint hi ha també ficoeritrina, que és vermella. Juntament amb la clorofil·la a (l’única) i diversos carotenoides, les ficobiliproteïnes són els responsables principals dels colors verd blau, verd maragda, verd fosc, etc., de les masses de cèl·lules (del grec cianos, blau, i phytos, planta). Més ajustada és la traducció literal del terme popular anglès: algues verd blaves ("blue-green algae").
A més d’aquests caràcters fonamentals, fotosíntesi evolucionada i ficobiliproteïnes (que també es troben als rodòfits i als criptòfits), hi ha als cianòfits altres trets que, si bé no són totalment generals, es poden considerar prou típics. En una primera enumeració, podríem incloure-hi: capacitat d’adaptació dels pigments a la llum, a través d’una regulació de la síntesi de ficobiliproteïnes; presència de vacúols gasífers particulars (en espècies planctòniques); canvis morfològics específics com a resposta a factors ambientals, que afecten els heterocists, acinets, allargament cel·lular, pèls terminals, etc.; emmagatzematge del polipèptid cianoficina com a substància de reserva; pigments fotosintetitzadors disposats en tilacoides provistos de ficobilisomes; presència de carboxisomes al citoplasma; i facilitat per a establir simbiosis amb diferents organismes superiors.
En el conjunt dels sers vius, els cianòfits ocupen una posició intermèdia entre els fotosintetitzadors més primitius, els bacteris fotosintetitzadors clàssics, i els fotosintetitzadors amb organització cel·lular eucariòtica (algues i plantes superiors). Per una banda, la cèl·lula dels cianòfits és de tipus procariòtic, però per l’altra, la fotosíntesi és de tipus superior, evolucionat. Les diferències entre cianòfits i bacteris fotosintetitzadors són, bàsicament, de tipus bioquímic o fisiològic, ja que aquests darrers disposen solament del fotosistema I, que pot captar energia, però no pas fabricar substàncies reductores. En canvi, els cianòfits (i tots els fotosintetitzadors eucariòtics) tenen, a més del fotosistema I, el fotosistema II, capaç de trencar l’aigua i sintetitzar un reductor. Amb els fotosintetitzadors eucariòtics, les diferències són sobretot de tipus estructural, encara que també hi ha alguns tipus de molècules que apareixen exclusivament en una part dels grups. De tota manera la majoria de les cèl·lules dels cianòfits tenen unes dimensions de 5 a 10 vegades més grans que les dels bacteris.
Estructura cel·lular
El citoplasma
Al microscopi òptic, el citoplasma de les cianofícies presenta un aspecte dens, sense vacúols típics, d’aspecte uniforme però granellut (a causa de les abundants inclusions de què parlarem després). La seva viscositat és molt superior a la del citoplasma dels eucariotes, i no presenta cap moviment de circulació, a diferència d’aquests. Com a procariotes que són, les cianofícies no presenten nucli delimitat per cap membrana, sinó una zona central, tradicionalment anomenada centroplasma, on hi ha el material nuclear, menys acolorida que la zona perifèrica, o cromatoplasma, on hi ha els pigments assimiladors. Amb el microscopi, el color de la cèl·lula s’observa, típicament, verd pur tirant a maragda, o verd grisós, rarament verd fulla o verd groguenc com a les clorofícies.
Amb el microscopi electrònic, es pot veure el centroplasma ocupat per material nuclear filamentos i algunes inclusions. Al cromatoplasma hi ha una concentració de membranes portadores dels pigments fotosintetitzadors: són els tilacoides, que es poden disposar perpendicularment o paral·lelament a la superfície externa.
Això es veu bé als gèneres Synechococcus o Pseudanabaena. En algunes cèl·lules hi ha tilacoides a tot el citoplasma, amb una lleugera tendència a concentrar-se a la perifèria (per exemple, a Merismopedia, Calothrix o Anabaena).
Els tilacoides de les cianofícies, com els dels cloroplasts de les plantes eucariotes, són sacs membranosos aplanats, i encara que fan el mateix paper de contenir els pigments de la fotosíntesi, són ben diferents dels cromatòfors presents en molts bacteris fotosintetitzadors. Típicament, els tilacoides de les cianofícies estan separats els uns dels altres per un espai citoplasmàtic clar, és a dir, no estan aparellats ni apilats, com veurem que passa als procloròfits i als cloroplasts de les plantes, on es troben aplegats de dos en dos o formant agregats de diverses capes. El curiós, però, el cas de Gloeobacter violaceus, una cianofícia de les roques alpines, que no té tilacoides. Tots els seus pigments fotosintetitzadors es concentren vora la membrana citoplasmàtica, tocant la seva cara interna.
Un aspecte molt típic dels tilacoides de les cianofícies és la presència de ficobilisomes. Es tracta d’orgànuls, només visibles al microscopi electrònic, que trobem adherits a la part externa del tilacoide. En ells hi ha les ficobiliproteïnes. Es componen d’unitats discoïdals, de 3 a 6 nm de gruix, apilades formant bastons d’uns 12 nm, d’amplada i longitud variables. La seva mida varia segons la natura i la intensitat de la llum que reben. També als cloroplasts de les algues vermelles o rodòfits hi ha ficobilisomes, amb algunes de les ficobiliproteïnes dels cianòfits, però també contenen altres substàncies peculiars. A més de la seva diferent composició molecular, les ficobiliproteïnes es poden distingir per llur absorció de la llum de diferents longituds d’ona. La funció d’aquests pigments és captar llum i transferir la seva energia a les clorofil·les que hi ha als centres fotosintetitzadors actius. Tant les cianofícies com les rodofícies contenen ficocianina i al·loficocianina, de manera general. L’eritrocianina, en canvi, no és present a totes les espècies.
Al centroplasma hi ha el material nuclear, d’estructura filamentosa, que conté el patrimoni genètic, i ribosomes, de natura procariòtica (coeficient de sedimentació 70 S), que també es troben, menys abundants, a la resta del citoplasma.
Les inclusions citoplasmàtiques
L’aspecte granellut del citoplasma de les cianofícies és degut a la presència de diversos tipus d’inclusions, visibles al microscopi òptic. N’hi ha d’altres solament visibles al microscopi electrònic.
Grànuls de glucogen. Totes les cianofícies poden sintetitzar glucogen, que actua com a reserva energètica. Aquest polisacàrid de glucosa, molt ramificat, forma grànuls de 30 a 70 nm de diàmetre, només visibles amb el microscopi electrònic. Quan n’hi ha molts, però, les cèl·lules es tenyeixen d’un color groc daurat fosc amb solució de lugol (iode solubilitzat). És curiós que algunes cianofícies, com Chlorogloeopsis, poden acumular poli-β-hidroxibutirat, un polímer de reserva típic de molts bacteris.
Cianoficina. Es tracta d’un polímer de reserva que només es troba a les cianofícies. Forma grànuls visibles al microscopi òptic que, a les cianofícies filamentoses, es disposen preferentment a la vora dels septes que separen les cèl·lules. La cianoficina és una substància nitrogenada, formada per un polipèptid que conté àcid aspàrtic i arginina. Amb el microscopi electrònic, els grànuls de cianoficina es veuen sovint estructurats.
Grànuls de volutina. Són la reserva de fosfat de les cianofícies. Són compostos de fosfat polimeritzat (metapolifosfat). Es tenyeixen bé amb blau de metilè i blau de toluïdina i, en aquestes condicions, es veuen bé al microscopi òptic. Observats amb contrast de fases positiu, tenen l’aspecte de grànuls clars i brillants, mentre que els de cianoficina apareixen com punts negres. Amb el microscopi electrònic, quan en fer la preparació s’han utilitzat sals de plom, la volutina resulta opaca als electrons, i la cianoficina és més transparent, però es presenta més estructurada.
Els carboxisomes. Invisibles amb el microscopi òptic, amb l’electrònic tenen aspecte de cossos de secció més o menys polièdrica. Estan limitats per una fina membrana de 3 a 5 nm de gruix. Es troben a molts altres procariotes autotròfics, i contenen importants quantitats de ribulosa-bifosfat-carboxilasa, una molècula clau de cara a la fixació del diòxid de carboni, al cicle de Benson-Calvin.
Vacúols gasífers. Es troben en moltes cianofícies planctòniques o en les formes de dispersió d’altres cianofícies. Al microscopi de contrast de fases, es veuen com cossos molt refringents. Cada vacúol gasífer és format per moltes vesícules plenes de gas, les quals només es veuen amb el microscopi electrònic. Són idèntiques a la resta de les vesícules dels procariotes. Tenen forma cilíndrica, amb un con apuntat a cada extrem. La paret dels vacúols gasífers és proteica, i hidròfoba a la part interna de la vesícula. Els gasos dissolts al medi arriben, per difusió, a l’interior de les vesícules. Quan, a causa d’una pressió hidrostàtica excessiva a la qual està sotmès el cianòfit, o d’un augment de la pressió de turgència cel·lular, aquestes vesícules es col·lapsen, els gasos es dissolen en el líquid cel·lular sense formar bombolles. Les cianofícies planctòniques fan servir els vacúols gasífers per a surar a l’aigua o per a mantenir-se a la profunditat més adient, segons les condicions ambientals. Si agafem una ampolla d’aigua que conté cianofícies amb vacúols gasífers, aviat s’acumulen a la part superior, on formen una capa ben visible. Les vesícules de gas no poden inflar-se o desinflar-se com un globus, ja que són rígides.
La paret cel·lular
L’estructura de la paret cel·lular de les cianofícies segueix el model dels bacteris gramnegatius. Però la microscòpia electrònica hi revela diferències en el gruix de les diferents capes, i les anàlisis químiques permeten comprovar que hi ha diferències importants en la composició dels polímers de la paret.
La capa de mureïna (peptidoglicà) que cobreix la membrana citoplasmàtica és, en general, més gruixuda que als bacteris típics; no hi ha cel·lulosa. Per fora, i una mica separada, hi trobem una membrana externa, formada per fosfolípids, proteïnes i lipopolisacàrids, anàloga a la dels bacteris gramnegatius. Però als cianòfits hi ha encara un cert nombre de capes, per fora d’aquesta membrana, que poden ser de natura fibrosa o amorfes i de tipus gelatinós. En algunes espècies, les capes exteriors poden ésser molt gruixudes, i estableixen continuïtat amb els materials de la càpsula i de les beines. Aquestes capes, formades per polisacàrids, són la base d’una gran varietat d’estructures morfològiques i agrupaments cel·lulars típics de les cianofícies.
Hom ha detectat, però, diferències menors entre la paret de les cianofícies i la dels bacteris gramnegatius: una composició més alta en sucres aminats a l’àcid muràmic dels primers, alguns sucres aminats, com la galactosamina i la mannosamina, l’absència de les lipoproteïnes que, als bacteris gramnegatius, uneixen la mureïna amb la membrana externa, diferències en els lipopolisacàrids, etc.
El moviment
Les cianofícies no presenten mai flagels de cap mena. La majoria no tenen moviment, però una bona part de les espècies filamentoses nues (sense beina o amb beina molt fina) i no ramificades, que viuen sobre els sediments dels fons de l’aigua o altres superfícies, presenten un curiós moviment de lenta reptació, que els permet d’avançar a una velocitat de 2 a 11 µm per segon, i evitar així d’ésser recobertes pels materials que es van sedimentant, o bé anar fins a llocs més favorables per al creixement. Aquest moviment no depèn tan clarament com el de les diatomees de la secreció d’un mucilag, sinó que es basa en l’acció de microfibril·les que, per fora de la capa de mureïna, es troben enrotllades sobre el filament. Llur acció produeix ondulacions a la paret, les quals, interaccionant amb el substrat, són causa d’un moviment de rotació i, alhora, de traslació. Per això, la punta encorbada de les Oscillatoria, que típicament presenten aquest moviment, en rodar, sembla oscil·lar d’una banda a l’altra. També hi intervé l’extrusió de mucilag a través de porus molt fins (uns 10 nm de diàmetre) que hi ha a la paret cel·lular. En considerar, més endavant, els hormogonis veurem que presenten la mateixa mena de moviment.
Diversitat morfològica
Morfologies tipiques dels sis ordres de cianofícies existents, segons l’ordenació botànica del grup, idealitzades a partir d’exemples representatius: 1/3 croococcals (Synechococcus i Merismopedia, respectivament); 2 pleurocapsals (Pleurocapsa ); 4 camesifonals (Chamaesiphon) 5/8 estigonematals (Stigonema i Fischerella, respectivament); 6 oscil·latorials (Oscillatoria); 7 nostocals (Nostoc).
Maber, original de Francesc Torrella
A nivell cel·lular, és més petita que als bacteris. Les formes cel·lulars bàsiques són molt senzilles: l’esfera, el cilindre amb les bases arrodonides (secció el·líptica) o gairebé planes (secció rectangular), amb diferents variacions degudes al contacte i mútua pressió entre cèl·lules, sovint com a resultat de la divisió cel·lular. Aquestes interaccions poden ser causa de cèl·lules en forma de disc o de moneda (com a moltes Oscillatoria), o de barrilet (com a Anabaena i Nostoc). Quan hi ha una diferenciació en base i àpex, trobem cèl·lules més o menys allargades, com a Chamaesiphon, que té el pol apical més ample que el basal.
Però on trobem més diversitat a les cianofícies és en la morfologia de les agrupacions pluricel·lulars. Les cèl·lules de les cianofícies es poden agrupar entre elles de dues maneres molt diferents. Per una banda, hi ha grups en els quals les cèl·lules, després de la divisió, no mantenen cap contacte directe entre elles. La intercomunicació entre les cèl·lules integrants de la colònia així formada només es pot fer, si es fa, a través de molècules químiques que han de travessar l’espai intercel·lular, normalment ocupat per mucilag. L’altra possibilitat consisteix en el fet que les cèl·lules, després de dividir-se, generalment en la mateixa direcció, quedin en contacte, formant així un rengle filamentós o tricoma. Existeixen tricomes envoltats per una beina més o menys conspicua, o per capes mucilaginoses i la membrana externa de la paret, formant el que en diem filaments.
La microscòpia electrònica ha descobert que, en molts gèneres, les parets transversals són travessades per petits porus ultramicroscòpics, els microplasmodesmes, que permeten com a mínim un contacte directe entre les membranes citoplasmàtiques de les cèl·lules adjacents, i potser, fins i tot, fan possible que hi hagi continuïtat entre els citoplasmes. Es evident que, en aquests casos, el grau d’integració dels tricomes és molt més gran que el de les colònies mucilaginoses.
En general, els tricomes presenten, com a Oscillatoria, un rengle únic de cèl·lules, són uniseriats. Però en alguns grups de morfologia més complexa, com les estigonematals, hi trobem tricomes pluriseriats i fins i tot ramificats. Hi poden haver també (entofisalidàcies, Hyella) sèries ordenades de cèl·lules que no mantenen un contacte íntim: són els pseudofilaments.
La morfologia de les colònies de les cianofícies ha estat utilitzada com a base per la sistemàtica clàssica, pels botànics i pels ecòlegs. És freqüent que, quan hom les cultiva al laboratori, no mostrin la seva capacitat per a sintetitzar els materials capsulars que són la base de moltes de les morfologies colonials, tan ben representades a la natura. Aquesta és una de les raons que han conduït els microbiòlegs, avesats a treballar amb material cultivat, a reduir considerablement el nombre de gèneres de cianofícies en les sistemàtiques proposades per ells. Fem a continuació un breu repàs dels principals tipus morfològics de les cianofícies, tal com els podem trobar a la natura.
Si partim de les cianofícies amb cèl·lules esfèriques, en podem trobar d’isolades i sense càpsula aparent (Synechocystis) o amb materials capsulars que formen beines fines i laminars (Chroococcus) o més amples i vesiculars (Gloeocapsa). Si les cèl·lules són cilíndriques o el·lipsoïdals, tenim, sense càpsula, Synechococcus, i amb càpsula clara i vesicular, Gloeothece. Pot passar també que les cèl·lules es trobin irregularment repartides en una mateixa colònia (no agrupades les filles d’una divisió cel·lular). Si les cèl·lules són esfèriques, tenim el gènere Aphanocapsa, i Aphanothece, si són allargades.
Quan les cèl·lules, petites i amb vacúols gasífers, es troben densament agrupades en el mucílag, formant colònies en general planctòniques, de forma irregular, tenim Microcystis. Però també poden formar, per divisió en dues direccions de l’espai, colònies tabulars planes (Merismopedia), mentre que si es divideixen en totes tres direccions de l’espai, formen paquets cúbics, molt ben ordenats (Eucapsis). També hi poden haver colònies buides, amb les cèl·lules situades a la perifèria (Gomphosphaeria, Coelosphaerium).
Seguint la línia d’augment de complexitat colonial, trobem encara els pseudofilaments, ja vistos, i els pseudo-parènquimes, estesos en dues direccions, però sense relació entre les cèl·lules (Hydrococcus). També podem trobar paquets de forma cúbica (sarcines) a Myxosarcina. Chlorococcidiopsis, unicel·lular al principi, per divisions successives dins d’una paret comuna acaba formant un grup cel·lular compacte. Una cosa semblant passa a Dermocarpa, però la colònia resultant presenta un pol basal i un pol distal. També presenta una clara polaritat Chamaesiphon, que té la cèl·lula basal dins d’una beina oberta, unida al substrat.
Entre els cianòfits que presenten filaments* veritables (estructura hormogonal), distingim les formes humorístiques, que no presenten cap diferenciació clara entre les cèl·lules, i les heterocístiques, que presenten heterocists. Al primer grup, tenim espècies sense beina, com Oscillatoria o Spirulina (que té forma d’hèlix), i espècies amb el tricoma ficat en una beina, com Lyngbya. Entre les heterocístiques, n’hi ha sense ramificacions veritables, com Anabaena, que forma rosaris de cèl·lules amb heterocists, i Nostoc, que també en forma, però que té els filaments inclosos en una beina mucilaginosa que forma colònies macroscòpiques, sovint fosques. En aquests dos gèneres, els heterocists són intercalars o terminals, però a Cylindrospermum, són terminals. Els membres del gènere Scytonema presenten falses ramificacions. En aquestes, el tricoma, en créixer a l’interior de la beina, la pressiona, la trenca en un punt, i surt a l’exterior en forma de nansa que acaba trencant-se, tot formant el que semblen dues branques. A Tolypothrix observem que la cèl·lula basal del filament vell s’ha convertit en un heterocist, un fet comú al procés de falsa ramificació. També poden semblar ramificats, sense ésser-ne, els filaments de Calothrix i de Rivularia, que presenten una clara diferenciació entre la base, on hi sol haver un heterocist, i l’àpex del filament, que pot acabar en un llarg pèl incolor.
* Anomenem filament, el conjunt de tricoma (o tricomes) i beina que l’embolica. Un filament veritable és aquell en el qual les cèl·lules estableixen un contacte directe entre elles. El tricoma és el grup de cèl·lules arrenglerades i en contacte.
Les formes heterocístiques de les estigonematals presenten tricomes amb ramificació veritable. En els filaments més vells, es poden distingir tricomes pluriseriats (amb diversos rengles de cèl·lules), com a Fischerella i Stigonema. En alguns gèneres, com Nostochopsis, els heterocists són pedicel·lats, i es formen en posició lateral, sobre una curta branca del tricoma principal. La ramificació veritable és la que afecta el tricoma, i no sols el filament. Sols es troba en les estigonematals, i pot ésser lateral o dicotòmica. Les cèl·lules poden dividir-se en més d’un pla, sense deixar de formar part del tricoma.
Divisió cel·lular i multiplicació
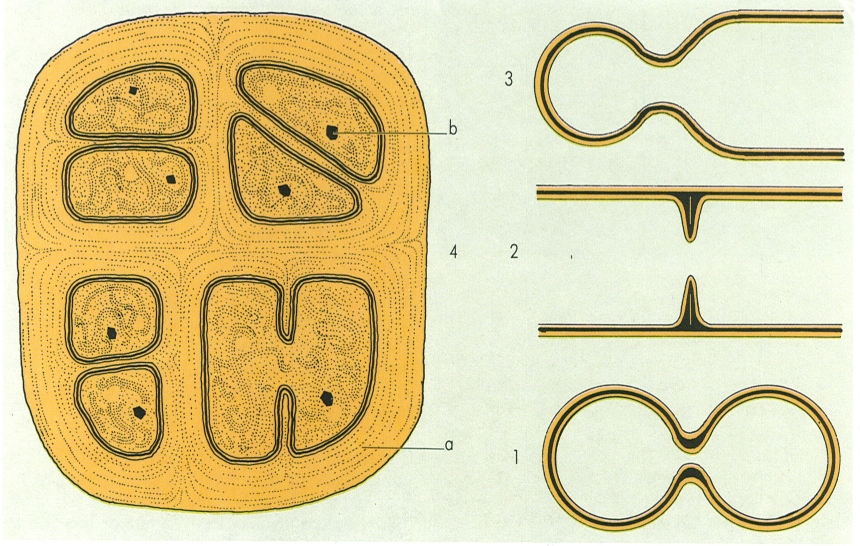
Tipus de divisió i separació cel·lular en les cianofícies: 1 escissió binària de tipus constrictiu; 2 escissió binària amb formació de septe; 3 gemmació; 4 divisió múltiple, en el cas d’una colònia de Chroococcidiopsis, amb beines gruixudes (a) i carboxisomes citoplasmàtics (b).
Maber, a partir de dades de l’autor
La forma bàsica de divisió cel·lular de les cianofícies es l’escissió, que es pot produir de quatre formes diferents. A la primera, dita escissió binària per constricció, després de la duplicació del material genètic, la zona equatorial de la cèl·lula mare s’estreny, i la separació de les cèl·lules es produeix per estrangulació (per exemple, a Synechocystis) o per creixement de la capa de material capsular (per exemple, a Chroococcidiopsis. Al segon tipus, es forma un septe, de la perifèria vers el centre, sostingut per la capa de peptidoglicà i per la membrana citoplasmàtica; un cop completat, resta sovint un contacte entre cèl·lules, mitjançant porus molt fins, els microplasmodesmes. La separació entre cèl·lules es pot fer per lisi de les capes interiors de la murrïna del septe (Nostoc, Anabaena) o per la formació de necridis, que són cèl·lules que, en morir, permeten la fragmentació dels tricomes (per exemple a Oscillatoria i a Phormidium); aquesta és la forma més freqüent de multiplicació dels tricomes. La tercera forma, pròpia de Chamaesiphon i gèneres pròxims, es produeix per escissió binària desigual, dita també gemmació; és una forma d’escissió per constricció, que apareix vora l’extrem apical i que separa una cèl·lula esfèrica, que funciona com una exòspora. Finalment, i en alguns escassos casos (Dermocarpa), l’interior de la cèl·lula es divideix en un cert nombre d’unitats iguals (endòspores o beòcits), que queden lliures per ruptura de la paret externa; ni per llur origen ni per llur estructura, tenen res a veure amb les endòspores dels bacteris.
Com els bacteris, les cianofícies no presenten reproducció sexual, encara que el capteniment d’algunes d’elles fa pensar en una possibilitat d’intercanvi genètic d’una cèl·lula a una altra, semblant al que es coneix als bacteris (conjugació) o transducció a càrrec de cianòfags (bacteriòfags especialitzats en cianofícies).
Formes cel·lulars especialitzades

Diferenciació cel·lular als cianòfits. A l'esquerra, necridis, cèl·lules col·lapsades, bicòncaves, a Oscillatoria margaritifera, de les aigües salades del litoral; al mig, heterocist a la base d’una falsa ramificació de Tolypothrix distorta, amb un proheterocist adjacent; a la dreta, heterocist de Gloeotrichia pisum, sobre vegetació submergida (Gandia).
Francesc Torrella i Xavier Tomàs / Francisco A. Comín
Les cianofícies, fins i tot les que viuen en medis efímers, no acostumen a presentar formes de resistència a les condicions adverses. Ni tan sols les endòspores (o beòcits) o les exòspores, que acabem de veure, presenten adaptacions especials a la resistència. Això és en bona part degut a l’extraordinària capacitat de resistència que té llur citoplasma, dens i viscós, que tolera bé la deshidratació. Al fang dels bassals secs, a les lleres dels rius, sobre les pedres i fins en suspensió en l’aire, hi ha cèl·lules deshidratades de cianofícies, en vida latent, que tornen a créixer quan disposen novament d’aigua. Un got d’aigua de l’aixeta posat a l’ampit d’una finestra presenta, al cap de pocs dies, un sediment verdós al fons, format principalment per cianofícies i clorofícies.
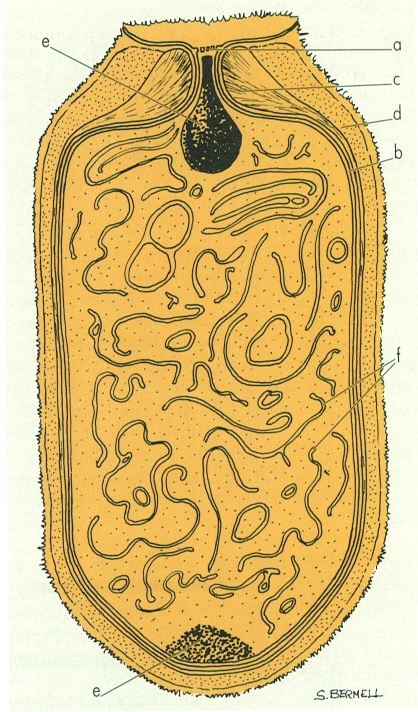
Organització d’un heterocist, en representació esquematizada: a connexions intercitoplasmàtiques; b membrana plasmàtica; c capa laminar de la paret; d capa homogènia de la paret; e grànuls polars de cianoficina; f sistema membranós intracitoplasmàtic.
Maber, a partir de dades de l’autor
Amb tot això, les cianofícies mostren una gran capacitat per a la diferenciació cel·lular, capacitat que ha produït diverses menes de cèl·lules especialitzades. Els acinets representen l’adaptació a condicions desfavorables. Són cèl·lules que es tornen més grosses, amb una paret més gruixuda que la de les cèl·lules vegetatives, a vegades amb petites protuberàncies. Es veuen bé amb el microscopi òptic pel seu citoplasma granellut, causa de l’acumulació, en gran quantitat, de cianoficina, com a substància de reserva. Entre la paret cel·lular i les capes mucilaginoses, presenten una nova capa fibrosa. Tenen un metabolisme més reduït. Quan les condicions ambientals són adequades, germinen. Són típics els acinets de Cylindrospermum.
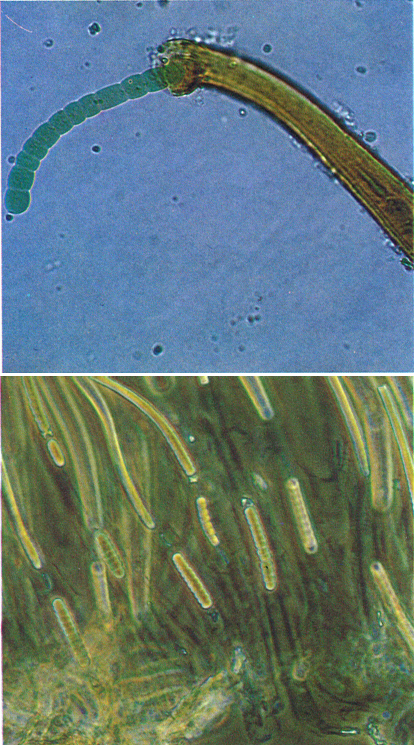
Hormogonis de Scytonema myochrous, eixint per la punta de la beina acolorida per l’escitonemina (Valls, Camp de Tarragona), a l’esquerra, i de Rivularia dura, de sobre les pedres dels rius (Prades), a la dreta.
Francesc Torrella
Les cianofícies fílamentoses poden presentar una mena de cèl·lules molt interessants i característiques, els heterocists. Són cèl·lules incolores, generalment més grosses que les normals, amb una paret reforçada, engruixida a les parets en contacte amb les cèl·lules adjacents, formant els corpuscles polars. Es tracta de cèl·lules especialitzades en la fixació de nitrogen en condicions aeròbiques, una funció de gran transcendència ecològica. Però també poden servir, en alguns casos, com a cèl·lules de resistència o per a facilitar la fragmentació dels tricomes. Hom ha trobat heterocists fòssils en roques del Precambrià, i segurament varen evolucionar quan l’atmosfera de la Terra va anarse enriquint en oxigen, com a resultat de la fotosíntesi de les mateixes cianofícies.
Els heterocists estableixen contacte amb les cèl·lules adjacents per una placa perforada per microplasmodesmes, a través dels quals reben aliments i altres molècules. No tenen sinó la part primitiva de la fotosíntesi (fotosistema I), que els serveix per a donar electrons al sistema fixador de nitrogen. Obtenen també energia oxidant sucres cedits per les altres cèl·lules, i així consumeixen l’oxigen que els arriba, el qual malmetria l’enzim nitrogenasa, molt sensible. Pels mateixos microplasmodesmes el nitrogen, ja fixat en forma de glutamina, un aminoàcid, passa a les cèl·lules vegetatives. Molts organismes s’han associat amb cianofícies, formant simbiosi, per tal d’aprofitar-se d’aquesta capacitat de fixació. En trobem casos en líquens (Peltigera), hepàtiques (Blasia), pteridòfits (Azolla) i plantes superiors (Cycas, Gunnera, etc.). La simple presència de cianofícies lliures al sòl o a l’aigua pot representar una aportació important de substàncies nitrogenades, com és el prou ben conegut cas del paper fertilitzant de les cianofícies que viuen a l’aigua dels conreus inundats d’arròs. D’altres estableixen simbiosi amb protozous, i hi viuen a l’interior de la cèl·lula, formant el que hom anomena cianelles. La diferenciació de les cèl·lules vegetatives en heterocists té lloc quan al medi no resta nitrogen combinat. Però quan el medi natural és ric en fonts de nitrogen combinat (nitrats, amoni) hi ha una desespecialització dels heterocists, que retornen a l’organització de cèl·lula vegetativa normal.
Els hormogonis són filaments nus i reptants de cèl·lules vegetatives, els quals es separen per la mort d’una cèl·lula (necridi). A les cianofícies ramificades, o fixes per un extrem, s’originen a l’àpex del filament, com en el cas de Rivularia, però més típics són els dels filaments amb beina, que surten de l’interior d’aquesta i es dispersen reptant com si fossin una petita Oscillatoria, cosa que el filament amb beina no podria fer.
Els hormocists, a mig camí entre hormogonis i acinets, són formats per diverses cèl·lules, de contingut molt granellut, envoltades per una beina acolorida, com si fossin hormogonis immòbils i protegits. Per això són anomenats també pseudohormogonis. En condicions favorables, germinen.
Hom parla encara dels nanòcits, que es formen a les croococcals, i sobretot a Gloeocapsa, Aphanothece, Mycrocystis, etc., per divisions successives de cèl·lules mares, sense que s’hi produeixi cap creixement. Són tan petits que sovint es fa difícil distingir-los de bacteris colonials.
Algunes rivulariàcies presenten a l’extrem del filament cèl·lules apigmentades, que formen un pèl i semblen mortes, però tenen citoplasma viu. Hom ha trobat que la deficiència de fòsfor indueix la formació de pèls llargs i, en general, l’allargament cel·lular de molts cianòfits filamentosos.
Peculiaritats fisiològiques
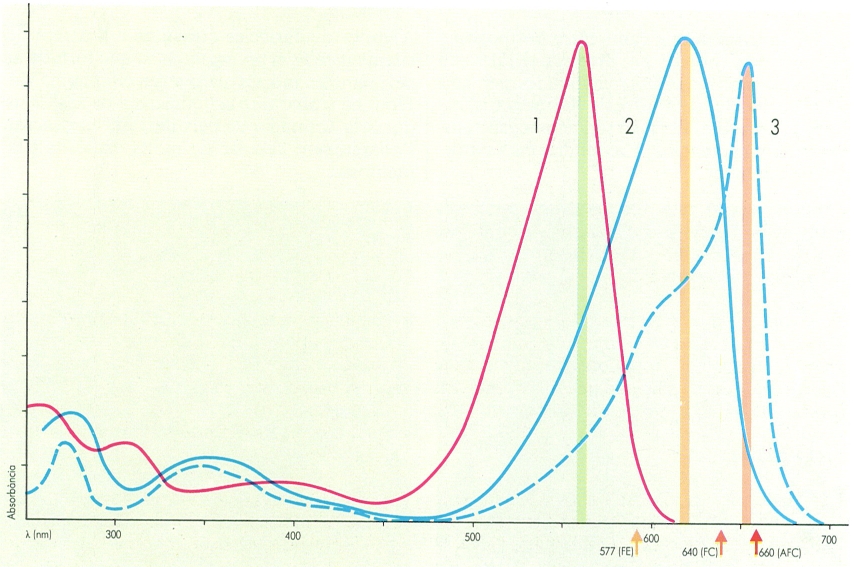
Espectre d’absorció i altres propietats òptiques de tres ficobiliproteïnes comunes en moltes cianofícies. El color de les corbes correspon aproximadament al del pigment concentrat quan reflecteix la llum. Les bandes verticals coincidents amb els pics d’absorció tenen un color semblant al de la llum absorbida pels pigments, és a dir, la longitud d’ona que poden aprofitar energèticament. Les sagetes de colors sota l’eix d’abscisses indiquen el color de la llum de fluorescència emesa pels pigments excitats.
Maber, original de Francesc Torrella
No deixa d’ésser sorprenent que uns organismes tan antics continuïn estant tan ben representats en ambients molt diversos de la biosfera. És clar que, a les primeres etapes de la història de la vida, a l’eó Proterozoic, en un llarguíssim tros de la història de la Terra, que va entre fa 2500 i 600 milions d’anys, les cianofícies dominaven entre els sers vius fotosintetitzadors, formant, arreu on hi havia aigua, a la mar, als pantans, etc., capes vives que anaven atrapant sediments, induint la precipitació de carbonat càlcic i formant roques constituïdes per múltiples capes superposades, els estromatòlits, que ara trobem fòssils. En aparèixer nous grups més evolucionats, les cianofícies foren eliminades de molts dels ambients que colonitzaven, però encara formen estromatòlits en maresmes salats, en llocs tèrmics (golf Pèrsic, Bahames, costa W de Mèxic, etc. i també a les salines de Santa Pola) i continuen, amb ple èxit, colonitzant múltiples ambients, molts d’ells extrems.
Hem vist algunes de les propietats que els asseguren l’èxit: la plasticitat adaptativa, la resistència a l’assecament sense danys al citoplasma, etc. Ja sigui per llur organització procariota o per l’acumulació de substàncies tòxiques (que poden arribar a emmetzinar el bestiar que beu aigua amb cianofícies), tenen pocs enemics naturals que se les mengin, ja que són difícils de digerir. A més, la seva capacitat per a fixar nitrogen i la seva adaptació a aprofitar matèria orgànica dissolta a l’aigua fan que puguin viure bé en aigües pobres en certs nutrients, com ara les aigües estratificades i calentes dels llacs i estanys, a l’estiu. Encara que no creixen massa de pressa, llur taxa de mortalitat reduïda permet que s’acumulin, a vegades en grans quantitats (flors d’aigua o "blooms"). En conjunt, les cianofícies viuen molt bé en tota mena d’ambients que resultin desfavorables per a les algues eucariòtiques. Citem, com a exemple ben conegut, les cianofícies de les aigües termals, que poden suportar temperatures de 60°C i fins de més de 70°C, temperatures que cap citoplasma normal, bacteris a part, no aguantaria.
Entre altres casos d’adaptació fisiològica interessants, podem citar l’adaptació cromàtica. Hi ha força espècies de cianofícies que poden modular la síntesi de ficoeritrina, i en alguns casos, també la de ficocianina, per adaptar llur composició en pigments al color de la llum que els arriba. Una substància química sensible, un fitocrom, "s’adona" de la mena de llum que arriba i actua sobre la transcripció dels gens, activant així la síntesi dels pigments que fan falta. Així, si la llum és verda, les cianofícies poden esdevenir més rosades o vermelloses.
També són molt plàstiques pel que fa a la mena de fotosíntesi que realitzen. A la superfície dels sediments dels llacs, bassals, salines, etc., passa sovint que, bona part del dia, les condicions són aeròbiques, mentre que de nit l’activitat bacteriana de descomposició esgota les reserves d’oxigen: les condicions esdevenen, doncs, reductores i comença a acumular-se gas sulfhídric, la presència del qual pel damunt d’un cert límit inhibeix el fotosistema II. Com a conseqüència, quan torna la llum al matí, les cianofícies comencen a funcionar com els bacteris fotosintetitzadors, i només fan servir H2S com a donador d’hidrogen. El mateix passa a les fonts pudents, riques en H2S. El fotosistema I és pobre en ficobiliproteïnes, responsables de la fluorescència de les cianofícies. Per això una mateixa cianofícia és fluorescent en medi oxidant (rica en ficobiliproteïnes del fotosistema II) i molt poc en medi reductor. Aquesta adaptació ha estat ben estudiada a Microcoleus, Spirulina, Phormidium, Oscillatoria limnetica (la més ben adaptada) i Aphanothece halophytica.
Pel que fa a la fixació de nitrogen, ja hem parlat del paper dels heterocists. Ara bé, recentment s’ha vist que algunes cèl·lules vegetatives també poden fixar nitrogen, sempre que evitessin acumular oxigen, el qual inactiva l’enzim nitrogenasa, la clau del procés. Això és possible si la cianofícia queda a les capes anòxiques de l’aigua, o si es protegeix de l’oxigen amb beines mucilaginoses prou gruixudes (com a Microcoleus i a Gloeothece) o amb altres estratègies metabòliques, capaces de mantenir una tensió d’oxigen baixa.
Un altre aspecte interessant és l’extraordinària capacitat d’adaptació a concentracions de sals i, per tant, a pressions osmòtiques àmpliament fluctuants. A les salines, estanys salabrosos endorreics, cubetes litorals, etc., la concentració en sal pot esdevenir saturant per evaporació, o baixar moltíssim després d’una pluja, de forma brusca. Les cianofícies han d’equilibrar la pressió osmòtica de llur citoplasma amb la del medi, i ho fan molt bé, acumulant sucres, poliols i altres compostos orgànics o, a vegades, ió potassi (cas d’Aphanothece halophytica), tal com ho fan els bacteris halòfils extrems. És evident que, en condicions tan dures, les cianofícies tenen poca competència.
Llur capacitat per a la fotosíntesi ha estat cercada per alguns protozous, que s’hi han associat, en vida endosimbiòtica. Inicialment, hom havia confós aquestes cianel·les, amb cloroplasts d’un grup estrany d’algues, que hom anomenava glaucòfits. Ara s’ha vist bé que no són sinó un grup heterogeni d’eucariotes que s’han associat amb cianofícies. Però aquests organismes (Cyanophora, Glaucocystis, Paulinella) ens donen la pista de l’origen dels cloroplasts per endosimbiosi.
Cada dia es descobreixen noves adaptacions i astúcies en la fisiologia d’aquest interessant grup. I, encara, els procariotes que han arribat fins a nosaltres, no són sinó una petita part dels que fa 1000 milions d’anys dominaven la Terra.
Sistemàtica
En començar a parlar de les cianofícies explicàvem com el seu estudi amb tècniques microbiològiques ha portat a una manera nova de considerar-les, reflectida en l’encunyació del nou nom de cianobacteris, nom que vol posar de manifest que es tracta d’un grup íntimament lligat als bacteris. No caldria parlar-ne gaire més, si no fos que totes dues aproximacions, l’algològica i la microbiològica, han portat a conceptes força diferents pel que fa a la taxonomia de les cianofícies.
En efecte, tant els estudis de Drouet, fets amb material d’herbari, com els de moltes escoles de microbiòlegs, han posat de manifest que molts caràcters morfològics de les cianofícies són deguts a l’efecte de l’ambient. Per això, moltes espècies, i fins i tot gèneres, no són sinó ecotips, o formes adaptatives no heredables, causades per l’efecte dels factors ambientals. Per això, moltes espècies que es poden diferenciar al camp, ja no es poden separar morfològicament quan s’han cultivat al laboratori. Això va portar els microbiòlegs a ajuntar molts gèneres i espècies en un nombre petit de nous tàxons.
Més recentment, hom ha vist que molts caràcters lligats al genotip (és a dir, bons caràcters per a diferenciar espècies) no s’expressen en cultius de laboratori, simplement perquè les condicions ecològiques naturals no hi són i no els fan necessaris. Però les diferències genètiques hi són igualment. Per tant, cal moderar en una bona part les simplificacions introduïdes pels microbiòlegs, sense oblidar, però, que és cert que sota l’efecte de factors ecològics diferents, una mateixa espècie es pot presentar en formes força distintes a les quals la sistemàtica clàssica havia atorgat una importància exagerada. Aquesta plasticitat s’ha vist que afecta principalment la síntesi de beines i de capes mucilaginoses i les formes d’agrupament entre les cèl·lules.
La sistemàtica de les cianofícies és, doncs, delicada, i camina a poc a poc vers una revisió que tingui en compte tot el que sabem de nou. Ara bé, la riquesa morfològica de les cianofícies és un bon indicador ecològic. La utilitat de les espècies clàssiques resulta encara d’utilitzar de cara a la caracterització florística dels ambients rics en algues. El recorregut descriptiu de les cianofícies més importants dels Països Catalans efectuat en aquesta obra s’haurà de basar, doncs, amb els retocs necessaris, sobre aquest tipus de sistemàtica, basada en tal com es presenten les cianofícies al camp, per a la qual cosa seguim obres bàsiques com les de Geitler (1932), Tilden (1910), Desikachary (1959), Bourrelly (1970), Friedmann (1982), reflectides en part a Strasburger (1983). Però serà també girat l’esguard vers la sistemàtica microbiològica de l’Escola de Stanier (Rippka i col·laboradors, 1979, 1981; Watterbury, 1978), tot intentant d’establir la correlació entre ambdues escoles. Més forta fou encara la simplificació introduïda perDrouet (1956, 1968, 1978, 1981) que, si bé no ha estat gaire seguida, actuà com a revulsiu sobre els taxònoms botànics i facilità el camí crític posterior.
La taxonomia botànica distingeix una sola classe, cianofícies, en el fílum cianòfits, bé que sigui gran la quantitat d’altres noms que hom ha donat a aquests organismes: mixofícies, esquizofícies, i darrerament, cianocloronts. Hom els divideix en dos grans grups, amb categoria de subclasse: les coccogonees i les hormogonees.
Les coccogonees (Coccogoneae) són cianofícies unicel·lulars o reunides en grups (cenobis) no filamentosos, agrupades en tres ordres: el de les croococcals (Chroococcales), el de les camesifonals (Chamaesiphonales) i el de les pleurocapsals (Pleurocapsales).
Les hormogonees (Hormogoneae) són cianofícies filamentoses, amb cèl·lules més diferenciades, agrupades també en tres ordres: el de les oscil·latorials (Oscillatoriales), el de les nostocals (Nostocales) i el de les estigonematals (Stigonematales).
A continuació es presenta la classificació microbiològica de les cianofícies (cianobacteris) cultivades. Dels 170 gèneres acceptats pels ficòlegs clàssics, els microbiòlegs només n’accepten 22 (el 1984), car en els cultius de laboratori s’assimilen "espècies" d’aparença prou diferent en estat espontani. Moltes espècies clàssiques no han pogut entrar en aquest sistema, perquè no han estat encara estudiades en cultiu (per exemple, perquè es troben fixades, conservades en herbaris). Per altra banda, hom ha isolat moltes soques de cianobacteris que mai no s’ha intentat relacionar amb espècies clàssiques.
- SECCIÓ 1. Inclou els cianobacteris unicel·lulars o reunits en colònies per material capsular, que es reprodueixen per escissió binària o per gemmació. Inclou la majoria de les croococcals i totes les camesifonals.
Chamaesiphon conserva en cultiu la capacitat de gemmació pel pol apical. Gloeobacler violaceus no té tilacoides i reté els pigments a la membrana. Synechococcus (en sentit microbiològic) comprèn diversos Anacyslis, Synechococcus (entre els quals S. lividus) algunes soques de Gloeocapsa alpicola, Microcystis aeruginosa, Agmenellum quadruplicatum i alguns Aphanothece. Synechocystis reuneix moltes soques d’Aphanocapsa, algunes Gloeocapsa alpicola, Eucapsis i alguns Microcystis aeruginosa. Aquests dos exemples seran suficients per a veure com va la correspondència entre sistemàtica microbiològica i clàssica. Hom distingeix també els gèneres Gloeothece (s.m.) i Gloeocapsa. - SECCIÓ 2: Inclou els cianobacteris unicel·lulars que es divideixen per escissió múltiple. Les cèl·lules són solitàries o formen agregats pseudofilamentosos, compactats per capes exteriors de la paret, de naturalesa fibrosa.
Dermocarpa presenta endòspores mòbils, i Xenocarpella les té immòbils, degut que presenten una coberta fibrosa. Dermocarpella, Myxosarcina, Chroococcidiopsis i Pleurocapsa poden presentar també divisió binària. Aquesta secció agrupa les pleurocapsals, incloent-hi les dermocarpàcies. - SECCIÓ 3. Comprèn els cianobacteris filamentosos que creixen per divisió cel·lular intercalar i en una sola direcció. El filament només té cèl·lules vegetatives, i la dispersió es fa per hormogonis. Correspon a l’ordre clàssic de les oscil·latorials.
De 25 gèneres clàssics, els microbiòlegs accepten Spirulina, Oscillatoria (on van a parar també moltes Lyngbya i Microcoleus, que perden la beina en cultiu) i Pseudanabaena. A més, creen dos grups, LPP-A i LPP-B, on van a parar representants de molts gèneres botànics poc definits, amb beina (un caràcter molt variable), com Phormidium, Plectonema i moltes Lyngbya. - SECCIÓ 4. Comprèn cianobacteris filamentosos de creixement intercalar en una sola direcció, que poden formar heterocists i acinets. Correspon a l’ordre clàssic de les nostocals i ha sofert una important reducció en el nombre de gèneres.
Hom distingeix els que poden produir hormogonis (Nostoc, Scytonema, Calothrix) i els que presenten solament fragmentació del tricoma (Anabaena, Nodularia, Cylindrospermum). Hom no atorga valor taxonòmic a les falses ramificacions ni a la morfologia colonial. Els gèneres es distingeixen per la posició dels heterocists i dels acinets, la morfologia de la cèl·lula i la del tricoma. - SECCIÓ 5. Correspon als cianobacteris filamentosos de creixement intercalar en més d’una direcció, ço que dóna lloc a l’aparició de ramificacions veritables. Poden formar heterocits i, en alguns casos, també acinets. Correspon a l’ordre clàssic de les estigonematals.
Hom distingeix dos gèneres. Chlorogloeopsis, de tricoma que, fragmentat, s’assembla a Gloeocapsa, però presenta heterocists. El segon gènere, Fischerella, és més antic, i conté també les formes cultivades de Stigonema i de Mastiglocladus.