Concepte de bacteri
Els bacteris són organismes cel·lulars procariòtics que, per llur mida microscòpica, pertanyen al conjunt dels microorganismes. L’estructura de llur citoplasma és molt simple i no hi trobem sistemes de membranes independents que els divideixin en compartiments funcionals, bé que hi ha l’excepció dels tilacoides de les cianofícies i dels procloròfits. Als bacteris pròpiament dits hi poden haver invaginacions de la membrana citoplasmàtica de diferents tipus i, en alguns casos, poden presentar-se orgànuls envoltats per membranes diferents de la citoplasmàtica (o membrana unitària). La majoria dels bacteris, de manera semblant a les cèl·lules de les plantes, estan envoltats per una paret cel·lular més o menys rígida, que sols manca en el cas dels micoplasmes, però que pot presentar una estructura ben diversa i sempre diferent de la de les plantes.
Morfologia i organització
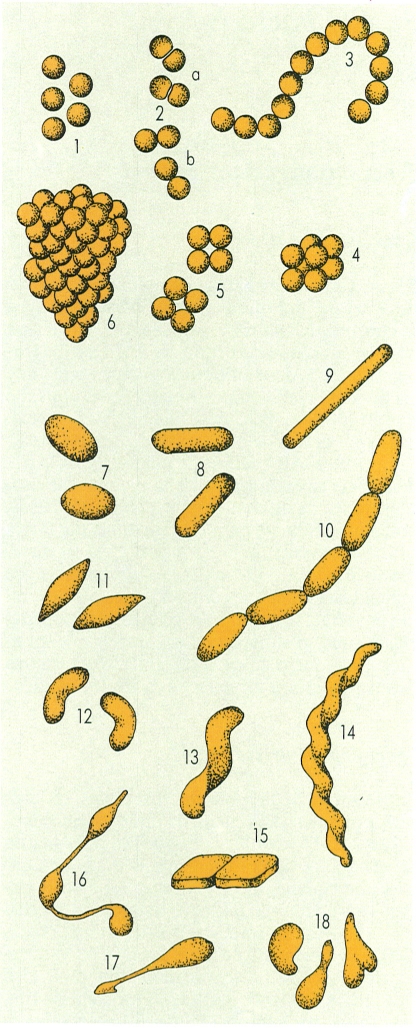
Morfologies bacterianes, idealitzades a partir de gèneres representatius: 1 cocs separats (Micrococcus); 2 parelles de cocs, aixafats al pla de contacte (a, Neisseria) i en forma de diplococ (b, Streptococcus o Diplococcus); 3 cadenetes de cocs (Streptococcus); 4 octets de cocs (Sarcina); 5 tètrades de cocs (Aerococcus); 6 raïms de cocs (Staphylococcus); 7 cocobacils (Escherichia); 8 bacils típics (Bacillus); 9 bacils filamentosos (Flexibacter); 10 cadenetes de bacils (Streptobacillus); 11 cèl·lules fusiformes (Fusobacterium); 12 vibrions (Vibrio); 13 espirils (Spirillum); 14 formes espirillars flexuoses (Spirochaeta); 15 formes laminars (Haoarcula); 16 cèl·lules amb prolongacions citoplasmàtiques (prosteques) (Hyphomicrobium); 17 cèl·lula amb prosteca amb funcions de peduncle i peu de subjecció (Caulobacter); 18 cèl·lules pleomòrfiques de tipus difteroide (Propionibacterium).
Maber, original de l’autor

Organització cel·lular dels bacteris, en el cas d’un bacteri ideal estàndard: a nucleoide, en contacte amb el citoplasma; b citoplasma amb ribosomes; c/d paret (en bacteris grampositius i gramnegatius, respectivament); e/f capa de peptidoglicà (en bacteris grampositius i gramnegatius, respectivament); g espai periplasmàtic; h membrana externa (en bacteris gramnegatius); i membrana plasmàtica; j invaginació de la membrana plasmàtica, amb pigments fotosintetitzadors o amb altres sistemes enzimàtics de membrana; k mesosoma; I/mcàpsula (de tipus compacte o fibrós, respectivament); n fímbries d’adherència; o pilus sexual de conjugació; p flagel procariòtic; q motor flagel·lar; r clorosoma (en bacteris verds del sofre); s vesícula gasífera; t carboxisoma (en bacteris autotròfics); u grànul de sofre elemental; v grànul de glucogen; x grànul de metapolifosfat (volutina); y grànul de poli-β-hidroxibutirat (PHB).
Maber, original de l’autor
Sense sortir de la mida microscòpica, els bacteris presenten diferències de dimensió molt notables. Hi ha cèl·lules petitíssimes, al plàncton de les aigües dolces i marines o a les comunitats microbianes del sòl, que gairebé no es poden veure amb els millors sistemes de microscòpia òptica. Es tracta dels ultrananobacteris, dels minibacteris i de certs estadis de vida latent que poden presentar diàmetres de 0,15 a 0,2 µm. A l’extrem oposat, hi ha bacteris filamentosos, com ara Beggiatoa gigantea, que poden atènyer 60 µm de diàmetre, i altres, més o menys esfèrics, com Thiovulum majus, en els quals el diàmetre pot arribar a 30 µm. Com que aquests darrers són nedadors, poden ser fàcilment confosos amb protozous per un observador poc experimentat. Molts apèndixs i materials que envolten els bacteris són de mida submicroscòpica i no poden ser resolts sense l’auxili d’un microscopi electrònic. Així doncs, molts detalls estructurals dels bacteris queden amagats al naturalista afeccionat, que empra com a peça més sofisticada d’observació el microscopi òptic.
La paret cel·lular

Paret buida d’un bacteri prostecat mort, del gènere Prosthecomicrobium, trobat en una bassa d’aigua dolça, vist al microscopi electrònic de transmissió (tinció amb acetat d’uranil). La paret bacteriana, que és rígida, ha conservat la forma del bacteri viu.
Francesc Torrella
La paret cel·lular, que és rígida a causa de la capa de mureïna que conté, dona forma a la cèl·lula bacteriana. Es fàcil de comprovar-ho, perquè quan la cèl·lula mor i el protoplasma desapareix, en resta la paret buida, que conserva la forma de la cèl·lula originària.
En els bacteris podem trobar dos tipus principals de paret, que es corresponen molt bé amb les propietats tinctorials de la cèl·lula bacteriana: la paret dels bacteris grampositius i la dels bacteris gramnegatius. Les parets grampositives, com les de l’estafilococ daurat (Staphylococcus aureus), tenen una capa de mureïna molt gruixuda, i no presenten membrana externa; en canvi, les parets dels bacteris gramnegatius presenten una capa de mureïna prima i, a més a més, estan envoltades per una membrana externa, com en el cas del bacil de la febre tifoide (Salmonella typhi). Aquesta membrana externa presenta una estructura molt semblant a la plasmàtica, però és ben diferent pel que fa a la composició i a les activitats enzimàtiques i d’altres tipus que hi són portades a terme. Entre aquesta i la membrana citoplasmàtica queda un espai molt ric en enzims, anomenat periplasma.
A la paret dels bacteris hi ha diversos tipus de molècules exclusives dels procariotes. n’és la més típica el peptidoglicà, que constitueix la substància anomenada mureïna, un polímer format per llargues cadenes de polisacàrid, constituït per la unió de n-acetilglucosamina i àcid α-acetilmuràmic, travades entre elles per curts polipèptids. Aquest polímer es troba tant als bacteris grampositius com als gramnegatius. Un altre tipus de molècula pròpia dels bacteris és el lipopolisacàrid, de la membrana exterior de les cèl·lules gramnegatives. El lipopolisacàrid és format per àcids grassos, n-acetilglucosamina, grups èster-difosfat i diferents glúcids, que varien molt segons el microorganisme de què es tracti.
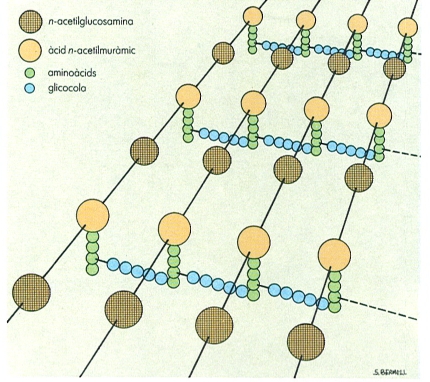
Estructura macromolecular del peptidoglicà o mureïna, el polímer estructural de paret més típic dels procariotes. En aquest cas es tracta del peptidoglicà de l’estafilococ daurat (Staphylococcus aureus), bacteri grampositiu.
Maber, a partir de dades de l’autor
Cada vegada es fa més evident que les parets dels bacteris són més variades del que podríem pensar, si només consideréssim la divisió en grampositives i en gramnegatives. En alguns bacteris, amb l’ajut de la microscòpia electrònica, hom ha detectat la presència de capes suplementàries a la paret, generalment de constitució proteica. En trobem per exemple, al coc Deinococcus (= Micrococcus) radiourans, un microorganisme especialment resistent a molts tipus de radiació. Cal pensar també que, als bacteris arcaics, la constitució de la paret és especial, ja que no hi ha gens de peptidoglicà.
Les capes situades al defora de la paret
Més enllà de la paret, molts procariotes tenen una càpsula més o menys conspicua, formada per llargues cadenes de polisacàrids o, a vegades, integrada per polipèptids, com en el cas del bacil del carbuncle (Bacillus anthracis). Les càpsules bacterianes compleixen moltes funcions. Poden actuar com a protecció contra la fagocitosi, com passa amb alguns patògens, com el pneumococ (Streptococcus pneumoniae). Hom pensa també que la càpsula protegeix la cèl·lula en els casos d’assecament ràpid. Per altra banda, les càpsules i els materials mucilaginosos que s’hi relacionen són molt útils per a fixar els bacteris sobre els diversos substrats. Així passa al tub digestiu de l’home i de molts altres animals, i a la superfície de les pedres i de qualsevol altre objecte viu o inanimat submergit a les aigües corrents, com ara la superfície dels peixos, les algues bentòniques, etc. Quan aquests materials de subjecció són prou consistents i formen una estructura definida a la base de la cèl·lula, constitueixen els glicocalzes.
El citoplasma
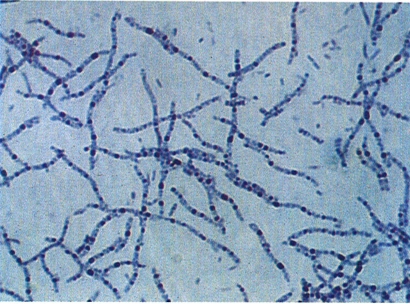
Inclusions de volutina prominents i metacromàtiques, després d’una tinció simple amb blau de metilè, en el bacteri Spirillum volutans d’un cultiu d’enriquiment amb aigua i restes vegetals en descomposició.
Francesc Torrella
El citoplasma dels bacteris es veu, al microscopi electrònic, finament granulós, a causa dels ribosomes procariòtics (d’uns 10 nm) que conté. En general, la zona central del citoplasma és ocupada pel nucleoide, que no està separat de la resta del protoplasma per membranes però se’n distingeix bé, a les micrografies electròniques, pel seu aspecte fibrós. Les tècniques de fixació i de tinció determinen que el genòfor del bacteri, constituït per l’ADN, es condensi formant el nucleoide. Però és freqüent que els bacteris tinguin també al citoplasma altres fragments d’ADN que també contenen informació genètica. Es tracta dels plàsmids. Tant el genòfor com els plàsmids es posen en contacte amb la membrana plasmàtica quan s’acosta el moment de la divisió cel·lular, i és sobretot en aquest període quan són més fàcils de distingir determinades invaginacions de la membrana que formen capes concèntriques, que recorden una ceba tallada pel mig. Se’n diuen mesosomes, i no és segur que en alguns casos no siguin simples artefactes (alteracions produïdes per les tècniques d’observació).
Com que als bacteris no hi ha reticle endoplasmàtic ni cap altre orgànul format per membranes comparable als que hi ha a la cèl·lula eucariòtica, la membrana citoplasmàtica dels bacteris, constituïda per fosfolípids i molt rica en proteïnes, i d’uns 7,5 nm de gruix, és la seu d’una intensíssima activitat enzimàtica. A la membrana i a les invaginacions que pugui formar, es troben les proteïnes responsables del transport dels aliments, de les cadenes respiratòries, els enzims relacionats amb un cert tipus de síntesi d’ATP, moltes menes de pigments fotosintètics, els llocs de reconeixement del genòfor per a la divisió cel·lular, etc.
A l’interior del citoplasma hi ha alguns petits orgànuls envoltats de finíssimes membranes proteiques, de 3 a 5 nm de gruix. En són un exemple els vacúols gasífers, idèntics als dels cianòfits, a l’interior dels quals s’acumulen gasos dissolts en el medi. Els bacteris els empren també per a la flotació. Altres exemples són els carboxisomes, que contenen enzims responsables de la fixació del diòxid de carboni, en el cas dels bacteris autotròfics, i els clorosomes o "vesícules de Chlorobium", que guarden els pigments fotosintetitzadors dels bacteris fotosintètics verds. A més d’aquests orgànuls, al citoplasma dels bacteris s’hi troben diferents inclusions. Els més freqüents són els grans de glucogen i els de polibetahidroxibutirat (PHB), tots dos amb paper de reserva energètica carbonada, els grans de sofre elemental, reserva energètica en els bacteris oxidadors de sofre, i els grans de volutina o metapolifosfat, que serveixen de reserva de fòsfor en una gran quantitat de bacteris.
Les prosteques
Són prolongacions del citoplasma i de la paret cel·lular que es poden trobar en alguns tipus de bacteris. En algun cas, sembla que les prosteques tenen com a finalitat l’augment de la superfície del microorganisme, per tal de poder aconseguir més aliments, ja que a tota la superfície de la membrana plasmàtica hi ha transportadors d’aliments. Aquest és el cas d’Hyphomicrobium, Ancalomicrobium, etc. Altres vegades, la prosteca, a més de la utilitat ja descrita, es fa servir com a estructura de subjecció al substrat, com passa a Caulobacter i a Asticaccaulis.
Els flagels i les fímbries
Els flagels bacterians són orgànuls filamentosos rígids, constituïts per una proteïna anomenada flagel·lina. Normalment, el seu diàmetre està comprès entre 12 i 18 nm, però els que porten beina poden tenir un gruix de fins a 30 nm. Tenen forma sinusoide (és a dir, uniformement ondulada), amb una longitud d’ona regular i fixa, ja que són rígids. Els flagels són implantats a la membrana plasmàtica mitjançant un corpuscle basal constituït per dos discs, el motor flagel·lar, que fa rodar els flagels, sense que aquests tinguin cap altra capacitat pròpia de moviment. Segons sigui el sentit del seu moviment rotatori, produeixen diferents efectes translacionals en la cèl·lula. Constatem, doncs, que aquests flagels simples i rígids tenen poc a veure amb els flagels més gruixuts, amb activitat pròpia d’ondulació i envoltats per membrana citoplasmàtica dels eucariotes, per als quals hom proposa el nom d’undulipodis.
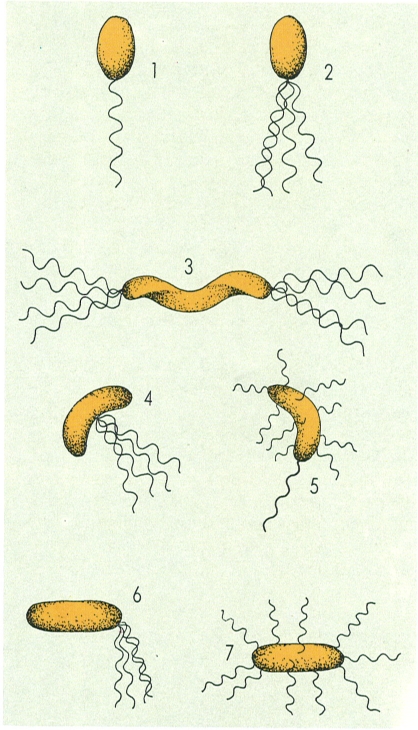
Disposicions flagel·lars bacterianes, idealitzades a partir d’exemples representatius: 1 polar lofòtrica (Pseudomonas); 2 polar lofòtrica (Pseudomonas); 3 polar amfítrica (Spirillum); 4 lateral i umbilical (Selenomonas ruminantium); 5 perítrica composta (Vibrio de procedència marina, amb flagels senzills perítrics i un flagel polar amb beina); 6 lateral i subterminal (Sphaerotilus natans, en cèl·lules dispersives); 7 perítrica clàssica (Escherichia coli).
Assumpció Cañadas/Maber, original de l’autor
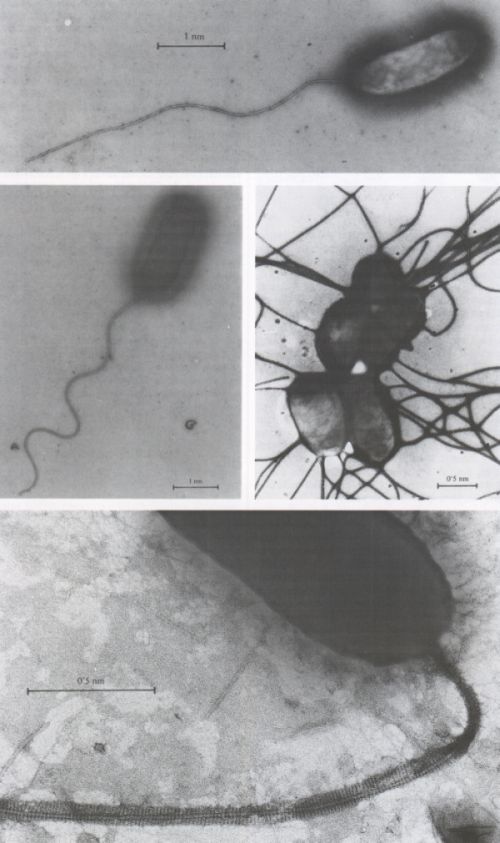
Flagels bacterians vistos al microscopi electrònic de transmissió (tinció amb acetat d’uranil). A dalt, flagel polar, sense beina, d’un Pseudomonas marí. Al centre, a l’esquerra, flagel polar amb beina, d’un Vibrio marí. Al centre, a la dreta, cèl·lules agrupades de Salmonella typhimmium amb flagels perítrics. A baix, una estructura de la beina d’un flagel polar d’un bacteri d’aigua dolça.
Francesc Torrella
Les fímbries són filaments més fins que els flagels, però també de natura proteica. Solen tenir entre 4 i 8 nm de diàmetre i el seu aspecte es rígid, i no ondulat. Compleixen diferents funcions, però la més freqüent és la de servir d’òrgans de reconeixement dels substrats físics facilitant que es realitzi la fixació mitjançant adsorció. Hi ha un cert tipus de fímbries, els pèls sexuals bacterians o pili (singular, pilus), que serveixen per a fer possible el pas de l’ADN d’una cèl·lula a l’altra durant el procés de la conjugació bacteriana.
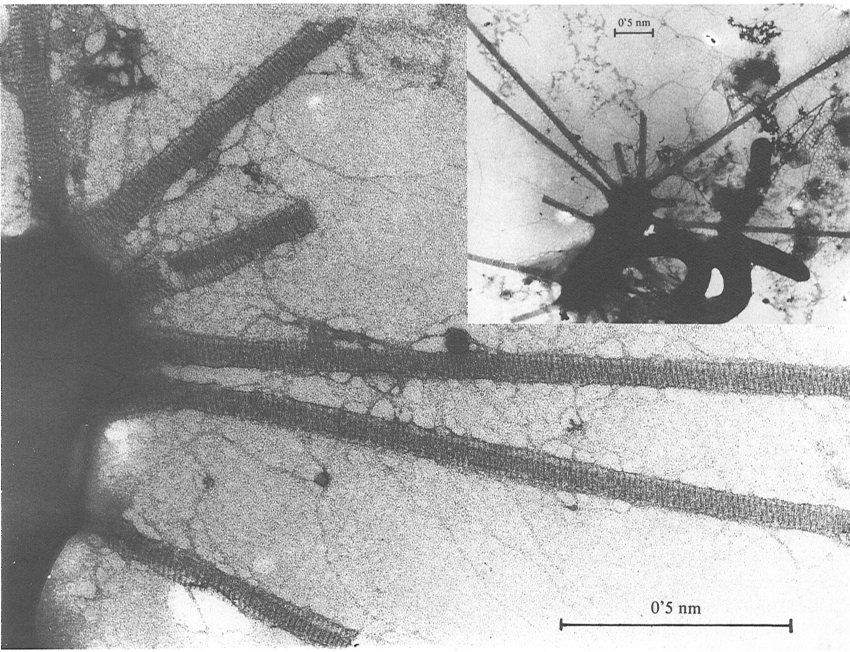
Bacteri planctònic amb espines procedent d’una bassa d’aigües eutrofitzades, vist al microscopi electrònic de transmissió (tinció amb acetat d’uranil). Al requadre, aspecte general; a la figura grossa, detall de la ultrastructura de les espines.
Francesc Torrella
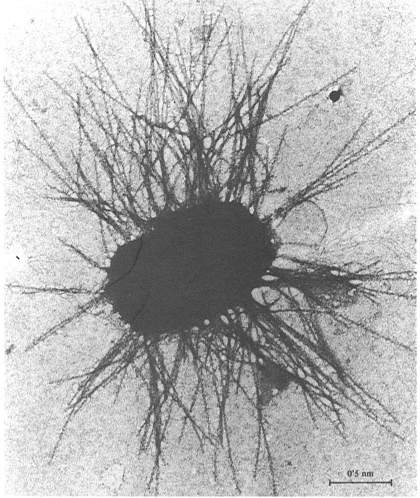
Fímbries en un bacteri planctònic d’una bassa eutròfica d’aigües dolces, vistes al microscopi electrònic de transmissió (tinció amb acetat d’uranil). Les fímbries poden servir al bacteri per a mantenir-se en l’ambient planctónic o per a reconèixer el substrat abans de fixar-s’hi.
Francesc Torrella
Alguns bacteris presenten unes estructures rígides, acabades en punta vers llur extrem i d’amplada considerable a la base (20-45 nm). Es tracta de les espines bacterianes, trobades en bacteris planctònics marins, les quals també poden aparèixer en bacteris planctònics d’aigua dolça. El seu paper podria ser el de frenar la sedimentació del bacteri, però també poden representar un mecanisme de defensa contra la depredació per petits protozous devoradors de bacteris que són molt abundants on es troben els bacteris amb espines.
Motilitat
El moviment actiu dels bacteris és de dos tipus: mitjançant flagels o per lliscament. En el cas dels bacteris amb flagel·lació polar, la rotació dels flagels en un sentit o en un altre fa que la cèl·lula avanci cap endavant o tiri enrera, segons un mecanisme de control de base bioquímica molt complexa, anomenat quimiotaxi o fototaxi. Als bacteris amb flagels perítrics, un sentit de rotació fa que la cèl·lula avanci, ja que provoca l’agrupació dels flagels en un pol cel·lular. En canvi, el sentit de rotació contrari escampa els flagels, i la cèl·lula es detura i es posa a girar sobre ella mateixa.
El moviment flagel·lar és molt comú entre els bacteris, els quals poden així traslladar-se en el si dels medis aquàtics, sempre que hi hagi energia suficient per a fer rodar els flagels. En llocs pobres en aliments es veuen freqüentment molts bacteris planctònics sense flagel, d’una mida molt petita i amb més o menys estructures fibrillars que els ajuden a romandre en el si de les aigües, aprofitant així els remolins tèrmics i de difusió que s’hi formen. En aquests casos sembla, doncs, que sigui més econòmic "deixar-se portar que anar a buscar".
Les espiroquetes presenten un cas de moviment flagel·lat molt especial, ja que en aquests microorganismes els flagels polars (fibres axials) es disposen helicoïdalment entorn del cos cel·lular, formant l’anomenat filament axial. Els flagels queden protegits per una membrana trilamel·lar i les seves puntes arriben a tocar-se a l’equador de la cèl·lula. Aquesta estructura tan complexa permet una rica gamma de moviments: translació endavant i endarrere, moviment com el d’un llevataps en medis viscosos, i diverses modalitats de moviment flexuós en medis líquids.
El moviment de lliscament es troba amb freqüència en els bacteris adaptats a la vida sobre les superfícies i a les interfases que poden canviar les seves propietats físico-químiques i nutricionals en períodes breus de temps. Aquest moviment és lent, però suficient perquè els bacteris, que solen ser clarament filamentosos o allargats, puguin traslladar-se d’un punt a l’altre de llur microambient, aprofitant així les condicions més favorables en cada moment.
Fisiologia
En el decurs dels milions d’anys d’evolució sobre la Terra, els procariotes no han donat lloc a una diversitat morfològica i estructural tan gran com la que trobem en els eucariotes. En canvi, els organismes de cèl·lula primitiva sí que han tingut un èxit extraordinari pel que fa a la multiplicitat de possibilitats d’adaptacions fisiològiques i metabòliques a la gran quantitat de macro i de microhàbitats que han anat apareixent sobre el planeta des de la seva formació. La diversitat nutricional dels bacteris va molt més enllà dels esquemes clàssics que divideixen els sers vius superiors en autòtrofs (vegetals amb clorofil·la) i heteròtrofs (fongs, protozous i metazous). A continuació, passem revista a la gran varietat de tipus tròfics que es troben en el món bacterià.
Els tipus tròfics
Des del punt de vista del seu nodriment (trofisme) i segons la font de carboni que utilitzin, els sers vius poden dividir-se en autòtrofs (principal font de carboni, el CO2) i heteròtrofs (molècules orgàniques com a principal font de carboni). Per altra banda, i segons la font d’energia, la divisió dels sers vius s’estableix en dos grups: fotòtrofs (principal font d’energia, la llum) i quimiòtrofs (principal font d’energia, un compost químic que s’oxida). En el metabolisme energètic d’una cèl·lula, és tan important la mateixa font d’energia com aquelles molècules que funcionen com a acceptors terminals d’electrons o equivalents reductors (àtoms d’hidrogen). La seva presència en una cèl·lula és la que permet tota una sèrie de reaccions bioquímiques lligades a l’obtenció d’energia en una forma fàcilment utilitzable, l’ATP.
Atenent a les categories esmentades més amunt i combinant totes les possibilitats, el món bacterià presenta una diversitat remarcable, que es concreta i es descriu amb els següents termes: fotoautòtrofs, fotoheteròtrofs, quimioautòtrofs i quimioheteròtrofs. Per altra banda, la relació dels diferents metabolismes bacterians amb l’oxigen dona peu a una rica terminologia en la qual destaquen termes com aerobi, anaerobi, microaeròfil i aerotolerant. A més, molts bacteris, a nivell individual, són versàtils en el tipus de metabolisme que poden presentar. Aquesta versatilitat és induïda i modulada pels paràmetres ambientals, que permeten que funcioni un tipus o altre de metabolisme. Tot això, com es veurà més avall, es concreta en els termes estricte i facultatiu afegits als tipus tròfics bàsics.
Els tipus de metabolisme microbià més importants, amb alguns exemples concrets, són els que segueixen. Els fotoautòtrofs són bacteris que utilitzen la llum com a font d’energia i el CO2 com a principal font de carboni; entre els bacteris fotosintètics n’hi ha molts que pertanyen a aquesta classe, com ara els Chromatium (bacteris vermells del sofre) i Chlorobium (bacteris verds del sofre). Els fotoheteròtrofs són bacteris que empren la llum com a font d’energia i molècules orgàniques com a font de carboni; hi ha representants d’aquest grup entre els bacteris vermells fotosintetitzadors (Rhodospirillum) i entre els bacteris verds fotosintetitzadors (Chloroflexus). Els quimioautòtrofs són també coneguts amb el nom de quimiolitòtrofs, i utilitzen compostos inorgànics reduïts com a font d’energia (H2, CO, NO-2, NH+4, SH2, S°, S2O3=, Fe+2) i el CO2 com a font de carboni principal (el terme litòtrofs —del grec lithos, roca— prové de la seva capacitat de créixer en medis estrictament minerals); són exemples d’aquest grup els oxidadors d’hidrogen (Hydrogenomonas), els oxidadors de nitrits (Nitrobacter), els oxidadors de compostos reduïts de sofre (Thiobacillus) i els oxidadors de l’ió ferrós (Gallionella). Els quimioheteròtrofs utilitzen un compost químic orgànic com a font de carboni (la majoria de vegades aquest mateix compost és font d’energia); la major part dels bacteris cultivats al laboratori pertanyen a aquest grup, juntament amb els patògens de plantes i animals, i en són exemples el bacil coliforme (Escherichia coli), el vibrió del còlera (Vibrio cholerae), el bacil de Koch (Mycobacterium tuberculosis) i molts d’altres.
Quan es parla del CO2 com a font de carboni cal tenir en compte que tots els sers vius poden incorporar aquesta molècula mitjançant les reaccions metabòliques de carboxilació, però només els autòtrofs poden emprar el diòxid de carboni com a font principal del carboni cel·lular.
Els tipus respiratoris
Des del punt de vista de l’acceptor terminal d’electrons, que permet el funcionament de les cadenes respiratòries, es distingeixen entre els bacteris diferents categories metabòliques.
Són respiradors aerobis els bacteris que poden emprar l’oxigen com a acceptor terminal d’electrons, transformant-lo en aigua, per reducció, en el procés respiratori; en són exemples típics els bacteris del gènere Pseudomonas, molt abundants en sòls i aigües, els Bacillus, comuns a la microflora edàfica, els estreptomicets del gènere Streptomyces, productors d’antibiòtics i propis del sòl, etc. Els bacteris microaeròfils poden respirar l’oxigen, però no resisteixen tensions parcials altes d’aquest gas, de manera que viuen en interfases en què l’oxigen encara és present, però no pas abundant; en són un exemple algunes espècies del gènere Spirillum.
Els anaerobis són organismes que poden viure (anaerobis facultatius) o han de viure (anaerobis estrictes) sense oxigen. Són anaerobis facultatius els bacils coliformes i el vibrió del còlera, citats abans. Els bacteris que no poden resistir la presència de l’oxigen són abundants al tracte intestinal dels mamífers. Posem per exemple els Bacteroides, Clostridium, etc. Encara hi ha un altre grup d’anaerobis, els anaerobis aerotolerants, que no poden utilitzar l’oxigen, però no moren en presència d’aquest, microorganismes com els bacils i cocs de la fermentació làctica del iogurt (Lactobacillus bulgaricus i Streptococcus thermophilus) són exemples d’aquest grup.
Els microorganismes anaerobis que fan servir substrats orgànics com a font de carboni i d’energia s’anomenen fermentadors. Obtenen el seu ATP per les fosforilacions a nivell de substrat que es duen a terme en el citoplasma. En són un bon exemple els clostridis (Clostridium), responsables de les fermentacions butíriques, les activitats dels quals van ser estudiades per Pasteur i li permeteren de descobrir el que ell anomenà "la vida sense aire".
A més dels que són fermentadors, molts bacteris anaerobis poden també respirar, ja que no fan servir l’oxigen, sinó altres acceptors terminals d’electrons, de tipus inorgànic. Tenim entre ells els reductors de sulfats, que fan servir l’ió sulfat, i el redueixen fins a sulfur. Són molt abundants als fangs anaeròbics de la mar i de les aigües dolces, que presenten un color negre a causa de llur activitat, ja que el sulfur que es forma precipita l’ió ferrós en forma de sulfur de ferro, de color negre. Diferents gèneres tenen aquests tipus de metabolisme. Un dels més estudiats és Desulfovibrio. Els bacteris desnitrificants fan servir nitrats i nitrits com a acceptors terminals d’electrons. Converteixen aquestes molècules en els gasos N2 i N2O, que es perden a l’atmosfera i representen una pèrdua més o menys important del nitrogen combinat que hi ha en els medis naturals. Moltes soques dels gèneres Pseudomonas i Bacillus són desnitrificants. Els bacteris productors de gas metà o metanògens fan servir el diòxid de carboni com a acceptor d’electrons i el converteixen en metà (CH4), Aquests microorganismes abunden en ambients anaeròbics molt reductors, com ara els fangs del fons de les aigües, els tancs de digestió de residus fecals, etc. Els gèneres Methanobacterium i Methanococcus són ben coneguts.
Un tipus de grup fisiològic molt important entre els procariotes és el dels fixadors de nitrogen molecular (N2). Llur capacitat metabòlica és sorprenent, ja que aquests microorganismes, alguns de vida lliure, com els Azobacter, i altres de vida simbiòtica, com els Rhizobium de les lleguminoses, poden reduir el nitrogen en forma de gas fins al nivell d’amoníac, que queda incorporat al material cel·lular en forma d’aminoàcids.
Els pigments

Els Pigments fotosintetitzadors dels diferents procariotes es manifesten per les coloracions distintes dels flascons que en contenen, en cultiu enriquit: a dalt, Oscillatoria redeckei, cianòfit de color verd (clorofil·la a) i Haloarcula, bacteri halòfil de color rosat (bacteriorodopsina); a baix, columna d’aigua de mar, sobre un fang reductor negre, amb bacteris verds del sofre (bacterioclorofil·la i carotenoides).
Francesc Torrella
La capacitat fotosintetitzadora dels sers vius, fins fa pocs anys, es considerava que sempre anava lligada als pigments clorofíl·lics: les clorofil·les dels vegetals superiors i les bacterioclorofil·les, en el cas dels procariotes fotosintetitzadors. Darrerament, hom ha descobert que hi ha un grup de procariotes que tenen un pigment anomenat bacteriorodopsina i que són capaços de portar a terme la transformació d’energia lumínica en energia química. Es tracta del grup dels bacteris halòfils extrems (Halobacterium i Haloarcula) on la bacteriorodopsina i altres pigments similars, situats a la membrana, són excitats pels fotons i originen un gradient de protons que fa possible la síntesi d’ATP.
Reproducció
La multiplicació de les cèl·lules bacterianes es realitza per escissió vegetativa. En els procariotes no s’han trobat sistemes de reproducció sexual semblants als que coneixem en els eucariotes. Això no obstant, els bacteris presenten sistemes primitius que permeten una certa barreja de la informació genètica, mitjançant el pas de fragments del genòfor d’unes cèl·lules a les altres, seguits de fenòmens de recombinació que estabilitzen els nous gens en el genoma de la cèl·lula receptora. D’aquesta manera, la cèl·lula receptora enriqueix el seu patrimoni genètic.
El sistema més comú de divisió de la cèl·lula bacteriana, que és el que normalment s’observa en els microorganismes que es cultiven en el laboratori, és l’escisió binària. En aquesta, la cèl·lula mare, que va augmentant la superfície de la paret i de la membrana de forma homogènia, tot afegint nou material a les diferents zones de la perifèria cel·lular, forma un septe en posició equatorial que origina dos bacteris fills de la mateixa mida. En aquesta situació pot dir-se que les cèl·lules bacterianes no envelleixen, ja que les dues cèl·lules filles s’han renovat i tenen les mateixes característiques. El septe es forma per invaginació de la membrana plasmàtica, en forma d’anell, que es va tancant cap al centre. Durant aquest procés d’escissió, és freqüent veure mesosomes lligats a la membrana, vora el septe en formació. El genòfor del bacteri que ja s’ha duplicat abans de la constitució del septe, és segregat en ambdues cèl·lules filles, ja que resta unit a les membranes plasmàtiques que queden separades pel septe. Molts cops, i principalment quan el medi és ric, la duplicació del genòfor bacterià és més ràpida que la formació del septe. A causa d’això és freqüent trobar a l’interior d’un mateix bacteri diversos genòfors. Aquests queden separats en sengles cèl·lules filles quan el bacteri passa a un medi pobre.
Quan hom observa directament els bacteris de les aigües o dels sòls amb un microscopi de contrast de fases o electrònic, de seguida es fa patent que el tipus de divisió per escissió binària no és pas l’únic comú en el món bacterià. Es molt freqüent que els bacteris, sobretot els que resten adherits a substrats sòlids, presentin una escissió desigual, que dona lloc a formes que recorden les gemmes dels llevats. Aquest tipus de divisió es produeix quan la cèl·lula mare fa crèixer la paret només en un dels pols de la cèl·lula. Quan ja s’està formant la cèl·lula filla, una còpia del genòfor va a parar al nou volum de citoplasma. En acabar la divisió la cèl·lula filla és més petita què l’originària, que resta doncs com una cèl·lula "vella". És freqüent que els bacteris que són fruit d’una escissió desigual presentin flagels o altres tipus de motilitat que afavoreixen la dispersió de l’espècie.
En alguns bacteris filamentosos, com en el cas del bacteri verd Pelodictyon clathrathiforme, hi ha divisió ternària, que fa possible la formació de malles de filaments.
Cèl·lules de dispersió i de resistència
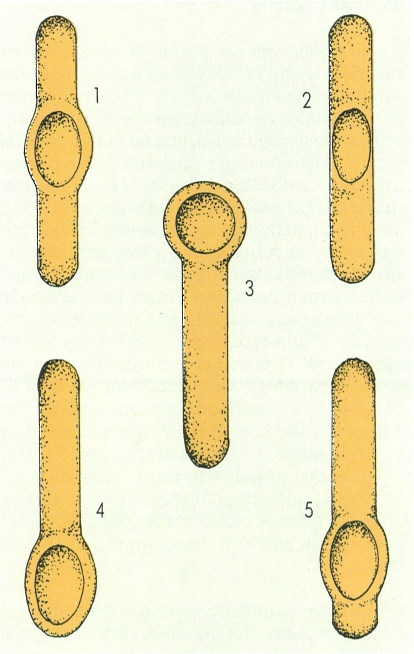
Morfologia i disposició de l’endòspora a l’interior de la cèl·lula bacteriana mare o esporangi, idealitzades a partir d’exemples representatius: 1 central, amb esporangi dilatat (Bacillus popilliae); 2 central (B. licheniformis); 3 terminal i esfèrica, amb esporangi dilatat (B. pasteurii); 4 terminal i el·líptica, amb esporangi dilatat (B. macerans); 5 subterminal, amb esporangi dilatat (B. polymyxa).
Assumpció Cañadas/Maber, original de l’autor
Tots els bacteris presenten una forma de dispersió de l’espècie que els permet de superar condicions ambientals adverses i mantenir un estat de vida latent (criptobiosi) a l’espera d’una situació favorable. En uns casos, el procés morfogenètic que porta a la formació d’una cèl·lula dispersiva és molt elaborat, i implica el joc de controls moleculars molt sofisticats. Un cas extrem és el dels bacteris que formen endòspores. En altres casos, com ara en molts bacteris aquàtics, les cèl·lules vegetatives, quan es troben en condicions de pobresa en substàncies nutritives, es fan molt petites i mantenen uns nivells baixíssims d’activitat metabòlica. En alguns bacteris edàfics que no formen endòspores, l’estratègia consisteix a embolcallar el citoplasma amb capes de material capsular, alhora que es compacten i engruixeixen les capes de la paret cellúlar , a fi i efecte d’evitar al màxim la deshidratació del protoplasma.
El gran grup dels actinomicets presenta una fragmentació dels filaments vegetatius en els casos en què no hi ha formació de cossos fructífers. Les espècies més evolucionades presenten conidis, que es formen a l’àpex de rudimentaris cossos fructífers (conidiòfors). Les espècies terrestres tenen conidis de superfície molt estructurada (espícules, costelles, etc), adaptats a la dispersió aèria. Les espècies aquàtiques tenen conidis dotats de flagels procariòtics.
Les cèl·lules vegetatives dels mixobacteris, quan es troben en situació de pobresa de substàncies nutritives, es transformen en mixòspores, que són cèl·lules de vida latent que presenten un engruiximent de la paret i que encara poden embolcallar-se amb una càpsula densa. En aquest darrer cas reben el nom de microcists. Tant les unes com les altres s’acumulen en cossos fructífers fets de mucílag, que presenten una morfologia típica de cada espècie. Els bacteris del gènere Azotobacter també formen cists, en resposta a una situació ambiental adversa. Aquests cists són plens de materials de reserva i estan envoltats de capes suplementàries, a l’exterior de la paret bacteriana.
Tant els conidis dels actinomicets com els cists esmentats més amunt són més resistents a la deshidratació, a la radiació ultraviolada i ionitzant, i a la desintegració mecànica, que les cèl·lules vegetatives. En canvi, la seva resistència a les temperatures elevades no és especialment notable. Passa el contrari en el cas de les estructures de vida latent originades pels bacteris dels gèneres Bacillus, Clostridium i alguns altres.
Aquestes cèl·lules de resistència i de dispersió, anomenades endòspores, a més de la resistència pròpia dels cists, presenten una gran resistència a la calor. En alguns casos, són necessàries temperatures de 121 °C mantingudes durant 15 minuts per a poder matar algunes endòspores de bacteris. L’ultrastructura de les endòspores és molt complexa, i hi podem distingir, de dins a enfora, el cos cel·lular, amb la membrana i la paret corresponent, el còrtex, la coberta de l’espora i una fina capa exterior o exospori. Hom creu que l’extraordinària resistència de les endòspores a la calor es produïda per la riquesa del seu citoplasma en una substància anomenada dipicolinat de calci. La posició de l’endòspora dins de la cèl·lula és variada i característica de cada microorganisme.
Els microorganismes dels gèneres Leucothrix i Thiothrix, que formen cadenetes de cèl·lules fixades sobre substrats submergits, donen lloc a cèl·lules dispersives, anomenades gonidis, amb capacitat de moure’s per lliscament. Hi ha encara molts altres casos de cèl·lules dispersives pròpies dels bacteris aquàtics i també d’alguns patògens, totes caracteritzades per la reducció de mida de la cèl·lula vegetativa i el pas a un metabolisme molt alentit. El cas dels espirils d’aigua dolça i marina (Spirillum i Oceanospirillum) i el dels vibrions marins (Vibrio) és ben documentat. També entre els patògens, els micoplasmes i les clamídies formen cèl·lules molt petites, que a vegades no arriben ni als 0,2 µm. Reben el nom de corpuscles elementals en el primer cas, i de formes coccoides en el segon.
Intercanvi genètic
Ja hem dit més amunt que els bacteris no tenen una reproducció sexual que es pugui comparar amb la típica dels eucariotes, però que presenten possibilitats de transferència gènica. La conjugació bacteriana consisteix en el pas d’ADN d’una cèl·lula donadora (que actua com a masculina), a través d’un pilus sexual o d’un pont intercel·lular, a una cèl·lula receptora (que actua com a femenina). Mitjançant aquest sistema poden ésser transferits, tant un fragment del genòfor, com un element genètic de rèplica autònoma o plàsmid. Aquest darrer pot continuar la seva dinàmica de duplicació independent un cop a la cèl·lula receptora, o bé integrar-se en el genòfor de l’hoste. Per conjugació es formen diploides parcials o merozigots, que desapareixen un cop ha tingut lloc la recombinació. S’ha comprovat que la conjugació entre bacteris té lloc normalment a la natura i que, entre altres, és el principal responsable que la resistència als antibiòtics, que generalment es troba codificada als plàsmids, s’escampi a les poblacions naturals de bacteris (on es compten mols patògens).
La transducció, un altre sistema d’intercanvi genètic, es basa en el fet que certs virus bacterians (bacteriòfags atenuats) poden establir una infecció latent en certes soques bacterianes que han estat atacades pel virus, originant cèl·lules lisogèniques. Aquestes poden acceptar fragments del genoma de la cèl·lula que donà origen al virus. D’entre totes les cèl·lules de la població lisogènica receptora, només algunes sofreixen la lisi causada pel virus. La resta conserva, i estabilitza en molts casos, trossos de genoma de la cèl·lula donadora. Es creu que la transducció juga un paper molt significatiu a la transferència de material genètic a les poblacions naturals de bacteris. La transformació consisteix en el pas d’ADN cel·lular que ha quedat lliure en el medi cap a l’interior d’una cèl·lula receptora (cèl·lula competent). La recombinació gènica, que es produeix més o menys fàcilment segons els sistemes, serà la barrera que en últim terme determinarà si el tros de l’ADN estrany que ha entrat al citoplasma serà incorporat al genòfor del bacteri receptor. Aquest sistema de transferència gènica, comprovat a la natura en el cas de diversos bacteris patògens, sembla que pot tenir importància en els casos en què les poblacions microbianes són molt denses i en què la lisi d’algunes cèl·lules, amb el subsegüent alliberament del seu ADN al medi, no és un fenomen rar.
Sistemàtica
Com ja hem dit anteriorment, els coneixements sobre la filogènia dels bacteris no són encara prou madurs per a poder permetre un ordenament "natural" d’aquests organismes. En la visió sistemàtica que oferim a continuació hem respectat les afinitats entre grups que la majoria d’autors considera com de base filogenètica. Així passa amb els bacteris arcaics, que es separen en una divisió a l’interior del regne procariotes. Les altres divisions dels procariotes, que fan referència a la composició i l’estructura dels embolcalls cel·lulars, no gaudeixen encara d’una comprovació genètico-molecular exhaustiva.
Els aspectes més discutits de tota classificació bacteriana han estat sempre les categories taxonòmiques superiors, principalment les classes i els ordres. Per això són ben poques les unitats taxonòmiques d’aquest ordre que són acceptades per una part important dels bacteriòlegs, i el lector en trobarà poques en el text que segueix. D’acord amb els texts moderns de bacteriología i, més concretament, amb les darreres edicions del Manual de Sistemàtica Bacteriana (Bergey, 8a i 9a edicions, encara [1985] en curs d’aparició), seguirem la classificació dels bacteris en grans parts o seccions. Aquestes són anomenades d’una manera inusual per a una sistemàtica de plantes o d’animals, però operativa per al bacteriòleg. A cada una d’aquestes parts s’inclouen els ordres i famílies acceptats, tot afegint-hi una sèrie de gèneres de filiació incerta. Cal dir també que, dins la brevetat del tractament de cadascun dels grups, hem donat més importància als que tenen un paper més aparent en el context de les ciències de la natura o als que poden ser fàcilment identificables amb els sistemes de microscòpia a l’abast dels afeccionats.