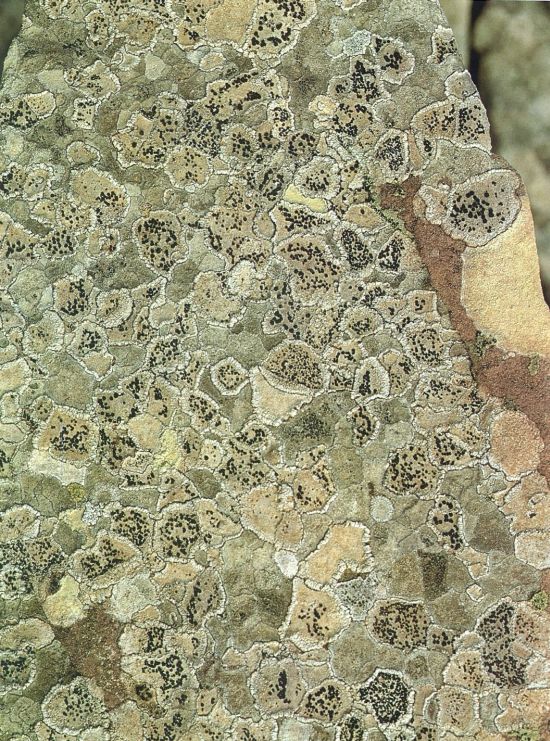
Els líquens són fongs que, gràcies a associar-se amb algues o cianofícies, han esdevingut autòtrofs com les plantes verdes i, alhora, s’han adaptat molt bé a les condicions extremes. Per això, com en la fotografia, poden colonitzar superfícies inhòspites per a altres plantes, com ara aquest bloc de gres roig, que gairebé desapareix del tot sota el mosaic polícrom dels tal·lus juxtaposats de diversos líquens, entre els quals veiem diverses espècies de Porpidia, Hafellia, Aspicilia, Rhizocarpon, Pertusaria, Lecidella, etc. En molts casos, el creixement dels tal·lus resta blocat quan dos d’immediats estableixen contacte. En d’altres, veiem com els tal·lus de creixement més ràpid comencen a recobrir-ne d’altres.
AISA.
Els líquens, ja coneguts pels grecs, han estat considerats com a organismes independents fins que S. Swendene demostrà, en dos treballs (1867, 1869), llur natura complexa, amb la intervenció d’un fong i una alga (com a mínim). Poc diferenciats pel poble, que els confon sovint amb molses i rarament els atorga noms populars, han estat objecte d’una especialitat, la liquenologia, i de l’estudi per part de botànics que només rarament s’ocupaven també dels fongs lliures. La situació continua igual en bona part, sobretot a causa de les diferències ecològiques i fisiològiques entre fongs i líquens, i també del pes de la tradició. Però des d’un punt de vista estrictament científic, l’estudi dels líquens és una part de la micologia, ja que no són altra cosa que el producte d’un procés evolutiu, la liquenificació, que ha afectat molts fongs: un de cada cinc fongs ha adoptat la forma liquen com a mètode nutricional. Els líquens que trobem ara a les roques, els arbres, els sòls, sobre les cases etc., són el resultat de la liquenificació de fongs molt diversos i de l’evolució posterior dels líquens així originats. Només associantse amb algues, els fongs liquenificats han aconseguit alliberar-se de la necessitat d’humitat de llarga durada i de matèria orgànica, i sortir a colonitzar medis aparentment inhòspits, on la majoria d’altres vegetals no poden viure. Llur èxit evolutiu els ha conduït a una forta diversificació. Les estimacions més restrictives n’accepten més de 13 500 espècies.
Un liquen és format, doncs, per un component fúngic, el micobiont, i un component fotosintetitzador, el fotobiont, que pot ésser una alga (ficobiont) o una cianofícia (cianobiont). Junts, formen una associació, un tal·lus, autosuficient, amb estructura, fisiologia i ecologia pròpies. Aquesta originalitat estructural, fisiològica i ecològica fa difícil d’explicar els líquens com a part integrada dins els fongs. A més, en grups diferents de fongs (sobretot d’ascomicets, però també de basidiomicets) trobem, poques o moltes, espècies liquenificades. Per això és més còmode i clar d’exposar els líquens com un tipus d’organització particular, recordant, sempre que calgui, les seves íntimes relacions amb diversos grups de fongs no liquenificats.
El micobiont
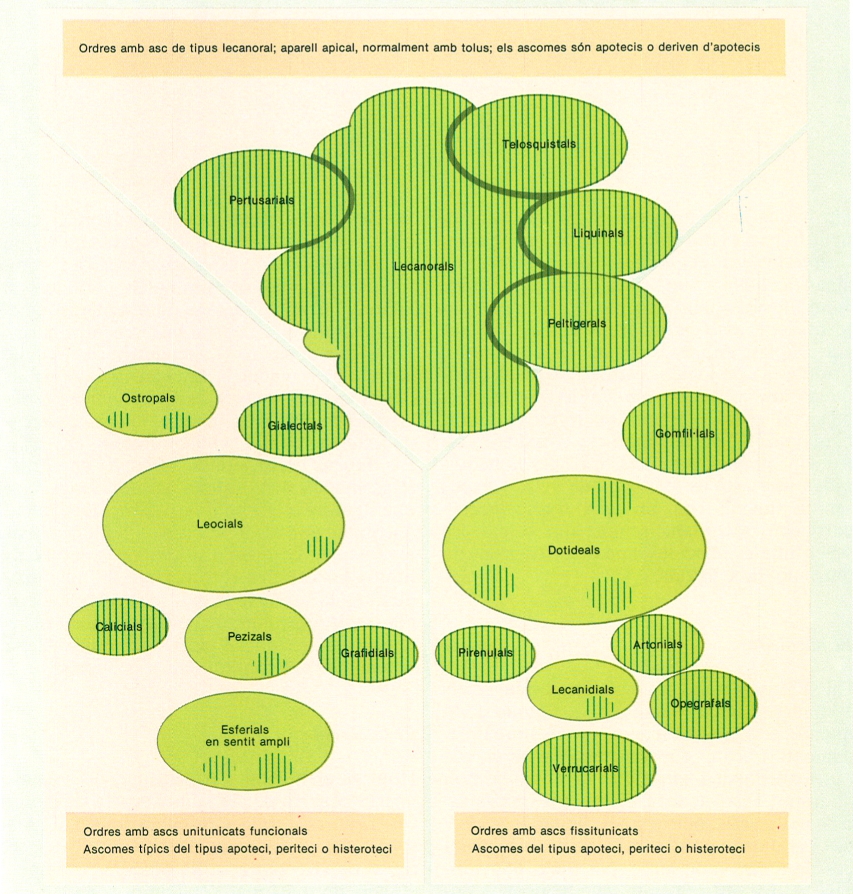
Els micobionts o espècies de fongs capaces de formar líquens són gairebé tots ascomicets. La figura mostra la distribució dels fongs liquenificats entre els ascomicets (no s’hi han inclòs els ordres que no contenen cap espècie de fong liquenificat); la superfície de les taques de color és indicadora del nombre de gèneres i espècies liquenificats de cada grup.
Biopunt, modificat a partir de J. Hafellner, en M. Galun.
Un 98% dels fongs liquenificats pertanyen als ascomicets (un 46% d’aquests). El grau de liquenificació que observem en els aproximadament 37 ordres d’ascomicets és molt divers, i probablement s’ha produït en èpoques molt diferents. Així, trobem sis ordres (telosquistals, peltigerals, pertusarials, liquinals, gialectals i grafidals) només formats, probablement des de temps molt antic, per fongs liquenificats. Altres ordres comprenen de manera dominant fongs liquenificats (lecanorals, calicials, artonials, opegrafals, verrucarials), un tercer grup (ostropals, leocials, pezizals, gomfil·lals, patel·larials, pirenulals, dotideals i xilarials) té només alguns representants liquenificats, potser en època més recent, i la resta ni han encetat la liquenificació. També sembla més recent la liquenificació d’alguns basidiomicets (unes 50 espècies, entre agàrics i afil·loforats). Així mateix, alguns deuteromicets han adoptat la liquenificació (uns 200). Els líquens estan, doncs, ordenats en el mateix canemàs de la sistemàtica dels fongs, dels quals representen una part. Els caràcters del micobiont són els essencials a l’hora de situar el liquen en l’esquema sistemàtic general dels fongs. La dràstica reforma que es va introduint en la sistemàtica dels fongs els afecta de ple, i el progrés en el seu coneixement detallat ha estat essencial per a l’aclariment de la sistemàtica del conjunt dels ascomicets.
El fotobiont

Els fotobionts, és a dir, els organismes fotosintetitzadors que s’associen amb fongs per a formar els líquens, són relativament poc nombrosos, i es troben tots entre un nombre petit de grups d’algues i de cianofícies. Sovint perden la capacitat de reproduir-se per via sexual. A Principals gèneres d’algues capaços d’actuar de fotobiont en una simbiosi liquènica: A1 Trebouxia (la més abundant), A2 Myrmecia, A3 Coccomyxa i A4 Trentepohlia (clorofícies); i A5 Dichothrix, A6 Gloeocapsa, A7 Nostoc i A8 Stigonema (cianofícies). B Altres aspectes del fotobiont dels líquens. B1 Formació del protal·lus de Xanthoria parietína; noteu com les hifes sorgides de la germinació d’una espora (1) van a trobar les algues i les envolten. B2 Relació entre les hifes i el fotobiont (Gloeocapsa) en el liquen Pyrenopsis conferta; es formen haustoris que penetren poc en l’alga. B3 Haustori intracel·lular penetrant en el fotobiont (Nostoc) en el liquen Lempholemma chalazanum. Hom hi ha indicat: 1 espora, 2 hifes de retenció, 3 hifes de recerca, 4 haustori, 5 alga clorofícia, 6 cianofícia, 7 beina mucilaginosa.
Biopunt, a partir de R. G. Werner i altres fonts.
Hom ha identificat, formant part de líquens, uns 13 gèneres de cianòfits i uns 24 de cloròfits. Les transformacions que sofreixen els fan sovint difícils de reconèixer, a nivell específic, si no se’n fa un cultiu pur. Per exemple, els gèneres que, quan són lliures, són filamentosos, poden adoptar la forma de cèl·lules isolades o de curtes cadenes quan viuen liquenificades. Només algunes espècies són corrents: Nostoc i Trentepohlia, que hom pot trobar també sovint lliures; Trebouxia i Myrmecia, molt rares lliures. És normal que una mateixa espècie pugui formar part de líquens diferents. També un mateix fong es pot associar amb fotobionts diferents, com a mínim en les seves primeres etapes, però al final només en predomina un. La liquenificació impedeix en general la reproducció sexual dels fotobionts, que es reprodueixen gairebé sempre asexualment, mitjançant aplanòspores, sovint sota el control del fong. Cultivats per separat, tant els micobionts com els fotobionts demostren que el creixement lent és l’expressió d’un caràcter lligat al genotip. S’han fet moltes vegades intents de resíntesi de líquens a partir de cultius de micosimbiont i de fotosimbiont; les primeres fases del desenvolupament del nou tal·lus (fins a una certa estratificació) no són gaire difícils d’aconseguir, però només en el cas del Cladonia cristatella hom ha arribat a obtenir podecis amb himeni, i picnidis.
L’organització i l’estratègia biològica dels líquens
El tal·lus
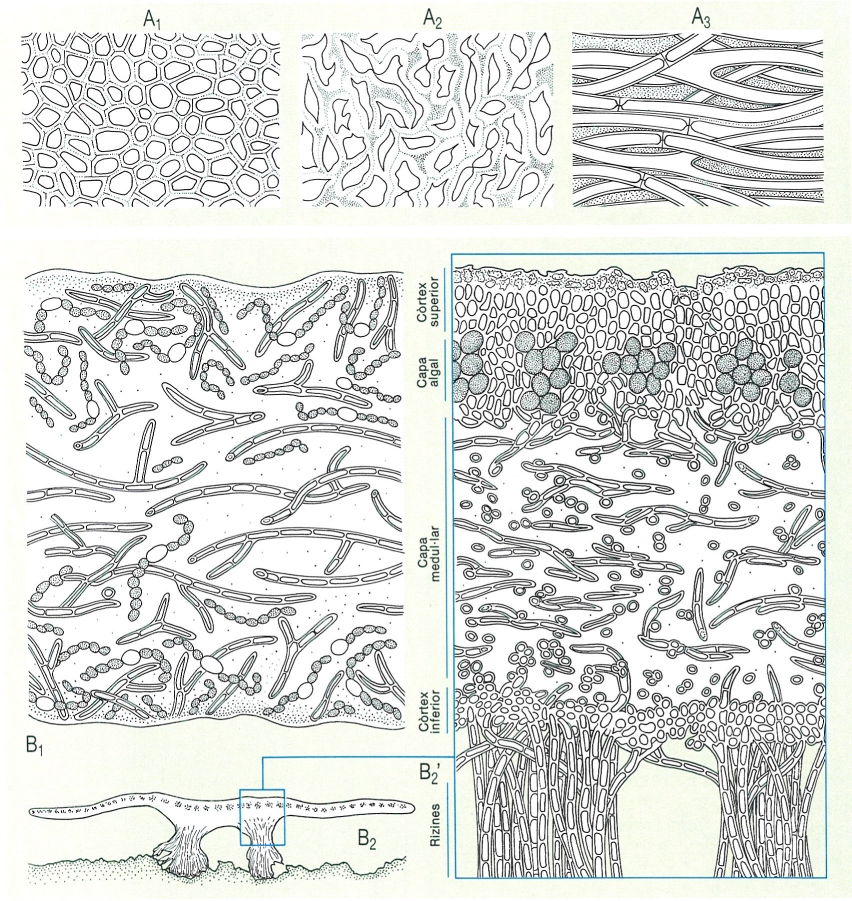
Estructura anatòmica dels tal·lus en els líquens. A dalt, principals tipus de falsos teixits (plectènquimes) en els líquens: A1 paraplectènquima, A2 prosoplectènquima, A3 calaroplectènquirna (aquest, amb hifes laxes o aracnoides, que deixen molts espais intercel·lulars). A baix, tal·lus homòmers i heteròmers. B1 Secció d’un tal·lus homòmer, de Collema tenax; les cadenetes de Nostoc estan barrejades més o menys uniformement amb les hifes (el puntejat indica la substància intercel·lular mucilaginosa). B2 Aspecte d’un tal·lus de Physconia grisea seccionat, i detall, augmentat (B2’), que mostra un tal·lus heteròmer.
Biopunt, a partir de fonts diverses.
El micobiont s’associa amb el fotobiont per formar l’aparell vegetatiu o tal·lus. Ho pot fer de maneres diferents, sempre orientades a facilitar la fotosíntesi i a assegurar la protecció de l’alga, la fixació al substrat i també la formació d’ascomes i de propàguls. Hi ha casos, com ara el gènere Collema, en què les cèl·lules de fotobiont es troben gairebé uniformement barrejades amb les hifes del fong. Són els líquens homòmers. Però és molt més freqüent que els components es disposin en capes: una capa superior, densa i protectora, només formada per fongs, el còrtex superior, sota el qual hi ha la capa que conté el fotobiont, la capa algal, la qual cobreix una capa laxa, també només fúngica, la capa medul·lar; sovint hi ha encara una capa inferior, fúngica, el còrtex inferior, amb paper de protecció i de fixació. Els líquens així diferenciats en estrats s’anomenen heteròmers.
Entre els tal·lus d’estructura més simple, podem esmentar els leprarioides (com ara les Lepraria), formats per una capa pulverulenta de petites unitats, sense còrtex, que sempre contenen algues i hifes del fong. També són secundàriament simples els tal·lus immersos al substrat, sovint només visibles a l’exterior per l’aparició d’ascomes: parlem de tal·lus endofleus quan el substrat és escorça (com en el cas d’Arthonia pinastri) i d’endolítics, quan és roca, freqüentment calcària (com en Verrucaria parmigera, Clauzadea immersa); sovint aquests tipus de tal·lus es consideren entre els crustacis (tipus en crosta interna).
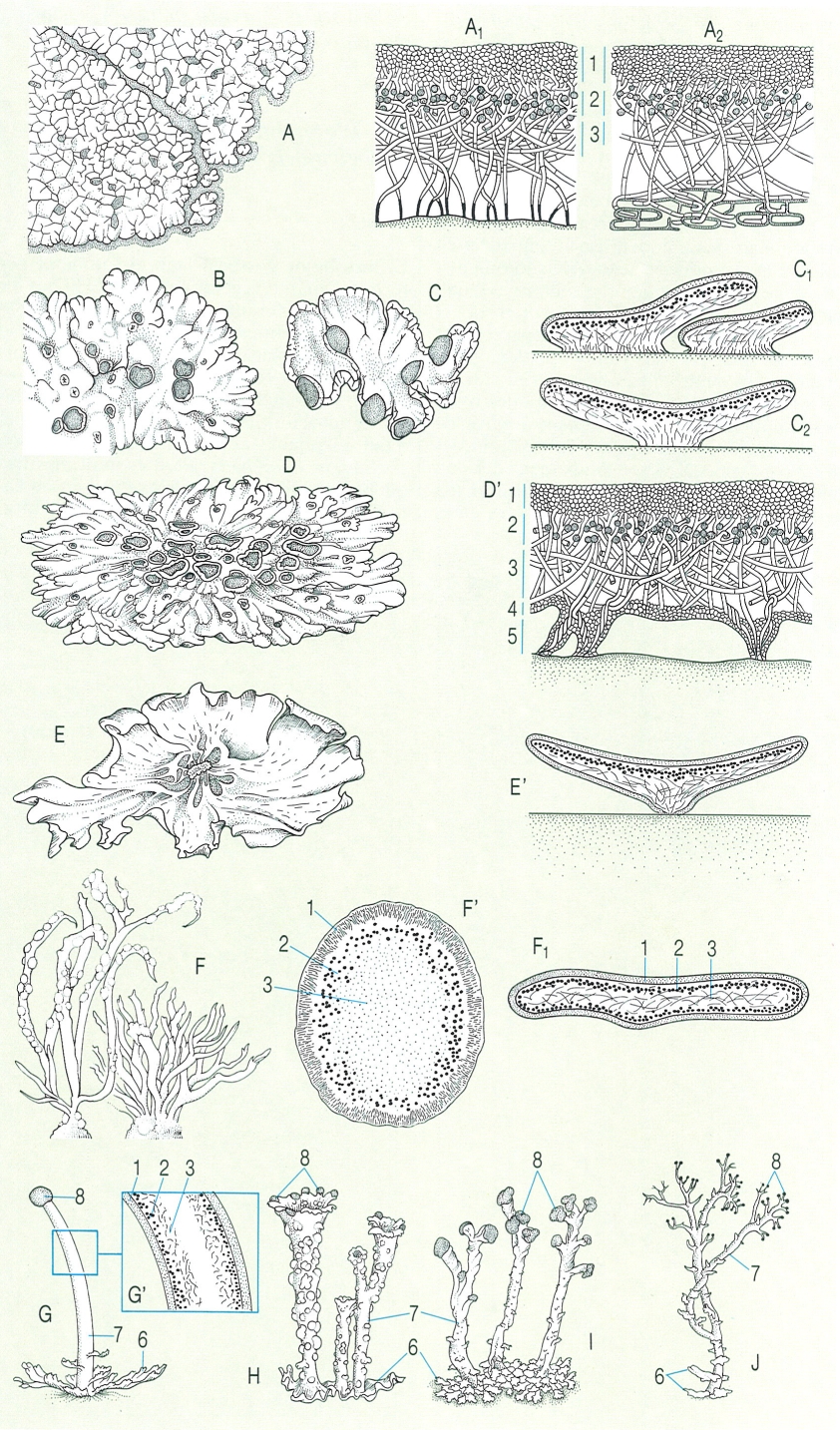
Tipus de tal·lus en els líquens. A Tal·lus crustaci fissurat i areolat, amb línia hipotal·lina al voltant i apotecis situats entre les arèoles; A1 secció d’un tal·lus crustaci epilític; A2 secció d’un tal·lus crustaci epifleu. B Tal·lus crustaci efigurat o placodioide, amb lòbuls (Solenopsora candicans). C Tal·lus esquamulós (Psora decipiens);C1 secció d’esquàmules d’una altra espècie (Squamarina cartilaginea), en la qual cal notar l’absència de còrtex inferior; C2 secció d’un tal·lus esquamulós peltat (Peltula) per a veure la fixació central i l’absència de còrtex inferior. D Tal·lus foliaci (Xanthoparmelia taractica) (D’ en secció, amb el còrtex inferior i les rizines que el fixen al substrat). E Tal·lus umbilicat (Umbilicaria virginis), vist per la cara inferior, per a mostrar el llombrígol central de fixació (E’ en secció, on es veu el còrtex inferior). F Tallus fruticulós (Roccella phycopsis,) (F’ en secció, per a veure-hi l’estructura estratificada i radiada); F1 secció d’un tal·lus fruticulós laciniat de Ramalina. G-J Tal·lus compostos: G estructura ideal d’un tal·lus compost, amb tal·lus primari (6) i podeci (7), que sosté un apoteci (8) (G’ anatomia del podeci); H podecis en trompeta (escifats) de Cladonia pyxidata; I podecis poc ramificats, amb discs himenials (fragment d’apoteci de Cladonia cristatella); J podecis ramificats (Cladonia furcata). Hom hi ha indicat: 1 còrtex superior, 2 capa algal, 3 medul·la, 4 còrtex inferior, 5 rizina, 6 tal·lus primari, 7 podeci, 8 apoteci (o disc himenial). El substrat, representat en puntejat en el dibuix, és de roca en tots els exemples excepte en A2, en què representa una escorça.
Biopunt, a partir de G. Clauzade, C. Roux, i J. Poelt.
Els tal·lus crustacis típics formen, com indica el seu nom, crostes sobre els substrats colonitzats, sovint amb còrtex superior diferenciat, que protegeix una capa algal i una capa medul·lar fermament fixada al substrat. És comprensible que quan volem recol·lectar aquests líquens ens haguem d’emportar un tros del substrat sobre el qual viuen. Els tal·lus crustacis formats sobre les escorces són els epifleus, i els que neixen sobre pedra són els epilítics. Quan són prims, els tal·lus crustacis poden ésser llisos, però en augmentar una mica el gruix formen fissures, que aviat delimiten un reticle (tal·lus fissurato-areolats), com passa en Rhizocarpon geographicum; en aquesta darrera espècie (i en moltes d’altres) el marge del tal·lus és delimitat per una línia fosca, integrada només per hifes del fong: és el protal·lus o línia hipotal·lina. Però el límit pot ésser no lineal sinó format per lòbuls radials, que es van ramificant a mesura que creixen; es tracta dels líquens placodioides o efigurats. En bastants casos, i sobretot en espècies terrícoles, les arèoles se separen en part del substrat, i formen esquàmules imbricades, sense còrtex inferior. Parlem llavors de tal·lus esquamulosos. Un cas particular són els tal·lus peltats, formats per esquàmules unides al substrat per un punt o curt peduncle central. Els dos tipus principals que veurem a continuació, els tal·lus foliacis i els fruticulosos, corresponen als líquens més grossos i visibles, que tradicionalment han estat designats amb el nom de macrolíquens. Els líquens foliacis són sempre lobulats i no estan units al substrat per tota la cara inferior sinó per punts o, més sovint, mitjançant òrgans de fixació anomenats rizines. Això fa possible, en general, de desprendre’ls del substrat fent passar un ganivet entre aquest i el còrtex inferior. Un cas particular és el dels tal·lus umbilicats, més o menys discoïdals, units al substrat només per un llombrígol central.

Òrgans de fixació i altres característiques del tal·lus liquènic: venes, cilis, pseudocifel·les, cifel·les, cefalodis i pruïna. A Rizinohifes o xarxa hifal (Psora decipiens), B cordó rizinoide (Buellia pulchella), C rizines cilíndriques simples (Melanelia septentrionalis), D rizines bifurcades, E rizines ramificades en angle recte (Peltigera), F venes en la cara inferior (Peltigera praetextata; F’ detall d’una rizina, que surt d’una vena), G cilis vitris (Phaeophyscia hirsuta), H cilis marginals (Anaptychia ciliaris), I papil·les (les prominències curtes) i fibril·les (les llargues) d’Usnea lapponica, J aspecte del córtex inferior de Sticta, amb cifel·les (J’ secció d’una cifel·la, on es veuen les cèl·lules arrodonides i laxament unides que la delimiten); K arèola de Caloplaca gloriae amb nombroses pseudocifel·les (K’ en secció, feta al nivell d’una pseudocifel·la; K" detall per a veure com les hifes medul·lars, laxament unides, afloren per una discontinuïtat del còrtex); L secció d’un cefalodi extern (Peltigera variolosa), M cefalodi intern (Lobaria laetevirens), N pruïna (Physconia detersa) i N’ detall del material blanquinós, pulverulent (cristalls d’oxalat càlcic), vist al microscopi electrònic d’escandallatge. Hom hi ha indicat: 1 còrtex, 2 capa algal, 3 medul·la, 4 còrtex del cefalodi, 5 cianofícies del cefalodi.
Biopunt, a partir de fonts diverses.
Els tal·lus fruticulosos són els fixats al substrat per un sol punt, el disc basal, i més o menys ramificats. N’hi ha de rígids, erectes, i de penjants, pèndols. Les branques poden ésser aplanades i d’estructura estratificada, i s’anomenen lacínies, o bé cilíndriques, i llavors tenen una estructura en capes concèntriques (estructura radiada). Un cas particular, ben exemplificat pel gènere Cladonia, és el dels tal·lus compostos, formats per esquàmules d’un tal·lus primari que emeten apoteciòfors cilíndrics (simples o ramificats) o en forma de trompeta, anomenats podecis, que constitueixen el tal·lus secundari, el qual pot ésser totalment dominant. També hem d’esmentar els tal·lus filamentosos o capil·lars (Ephebe, Coenogonium, Spilonema), formats per fotobionts filamentosos revestits per una beina fúngica. Cada tipus de tal·lus s’ha originat diverses vegades, i moltes famílies reuneixen líquens de tal·lus molt diversos. L’aspecte extern té, doncs, cada cop menys importància per a la delimitació de les famílies, i fins i tot dels gèneres. A això cal afegir la gran plasticitat de molts tal·lus davant els diferents ambients, la qual cosa fa que moltes espècies siguin considerablement polimòrfiques.
És bastant important l’anatomia del tal·lus. El còrtex, per exemple, pot ésser format per hifes de cèl·lules curtes i ben soldades entre elles, amb un aspecte cel·lular (paraplectènquima), o bé per hifes de cèl·lules més llargues, paral·leles, de paret gruixuda i soldada amb les parets veïnes fins a fer-ne difícil l’apreciació dels límits (prosoplectènquima). Hi ha també diferenciacions corticals formades per hifes perpendiculars a la superfície (com en Roccella). No és rara la presència de pigments protectors al còrtex i/o d’una capa protectora de polisacàrids (l’epicòrtex), sovint amb perforacions, o de cèl·lules mortes (capa epinecral). Típicament, la medul·la és formada per hifes gruixudes, laxament entreteixides, sovint incrustades de cristalls de substàncies liquèniques i amb molts espais entre elles. Hom ha provosat per a aquest «teixit» aracnoide el nom de calaroplectènquima.

Tal·lus compost de Cladonia fimbriata, format per un tal·lus primari (esquàmules aplicades al substrat, que aquí és fusta molt descomposta) i uns podecis, que aquí són escifats (en forma de trompeta) i pulverulents (sorediats). En el marge dels podecis hi ha picnidis i, ocasionalment (no en la fotografia), ascocarps (concretament, discs himenials).
Antoni Agelet.
Els líquens es fixen al substrat sia per tota la medul·la (líquens crustacis), o en el cas de líquens esquamulosos, amb xarxes d’hifes (rizinohifes) o feixos d’hifes (cordons rizinoides), i en el cas de molts líquens foliacis, mitjançant rizines, amb aspecte d’arreletes, simples o ramificades. Ja hem dit que molts líquens fruticulosos formen un disc de fixació (Usnea, Ramalina), però d’altres (Alectoria) es poden fixar al substrat per les seves mateixes branques. No hem de confondre les rizines, sempre producte del còrtex inferior, amb els cilis, emesos pel marge del tal·lus (Physcia adscendens), robustos (Anaptychia ciliaris), o rígids, fins i hialins (cilis vitris o de vidre, com en Physconia servitii). Moltes espècies de Sticta presenten en el còrtex inferior uns clotets blancs, les cifel·les, entapissats de cèl·lules especialitzades, laxes. Són molt més freqüents les pseudocifel·les (per exemple, en Cetraria islandica), que poden aparèixer tant en el còrtex inferior com en el superior. Són taquetes, en general de color mésclar, planes, arrodonides o allargades (Parmelia saxalitis) i representen discontinuïtats del còrtex, per les quals les hifes medul·lars emergeixen a la superfície. Tant les cifel·les com les pseudocifel·les s’interpreten com a diferenciacions destinades a facilitar l’intercanvi de gasos; per això no apareixen en els líquens que tenen porus a l’epicòrtex.
En alguns líquens (unes 520 espècies, pertanyents a uns 21 gèneres) formats amb algues verdes s’estableix una simbiosi amb 3 components, ja que s’hi afegeix una cianofícia. Aquest segon fotobiont pot formar una segona capa verda, sovint sota la de les clorofícies (com passa en Solorina crocea), o bé pot originar un òrgan extern pustuliforme o coral·loide, de color de plom, negrós, brunenc (Peltigera leucophlebia, Lobaria amplissima). Aquestes diferenciacions són anomenades cefalodis (interns i externs, respectivament). En tots aquests casos, i tenint en compte que la cianofícia dels cefalodis té sempre heterocists, amb capacitat per a fixar nitrogen, podem comprendre que molt probablement els cefalodis funcionen com a òrgans de fixació de nitrogen, en part comparables als nòduls radicals de les lleguminoses.
Els òrgans reproductors: ascomes
Els ascomicets liquenificats es reprodueixen formant fructificacions o ascomes de tipus i mida molt diferents. Un liquen que presenta ascomes s’anomena fèrtil. Quan no en té, diem que és estèril. Un liquen pot ésser estèril a causa de condicions de vida inadequades o quan s’ha especialitzat a reproduir-se asexualment.
Els apotecis i els peritecis
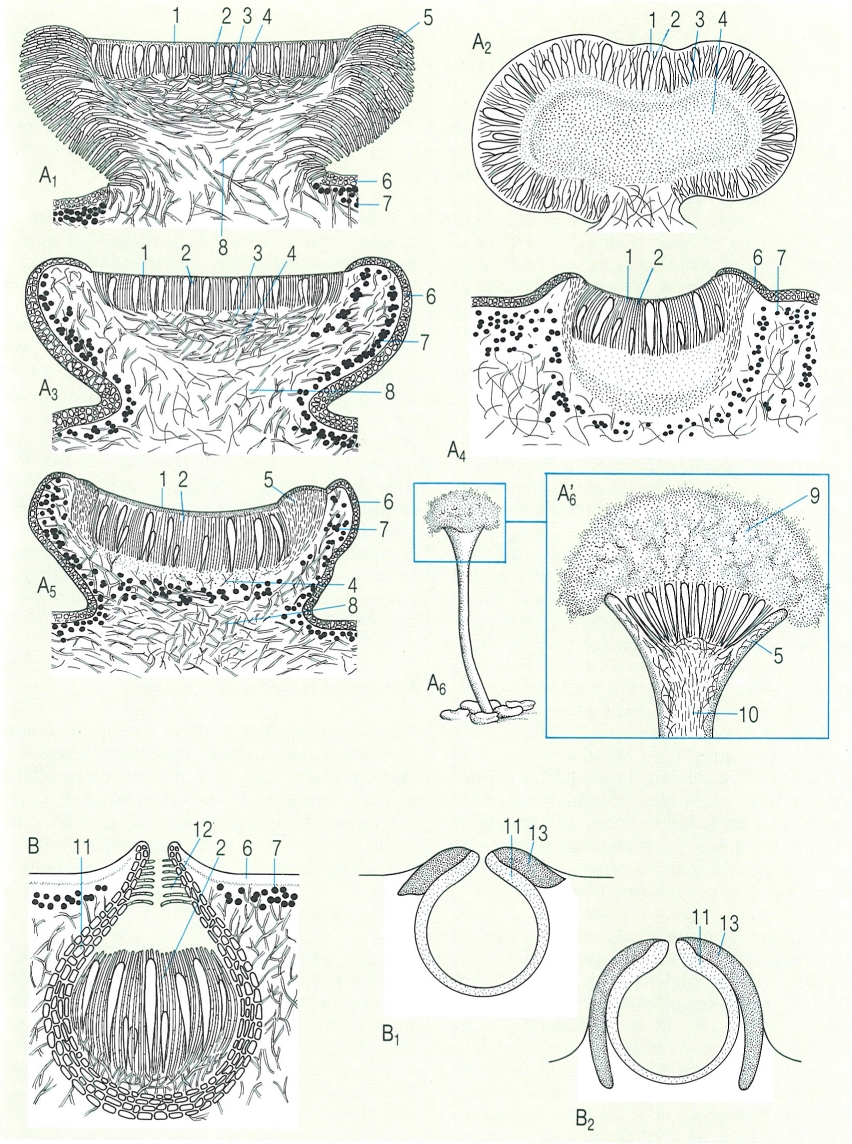
Tipus principals d’ascomes en els líquens: apotecis (A) i peritecis (B): A1 apoteci lecideí, amb marge propi (excípul fúngic); A2 apoteci biatorí, sense excípul; A3 apoteci lecanorí, amb marge tal·lí que conté algues; A4 apoteci criptolecanorí o aspicilioide, semiimmers en el tal·lus, però envoltat d’algues; A5 apoteci zeorí, amb marge propi i marge tal·lí, A6 mazedi (Calicium) estipitat (A6 secció, amb la massa pulverulenta on hi ha les espores). B Secció d’un periteci, envoltat per l’excípul; B1 B2 secció de dos peritecis amb involucre, que en el primer (B1) en cobreix un terç de l’alçada, i en el segon (B2) el cobreix fins a la base. Hom hi ha indicat: 1 epiteci, 2 teci o himeni, 3 subhimeni, 4 hipoteci, 5 marge propi, 6 còrtex, 7 capa algal, 8 medul·la, 9 mazedi, 10 estípit, 11 excípul o paret del periteci, 12 perífisi, 13 involucre.
Biopunt, a partir de fonts diverses.
Els dos grans tipus d’ascomes són els apotecis, els més abundants, en els quals la capa fèrtil on hi ha els ascs és en contacte amb l’exterior, i els peritecis, sovint més petits, en què els ascs estan tancats en una cavitat només oberta a l’exterior per un porus anomenat ostíol.
Segons l’ontogènia de cada ascoma (ascohimenial o ascolocular, vegeu els ascomicets) tenim diverses menes d’apotecis i de peritecis, que no es poden distingir a simple vista ni amb una lupa. Segons llur posició, els ascomes poden estar més o menys immersos en el tal·lus, inclusos (a nivell del tal·lus), sèssils, prominents i fins i tot estipitats. Els podecis de les Cladonia no són sinó estípits liquenitzats. Hi ha ascomes allargats, amb el disc en forma d’escletxa, de vegades ampla, sovint sinuosos: són les lirel·les, freqüents en les opegrafals i en el gènere Graphis. En els mazedis, els ascs i les paràfisis aviat es converteixen en una pols desbordant, barrejada amb espores.
Les hifes que delimiten els ascomes formen una paret sovint ben diferenciada, l’excípul, també anomenat marge propi o parateci en els apotecis, i, en general, té un color diferent del del tal·lus. Sovint, en els apotecis s’origina també, per fora, un marge amb algues i còrtex, el marge tal·lí, excípul tal·lí o amfiteci, que presenta el mateix color del tal·lus.
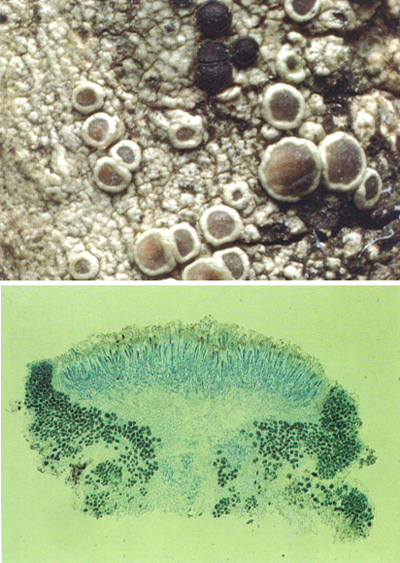
Els apotecis lecanorins i lecideïns són els dos tipus principals d’apotecis presents en els líquens. Negres i amb marge propi, també negre, els apotecis lecideïns de Buellia disciformis apareixen envoltats, en la fotografia de dalt, pels apotecis més grossos i amb marge tal·lí del mateix color del tal·lus i amb disc brunenc, del tipus lecanori, de Lecanora chlarotera forma meridionalis. Vegeu les diverses etapes del desenvolupament i, sovint, la deformació per pressió mútua. A baix, secció (tenyida) d’un apoteci lecanori, on veiem l’epiteci, cobrint l’himeni (hi veiem sobretot ascs immadurs), el qual recobreix l’hipoteci. Pels costats, hi ha el marge tal·lí, amb nombroses algues (molt tenyides de blau).
Jordi Vidal i Bukhard Büdel.
Els apotecis amb marge propi s’anomenen lecideïns; els que tenen marge tal·lí són els lecanorins; i els apotecis lecanorins immersos al tal·lus, de forma que estan envoltats d’algues però el marge tal·lí es veu poc o gens diferenciat, són els criptolecanorins (o aspicilioides). Quan es veuen bé el parateci i l’amfiteci, junts, es parla d’apotecis zeorins. En diversos líquens es veuen bé els dos marges; a vegades (Diploschistes), estan més o menys separats. Quan l’himeni d’apotecis inicialment lecideïns creix tan de pressa que es torna convex i elimina el marge propi, hom parla d’apotecis biatorins. La capa fèrtil s’anomena, com en els ascomicets no liquenificats, himeni o teci; just a sota hi ha un subhimeni, que reposa sobre un hipoteci, el qual pot ésser hialí, brunenc o carbonaci. Les hifes situades entre els ascs (hamateci) poden formar filaments ascendents, poc o gens ramificats (paràfisis), o descendents, més o menys anastomitzats i fixats després a la base de l’himeni (pseudoparàfisis). També es poden formar, per esponjament del teixit preexistent, fins a donar lloc a una sèrie de trabècules fines, ramificades i anastomitzades (parafisoides). Les paràfisis presenten sovint un àpex dilatat (paràfisis capitades). Per reunió, amb incrustació, dels àpexs, es forma sovint una capa acolorida, protectora de l’himeni, l’epiteci. Entre els ascs i les paràfisis és característic que hi hagi una gelatina himenial, que normalment es tenyeix de blau amb els reactius iodats. A vegades hi ha gotetes de lípids atrapades en aquesta gelatina (himeni inspers). Les paràfisis poden ésser coherents, sovint anastomitzades i fermament retingudes per la gelatina himenial, o lliures i no coherents. En els peritecis, cal fixar-se si l’excípul o paret del periteci l’envolta completament (és enter) o només es veu a la part superior (és dimidiai). En alguns casos (per exemple en moltes Verrucaria), s’hi afegeix un escut negrós i unit a l’àpex, l’involucrel, que pot cobrir la part apical, un terç, la meitat o la totalitat del periteci (fins a arribar a tancar-se a la base); per veure-ho bé, cal examinar una bona secció de periteci al microscopi. En alguns casos (per exemple, en els gèneres Endocarpon, Staurothele) l’himeni conté algues himenials, diferents de les que hi ha en el tal·lus.
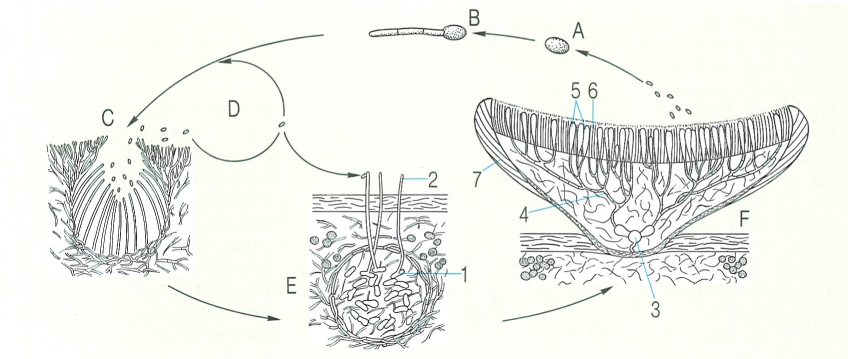
Cicle vital dels líquens. A Espora haploide. B Germinació d’una espora (la incorporació d’algues i la formació del tal·lus liquènic no figuren en el dibuix). C Picnidi, que produeix pienidiòspores o espermacis. D Germinació de les pienidiòspores (dubtosa). E Ascogonis, amb tricògines emergents, situats en el primordi de l’ascocarp (el centre), i fecundació (no sempre realitzada) per pienidiòspores. F Ascocarp de tipus apoteci; a partir dels ascogonis fecundats del centre (3) s’han format hifes ascògenes, dicariòtiques, que aniran originant ascs durant molt de temps. Hom ha indicat en el dibuix: 1 ascogoni, 2 tricògina, 3 centre, 4 hifes ascògenes, 5 ascs, 6 paràfisis que formen l’himeni (la meiosi té lloc en madurar els ascs), 7 excípul.
Biopunt, a partir d’originals de M. A. Letrouit, publicats per M. Galun.
El desenvolupament dels ascomes és força diversificat i complex. En general, tot comença amb la formació d’un petit cabdell d’hifes ascogonials (o ascogoni), d’on hom pot veure emergir tricògines. En alguns líquens (Collema, Cladonia, etc.), sembla evident que cada tricògina pot rebre un espermaci (en realitat, un conidi), el nucli del qual fecunda l’ascogoni i fa possible que en derivin hifes ascògenes, dicariòtiques, que es ramifiquen dins el primordi de l’ascoma, i després pel que serà l’himeni en l’esboç de l’ascoma, una fase més complexa però encara soterrada al tal·lus. En la majoria dels líquens, l’apoteci jove emergeix aviat (gimnocàrpic), però en les pertusarials, les peltigerals i les grafidals no ho fa fins que és plenament madur (hemiangiocàrpic). Si només s’obre a l’exterior per un porus, que s’obre pas per un tap superior o con epicentral, es produeix un periteci de desenvolupament angiocàrpic. No és pas segur que en tots els líquens l’ascogoni sigui realment fecundat. L’aparició d’hifes ascògenes per partenocàrpia és probable, i la variabilitat genètica es deu mantenir per parasexualitat. Com en els ascomicets no liquenificats, el desenvolupament pot ésser ascohimenial (amb els ascs formant part d’un himeni des d’un bon començament), o ascolocular (amb els ascs que s’obren pas i s’encabeixen en un lòcul excavat en un estroma preexistent). Malgrat la freqüent correlació entre l’existència de desenvolupament ascohimenial i la d’ascs unitunicats, hi ha líquens amb ascomes ascohimenials que tenen ascs bitunicats.
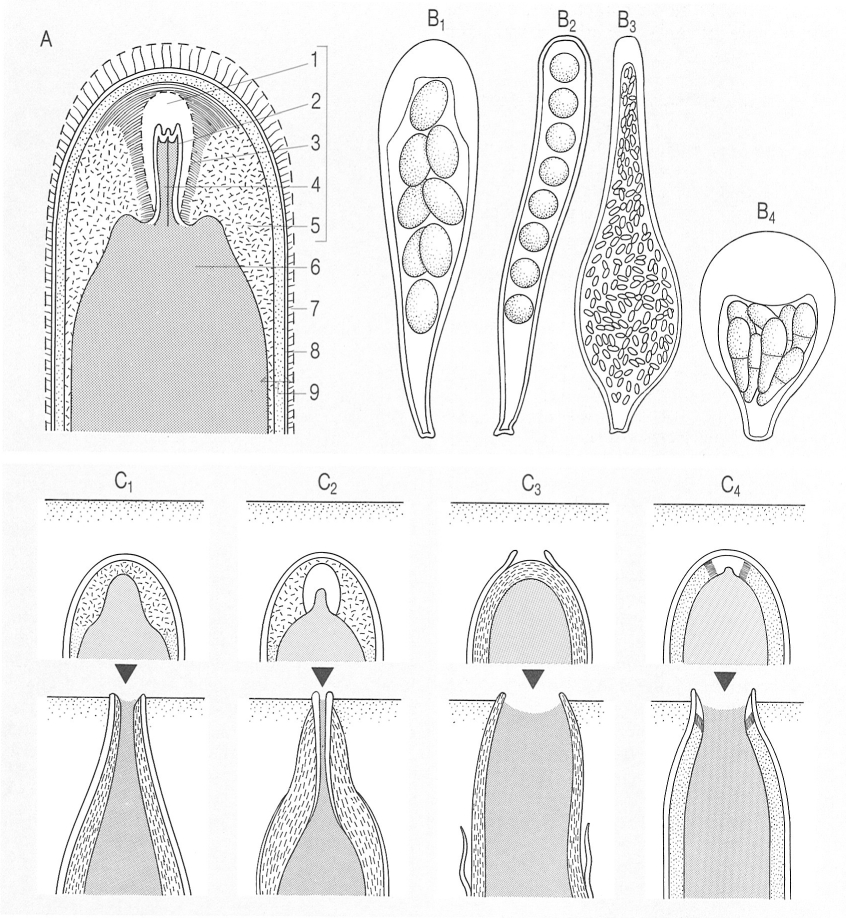
Aspectes de la morfologia dels ascs dels líquens. A Esquema generalitzat d’un aparell apical típic d’un asc de liquen; l’engruiximent apical (1 a 5) és el tolus. 6 Tipus d’ascs: B1 claviforme (Aspicilia chadefaudiana), B2 cilíndric (Schaereria cinereorufa), B3 ampul·liforme (Thelocarpon laureri) B4 piriforme (Arthonia vinosa). C Tipus de dehiscència: C1 tipus Teloschistes, C2 tipus rostrat (amb una certa extensibilitat de la túnica interna), C3 tipus fissitunicat (la túnica interna és molt extensible), C4 tipus amb eversió. En el dibuix, la línia negra, fina, horitzontal, representa la superfície de l’himeni. Hom hi ha indicat: 1 cos axial o manubri, 2 nansa apical, 3 anell, 4 cambra ocular, 5 engruiximent subapical, 6 cambra subapical, 7 capa gelificada externa, 8 exoasc o túnica externa, 9 endoasc o túnica interna (amb dues capes).
Biopunt, a partir de M. A. Letroit i altres fonts.
Els ascs dels ascomicets són normalment petits (unes 50 µm) i claviformes (en forma de porra), però també poden ésser grossos (fins a 300 µm en algunes Pertusaria), subglobosos (Arthonid), cilíndrics (Bactrospora), etc; la paret és gruixuda i, al microscopi electrònic, es resol en quatre capes, dues d’externes (a, la capa externa de la qual sol ésser gelificada, i b), que formen l’exoasc, i dues d’internes (la c, gruixuda, i la d), que formen l’endoasc.
Sobretot les capes internes es presenten més o menys dilatades a la part apical, i formen un aparell apical, que incorpora sovint substàncies amiloides, les quals es tenyeixen de blau amb reactius iodats (reacció I + blau).
Per a estudiar l’aparell apical hom estova una mica d’himeni amb una solució de KOH, el renta diverses vegades amb aigua destil·lada i el tracta després amb lugol (solució iodoiodurada) diluït. Tant al microscopi òptic, després d’aquest tractament, com al microscopi electrònic, es veu que l’aparell apical és molt divers. Hom li concedeix actualment una gran importància per a establir una taxonomia més natural dels líquens. Així, podem veure que (sobretot en el gran ordre de les lecanorals), l’endoasc forma un engruiximent apical, anomenat tolus (del grec thólos, cúpula) o dom (del grec, doma, mateix significat), que pot ésser més o menys amiloide (I+ blau), sia uniformement (tipus Catillaria) o amb una zona axial o cos axial (també dita manubri) diferenciada, més clara (tipus Lecanord). Cal fixar-se, també, en la possible presència d’una penetració de la cambra esporal en el tolus, que forma la cambra ocular (ben visible i ampla en el gènere Rhizoplaca). En el tipus Psora hi ha un característic tub amiloide, prominent cap a la cambra esporal, formant un penjant.
A diferència dels ascomicets no liquenificats, l’activitat dels ascomes dels líquens és de llarga durada, i constantment poden madurar nous ascs i noves ascòspores; hi ha, de tota manera, moments privilegiats per a l’expulsió de les espores (nit, tardor, després de les pluges, etc.).
Un dels tipus de dehiscència més freqüent és el rostrat (per exemple, en les lecanoràcies, les parmeliàcies, les fisciàcies, i les cladoniàcies), en el qual les capes externes de l’asc es trenquen i la interna s’allarga, per formar un rostre, fins a la superfície de l’himeni. En el tipus Teloschistes (propi de les telosquistals i potser també de Pertusaria), totes les capes de la paret s’estiren fins a la superfície hime nial i es trenquen a l’àpex. La dehiscència fissitunicada és freqüent en els líquens bitunicats: primer es trenquen les parets externes de l’asc, i la més interna, elàstica, es desenganxa més o menys i s’estira, a vegades més enllà de la superfície himenial; la presenten molts líquens amb peritecis (pirenulals, verrucarials), juntament amb líquens amb apotecis, com les artonials i les opegrafals, Rhizocarpon i Peltigera (aquests dos, amb un allargament apical limitat).
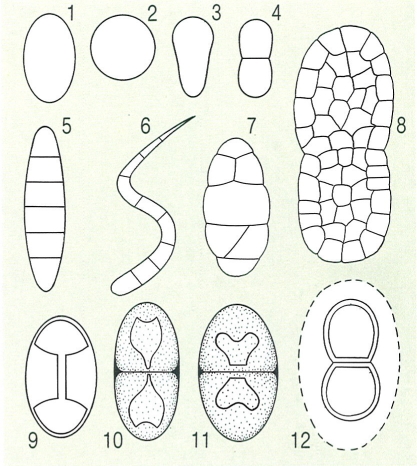
Variabilitat morfològica en les espores dels líquens. Espores simples: 1 el·lipsoïdal, 2 globosa, 3 claviforme heteropolar. Espores amb septes transversals (transseptes): 4 bicel·lular constricta, 5 pluriseptada, 6 vermiforme. Espores amb septes transversals i longitudinals: 7 submural, 8 mural. Espores especials: 9 polardiblàstica o polarilocular, 10 bruna, de parets irregularment engruixides (tipus Physcia) 11 polardiblàstica o polarilocular, però amb la llum cel·lular en forma de cor (tipus Mischoblastia) 12 bicel·lular constricta i amb halo (halonada).
Biopunt, a partir de fonts diverses.
A mesura que les espores van madurant, van diferenciant en llur paret diverses capes: a la paret pròpia, no sempre es diferencia un endospori; posteriorment, es diposita, per fora, un perispori, força visible, que pot ésser reduït (Lecanora) o gelificat i inflat, formant un halo (espores halonades, per exemple de Porpidia); també es pot originar una ornamentació (visible al microscopi d’escombratge), principalment derivada de l’heterogeneïtat de la capa mitjana del perispori. Les espores poden ésser incolores (hialines) o pigmentades, i en aquest cas el pigment, verdós o brunenc (probablement es tracta de melanines), apareix més o menys tard, uniformement o no (bandes, en algunes Rinodina); hom qualifica de fuliginoses les espores brunenques (Buellia, Physcia, Rhizocarpon, Diploschistes). Les espores poden ésser de formes molt diverses, globoses, el·lipsoïdals (molt freqüents), allargades, heteropolars (més amples d’un costat que de l’altre) i filamentoses. Moltes són unicel·lulars, però hi pot haver septes, que es van formant en madurar; els més freqüents són els transversals (transseptes). Si, a més de transversals, n’hi ha algun de longitudinal (longiseptes), parlem d’espores submurals, i de murals, si n’hi ha molts. Quan hi ha un septe transversal gruixut i perforat per un canal es parla d’espores polariloculars o polardiblàstiques (Caloplaca, Xanthoria).
Els picnidis
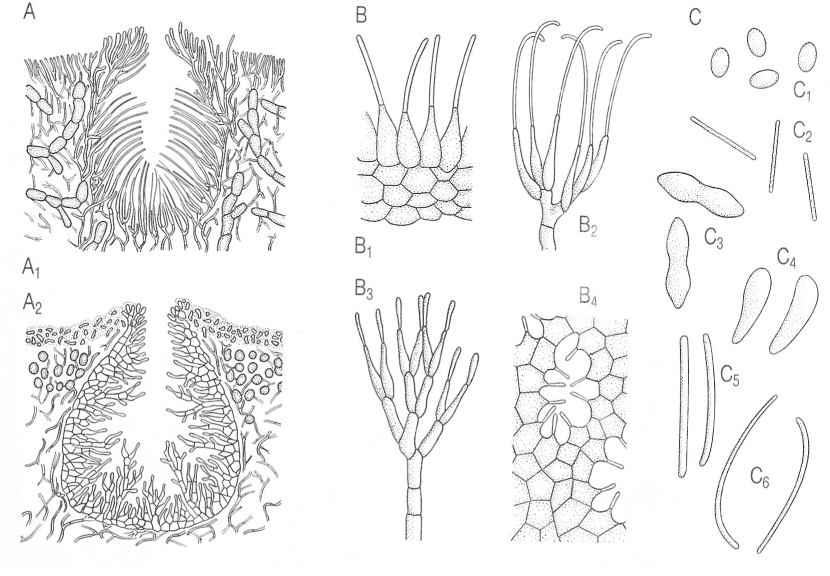
Tipus principals de picnidis (a), de picnidiòfors o conidiòfors (b) i de picnidiòspores o conidis (c) en els líquens. A Picnidis: A1 del tipus Roccella, amb conidiòfors simples i picnidiòspores filiformes i terminals; A2 del tipus Umbilicaria, amb conidiòfors ramificats (les diverses cèl·lules poden emetre picnidiòspores). B Picnidiòfors:
Biopunt, a partir de fonts diverses.
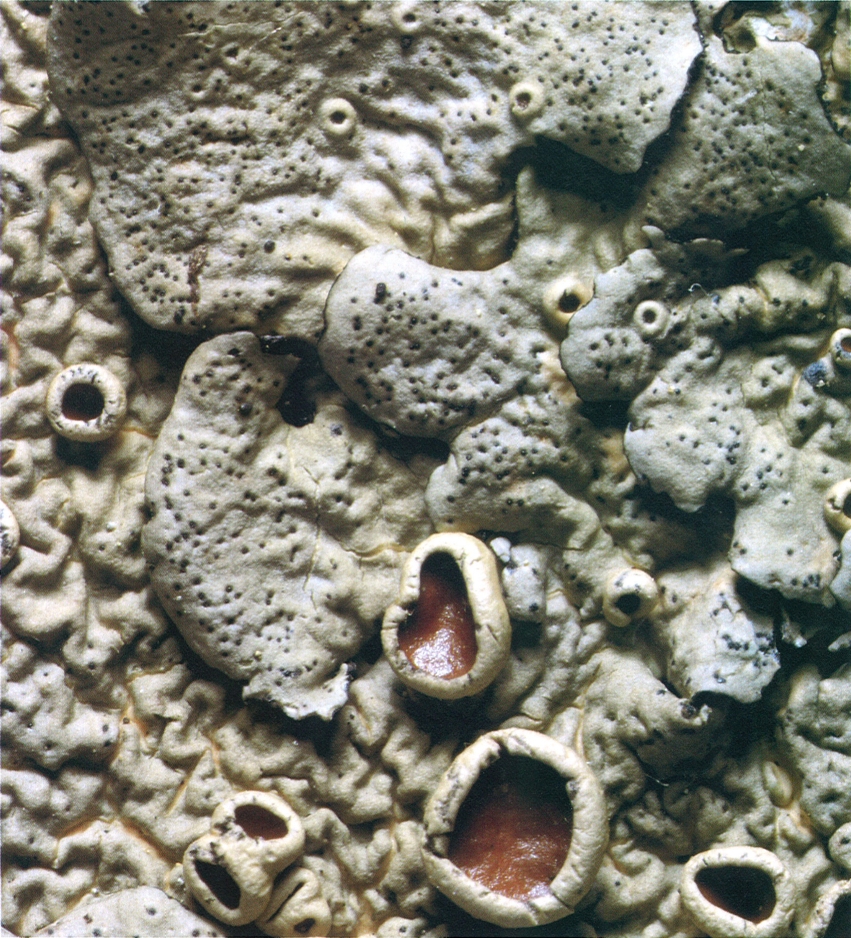
Picnidis, només visibles pels seus ostíols enfosquits (ja que la resta del pienidi és immersa al tal·lus), en un fragment de Parmelina quercina; són situats vora els lòbuls i són ben fàcils de diferenciar dels apotecis joves. En la fotografia destaquen especialment els apotecis madurs, amb marge tal·lí (lecanorins) i himeni bru, lluent. Els picnidis produeixen picnidiòspores (o conidis), que fan un paper sovint obscur en el cicle vital dels líquens.
Jordi Vidal
Sovint, és possible de veure amb una lupa, i sobretot a la part jove del tal·lus (examinem, per exemple, un lòbul de Parmelia taractica), uns puntets negres dispersos. Són les obertures (ostíols) dels conidiomes (en aquest cas, anomenats també picnidis) immersos al tal·lus. Els picnidis són els òrgans més freqüents de producció de conidis, tradicionalment anomenats picnidiòspores. Gairebé un 60% dels líquens emeten conidis, que en alguns casos funcionen com a espermacis i poden fecundar les tricògines, en uns altres casos poden actuar com a espores asexuals (i hi ha espècies que no disposen de cap altre medi de propagació) i, potser sovint, són simples estructures residuals, sense cap funció coneguda, però útils per a la distinció de molts gèneres i espècies. Hi ha líquens, més rars, que en lloc de picnidis poden formar conidiomes en forma d’acèrvul (cupuliforme), d’esporodoqui (pulvinulat), de sinnema (anomenats hifòfors, amb un peu hifal que sosté un feix de conidis), i de campilidi (en forma de casc). En el gènere Cladonia, els picnidis són ben visibles al marge de les trompetes o a l’àpex de les ramificacions dels podecis. Els conidis en surten sovint acompanyats d’una substància gelatinosa, hialina o acolorida. El tipus de picnidis i la forma i les dimensions (sempre petites, com a mínim en amplada) dels conidis tenen importància taxonòmica. Així, per exemple, els que contenen una xarxa esponjosa de conidiòfors curts (tipus Xanthoria) són característics de l’ordre de les telosquistals.
Els propàguls vegetatius: soredis i isidis
Fruit de l’evolució dels fongs en una etapa posterior a llur liquenificació és la capacitat, lligada genèticament a determinades espècies, de formar soredis o isidis, que funcionen essencialment com a propàguls vegetatius i no apareixen en els micobionts (fongs) ni en els fotobionts (algues) quan els cultivem per separat.
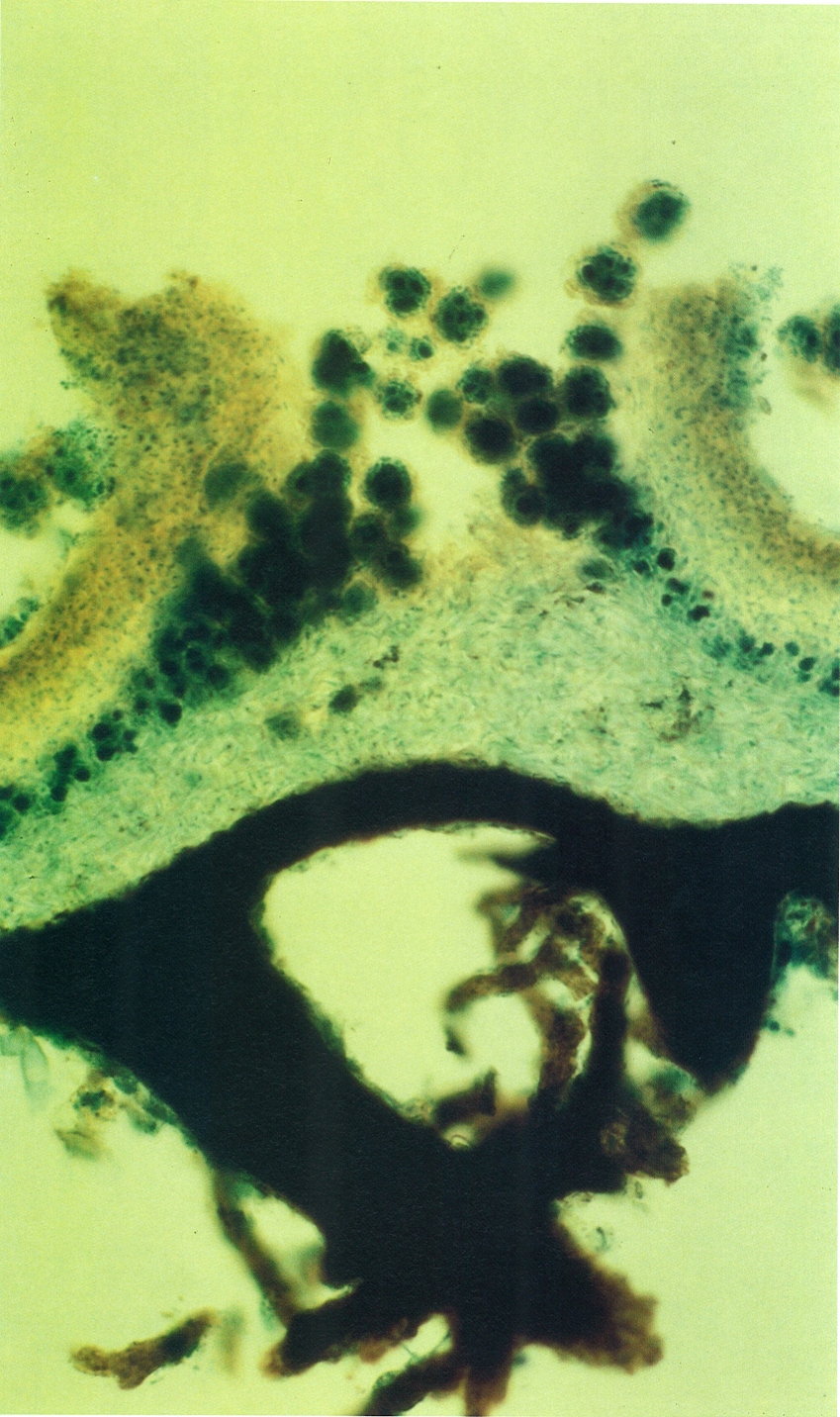
Els soredis i els isidis són dues menes de propàguls gràcies als quals molts líquens realitzen una multiplicació vegetativa. La fotografia mostra una secció tenyida d’un sorali de Parmelia sulcata (boletes amb les algues molt tenyides de blau), en el qual els soredis, després d’acumular-se, han trencat el còrtex i han sortit a l’exterior. Noteu que hi ha dues rizines sortint del còrtex inferior, ennegrit.
Burkhard Büdel.
La multiplicació vegetativa es pot fer en els líquens per fragmentació del tal·lus o bé mitjançant la formació de soredis i isidis, unitats de dispersió que contenen cèl·lules algals i hifes del fong; en el cas dels soredis, formen un propàgui esfèric, no corticat, mentre que els isidis són expansions recobertes pel córtex, que es trenquen i se separen fàcilment del tal·lus, de manera que es converteixen també en propàguls. A Fragmentació del tal·lus: un tal·lus ramificat, com aquests podecis de Cladonia, és fràgil en temps sec i, sota qualsevol acció mecànica (aquí, la caiguda d’una branca), es fragmenta en molts propàguis, que contenen fongs i algues. B Tipus de soredis i soralis: B1 soralis maculiformes, convexos, d’una Pertusaria (B’1 secció d’un sorali, mostrant els soredis, que es formen en la capa algal, trenquen el còrtex i surten a l’exterior; B1" detall de dos soredis d’espècies diferents, formats per una o més algues, revestides per hifes del fong); B2 soralis maculiformes, plans, de Pertusaria amara (cada boleta visible és un soredi) maculiformes, plans, de ; B3 soralis capitiformes, d’Hypogymnia tubulosa, situats a l’àpex de lòbuls cilíndrics; B4 soralis de fissura, de Parmelia sulcata (els soredis surten a través de fissures més o menys ramificades); B5 soralis marginals, de Cetraria pinastri. C Tipus d’isidis: C1 isidis cilíndrics o digitiformes de Xanthoparmelia conspersa (noteu el còrtex que els revesteix i la continuïtat amb la capa algal); C2 isidis esfèrics, de Xanthoparmelia tinctina; C3 isidis claviformes, de Melanelia exasperatula; C4 isidis coral·loides, de Lasallia pustulata, negres i ramificats; C5 fibril·les de Bryoria bicolor, que es trenquen fàcilment per la base i actuen com a propàguls. Hom hi ha indicat: 1 còrtex, 2 capa algal, 3 medul·la.
Biopunt, a partir de fonts diverses.
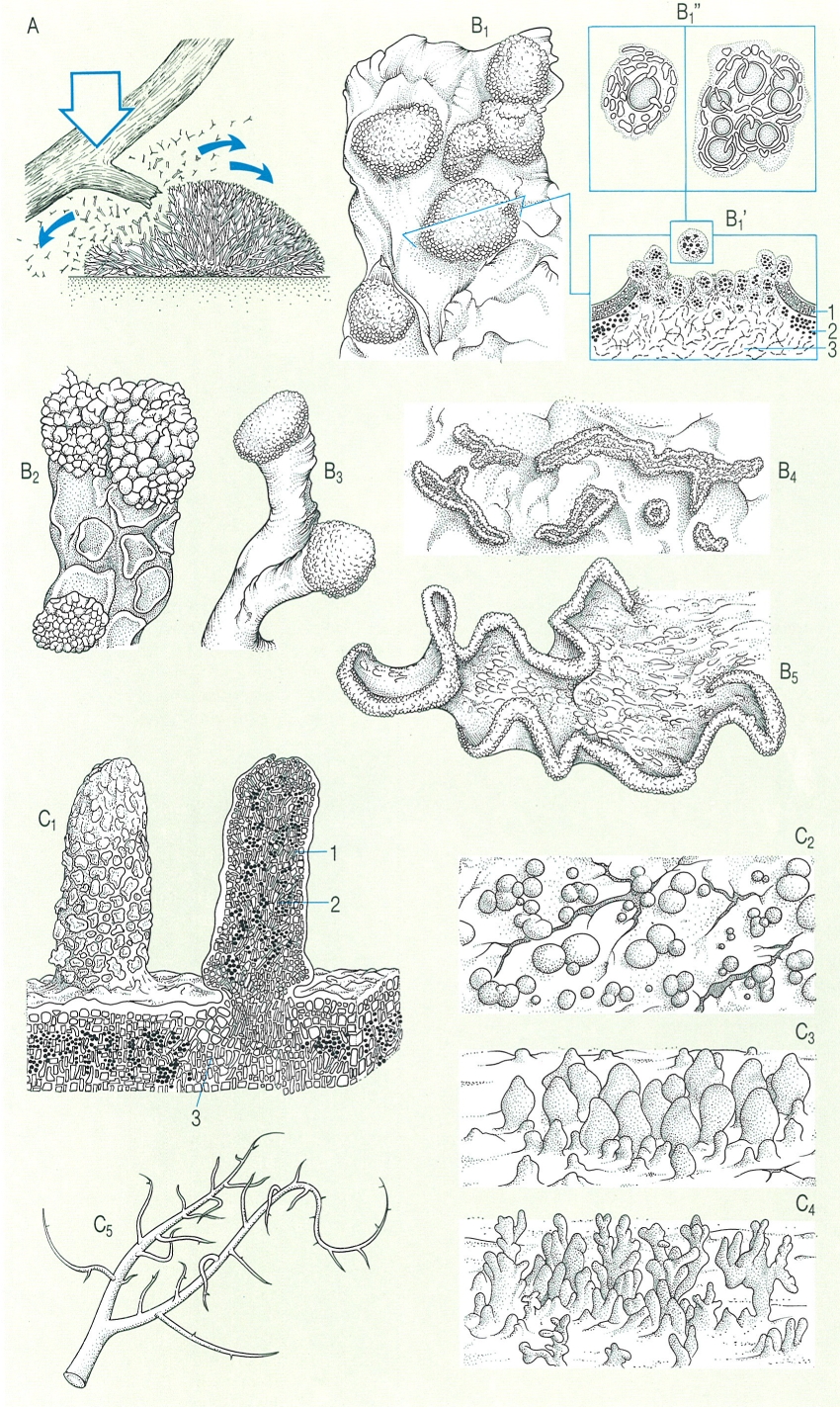
Els soredis són unitats de dispersió formades per una o diverses cèl·lules algals, laxament embolicades per hifes del micobiont. Llur formació s’inicia en la capa algal, però aviat emergeixen, trencant el còrtex, i formen una massa convexa, on es veuen soredis en forma de boletes pulverulentes. Aquesta massa de soredis, seca i hidròfuga (adaptada, doncs, a la dispersió pel vent i per l’impacte de gotes de pluja) s’anomena sorali. Els soralis són visibles a simple vista, o amb una lupa, i n’hi ha de maculiformes (de forma arrodonida, sovint convexa), capitiformes (marginals i arrodonits), labriformes (marginals i allargats), de fissura (emergents per una escletxa) i d’altres menes. A l’aeroplàncton, no és rar de trobarhi soredis; quan cauen en un lloc apropiat, creixen aviat i originen un nou tal·lus, ja que els dos simbionts, fong i alga, s’han desplaçat conjuntament.
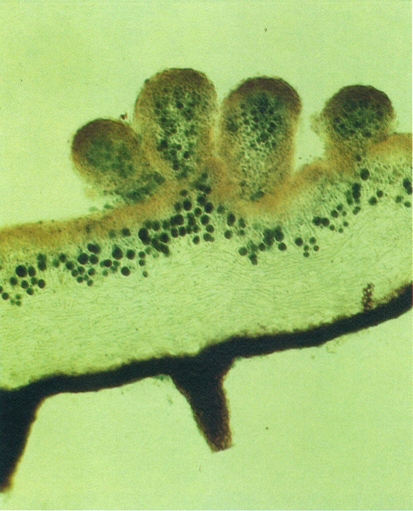
Secció d’un tal·lus de Parmelia saxatilis, amb quatre isidis cilíndrics. Noteu la continuïtat del còrtex superior que envolta els isidis. És ben visible la capa algal (amb algues tenyides de blau), la capa medul·lar i el còrtex inferior, ennegrit i amb dues rizines. Com que la base dels isidis és fràgil, aquests queden fàcilment separats del tal·lus i es dispersen.
Burkhard Büdel.
Els isidis són emergències del tal·lus de forma cilíndrica, ramificada o esquamulosa, revestides pel còrtex. Per això llur superfície és llisa. Fàcilment es trenquen per la base i es converteixen en eficaços propàguls, ja que també porten els dos components. Hi ha isidis esfèrics (Parmelia tinctina), cilíndrics o digitiformes (Pertusaria pseudocorallina), claviformes (Parmelia exasperatula), coral·loides (Lasallia pustulata), o esquamulosos, dits fil·lidis (Collema flaccidum). Els esquizidis són trossos esquamulosos de còrtex i capa algal que se separen de la medul·la i poden ésser dispersats per les gotes de pluja (Fulgensia desertorum). Les fibril·les (Usnea) o espínules (Bryoria) són curtes branques laterals, fàcilment dispersables.
Malgrat la complexitat de moltes d’aquestes diferenciacions, cal no oblidar que els líquens són fràgils en temps sec. Per això, els fruticulosos i alguns foliacis es fragmenten fàcilment, i els trossets així originats poden funcionar com a propàguls (Cladonia, Bryoria).
L’evolució ha actuat sovint en el sentit d’una pèrdua progressiva dels apotecis en el cas dels líquens dotats d’una bona capacitat de multiplicació asexual, seguida d’un isolament respecte de l’espècie progenitora (espècie primària), que ha conduït sovint a una diferenciació específica. Trobem sovint parelles d’espècies: Diploicia subcanescens-D. canescens, Fulgensia fulgida-F. subbracteata; la primera és l’espècie primària i la segona, la derivada o secundària, amb soredis o isidis. També podem trobar grups de tres espècies relacionades, una de primària, una amb soredis i una altra amb isidis. Una espècie derivada no pot tenir els dos tipus de propàguls a la vegada i, en general, té una amplitud ecològica i una àrea de distribució més grans. Els líquens que només tenen apotecis o peritecis es reprodueixen dispersant espores. En germinar, les que han caigut en un lloc adequat s’associen aviat amb algues preexistents, lliures o escapades del tal·lus o de propàguls d’altres líquens. Aviat el nou liquen només conserva les algues del fotobiont habitual, i les altres són eliminades.
Aspectes, de fisiologia i ecologia
Els metabòlits secundaris
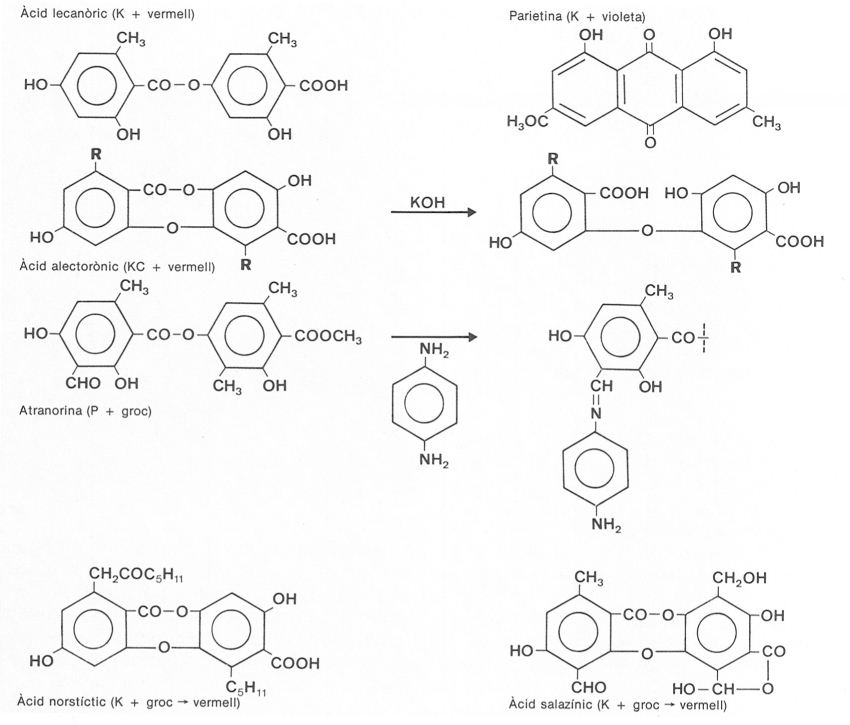
Fórmula, desenvolupada d’algunes substàncies químiques presents en els líquens, i exemples de reaccions químiques de taca. Els líquens poden contenir substàncies liquèniques específiques. Entre les de la figura, l’àcid lecanòric i l’atranorina són dèpsids, els àcids alectorònic, norstíctic i salazínic són depsidones (vegeu el doble pont d’unió entre els cicles) i la parietina és una antraquinona. Els liquenòlegs fan servir molt sovint les reaccions de taca per a determinar les espècies de líquens. Algunes de les més típiques són indicades al costat de les substàncies respectives, en el dibuix. En el cas de la reacció KC, l’aplicació inicial de KOH provoca el canvi químic indicat, i la nova substància dóna reacció acolorida amb C. En el cas de la parafenilendiamina (P+ groc) hi ha un trencament de la molècula i una unió del reactiu amb una de les parts, per a donar un pigment groc.
Biopunt, a partir de fonts diverses.
Moltes de les substàncies que contenen els líquens són les normals en molts éssers vius, o en els fongs i les algues (quitina, clorofil·la, cel·lulosa, etc.); tanmateix, alguns líquens contenen uns polisacàrids particulars, les liquenines, que produeixen reacció I+ blau (donen color blau amb els reactius iodats) i són comestibles. Però ens aturarem breument en els aproximadament 350 metabòlits secundaris (dits substàncies liquèniques) que poden contenir, perquè, ultra llur originalitat (la majoria només els saben fabricar els líquens), molts d’ells tenen un gran interès taxonòmic. Aquest interès ja començà a ésser explotat per Nylander quan, el 1866, proposà la utilització de les reaccions de taca, emprant com a reactius una solució d’hipoclorit càlcic, que conté clor, i una d’hidròxid potàssic. Hom utilitza també altres reactius, com el Lugol o la parafenilen-diamina en solució alcohòlica. L’aplicació d’una goteta de reactiu sobre el còrtex, o la medul·la, l’epiteci, etc., pot anar acompanyada (reacció positiva) d’un canvi de color o no (reacció negativa). Els liquenòlegs expressen el resultat de la reacció escrivint primer la part del liquen tractada, després el símbol del reactiu seguit del signe + i el color de la reacció, o, si no hi ha hagut canvi de color, el signe−. Així, en el cas de Pertusaria amara, si diem que té els soralis KC+ vinós, indiquem que aplicant als soralis primer KOH i després hipoclorit, apareix un color roig vinós. Còrtex K− vol dir que no hi ha reacció.
El reactiu K és una solució concentrada de KOH en aigua. Pot produir color groc, estable, o lentament canviant vers el vermell. El reactiu C és lleixiu comercial, millor si és concentrat. Pot donar color taronja o vermell carmí. El reactiu I és lugol (iode dissolt en una solució aquosa de iodur potàssic); pot donar color vermellós o blau. El reactiu P és una solució de para-fenilen-diamina (10 mg) en alcohol etílic (1 cm3); es manté utilitzable només durant algunes hores, i pot donar color groc taronja o vermell (convé evitar-ne el contacte amb la pell, perquè és cancerigen). El reactiu N es prepara amb 1/3 d’àcid nítric comercial i 2/3 d’aigua.
Les substàncies liquèniques es troben fora de les cèl·lules, sovint incrustant les hifes medul·lars, el còrtex superior o l’epiteci. Són solubles en acetona o cloroform. Les més freqüents són substàncies amb dos cicles fenòlics, com ara els dèpsids (para i meta), amb un pont —COO—, i les depsidones (amb un pont —O— a més del pont anterior). Hi ha també antraquinones, àcids úsnics, derivats de l’àcid pulvínic, terpenoides, etc. Cada espècie pot contenir una o unes poques substàncies liquèniques (a vegades, cap). Determinades espècies presenten una certa variabilitat (races químiques) Hom procedeix a isolar les substàncies liquèniques i a identificar-les mitjançant tècniques senzilles de cromatografia de capa fina (TLC), o de microcristal·lització (a partir d’extrets en acetona) o, més complexes, de cromatografia de gas o espectrometria de massa. El contingut químic de metabòlits secundaris de cada espècie és un caràcter molt important a l’hora de separar espècies i, fins i tot, gèneres o grans grups. Per exemple, la presència d’antraquinones és un caràcter freqüent en les telosquistals. Les substàncies liquèniques poden ésser responsables de la poca acceptació que tenen molts líquens per part dels herbívors, o poden contribuir a atacar el substrat (acció important en l’edafogènesi) i a solubilitzar determinats ions. En molts casos, com passa amb molts metabòlits secundaris, no se sap de moment per què serveixen. Algunes han estat emprades des de temps antic en tintoreria (per exemple, l’orxella) i en perfumeria («mousse de chêne»).
Creixement i requeriments

Una branca de roure abundantment colonitzada per líquens diversos. Hi ha els tal·lus fruticulosos de Ramalia farinacea (groc pàl·lid, sense apotecis) i de R. fastigiata (del mateix color, amb apotecis terminals), els tal·lus foliacis de Pleurosticta acetabulum (tal·lus verd olivaci), de Parmelina quercina (gris, amb apotecis bruns), de Xanthoria parietina (taronja) i petits tal·lus crustacis, de Lecidella elaeochroma, Rinodina, etc.
Antonio Gómez-Bolea.
Els líquens creixen molt més lentament que els fongs no liquenificats: increments anuals de radi de 0,5 a 1 mm són normals entre els líquens crustacis; els foliacis poden créixer de 2 a 7 mm per any, amb valors extrems d’uns 25 mm; els fruticulosos augmenten de llargada a una velocitat semblant. Aquesta lentitud és un caràcter intrínsec, genètic, que es conserva en els simbionts un cop isolats. La successió, en les comunitats de líquens, és, doncs, molt més lenta. La mida dels líquens pot ajudar a datar l’antiguitat del substrat (liquenometria, aplicada en la datació de les morenes).
Les hifes del micobiont es poden relacionar amb les cèl·lules del fotobiont d’una manera superficial, o formant apressoris, o penetrant a través de la paret i formant haustoris que, com els dels fongs no liquenificats, mai no trenquen el plasmalemma de la cèl·lula on entren. En la proximitat del micobiont, o per efecte dels haustoris, les cèl·lules del fotobiont augmenten la permeabilitat de llur membrana, i arriben a cedir un 80% del que fotosintetitzen, en forma de sorbitol o arabitol (algues) o de glucosa (cianofícies), que el micobiont s’encarregarà de transformar aviat en mannitol. Si allunyem l’alga de la influència del fong, la paret perd la major part de la seva permeabilitat i l’alga es queda per ella gairebé tots els seus fotosintetitzats. Els líquens que posseeixen cianofícies amb heterocists al tal·lus o als cefalodis tenen capacitat de fixació de nitrogen.
L’aigua és un factor essencial per a la vida dels líquens, que es comporten com a organismes poiquilohidres i toleren sense dany assecaments periòdics. De fet, el pas freqüent de la forma deshidratada (que comporta criptobiosi, o atenuació extrema de les funcions vitals) a la forma hidratada, biològicament activa, és una necessitat per al bon funcionament del liquen. Un tal·lus plenament hidratat pot contenir del 140% (en líquens crustacis) al 300% (en líquens foliacis) i fins al 600% (en líquens gelatinosos) d’aigua, en percentatge de pes sec; l’activitat és, però, òptima amb un contingut d’aigua inferior al màxim, que permet la circulació dels gasos. Molts líquens, i especialment els que tenen cianofícies, necessiten aigua líquida per a hidratarse, però d’altres (sobretot els formats amb Trentepohlia) poden absorbir aigua a partir de l’aire bastant humit (80-95%), de la condensació de rosada o de la boira; alguns d’aquests poden viure en extraploms i coves, on no plou, i llavors parlem de líquens ombròfobs o aerohigròfils. D’altres toleren una immersió més o menys llarga; són els hidròfils. Els líquens dels llocs àrids (xeròfils) rarament han de suportar llargs períodes sense hidratació, ja que absorbeixen sovint rosada o humitat a partir de l’aire humit nocturn. Els deserts litorals, que reben boires i rosada, acostumen a ésser rics en líquens; no passa el mateix, en canvi, en les zones interiors del Sàhara, per exemple.
Malgrat llur resistència a les temperatures extremes quan el tal·lus és sec, hi ha líquens de llocs freds (psicròfils, sobretot a les muntanyes altes) i espècies que exigeixen hiverns temperats (termòfils, freqüents a les nostres àrees costaneres; aclarim aquí que qualifiquem de tèrmica l’àrea que gaudeix d’hiverns temperats, on només molt rarament gela).
Pel que fa a la llum, hi ha líquens fotòfils, molt exigents d’il·luminació, i esciòfils, que poden viure en llocs poc il·luminats, com ara els boscos densos o l’entrada dels avencs. L’exposició al sol, sobretot durant el matí, fa evaporar aviat l’aigua absorbida durant la nit. Els líquens que ho toleren són els heliòfils, i els que defugen la insolació i es refugien a les superfícies obagues són els heliòfobs. Alguns líquens de llocs alts necessiten, durant l’hivern, la protecció de la neu; són els líquens quionòfils (del grec quionos, neu). Els que viuen en lloc sempre lliures de neu i glaç són els quionòfobs
Els nutrients que, en petita quantitat, necessiten els líquens, els arriben principalment amb la pols o amb els esquitxos de la pluja. Els líquens que s’adapten a una reduïda aportació de nutrients s’anomenen anitròfils, per contraposició als nitròfils, que viuen en superfícies ben adobades. Quan l’origen dels nutrients és la pols, sobretot la que s’origina a les zones poblades, on hi ha bestiar, residus orgànics, etc., s’afavoreix l’aparició de líquens coniòfils. Quan els nutrients d’una superfície procedeixen dels excrements dels ocells que hi freqüenten, s’hi instal·len líquens ornitocopròfils.
Pel que fa a la natura del substrat on viuen, si es tracta de pedra, tenim els líquens saxícoles, que poden ésser acidòfils, neutròfils o basòfils. Sovint els acidòfïls són silicícoles, si viuen sobre silicats més o menys àcids. Els basòfils viuen sovint sobre roques carbonatades (que bullen amb una gota de ClH diluït) i s’anomenen caldcoles. Els líquens epífits, que viuen sobre arbres o arbusts (els foròfits) són corticícoles. La fusta descorticada, de troncs o branques, o bé la fusta de pals, cledes i d’edificis situats a la intempèrie, porta, sobretot a la muntanya, líquens lignícoles. També a la muntanya, podem trobar líquens briòfils, sobre les molses, o turfòfils, sobre el sòl amb restes orgàniques més o menys descompostes.
Els sòls de les comunitats vegetals prou obertes (boscos clars, landes, brolles, timonedes) presenten líquens terrícoles, molt especialitzats en el cas dels gipsícoles, que viuen sobre sòls guixencs.
La sensibilitat a la composició atmosfèrica
Els líquens, i especialment certes espècies, són reconeguts mundialment com els organismes més sensibles davant els efectes nocius de la contaminació atmosfèrica. Això és així gràcies a la natura peculiar dels simbionts, que depenen gairebé exclusivament de l’atmosfera o de l’aigua que circula sobre el substrat. A més, el metabolisme i, per tant, el creixement, són lents. En un tal·lus de liquen, una biomassa important de fongs depèn de comparativament pocs fotobionts. Per això, qualsevol substància que redueixi la capacitat de fotosíntesi d’aquests darrers pot portar al liquen a consumir més matèria orgànica de la que fabrica, amb el resultat de la seva mort.
Algunes de les raons per les quals hom utilitza els líquens com a monitors biològics es poden resumir en els següents punts. En primer lloc, tenen un tal·lus perenne, sense cutícula ni estomes i, per tant, amb la superfície constantment oberta al pas de líquids i gasos durant tota llur vida, que és llarga. En segon lloc, hi ha una relació fisiològica íntima entre els tal·lus i l’ambient, i no depenen directament del substrat per a llur nutrició. En tercer lloc, són molt estenoics, és a dir, d’exigències ecològiques precises, i molt sensibles als canvis que es poden presentar a l’ambient, ja que no disposen de mecanismes de defensa tan importants com pot ésser el control del seu contingut d’aigua (són poiquilohidres), entre altres; en canvi, són capaços de concentrar i acumular diferents compostos en el seu tal·lus. Hi ha tota una escala d’espècies de líquens que tenen requeriments i sensibilitats diferents, de manera que el mal causat pels diferents contaminants sobre cadascuna és selectiu i diferenciat; per això, els líquens actuen com a monitors continus i integradors de les condicions ambientals i de llurs canvis.
L’anhídrid sulfurós (SO2) és el contaminant gasós que afecta més el funcionament normal dels tal·lus liquènics, i és també el més difós i el que presenta concentracions més altes. Es calcula que, com a resultat de l’activitat humana, en són abocades unes 200 t a l’any a l’atmosfera, a més d’òxids de nitrogen, hidrocarburs, derivats fluorats o clorats, etc., juntament amb partícules en suspensió, metalls pesats i molts altres compostos. La transformació d’aquestes substàncies en àcids a l’atmosfera provoca les pluges àcides, que tenen un efecte negatiu sobre els líquens, ja que incrementen els efectes directes del SO2, perquè redueix el pH dels substrats i això provoca una pèrdua de la capacitat d’amortiment que aquests tenen de natural. Cal precisar, però, que a la península Ibèrica només ha estat possible detectar fins ara pluges àcides en alguns punts de i del massís del Montseny.
Si bé ha estat menys estudiada, al nostre país pot ésser més important encara l’anomenada contaminació fotoquímica, és a dir, la produïda per l’aparició a l’atmosfera d’agents oxidants que s’originen quan reaccionen entre ells els òxids de nitrogen, els hidrocarburs i l’oxigen, en presència de la radiació ultraviolada del sol.
Les diferències en el grau de sensibilitat que cada espècie de liquen ofereix a la contaminació atmosfèrica són el resultat d’un equilibri entre la seva capacitat amortidora dels efectes del SO2 i la seva tolerància. Els mecanismes d’acció són complexos, i és més senzill fixar-se en algunes de les respostes dels líquens al SO2 i els seus mecanismes de resistència. Per exemple, una de les primeres reaccions de tal·lus orientades a minimitzar els efectes es basa en intentar reduir la captació d’aquest gas, cosa que aconsegueixen limitant la superfície de contacte amb l’aigua que porta el contaminant dissolt, sia desenvolupant soredis (que no s’hidraten fàcilment amb aigua líquida) o elaborant concentracions més altes de substàncies hidròfobes (àcid fumarprotocetràric) o encongint el tal·lus a base de fer que els lòbuls siguin més estrets i convexos, o tractant de reduir l’efecte dels ions tòxics gràcies a la capacitat de tamponament del tal·lus i del substrat. En canvi, la tolerància depèn de la resistència del protoplasma de les cèl·lules, de l’estat del metabolisme, de la vitalitat, de la capacitat d’inhibició dels efectes i del grau de maduresa del tal·lus. En un mateix lloc, la mateixa espècie es comporta com a més sensible durant les seves fases juvenils que quan el tal·lus es troba en plena maduresa. Aquesta és una de les raons de la minva de la diversitat i la biomassa dels líquens a les zones contaminades: simplement, o bé no es poden reproduir o bé els tal·lus més joves deixen de progressar.
Els efectes més importants són els que afecten la fotosíntesi i la respiració: la captació de CO2 és inhibida, la composició de les membranes cel·lulars s’altera, permetent la pèrdua d’ions potassi, la clorofil·la es transforma en feofitina (bruna) per pèrdua d’ions magnesi, comencen les decoloracions del tal·lus i diverses modificacions; més endavant, també es perd la respiració i, al final, tant el fotobiont com el fong degeneren, es malmet l’estructura del tal·lus i es perd la capacitat de reproducció. Algunes substàncies liquèniques es poden veure afectades pels compostos que es desprenen de les membranes alterades, originant així substàncies acolorides. Un cas molt conegut és el d’algunes espècies de parmèlies, com ara Parmelia sulcata, P. tiliacea i P. quercina, que adquireixen una tonalitat vermella, de manera que si en un bosc veiem sobre els troncs líquens foliacis amb tonalitats vermelloses, això ens pot indicar la presència de SO2 a l’ambient. La sensibilitat d’una mateixa espècie també es pot modificar segons les condicions i la situació geogràfica. Per exemple, Parmelia quercina, Physconia distorta o Anaptychia ciliaris, que són considerades com espècies extremament sensibles al nord d’Europa, es comporten com a resistents a la península Ibèrica, on viuen òptimament.
La importància de l’acció del contaminant depèn també de la seva persistència i concentració, del règim climàtic, del pH del substrat, etc. Es produeix una associació complexa d’efectes fisiològics difícil de reproduir al laboratori. La naturalesa del substrat és molt important perquè, segons la seva capacitat per a aminorar els efectes de l’aciditat, la contaminació podrà afectar més o menys els líquens. A les ciutats, no és difícil trobar monuments fets de pedra calcària o murs amb morter que presenten líquens, ja que són capaços de neutralitzar les substàncies àcides. Si les roques són silícies això ja és més difícil. Les escorces dels arbres tenen un pH molt variat: els roures i les alzines s’apropen a la neutralitat, els pollancres i els oms la tenen clarament bàsica i les coníferes, àcida. Els líquens que viuen sobre l’escorça són els més adequats per a realitzar una valoració de la qualitat de l’aire, però la flora que contenen és diferent segons el tipus d’escorça. Això es veu bé quan les escorces bàsiques o neutres comencen a acidificar-se a causa de l’efecte dels contaminants atmosfèrics: a poc a poc, la vegetació liquènica normal va essent substituïda per una altra, formada per espècies més acidòfiles. Així, si trobem Pseudevernia furfuracea o Hypogymnia physodes sobre alzines, podem pensar que és un indici de la presència de SO2 a l’atmosfera.
Hom ha creat mètodes de treball molt diferents per a estudiar la distribució dels líquens a les zones d’igual contaminació que apareixen al voltant de les fonts de pol·lució i per relacionar-la amb el dany causat als líquens en augmentar la concentració dels contaminants. Ja l’any 1965, a Gran Bretanya, hom estudià la distribució de Parmelia saxatilis i es va trobar que el contingut mitjà de SO2 en l’aire, a l’hivern, anava més correlacionat amb la distribució dels líquens que els valors mitjans anuals; també es va veure la importància de la configuració del terreny i dels vents dominants com a factors que influïen en la sensibilitat de l’espècie.
Algunes anàlisis qualitatives es basen en la distribució individual de les espècies que, un cop correlacionades amb les quantitats mitjanes de SO2, indiquen la diferent resistència de cadascuna. Però no tenen en compte ni la vitalitat ni la cobertura dels tal·lus. Són molt més interessants els mètodes que estudien les associacions de líquens, ja que és així com acostumen a desenvolupar-se en la natura. Així, Hawksworth i Rose aconseguiren (1970) mercès a una xarxa d’estacions de mesura de SO2 en nuclis urbans industrials, realitzar una escala de sensibilitat dels diversos líquens en relació amb les concentracions mitjanes de SO2 que podia tolerar cada espècie a l’hivern, a la Gran Bretanya. Aquesta escala fou desdoblada en dues, segons si l’arbre era d’escorça neutra o àcida. La primera adaptació d’aquesta escala a la península Ibèrica, fou aplicada a Madrid, i posteriorment, a València, Galícia i Astúries. En qualsevol d’aquestes escales, la sensibilitat dels líquens permet apreciar concentracions tan baixes com de 30 µg/m3 de SO2. Les espècies hi apareixen agrupades en diferents nivells, i ordenades de més resistents a més sensibles; la zona 1 correspon sempre al «desert liquènic», i hom distingeix fins a un nivell 10, que és el que presenta la màxima diversitat d’espècies i la presència de les espècies més sensibles.
Partint també de les comunitats de líquens, podem obtenir un índex de puresa atmosfèrica (IPA), proposat per De Sloover i Leblanc. El càlcul d’aquests índexs per a cada punt exigeix prendre diversos inventaris fitosociològics en un grup d’arbres, en condicions ecològiques homogènies, prenent nota de la cobertura, l’abundància i la freqüència de les espècies en cada comunitat i en uns quants arbres de cada punt. Amb els valors obtinguts es poden traçar línies d’isocontaminació i obtenir mapes que expressen de manera senzilla el grau de contaminació dels diferents punts de l’àrea estudiada. L’aplicació dels IPA als Països Catalans s’inicià amb dos treballs, un sobre els voltants de Sabadell i l’altre sobre el Tarragonès.
Hi ha també una tècnica basada en el transplantament de discs d’escorça amb líquens des de llocs on l’atmosfera és pura (teòricament), a zones contaminades. Hom observa després els efectes que es produeixen sobre els líquens en ambient diferents.
Altres tipus de contaminació, com els deguts als nitrats de peroxiacil 2, l’ozó o als derivats fluorats, poden ésser detectats pels líquens. Pel que fa als metalls pesats, els líquens els poden acumular tant a la superfície de les hifes com a les parets cel·lulars, concentrant-los a partir de solucions diluïdes. Per això va bé estudiar-los per a controlar l’evolució d’aquests contaminants i inferir els diversos models d’emissió a l’atmosfera. Els seus efectes desapareixen ràpidament en augmentar la distància a la font contaminant i només els que poden penetrar les cèl·lules tenen acció directa sobre la respiració o la fotosíntesi. El plom i el mercuri són els més perjudicials, però el zenc té molt influència sobre els líquens amb cianofícies.
Els grans grups de líquens
Actualment una gran part dels líquens són considerats dins els ascomicets, que podem anomenar ascomicets liquenificats, mentre que una petita part és considerada dels basidiomicets, els basidiomicets liquenificats. En trobem, principalment entre els ascomicets, també entre alguns basidiomicets i algun cop entre els deuteromicets i els ficomicets (cas de Geosiphon pyriforme), amb quatre possibles situacions: grups totalment liquenificats (només comprenen líquens), grups predominantment liquenificats (la major part dels representants són líquens), grups amb només alguns representants liquenificats i grups totalment no liquenificats. Ja podem imaginar que quan ens toqui explicar els líquens dels grups segon i tercer haurem de tornar a considerar alguns ordres ja vistos dins el gran grup de les ascomicètides. Es tracta dels ordres de les ostropals, les calicials, les leocials, les pezizals, les patel·larials, les esferials i les dotideals.

Separades pels seus caràcters microscòpics de les antigues Lecidea, les Lecidella, amb apotecis lecideïns, tenen afinitat amb les lecanores. Una de les més freqüents sobre roques enriquides amb nitrat és L. carpathica, de compartiments tal·lins convexos i de color gris blanquinós.
Javier Etayo.
Una de les lecanores més freqüents al nostre país, sobre escorça de tota mena de vegetals lignificats, és Lecanora sienae, que prefereix els llocs secs i assolellats. Noteu el disc dels apotecis, de color bru fosc, i el marge tal·lí, propi dels apotecis lecanorins.
Albert Masó.
Igual que la resta dels micòlegs, els liquenòlegs també s’han adonat que diversos caràcters molt aparents, com l’aspecte del tal·lus o les fructifïcacions, el nombre de septes a les espores, els sistemes de reproducció asexual, etc., sobre els quals s’havia basat fins no fa gaire l’ordenació (és a dir, la sistemàtica clàssica dels líquens), han aparegut, sovint d’una manera independent, en diverses línies evolutives. Per això, dos líquens amb el tal·lus crustaci, amb apotecis lecanorins o amb espores murals, poden pertànyer a grups molt separats evolutivament. En llur recerca dels caràcters més estables i segurs a l’hora de reflectir veritables afinitats naturals (és a dir, evolutives), els micòlegs i els liquenòlegs s’han centrat cada cop més en l’estudi de l’aparell reproductor: tipus de desenvolupament (des del primordi), tipus d’hamateci (els filaments i elements que acompanyen els ascs) i, sobretot, organització dels ascs, tenint en compte principalment si són o no fissitunicats i quin tipus tenen d’aparell apical; aquest és estudiat al microscopi òptic, amb un tractament previ amb KOH, un rentatge i una tinció amb solució iodada (lugol), o al microscopi electrònic. La consideració d’aquests caràcters (no sempre fàcils d’apreciar i de valorar) ha comportat un capgirament de la sistemàtica dels ascomicets, tant dels liquenificats com dels no liquenificats.


Dues parmèlies freqüents al nostre país. A l’esquerra, Parmelia saxatilis, una espècie preferentment saxicola, de muntanya, amb pseudocifel·les reticulades i isidis; la posem com a exemple del gènere en el sentit actual, restringit. A la dreta, Parmelina tiliacea, representant del gènere Parmelina, de tal·lus gris clar. És abundant tant sobre roques silícies com sobre escorces d’arbre, en llocs amb elevada humitat atmosfèrica (boira, rosada); de color gris clar, té la part central del tal·lus coberta d’isidis de punta bruna, que li fan agafar un cert aspecte brutós, i només quan les condicions són òptimes pot presentar apotecis, bruns.
Volkmar Wirth i Jordi Vidal
Seguint les propostes d’Eriksson, Hawksworth i Hafellner, disposarem els ascomicets liquenificats de la manera següent. En primer lloc tractarem els ordres totalment o gairebé totalment liquenificats, amb l’àpex de l’asc típicament engruixit formant un tolus amiloide i amb apotecis: lecanorals (Lecanorales), telosquistals (Teloschistales), peltigerals (Peltigerales), pertusarials (Pertusariales) i liquinals (Lichinales), que en conjunt constitueixen el 78% dels nostres líquens. En segon lloc, els ordres amb ascs funcionalment unitunicats, amb apotecis, peritecis o histerotecis: gialectals (Gyalectales), ostropals (Ostropales), calicials (Caliciales), leocials (Leotiales), pezizals (Pezizales), grafidals (Graphidales) i esferials (Sphaeriales), que constitueixen el 3,6% dels nostres líquens. Finalment i en tercer lloc, els ordres amb ascs fissitunicats i amb ascomes apotecioides, peritecioides o histerotecioides: artonials (Arthoniales), opegrafals (Opegraphales), gomfil·lals (Gomphillales), patel·larials (Patellariales), verrucarials (Verrucariales), pirenulals (Pyrenulales) i dotideals (Dothideales), que constitueixen el 17,4% de les espècies trobades al nostre país. Hi ha algunes famílies que encara no han pogut ésser atribuïdes amb seguretat a cap dels ordres esmentats. A més, resta un grup de líquens que no formen mai fructificacions sexuals, que estudiarem en el grup anomenat líquens estèrils. Els pocs basidiomicets liquenificats pertanyen als ordres de les atelials (Atheliales), les faneroquetals (Phanerochaetales), les cantarel·lals (Cantharellales) i les tricolomatals (Tricholomatales). L’amplitud dels ordres en nombre d’espècies és molt desigual. Només els ordres de les lecanorals (61%), les telosquistals (8,6%) i les verrucarials (6,8%) representen més de les tres quartes parts de les gairebé 1280 espècies inventariades fins ara (1989) als Països Catalans.