Els fongs lisotròfics: el tipus d’organització fong

Els fongs lisotròfics poden originar, per simple combinació d’hifes especialitzades, aparells fructífers molt especialitzats. En aquesta visió parcial d’un cos fructífer (basidiocarp) jove d’un cortinari (Cortinarius purpurascens), veiem la carn del barret i del peu, i les làmines (on hi ha l’himeni fèrtil); les hifes hi estan massa juntes per a ésser visibles. En canvi, veiem prou bé les que reuneixen el marge del barret amb el peu, que formen el que hom anomena una cortina. Les hifes superiors de la cortina s’han tenyit de bru a causa de les espores, ja madures, que van caient des de les làmines.
Josep M. Vidal / SCM.
Els fongs són organismes heteròtrofs i, ja que no poden realitzar la fotosíntesi, necessiten captar i oxidar molècules riques en energia. Aquestes substàncies orgàniques, els aliments, han d’ésser presents al substrat colonitzat pel fong. Hom acaba de veure els organismes que capten l’aliment per fagotròfia, els fongs ameboides. Però la majoria dels organismes estudiats pels micòlegs els capten per digestió externa o lisotròfia, que comporta la degradació, fora de la cèl·lula, dels aliments continguts al substrat, mitjançant l’activitat d’enzims digestius, dissolts en aigua, emesos pel fong. Aquests enzims trenquen les molècules massa grosses del substrat, que queda reduït a d’altres prou petites per poder ésser absorbides pel fong. Hi ha d’haver, doncs, una difusió, en medi aquós, d’enzims des del fong fins al substrat, i de molècules producte de la digestió, en sentit invers. Res d’això no funcionarà si no hi ha prou aigua, i d’ací ve la relació entre l’activitat dels fongs i la disponibilitat d’aigua.
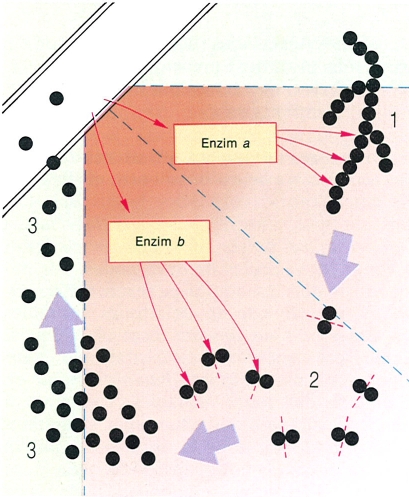
La digestió externa del substrat (lisotròfia) i l’absorció dels productes resultants en ambient aquós, base del sistema alimentari dels fongs, ha estat adoptada per dos grans branques de fongs: els pseudofongs i els fongs veritables. L’hifa del fong (a dalt, a l’esquerra) allibera enzims digestius al substrat extern, i aquests actuen sobre els polímers insolubles (1), fent-los passar a unitats solubles més petites (3) a través d’un o més estadis intermedis (2); finalment, l’hifa els absorbeix. El gradient de color indica els gradients de concentració dels enzims.
Biopunt, a partir de E. Moore-Landecker.
És comprensible que aviat s’esgotin els aliments del substrat proper al fong, i aquest es vegi obligat a continuar creixent, allargant-se i explorant nous substrats, penetrant-hi profundament. Per això, la majoria dels fongs tenen un aparell vegetatiu filamentós, en forma d’hifes, fines, tubulars, amb una gran capacitat de penetració, de captació d’aliments i de transport del que han absorbit. El conjunt de les hifes, és a dir, el miceli dels fongs està en contínua activitat de creixement, ramificació i exploració, sempre que no li falti un mínim d’humitat. A vegades, el substrat ja conté molècules d’aliment prou petites (com passa en el cas dels llevats, o dels fongs paràsits, que penetren entre les cèl·lules de l’organisme parasitat o al seu interior), que seran absorbides directament, sense necessitat de digestió. Per això, el cos vegetatiu pot ésser format per unitats petites, separades entre elles.
A primera vista, i a causa del predomini de l’aparell vegetatiu format per hifes, el grup dels fongs lisotròfics sembla força homogeni. Però recentment, s’han anat acumulant dades, sobretot bioquímiques i ultrastructurals, que han portat al convenciment que, en realitat, reuneix artificialment els representants de dues grans línies evolutives, una derivada d’un grup d’algues, i una altra de més propera als protozous i als animals. Ambdós grups, partint d’un origen tan diferent, s’han adaptat a la vida lisotròfica, és a dir, a alimentar-se per digestió externa. No és estrany que la mateixa necessitat d’explorar el substrat i de captar aliments hagi conduït a un tipus d’hifes d’aspecte força semblant, que donen així la impressió d’un sol grup homogeni. Però entre els dos grups, que hom anomenarà des d’ara el dels pseudofongs (Pseudofungi), o fongs algals, i el dels fongs veritables (Fungi), hi ha diferències considerables, que posen de manifest llur distància evolutiva. Vegem-ne les principals.
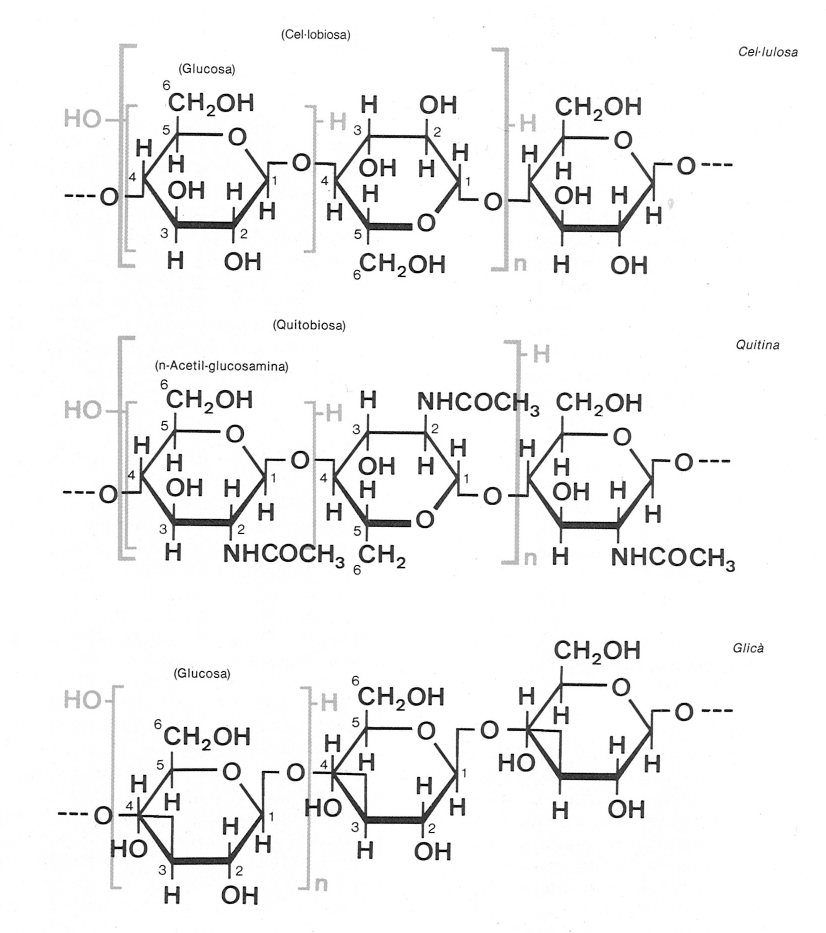
L’estudi de l’estructura i la composició de la paret cel·lular ha permès de detectar l’existència de dues grans vies filètiques dins els fongs: d’una banda, els pseudofongs, provinents d’algun tipus d’alga unicel·lular, que contenen cel·lulosa com a component fibril·lar de la seva paret; de l’altra, els fongs veritables, en els quals la mateixa funció és a càrrec de la quitina i tenen, per això, un origen més pròxim a algun grup d’animals unicel·lulars. El dibuix presenta també l’estructura del glicà, un dels components més freqüents (ascomicets, basidiomicets, etc.) de naturalesa no fibríl·lar. La diferència entre les tres substàncies és deguda al monòmer i al tipus d’enllaços. Observem com, en els components fibríl·lars, els monòmers estan girats 180 graus l’un respecte de l’altre, equilibrant així la molècula.
Biopunt, a partir de fonts diverses.
En primer lloc, cal fixar-se en la paret cel·lular, que consta en tots dos grups d’un component fibril·lar (comparable a les tiges de ferro del formigó armat) i un componentmatricial (comparable al formigó que engloba el ferro). Com a component fibril·lar, els pseudofongs tenen cel·lulosa, igual que les algues. En canvi, els fongs veritables tenen quitina, un polímer semblant al que produeixen molts animals (insectes, crustacis). Una altra diferència notable rau en les cèl·lules flagel·lades, que en els pseudofongs tenen dos flagels eucariòtics (més propiament anomenats cilis o undulipodis), un d’anterior, tractor (que tiba la cèl·lula cap endavant) gràcies als mastigonemestubulars (varetes laterals buides), i un de posterior, llis i una mica més curt (a vegades, reduït al corpuscle basal). Aquest tipus de dotació flagel·lar és més que semblant a la dels heterocontòfits. En canvi, en els fongs veritables, les cèl·lules flagel·lades (només presents en els grups més primitius) presenten un sol flagel, llis i posterior, que actua com la cua d’un peix o d’un espermatozoide.
Hi ha d’altres diferències ultrastructurals que afecten la zona de transició entre els flagels i els cinetosomes (corpuscles basals dels flagels, de tipus centriolar), les rels flagel·lars, i l’aparellapical de les hifes, que, en els pseudofongs, té vesícules de dues mides. Els mitocondris dels pseudofongs tenen crestes tubulars, mentre que els dels fongs veritables les tenen de tipusaplanat, com els animals. També cal esmentar el cicle vital, que és diplont en els pseudofongs i haplo-dicariòtic en la majoria dels fongs veritables. També en el camp del metabolisme s’han anat acumulant dades que donen fe de la distància evolutiva existent entre els dos grups. Hom esmentarà només la diferent via metabòlica (la de l’àciddiaminopimèlic, en els pseudofongs i en les plantes, i la de l’àcidaminoadípic, en els fongs veritables) que condueix a la biosíntesi de l’aminoàcid lisina (que els animals no saben sintetitzar), la síntesi d’àcid nicotínic a partir de compostos de tres àtoms de carboni (en lloc de fer-ho partint del triptòfan, com ho fan els fongs veritables i els animals) i també la presència d’hidroxiprolina a la proteïna de la matriu de la paret, el tipus d’ARN ribosòmic (de pes molecular diferent al dels fongs veritables), els enzims diferents que intervenen en la síntesi del triptòfan, i la incapacitat de formar pigments al miceli vegetatiu i de sintetitzar esterols.
Per aquestes i altres raons, hom fa derivar els pseudofongs de les algues heterocontòfites, potser de xantofícies filamentoses semblants a Vaucheria, o d’alguna crisofícia antiga. Cavalier-Smith proposa incloure’ls, al costat dels heterocontòfits i de les algues brunes, en un sisè regne, el de les cromistes (Chromista). En aquest esquema, que consta de cinc regnes (i no quatre, com el de Whittaker i Margulis) d’eucariotes, els fongs veritables formen, des dels quitridiomicets als deuteromicets, un sol regne, el dels fongs (Fungi); les algues verdes i les vermelles, juntament amb les molses i les plantes vasculars, configuren el de les plantes (Plantae); i els euglenòfits, els dinòfits, les craspedofícies i els fongs ameboides resten en el regne dels protozous (Protozoa). Els animals continuen en el regne d’aquest nom (Animalia). És consolidi o no aquest esquema en el futur, cal tractar primer, per separat, els pseudofongs, lligats a les algues, i passar després als fongs veritables (o, simplement, fongs), molt més propers als animals i, com ells, derivats d’un petit grup de protozous flagel·lats, amb un sol flagel llis i caràcters molt animalitzats, el de les craspedofícies que, per originar els fongs haurien passat d’una vida microfagotròfica o autotròfica a la lisotròfia, canviant llur forma i incorporant quitina a llur paret.
El concepte de pseudofong
Els pseudofongs es configuren com un grup molt particular, que conserva molts caràcters algals. Molts viuen en aigua dolça, com a sapròfits o com a paràsits d’algues, de fongs i fins i tot d’animals diversos. L’aparell vegetatiu pot ésser unicel·lular (com en moltes lagenidials i en molts hifoquitriomicets), o format per cèl·lules disperses en una xarxa filamentosa (en els labirintulomicets), però en general està constituït per filaments sifonats (sense septes, excepte a la base dels òrgans reproductors). Les zoòspores tenen dos flagels, l’anterior, amb mastigonemes tubulars, i el posterior, llis, a vegades (hifoquitriomicets) absent i reduït al cinetosoma.
El concepte de fong veritable
Els fongs veritables, que anomenarem simplement fongs, se’ns presenten com un grup homogeni i monofilètic, i per això ha estat tractat per alguns autors dins d’un regne propi, fongs (Fungi). Si en els pseudofongs (grup molt particular, que conserva molts caràcters algals) ens trobàvem amb una tendència evolutiva, tardana, a deixar el medi aquàtic i a adaptar-se a la dispersió de propàguls pel vent, aquí tornem a trobar, i amb més força encara, aquesta tendència. De fet, només el grup més primitiu, el dels quitridiomicets, s’ha adaptat a la vida aquàtica i té cèl·lules reproductores (gàmetes i espores) flagel·lades, però que ara són opistocontes, és a dir, que tenen un sol flagel, llis, dirigit enrere, que impulsa la cèl·lula com la cua d’un peix. Aviat, en aquesta línia evolutiva, assitim a la desaparició de les cèl·lules flagel·lades (ja en els zigomicets), al triomf de l’adaptació a la vida terrestre i a la dispersió d’espores transportades sobretot per l’aire.
La paret cel·lular conté gairebé sempre quitina, i mai cel·lulosa. Secundàriament, poden perdre la paret, per adaptació a la vida parasitària endocel·lular, en grups primitius. Només els quitridiomicets poden presentar reproducció per isogàmia, anisogàmia o oogàmia, i durant l’evolució observem una tendència a simplificar la reproducció sexual, que condueix, des de casos d’espermatització (fecundació d’un oogoni per gàmetes no flagel·lats) a la gametangiogàmia (amb fusió de gametangis, però sense diferenciació d’ovocèl·lules), fins a arribar a la somatogàmia (fusió de cèl·lules somàtiques que actuen com a gàmetes). Els cicles vitals són haplonts o haplo-dicarionts, rarament haplo-diplonts. Els diplonts són excepcionals. La fase que va tenint cada cop més èxit és la dicariòtica (amb dos nuclis haploides, heterogenis, per cèl·lula), que arriba a predominar del tot en els basidiomicets, el grup més evolucionat. El metabolisme dels fongs és molt més proper al nostre i al dels animals en general que al dels pseudofongs. Sintetitzen la lisina per la via de l’àcid aminoadípic; sovint sintetitzen carotenoides, que els poden conferir sensibilitat a la llum. Poden fabricar també una gran varietat de pigments (sovint fenòlics), una de les manifestacions d’un metabolisme molt ric en productes secundaris.
Pel que fa a espècies, un 99% dels fongs lisotròfics pertanyen a aquest grup; estan repartits de la següent manera (segons estimació del «Dictionaryof the Fungi», 1983): quitridiomicets (Chytridiomycetes): 575 espècies; zigomicets (Zygomycetes): 665; ascomicets (Ascomycetes): 28 000; basidiomicets (Basidiomycetes): 16 000; i deuteromicets (Deuteromycetes): 17 000. Seguint l’obra estàndard de Strasburger (1986) atribuïm a aquests grups, ben definits, la categoria de classe. Molts autors recents, però, atorguen als fongs veritables la categoria de divisió, amb el nom d’eumicots (Eumycota), i als grans grups esmentats la de subdivisió (Chytridiomycotina, Zygomycotina, etc.).
Característiques dels fongs veritables
Estructura i ultrastructura de l’aparell vegetatiu: les hifes

El miceli, l’aparell vegetatiu de la majoria dels fongs, és un conjunt de tubets filamentosos microscòpics, les hifes, que formen el «cos del fong», capaç d’alimentar-se i de formar els òrgans reproductors. Normalment, el miceli resta amagat dins el substrat nutritiu, però en el cas de la fotografia, es formen agregats d’hifes fora del substrat. Però no tots formen hifes: n’hi ha d’unicel·lulars que viuen dins altres cèl·lules, d’altres formen cadenes de cèl·lules, etc.
Joaquim Reberté.
L’aparell vegetatiu de la majoria dels fongs és filamentós. No ho és per casualitat: la forma de filament estret (normalment, de 5 a 15 μm de diàmetre) està ben adaptada per a penetrar el substrat, cercar-hi aliments i absorbir-los després per lisotròfia. Els filaments s’anomenen hifes i el conjunt de les hifes constitueix el miceli, un aparell vegetatiu gairebé sempre immers al substrat i difícil de veure.
La majoria dels fongs són, doncs, miceliars, i d’aspecte no pas gaire variat. Hi ha micelis sense septes (anomenats sifonats o cenocítics) i micelis amb parets transversals o septes, que típicament presenten una perforació, de forma simple o complexa. Si només disposéssim dels caràcters de l’aparell vegetatiu no podríem pas reconèixer la majoria dels fongs.
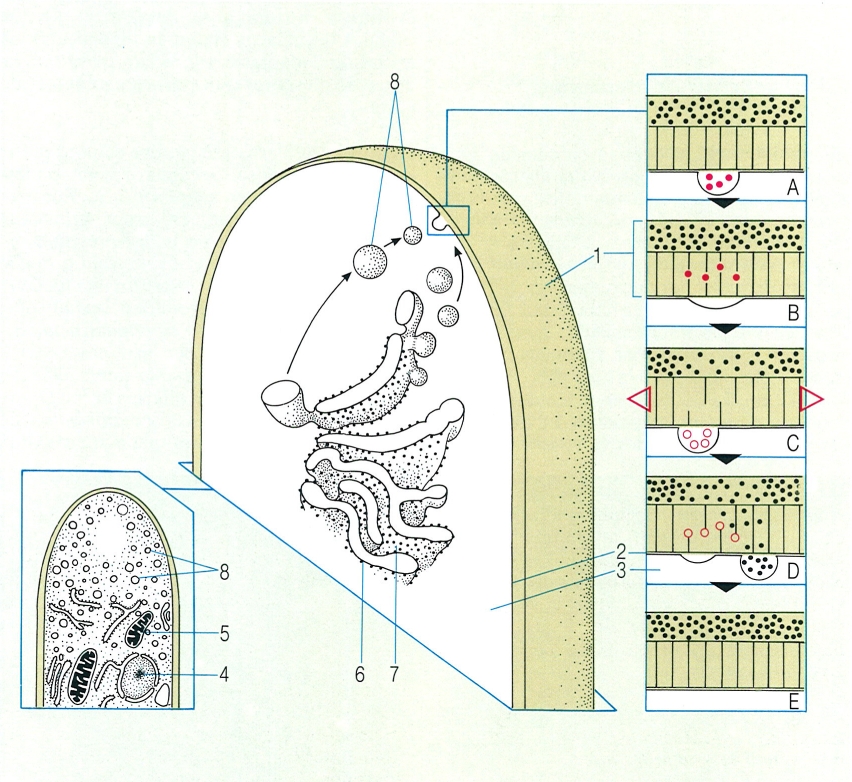
Esquema de la ultrastructura de l’àpex d’una hifa i detall de la transferència de materials durant el procés d’allargament de l’hifa. El reticle endoplasmàtic emet petites vesícules, que contenen les molècules necessàries per a formar noves estructures. Les vesícules emigren fins al pol apical de l’hifa, on es fusionen a la membrana cel·lular i aboquen el seu contingut per a formar la nova paret. El procés té una primera etapa d’estovament de les microfibril·les per acció d’enzims que les fragmenten (A, B), seguida d’un estirament de la membrana (C) i arribada d’enzims de síntesi de paret i de components amorfs de paret, que reconstitueixen l’estructura primitiva (D, E). Hom ha indicat en el dibuix: 1 paret, 2 plasmalemma, 3 citoplasma, 4 nucli, 5 mitocondri, 6 ribosoma, 7 reticle endoplasmàtic, 8 vesícula apical.
Biopunt, a partir de fonts diverses.
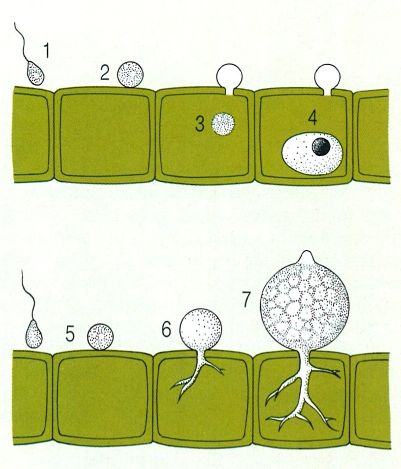
Exemples de fongs que no formen hifes. A dalt, el cas dels fongs paràsits endobiòtics: arribada (1), encistament (2), entrada del protoplast d’una zoòspora en una cèl·lula d’una alga (3), i creixement endocel·lular del cos vegetatiu, arrodonit, del fong (4). A baix, el cas dels fongs paràsits epibiòtics: la zoòspora s’enquista (5), penetra (6) la cèl·lula parasitada i forma ramificacions (rizomiceli) destinades a absorbir substàncies nutritives; la part externa, l’única reproductora, té forma d’olleta (quitridi) i es tornarà un esporangi epibiòtic (7).
Biopunt, a partir de fonts diverses.
D'altra banda,, no tots són filamentosos. N’hi ha d’unicel·lulars i sense paret, adaptats a viure a l’interior d’altres cèl·lules (paràsits endocel·lulars). D’altres formen un cist amb aspecte d’olleta sobre l’organisme que parasiten, i envien a l’interior branques sense nucli, molt ramificades, que formen el rizomiceli. Molts altres fongs poden formar, durant una etapa de llur vida o sempre, un cos vegetatiu integrat per cèl·lules disperses o reunides en cadenes curtes, que en general es reprodueixen activament per gemmació. Són els llevats o fongs en fase levuriforme, sovint adaptats a medis líquids i/o molt rics en aliments solubles.
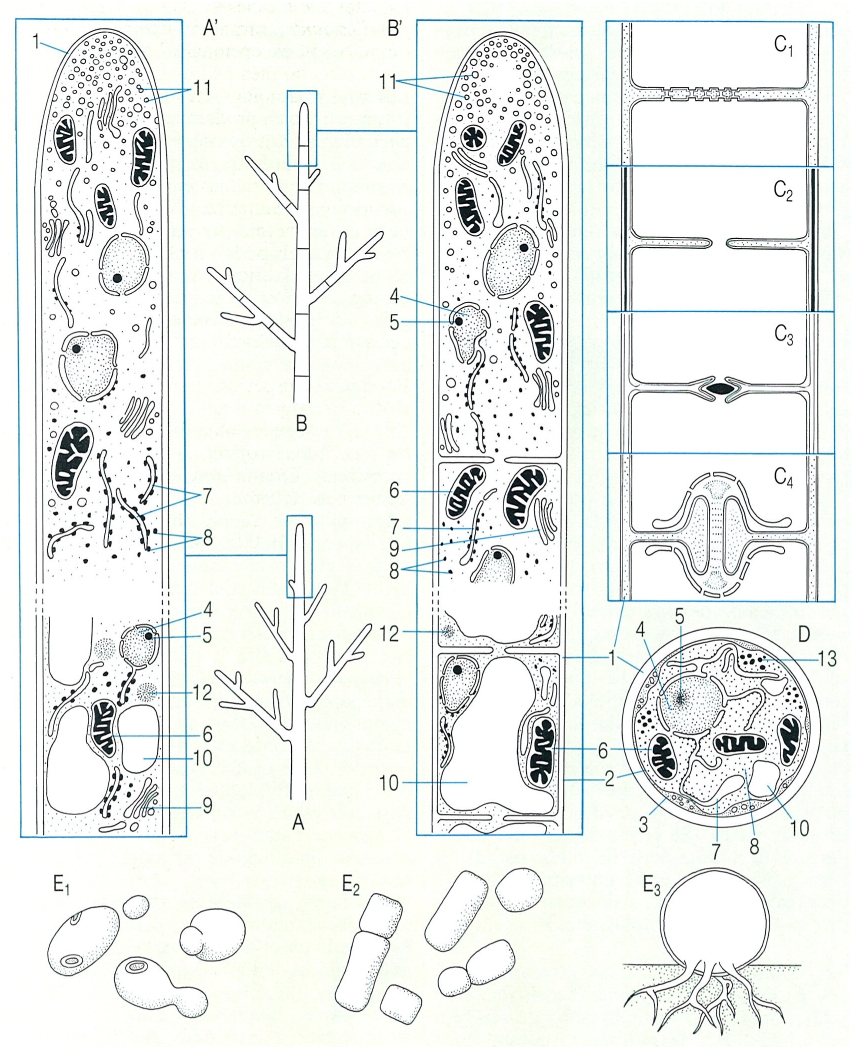
Morfologia general de les hifes i altres tipus d’organització en els fongs. A, A’ hifa ramificada i no septada de zigomicet. B, B’ hifa ramificada i septada d’ascomicet (noteu que a mesura que ens allunyem de l’àpex la vacuolització és cada cop més important, i que hi ha una àrea lliure de vesícules característica en la zona apical). C Estructura dels diferents tipus de septe en les hifes dels fongs: C1 septe amb micròpors, C2 septe amb un porus simple (típic de la majoria dels ascomicets), C3 septe dels tricomicets, C4 septe amb dolípor, amb parentesoma de reticle endoplasmàtic modificat, típic dels basidiomicets. D Secció transversal d’una cèl·lula de fong. E Altres tipus d’organització en els fongs: E1 cèl·lules disperses, amb multiplicació per gemmació (llevats); E2 cèl·lules disperses, amb multiplicació per fissió (llevats); E3 cos epibiòtic, amb rizomiceli (quitridiomicets). Hom ha indicat en el dibuix: 1 paret cel·lular, 2 plasmalemma, 3 lomasoma, 4 nucli, 5 nuclèol, 6 mitocondri, 7 reticle endoplasmàtic, 8 ribosoma, 9 dictiosoma, 10 vacúol, 11 vesícula apical, 12 corpuscles lipídics, 13 glicogen.
Biopunt, a partir d’Ingold, Deacon i altres fonts.
Com hem vist, les hifes són la base estructural de l’aparell vegetatiu de la majoria dels fongs. Ens les hem d’imaginar com a tubs de diàmetre gairebé constant, septats o no, molt ramificats, delimitats per una paret cel·lular rígida. Per l’interior, hi circula el protoplasma. La part apical té la paret més prima i és l’única que pot créixer i fer possible l’allargament i la ramificació de l’hifa. Darrera aquesta zona apical d’extensió, la paret es va fent més gruixuda (0,1 - 0,2 μm). Per dins, sovint força adherit a la paret, hi ha el plasmalemma (o membrana plasmàtica), que, en alguns punts, se’n separa i forma complicats plecs amb aspecte de vesícules, els lomasomes, de funció poc coneguda. Si no tenen septes, contenen molts nuclis distribuïts d’una manera més o menys regular. Si hi ha septes, cadascun dels compartiments que delimiten pot contenir un o dos nuclis, excepte el compartiment apical, implicat en l’allargament, que pot fer de 150 a 500 μm de llarg i contenir diversos nuclis. Es tracta sempre de nuclis eucariòtics típics, però de mida petita. El citoplasma conté també mitocondris, reticle endoplasmàtic, vacúols, ribosomes, cisternes de Golgi, gútules de lípids, etc.
El compartiment apical es caracteritza pel seu citoplasma dens i poc vacuolitzat, amb una acumulació prop de la punta de vesícules delimitades per membrana. Aquesta regió apical, constituïda per les vesícules, varia d’aspecte segons el gran grup, però també s’assembla més o menys al que podem trobar en els pseudofongs, en algunes algues o en els tubs pol·línics. A mesura que ens allunyem de l’àpex, anem trobant vacúols cada cop més grans. Arriba un moment que aquests ocupen la part central del compartiment. Els porus dels compartiments més vells es poden obturar. D’aquesta manera, el creixement dels vacúols empeny el citoplasma en direcció a la part més jove de l’hifa i en facilita l’allargament apical. A les cèl·lules madures, hi podem trobar altres substàncies de reserva, com ara glicogen, esterols, etc., a més de gotetes de lípids, si bé una part dels constituents de la paret poden servir també de substàncies de reserva. Les hifes velles, després de trametre a les joves la part més important de llur contingut, es poden desintegrar per autòlisi.
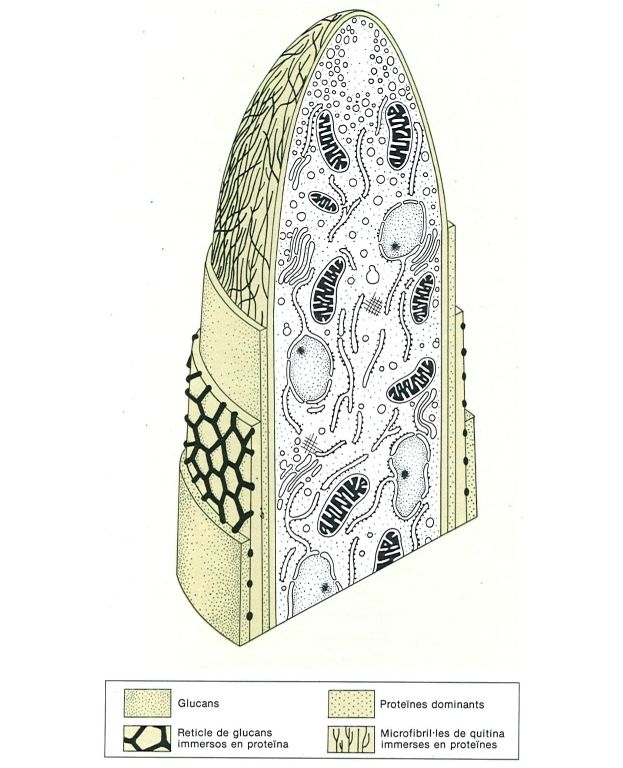
Estructura de la paret d’una hifa, amb indicació de les principals regions, en el cas de Neurospora (ascomicet). Només la capa interior porta microfibril·les de quitina. La composició varia, però, en els diferents grups de fongs.
Biopunt, a partir d’Hunsley i Kay, 1976, i altres fonts.
Les hifes tenen, doncs, una gran capacitat de transport, i això és un dels fenòmens que explica el gran èxit evolutiu dels fongs. Però, a més, tenen molta facilitat per a soldar-se o anastomitzar-se i intercanviar citoplasma, nuclis, reserves, etc. El que era un miceli ramificat es pot convertir aviat en una xarxa. Aquesta capacitat és encara poc desenvolupada en els grups més primitius. Les hifes són hialines, però hi ha grups que les tenen grogues, violàcies o brunenques (sovint, tenyides de melanina). En general, però, el color de les colònies en cultiu és causat per l’acumulació d’espores.
La paret és responsable de la forma de les hifes i de llur resistència a determinats canvis ambientals (com la pressió osmòtica) i conté sobretot polisacàrids, però també molts pròtids i lípids. La composició pot variar quantitativament, i fins i tot qualitativament, segons el gran grup o el moment del desenvolupament. Ja ha estat comentada la gran diferència que hi ha en la composició de la paret dels pseudofongs, però també s’observen diferències notables entre la paret dels zigomicets, la dels ascomicets miceliars, la dels llevats i la dels basidiomicets. Podem distingir entre els components fibril·lars, que en els fongs veritables corresponen en general a la quitina (un polímer d’acetilgalactosamina), i els matricials, que no formen microfilaments i omplen part dels espais i donen gruix; esmentem-ne els glicans (ramificats i no ramificats), els mannans (polímers de mannosa) i les proteïnes. Aquests components s’estructuren en diverses capes, de diferent composició. Un model especialment complex, però ben conegut, és el de la paret de Neurospora. La paret de la part apical de l’hifa és prima i conté sobretot quitina i proteïnes; després s’hi afegeixen altres capes, que n’augmenten progressivament el gruix i la rigidesa. A les espores, les parets són més gruixudes i complexes i contenen també lípids i, sovint, melanines, que les protegeixen contra les radiacions.
Pel que fa als orgànuls, el plasmalemma dels fongs conté esterols, sobretot ergosterol, a diferència del dels animals, que presenta colesterol, i del de les plantes, que conté substàncies semblants al colesterol. Per això, els compostos que afecten la síntesi del colesterol no els alteren (i, al contrari, els compostos que afecten la síntesi de l’ergosterol no afecten els animals). Els fongs veritables no presenten aparell de Golgi format per dictiosomes, però hi ha una o més cisternes de Golgi, que poden originar, per gemmació, sovint en col·laboració amb el reticle endoplasmàtic, les vesícules de la regió apical, riques en enzims i probablement relacionades amb la síntesi de paret. També hi ha els quitosomes, uns grànuls sense membrana que intervenen en la síntesi de quitina. Els altres orgànuls cel·lulars són semblants als de la resta dels eucariotes. Cal recordar, però, que els mitocondris tenen crestes aplanades (no discoïdals) i no tubulars com els pseudofongs. El reticle endoplasmàtic és dispers.
És característic dels fongs que tinguin el nucli petit (2-3 μm de diàmetre), molt sovint haploide. Els nuclis diploides són de curta durada (ascs i basidis joves). La mitosi és endonuclear, i el cariolemma és persistent i sofreix una constricció equatorial, que condueix a la separació dels dos nuclis fills. Només els fongs que tenen zoòspores tenen centríols. En els altres, que són la majoria, hi ha només una placa centriolar o corpuscle polar del fus, d’on irradien els microtúbuls de l’aparell mitòtic. Els cromosomes no són fàcils de veure i no resten ordenats durant la metafase. En canvi, la meiosi segueix el procés típic dels eucariotes, i els cromosomes es fan més visibles. Els microtúbuls, però, semblen diferents als dels altres eucariotes, ja que són insensibles a l’acció bloquejadora de la colquicina.
El creixement dels fongs
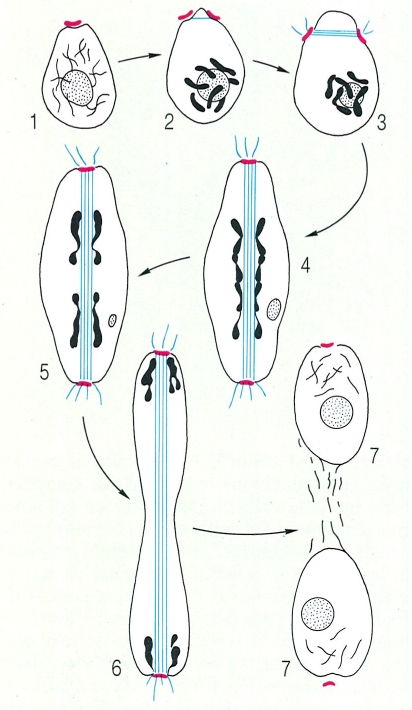
Esquema general de la divisió nuclear en els ascomicets, on es veu la formació dels microtúbuls (en blau) de l’aparell mitòtic a partir de les plaques centriolars (en vermell), la progressiva separació dels cromosomes (taques negres) i l’escanyament continuat del nucli fins a formar dos nuclis fills. 1 Inici de la condensació dels cromosomes, 2 divisió de la placa centriolar, 3 formació del fus mitòtic i migració de les plaques centriolars als àpexs del nucli, 4 inici de la separació dels cromosomes, 5 continuació de la separació dels cromosomes i estirament del nucli, 6 arribada dels cromosomes i inici de l’escanyament del nucli, 7 separació dels dos nuclis fills.
Biopunt, a partir de J. H. Burnett.
L’allargament de les hifes es limita exclusivament a la zona apical, de manera que tot l’increment del volum de les hifes és causat per llur augment de longitud. El creixement dels filaments dels fongs és, doncs, molt diferent al que presenten moltes algues filamentoses (de creixement intercalar), però es pot comparar amb el de les algues sifonades (caulerpals, vauquerials), dels pèls radicals o dels tubs pol·línics. El creixement es produeix per combinació de dos efectes principals: la pressió de turgència i la plasticitat i activitat de la part apical, que permeten de sintetitzar nova paret. La pressió de turgència també activa el moviment del citoplasma en direcció a l’àpex, que serà així abastit de nutrients procedents d’un gran volum d’hifes situades més enrere. Si col·loquem el miceli en un medi hipertònic (més concentrat que el citoplasma), les hifes deixen de créixer. Si les tallem massa arran de l’àpex (menys de 0,2-10 mm, segons l’espècie) moren, perquè no reben prou nutrients. La velocitat de creixement, en condicions òptimes, pot ésser molt notable, ja que sovint superen 1 mm/h i, en Neurospora, ateny gairebé 4 mm/h. La velocitat de creixement, en condicions estàndard (de temperatura, medi de cultiu), expressada en mil·límetres d’augment del radi per dia, constitueix un caràcter important a l’hora d’identificar molts fongs. La regió apical és essencial en la formació de paret nova, per a l’allargament de les hifes. A la zona d’extensió, un 80% del volum citoplasmàtic pot correspondre a vesícules. Constantment i en gran nombre (fins a 37 000 per minut) es formen, viatgen vers l’àpex i es fusionen amb el plasmalemma, que agafa un aspecte ondulat i crenulat mentre va augmentant de superfície. Aboquen, a l’espai que hi ha entre el plasmalemma i la paret, enzims i, possiblement, substàncies que han de passar a formar part de la nova paret. Uns primers enzims estoven la paret preexistent (trencant enllaços) i fan possible que aquesta es doni per efecte de la pressió de turgència. Després se sintetitzen nous components de paret per a reomplir els buits generats. Més endarrere, la paret només augmentarà de gruix i, a unes 75 μm de l’àpex, ja no hi haurà cap mena de creixement.
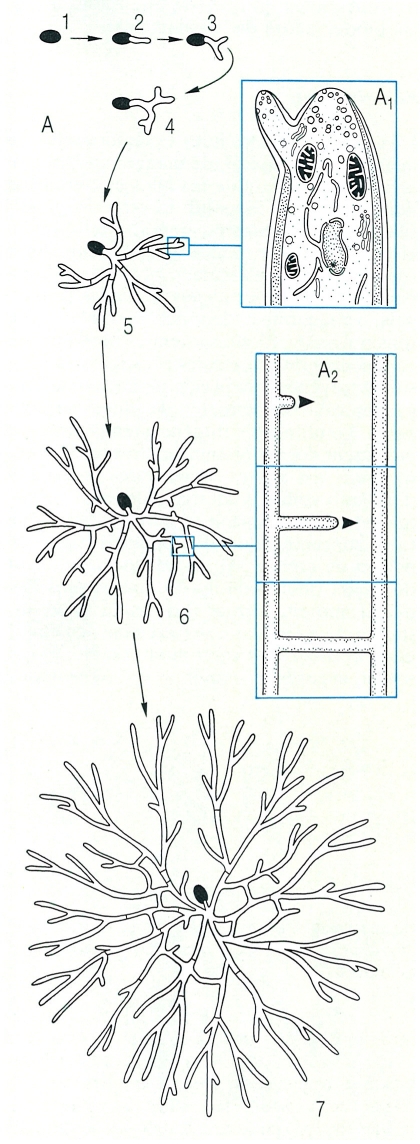
Procés de desenvolupament d’un miceli fúngic (A): germinació de l’espora (1-2) i detalls del procés de ramificació de les hifes (3-7): A1 aparició d’una ramificació en la zona subapical, mitjançant l’aparició d’un aparell apical nou; A2 anastomosi de dues hifes, que fa possible que es formi una xarxa miceliar (7).
Biopunt, a partir de fonts diverses.
Si hom observa la germinació d’una espora, es pot apreciar que primer s’infla i després genera un tubgerminatiu, gràcies a l’activitat d’un aparell apical. Aviat, a la zona subapical, es produeixen ramificacions, essencials per a explorar bé el substrat. El mecanisme és semblant al de l’estirament apical: es produeix un reblaniment local de la paret per acció d’enzims, i aquesta, ara elàstica, s’infla per efecte de la pressió de turgència, alhora que es forma un nou aparell apical, que assegurarà l’allargament de la nova branca. La branca va creixent, primer tendint a allunyar-se de les altres (ja que cerca la màxima concentració de nutrients o la màxima pressió d’oxigen). La densitat de la colònia dependrà, doncs, de la concentració de nutrients. Després, les hifes formaran una maranya i, per anastomosi, acabaran generant una complicada xarxa. Si el fong creix en un medi pla (per exemple, de cultiu en placa de Petri, creixement en un prat, etc.) el miceli adopta la forma d’un disc i, si té temps d’envellir, mor pel centre (per autòlisi) i queda formant un anell. Aquest anell es manifesta en la natura per l’ordenació de les fructificacions en forma de sardanes o erols de bolets, el diàmetre dels quals augmenta cada any. La persistència de molts micelis i, per tant, dels bolets que produeixen, permet de localitzar, encara que el miceli no es vegi, les rovelloneres o les llenegueres, i anar periòdicament a recol·lectar-hi els bolets (que són les fructificacions) que han produït, quan les condicions són favorables (temporada, pluviometria, temperatura). És una informació que sovint es guarda i es transmet de pares a fills, ja que molts micelis són de molta durada. Hi ha erols de bolets molt típics, com els dels cama-secs (Marasmiusoreades), que poden créixer fins a 20 cm per any, en radi, i atènyer dimensions d’1 a 10 m de diàmetre. S’han trobat, a les closes dels aiguamolls de l’Empordà, erols de Leucoagaricus de més de 30 m de diàmetre. Sovint, l’activitat del miceli provoca primer un creixement actiu de l’herba, en mineralitzar el nitrogen i el fòsfor del sòl. L’herba, més verda, destaca formant un cercle (un efecte indesitjat a les gespes dels jardins). Un cop passat el miceli, l’herba es pot veure recremada, per falta d’aigua o per acumulació de substàncies tòxiques excretades pel fong. Si el fong creix sobre un bocí de matèria orgànica submergit (per exemple, penjat) en un líquid, el miceli pren la forma d’una esfera de filaments.

La formació d’erols de bolets és un reflex de la manera que té de créixer el miceli del fong: el miceli s’estén d’una manera radial cobrint una àrea circular i comença a morir pel centre. La fotografia mostra un erol de cama-secs (Marasmius oreades) en un prat, i permet de veure les diferències en el creixement de l’herba degudes a l’activitat del fong. El dibuix presenta una secció de l’erol, on es veu la zona ocupada pel miceli (en blanc). El creixement de l’herba resta estimulat a dins i a fora de l’anella de fructificacions del bolet. Vora els carpòfors, l’herba es pot assecar.
Biopunt, a partir de fonts diverses; foto: August Rocabruna / SCM.
La ramificació del miceli permet als fongs d’explorar el substrat i interaccionar-hi a través d’una gran superfície d’intercanvi. Així, s’ha calculat que un fragment de miceli d’1 g de pes sec representa de 4 a 5 m2 de superfície. Els bacteris, freqüents competidors dels fongs, es divideixen activament i generen aviat molta superfície, però no tenen la capacitat de penetració dins el substrat amb què estan dotats els fongs, i s’han d’acontentar amb atacar les superfícies lliures. Però, com a contrapartida inevitable de l’organització filamentosa, les hifes són poc resistents als ambients extrems. La paret ha de tenir porus i per això perden fàcilment l’aigua si estan en contacte amb aire no saturat d’humitat. Moltes hifes aèries, espores i òrgans de resistència, acumulen un pigment negre, la melanina, per a protegir-se dels ambients massa secs o assolellats.
Igual que en els bacteris, després de sembrar un fong en un medi adient, observem diverses etapes en el seu creixement (dades referides al pes sec): una de latència, una de creixementexponencial, i una de progressiva atenuació, que condueix a una faseestacionària i a la mort.
Les diferenciacions de l’aparell vegetatiu dels fongs
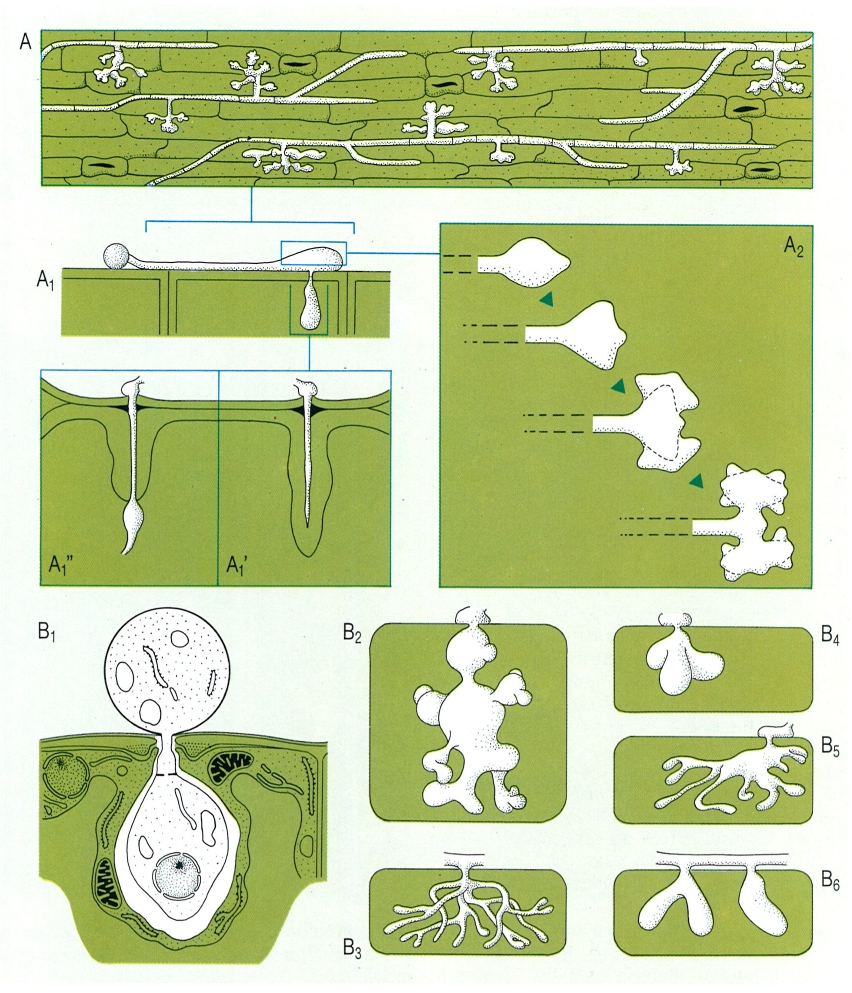
Apressoris i haustoris són estructures especialitzades que es poden formar en l’aparell vegetatiu dels fongs paràsits. El dibuix en recull alguns exemples. A Desenvolupament d’apressoris en les hifes d’un fong paràsit (Rhizoctonia solani) que penetra la superfície d’una tija de tomaquera; A1 detall de l’hifa en el moment de penetrar la cèl·lula a través de la paret, amb engruiximents (lignitúber) induïts per la cèl·lula a la seva paret per a defensar-se (A1’ amb èxit, A1" sense èxit); i A2 desenvolupament dels apressoris per ramificació apical repetida. B Formació d’haustoris en diversos fongs paràsits: B1 Erysiphe graminis, B2 Hemileia holstii, B3 Peronospora calotheca, B4 Hemileia vastatrix, B5 Puccinia purpurea, B6 Peronospora parasitica.
Biopunt, a partir de fonts diverses.
Molts fongs presenten en el seu cicle vital no solament una fase miceliar, sinó també una altra de llevat, que pot alternar amb la primera (dimorfisme). El pas a la fase de llevat es pot produir com a resposta a un o diversos factors, sovint de tipus ambiental (temperatura semblant a la corporal, en el cas de fongs paràsits de l’home; medi líquid; entrada en l’hemolimfa, en el cas de fongs paràsits d’insectes; ambient molt ric en sucres, etc.). En alguns casos, l’adaptació a la forma de llevat és permanent, i la forma filamentosa es deu haver perdut. Les cèl·lules dels llevats es multipliquen molt de pressa, en general per gemmació i, quan s’esgota l’oxigen del medi, poden viure per fermentació. En molts zigomicets, i en els basidiomicets gelatinosos, es dona sovint el dimorfisme.
La penetració de les hifes infectants o dels tubs de germinació produïts per les espores dels fongs paràsits en els organismes vius es fa sovint mitjançant apressoris, que són inflaments hifals, lobulats o no, fermament enganxats a la superfície de l’hoste. Permeten al fong de perforar l’obstacle de la paret cel·lular, continuar creixent a l’altre costat i propagar la infecció. La penetració s’aconsegueix combinant un efecte físic de pressió amb l’actuació d’enzims.
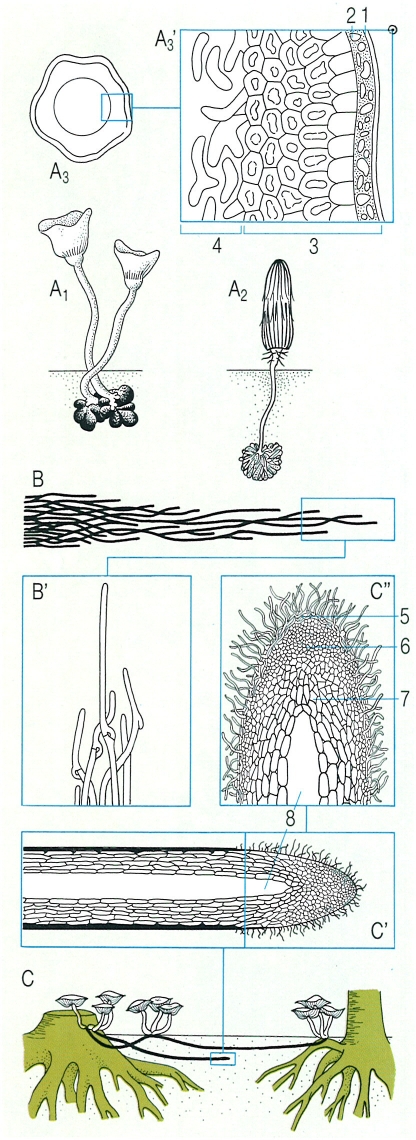
Altres diferenciacions de l’aparell vegetatiu dels fongs: esclerocis, rizomorfs i cordons miceliars. A Esclerocis de Sclerotinia tuberosa (A1) i de Coprinus stercorarius (A2) produint carpòfors, i detall d’un escleroci de Sclerotium rolfsii (A3), d’aproximadament 2 o 3 mm de diàmetre, amb les seves parts principals en secció (A3’). B Formació de cordons miceliars en Serpula lacrymans, formats per hifes amb branques laterals que creixen paral·lelament a la direcció de l’hifa principal (B’). C Formació de rizomorfs subterranis en Armillariella mellea, que permeten al fong d’arribar fins als troncs d’arbres vius, partint d’una soca morta; i detalls de l’àpex del rizomorf (C’, C"), amb la seva cavitat central i la seva estructura apical, que imita la caliptra d’una arrel. Hom hi ha indicat: 1 cutícula, 2 epidermis, 3 còrtex, 4 medul·la, 5 còrtex lax i gelificat, 6 pseudoparènquima consistent, 7 pseudoparènquina de cèl·lules grosses i laxes, 8 cavitat central
Biopunt, a partir de fonts diverses.
Alguns fongs saben formar estructures pluricel·lulars de resistència, sovint hipogees, els esclerocis, que poden ésser simples agregats d’hifes brunes (amb melanines), com en Rhizoctonia solani, o poden tenir una complexa organització i especialització, i mides de fins a 5 mm o més. Contenen substàncies de reserva, com mannitol, trehalosa, glicogen i lípids, i, proporcionalment, poca aigua. Quan es formen, recullen la majoria dels nutrients del miceli. Són, doncs, una forma de supervivència i de reserva de nutrients. Són resistents als períodes secs i a les temperatures extremes, i alguns són molt longeus (4-14 anys). Els de Claviceps no poden reprendre l’activitat si no han travessat abans un període d’hivernació.
En general, totes les hifes vegetatives tenen una gran capacitat de transport de nutrients, mitjançant el flux citoplasmàtic. Però alguns fongs formen estructures especialitzades, de gruix visible (de 100 μm a 5 mm), que sobretot els faciliten la colonització ràpida de nous substrats, situats a una certa distància de la font primària d’aliments. Els cordons miceliars són les estructures més freqüents, relativament simples; es formen a partir d’una o més hifes principals o conductores, que es ramifiquen i alhora creixen activament, però de tal manera que les noves branques, en comptes de divergir, creixen paral·leles, molt a la vora, s’anastomitzen i arriben a diferenciar-se en hifes connectives, que les mantenen lligades entorn de les hifes centrals, que es van diferenciant fins a tenir un actiu paper conductor. Són les hifestràquea. Són molt coneguts els cordons miceliars que forma Serpula lacrymans, que ataca la fusta de les cases. Poden arribar a travessar parets toves i atènyer així fusta encara no colonitzada. Però són molts el’s bolets que tenen cordons miceliars a la base del peu (Agaricus, Collybia, Clitocybe vermicularis, Phallus, Clathrus). D’altra banda, els rizomorfs s’estructuren a partir d’un àpex format per hifes ja agregades, que recorda el meristema apical de les rels. Fins i tot hi ha hifes que creixen cap endavant, es gelifiquen i formen una mena de piloriza protectora. El gruix és sempre considerable, i és degut a l’agregació de milers d’hifes paral·leles que han crescut de 5 a 6 cops més de pressa que les vegetatives no especialitzades. Hi ha un còrtex protector, sovint melaninitzat, i una medul·la que, en madurar, origina una zona central buida, tubular, que facilita el transport d’oxigen. El cas més conegut de rizomorfs és el de l’alzinoi (Armilariella mellea i espècies afins): es poden estendre pel sòl o sota l’escorça, fins a uns quants metres de distància, de manera que augmenten la capacitat invasora d’aquest bolet tan perjudicial dels arbres forestals i fins i tot dels fruiters; en l’obscuritat, els rizomorfs de l’alzinoi poden desprendre una lluminositat violàcia.
Les clamidòspores són diferenciacions de l’aparell vegetatiu, que reacciona davant condicions desfavorables. Algunes cèl·lules vegetatives, o una porció de miceli cenocítica delimitada per noves parets, acumulen substàncies de reserva i formen una nova paret, gruixuda i sovint fosca, per dins de la preexistent. Molts grups diferents formen clamidòspores, que tenen un paper de resistència, funció equivalent al dels cists de molts altres organismes. Són molt conegudes les clamidòspores de les endogonals. També els zigomicets en fan sovint. Alguns fongs paràsits, com Sepedonium o Mycogone les fan en gran nombre, de color viu i molt ornamentades. Tot el barret de les Asterophora es descompon en una massa pulverulenta de clamidòspores.
La reproducció dels fongs
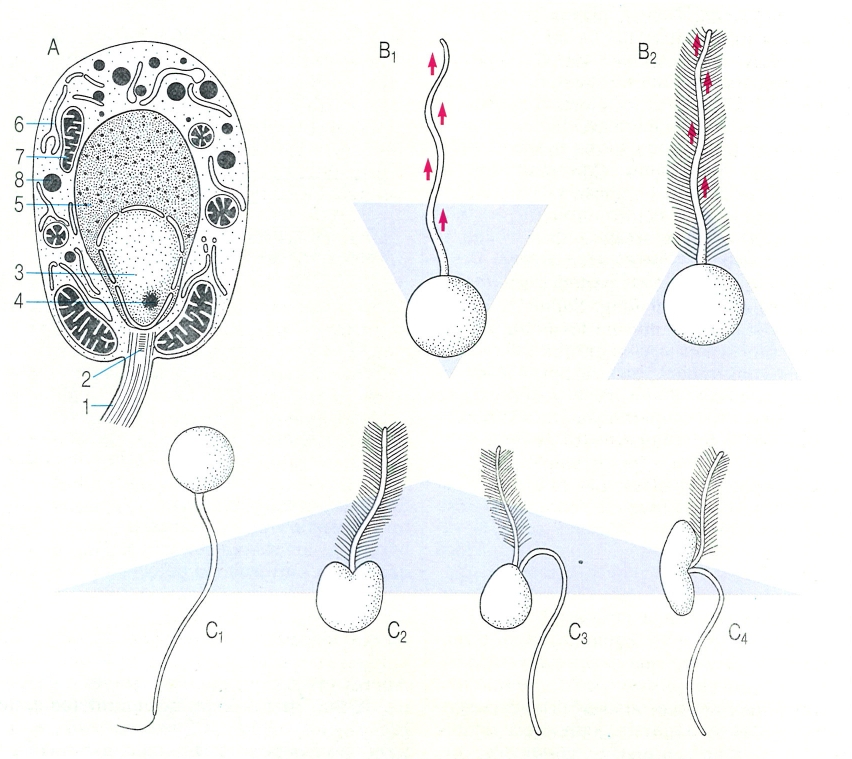
Estructura, variabilitat i moviment de les zoòspores dels fongs. A Estructura d’una zoòspora (en aquest cas, de blastocladial). B Sentit del moviment (representat per les fletxes morades en el dibuix), segons el tipus de dotació flagel·lar que presenten les zoòspores. Les sagetes vermelles indiquen el sentit de Tona sinusoidal en el flagel. B1 flagel llis, B2 flagel barbulat, amb mastigonemes. Noteu com el sentit del moviment s’inverteix quan hi ha mastigonemes (retronemes). C Principals tipus de zoòspores: C1 zoòspores amb flagel llis, posterior; C2 zoòspora amb flagel barbulat, anterior; C3, C4 zoòspora amb flagel barbulat anterior i llis posterior. Hom hi ha indicat: 1 flagel, 2 cinetosoma (blefaroplast), 3 nucli, 4 nuclèol, 5 casquet nuclear (regió ribosòmica), 6 reticle endoplasmàtic, 7 mitocondri, 8 corpuscle lipídic.
Biopunt, a partir de fonts diverses.
Alguns fongs primitius transformen la totalitat de llur aparell vegetatiu en diferenciació reproductora. Són els fongs holocàrpics. És molt més freqüent, però, que només una part de l’aparell vegetatiu intervingui en la reproducció, com és propi dels fongs eucàrpics. Hom parlarà primer de la reproducció o multiplicació asexual i després de la sexual.
La reproducció asexual
Qualsevol fragment de miceli o de fructificació, fresc o sec, pot servir com a propàgul dels fongs. Aquest fet s’aprofita per a repicar (replantar en medi de cultiu nou) els fongs cultivats en placa de Petri, o per a obtenir cultius a partir de fragments de bolet. En estat natural, els fragments secs de miceli també deuen servir sovint com a propàgul.
Les clamidòspores afegeixen també una funció de propàguls a la de formes de resistència, però és més freqüent que els fongs es propaguin per espores, que poden ésser planòspores o zoòspores (en els quitridiomicets) o aplanòspores (en la resta), formades independentment de la reproducció sexual (mitòspores) o bé com a conseqüència de la reproducció sexual (meiòspores).
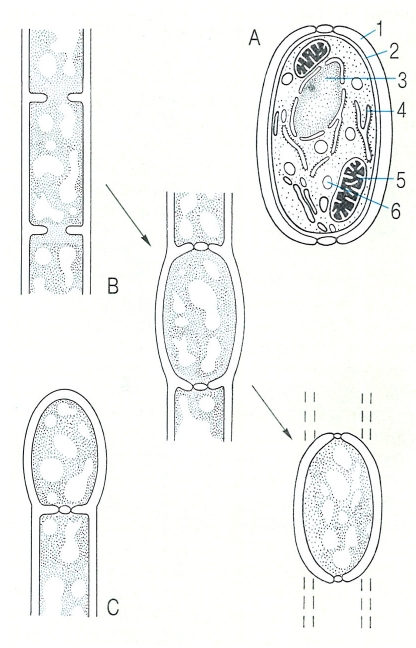
Estructura d’una clamidòspora (A) i formació intercalar (B) i terminal (C) de clamidòspores en les hifes dels fongs. En el procés de formació d’una clamidòspora s’obturen els porus, la paret s’engruixeix i la resta de l’hifa sofreix una autolisi. Hom ha indicat en el dibuix: 1 paret primària, 2 paret secundària, 3 nucli, 4 reticle endoplasmàtic, 5 mitocondri, 6 cos lipídic.
Biopunt, a partir de fonts diverses.
Les espores asexuals són de dues menes, les esporangiòspores i els conidis. Les esporangiòspores es formen a l’interior d’una cèl·lula, l’esporangi (o esporocist), després de la divisió del nucli per mitosi. En l’exemple més característic, els zigomicets, veiem aparèixer en els esporangis joves un gran nombre de vesícules de segmentació, que es van ordenant entorn dels diversos nuclis producte de la mitosi, potser amb la col·laboració de microtúbuls. A continuació hom les veurà fusionar-se per formar progressivament una xarxa de sacs que delimita porcions de citoplasma. Els forats de la xarxa es van tancant i acaben formant membranes completes. Després, es formarà la paret esporal, que pot ésser força gruixuda i complexa. El nombre d’espores és usualment elevat, i són alliberades per ruptura de la paret de l’esporangi. Hi ha esporangis que contenen poques espores: són els esporangíols. Quan en tenen una de sola, conserven per fora la paret de l’esporangi i per dins la de l’espora. En els zigomicets, les espores no tenen flagels, però en la majoria dels quitridiomicets són flagel·lades, nedadores (planòspores) i no se’ls veu paret.
Les esporangiòspores són endòspores (es formen dins d’una cèl·lula). En canvi, les conidiòspores o conidis (del grec conis, pols, per les masses polsoses que formen) són exòspores, i es formen també per mitosi, però mai a l’interior d’una cèl·lula. Segons el procés de formació (conidiogènesi), distingim els conidis tàl·lics (anomenats també tal·loconidis o artroconidis), originats per septació i posterior segmentació d’una hifa preexistent, i els blàstics o blastoconidis, originats per gemmació, d’una manera que recorda la dels llevats, però sovint de forma repetida, formant cadenes i separats per septes. Com que les conidiòspores tenen una importància especial en els deuteromicets, a la introducció dedicada a aquest grup hom les tracta amb més detall. Les conidiòspores s’acostumen a produir en gran quantitat durant els períodes favorables per al creixement del fong. Serveixen per a dispersar-lo i augmentar-ne les poblacions a curt termini. El cicle vital en què intervenen és ràpid i no implica, en principi, cap mena de recombinació d’informació genètica (tret del fenomen de la parasexualitat). El cicle asexual (dit també imperfecte) és important en uns grups de fongs (per exemple, als ascomicets) i poc en d’altres. Les fases que presenten reproducció asexual s’anomenen anamòrfiques (anamorf).
La reproducció sexual
La reproducció sexual es realitza amb la intervenció de cèl·lules haploides de diferent origen, els gàmetes, que es produeixen en òrgans sexuals, els gametangis (o gametocists), i es fusionen en dues etapes: primer el citoplasma (plasmogàmia) i després els nuclis (cariogàmia). S’origina així un zigot, diploide, que pot actuar o no com a òrgan de resistència. Tard o d’hora, el nucli es divideix, gairebé sempre per meiosi, i origina un esporangi, amb espores que, si s’han originat per meiosi, com passa normalment, s’anomenen meiòspores. El cicle on intervé la reproducció sexual és aquell que, de manera primària, permet la recombinació genètica. Per això s’anomena cicle perfecte, i en la fase que presenta la reproducció sexual, fase perfecta o teleomòrfica (teleomorf).
En el cas dels fongs més evolucionats (ascomicets, basidiomicets), assistim a un fenomen biològic únic: un cop realitzada la plasmogàmia, els dos nuclis triguen molt a unir-se i es van dividint, tots dos alhora, de manera que cada compartiment cel·lular conté una còpia de cada nucli. Cada una d’aquestes unitats s’anomena dicàrion, i forma part d’hifesdicariòtiques. De fet, la divisió i multiplicació dels nuclis sexuals per a formar hifes dicariòtiques representa una forma enginyosa de multiplicar els zigots, que es completa quan per fi es produeix la cariogàmia. Les hifes dels ascomicets destinades a produir ascs (hifesascògenes) són dicariòtiques i, encara més, en són gairebé totes les hifes dels basidiomicets. Un plat de rovellons o de llenegues és un plat d’hifes dicariòtiques.
Hi ha fongs que es poden autofecundar: són els hermafrodites o monoics. La majoria necessiten dos tal·lus diferents, compatibles, per a fer possible la reproducció sexual. Si presenten òrgans masculins i femenins diferenciats, en tal·lus separats, s’anomenen dioics. Però sovint no és possible distingir quin tal·lus és el masculí i quin el femení. Ara bé, no per això deixen d’actuar sistemes genètics, que s’explicaran més endavant, per excloure la consanguinitat de la reproducció sexual.
En els fongs més primitius (quitridiomicets) sovint es troben gametangis que produeixen gàmetes flagel·lats (planogàmetes), que són alliberats a l’aigua, on poden copular i donar origen a un zigot nedador. Si són indiferenciables, parlem d’isogàmetes. Però pot passar que els masculins i els femenins siguin morfològicament diferents. Aquests heterogàmetes poden ésser tots dos mòbils (anisogàmetes) o un d’ells pot ésser immòbil (oogàmetes). En aquest darrer cas, el gametangi masculí (el que dona gàmetes nedadors) s’anomena anteridi, i el femení, oogoni. Però, ja en el mateix grup dels quitridiomicets, trobem una tendència a la simplificació de la reproducció sexual. Primer deixen d’independitzar-se els gàmetes, i són els gametangis els que intercanvien els nuclis gamètics a través d’un porus o d’un curt tub de fecundació (contacte gametangial), o bé es fusionen del tot (copulació gametangial o gametangiogàmia). El contacte gametangial es dona en bastants ascomicets, i la gametangiogàmia és característica dels zigomicets. Molt sovint s’han pogut descobrir complexos mecanismes de regulació, en els quals diverses hormones van apareixent per a produir la diferenciació dels gametangis només quan hi ha presents micelis compatibles.
En alguns ascomicets trobem l’espermatització, en la qual els oogonis, que tenen una prolongació filamentosa (la tricògina) són fecundats per gàmetes no nedadors masculins, els espermacis. Però la culminació del procés de reducció és la somatogàmia, típica dels basidiomicets, en la qual cèl·lules vegetatives indiferenciades es fusionen entre elles (o amb una espora) per a donar origen a un micelidicariòtic, després d’una plasmogàmia. En la introducció a cada gran grup de fongs veurem millor diversos exemples d’aquests tipus de reproducció. La ràpida tendència a abandonar els gàmetes flagel·lats en l’evolució dels fongs veritables es pot interpretar com el resultat d’una progressiva adaptació a la vida fora de l’aigua.
Els carpòfors
És sobretot en els ascomicets i en els basidiomicets on es verifica que les cèl·lules que produeixen espores després de la reproducció sexual (els ascs i els basidis), sovint les ajuden a complir millor la seva comesa hifes estèrils que s’ajunten i formen falsos teixits, anomenats plectènquimes (prosènquimes, quan encara s’hi reconeixen les hifes, i pseudoparènquimes, en el cas contrari).
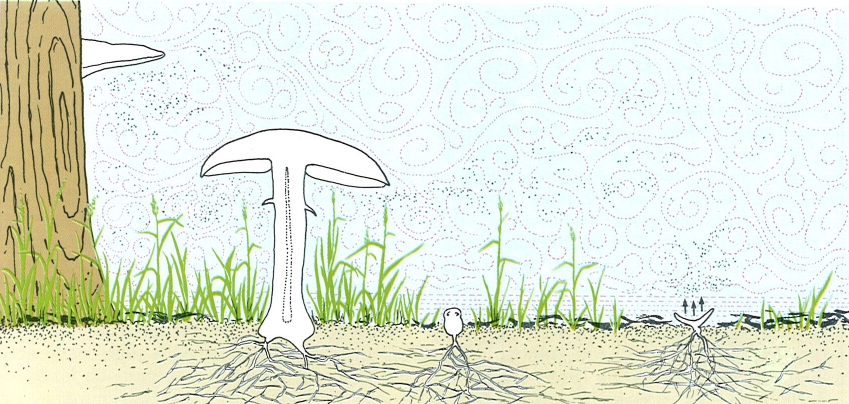
Els carpòfors dels fongs estan adaptats a deixar anar les espores (punts negres) en la capa d’aire amb flux turbulent, que se les emporta lluny, ja que en la capa inferior, de flux laminar, caurien aviat a terra. Els dos carpòfors de basidiomicets del dibuix (esquerra i centre) tenen l’himeni fèrtil en la zona turbulenta. El d’ascomicet, una cassoleta (dreta), és en la zona laminar, però envia les espores a la zona turbulenta.
Biopunt, a partir de fonts diverses.
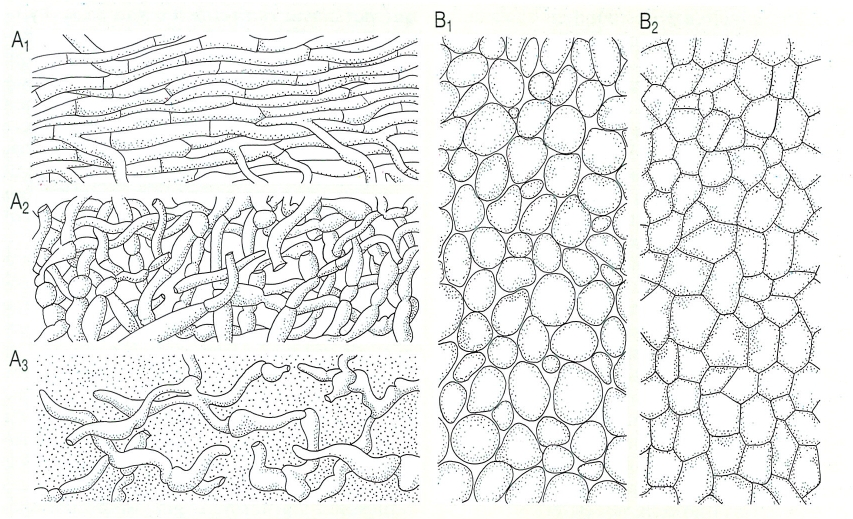
Tipus de plectènquimes o falsos teixits i tipus de carpòfors en els ascomicets i els basidiomicets. Hom ha representat, d’entre els plectènquimes, els prosènquimes (A), on encara és possible de distingir les hifes, entrellaçades o soldades per punts, però més o menys individualitzades, i els pseudoparènquimes (B), amb cèl·lules més curtes i pròximes, que no permeten de distingir les hifes. A1 Prosènquima en el qual les hifes resten diferenciades i més o menys paral·leles; A2 prosènquima en el qual les hifes estan entrellaçades; A3 prosènquima en el qual les hifes estan disperses en un gel. B1 Pseudoparènquima d’hifes arrodonides, amb espais intercel·lulars; B2 pseudoparènquima de cèl·lules molt juntes, sense espais intercel·lulars. A la meitat inferior del dibuix hom ha representat diversos exemples de carpòfors d’ascomicets, o ascocarps (1-11), i de basidiomicets, o basidiocarps (12-22): 1-5 discomicets (1 Peziza, 2 Helvella, 3 Mitrula, 4 Morchella, 5 Sarcoscypha); 6-8 pirenomicets (6 Sordaria, 7 Daldinia, 8 escleroci a la base, amb estromes, de Claviceps, i 8’ secció d’un estroma, amb peritecis); 9-11 formes hipogees (9 Genea, 10 Tuber, 11Balsamia); 12-13 carpòfors amb porus (12) Boletus i 13 Phellinus); 14 carpòfor de làmines (Agaricus); 15 carpòfor amb agulles Hydnum); 16 carpòfor efuso-reflex, amb l’himeni llis (Stereum); 17 carpòfor en forma de niuet (Cyathus), amb el detall d’un peridíol; 18 carpòfor coral·loide de peu de rata (Ramaria); 19 carpòfor gasteroide amb peridi doble d’estrelleta (Geastrum); 20 carpòfor gasteroide de pet de llop (Lycoperdon); 21 carpòfor gasteroide amb receptacle expansionable (Phallus); 22 carpòfor hipogeu de fals fetjó (Hymenogaster); 23Sphaerobolus, que projecta un peridíol esfèric, portador de les basidiòspores.
Biopunt, a partir de fonts diverses.
Els òrgans portadors d’acs i de basidis s’anomenen carpòfors (del grec carpós, fruit) i, respectivament, ascocarps (o ascomes) i basidiocarps. De fet, la majoria de les produccions naturals que popularment s’anomenen bolets són basidiocarps (rovellons, llenegues, surenys, sabateres, bolets de soca, rossinyols, trompetes, peus de rata) o ascocarps (múrgules, orelles de gat, cassoletes). Tota la monotonia que hom trobava a l’aparell vegetatiu es transforma ara en varietat de formes, colors i olors, quan es tracta de formar carpòfors. Aquests, en els fongs superiors, són els que presenten els caràcters que permeten identificar les diferents espècies. Tot el carpòfor està destinat a aconseguir que les cèl·lules esporògenes contingudes a la fina capa fèrtil anomenada himeni puguin dispersar les espores amb la màxima eficàcia.
La dispersió de les espores
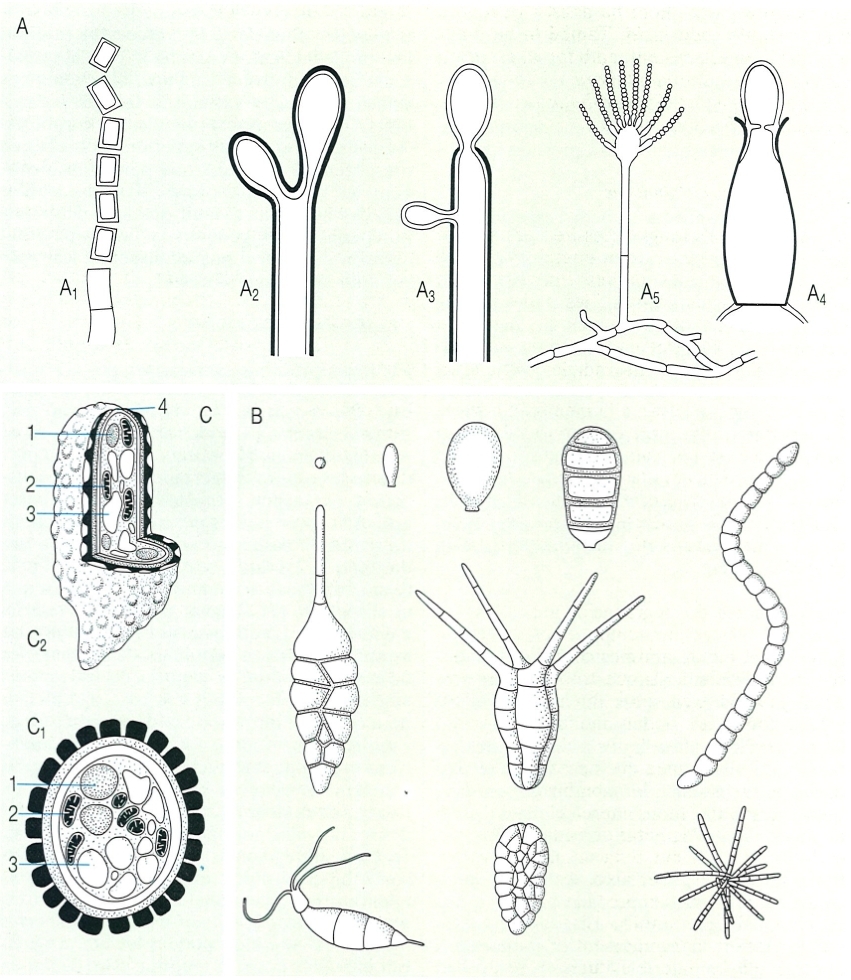
Aplanòspores dels fongs, adaptades a la dispersió per l’aire. A Tipus de desenvolupament dels conidis: A1 tàl·lic, A2 holoblàstic, A3 enteroblàstic trètic, A4 enteroblàstic fialídic, A5 conidiòfor d’Aspergillus, amb fiàlides productores de cadenes de conidis. B Diversitat de mides i formes de conidis. C Estructura de les espores: C1 estructura d’una ascòspora de paret robusta, biestratificada i ornamentada, C2 estructura d’una basidiòspora, de superfície verrucosa. Hom ha indicat en el dibuix: 1 nucli, 2 mitocondri, 3 vacúol, 4 porus germinatiu.
Biopunt, a partir de fonts diverses.
Els fongs primitius es dispersen per via aquàtica, mitjançant espores nedadores (planòspores). Però, com ja s’ha vist, l’adaptació dels fongs veritables a la vida fora de l’aigua es va produir ja en els zigomicets i conduí a la pèrdua de les espores flagel·lades i a la seva substitució per espores (aplanòspores) de paret gruixuda i prou petites per a poder restar molt de temps en suspensió en l’aire (cauen a raó de 0,003 a 2,8 cm/s, segons llur radi). Però, secundàriament, tornem a trobar espores que es dispersen per l’aigua, però sense flagels, passivament. La dispersió per l’aire permet de sembrar espores a centenars de quilòmetres de distància, si bé la majoria de les espores emeses cauen força més a la vora. D’una manera o altra, l’eficàcia de la dispersió és gran, i molts fongs viuen a tot arreu si les condicions ecològiques són adients, o en àrees molt extenses. Sempre hi ha espores viatjant en l’aire, i en podem trobar unes 10 000 en 1 m3 d’aire, un valor normal, i fins a unes 200 000, en casos excepcionals. Ho saben bé les persones hipersensibles, que pateixen afeccions respiratòries quan inhalen determinades espores de l’aire. Si bé són moltes les espècies sembrades, són molt poques les que originaran individus nous; la majoria, o no cauen en un lloc adequat, o el troben ja ocupat.
Però, ¿com arriben les espores als corrents turbulents de l’aire? Els conidis, per exemple, sovint són hidròfugs (repel·len l’aigua i no es deixen mullar). Són les gotes de la pluja o el vent fort els que mobilitzen les espores. Els esporangis acostumen a ésser portats per llargs esporangiòfors, i quan se’n trenca la paret, el vent se n’endú les espores. En el cas dels ascomicets, els ascs acostumen a funcionar com a petits canons, i disparen les espores a una certa distància (d’1 mm a 40 cm), en direcció a la llum. Els basidiomicets comuniquen també a les espores un cert impuls, que els permet d’arribar a distàncies de 0,1 a 0,2 mm. Com que l’himeni es troba a una certa alçada, sovint gràcies al peu del bolet, les espores poden caure en l’aire, animat per un moviment turbulent que se les emporta. Els pets de llop tenen les espores tancades en una cavitat elàstica, i qualsevol impacte, sobretot el de les gotes de pluja, fa sortir manxades d’aire carregat d’espores; en podem fer la prova deixant caure gotes d’aigua sobre pets de llop ben madurs. Són també les gotes d’aigua les que, en caure dins els niuets (nidularials) esclaten en esquitxos i expulsen les unitats portadores d’espores (els peridíols) a una bona distància. Però hi ha també fongs que utilitzen els animals com a agents dispersadors. Hom recordarà aquí les fal·lals, que, amb la pudor de carn en descomposició que desprenen, atrauen dípters, que s’embruten d’espores enganxoses i se les emporten; i les tòfones i altres fongs hipogeus (amagats sota terra), que es fan descobrir per nerbívors i rosegadors gràcies al perfum que desprenen, fins que són desenterrats i menjats; les espores sortiran més tard, amb els excrements. En aquests dos casos, hom ha assistit gairebé a una veritable coevolució. També hi ha fongs que disparen activament, amb força, esporangis (com ara, Entomophthora i Pilobolus) o peridíols (com ara Sphaerobolus), gràcies a mecanismes força complexos.
La nutrició i el metabolisme dels fongs

Cultiu d’un fong en placa de Petri. El fong ha estat sembrat en el marge del medi de cultiu i ha format una colònia de creixement radial, esporulada a la part més vella (noteu el canvi de color).
Rafael Campillo / Zardoya Press.
Ja sabem que els fongs necessiten per a viure compostos orgànics rics en energia. Quan es tracta de molècules petites (monosacàrids, aminoàcids, àcids grassos), les poden absorbir directament. Però és més freqüent que trobin com a substrat molècules massa grosses, que han d’ésser degradades mitjançant enzims secretats (probablement continguts en vesícules que s’uneixen a la membrana cel·lular, i surten a fora a través de porus de la paret), o que resten units a l’exterior de la paret. La producció d’aquests enzims és estimulada per la presència dels substrats i frenada per la dels productes finals. Cap de tots aquests mecanismes no funcionaria si no hi hagués prou aigua.
Els productes de degradació, abans d’ésser absorbits, resten un temps a disposició de fongs i d’altres organismes competidors. Les condicions ambientals que troben els enzims en sortir influeixen sobre llur activitat. Però alguns fongs les poden modificar: Serpula lacrymans, que ataca la fusta de les cases, es fabrica part de l’aigua que necessita, per oxidació del substrat, i la subministra als llocs massa secs; altres fongs alteren el pH del medi on viuen, etc. L’activitat de cada hifa provoca la formació d’una zona immediata empobrida en aliments. Per això, les hifes han de continuar creixent sempre, i la part vella pot acabar morint per autòlisi. Els fongs han d’estar explorant contínuament el substrat, i avancen a salts, ja que s’aturen quan arriben a una zona particularment rica en aliments.
La font bàsica d’aliment per als fongs són els glúcids d’origen vegetal. En degradar-los, col·laboren a netejar la biosfera de residus vegetals, que es formen en grans quantitats i aviat s’acumularien. Tots els fongs saben utilitzar la glucosa, i gairebé tots la maltosa, la sacarosa i el midó. Alguns poden utilitzar greixos i proteïnes, però a vegades necessiten una aportació suplementària de sucres per a poder-ho fer. Del 40 al 50% de la paret de les cèl·lules de les plantes és cel·lulosa. També conté una bona quantitat d’hemicel·luloses, més fàcils de descompondre. Però la cel·lulosa és el principal aliment dels fongs, que fabriquen tres enzims cel·lulolítics. L’endogluconasa, separa de la cel·lulosa residus de cel·lobiosa (un disacàrid), que seran atacats a continuació per la glucosidasa, que els deixarà reduïts a molècules de glucosa. Però la cel·lulosa té parts on el polímer es disposa fortament empaquetat (cel·lulosa cristal·lina). Cal llavors un tercer enzim, l’exogluconasa, que ataca les molècules pels extrems i en va separant també cel·lobiosa. El conjunt d’aquests enzims constitueix el complex de la cel·lulosa. S’observa doncs com, per aprofitar un sol substrat, es necessiten tres enzims, que són activats per la presència de la cel·lulosa i frenats per l’acumulació de glucosa. Bé que molt més rars, hi ha fongs que saben degradar substrats «difícils», com ara lignina, hidrocarburs, plàstics (degradant primer els plastificants), pesticides, etc. El seu valor en la neteja dels ecosistemes pot ésser molt important.
Els fongs es poden cultivar en medis naturals, gelificats amb agar-agar, als quals hom afegeix extrets vegetals (patata, extret de malta, farina de civada). Quan cal precisar millor els requeriments dels fongs, hom empra medis artificials, que contenen diversos nutrients (nitrogen, fòsfor, potassi, sofre, etc.), i una font de matèria orgànica, que pot ésser glucosa o sacarosa. Alguns fongs necessiten, a més, vitamines, com tiamina o biotina, o certs aminoàcids. El medi de cultiu es disposa, en capes primes, en platets de vidre o plàstic amb tapadora (plaques de Petri) o en tubs inclinats. Les tècniques d’isolament, de sembra i de ressembra (repicatge) són molt semblants a les que utilitzen els microbiòlegs. Molts fongs superiors es poden cultivar en compost (preparat de matèria vegetal o fems, predigerit per bacteris) esterilitzat.
Tots els fongs poden utilitzar aminoàcids, la majoria NH4+ i alguns NO3−, com a font de nitrogen. La majoria poden utilitzar els ions sulfat com a font de sofre. En condicions especials, les necessitats poden variar. Així, el llevat del vi, quan fermenta, necessita diverses vitamines, que en condicions aeròbiques ell mateix sap sintetitzar. L’absorció de substàncies és regulada per diversos mecanismes que, en bona part, depenen de les molècules de permeasa de la membrana cel·lular, que presenten una certa especificitat envers determinats sucres o aminoàcids. D’altres presenten certes preferències. Així, un fong pot absorbir d’una manera preferent glucosa, i només quan aquesta s’esgota en el medi, absorbeix altres sucres.
Els sucres són metabolitzats seguint sobretot dues de les vies de la glucòlisi conegudes, la d’Embden-Meyerhof i la de l’hexosa-monofosfat. Si hi ha suficient oxigen, l’àcid pirúvic produït per aquestes vies s’oxida en el cicle de Krebs, i dona CO2 i aigua. Però molts fongs filamentosos i llevats poden créixer anaeròbicament quan falta oxigen, fermentar els sucres, i produir aleshores productes orgànics com alcohol etílic, àcid làctic, acetaldehid, etc. Mentre puguin ésser absorbits per les cèl·lules, molts altres compostos orgànics són susceptibles d’ésser metabolitzats. Aquest és el cas dels aminoàcids. Molts compostos intermediaris de les vies que metabolitzen els sucres s’utilitzen com a base per a la biosíntesi de compostos importants, com àcids grassos o aminoàcids, o metabòlits secundaris.
L’extracció d’aquests intermediaris és compensada per mitjà de vies alternatives que continuen subministrant energia. Una de les més importants és la de la carboxilació (combinació de piruvat amb CO2) en la qual intervé la biotina com a coenzim. Un altre cicle alternatiu és el del glioxilat, important quan els fongs disposen d’àcid acètic com a font d’àcid carbònic. Alguns fongs tenen una elevada capacitat per a secretar àcids orgànics al medi de cultiu. Aquestes propietats són aprofitades per l’home per a la síntesi d’àcid cítric (en la qual un 95% de la glucosa subministrada és transformada per Aspergillus nidulans en aquest àcid), i d’àcid fumàric (per Rhizopus nigricans). Aquesta acumulació va lligada en general a condicions de creixement anormals, per exemple, a la falta de nitrogen.
Substàncies de reserva
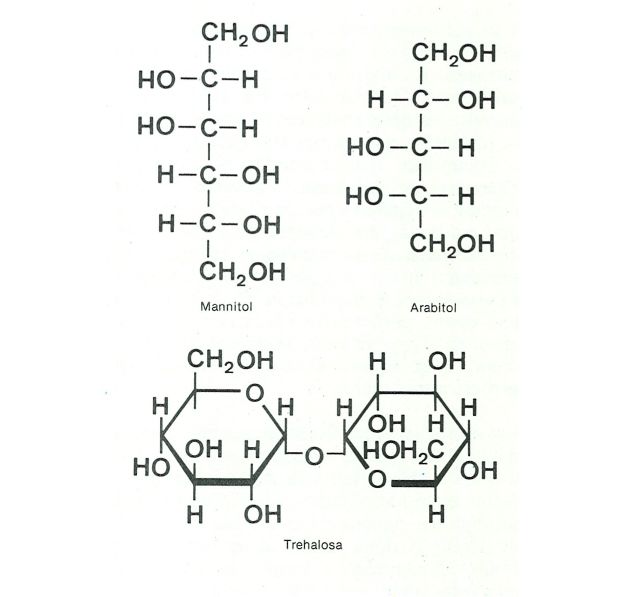
Estructura de la molècula de tres de les principals substàncies de reserva en els fongs.
Biopunt, a partir de fonts diverses.
S’assemblen molt més a les dels animals que a les de les plantes: lípids (ormant gotetes anomenades sovint gútules), o glicogen (formant rosetes només observables al microscopi electrònic). Els glúcids, que són fàcilment interconvertibles, s’acumulen i es transporten des de les hifes en forma de trehalosa (un disacàrid) i de sucres-alcohol, com mannitol i arabitol. Els zigomicets tenen principalment trehalosa. Els fongs paràsits, sobretot els de les plantes vives, es beneficien de la ràpida transformació dels sucres que capten de la planta (glucosa, sacarosa, fructosa) en els sucres fúngics que hom acaba d’esmentar, que no poden ésser metabolitzats per les plantes. S’estableix així un flux sense retorn, que va de la planta parasitada al fong.
Metabolisme secundari
Es produeix per la desviació de les vies que condueixen al creixement normal, que produeix substàncies, els metabòlits secundaris, amb més activitat quan el creixement normal del fong es veu dificultat per alguna raó. Són coneguts químicament més de 100 d’aquests metabòlits fúngics (si deixem de banda els dels líquens); alguns tenen un gran interès aplicat i, per tant, comercial (antibiòtics, hormones vegetals com la gibberel·lina, saboritzants, etc.), o són molt tòxics i poden constituir un problema sanitari (micotoxines, alcaloides, tòxics dels bolets mortals i metzinosos). Els metabòlits secundaris són molt específics i no tenen una funció clara en la vida dels organismes productors; són sintetitzats per organismes que han deixat de créixer (es troben en el principi de la fase estacionària). Això s’ha de tenir en compte quan hom els vol produir industrialment.
Pot ésser que els metabòlits secundaris no tinguin una importància directa per al fong, però que es produeixin en vies metabòliques que sí que en tenen. Per això s’acumulen quan, a causa de la reducció de l’activitat cel·lular, aquests processos metabòlics ja no tenen interès.
Els factors ambientals que afecten els fongs
La influència de diversos factors ambientals (temperatura, pH, aigua, llum, oxigenació) s’ha estudiat sobretot al laboratori, observant les variacions en el creixement de cultius purs amb diversos graus d’intensitat de cada factor. Probablement, a la natura, amb la competència dels altres organismes, les coses no van de la mateixa manera.
La majoria dels fongs creixen a temperatures que oscil·len entre 10 i 40°C, essent òptima la que va de 25 a 35°C, però mantenen un bon creixement a temperatura ambient (si no gela). Pel que fa a la temperatura, doncs, són mesòfils. Els que penetren en els animals de sang calenta, que no són gaires, neixen a 37°C. Alguns fongs són termòfils i poden créixer entre 20 i 50°C, essent la temperatura òptima vers 40°C. Poden viure en les fulles en descomposició, nius d’ocells o sòls escalfats pels raigs del sol. Els podem distingir dels que només són termotolerants, la temperatura òptima dels quals se situa a 40°C, tot i que poden créixer encara entre 12 i 20°C; Aspergillus fumigatus n’és un exemple, i es pot instal·lar als pulmons, on causa aspergil·losi. També hi ha alguns fongs, els psicròfils (i els psicrotolerants) que viuen a temperatures baixes, i no en toleren de superiors a 20°C. Poden viure en llocs freds, o, a l’hivern, fins i tot sota la neu. Alguns constitueixen un problema perquè poden colonitzar els aliments conservats als frigorífics. Pel que sembla, els fongs termòfils tenen els lípids de la membrana més saturats (i, per tant, la temperatura de fusió més alta) i els enzims més estables.
Els boletaires saben que alguns bolets prefereixen sòls calcaris (Amanita ovoidea) i d’altres viuen millor en sòls silicis (Amanita mairei, A. caesarea). Això pot dependre en part de la reacció del sòl. De fet, la majoria dels fongs viuen en un pH tirant vers l’àcid, entre 5,5 i 7,5. Els pH més alts afavoreixen la vida bacteriana. Molts fongs són, doncs, acidòfils, i pocs són clarament basòfils. En qualsevol cas, les hifes treballen per mantenir al citoplasma l’equilibri iònic i, quan cal, expulsen l’excés d’ions H+, ja que l’activitat dels seus enzims depèn estretament pel pH citoplasmàtic. Ja hem vist també com alguns fongs poden influir sobre el pH del medi que els envolta.
La majoria dels fongs són aeròbics estrictes, però n’hi ha alguns (llevats, Fusarium oxysporum, Mucor hiemalis) que, en absència d’oxigen, poden realitzar la fermentació (però amb una producció de menys del 10% de la que obtenen en condicions aeròbiques). Són curiosos alguns fongs que viuen en aigües embassades, que, com el quitridiomicet Blastocladiella ramosa, sempre realitzen la fermentació, disposin o no d’oxigen, i, encara, alguns quitridiomicets que viuen al rumen de les ovelles, que són anaeròbics obligats, com els bacteris del gènere Clostridium, i moren en presència d’oxigen.
Qualsevol micòfil mediterrani coneix la importància de l’aigua en l’activitat dels fongs. L’aigua és indispensable en els mecanismes de la lisotròfia. Alguns bolets (sapròfits i petits en general) surten de seguida després de les pluges. D’altres, entre els quals hi ha els més apreciats, que van lligats a l’activitat dels arbres amb els quals estableixen simbiosi, no ho fan fins uns 21 dies després (segons creença popular raonablement exacta). Hi ha fongs xeròfils que poden créixer i fructificar en substrats bastant pobres en aigua. Aquest contingut es pot mesurar en termes d’humitat relativa en equilibri (límit inferior habitual, 70%), activitat de l’aigua o potencial de l’aigua, etc. Si hom posa restes vegetals (fulles, fusta, etc.) recollides al bosc, o fems en una cambra humida (on mantenim una humitat relativa del 100%) hom veu aparèixer molts fongs i mixomicets. Al sòl, el límit superior del contingut d’aigua compatible amb el creixement dels bolets és el que impedeix l’airejament. Per això, pocs sòls embassats presenten bolets. Les èpoques plujoses són favorables per al creixement dels bolets, no solament per les precipitacions líquides, sinó perquè la humitat relativa de l’aire es manté alta molt de temps i s’evita la perillosa evaporació. L’aigua pot ésser escassa per al fong si hi ha un excés de soluts que la mantinguin segrestada. Els fongs que toleren pressions osmòtiques elevades, com Eremascus albus, o les floridures que surten als pots de melmelades, s’anomenen osmòfils, i sovint acumulen al citoplasma compostos osmòticament actius (glicerina).
A continuació, la taula presenta el grau de comestibilitat i toxicitat de diferents bolets, preparada per la Societat Catalana de Micologia. Aquesta llista comprèn, entre altres, les espècies que tenen nom popular català. Les comestibles han estat repartides en quatre grups, segons llur qualitat, d’acord amb un cert consens, entre experts de la Societat Catalana de Micologia, coordinats per August Rocabruna. Dins de cada grup, s’han endreçat per ordre alfabètic. Algunes espècies no s’inclouen entre les comestibles per la facilitat d’ésser confoses amb d’altres de tòxiques. Tampoc hi són les que són comestibles però rares, poc conegudes o massa petites. Moltes d’altres, sense arribar a ésser tòxiques, s’han de refusar pel fet que tenen sabors desagradables (picant, amarg, etc). Només n’hem fet la llista de les principals. Hem indicat amb un asterisc (*) les que són bones degudament tractades, i amb dos (**) les que resulten tòxiques si hom les consumeix amb begudes alcohòliques. Les espècies tòxiques no són gaires, i n’hem esmentat 26 de les més importants. El consum d’uns deu d’aquestes pot comportar emmetzinaments mortals.
Genètica dels fongs
Molts fongs, com Neurospora crassa, Sordaria fimicola, Saccharomyces cerevisiae, Aspergillus nidulans i Phycomyces blakesleeanus, han estat molt emprats en estudis de genètica haploide. Quan hom provoca mutacions en nuclis haploides, els efectes es manifesten de seguida. A més, els cultius de fongs són fàcils de fer, barats, i ocupen poc espai. D’altra banda, el seu cicle de vida és curt. En els ascomicets, el contingut d’un asc comprèn la totalitat dels productes de la meiosi, de manera que les espores es poden prendre d’una en una i analitzar.
La unió de gàmetes o de nuclis gamètics (per simplificació dels processos sexuals) condueix essencialment a la recombinació genètica. Per una banda, es recombinen els caràcters en redistribuir-se entre els nuclis els cromosomes que en són portadors. Però també ho fan gràcies a l’intercanvi de fragments de cromosomes que es produeix durant l’encreuament. Així es barreja informació entre cromosomes homòlegs. Tots aquests processos tenen com a conseqüència biològica bàsica la conservació o augment de la variabilitat en les poblacions. Però ja veurem com els fongs hi han afegit innovacions pròpies.
La compatibilitat
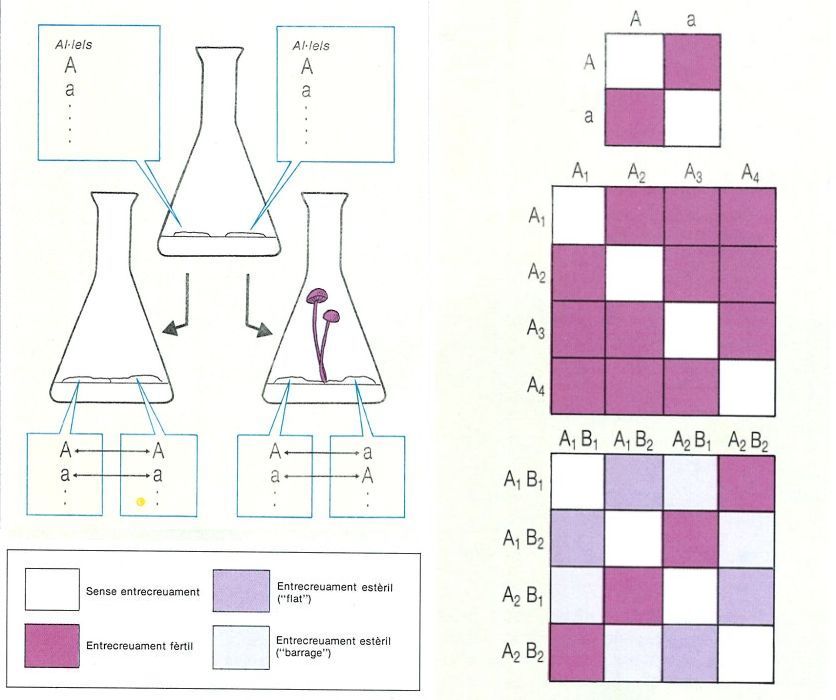
L’heterotal·lia és una barrera genètica amb la qual molts fongs eviten l’endogàmia (encreuament entre micelis massa semblants genèticament). El dibuix ens mostra un experiment de confrontació de dos micelis de Stropharia semiglobata, un fong que mostra heterocariosi bipolar, i les dues possibilitats: en primer lloc, el cas en què no hi ha somatogàmia (esquerra); i en segon lloc, quan la somatogàmia es produeix i apareixen carpòfors (dreta, en morat). A sota, possibles encreuaments en dos casos d’heterocariosi bipolar i en un de tetrapolar.
Biopunt, a partir de fonts diverses
Alguns fongs són homotàl·lics i es poden autofecundar; això però, no afavoreix la recombinació genètica. Per aquesta raó, la majoria dels fongs han desenvolupat barreres tendents a evitar l’endogàmia, i han esdevingut heterotàl·lics. Hi pot haver micelis masculins, amb gametangis masculins, i micelis femenins, amb gametangis femenins. Però quan aquests no es poden diferenciar, hom parla simplement de micelis tipus + i tipus −, i llur comportament és determinat per dos al·lels pertanyents a un sol locus. En el cas dels zigomicets, per exemple, només conjugaran gametangis d’un miceli + amb els d’un miceli −. Una cosa semblant passa en els ascomicets. Tots aquests són casos d’heterotàl·lia dipolar. No és rar, però, trobar en aquest darrer grup casos de partenogènesi, on un gametangi femení origina directament hifes dicariòtiques sense fecundació prèvia. Cal interpretar-ho com un mecanisme de supervivència en casos en què la reproducció sexual és difícil. En els basidiomicets podem trobar també casos d’heterotàl·lia dipolar, però sovint és multial·lèlica, és a dir, regida per un sol gen que presenta diversos al·lels (per exemple, A1,A2, A3, A4). Les combinacions que es poden presentar per encreuament, en el cas en què hi ha 4 al·lels, són 16, 4 de les quals són incompatibles; les 12 restants condueixen a la formació d’un miceli secundari (dicariòtic), que es manifesta formant fructificacions. Bolets tan coneguts com el bolet de tinta (Coprinus comatus) o Stropharia semiglobata es comporten així. Però és més freqüent l’heterotal·lisme tetrapolar amb al·lels múltiples, regulat no per un sinó per dos gens, A i B, que poden presentar al·lels (A1, A2, A3 i B1,B2, B3...). En el cas de 4 al·lels, de les 16 combinacions possibles, només 4 són compatibles, 4 són incompatibles i les altres condueixen a inicis de reproducció, que no porten, però, a l’aparició final de fructificacions. Aquestes reaccions tan complexes serveixen per a evitar la consanguinitat i per assegurar que l’encreuament es produeixi entre micelis genèticament al més diferents possible.
L’heterocariosi
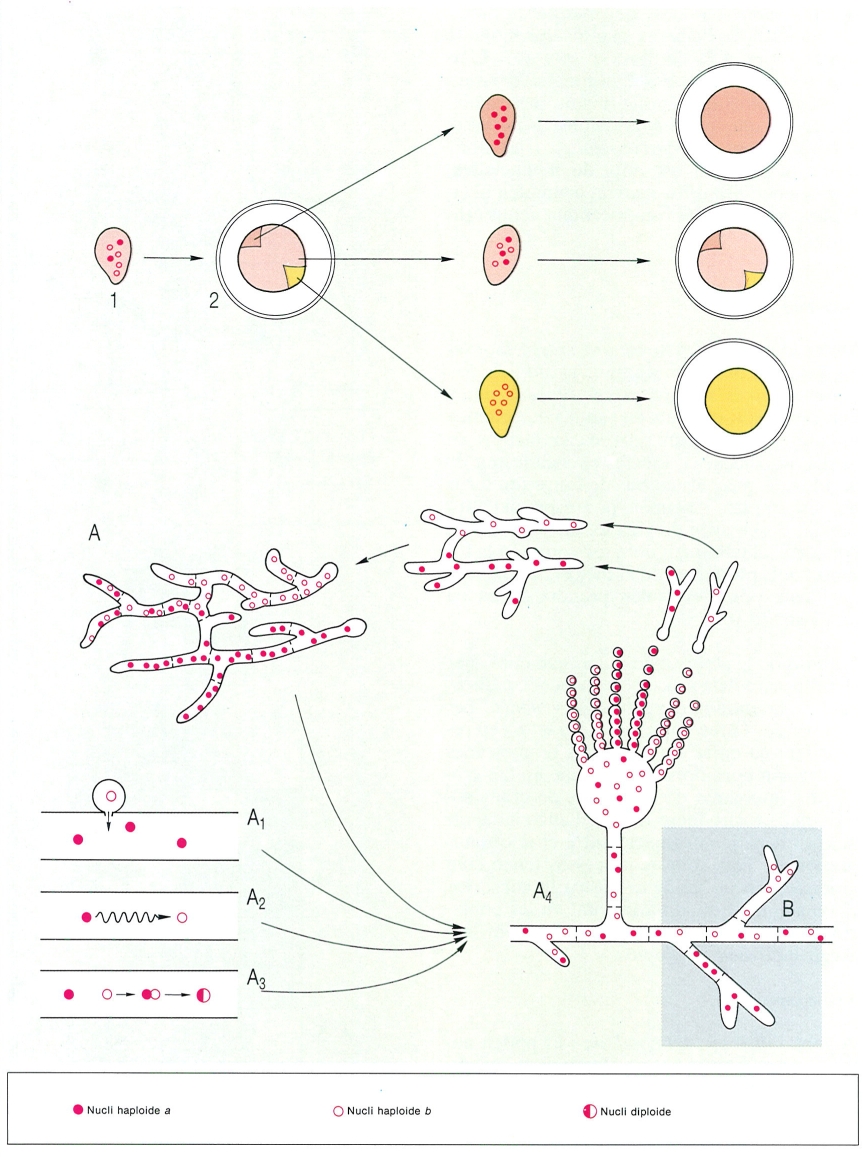
Heterocariosi en els fongs. A dalt, l’esquema mostra en un exemple senzill el fenomen de i’heterocariosi pel qual, a partir d’una espora emesa per un asc (1), i com a conseqüència del procés il·lustrat a baix (B) (en què, a partir d’un miceli heterocariòtic, es poden originar hifes amb un sol tipus de nucli) es forma una colònia (2) en la qual apareixen dos sectors de característiques diferents de les de la resta de la colònia, com a conseqüència del desenvolupament d’hifes amb un mateix tipus de nucli a cadascun d’aquests sectors. Si resembrem per separat el miceli de cadascun d’aquests sectors, obtindrem micelis de característiques uniformes. En canvi, si fem el mateix amb la colònia principal, tornen a aparèixer sectors, com a conseqüència de l’heterogeneïtat dels nuclis. Hom ha representat els ascs, amb els diferents tipus de nuclis que contenen. A Mecanismes que poden intervenir en la formació d’un miceli heterocariòtic: A1 incorporació d’un nucli procedent d’una espora, A2 mutació, A3 diploïdització, A4 aspecte d’un miceli heterocariòtic (Aspergillus) amb dos tipus de nuclis genèticament diferents, que originen dos tipus de conidis. Cada tipus de conidi dona lloc a una hifa monocariòtica; aquestes es poden fusionar i donar lloc, novament, a un miceli heterocariòtic. Quan el miceli es ramifica (B), és possible que s’originin branques amb un sol tipus de nucli, que es convertiran en els sectors al·ludits anteriorment en A.
Biopunt, a partir de fonts diverses
Si l’aparell vegetatiu d’un fong s’ha originat a partir d’una espora, hom pot imaginar que tots els seus nuclis seran iguals (miceli homocariòtic). A la realitat però, molt sovint trobem en un mateix miceli dues o més menes de nuclis, genèticament diferents, que es distribueixen a l’atzar i es multipliquen independentment els uns dels altres. Que el miceli sigui heterocariòtic significa que, des del punt de vista genètic, els al·lels es complementen i el miceli funciona com si fos dicariòtic o diploide. El nombre relatiu de cada mena de nuclis en el conjunt pot variar sota l’efecte de la pressió de selecció ambiental. Hom obté així una millor adaptació del miceli segons les circumstàncies. Un miceli heterocariòtic no es comporta, doncs, com un individu, sinó com una veritable població. L’heterocariosi es pot donar per la intervenció de diversos fenòmens, com ara: mutació d’algun dels molts nuclis del miceli; fusió d’una hifa amb una altra (o d’una espora amb una hifa), acompanyada de cessió de nuclis; diploïdització per fusió de dos nuclis (un fenomen rar, però existent); germinació d’una espora portadora de més d’un nucli, etc. Els fongs heterocariòtics poden tornar a ésser homocariòtics per mitjà de la formació i germinació d’espores unicel·lulars o per emissió de branques amb una sola mena de nuclis. Descoberts al laboratori, no se sap encara fins a quin punt els micelis heterocariòtics són importants en condicions naturals.
La parasexualitat
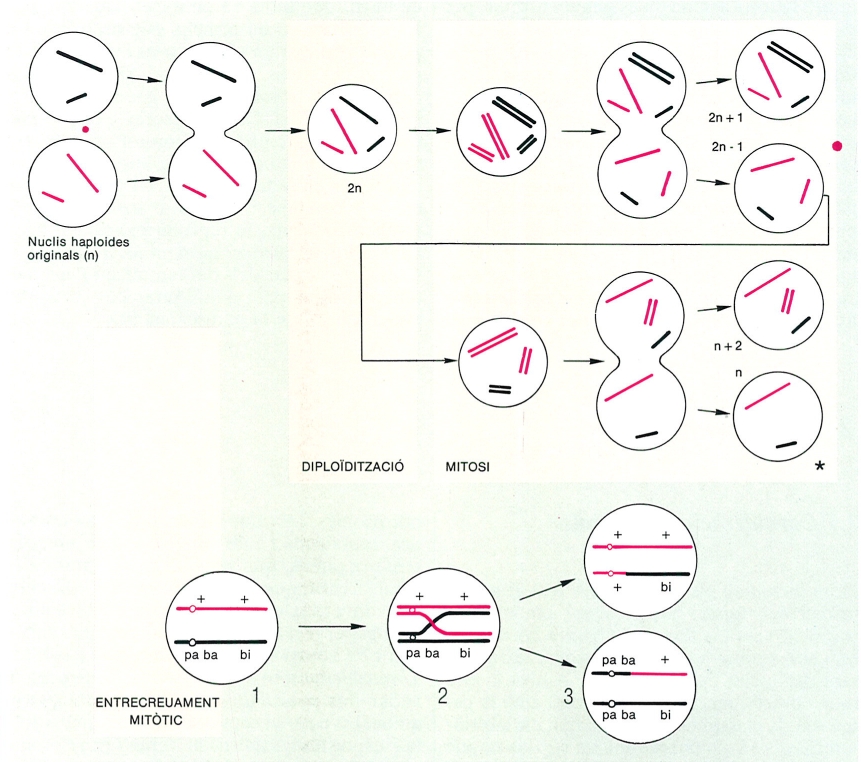
Parasexualitat: dos exemples del tipus de processos implicats. A dalt, seqüència de canvis que pot conduir a la recombinació de cromosomes, és a dir, a l’aparició de nuclis (*) dotats d’una combinació de cromosomes diferent de les existents al començament del procés. Fixem-nos que en les dues mitosis es produeix una mala disjunció, que afecta un dels cromosomes. A baix, seqüència de canvis que pot conduir a la recombinació, per entrecreuament mitòtic, de segments de cromosomes homòlegs. 1 Dos cromosomes homòlegs, un dels quals té dos al·lels mutants; 2 el procés de duplicació pot implicar, també en la mitosi, un entrecreuament; 3 després de la separació es produeix un trencament, seguit de soldadura i, com a conseqüència, apareixen cromosomes amb una informació genètica diferent a la que teníem en 1.
Biopunt, a partir de fonts diverses
Durant l’estudi de l’heterocariosi en Aspergillus nidulans, es trobaren algunes espores «inesperades», que posseïen informació genètica «recombinada», o sigui, tenien nuclis que no eren iguals a cap dels que hi havia al miceli heterocariòtic. Això portà al descobriment d’un fenomen que només es dona entre els fongs superiors, la parasexualitat, que implica una sèrie de processos que, sense que es fusionin dos gàmetes, permeten recombinar la informació genètica dels nuclis preexistents. Els processos no són gaire ben coneguts, però exigeixen la presència prèvia d’un miceli heterocariòtic on es puguin formar, per diploïdització (deguda a la fusió entre dos nuclis amb informació genètica diferent), nuclis diploides. En aquests es poden donar fenòmens d’encreuament durant la mitosi, que recombinen la informació genètica dins els cromosomes homòlegs. Però encara que aquests encreuaments, poc probables, no es donessin, es produeixen de tant en tant processos d’haploïdització complexos, que condueixen a l’aparició de nous nuclis haploides, per pèrdua de cromosomes amb la intervenció de fenòmens de no disjunció. Es tracta, doncs, d’un cicle irregular, no controlat. Que arribi a realitzar-se és possible gràcies al nombre enorme de nuclis que hi ha en un miceli, que fan que transformacions en principi poc probables es puguin realitzar en alguns casos i arribar a l’aparició de nuclis «nous», és a dir, amb una combinació d’informació genètica abans inexistent. Aquest curiós procés ha aparegut evolutivament en fongs (sobretot ascomicets) que passen una bona part de llur vida en forma de miceli heterocariòtic. La seva eficàcia en la recombinació genètica ha fet que en molts ascomicets, la reproducció sexual hagi anat perdent protagonisme fins a arribar a desaparèixer (cas dels deuteromicets), que no tenen altra manera d’assegurar llur variabilitat genètica que la parasexualitat.