Els protosteliomicets
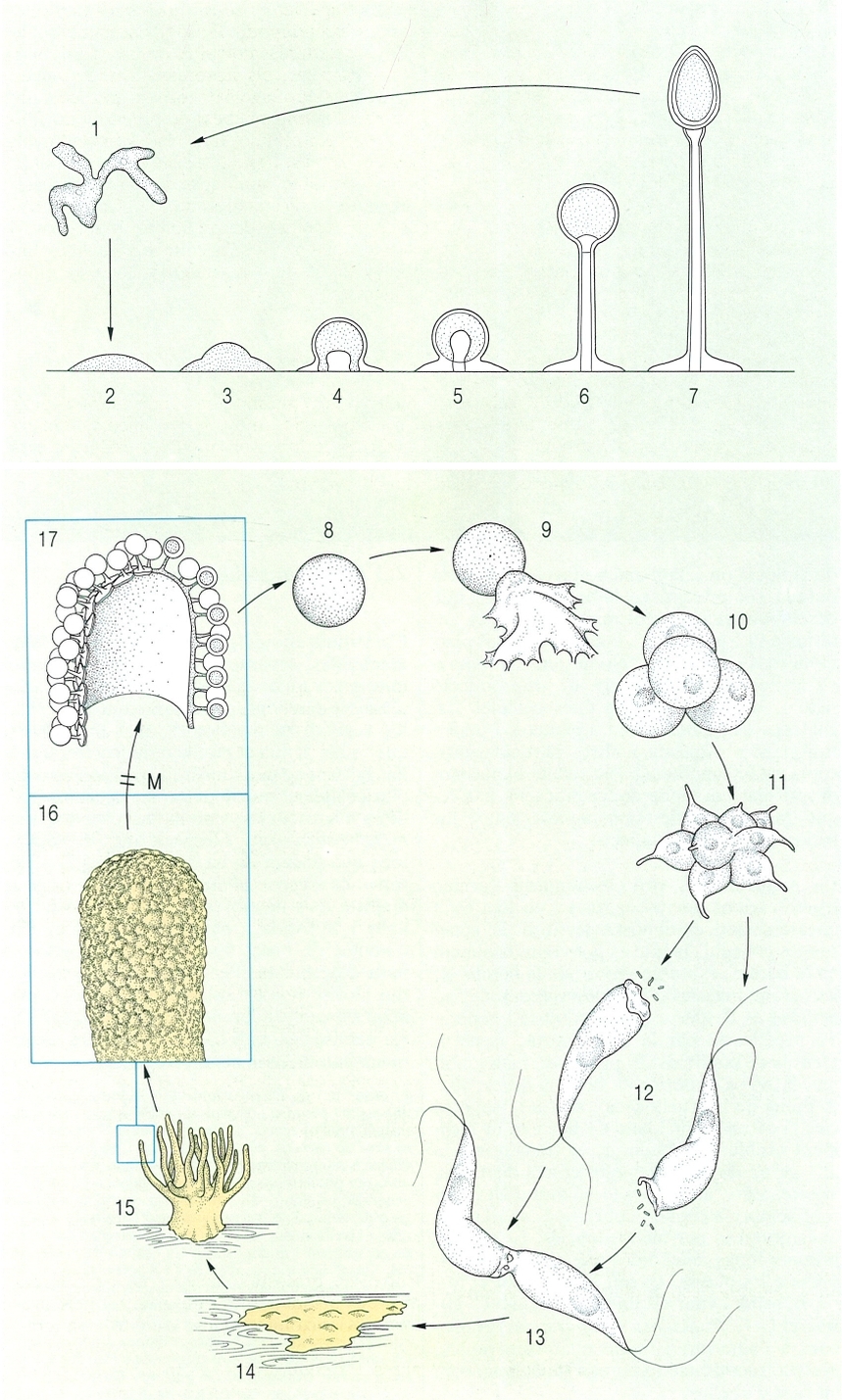
Processos biològics característics dels protosteliomicets: a dalt, esporogènesi (en Nematostelium ovatum); a baix, cicle vital (de Ceratomyxa fruticulosa). Hom hi ha indicat: 1 cèl·lula vegetativa ameboide (procedent de la germinació de l’espora), 2 estadi inicial de prespora, 3 estadi en forma de barret, 4 aparició de l’esteliogen, 5 inici de la formació de l’estípit, 6 darrer estadi de la formació de l’estípit (l’esteliogen penetra dins el tub de l’estípit), 7 esporocarp madur, amb una espora terminal envoltada per una coberta exterior (a la part alta de l’estípit hi ha una apòfisi), 8 espora, 9 germinació de l’espora, 10 tètrade, 11 octet, 12 cèl·lules flagel·lades (cèl·lules en eixam), 13 plasmogàmia o fusió de dues cèl·lules flagel·lades, que actuen com a gàmetes; 14 formació del plasmodi en un tronc en descomposició, 15 esporòfor ramificat, 16 detall d’una branca de l’esporòfor, coberta de protòspores (en les quals té lloc la meiosi); 17 protòspores sobre els seus peduncles respectius, formades per un esporangi que conté una sola espora. Hom indica en el dibuix en color blanc la fase haploide, i en el taronja la fase diploide.
Francesc Xavier Yuste, a partir de L. S. Olive i R. W. Scheetz
Aquests organismes, també anomenats protostèlids, són majoritàriament microscòpics i, per tant, és molt fàcil que al camp ens passin per alt. Avui dia en coneixem unes 25 espècies, repartides extensament pel món. Molts necessiten de 3 a 5 dies per arribar a la fructificació. Si volem fer un estudi acurat, un cop aconseguida la germinació hem de resembrar els estadis vegetatius en plaques de cultiu amb medis pobres en nutrients, per tal d’evitar que s’hi desenvolupin altres espècies contaminants; després, s’hi afegeixen bacteris o llevats com a aliment.
Els cicles vitals són molt diversos, tot i que segueixen un model general que comprèn els estadis d’espora, ameboflagel·lat, ameboide obligatori, fructificació i espora. L’estadi ameboflagel·lat és constituït per protoplasts ameboides, acompanyats o no de cèl·lules flagel·lades, els quals es poden encistar quan les condicions es tornen desfavorables. La majoria d’espècies estudiades presenten estadi ameboide, i és situat en una posició del cicle vital anàloga a l’estadi plasmodial dels mixomicets.
En els cicles vitals més senzills, com el de Protostelium mycophaga, no hi ha un estadi ameboide obligat clar. La germinació de les espores dona pas a un estadi vegetatiu constituït per cèl·lules ameboides, que probablement correspon a un estadi ameboflagel·lat de llarga durada. Les cèl·lules ameboides, de color taronja, repten sobre el substrat emetent pseudopodis, es nodreixen de llevats, i es divideixen per plasmotomia (una divisió cel·lular no sincronitzada amb la del nucli, que dona cèl·lules filles de mida desigual). Passats gairebé tres dies de vida, les cèl·lules deixen de nodrir-se, aturen el moviment de translació i concentren el protoplasma a la part central, formant un estípit tubular, fi i fixat al substrat, que eleva gradualment la massa protoplasmàtica. Al final de la maduració, observem un cos fructífer o esporocarp (de 50 a 150 µm d’alt) que consta d’un estípit que conté cel·lulosa i sosté, a la part apical, una sola espora, de 15 a 18 µm de diàmetre.
Més complex és el cicle vital de Schizoplasmodium cavosteloides. L’estadi ameboflagel·lat, de llarga durada, és representat per protoplasts ameboides. L’estadi ameboide obligat és un plasmodi reticulat on, a diferència dels mixomicets, no són observables corrents protoplasmàtics rítmics. El cos fructífer presenta bal·listòspores, que són expulsades a una certa distància mercès a l’ejecció d’una substància (potser un gas a pressió) retinguda en una vesícula. És un caràcter que no apareix en cap altre dels fongs ameboides.
En els cicles de Cavostelium i Ceratiomyxella apareixen cèl·lules flagel·lades en l’estadi ameboflagel·lat, que presenten un flagel anterior llis i, sovint, un altre de semblant, però més curt. Els flagels estan connectats amb el nucli.

El protosteliomicet més fàcil de veure és Ceratiomyxa, que forma, sobre fusta en descomposició, una gespa d’estructures blanques, eriçades de peduncles, portadors cadascun d’una exòspora. La fotografia correspon a C. porioides, de superfície porada, si bé l’espècie més freqüent és C. fruticulosa, que forma estructures arborescents en forma de ditet, força semblants a primera vista a les de la fotografia.
August Rocabruna / SCM
Entre els protosteliomicets que considerem més evolucionats hi ha un grup, el dels protostèlids mixomicetoides, que té una gran afinitat amb els mixomicets. Comprèn els gèneres Ceratiomyxa, Protosporangium i Clatostelium. La germinació de llurs espores dona lloc a un estadi ameboflagel·lat de curta durada, sempre amb cèl·lules flagel·lades, que desemboca en un estadi ameboide obligat, anterior a la fructificació, que és anàleg al plasmodi dels mixomicets. Només en Ceratiomyxa coneixem l’existència de reproducció sexual, representada per dues cèl·lules flagel·lades que actuen com a gàmetes i poden fusionar-se. Aquesta i altres particularitats, com l’estadi ameboflagel·lat reduït, el plasmodi reticulat i el mode de fructificació, han fet que durant molt de temps aquest gènere fos inclòs entre els mixomicets.
La majoria dels protosteliomicets formen fructificacions microscòpiques i no han estat encara cercats en el nostre territori. El gènere més conegut, ja que es pot veure fàcilment en la natura, és Ceratiomyxa, de la família de les ceratiomixàcies (Ceratiomyxaceae). C. fruticulosa és l’espècie més corrent del gènere. Té predilecció per la fusta morta, especialment la de coníferes. El plasmodi, hialí i molt mucós, viu a l’interior de fusta ja degradada. A l’inici de la fructificació, surt a la superfície, pren un color blanc de llet i es fa fàcilment visible. En madurar, es transforma en una gespa de múltiples estructures digitades, arborescents, acel·lulars (1-10 mm d’alt), gairebé sempre blanques, cobertes d’espores hialines portades per fins peduncles. És un dels primers fongs ameboides que apareixen després de les pluges. Es pot trobar a tot el territori, sense evitar els llocs més càlids. C. porioides (=C. fruticulosa var. porioides) és propera a l’anterior, i sovint apareixen juntes, però la fructificació és porada reticulada, com un rusc d’abelles.
Els dictiosteliomicets
Constitueixen un grup homogeni de fongs ameboides, separat recentment dels acrasiomicets per raons que hom explicarà en parlar d’aquest darrer grup. Presenten només cèl·lules vegetatives ameboides, mai flagel·lades, que, quan arriba el moment de fructificar, solen agregar-se per constituir pseudoplasmodis pluricel·lulars, que donaran lloc a cossos fructífers microscòpics, amb moltes espores, anomenats sorocarps. L’espècie més estudiada, tot i que encara no ha estat observada a Europa, és Dictyostelium discoideum. Com la majoria dels membres del grup, ha estat aïllada i cultivada a partir de mostres de sòl, d’humus de bosc, fusta en descomposició o fems d’herbívors. Per a dur a terme els cultius es disposen els substrats que s’han d’analitzar en cambra humida o sobre plaques d’agar sembrades amb bacteris (com ara Escherichia coli o Aerobacter aerogenes).
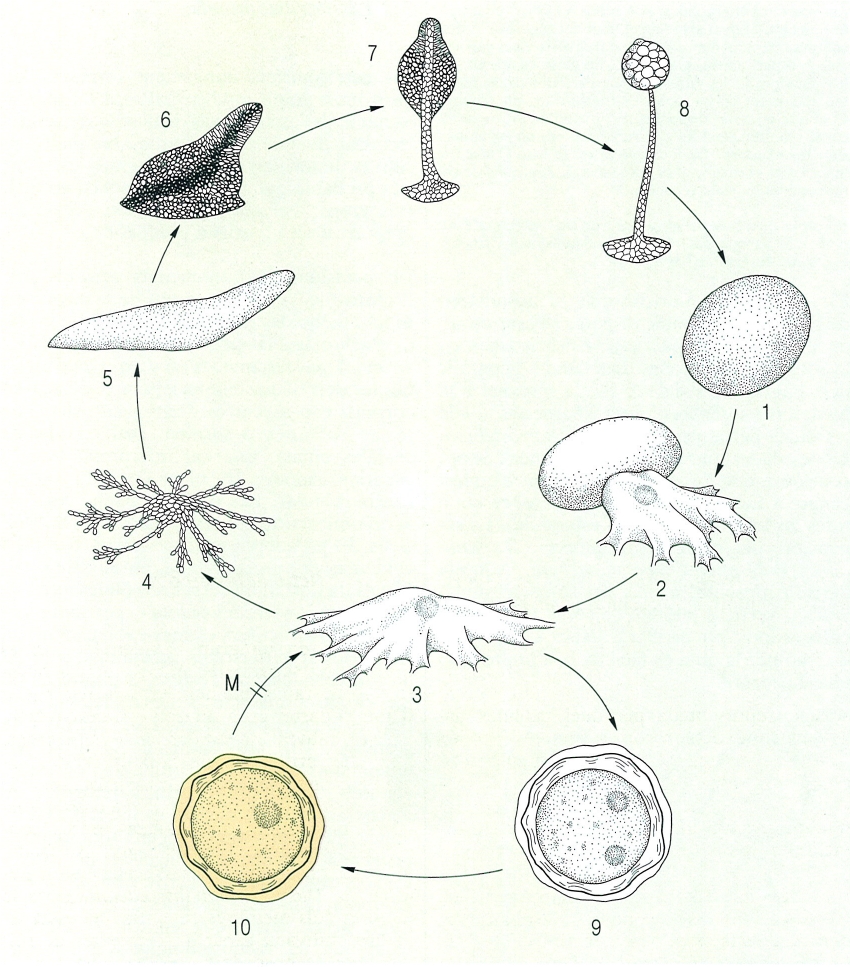
Cicle vital del dictiosteliomicet Dictyostelium discoideum, capaç de completar-se en un temps curt, d’algunes hores. Segons la disponibilitat d’aigua, el cicle es tanca sense cap període de repòs (1-8), o inclou la formació de cists de resistència (9-10). Hom ha representat en el dibuix en blanc la fase haploide, i en el taronja, la fase diploide, i hi ha indicat: 1 espora, 2 germinació de l’espora, 3 cèl·lula ameboide o vegetativa, 4 agregació de cèl·lules ameboides (a escala molt més petita), 5 pseudoplasmodi, 6 inici de la fructificació o culminació (les amebes de la part alta han originat el soròfor, que ha de sostenir les espores), 7 inici de la formació del soròfor (aquest, anirà elevant el futur sorus), 8 sorocarp madur (duu el sorus madur a l’extrem format per espores embolicades per un mucilag, i el soròfor pluricel·lular i tubular, format per cèl·lules mortes i recobert d’una capa cel·lulòsica), 9 formació d’un macrocist haploide, 10 macrocist diploide (després de la cariogàmia; prèvia meiosi, donarà lloc a una cèl·lula ameboide).
Francesc Xavier Yuste, a partir de R. W. Scheetz
El cicle vital de Dictyostelium discoideum s’inicia quan una espora germina i origina una cèl·lula ameboide única (estadi ameboflagel·lat), que repta sobre el substrat a la recerca de bacteris, els quals fagocita. Les amebes es divideixen mitòticament, una i altra vegada, fins a formar una gran població de cèl·lules independents. El temps de generació és d’entre 2 i 3 hores. Quan arriben a un cert grau de densitat, o quan la població bacteriana s’ha reduït a nivells massa baixos, les amebes frenen llur moviment, nutrició i multiplicació. És l’estadi de preagregació, que té una durada d’entre 4 i 8 hores, al cap de les quals s’observa un moviment d’atracció entre les amebes. Aquestes, primer s’associen en petits grups, i posteriorment actuen com a centre d’agregació, vers on és atreta la resta de les amebes. Només responen a aquesta crida les amebes en estadi de preagregació, mentre que les que estan en fase tròfica hi són insensibles. Durant el procés d’agregació pot detectar-se en el medi la presència d’una substància, l’acrasina, identificada després com AMP-cíclic (3’, 5’-adenosín monofosfat cíclic), que actua com a missatger quimiotàctic. El centre d’agregació deixa anar aquesta substància al medi i això fa que centenars o milers d’amebes formin corrents que convergeixen en el mateix punt. Allí es crea una massa de cèl·lules individualitzades i adherides entre elles, que actua com un organisme pluricel·lular, el pseudoplasmodi, que és allargassat, cilíndric i té moviments de reptació, però, a diferència dels plasmodis dels mixomicets, no té corrents protoplasmàtics interns ni capacitat de fagocitosi. Si hom fragmenta un pseudoplasmodi i en posa una porció en un medi ric en bacteris, les cèl·lules que el formen reprenen la vida lliure. Si hom el deixa en el mateix medi on era, cada segment actua com una unitat, després d’un cert període de latència. El desplaçament del pseudoplasmodi sobre el substrat deixa al seu darrere un rastre constituït per restes d’una coberta mucosa que l’envolta. Aquesta es va formant de nou contínuament. La direcció del moviment indica una atracció per la llum i que és sensible a la temperatura, fet que no es dona en les amebes en solitari. El pseudoplasmodi defuig, però, il·luminacions i temperatures altes. El mecanisme del moviment és encara poc conegut. Se sap, però, que la coberta és estacionària, i no efectua traccions sobre el substrat. En canvi, sembla que sí que existeixen traccions entre les cèl·lules ameboides i la coberta. Per tant, el pseudoplasmodi es mouria de manera semblant a com ho fa la cinta d’un vehicle eruga. La migració del pseudoplasmodi té poca durada i el recorregut és curt. Entre els factors que influeixen en la durada del desplaçament, el més important sembla ésser la humitat relativa. Quan aquesta baixa per sota d’un nivell crític, el pseudoplasmodi s’atura i entra en fructificació. En iniciar-se aquesta, es fa patent que en el pseudoplasmodi hi havia una certa diferenciació entre les cèl·lules, ja que les anteriors són grans, consumidores d’energia, i estan destinades a formar el peu del sorocarp, i les posteriors són petites, estalviadores d’energia, i acabaran esdevenint espores. El procés de fructificació, també anomenat de culminació, és complex. S’inicia amb una densificació del pseudoplasmodi, que s’arrodoneix i pren l’aspecte d’una gota més o menys cònica. Les cèl·lules generadores de l’estípit es disposen a la part culminal i les generadores d’espores resten a la base. Les cèl·lules culminals van formant internament l’estípit, que es dirigeix vers el substrat travessant la massa cel·lular i, quan entra en contacte amb el substrat, empeny cap amunt tota la massa cel·lular, el sorogen. En arribar a la maduresa, l’estípit és pluricel·lular, tubular i de coberta cel·lulòsica, format per cèl·lules mortes, de parets una mica engruixides. Això dona a l’estípit un aspecte reticulat, al qual al·ludeix el nom del gènere. L’estípit continua creixent a mesura que noves cèl·lules es van afegint a la part apical. Quan totes les cèl·lules apicals s’hi han incorporat, les altres amebes s’arrodoneixen, secreten una paret cel·lulòsica i es transformen en espores. Com que la massa d’espores no prové d’un protoplasma comú i multinucleat, el cos fructífer no és un esporangi, sinó un sorocarp. La localització de la cariogàmia i la meiosi dins el cicle vital es desconeix. L’existència de parasexualitat ha estat reconeguda.
Els altres membres del grup tenen un cicle vital semblant, però n’hi ha que no formen pseudoplasmodi i fructifiquen just després de l’agregació.
El coneixement florístic dels dictiosteliomicets europeus és molt dispers, i basat en algunes campanyes puntuals. No existeixen citacions d’aquests fongs dins l’àmbit geogràfic dels Països Catalans. A escala mundial n’hi ha descrits 4 gèneres i unes 30 espècies, 20 de les quals pertanyen al gènere Dictyostelium. De les dades que es tenen actualment es pot concloure que l’espècie més corrent d’aquest gènere a Europa és D. mucoroides, present a totes les localitats fins avui estudiades. La segueixen, per ordre de freqüència, D. minutum i Polysphondylium violaceum. Aquesta darrera presenta, a més del sorus terminal, violaci, verticils laterals de sorus portats per curts estípits; pot arribar a assolir 2 cm d’alçada, raó per la qual és fàcil de confondre amb una mucoràcia.
Els acrasiomicets
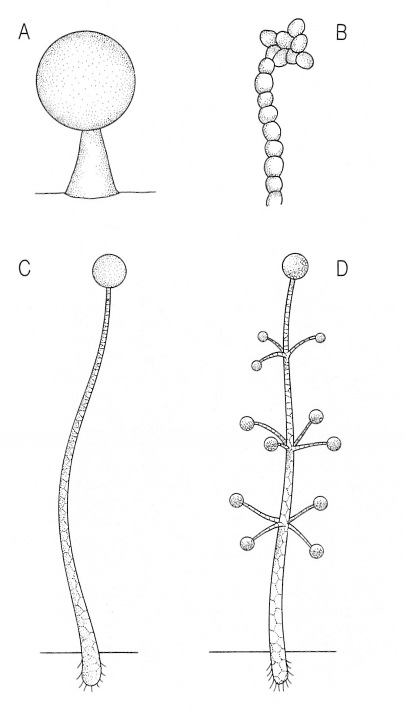
Sorocarps de dos acrasiomicets (A, B) i de dos dictiosteliomicets (C, D). El soròfor dels acrasiomicets és format per cèl·lules vives, i el dels dictiosteliomicets ho és per cèl·lules mortes. A Sorocarp de Guttulinopsis (0,3 mm d’alçada, peu inclòs), B sorocarp d’Acrasis(0,1 mm d’alçada, peu inclòs), C sorocarp de Dictyostelium (4 mm d’alçada, peu inclòs) D sorocarp de Polysphondylium (4 mm d’alçada, peu inclòs).
Francesc Xavier Yuste, original de K. B. Raper
Els acrasiomicets antigament formaven un grup molt ampli, que actualment ha quedat reduït amb l’escissió dels dictiosteliomicets (el lector ha de tenir present aquest fet quan consulti la major part de la bibliografia existent) i és probable que encara s’escindeixi en d’altres grups. Pel que sembla, són encara un grup artificial i d’origen polifilètic.
Els acrasiomicets (anomenats acràsids pels prototistòlegs) es diferencien de la resta dels fongs ameboides perquè les amebes de l’estadi ameboflagel·lat presenten pseudopodis lobosos. Aquestes amebes no s’agrupen com ho fan les dels dictiosteliomicets, ni s’alineen en corrents cap a centres d’agregació, sinó que resten solitàries o formant petits grups de cèl·lules aïllats, que cal interpretar com a pseudoplasmodis. No hi ha, per tant, adherència cel·lular. les cèl·lules ameboides dels acrasiomicets no són estimulades per l’AMP cíclic. El pseudoplasmodi no migra. No hi ha evidència del procés sexual en el cicle vital. En els dictiosteliomicets, les cèl·lules que formen l’estípit secreten parets i, per tant, moren. El cos fructífers és més evolucionat. En els cossos fructífers dels acrasiomicets no hi ha tub cel·lulòsic de l’estípit, a diferència de tots els altres mixomicots. Les cèl·lules de l’estípit es mantenen viables, i són capaces de germinar. Els mitocondris de la resta dels mixomicots tenen crestes tubulars; en canvi, els dels acrasiomicets són heterogenis en aquest aspecte. El nuclèol és diferent. Hi ha indicis que com a mínim alguns acrasiomicets provenen d’un ancessor flagel·lat; per aquesta raó s’explica que en el cicle vital de Pocheina rosea apareguin cèl·lules flagel·lades (és l’únic acrasiomicet que en té). Apareixen, igual que els dictiosteliomicets, sobre una gran varietat d’hàbitats (excrements, escorces, fusta d’arbres i altres restes vegetals) i tenen una àmplia distribució.
Les famílies d’acrasiomicets es distingeixen primàriament les unes de les altres per l’estructura del sorocarp. En algunes, les espores són diferents de les cèl·lules de l’estípit, mentre que en altres són semblants. Entre les espècies més conegudes hi ha Pocheina rosea, Acrasis rosea i Guttulinopsis vulgaris. Als Països Catalans només s’ha trobat la primera, que és corrent sobre mostres d’escorça posada en cambra humida. Les escorces que en produeixen més fàcilment són les de pi, càdec, xiprer, olivera i vinya. Aquesta espècie té els esporocarps estipitats, amb un sorus terminal globós, d’entre 0,04 i 0,1 mm de diàmetre, sovint de color rosaci, i sol aparèixer al cap de pocs dies d’iniciar-se el cultiu (gairebé sempre entre 2 i 4 dies).