Adaptació al medi terrestre: les plantes vasculars
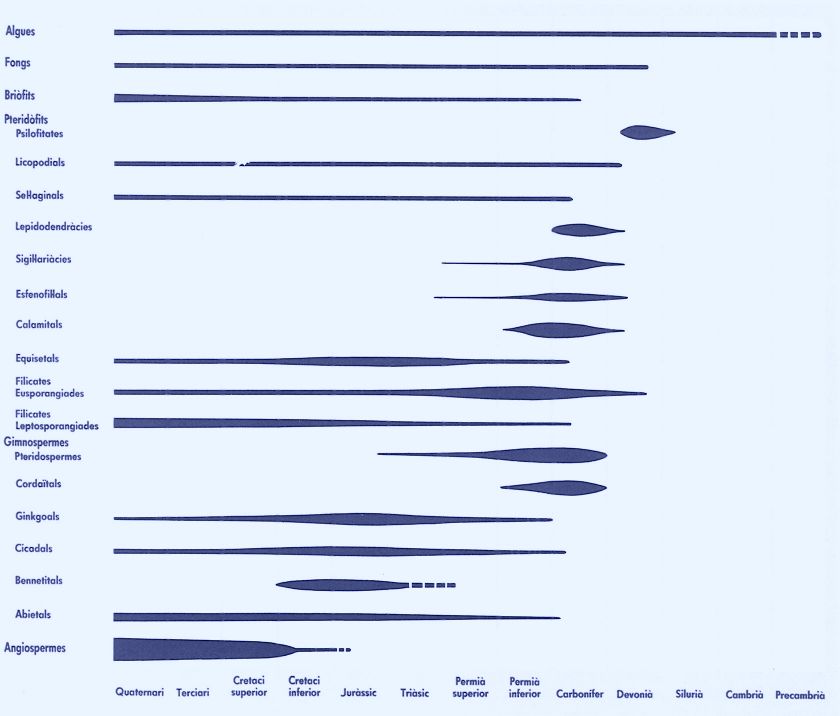
Origen i desenvolupament de les plantes al llarg dels diversos períodes geològics. El gruix de les corresponents línies expressa la importància quantitativa del grup, i la llargada el seu període de presència sobre la Terra.
Maber, a partir de Strasburger.
A la introducció dedicada als briòfits és evocada l’aventura que, per a l’evolució de les plantes, representà l’adaptació a un medi radicalment nou, el medi terrestre, i com resolgueren el problema els tres grans grups bàsicament terrestres (dits també embriòfits) que són els briòfits, els pteridòfits i els espermatòfits. Recordem ara com el grau d’adaptació dels briòfits al medi no aquàtic és imperfecte, de manera que molts d’ells es veuen obligats a viure en ambients permanentment humits o bé, si viuen en llocs secs, només tenen l’alternativa d’aturar llur activitat quan perden l’aigua per evaporació (i això passa sovint, ja que llur cutícula és insuficient per a retenir-la).
Els pteridòfits, que degueren aparèixer si fa no fa al mateix temps que els briòfits (potser a començament del Devonià o abans), seguiren una estratègia diferent en llur evolució: la millora progressiva de la barrera que s’oposa a la pèrdua d’aigua, la cutícula impermeable que recobreix l’epidermis i, com a conseqüència, la conservació, dins d’uns límits estrets i òptims, de la hidratació del citoplasma de les cèl·lules. Els pteridòfits, com els espermatòfits, les plantes terrestres més abundants, són organismes homeohidres (és a dir, que conserven constant el seu contingut intern d’aigua); si perden massa aigua, per tant, les seves cèl·lules moren.
Imaginem ara un model simplificat, mínim, de planta terrestre homeohidra. Ens la podem figurar com un cilindre, amb un pol de fixació, clavat en terra, i l’altre pol, assimilador, dirigit vers la llum. Les seves cèl·lules verdes no són pas gaire diferents de les de les algues clorofícies. Però estan separades de l’aire per una epidermis amb cutícula impermeable. Ara bé, per a la vida d’una cèl·lula vegetal, no n’hi ha prou amb l’aigua i la llum. Cal també gas carbònic i sals minerals (que representen l’aportació de nitrogen, fòsfor, sofre, ferro, etc.). A l’epidermis hi ha d’haver perforacions (els estomes), que permetin l’intercanvi de gasos amb l’atmosfera. Però, llavors, serà inevitable que una part de l’aigua s’escapi per evaporació (transpiració). I, d’on poden poar més aigua que substitueixi la transpirada? Només del sòl, on també hi ha les sals minerals, dissoltes. Cal doncs salvar la distància que separa el pol de la planta enfonsat en el sòl del pol aeri, que rep la llum i transpira. Aviat apareixen uns cordons de cèl·lules, estirades en direcció vertical, que s’especialitzen en la conducció d’aigua i de substàncies dissoltes. El mecanisme de la transpiració serà el motor de la bomba. El nostre cilindre verd, el corm i el seu pol aeri, el teloma, funcionen ja com una planta ben adaptada al medi terrestre, capaç de conservar el seu citoplasma adequadament hidratat i aprovisionat en gas carbònic i sals minerals.
La millora d’aquesta organització bàsica vindrà per un augment de la superfície aèria (per exemple, per ramificació del teloma), per un augment de la superfície subterrània, de fixació i captació (evolució de l’arrel, ramificació), i, sobretot, per un augment de la importància i de l’eficàcia dels vasos conductors. Les plantes amb sistema vascular diferenciat, anomenades plantes vasculars o traqueòfits (els pteridòfits i els espermatòfits) són les plantes dominants, de molt, al medi terrestre.
L’evolució dels òrgans subterranis parteix, al principi, de l’existència d’eixos subterranis, de desenvolupament horitzontal, els rizomes, dotats de rizoides fins, encarregats de l’absorció d’aigua i de sals. Les arrels típiques, ramificades, amb geotropisme positiu, apareixen més tardanament. L’evolució de la part aèria és esperonada per la necessitat d’augmentar la superfície de captació de llum, però també ho és la competència i la consegüent lluita per l’espai i per la llum, que s’estableix quan augmenta la densitat de les plantes. L’augment de la superfície de fotosíntesi es pot produir per ramificació dels telomes. Les primeres plantes vasculars (els primers pteridòfits), s’assemblaven a una petita planta de ginesta, i les branquetes feien de fulles. En una línia evolutiva, aquestes branquetes terminals o laterals es van anar aplanant, tot donant fulletes amb un sol nervi, els microfil·les. En una segona línea evolutiva, un grup de branquetes contigües es disposaren primerament en un pla perpendicular a la llum (procés de planació) i després se soldaren entre elles (concrescència), formant una làmina amb diversos nervis (els feixos de vasos conductors de les branquetes), tot originant així les fulles típiques, els megafil·les.
La creixent importància de la part aèria exigia alhora un augment de la resistència mecànica de la tija i de la capacitat dels vasos conductors. Al principi, el teixit conductor era també l’encarregat principal d’assegurar la resistència mecànica, i només força més tardanament, aparegué un teixit mecànic especialitzat. La distribució dels teixits conductors a la tija, visible en un tall transversal, constitueix l’estela, que sofrí també una interessant diversificació. A les plantes vasculars més primitives, el xilema (és a dir, els vasos encarregats de transportar l’aigua amb sals minerals captada del sòl) se situa a l’eix de la tija, envoltat pel floema (els vasos encarregats de transportar, per tota la planta, aigua amb substàncies orgàniques elaborades, ions, etc.): hom diu que les plantes que presenten aquesta distribució tenen protostela. Però de l’estela surten sovint ramificacions dirigides a les fulles, els rastres foliars. La disposició d’aquests pot donar una estela amb braços radials, en forma d’estrella (en secció): es tracta d’una actinostela, també primitiva. Ara bé, una ubicació del xilema tan lluny com sigui possible de l’eix, fa que la seva eficàcia mecànica sigui molt més elevada. Molt èxit evolutiu han tingut, doncs, les sifonosteles, amb la part central buida, o farcida per una medul·la de cèl·lules grosses. La sortida dels rastres foliars crea aviat un espai buit o interstici foliar. Com a culminació del creixement d’aquests intersticis, apareix una estela en forma de xarxa, la qual, en secció, es veu com una colla de feixos formats per floema i xilema (feixos libero-llenyosos) disposats en anell. Aquesta disposició, dita eustela, és la més freqüent a les plantes superiors. Una mena d’eustela dispersa és l’atactostela, pròpia de les monocotiledònies.
Organització i cicle biològic
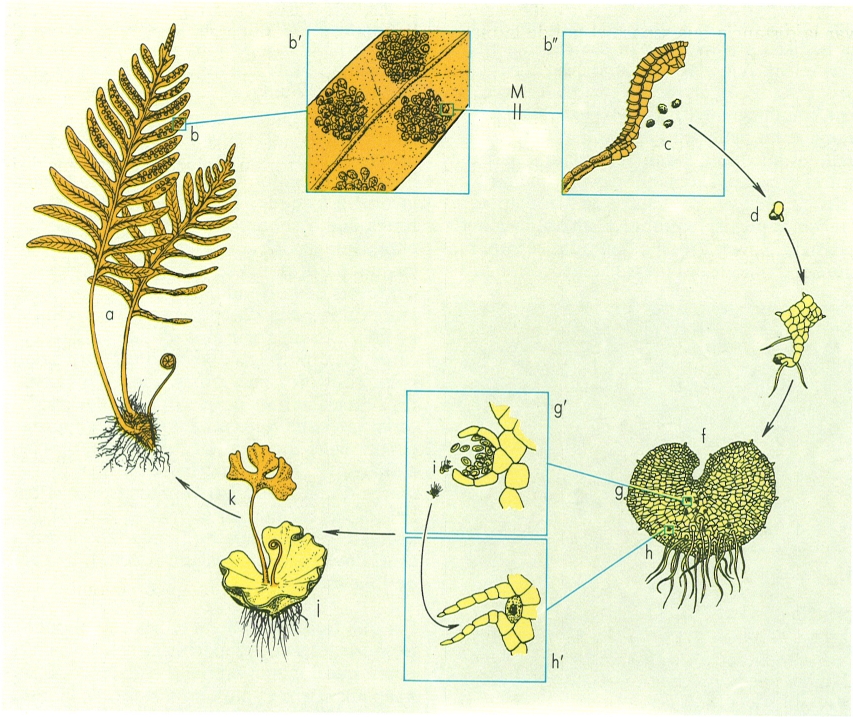
Cicle vital d’un pteridòfit, concretament de Polypodium vulgare (to clar, fase gametofítica, (n) haploide; to fosc, fase esporofítica, (2n) diploide; M reducció meiòtica): a esporòfit (frondes fèrtils, rizomes i arrels); b sorus (b’ grup de sorus augmentats, b" esporangi madur alliberant les espores, més augmentat encara); c espores; d espora iniciant la germinació; e protal·lus en formació; f protal·lus madur o gametòfit; g anteridi (g’ anteridi augmentat alliberant espermatozoides); h arquegoni (h’ arquegoni augmentat); i espermatozoide; j protal·lus en involució; k esporòfit jove.
Maber, a partir de dades dels autors.
Els pteridòfits no estan encara tan ben adaptats a la vida terrestre com els espermatòfits. Ni llur cutícula, ni llur sistema vascular (dotat només de traqueides) no estan encara prou perfeccionats. Per això els trobem preferentment als llocs humits o poc assolellats i, amb algunes excepcions, no atenyen actualment mides gaire grans. La culminació de l’adaptació a la vida terrestre es produeix amb les plantes amb llavors o espermatòfits.
Els pteridòfits comprenen les plantes vasculars més primitives, els licopodis, les selaginel·les, els isòets, les cues de cavall, les falgueres típiques i les falgueres aquàtiques. Molts representants d’aquest grup són extingits i solament es coneixen a través de llurs fòssils. Com acabem de veure, els pteridòfits corresponen a la primera fase de l’adaptació de les plantes homeohidres a la vida terrestre. Llur evolució degué produir-se a partir d’algues clorofícies, que comparteixen amb ells la mateixa mena de cloroplasts i de pigments, la mateixa mena de material de paret (la cel·lulosa) i el mateix tipus de cèl·lules mòbils, amb flagels llisos. Però els pteridòfits tenen ja anteridis i arquegonis, és a dir, òrgans reproductors protegits per una capa de cèl·lules estèrils, no simples cèl·lules reproductores, com les algues. En això, s’assemblen a les molses, però se’n diferencien perquè han orientat llur evolució cap al perfeccionament de l’esporòfit.
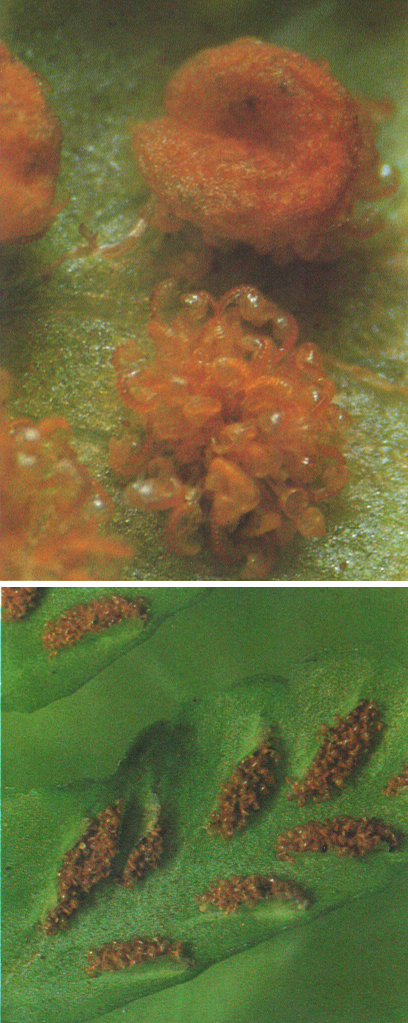
Sorus laminars, és a dir situats sobre el limbe de la fronda, de Polystichum (a dalt) i d’Asplenium (a baix). Ultra els esporangis, s’hi distingeix perfectament l’indusi, central en el primer cas i lateral en el segon.
Enric Curtó.
La fase esporofítica és la que normalment veiem. El gametòfit és força reduït, però típicament és lliure, no lligat a l’esporòfit com passa a les molses i a les hepàtiques. Només l’esporòfit, que és diploide, podia permetre que l’evolució hi acumulés una complexitat d’estructura i de sistema conductor, adequada per a viure en el medi aeri sense deshidratar-se.
Una falguera prou coneguda, el polipodi o herba pigotera (Polypodium vulgare) ens servirà d’exemple per a descriure el cicle vital típic d’una falguera. En germinar sobre un sòl humit i ombrívol, l’espora origina un gametòfit petit i poc diferenciat, el protal·lus, en forma de làmina verdosa, fixada per alguns rizoides. No es diferencia pas massa d’una hepàtica tal·losa. A la cara ventral es formen anteridis i arquegonis, més simplifïcats (amb menys cèl·lules) que els dels briòfits.
En temps humit, els anteridis s’obren i deixen anar els espermatozoides, que tenen forma espiralada i porten diversos flagels llisos. Els espermatozoides fecunden l’ovocèl·lula d’un dels arquegonis, es forma primer un zigot, diploide, i aviat un embrió. Els pteridòfits no poden aturar aquí el desenvolupament de l’embrió (a l’inrevés dels espermatòfits, que produeixen llavors amb un embrió en repòs, molt ben adaptades a la vida terrestre). Per tant, aviat apareix sobre el protal·lus una plàntula amb arrel i una primera fulla, el jove esporòfit diploide. Aquest origina una tija, coberta d’esquames i portadora d’arreletes, que creix entre les pedres o penetra a les fissures. El seu creixement es produeix gràcies a l’activitat d’una sola cèl·lula apical, tetraèdrica (com als briòfits).

Prefoliació circinada de les frondes joves d’una falguera, concretament de Dryopteris filix-mas, la falguera mascle.
Ramon M. Masalles.
Del rizoma van sortint les fulles o frondes que, quan són joves, tenen forma de bàcul de bisbe (prefoliació circinada). Les frondes de la falguera que ens serveix d’exemple són megafilles. A més, serveixen a la vegada per a realitzar la fotosíntesi i per a portar esporangis (són alhora trofofil·les i esporofil·les, és a dir, trofosporofil·les). Per això, quan comencen a madurar, veiem com, al revers, apareixen unes taques grogues, discoïdals: són els sorus (del grec soros, amuntegament) d’esporangis. Cada esporangi té un petit peu i una coberta estèril. Les cèl·lules que hi ha a dins formen una part central, l’arquespori, envoltades per una part perifèrica, el tapet. Les cèl·lules de l’arquespori sofreixen la meiosi i s’embolcallen d’una coberta resistent, sovint bruna o daurada. Mentre maduren, són nodrides pels aliments continguts a les cèl·lules del tapet. La paret de l’esporangi madur s’obre en temps sec, a causa de la tensió que s’origina a les cèl·lules de l’anell, una part especialitzada de la paret. Les espores són dispersades pel vent.
Les principals variacions entorn d’aquest exemple corresponen al protal·lus, que pot ésser cilíndric, o irregular, o molt petit, i no rarament incolor. En aquests casos, viu en simbiosi amb fongs que li subministren aliments. Per altra banda, els esporangis del nostre exemple s’originen a partir d’una sola cèl·lula de l’epidermis del revers de la fronda, no creixen gaire i presenten una paret d’una sola capa de cèl·lules i no gaires espores: són leptosporangis. En altres casos, els esporangis deriven d’un grup de cèl·lules, són força més grossos, tenen més espores i estan limitats per una paret pluristratificada: són eusporangis. La posició dels esporangis pot ésser terminal (al final d’una branca), axil·lar (a l’axil·la d’una fronda), marginal (al marge d una fronda) o laminar (sobre el limbe). No és rar que els sorus laminars estiguin protegits per una làmina fixada lateralment o centralment (com si fos una xinxeta): es tracta de l’indusi.
Pel que fa a les espores, el nostre polipodi és isospòric, és a dir, produeix una sola mena d’espores, que donaran protal·lus amb anteridis i arquegonis. Però és molt important, en la història evolutiva dels pteridòfits, l’aparició de l’heterospòria. Els pteridòfits heterospòrics tenen dues menes d’esporangis. Els microsporangis produeixen només micròspores, en gèneral petites, que en germinar produeixen microprotal·lus, masculins, que només tenen anteridis. Ja podem imaginar que els megasporangis produeixen només megàspores, més grosses, que donen megaprotal·lus, femenins, que només porten arquegonis. Sembla que l’heterospòria tingué èxit en algunes línies evolutives de pteridòfits, de tal manera que arribà a conduir a l’origen, per evolució, dels espermatòfits (que també són heterospòrics).
Sistemàtica
El conjunt dels pteridòfits correspon a un grup de línies evolutives de desigual antiguitat i desigual amplitud. La classe de les psilofïtates (Psilophytataé), la més antiga, s’extingí ja al Devonià, mentre que la classe de les psilotates (Psilotatae) també molt simple, és actual. Més modernes són les classes de les licopodiates (Lycopodiataé) i de les equisetates (Equisetatae), amb microfil·les, i la de les filicates (Filicatae), amb megafil·les; aquesta darrera correspon a les falgueres típiques. Les falgueres aquàtiques (Hydropterides), relacionades amb aquestes darreres, són un grup difícil de situar, i seran considerades per nosaltres com un apèndix de les filicates.