Concepte de mamífer

Un dels trets més característics que participen en la definició de mamífer és la capacitat que tenen d’alletar les seves cries durant les primeres fases de la vida extraembrionària. La fotografia ens mostra una femella de liró gris (Glis glis) amb els seus petits.
Joaquim Reberter.
La definició de mamífer és fàcil d’establir quan es consideren exclusivament les formes vivents. No s’esdevé el mateix quan es consideren les nombroses formes fòssils que testimonien un trànsit progressiu des dels rèptils fins als mamífers. Aquest pas és tan gradual que, realment, es fa difícil, davant de certs fòssils, de poder dir si es tracta d’un rèptil o d’un mamífer. En última instància, la classificació es fa a partir d’un sol os, el dentari, que, en els mamífers, forma la mandíbula inferior. Els altres ossos que presentaven els rèptils s’incorporen en els mamífiers a l’oïda mitjana: l’estrep d’aquests es correspon amb la columel·la dels rèptils i l’hiomandibular dels peixos; l’articular dels rèptils ha esdevingut el martell dels mamífers i el quadrat, l’enclusa; aquest dos ossos dels mamífers representen l’articulació mandibular reptiliana incorporada a l’oïda, incorporació que suposa una millora de l’audició. D’altra banda, l’angular dels rèptils esdevé el timpànic.
Aquest és, en realitat, per excel·lència, el caràcter definitori dels mamífers, però aquests presenten altres característiques diferencials.
El diafragma, entre les cavitats toràcica i abdominal, és completament muscular. L’arc aòrtic esquerre és l’únic existent i la sang porta en suspensió eritròcits no nucleats. En els mamífers vivents, és característica la presència de pèl en algun dels seus estats de desenvolupament, presència que tan sols no es produeix en els adults d’algunes espècies. Els joves són alimentats per les secrecions de les glàndules mamàries de les femelles.
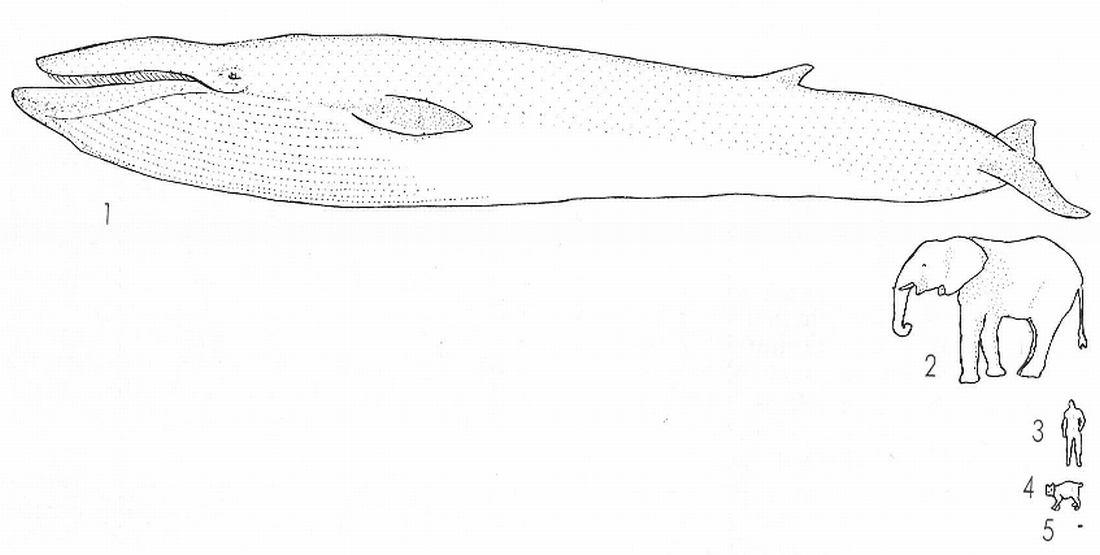
L’àmbit de variació de les dimensions del cos, mentre que és relativament petit en la majoria de classes de vertebrats (especialment en els terrestres), és molt ampli en els mamífers: oscil·la entre els quasi 2 g d’una musaranya i alguns rats-penats fins a les 7 tones d’alguns mascles d’elefant africà. Els mamífers marins tendeixen a augmentar la seva talla fonamentalment per pal·liar problemes de control calòric; així, hom diu que algun exemplar de balena blava pot atènyer les 150 tones de pes, que constituirien el límit superior de massa corporal en els mamífers. En el dibuix hom compara les dimensions d’un rorqual (1), amb les d’un elefant (2), de l’home (3), d’un linx (4) i d’una musaranya (5); aquesta apareix representada només per un punt.
Ferran Rodríguez, a partir d’originals diversos.
Alguns altres caràcters que poden presentar, no estrictament definitoris però de presència molt general, encara que de vegades tampoc exclusiva, són: el viviparisme, que sembla norma general, però que, en realitat, falta en els anomenats monotremes australians, que són ovípars; l’amni, la membrana més interna que embolcalla els fetus, i l’al·lantoide, sac que neix en l’extremitat posterior de l’embrió i contribueix a formar els annexos fetals; la bufeta urinària i l’urac, que es presenten també en els ocells i en els rèptils, presència que mostra, una vegada més, l’origen dels mamífers a partir dels rèptils teràpsids. Aquestes membranes foren les que, en crear un medi aquós entorn de l’ou, possibilitaren la independència dels vertebrats amniotes respecte al medi aquàtic. Són, per tant, una característica comuna dels rèptils, els ocells i els mamífers, que els diferència dels peixos i dels amfibis, que són anamniotes. El manteniment (excepte en algunes espècies, en els joves o durant la hivernació, l’estivació o els endormiscaments) d’una temperatura interna alta, l’homeotèrmia, es considera característic dels mamífers, encara que, molt probablement, es va donar en alguns rèptils extints.
El cervell és gran en comparació amb el d’altres vertebrats i, en els placentaris, hi existeix un cos callós, "corpus callosum"; a més, el cervell, especialment el neopali, creix enormement i, de vegades, pot arribar a cobrir el cerebel i els lòbuls olfactius, la qual cosa permet que aquests animals puguin tenir una vida intel·ligent. Aquest cervell dona forma a un crani relacionat per dos còndils amb la vèrtebra atlas, la primera de la columna vertebral. Tret de comptades excepcions, hi ha dents en el marge mandibular que es canvien una vegada i que, d’ordinari, són diferenciades segons la seva funció.
Es poden donar, encara, altres característiques com la de presentar set vèrtebres cervicals (a excepció d’alguns desdentats i sirenis) i, comparada amb la dels rèptils, la millor estructuració de la caixa toràcica, que permet una respiració més eficaç i, per tant, un metabolisme més elevat. També és molt notable la verticalització dels apèndixs locomotors, aconseguida en doblegar-se cap enrere el colze i cap endavant el genoll.
La perfecta mecànica de l’aparell locomotor i l’elevat rendiment energètic del cos, ha permès als mamífers un domini absolut de la vida continental. Animals amb una temperatura elevada i constant, fruit d’un actiu metabolisme, s’han independitzat molt del medi. El seu cos es manté a la temperatura òptima per al conjunt de funcions orgàniques i aquesta es regula de tal manera que és independent de la de l’ambient. Aquesta capacitat els permet de tenir una àmplia àrea de distribució i, fins i tot, de resistir el fred de les regions polars, on s’enfronten a les condicions adverses causades per l’alt desnivell tèrmic que hi ha entre la temperatura pròpia i la del medi mantenint una gran activitat.
Malgrat que formen una classe amb un nombre d’espècies (poc més de 4000) més petit que el dels ocells, aquelles són d’una diversitat extraordinària. La seva mida oscil·la entre la d’una petita musaranya o rat-penat, amb un pes al voltant de 2 g, fins al d’una balena blava, que arriba a pesar 150 000 kg (pes que representa més de quaranta vegades el d’un elefant adult) i a mesurar 34 m de longitud.
Entre les causes del seu notable èxit, cal considerar el grau d’homeotèrmia assolit, l’hàbit emprenedor, l’enginy i l’atenció parental, que permeten un acurat desenvolupament, i la protecció de les cries.
Les característiques i les adaptacions anatòmiques dels mamífers
El tegument
La pell presenta una epidermis, derivada de l’ectoderma, i una dermis, que deriva de la mesoderma i se suporta en la hipodermis, que recobreix els músculs i els ossos. Inserida a la pell, s’hi troba una estructura, el pèl, característica dels mamífers i, en tot cas, d’alguns rèptils fòssils.
L’epidermis no és irrigada i, des de la seva capa més profunda, l’estrat germinatiu, les seves cèl·lules es divideixen i, en conseqüència, migren cap a la superfície, on es van desprenent amb regularitat. Aquestes cèl·lules fan i acumulen grànuls de ceratina i formen, així, una capa aïllant superficial. Un equilibri adequat entre la velocitat de la divisió cel·lular i el despreniment en la superfície fa que s’estructurin els diversos tipus de pell que es presenten en els mamífers.
Les espècies de mida més gran i amb poc pèl són les que han de tenir la pell més gruixuda. L’elefant, l’hipopòtam, el rinoceront i el porc la tenen d’uns 2 mm de gruix; per aquest motiu, Cuvier els va classificar com a paquiderms, denominació vulgar que encara s’admet però que agrupa animals filèticament distants i que no té, per tant, cap validesa zoològica. De tota manera, el gruix de pell d’aquests animals no és el màxim que s’ha donat en els mamífers; la vaca marina, extingida a causa de l’acció humana, tenia una pell de fins 7,5 cm i hom l’emprava per fer barques. Una de les més primes es dona en els cetacis, en què forma una delicada capa que es desprèn en refregar-la amb els dits; tanmateix, en la beluga arriba a tenir 9 mm de gruix i és, pel seu contingut alimentari, molt apreciada pels esquimals.
La dermis és vascularitzada i presenta, a més de fibres conjuntives, terminacions nervioses i, fins i tot, fibres musculars. En determinades zones paral·leles a la superfície del cos, poden existir reticles de capil·lars, que tenen una estreta relació amb el control tèrmic. Quan hi ha un excés de calor, es dilaten i el flux de sang a la perifèria augmenta i es refreda i aconsegueix, així, augmentar la dissipació del calor. Quan l’individu té fred es contrauen els capil·lars, el flux de sang minva i disminueix la pèrdua de calor. Això es pot observar bé en l’home, que quan té calor mostra bons colors i que, en canvi, empal·lideix en cas de fred i, fins i tot, es posa de color morat, en darrer terme, per reducció i enfosquiment de la sang perifèrica. A la dermis, hi ha un gran nombre d’estructures sensorials i nervis associats amb la detecció del dolor i els canvis tèrmics i de pressió, que permeten de reaccionar enfront dels canvis del medi extern. A més, uns nervis motors inerven estructures vasculars, musculars o secretores de la pell.
La pell dels mamífers intervé en la formació d’importants estructures: l’incubador de l’equidna, la bossa marsupial, la membrana interdigital —en espècies amb intensa vida aquàtica—, les expansions cutànies —que permeten el planatge als dermòpters, marsupials i alguns rosegadors—, el patagi dels rats-penats i, fins i tot, les aletes dorsal i caudal dels cetacis.
En alguns rosegadors, la pell de la cua es desprèn fàcilment perquè no presenta fibres longitudinals i no està fortament adherida a la capa muscular tendinosa. Aquesta disposició és una defensa enfront dels depredadors, que, de vegades, es queden només amb la beina de la cua. En el rosegador, la part de cua sense beina s’asseca i cau.
Implantat a la pell es troba el pèl, esquama modificada característica dels mamífers actuals. N’hi ha de dos tipus: de protecció i de revestiment. Els de protecció són mes forts, llargs i més reduïts en nombre, que no els de revestiment, que són fins i curts. En algunes espècies, com en els pinnípedes, el pèl de protecció és molt reduït o no es presenta. En els insectívors, en canvi, és tan fi que no es veu la diferència entre ambdós tipus de pèl. La missió pristina del pèl és la de formar una capa d’aire tèrmicament aïllant al voltant del cos del mamífer. En espècies com els cetacis, en què el pèl desapareix perquè molestaria la natació, l’efecte d’aïllant tèrmic el fa la capa subcutània de greix.

Les banyes dels mamífers són derivats tegumentaris destinats a la defensa de l’individu. Les fotografies il·lustren diferents tipus de banyes. A l’esquerra, les d’un bòvid, concretament una cabra, que es caracteritzen perquè tenen el nucli ossi central, mesodèrmic, perenne, i recobert d’un estoig corni. A la dreta, les astes o banyes d’un cèrvid (un cabirol), que tenen el nucli ossi, mesodèrmic i caduc, i que cada any es renoven seguint un cicle relacionat amb el reproductor; en el cas de la fotografia, la banya encara és recoberta del «vellut» o capa tegumentària que cau en acabar el seu creixement.
Josep M. Barres.
Existeixen importants derivats cutanis. Els més característics són les glàndules mamàries, que es formen en la denominada cresta mamària o cordó làctic. D’ordinari, a partir d’esbossos que creixen en profunditat per tot el cordó, es van desenvolupant diverses glàndules que són necessàries per a alimentar les nombroses cries. No totes les glàndules secreten la mateixa quantitat de llet; això provoca que els joves es barallin per escollir la millor i que s’estableixi un ordre jeràrquic en què cadascun d’ells s’alleta, sempre, en el mateix mugró. En els primats, en els cetacis i en alguns quiròpters, només es desenvolupen les glàndules situades a la regió pectoral. Els nadissos xuclen la llet, excepte els cetacis, en què existeix una musculatura que permet a la femella dirigir la llet cap a la boca del petit, que no podria estar molt temps sota l’aigua alletant-se i que, així, pot prendre-la de pressa i sortir a la superfície a respirar. Els derivats frontals i parietals poden ésser permanents, com les banyes del bestiar, o renovar-se cada any, com les defenses dels cèrvids; la mida d’aquestes últimes, presents sols en els mascles, ens dona el grau de dominància dels exemplars en els grups i, per tant, la jerarquia d’aquests. El que les té més esplèndides és el cap de colla. Aquestes estructures també permeten determinar, encara que no acuradament, l’edat dels animals.
Altres derivats cutanis són les ungles i els sabots o unglots. També ho són les estructures que permeten el planatge o el vol, en què, encara que hi pot haver un suport ossi, la membrana del vol és formada per pell.
L’esquelet
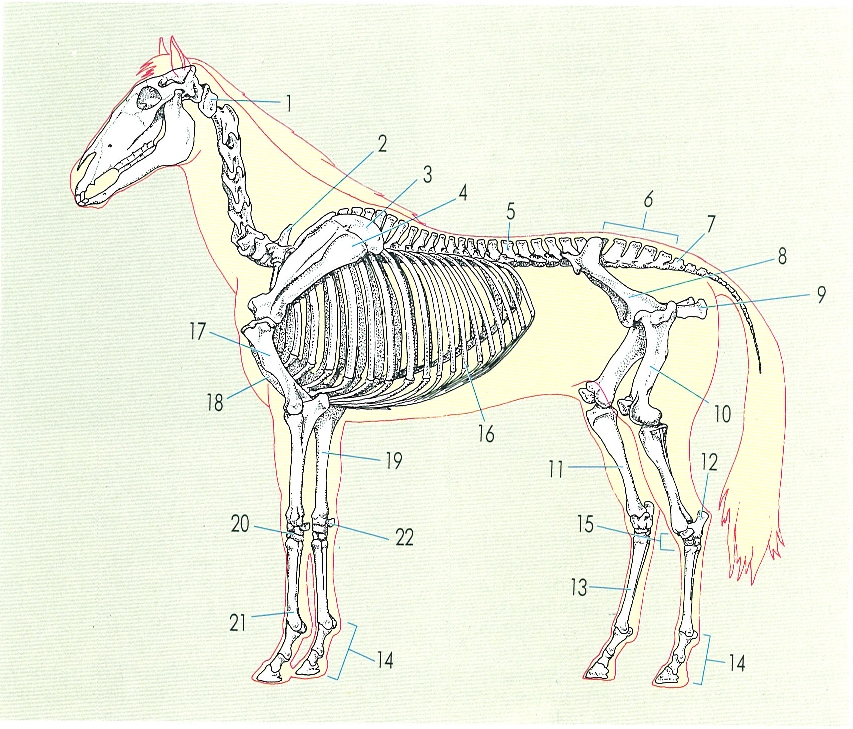
Esquelet d’un mamífer, concretament d’un cavall, que mostra les peces òssies principals. En el dibuix s’observa com el colze es doblega cap enrere i el genoll cap endavant, i també la fortalesa de les extremitats posteriors, responsables de l’impuls cap endavant i del salt. Hom hi ha indicat: 1 atlas, 2 primera vèrtebra toràcica, 3 cartílag de l’escàpula, 4 escàpula, 5 primera vèrtebra lumbar, 6 sacre, 7 primera vèrtebra coccígia, 8 ili, 9 isqui, 10 fèmur, 11 tíbia, 12 calcani, 13 metatars, 14 dit, 15 tars, 16 costella, 17 húmer, 18 cartílag xifoide, 19 radi, 20 carp, 21 metacarp, 22 os accessori del carp.
Ferran Rodríguez.
L’esquelet dels mamífers és molt ben ossificat; presenta, a més, una disminució del nombre d’ossos dèrmics i de substitució que es manifesta clarament en el crani. Un bon exemple d’això el trobem a la mandíbula inferior, formada exclusivament pel dentari. Tant l’estrep, que deriva de la columel·la dels rèptils, com l’enclusa, derivada del quadrat, i el martell, derivat de l’articular, s’incorporen a l’oïda mitjana i no queden altres ossos dels nombrosos que recobrien el cartílag de Meckel, l’eix estructural de la mandíbula inferior.
No és, contra el que se suposa popularment, un esquelet més pesant que el dels ocells; en els ocells el pes es desplaça cap a la part posterior, la qual cosa fa baixar el centre de gravetat i dona estabilitat al vol. Sí que es veritat, encara que sigui poc conegut, que, en determinats grups, com els sirenis, es produeix un fenomen de paquiostosi, és a dir, un augment de gruix i, per tant, del pes dels ossos, que, sens dubte, facilita els moviments dins l’aigua i minva la tendència d’anar cap a la superfície, afavorida per l’especial estructura pulmonar, amb voluminosos alvèols perifèrics; pot ésser que aquesta estructuració faciliti la seva postura normal, que tendeix a mantenir el cap fora de l’aigua.
El crani
El crani s’articula amb l’atlas, la primera vèrtebra cervical, a través de dos còndils. Les vèrtebres, que, pel seu origen, tenen una disposició intersegmentària, presenten els seus centres aplanats, són amfiplanes. Separant les vèrtebres entre elles, hi ha els discos intervertebrals, que tenen, en el seu interior, els cossos polposos, que són restes de notocordi. Les hèrnies discals que es donen en l’home consisteixen en el trencament d’un disc intervertebral, la pèrdua de la massa interna, i la pressió de les arrels nervioses i el dolor consegüent, per disminució de l’alçada del cos intervertebral. Això ens demostra l’existència de restes de notocordi en els mamífers.
Cal considerar el crani a partir del seu origen doble. És constituït per un neurocrani, que protegeix l’encèfal i un esplacnocrani, format en l’evolució a partir de l’esquelet visceral i funcionalment relacionat —en el seu origen— amb les funcions respiratòria i digestiva. El neurocrani té elements ossis d’origen endocondral i d’origen dèrmic.
La columna vertebral
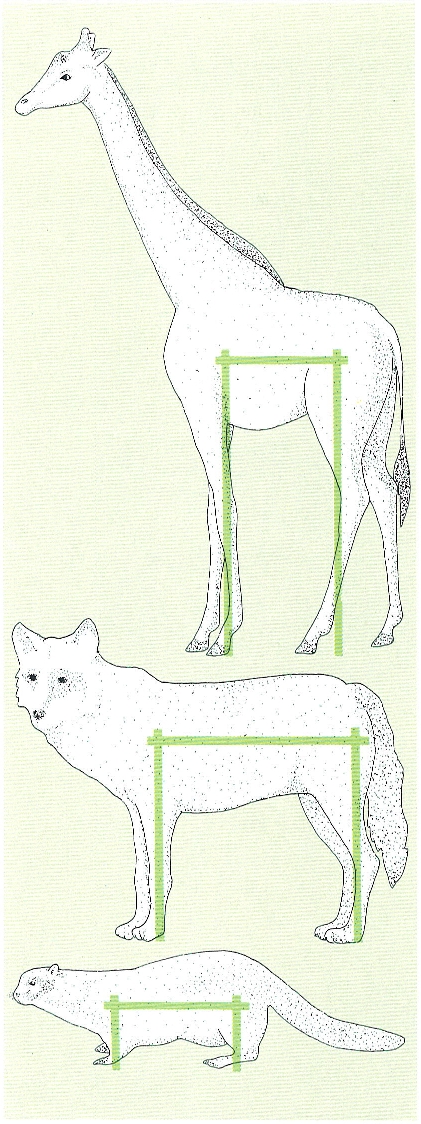
L’anomenada llei del pont regeix l’estructura de l’esquelet dels mamífers: les extremitats llargues obliguen a tenir una columna curta i una columna llarga obliga a tenir les extremitats curtes. El dibuix visualitza aquest principi a través de dos exemples oposats, el de la girafa (a dalt) i el de la fura (a baix), i un d’intermedi, el llop (al mig).
Ferran Rodríguez, a partir de fonts diverses.
El conjunt de les vèrtebres forma la columna vertebral amb la qual s’articulen, a través de cintures, els apèndixs locomotors. Una gran distància entre les cintures obliga al mamífer a tenir unes potes curtes; i, al contrari, unes potes altes obliguen que hi hagi una curta distància entre les cintures. És la denominada llei del pont i el seu compliment i els seus avantatges es veuen clarament comparant la forma d’una fura amb la d’una girafa o la d’un antílop. La fura és allargada, té les potes petites, està perfectament adaptada per a entrar en caus i llançar-se sobre les seves petites preses; la seva agilitat —dit en termes breus— és fabulosa. La girafa i els antílops, amb potes altes, són ràpids i resistents en els seus desplaçaments, però si no calculen bé la distància de fugida són atrapats pels carnívors, pel seu específic depredador. De fet, en això es palesa l’efecte seleccionador del carnívor sobre les poblacions d’herbívors, ja que la distància de fugida és sempre la mateixa i quan l’herbívor fuig més lentament per malaltia o vellesa és quan és depredat pel carnívor.
En els mamífers, el moviment de la columna vertebral és dorsiventral, mai transversal, a diferència de la dels rèptils. En les formes marines, cetacis i pinnípedes, l’impuls natatori és donat pel moviment dorsiventral de la cua, no transversal, a diferència dels peixos o dels rèptils que són o eren aquàtics.
En la regió toràcica, les costelles, en articular-se dorsalment amb les vèrtebres i unir-se ventralment a l’estèrnum, formen la caixa toràcica, que s’ha fet imprescindible pels moviments destinats a la ventilació pulmonar i per la protecció d’altres vísceres. Cap a la regió anterior, es diferencia el coll, format normalment per set vèrtebres cervicals, de mida molt diversa, però iguals en nombre tant en les musaranyes com en el herbívors o en els primats. Aquesta diferenciació permet, a la majoria dels mamífers, acostar la boca a l’aliment, per bé que l’alliberació de la mà permet a alguns marsupials, als rosegadors i als primats d’acostar l’aliment a la boca.
La regió distal de la columna vertebral té un grau de desenvolupament variable i una funcionalitat molt diversa. En les mones aranya, per exemple, serveix d’aparell de fixació en la vida arborícola; en altres espècies, per exemple l’esquirol, de compensador d’equilibri en el salt. En algunes espècies, com l’home, el grau de reducció és molt notable.
Les extremitats i les cintures
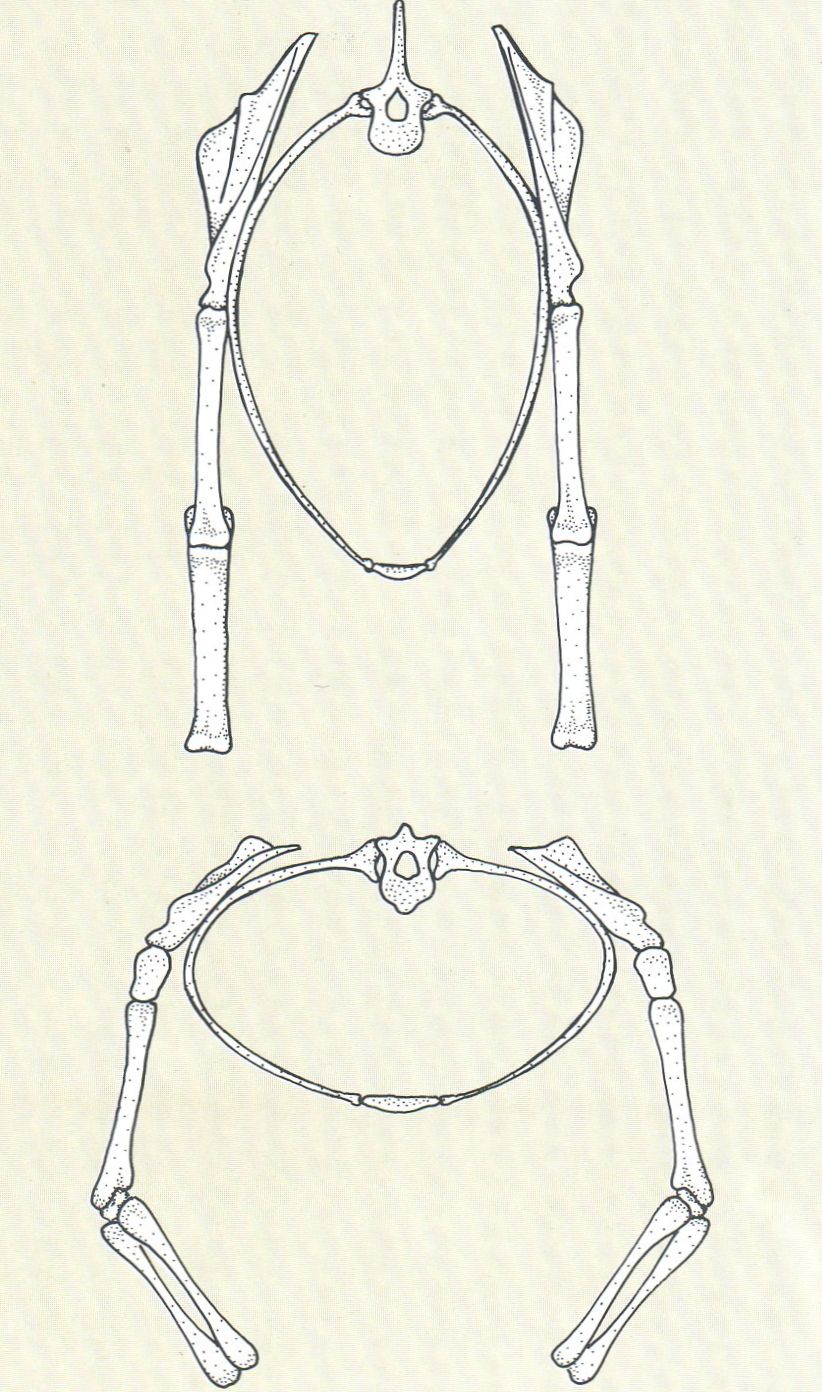
La forma de la caixa toràcica i la disposició de la cintura escapular i de les extremitats locomotrius responen a necessitats mecàniques. En els herbívors (a dalt), la caixa toràcica és allargada en sentit dorsiventral i les extremitats es disposen d’una manera paral·lela. En els primats (a baix), la caixa toràcica s’eixampla transversalment, carácter que va lligat a la braquiació, és a dir, al desplaçament, gràcies a la presència de membres llargs en relació a la vida arborícola.
Ferran Rodríguez, a partir de fonts diverses.
L’extremitat toràcica és formada per la cintura escapular i els tres segments del membre anterior: el braç, l’avantbraç i la mà. La cintura escapular, en els mamífers, consta, genèricament, de tres ossos: escàpula o omòplat, coracoide i clavícula. El coracoide és soldat a l’escàpula i forma en aquesta l’apòfisi coracoïdal.
El grau de desenvolupament de la clavícula és molt variable; és gran en les espècies braquiadores, que es desplacen pels arbres amb les mans, per exemple en els primats; falta completament quan la cintura no és sotmesa a fortes traccions i l’extremitat anterior no té desplaçaments transversals, per exemple en els herbívors. Pel que fa a aquest darrer cas, cal tenir en compte el pes que ha de suportar la cintura, lligada sols muscularment amb la columna vertebral.
La caixa toràcica és allargada en els herbívors; en canvi, en els primats i, especialment, en l’home, s’eixampla. En l’home, a causa de la verticalització, l’húmer es torça; contràriament, per la posició de la cintura, els braços no podrien posar-se en contacte i ens faltaria un dels nostres moviments més característics i útils. Aquest moviment s’ha anat adquirint al llarg de l’evolució; en l’home de Neandertal, la torsió era menor.
La posició bípeda, que allibera la mà i que tanta importància ha tingut en l’evolució humana, provoca una sèrie de transformacions en la columna vertebral i fa que algunes vèrtebres hagin de resistir esforços de tracció molt importants.
L’extremitat pelviana, com la toràcica, consta d’una cintura pelviana i dels tres segments del membre posterior: cuixa, cama i peu. En els mamífers primitius, el suport de l’extremitat es fa a la planta, però la pressió de selecció que actua progressivament en el procés evolutiu fa que l’extremitat s’allargui i que els dits i, finalment, les ungles esdevinguin el suport. Tenim doncs formes plantígrades, digitígrades i ungulígrades; bons exemples d’aquestes són, respectivament, l’ós i l’home —primitiu en aquest aspecte, malgrat la seva intel·ligència—, els fèlids i els herbívors.
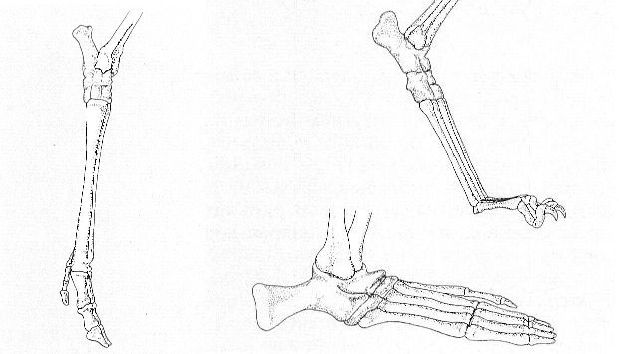
Últim segment, o peu, de l’extremitat posterior en els casos d’un mamífer ungulígrad (a dalt, d’un cérvol), d’un digitígrad (al mig, d’un gos) i d’un plantígrad (a baix, d’un goril·la). Noteu el desplaçament del tars i l’increment de longitud de les extremitats.
Ferran Rodríguez, a partir de fonts diverses.
En els sòls tous, s’hi troben mamífers amb extremitats amples, mentre que en els secs tenen la part distal de l’extremitat estreta. No és d’estranyar que els artiodàctils requereixin prats més tous que els perissodàctils. És coneguda la resistència i el peu ferm de l’ase; ara bé, també hi ha artiodàctils antílops en zones pràcticament desèrtiques, i és cert que els artiodàctils, amb el temps, han anat desplaçant els perissodàctils i, fins i tot, és possible que alguns d’ells (com el cavall) s’haguessin extingit sense la protecció i l’explotació humana, per bé que, en aquest cas, intervenen altres factors com el tub digestiu (l’estómac plurilocular del remugant aprofita molt més la digestió de la cel·lulosa que no el gran cec intestinal dels perissodàctils).
L’extremitat pentadàctila fa menys eficaç l’impuls que una extremitat didàctila o monodàctila, en què l’impuls és més aprofitat. Sempre que hi ha una tendència a caminar, hi ha també una reducció en el nombre de dits; en l’home, s’observa la reducció dels dits externs del peu. Aquest fenomen no es dona només en els mamífers: només cal observar la reducció dels dits en els estruços.
La musculatura
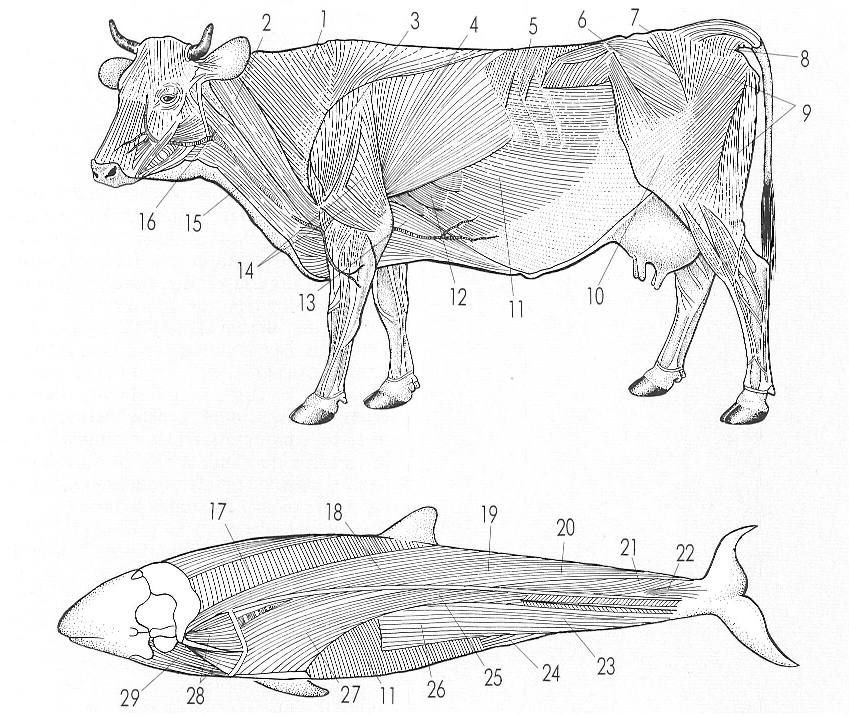
La musculatura apendicular s’imposa, en l’evolució dels vertebrats, sobre la musculatura segmentària, en relació amb les seves funcions principals: de sosteniment i de moviment. Els dibuixos il·lustren el paral·lelisme que hi ha entre la musculatura de dos mamífers de tipus de vida ben diferent, un remugant (a dalt) i un cetaci odontocet (a baix), de vida aquàtica; en aquest, la musculatura és més semblant a la d’un mamífer terrestre que no a la d’un peix; els músculs de la cua es formen a partir dels del tronc 1 trapezi, 2 cleidomastoïdeu, 3 deltoide, 4 dorsal ample, 5 «serrato dorsal», 6 tensor de la «fascia lata», 7 gluti mitjà, 8 coccigi, 9 bíceps femoral, 10 «fascia lata», 11 oblic abdominal extern, 12 «serrato ventral», 13 pectoral profund posterior, 14 cap del tríceps, 15 esternocefàlic, 16 cleidooccipital, 17 espiral, 18 dorsal llarg, 19 aponeurosi dorsal, 20 extensor caudal mitjà, 21 extensor caudal lateral (part mitjana), 22 extensor caudal lateral (part lateral), 23 flexors caudals, 24 elevador anal, 25 iliocostal lumbar, 26 hipaxial lumbar, 27 ilicostal dorsal, 28 escalè, 29 estilohioïdeu.
Amadeu Blasco, a partir de fonts diverses.
La musculatura acostuma a representar entre un 30 i un 50 % del volum total del cos. Al voltant del 80 % de la seva energia és dissipada en forma de calor que, a través de la sang, va circulant per les diferents parts del cos i ajuda, així, a mantenir la temperatura corporal.
La disposició segmentària, en miòtoms, que és tan evident en els vertebrats inferiors, és difícilment observable en els mamífers, en què predomina la musculatura apendicular. La musculatura paraaxial, derivada dels metàmers, es manté segmentària; passa, formant un pont, d’un cos vertebral a l’altre, la qual cosa dona mobilitat a la columna vertebral. L’evolució dels segments és encara clara en el tòrax. Els miòtoms s’estenen en sentit dorsiventral i, en ells, es distingeix l’epímer, en posició dorsal, que origina la musculatura extensora, i l’hipòmer, que dona lloc als músculs flexors laterals i ventrals. Els extrems anteriors dels hipòmers creixen en direcció ventral i els d’un segment se solden amb els del següent formant el recte abdominal, que reflecteix, clarament, el seu origen segmentari. En els mamífers, de caixa toràcica ben estructurada, sol estar ben desenvolupat i entapissa la cavitat abdominal; en canvi, en els urodels es troba al llarg de tota la regió ventral.
En general, i com en tots els vertebrats, la musculatura estriada que s’insereix, gràcies al teixit connectiu, entre les diverses peces òssies, és la responsable del moviment d’aquestes i, per tant, de l’individu. Ara bé, no sempre és així, com, per exemple, en la musculatura cutània, que permet el moviment de la pell (qui no ha vist l’expressió d’una cara arrugant el front o a un cavall traient-se les mosques de sobre?). La musculatura de l’ull permet el moviment dels globus oculars en totes les direccions de l’espai i és originada, com es veu seguint el seu desenvolupament embrionari, a partir de diferents segments. En el seu dia, aquest fet fou un argument de pes a favor de la teoria sobre l’origen segmentari del crani.
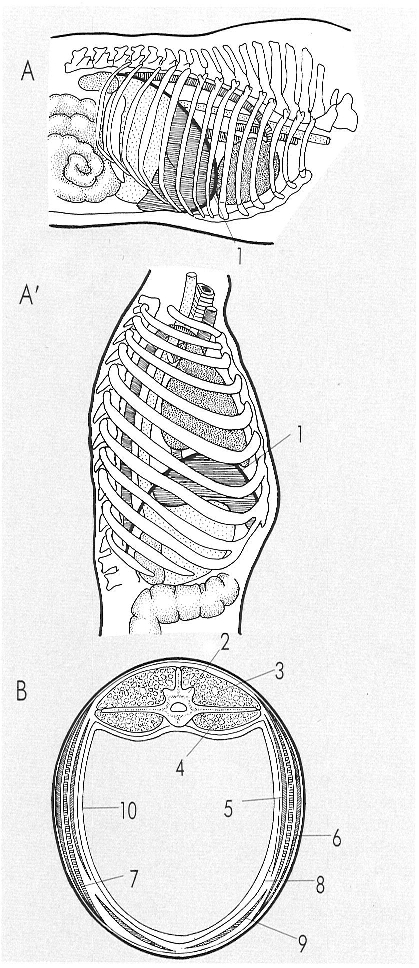
El diafragma és el múscul que separa la cavitat toràcica de l’abdominal (vegeu A i A’); és important en la mecànica respiratòria i és innervat pel nervi frènic, cosa que constata que el seu origen, en els mamífers, és en relació amb la musculatura del coll. En l’esquema B, corresponent a la secció transversal de la regió lumbar d’un cavall, veiem els principals músculs que envolten la cavitat abdominal. Hom ha indicat en el dibuix: 1 diafragma, 2 vèrtebra, 3 «longissimus dorsi», 4 musculatura sublumbar, 5 múscul transvers abdominal, 6 múscul oblic abdominal extern, 7 múscul abdominal intern, 8 «fascia transversa», 9 múscul recte abdominal, 10 peritoneu.
Amadeu Blasco, a partir de fonts diverses.
Un múscul especialment característic és el diafragma, que separa, en els mamífers, la cavitat toràcica de l’abdominal. Se suporta en l’apèndix xifoide, en els cartílags costals, en les costelles i en les vèrtebres. Intervé en el mecanisme respiratori i és inervat pel nervi frènic, la qual cosa ens demostra que s’originà a partir de la musculatura del coll i que és anàleg i no homòleg del diafragma dels ocells, en els quals s’origina a partir de la musculatura costal.
La musculatura branquiomèrica, d’origen branquial, és inervada per nervis cranials i, com és lògic, relacionada amb el crani visceral. La majoria dels músculs són funcionalment responsables de la incorporació d’aliment o estan lligats a fenòmens respiratoris. El pas rèptil-mamífer es defineix per les modificacions que es donen a la mandíbula; en el sistema muscular, aquestes modificacions es concreten en la incorporació a l’oïda del múscul tensor del timpà i de l’estapedial. Altres músculs, com el trapezi i l’esternocleidomastoïdeu, són d’origen branquial.
S’observen modificacions especials de la musculatura en els rats-penats, els talps, les balenes i els dofins, relacionades amb el seu especial funcionalisme. Una estructura molt particular és la trompa dels elefants (formada, principalment, per músculs que, normalment, envolten els conductes nasals i el llavi bucal superior) i la musculatura que permet la mobilitat del morro de la rata mesquera.
Els sistemes de relació i control
El sistema nerviós
El sistema nerviós es troba molt ben desenvolupat, sobretot els hemisferis cerebrals, que, en algunes espècies, presenten gran nombre de circumvolucions. Això fa que la perspicàcia, l’hàbit inquisitiu i, en definitiva, la vida intel·ligent adquireixin una importància extraordinària. Aquest alt grau de desenvolupament dels lòbuls cerebrals és un fet molt important però tampoc tan rar, ja que, en altres nivells de l’escala vertebrada, s’ha donat en altres regions del sistema nerviós (n’és un bon exemple l’enorme cerebel que presenten els peixos mormiriformes, que, comparativament, respecte a la totalitat de l’encèfal, és més gran que el cervell de l’home respecte al seu encèfal). Però no sols augmenta el volum sinó que, en molts casos, també s’incrementa la superfície gràcies a la presència d’una sèrie de circumvolucions. La capa externa de la regió cortical és formada per substància grisa.
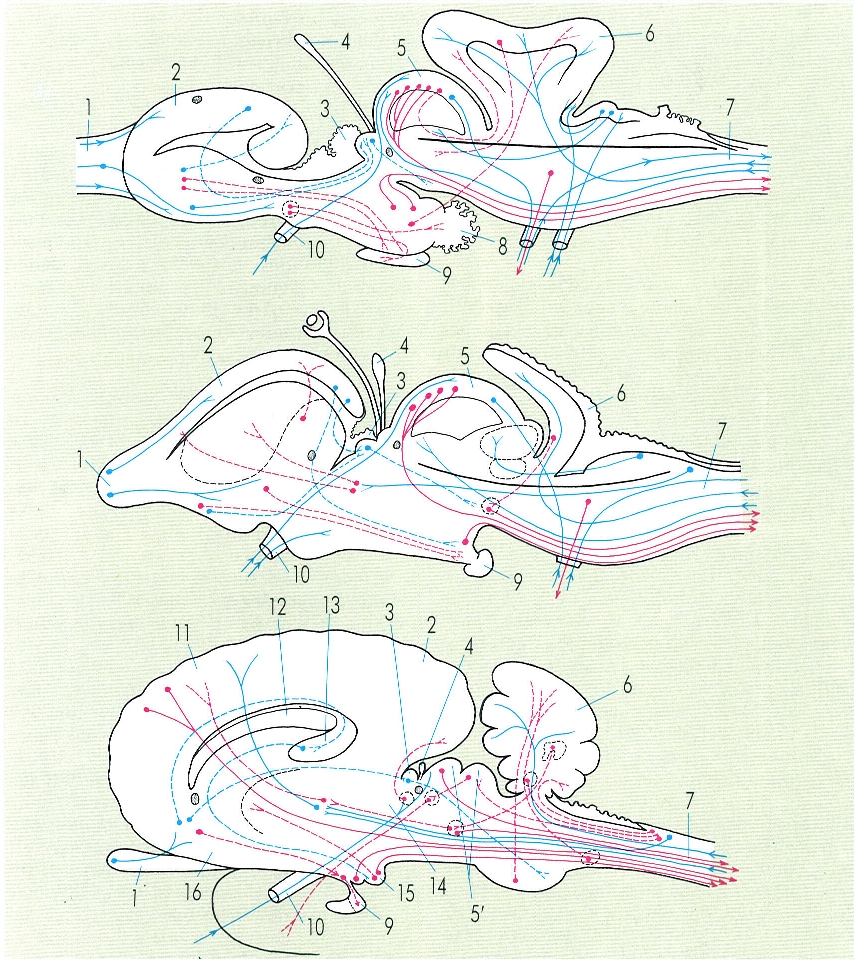
Esquema de les principals connexions de l’eix neural d’un peix cartilaginós (a dalt), d’un rèptil (al mig) i d’un mamífer (a baix), on s’observa la diferent importància que hi atenyen cadascuna de les parts de l’encèfal. En els mamífers s’observa un gran desenvolupament del cervell, la reducció de l’epífisi, la incorporació dels «corpora postica» als tubercles bigemins (que ja s’observa en determinats rèptils) per formar els tubercles quadrigemins i el gran desenvolupament del cerebel. Estudis recents efectuats amb metodologia bioquímica han demostrat la formació de neurones en els rosegadors adults, cosa que fins ara no s’ha estudiat en altres grups de mamífers, l’home inclòs. Hom ha indicat en el dibuix: 1 bulb olfactori, 2 hemisferis cerebrals, 3 habènul·la, 4 epífisi, 5, 5’ tubercles bigemins i quadrigemins, respectivament, 6 cerebel, 7 bulb, 8 sac vascular, 9 hipòfisi, 10 fibres òptiques, 11 neopal·li, 12 cos callós, 13 hipocamp, 14 tàlem, 15 cossos mamil·lars, 16 paleopal·li.
Gustavo Hormiga, a partir de fonts diverses.
Els respectius hemisferis cerebrals són units entre ells en els placentaris —però no en els monotremes ni en els marsupials— per una ampla comissura blanca, el cos callós. Els lòbuls olfactius són petits, si se’ls compara amb els dels vertebrats inferiors, encara que, en determinats grups, com en els èquids, la seva mida segueix essent important en relació a la seva capacitat d’olfacció (un semental és capaç de reconèixer una euga en zel a una vintena de quilòmetres de distància). El diencèfal consta d’un epitàlem dorsal, un tàlem lateral i un hipotàlem ventral. En el sostre del diencèfal, hi ha també una glàndula pineal relacionada filèticament amb un òrgan fotosensible, fet que justifica que, actualment, conservi funcions de rellotge intern. L’hipotàlem consta de quatre parts: l’infundíbul, que forma —en especial— el lòbul posterior de la hipòfisi, el quiasma òptic, on es creuen els nervis òptics, el "tuber cinereum", que sobresurt de la base del quart ventricle, i els cossos mamil·lars; els nuclis hipotalèmics s’encarreguen del control visceral, per exemple de la regulació del balanç d’aigua, de la pressió arterial, de la son i de la temperatura corporal. El mesencèfal és completament recobert pels hemisferis cerebrals i pel cerebel i, per tant, proporcionalment, és més petit que el d’altres vertebrats; a la part dorsal, s’hi diferencien els tubercles quadrigemins: els tubercles anteriors, relacionats amb la visió, i els posteriors, amb l’audició.
El cerebel, relacionat amb la coordinació dels moviments i el manteniment de l’equilibri corporal, és ben desenvolupat en els mamífers. Presenta sempre dos hemisferis laterals i una part mitjana, el vermis, que és la més primitiva, el paleocerebel; sota el cerebel, hi ha la protubèrancia anular o pont de Varoli, i la part posterior és la regió bulbar, que continua directament amb la medul·la espinal i és el lloc a través del qual passen les línies cerebrospinals.
En els mamífers, hi ha una clara tendència a l’escurçament de la medul·la espinal i, en algunes espècies s’estén fins al sacre. A partir d’aquest, s’estén el que es denomina "phyllum terminale", que no adquireix diferenciació nerviosa. En l’home, la longitud de la medul·la sol ésser de 45 cm. La columna vertebral, per diferència en el creixement, arriba, a uns 75 cm. Per això, en l’adult, la medul·la arriba només, fins entre la primera i segona vèrtebra lumbars.
S’ha parlat molt de la importància pel que fa a la intel·ligència del pes de l’encèfal. La té, però també s’ha de tenir en compte el pes del cos. Aquesta relació no pot ésser una senzilla fracció en què el numerador sigui el pes de l’encèfal i el denominador el pes del cos, ja que es primarien les espècies de mida petita; si això fos així, un petit tití tindria un coeficient superior a l’humà; aquesta relació és d’1:560 en l’elefant índic, en l’home d’1:45, en el tití d’1:26 i en la mona aranya d’1:15. Aplicant la relació E (pes encèfal)/ P0.56 (pes cos) s’obtenen els següents valors: home 2,8, mones superiors 0,7-0,8, mones 0,3-0,4, fèlids 0,37, conill 0,18, rates i ratolins 0,06-0,09.
El sistema endocrí
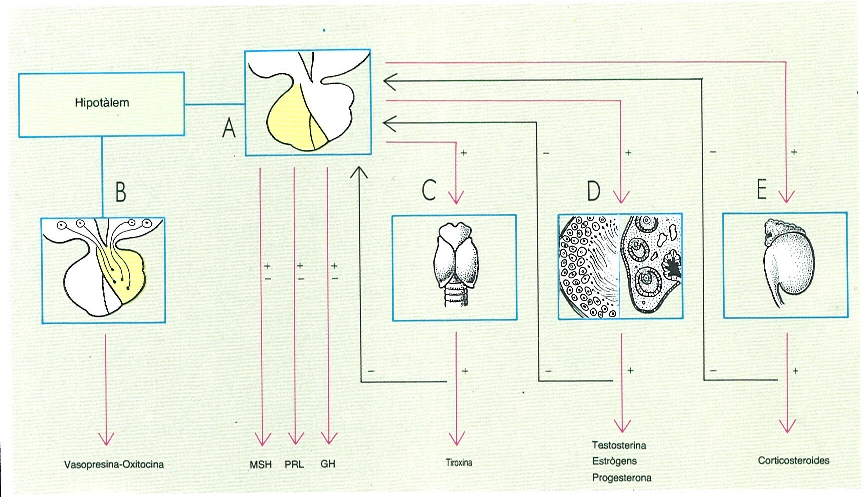
Esquema de la regulació hormonal en els mamífers, que mostra la importància de l’hipotàlem i la hipòfisi. Hom ha indicat en el dibuix la relació funcional entre l’hipotàlem, l’adenohipòfisi (A), la neurohipòfisi (B), la glàndula tiroide (C), les gònades (D) i l’escorça adrenal (E). Hom ha indicat, també, abreujadament, l’hormona estimulant dels melanòcits (MSH), la prolactina (PRL) i l’hormona de creixement (GH).
Gustavo Hormiga, a partir de fonts diverses.
Les glàndules que constitueixen el sistema endocrí no presenten conductes excretors per a eliminar els productes que elaboren, les hormones: aquestes substàncies van directament a la sang o a la limfa, on són vehiculitzades vers les diferents parts del cos. Juntament amb el sistema nerviós, aquest sistema és responsable de la coordinació funcional dels organismes. Algunes glàndules tenen una funció mixta. El pàncreas és, ensems, glàndula de secreció externa i interna, i les glàndules productores d’espermatozoides i d’òvuls tenen, a més, una funció endocrina.
La glàndula tiroide, homòloga de certa part de l’endostil dels procordats, presenta en els mamífers dos lòbuls units per un estret istme que passa per la cara ventral de la tràquea. Es forma a l’embrió com a diverticle ventral mitjà de la faringe entre el primer i segon sac faringi. La presència de goll (hiperplàsia de la glàndula tiroide) no sols es dona en els humans sinó també en altres espècies animals, almenys en les domèstiques, lligat moltes vegades a la ingestió de brasicina, molt abundant a la col.
Les glàndules paratiroides deriven dels extrems dorsals dels sacs faringis tercer i quart; n’hi ha dues a cada costat i estan més o menys dintre de la massa tiroide. Regulen la quantitat de calci i fòsfor de la sang i la seva extirpació o, fins i tot, lesió, provoca tetània i, en pocs dies, la mort.
Les glàndules adrenals o suprarenals són cossos compactes situats a la massa de teixit adipós que existeix a l’extrem cefàlic del ronyó. Presenten una part cortical derivada del mesoblast i una part medul·lar derivada de l’ectoblast. La medul·la segrega adrenalina o epinefrina elevadora de la tensió sanguínia, dels batecs del cor i del nivell de glucèmia i és responsable de la dilatació de les vies respiratòries; en definitiva, prepara l’organisme per a la lluita. La regió cortical segrega esteroides amb propietats hormonals, els adrenocorticoides, que intervenen en la regulació de l’equilibri sodi/potassi, el metabolisme glucídic i proteic, i d’altres relacionades amb hormones sexuals i hipofisàries.
Com a glàndula endocrina, el pàncreas està limitat en els illots de Langerhans, que presenten cèl·lules que segreguen glucagó, d’acció hiperglucemiant, i cèl·lules β, que segreguen insulina, d’acció hipoglucemiant.
Els testicles, les gònades masculines, a més de produir espermatozoides, són glàndules endocrines responsables, amb la seva secreció, de l’aparició dels caràcters sexuals secundaris dels mascles. El desenvolupament i activitat normal dels òrgans sexuals accesoris són controlats per la testosterona, l’androgen natural més potent, produït per les cèl·lules de Leydig, que se situen als intersticis existents entre els tubs seminífers. En els casos de criptorquídia, els tubs seminífers degenerats no formen espermatozoides, però les cèl·lules intersticials segreguen testosterona, la qual cosa fa que els caràcters sexuals secundaris dels individus criptorquídics corresponguin als d’un individu normal.
Els ovaris, les gònades femenines, a més de produir òvuls, són glàndules endocrines que segreguen estrògens responsables dels caràcters sexuals secundaris de les femelles, progesterona i relaxina. L’extirpació dels ovaris a eugues i conills d’índies durant l’última fase de gestació no provoca l’avortament del fetus i cal interpretar que en aquests animals, i segurament en altres, la placenta pot substituir els ovaris en la secreció de progesterona.
La hipòfisi situada a la sella turca de l’esfenoide és unida al diencèfal per un pedicle; consta de dues parts principals, el lòbul anterior (adenohipòfisi), evaginació del sostre de la boca primitiva, i el lòbul posterior (neurohipòfisi), que deriva del terra del diencèfal. El lòbul intermedi presenta vesícules plenes de substància col·loide. És coneguda també com glàndula pituïtària i les seves secrecions són d’importància vital per al creixement, la maduració i la reproducció. Dirigeix el funcionament de moltes altres glàndules de secreció i, al mateix temps, és influïda per elles: els impulsos nerviosos s’hi transformen en hormonals i les hormones actuen, també, sobre centres nerviosos.
La placenta també és una important estructura endocrina; hi ha proves que indiquen que és l’origen de l’estrogen extraovàric, de progesterona i d’hormones gonadotropes. El còrion placentari elabora la coneguda gonadotropina coriònica. Han estat detectades altres hormones a la placenta de determinades espècies, la qual cosa vol dir que un estudi més profund permetria de detectar-les segurament en la majoria, sinó en tots, dels mamífers. La placenta, doncs, té, a més del seu paper principal de relació entre la mare i el fetus, el de secreció d’hormones.
Els òrgans dels sentits

Els òrgans dels sentits dels mamífers són ben desenvolupats i amb adaptacions concretes als seus tipus de vida i hàbitat. No sols permeten rebre informació del medi sinó també establir una relació familiar i social que posa de manifest l’alt psiquisme assolit pels mamífers. A la fotografia veiem un conill amb els pavellons de l’orella en posició d’escoltar.
Xavier Parellada.
En tots els vertebrats, la consciència de la situació de l’individu en l’espai ve donada pel sistema nerviós central. Ara bé, les condicions del medi, les informacions respecte a l’entorn, són donades pels òrgans dels sentits. Aquests es troben fonamentalment en el cap, perquè així la informació és donada a l’encèfal al més ràpidament possible i perquè el cap és la zona de l’individu que es troba més aviat en un nou medi.
Els lòbuls olfactius no són tan grans com en els vertebrats inferiors, però tenen gran superfície. El sentit de l’olfacte és molt ben desenvolupat en quasi tots els mamífers. L’utilitzen per buscar aliment, situar l’enemic i reconèixer el sexe. Un egüer és capaç de detectar una euga en zel a quilòmetres de distància. El món de les olors és difícil de comprendre per l’home, ja que tenim molt limitat l’òrgan olfactiu, però hem d’acceptar l’existència d’un relleu olorós. En els mamífers adaptats al medi aquàtic, aquest sentit deixa d’ésser funcional.
A l’ull dels mamífers, no s’observa cap estructura especial. Pel que fa a la precisió de la imatge, s’ha d’acceptar que l’ull dels ocells és més perfecte que el dels mamífers i això es vàlid, fins i tot, per als primats, en què el sentit de l’olfacte s’ha reduït força i, en canvi, s’ha desenvolupat més la visió. Per això, els primats han pogut reduir el morro, la qual cosa els permet de presentar un aspecte menys bestial i han desplaçat els ulls a una posició frontal, que els permet una bona visió en relleu. En la vida arborícola dels primats, és de vital importància poder calcular la resistència de les branques i perd interès l’olfacte, ja que la seva alimentació, genèricament parlant, és frugívora i insectívora. Certes espècies s’han adaptat a la visió nocturna; moltes presenten grans vibrisses en el morro, sobre els ulls, que es posen en contacte amb els objectes quan de nit l’animal avança progressivament, de manera que la informació que reben aquestes espècies és doble: visual i tàctil. Altres espècies, com els cetacis, presenten una visió relativament desenvolupada, encara que les que viuen en aigües tèrboles, com el dofí del Plata o del Ganges, tenen els ulls petits; la principal font d’informació dels cetacis és l’acústica. Les espècies molt adaptades a excavar, com els talps, tenen els ulls petits.
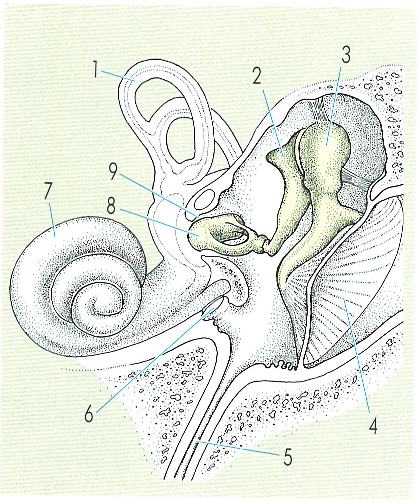
Esquema del sistema auditiu dels mamífers. L’oïda interna és un òrgan d’audició i d’equilibri. El pas de la vida aquàtica a la vida terrestre va exigir la presència d’un sistema que permetés la transmissió de l’estímul des d’un medi aeri a un medi aquàtic, intern; així aparegué l’oïda mitjana. Hom ha indicat en el dibuix les parts següents: 1 conducte semicircular, 2 enclusa, 3 martell, 4 membrana timpànica, 5 trompa d’Eustaqui, 6 finestra rodona, 7 caragol o còclea, 8 estrep, 9 finestra oval.
Amadeu Blasco, a partir de fonts diverses.
L’oïda ha millorat anatòmicament i funcionalment respecte als rèptils i als ocells. El cargol ha augmentat en longitud, s’han incorporat dos ossos més a la cadena d’ossets que transmeten el so de la membrana timpànica a l’oïda interna i ha aparegut un pavelló de l’orella ampli i, normalment, orientable a voluntat, que dirigeix les ones sonores al conducte auditiu.
Els quiròpters, els cetacis i els pinnípedes, depenen de la captació del so que ells mateixos produeixen per descobrir objectes en el medi en què es mouen. Els rats-penats descobreixen així els objectes que s’interposen en el seu vol o els insectes de què s’alimenten. Els cetacis, almenys els odontocets, es dirigeixen per ecolocalització; s’han fet experiments que han demostrat que amb els ulls tapats poden evitar obstacles i localitzar petits objectes amb gran precisió. Els pinnípedes localitzen objectes a les fosques, amb gran rapidesa i són capaços de diferenciar-los de forma, mida i composició.
S’ha treballat molt en l’estudi de la parla entre les bèsties salvatges, però no fa massa que es comprèn la natura i el paper dels mitjans d’intercomunicació utilitzats pels mamífers. Hi ha una sèrie de senyals, no solament acústics sinó també visuals i olfactius, que desencadenen en l’individu que els rep una reacció característica. Les ovelles reconeixen per l’olfacte quin és el seu anyell. Els pastors, quan volen fer alletar un anyell que ha perdut la seva mare per una altra mare que ha perdut el seu anyell, treuen la pell d’aquest últim i la col·loquen sobre el primer; només així la femella supervivent accepta d’alletar l’altre anyell. També són conegudes les relacions de tipus acústic. Són típics els crits d’alarma donats per un individu i que tenen resposta per part de tota la població. El crit d’alarma és més o menys intens segons el tipus de perill; això ha estat ben estudiat en Citellus. En certes mones s’ha comprovat que tenen un vocabulari amb un important grau de vocalitzacions, que tenen el seu paper en el comportament de l’espècie. En el gibó, s’han descrit nou tipus de vocalitzacions que fan el seu paper en la coordinació del grup social. En el grup dels cetacis, la veu es diversifica extraordinàriament. Capaços de produir gran nombre de sons, es relacionen a través de la termoclina. Les seves veus poden ésser tan diverses que la beluga ha rebut la denominació de canari de mar. Hi ha proves que alguns misticets utilitzen l’ecolocalització per nedar prop de la costa. Les ones que emeten, les reben a través de les mandíbules. Els odontocets les reben a través del meló, estructura en forma de lent que tenen en el front; tenen un món sònic molt important que, a més de permetre’ls relacionar-se entre ells, els dona idea del relleu existent.
Els senyals olfactius són molt importants en els mamífers, encara que són mal compresos per l’home que té, com ja hem dit, un sentit de l’olfacte molt reduït. L’existència de nombroses glàndules productores de secrecions olfactives ja ens dona idea de l’extensió del fenomen. Les secrecions de les glàndules pèdies, carpianes, tarsals i metatarsals, preorbitals, inguinals, retrocornals, occipitals, caudals i prepucials dels remugants serveixen per a relacionar els sexes o bé per a marcar el territori; és per això que veiem a molts d’aquests animals en jardins zoològics refregant-se en els tancats i arbres a l’època de zel. Les glàndules anals són molt típiques dels carnívors i els donen una olor característica. Els gossos, quan orinen amb freqüència, no fan sinó anar marcant un territori. Les defecacions de les martes sobre les pedres són unes altres marques territorials.
Els senyals visuals han estat menys estudiats en els mamífers que en els ocells, ja que són menys espectaculars que en aquests. Però, indubtablement, existeixen i es troben descrits en gran nombre d’estudis de jerarquia social, en què es veu que aquesta es manté més per una sèrie d’actituds característiques que per autèntiques lluites. La coloració de la cara dels mandrils o les característiques manifestacions facials dels llops i, fins i tot, la posició del pèl en el cos, en són bons exemples.
L’aparell digestiu
Com a tots els grups, el seu origen és fonamentalment endoblàstic, tot i que la regió proximal —estomodeu— i la distal —proctodeu— són d’origen ectoblàstic. En el tub digestiu, s’han de diferenciar dues porcions: un intestí cefàlic, format per la boca i la faringe, amb els seus derivats: glàndules salivals, tiroide, tim i paratiroide; i un intestí troncal, format per la porció del tub digestiu i els seus annexos situats caudalment respecte l’esbós pulmonar. Embriològicament, el més correcte és diferenciar un intestí anterior i un de posterior, en relació al conducte onfaloentèric. Ara bé, gràcies a l’aparició de dos esfínters, el pilòric i l’iliocecal, s’hi diferencien tres parts: anterior, mitjana i posterior. La primera part comprèn l’esòfag i l’estómac; la segona, compresa entre els dos esfínters, és el budell prim amb el fetge i el pàncreas, derivats també endodèrmics, i la tercera és el budell gros, que s’estén des de l’esfínter iliocecal fins a l’anus. En l’embrió, hi ha un punt en què ectoblast i endoblast contacten; és la membrana faríngia, que, al final del període de segmentació, desapareixerà. Externament a ella i amb la col·laboració del mesoblast es formarà la boca. El mesoblast dividirà la cavitat de la boca primitiva en dues parts: una d’inferior, o boca definitiva, i una altra de superior, dividida en dos per un septe sagital i mitjà que es transforma en les fosses nasals. Això sols es dona en els mamífers i parcialment en els ocells i crocodilians, que tenen paladar secundari. En el paladar dels mamífers, hi ha una sèrie de rugositats més fortes que, en el cas dels carnívors, serveix per a fixar les preses. Sembla que les barbes de les balenes són homòlogues d’aquestes rugositats.
Les genives, l’esmalt de les dents i la mucosa de les galtes són també d’origen ectoblàstic. Les dents d’origen ectoblàstic i mesoblàstic diferencien dues parts: una de central, la boca pròpiament dita, i una altra de perifèrica, excèntrica, el vestíbul oral. Les galtes, els llavis i el septe submandibular acaben de limitar la boca.
La dentició
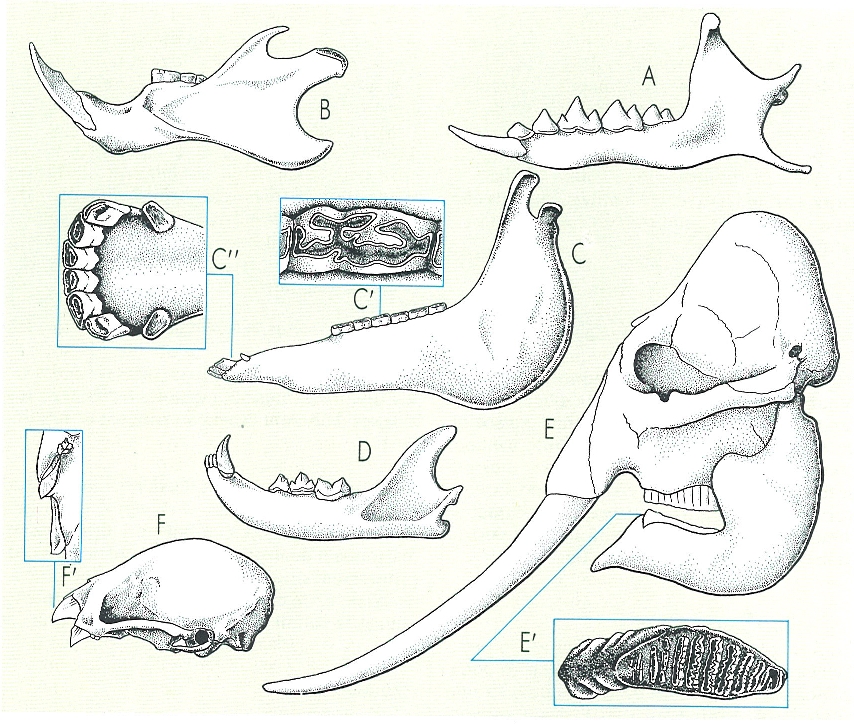
Diferents models de dentició en els mamífers. En la dentició dels insectívors (A, corresponent a la musaranya comuna, Crocidura russula) s’observa el caràcter primitiu en les molars tricodontes. En la d’un rosegador (B, corresponent a la rata negra, Rattus rattus) observem el diastema característic d’aquest grup i també d’altres, com els lagomorfs i els perissodàctils (cas del cavall, Equus caballus, que s’il·lustra en C, C’, C"). Els carnívors (D, corresponent al gat salvatge, Felis silvestris) presenten una clara reducció de les molars. L’especialització s’observa en les plaques trituradores de les molars de l’elefant (E, E’, Elephas indicus) o en la dentició tallant del vampir Desmodus rotundus (F, F’).
Gustavo Hormiga, a partir de fonts diverses.
Les dents adquireixen una diversitat de formes extraordinària, encara que, en algunes espècies, com ara les foques, el dofins o els catxalots, s’han fet totes iguals (isodòncia). El nombre de dents i la forma que presenten caracteritzen l’espècie; per aquest motiu, en mastozoologia es fa referència constant a aquests dos caràcters i s’elaboren les denominades fórmules dentàries. La fórmula completa de la dentició humana adulta és:
(M3) M2 M1 P2 P1 C I2 I1 ║ I1 I2 C P1 P2 M1 M2 (M3) / (M3) M2 M1 P2 P1 C I2 I1 ║ I1 I2 C P1 P2 M1 M2(M3)
Com sigui que, normalment, la boca és simètrica quant al nombre de peces i que la posició de les peces és sempre la mateixa, la fórmula es pot simplificar de diverses maneres: 2/2 1/1 2/2 3/3, I2, C1, P2, M3. Quan hom vol referir-se a una determinada dent es posa la inicial amb un exponent o subíndex. En el primer cas, es refereix a una dent de la mandíbula superior, en el segon, de la inferior. Les peces de la dentició de llet s’escriuen en minúscules, mentre que les definitives són escrites en majúscules. Les peces carnívores són la p3 i p4 de llet i les P4M1 definitives. En l’oclusió, les dents de la mandíbula superior se situen cap enfora i en la inferior cap endins. Així encaixen; en el recanvi, les dents superiors surten cap endavant i cap al costat, i les inferiors enrere i endins.
En l’ornitorinc, les plaques còrnies del bec substitueixen les peces transitòries. Els equidnes tenen un bec tubular i han perdut totes les dents. Els marsupials ofereixen una gran diversitat de dents segons el seu règim alimentari i les seves nombroses adaptacions responsables del seu paral·lelisme amb els placentaris. No canvien les dents de la part de davant de la boca i aquestes apareixen tard; això és una adaptació de l’animal per a poder estar fix al mugró matern, ja que les dents el lesionarien.
En els placentaris, els insectívors tenen unes dents primitives i punxegudes en relació al tipus alimentari. En els rats-penats, o no apareix dentició de llet o aquesta és guarnida de nombroses puntes que permeten al petit de fixar-se als pèls materns. Els primers s’acosten l’aliment amb la mà, que substitueix, en aquesta tasca, les mandíbules i les dents anteriors; com a conseqüència d’aquesta adaptació, s’ha reduït la longitud de les mandíbules i del coll. Quan el règim és insectívor, com en els óssos formiguers i els pangolins, no hi ha dents, sinó unes estructures especials en l’estómac per a triturar els insectes. Els lagomorfs, conills i llebres, presenten dues incisives a cada hemimandíbula superior, disposades anteroposteriorment. En els rosegadors, hi ha una tendència a la monofiodòncia, amb atròfia de la dentició de llet i un espai —diastema— entre incisives i premolars que també es dona en altres grups, com els mateixos logomorfs, i que els serveix per a passar l’aliment de les galtes a la boca i a l’inrevés. A les galtes presenten una carnositat que tapa el diastema. Els carnívors tenen una dentició heterodonta molt marcada, amb tendència a la reducció de les molars; en el cas extrem dels fèlids, les peces carnisseres, especialitzades a tallar tendrums, són, a la dentició definitiva, la premolar superior i la molar inferior. Entre els cetacis, en les balenes, els esbossos dentaris es recobreixen a la vida fetal; s’alimenten gràcies al filtre format per les barbes i a l’acció de pistó de la llengua. Els dofins presenten una dentició molt homogènia i en els catxalots s’atrofia la de la mandíbula superior. En els elefants, les incisives superiors es transformen en defenses i les premolars i molars se substitueixen en sentit horitzontal; només en tenen, simultàniament, una de funcional. L’última molar ja és desgastada entre els cinquanta i els seixanta anys i els animals d’aquesta edat sols es poden alimentar d’herba tendra. Mal alimentats i desnodrits, tenen un comportament agressiu i acaben per morir en aquelles àrees on es troba aliment fresc tot l’any. D’aquí que les millors peces de vori, que corresponen als mascles més vells, es trobin sols en determinades àrees conegudes com a cementiris d’elefants. Els perissodàctils presenten dents altes, premolars i molars molt iguals i tenen, en les incisives, unes marques i contramarques que desapareixen per desgastament amb el temps. Per aquest motiu, la seva observació permet de determinar l’edat de l’animal. Els artiodàctils tenen les dents més baixes; entre ells, hi ha diverses famílies, com els cèrvids i els bòvids, que perden les incisives de la mandíbula superior.
El tub digestiu
A la boca, la llengua, sempre molt mòbil, és coberta d’estructures gustatives i presenta, de vegades, una forma molt diversa. En els carnívors, és coberta de denticles que li permeten de netejar els ossos dels últims trossos de carn. En els formiguers, és extraordinàriament llarga i els permet de seleccionar els insectes de què s’alimenten. En alguns carnívors, com els gossos, l’evaporació que es dona a la llengua intervé en fenòmens de termoregulació. En tots, té una importància extraordinària en el fenomen de la deglució, ja que acompanyen l’aliment contra el paladar.
La faringe, la cavitat musculosomembranosa que és situada rere les fosses nasals i la boca, presenta una sèrie de transformacions limfoepitelials que s’estenen a cada costat, des de la base de la llengua fins al fòrnix, la part més alta de la faringe. Per ella passa tant l’aire com l’aliment. L’entrada d’aire a la tràquea és regulada per l’epiglotis, que és un cartílag en forma de fulla que cobreix la laringe durant la deglució i evita que l’aliment entri a la tràquea. L’aliment deglutit passa a l’esòfag.
L’esòfag es un tub musculomembranós que va de la faringe al càrdias de l’estómac. La quantitat de fibra muscular llisa i estriada varia segons l’edat i les espècies. En les cries, el vòmit és fàcil; es difícil, en canvi, a la major part dels adults. En els carnívors, és, però, fàcil, fet que indica que tenen una bona proporció de fibra muscular estriada de contracció voluntària en l’esòfag. En els cavalls, el vòmit es fa impossible, a causa, sobretot, de l’existència d’un càrdias extraordinàriament fort. La facilitat del vòmit en els carnívors adults es posa de manifest en observar els costums de cria dels llops que regurgiten l’aliment semidigerit per alimentar les cries.
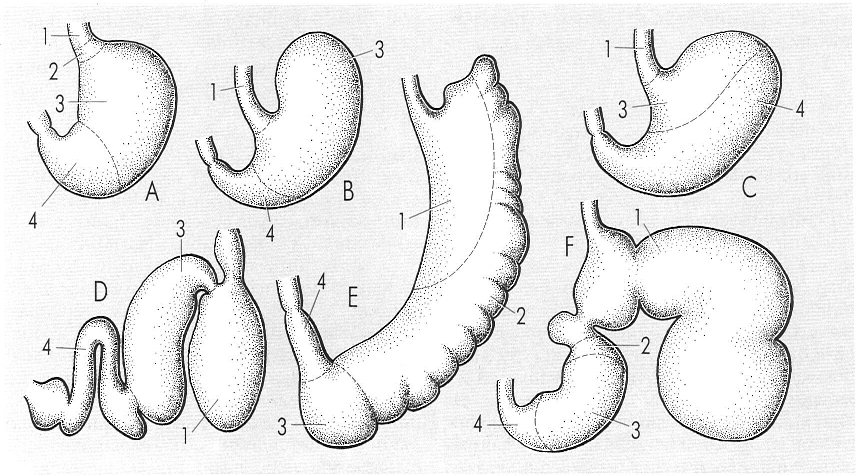
La importància de les diferents parts de l’estómac en els mamífers és en relació amb la seva funció. Hi ha, per això, una tendència a formar cambres diferents que, excepte en els cetacis, es pot explicar com una adaptació a la digestió de la cel·lulosa. En el dibuix s’il·lustren els estómacs corresponents a: A home (Homo), B llebre (Lepus), C rosegador (Citellus = Spermophilus), D dofí (Tursiops), E marsupial (Macropus) i F bou (Bos). Tipus d’epiteli: 1 esofàgic, 2 cardial (només en certs mamífers), 3 fúndic, 4 pilòric. Així mateix, la longitud del tub digestiu és en relació amb el tipus d’aliment ingerit; els herbívors el tenen llarg (el de la vaca fa uns 30 m, el del cavall 27, el de l’elefant 50 m) i els carnívors proporcionalment curt (el budell del lleó fa uns 3 m). L’home, omnívor, té un budell d’uns 10 m, el ximpanzé d’uns 14 m, i el teixó d’uns 4 m.
Gustavo Hormiga, a partir de fonts diverses.
A través del càrdias, l’aliment penetra a l’estómac, situat a la cavitat abdominal sota el diafragma. En els mamífers, existeixen diversos tipus d’estómacs, que són en relació amb el tipus d’aliment o amb la manera d’ingerirlo. En els marsupials herbívors, la cavitat de la regió fúndica augmenta molt de volum i, fins i tot, presenta uns inicis de cecs.
En els remugants s’arriba al grau de transformació més important. Hi ha quatre cavitats: rumen, reticle, llibret i quall. L’herba és acumulada al rumen. L’aliment és freqüentment regurgitat des del rumen i el reticle a la cavitat oral; una vegada mastegat, insalivat i de nou deglutit, torna a l’estómac on és digerida la cel·lulosa gràcies a l’acció de protozous i bacteris; durant aquest procés, gran quantitat de productes són absorbits a través de les parets i passen al torrent circulatori. Des del reticle, l’aliment passa al llibret, de parets musculoses, i, finalment, al quall, típica regió glandular, on s’inicia la digestió de les proteïnes.
A l’extrem de l’estómac, hi ha una glàndula, el pílor, que franqueja el pas del contingut estomacal a l’intestí prim, en el qual es distingeixen tres parts: el duodè, el jejú i l’ili, de característiques glandulars i absorbents. El contingut intestinal passa a través de la vàlvula iliocecal al budell gros, que presenta cec, còlon i recte. El cec pot ésser enorme, com en tots els herbívors no remugants, per exemple el cavall, o reduït, quan la digestió de la cel·lulosa es fa a l’estómac, com és el cas dels remugants. En l’evolució, quan hi ha un canvi de règim alimentari, el cec es redueix i queda un apèndix, resta d’aquest cec, més gran. En el còlon, que és ampli, s’absorbeix, fonamentalment, aigua i, en la part distal, es fan sòlides les defecacions, en les quals queden gravats els plecs de la part distal de l’intestí, per la qual cosa es pot saber a quina espècie pertanyen les defecacions.
Les glàndules annexes
Acompanyant el tub digestiu, hi ha una sèrie de glàndules qualificades d’annexes: les salivals, el fetge i el pàncrees.
Internament, tant als llavis com a les galtes, hi ha glàndules salivals que es poden notar com petits grumolls. En els mamífers, moltes vegades, són tan grans que se situen fora de la cavitat bucal i la drenen a través de conductes. Si l’aliment que prenen és molt sec, cas d’alguns herbívors, les glàndules es fan molt grans. En l’home, tenen una distribució àmplia en la cavitat bucal, darrere els llavis, en una sèrie de petits sortints que es poden trobar palpant amb la llengua, però, a més, hi ha les paròtides, les submaxil·lars i les sublinguals. Les primeres donen una saliva més fluïda i redueixen la seva secreció quan són inhibides per via sinàptica; és per aquest motiu que, en certes situacions de tensió, es nota la boca seca. Molts herbívors tenen glàndules salivals en posició molar, la secreció de les quals fluïdifica l’herba seca que han d’ingerir. Un insectívor, Blarina, segrega una saliva verinosa.
Quant al fetge, l’esbós hepàtic es forma a partir de l’endoderma que ha de donar el tub digestiu. Normalment, en els mamífers, és situat al costat dret, però en alguns primats (Colobus, Nasalis) i quiròpters es troba a l’esquerra. Les seves dimensions varien segons el règim alimentari: en les espècies carnívores, és més gran que en les herbívores. El nombre de lòbuls varia segons els moviments en sentit anteroposterior de la columna vertebral; així, en la llebre, el gos i el gat, el fetge presenta diversos lòbuls, mentre que, en canvi, en les balenes, els bradipòdids, els remugants, els elefants i en molts rats-penats, solament es troben lleugeres fissures; en alguns marsupials, com Phascolarctos i Sarcophilus, n’hi ha trenta o quaranta i en alguns pinnípedes, com Otaria byronia, uns quaranta. Quan hi ha molts lòbuls, com en aquests casos ara citats, es disposen com les teules en una teulada. La bilis es recull a la bufeta del fel, que presenta parets gruixudes en els carnívors, les ovelles i els primats, i és prima en el ratolí i en el conill d’índies. La bufeta del fel pot faltar en algun herbívor.
El pàncrees és una estructura similar a la d’una glàndula salival. En algun cas és difós, com en els ratspenats, les musaranyes i els lèmurs. En altres, és tan compacte que, fins i tot, hi ha una fenestració perquè hi passi la vena porta. En l’home desemboca pel conducte de Wirsung a l’ampolla de Vater, però en moltes altres espècies de mamífers ho fa directament al duodè. Pot haver-hi un conducte accessori, el de Santorini.
L’aparell respiratori
L’aparell respiratori és un derivat endodèrmic evolutivament originat, segurament, d’un diverticle de l’aparell digestiu que aparegué, en el Devonià, en peixos que iniciaven el camí a la tetrapòdia i els va permetre d’anar d’una llacuna a una altra en períodes de sequera.
La tràquea és el conducte aerífer que continua a la laringe i acaba bifurcant-se en dos bronquis. Se situa ventralment a l’esòfag, que la separa de la columna vertebral. És formada per anells de cartílag hialí que alternen amb franges de teixit conjuntiu i fibres elàstiques. Els anells són incomplets, la qual cosa dona certs avantatges en els cops i compressions. Són oberts en direcció al dors, on hi ha una membrana formada per fibres musculars llises. En els moments de tos, aquesta membrana s’enfonsa per dins de la tràquea i fa que l’aire circuli ràpidament per tal d’expulsar, així, les partícules o el material molest que existeix dins de les vies respiratòries.
Els pulmons, fixats a l’interior de la caixa toràcica pels bronquis i els vasos a l’alçada de l’ili, es mouen lliurement però son inerts. Els bronquis formen, en successives bifurcacions, els bronquíols, que acaben desembocant en els alvèols, el teixit respiratori actiu. Els moviments d’inspiració i espiració són producte del joc de les parets toràciques i del diafragma. Gràcies a les pleures derivades de la cavitat celòmica, la pleura visceral, aplicada al pulmó, i la pleura parietal, aplicada a la caixa toràcica, el moviment dels pulmons, solidaris amb la caixa toràcica, és suau. En la major part de mamífers, les dues pleures es mantenen independents l’una de l’altra, però, en els èquids, es fonen a la part posterior del mediastí i, en els elefants, la cavitat interpleural s’oblitera, ja, en l’embrió; segurament, l’intercanvi respiratori és tan fort que faria que les pleures s’esquincessin.
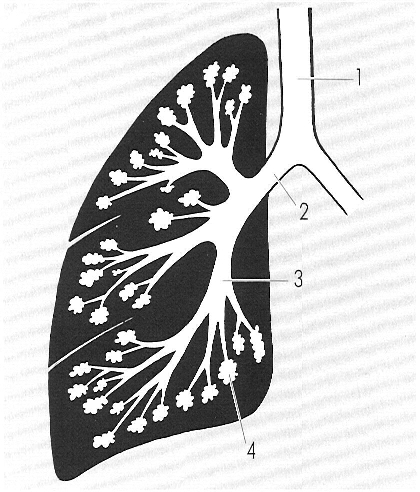
Esquema del pulmó dret de l’home. En els mamífers, la lobació pulmonar és en relació amb el moviment dorsiventral de la caixa toràcica. Prova d’això és que en els cetacis, en què el moviment d’aquesta és petit, la lobació és pràcticament inexistent. 1 tràquea, 2 bronqui, 3 bronquíol, 4 alvèol.
Gustavo Hormiga, a partir de fonts diverses.
Els pulmons són els òrgans essencials de l’aparell respiratori en què es realitza l’hematosi. El nombre de lòbuls que presenten no té valor sistemàtic sinó que es relaciona amb la capacitat de moviment dorsiventral de la columna. Així, el pulmó del cabirol en té molts i el del porc espí en té cinc o sis en els joves i deu o dotze en l’adult. En aquests casos, els lòbuls poden lliscar els uns sobre els altres i el tòrax acostuma a ésser comprimit i allargat. En els animals de gran talla, com els cetacis, elefants i hipopòtams, els problemes són d’una altra mena i els pulmons no presenten lobulació o la presenten molt senzilla.
S’ha comprovat que la mida dels alvèols depèn del tipus de vida de l’espècie i de l’exercici que realitzen el seus membres. Hi ha força dades sobre l’evolució de l’alvèol pulmonar en l’espècie humana i en determinades espècies de laboratori, en relació a l’edat i a l’exercici. Amb l’edat i el sedentarisme, el diàmetre de l’alvèol augmenta, per la qual cosa la superfície activa minva.
Sense formar part, pròpiament, de l’aparell respiratori, el diafragma juga un paper essencial en la respiració. Juntament amb la caixa toràcica, augmenta o disminueix el volum d’aquesta última, fa seguir els pulmons en el seu moviment, i provoca la inspiració i respiració. El diafragma és un múscul aponeuròtic, aplanat i prim, que separa completament la caixa torácica de la cavitat abdominal. La seva forma és, normalment, de cúpula, però s’inclinà diferentment segons les espècies, la qual cosa determina un major o menor volum de la caixa toràcica i, per tant, dels pulmons. En els mamífers aquàtics, és molt oblic, encara que s’ha exagerat molt sobre la capacitat de la caixa toràcica i el volum pulmonar dels mamífers marins. Antigament, es creia que els pulmons dels cetacis eren una reserva d’aire, d’oxigen; de fet, però en realitat, no són més grans del normal en les espècies terrestres i, a més a més, com és lògic, la major part de les espècies els buiden per fer les típiques immersions, perquè, així, la immersió és més fàcil i el poc aire que queda és en l’arbre bronquial, on no s’absorbeix el nitrogen i no es pot presentar, per tant, el fenomen d’embòlia gasosa que tant preocupava, fa uns anys, el món científic.
L’aparell circulatori
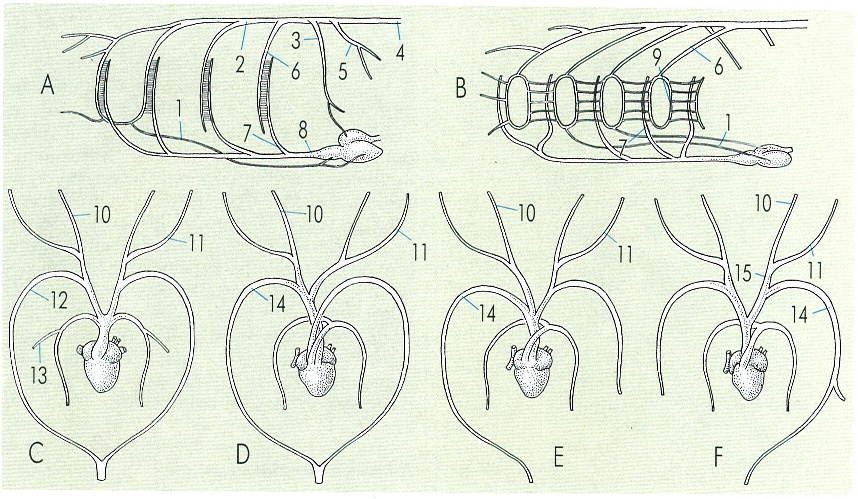
En l’evolució de l’aparell circulatori hi ha una progressiva pèrdua d’arcs aòrtics. Inicialment, en els peixos (A, un teleosti; B un condricti), anaven a irrigar les brànquies i a oxigenar la sang. En els amfibis (C) i els rèptils (D) l’arc sistèmic encara és complet, mentre que en els ocells (E) s’atrofia l’esquerre i en els mamífers (F), el dret; l’aorta dels mamífers és l’arc esquerre cinquè. En el dibuix hom ha indicat: 1 artèria hipobranquial, 2 arrel de l’aorta, 3 artèria subclàvia, 4 aorta dorsal, 5 artèria ilíaca, 6 artèria eferent, 7 artèria aferent, 8 aorta ventral, 9 anell, 10 caròtide externa, 11 caròtide interna, 12 arc sistèmic, 13 artèria pulmocutània, 14 aorta (arc sistèmic), 15 caròtide.
Amadeu Blasco, a partir de fonts diverses.
En els mamífers, els arcs aòrtics es desenvolupen d’una manera similar als dels rèptils, a excepció de l’arc sistèmic, el quart, ja que desapareix el dret i es manté l’esquerre (al contrari que en les ocells, en els quals es perd l’esquerre i queda el dret). El tercer arc forma les caròtides internes que irriguen el cap i el cervell; les extensions anteriors de l’aorta ventral formen les caròtides externes, que van a les mandíbules inferiors. El sisè arc forma les artèries pulmonars. El primer, el segon i el cinquè arcs aòrtics s’atrofien sempre, com passava en els grups inferiors.
El sistema venós difereix molt del que presenten les classes vertebrades inferiors, però al llarg de l’escala filètica dels vertebrats es pot veure la seva evolució fins que s’estructura el que es presenta en els mamífers.
En descriure els arcs aòrtics es veu que en els amfibis, els rèptils i els ocells hi ha dues circulacions: la major i la menor. En els mamífers, els pulmons són ben aconseguits; la circulació menor porta un volum de sang quasi igual al de la circulació major i, per tant, els ventricles poden estar ben septats. Juntament amb els ocells i els crocodilians, són els únics grups de vertebrats que tenen el septe interventricular complet. La compensació de volums circulants es fa en els pulmons, on els capil·lars de la circulació major i els de la menor són en contacte.
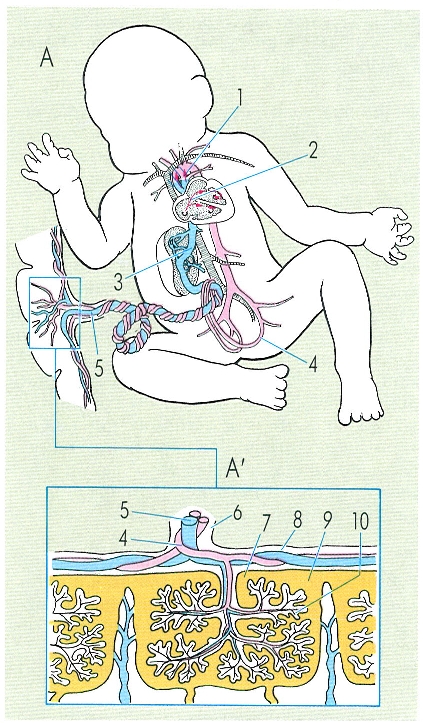
En la circulació del fetus d’un mamífer (A), la sang surt de la placenta per la vena umbilical, arriba a l’aurícula dreta i, gràcies al fet que l’envà interauricular no és complet, pot passar a través de l’aurícula esquerra a la circulació general. La circulació pulmonar no és eficient fins després del part; abans del part la sang impulsada pel ventricle dret passa a través del «ductus arteriosus» a l’aorta. En la placenta de tipus hemocorial (A’), que és la que es presenta en l’home, podem observar l’íntim contacte dels capil·lars de la part fetal de la placenta amb la sang materna. Hom ha indicat en el dibuix: 1 «ductus arteriosus», 2 forat oval, 3 «ductus venosus», 4 artèria umbilical, 5 vena umbilical, 6 cordó umbilical, 7 còrion, 8 amni, 9 espai subcorial (sang materna), 10 vellositats amb vasos sanguinis.
Amadeu Blasco, original de De Melloni.
En els mamífers, hi ha un canvi substancial entre la circulació que presenta el fetus i la que presenta l’adult. L’embrió agafa l’oxigen que necessita per a respirar de la placenta materna i els seus pulmons no són funcionals, ja que, malgrat estar dintre del claustre matern, l’intercanvi gasós és inviable. En el fetus, els pulmons estan col·lapsats i no els arriba sang a través de la circulació menor. Les preguntes que, de bell antuvi, ens fem, són: què passa en el fetus quan el cor es contrau?, es pot donar aquest fet? Bé, en el fetus hi ha dos mecanismes pels quals la sang no va a la circulació pulmonar; aquests mecanismes són: que el septe interauricular no és complet (forat oval) i que hi ha un vas, l’anomenat "ductus arteriosus", que uneix l’aorta amb l’artèria pulmonar. La circulació funciona, en el fetus, de la següent manera: la sang de l’embrió, oxigenada a la placenta, va, a través de les venes umbilicals, a l’aurícula dreta i, des d’aquesta, la sang passa a l’aurícula esquerra des d’on és impulsada al ventricle esquerre i a la circulació general. A l’aorta dorsal, s’originen les artèries umbilicals, que duen la sang a oxigenar-se a la placenta. Ara bé, encara que gran part de la sang vagi de l’aurícula dreta a l’esquerra, quan l’aurícula dreta es contrau, passa sang al ventricle respectiu que, en contraure’s, envia sang a la circulació menor. Com sigui que aquesta sang no pot anar als pulmons, on troba resistència perquè aquests òrgans no estan expandits, abandona la circulació menor i penetra a l’aorta mitjançant el "ductus arteriosus".
En néixer i tancar-se la circulació fetal, baixen els nivells d’oxigen en el nadís i augmentent els d’anhídrid carbònic. Aquest actua sobre els centres respiratoris bulbars, que estimulen els primers moviments del diafragma i dels músculs intercostals. Aquests moviments redueixen la pressió en l’interior de la caixa toràcica i forcen l’entrada de l’aire als pulmons, que segueixen passivament els moviments que fa aquesta. Són els moviments responsables de l’entrada de sang a les artèries pulmonars i redueixen la resistència del flux de sang cap al ventricle dret. El retorn de la sang que ha anat dels pulmons a l’aurícula esquerra fa que augmenti la pressió en aquesta última i força el tancament del forat oval. En l’home, normalment, onze hores després del naixement ja no es barregen les sangs a nivell auricular; durant els primers dies, aquesta vàlvula actua solament per pressió i es pot reobrir; però, al cap de poc temps, se solda i forma una barrera permanent. El "ductus arteriosus" dura més temps i en els humans no es tanca, normalment, fins després d’alguns mesos. En els cetacis, el "ductus arteriosus" es manté i, en les seves prologandes immersions, quan tenen els pulmons colapsats, la sang que hi aniria passa a la circulació general, que no augmenta gràcies al fet que, en aquestes immersions, es dona una marcada bradicàrdia.
En els mamífers, el sistema limfàtic no presenta cors sinó, exclusivament, ganglis. No té, doncs, la complexitat d’altres formes vertebrades.
L’aparell excretor
La regulació del medi intern és una funció de l’aparell excretor. El ronyó dels mamífers, com el de tots els amniotes, és de tipus metanèfric. Es desenvolupa, doncs, a partir de la part més posterior del mesòmer i presenta molts més túbuls que qualsevol altre tipus de ronyó. Cada unitat excretora és formada, en la seva regió anterior, per la càpsula de Browman, que és seguida per un coll, regió molt curta, un túbul convolut proximal, una regió intermèdia, la nansa de Henle i un túbul convolut distal. La nansa de Henle, extraordinàriament llarga, permet una gran absorció d’aigua. Aquesta nansa no és estructurada en els rèptils, de manera que la idea que normalment es té de l’adaptació més gran d’aquests animals a la sequera respecte dels mamífers és falsa; els mamífers, a nivell renal, són molt més adaptats.
Com en els rèptils, els túbuls col·lectors del metanefre no desemboquen en el conducte de Wolff, sinó a l’urèter, que es forma a partir de l’extremitat posterior del conducte de Wolff i penetra en el teixit metanèfric, dividint-se en una estructura en forma d’embut, els calzes renals. És en aquests calzes on entren els túbuls col·lectors. El ronyó té, doncs, un origen doble; d’una banda, el del teixit nefrogènic, mesoblàstic i, de l’altra, a partir d’un brot vesical. En l’home, es calcula que hi ha un milió d’elements excretors per ronyó.
Durant la vida embrionària, l’excreció es fa pel mesonefre, encara que, en algunes espècies, com el conill d’índies i el ratolí, la seva dimensió és tan petita que, difícilment, es pot considerar com una estructura funcional; en canvi, en l’equidna i en alguns marsupials, el mesonefre pot perdurar fins després del naixement. Mentre el mesonefre és actiu, el conducte de Wolff fa de conducte urinari, però, tant bon punt el ronyó metanèfric comença a funcionar, el conducte de Wolff degenera completament en les femelles i passa en els mascles a formar part d’estructures masculines, entre elles l’epidídim, el conducte deferent i les vesícules seminals.
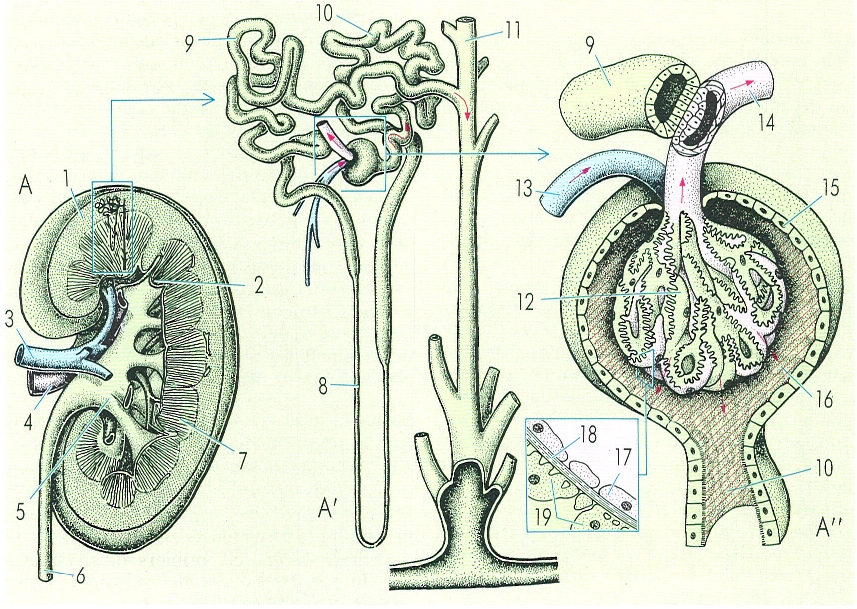
Esquema d’un ronyó de mamífer, concretament d’home (A), on es mostra com, a causa de la disposició dels elements excretors, s’hi diferencia una part cortical i una part medul·lar. L’esquema mostra en detall una nefrona humana (A’) i un corpuscle renal o de Malpighi (A"). Hom hi ha indicat: 1 escorça, 2 piràmide papil·lar, 3 artèria renal, 4 vena renal, 5 pelvis renal, 6 urèter, 7 medul·la, 8 nansa de Henle, 9 tub convolut distal, 10 tub convolut proximal, 11 tub col·lector, 12 glomèrul, 13 arteriola aferent, 14 arteriola eferent, 15 càpsula de Bowman, 16 espai urinari, 17 cèl·lula endotelial, 18 membrana, 19 podocit.
Gustavo Hormiga, a partir de fonts diverses.
Els ronyons són situats a la cavitat abdominal, units a la paret dorsal del cos. De manera característica, en alguns casos, presenten diversos lòbuls, els renculi, que no són altra cosa que la manifestació externa de l’estructura interna que presenten els ronyons dels mamífers. Així, en els remugants, en els óssos i en els cetacis, es troba un veritable ronyó compost per diversos ronyons simples (renculus), proveït del seu propi calze i del seu urèter. Aquests tipus de ronyons apareixen profundament lobats i els seus elements es mantenen units entre ells formant un òrgan compacte per teixit conjuntiu.
La bufeta urinària, present en tots els mamífers, és un sac que deriva d’una part de l’al·lantoide. La bufeta, que rep la secreció urinària, s’aprima per la seva part posterior fins unir-se a la uretra, que la comunica amb l’exterior. En el punt d’unió entre la bufeta i la uretra, hi ha un esfínter que regula la sortida de l’orina. En els mascles, l’uretra passa a través del penis; en les femelles, s’obre a l’extrem del clítoris o, en algunes espècies, en el sinus urogenital. L’urèter té un determinat trajecte dins de la bufeta, i aquest trajecte actua com a mecanisme valvular per impedir el reflux de l’orina en direcció a la pelvis renal.
L’aparell reproductor
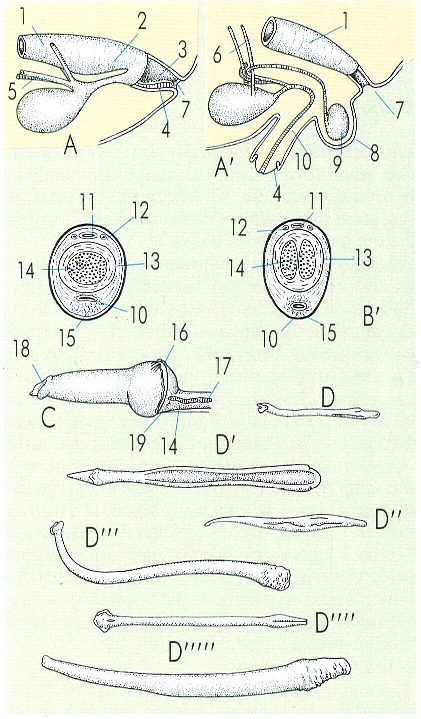
Morfologia de l’aparell copulador en els mamífers: esquema de la relació existent entre els aparells digestiu i urogenital en els monotremes (A) i en la majoria dels altres mamífers mascles (A’); secció transversal d’un penis de mona rhesus a l’àrea proximal (B) i a l’àrea distal (B’); gland del penis d’un gos en erecció (només la part lliure del penis, C); ossos del penis de fura (D), de gos (D’), de guineu (D"), de «mapache» (D’"), de toixó (D"") i d’ós (D’""). Hom ha indicat: 1 recte, 2 coprodeu, 3 proctodeu, 4 penis, 5 conducte deferent, 6 urèter, 7 anus, 8 escrot, 9 testicle, 10 uretra, 11 vena dorsal del penis, 12 artèries, 13 túnica fibrosa, 14 cos cavemós, 15 cos esponjós, 16 vena anterior, 17 vena posterior, 18 meat urinari, 19 secció dels teguments de la veina.
Amadeu Blasco, a partir de fonts diverses.
Dins del conjunt de sistemes que presenten els vertebrats, el sistema reproductor és únic. Així com les adaptacions que es donen en altres sistemes contribueixen a la supervivència de l’individu, aquest, el reproductor, assegura la supervivència de l’espècie.
Les unitats morfològiques de reproducció són, com en tot el regne animal, els gàmetes—espermatozoides i òvuls— que requereixen un òrgan, la gònada, en la qual es formen i s’emmagatzemen.
Les gònades són glàndules compostes que, a més de formar i guardar gàmetes, secreten hormones. Es formen a partir de masses mesoblàstiques, però, a diferència del ronyó, no mostren segmentació. La proliferació d’aquest teixit forma la cresta genital que penja en la cavitat celòmica. A la cresta genital, és ben definit l’epiteli germinal, que, eventualment, formarà les cèl·lules germinals. En aquest estadi, l’epiteli germinal prolifera i els gonòcits van a instal·lar-se en la cresta genital; aquestes masses es denominen cordons sexuals i seran els tubs seminífers dels mascles i els cordons medul·lars de les femelles. La gònada indiferenciada adopta progressivament l’aspecte definitiu. Si es desenvolupa la regió cortical i es reabsorbeix la porció medul·lar, dona un ovari; si, per contra, creix la part medul·lar i es reabsorbeix la cortical, dona un testicle.
En tots els vertebrats, l’estructura interna dels testicles és extraordinàriament constant; en els mamífers, la seva posició pot variar: inicialment situats a la cavitat abdominal, a la regió dorsal anterior, en alguns casos poden migrar fora de la cavitat abdominal per situar-se en una bossa, l’escrot. En línies generals i amb el grau de veritat que reflecteix tota afirmació general, es pot dir que els mamífers primitius tenen els testicles dins la cavitat del cos i que les formes més evolucionades els tenen fora. Així, en els monotremes, cetacis i sirenis, es troben, durant tota la vida, a la cavitat abdominal; en alguns marsupials, en molts rosegadors, en els insectívors, en els rats-penats, en alguns ungulats, com els camells, i en alguns primats, els testicles baixen a la bossa escrotal sols a les èpoques d’activitat sexual; en altres marsupials, en molts carnívors, en els ungulats i en els primats, els testicles baixen a l’escrot a l’última fase del desenvolupament fetal i hi romanen per sempre. En les espècies en què els testicles són fora de la cavitat del cos, els casos de criptorquídia—testicles situats dins de la cavitat abdominal— suposen una menor fertilitat. A l’escrot, els vasos sanguinis són organitzats de tal manera que, per sistemes de contracorrent, es refreda la bossa escrotal; això permet comprendre el fet que, en determinats grups i durant l’època de zel, els testicles se situïn en una bossa escrotal especial situada fora de la cavitat del cos.
Els espermatozoides, produïts en els testicles, són conduïts a l’exterior per una sèrie d’estructures derivades del conducte de Wolff, que prístinament tenia missió excretora, però, posteriorment, passa a estar al servei de l’aparell genital. Concretament els conductes eferents comuniquen els tubs seminífers del testicle amb l’epidídim, que continua amb el conducte deferent, que drena l’uretra a escassa distància de la bufeta urinària. En molts mamífers, el final del conducte deferent s’eixampla per emmagatzemar, durant un cert temps, els espermatozoides. Diverses estructures glandulars desemboquen a diferents parts d’aquest recorregut i, entre els espermatozoides i les secrecions d’aquestes glàndules, es forma un líquid especial, el semen o esperma. Cal dir que, en els mamífers, la vesícula seminal no té missió d’emmagatzemar sinó una funció glandular.
En tot el món vivent, la fecundació es dona sempre en un medi aquós. D’aquí la necesitat, en el medi terrestre, d’introduir el semen en la cavitat materna. El penis dels monotremes condueix exclusivament semen; l’orina és alliberada a la cloaca. En alguns marsupials el penis és bífid. En el penis de molts mamífers placentaris, hi ha un os en el septe que separa els cossos cavernosos.
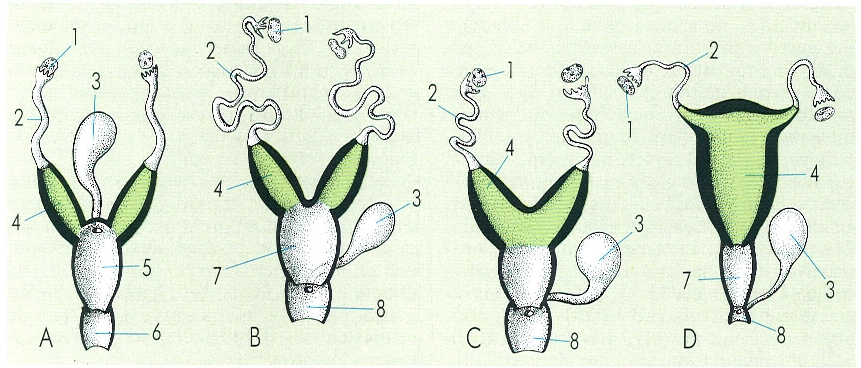
Diferents tipus d’úter dels mamífers: A úters separats d’un monotrema; B úter doble d’un conill; C úter bicorne d’una vaca, en què els corns són parcialment fusionats: i D úter senzill humà, amb una cambra central. Hom ha indicat en el dibuix: 1 ovari, 2 oviducte. 3 bufeta urinària, 4 úter, 5 sinus urogenital, 6 cloaca, 7 vagina, 8 vestíbul.
Amadeu Blasco, a partir de fonts diverses.
En les femelles, es poden diferenciar l’ovari o gònada productora d’òvuls i els conductes genitals. També en les femelles, paral·lelament al conducte de Wolff, es forma el conducte de Müller. Això també passa en els mascles, però, en aquests, aquest darrer conducte degenera. El conducte de Müller no està en continuïtat amb l’ovari; d’aquí que els òvuls en el moment de produir-se, hagin d’anar a la cavitat del cos i ésser aspirats per la part proximal del conducte de Müller. D’aquí també que, en determinats casos, alguns òvuls s’escapin en aquesta aspiració i es quedin en la cavitat celòmica. El conducte de Müller presenta diferents segments amb diverses funcions: l’òstium, l’oviducte, l’úter i la vagina. L’òstium recull l’òvul, que sorgeix envoltat d’una corona de cèl·lules radiades, quan el fol·licle es trenca; queda l’òvul entre plecs de mucosa que formen una superfície de recepció humida i tova i progressivament és transporta per l’oviducte gràcies a la capa muscular i als cilis de la mucosa de l’oviducte.
El transport dels òvuls dura alguns dies i els moviments de l’oviducte no deuen ésser continus sinó que deuen tenir una influència hormonal. L’espermatozoide fecunda l’òvul en el terç anterior de l’oviducte. Iniciat el procés de segmentació de l’ou, en l’espècie humana al sisè dia, en estat de blastòcit, s’implanta a l’úter. En els mamífers, els graus de soldadura dels conductes de Müller són variables en els diversos grups animals i es formen úters dobles, bicornes o simples.
En els mamífers, sols els monotremes tenen una cloaca per a rebre el penis. En la resta, les parts distals dels conductes de Müller es modifiquen per a formar l’orifici genital que rebrà el penis. La part externa de l’obertura genital, el sinus urogenital,és formada per una porció invaginada de la pell. En l’interior de la paret del sinus urogenital, s’hi troba el clítoris, estructura homòloga del penis masculí, que, en algunes espècies, com en les hienes, és tan gran que s’havia pensat en l’existència d’intersexes. Internament al sinus urogenital, s’hi troba la part més distal del conducte de Müller, la vagina. Entre el sinus urogenital i la vagina, hi ha un plec de teixit, l’himen, que es trenca i sagna a la primera còpula. En els marsupials, els extrems distals dels conductes de Müller, dret i esquerre, no s’uneixen i tenen, per tant, vagines parelles; també és comprensible, per això, que el penis dels marsupials sigui doble. En els placentaris, els extrems dels conductes de Müller es fusionen i formen una vagina imparella, de posició mitjana, que és la que rep el penis en el moment de la còpula.
Les característiques i les adaptacions fisiològiques dels mamífers: la regulació tèrmica
Una de les característiques més importants dels mamífers és la seva capacitat de regular la temperatura, que s’ha de veure dintre d’un fenomen més ampli, l’homeòstasi, la constància del medi intern. La conservació de la temperatura exigeix el manteniment d’un balanç calòric, fet que no es dona del tot en les cries; d’aquí, entre altres motivacions, la necessitat del contacte amb la mare, que els dona l’escalfor que els falta. La calor es produeix per l’activitat metabòlica i per la captació de la calor solar. Les pèrdues de calor són resultat de la radiació i també resulten de l’evaporació a través de la superfície de la pell i dels processos respiratoris. Com és natural, el mamífer ha de recórrer a uns diferents mecanismes de control segons sigui hivern o estiu. A l’hivern, el pelatge és gruixut i s’aclareix a l’estiu. L’excés de calor pot ésser eliminat pel calor d’evaporació gràcies a la sudoració, a l’evaporació a través de la llengua o a l’assecament de la superfície del cos, que s’ha humitejat prèviament amb saliva.
A les zones polars o a l’hivern a l’alta muntanya, les espècies de mida petita eviten les temperatures excessivament baixes amagantse sota la neu. Les espècies de mida gran tenen una superfície aïllant important; a més, la seva mida ja és una condició favorable, ja que, proporcionalment, la superfície d’irradiació es fa proporcionalment menor a la producció de calor, donat que aquesta és en relació amb el volum (amb l’augment de mida, la superfície creix al quadrat i el volum al cub). A les parts distals, poden haver-hi fenòmens d’hipotèrmia, la qual cosa produeix la minva del gradient tèrmic i, per tant, la pèrdua de calor.
Les regions càlides i seques ofereixen dificultats més serioses, ja que l’individu ha de disposar d’aigua per a poder regular la seva temperatura i evitar el sobreescalfament. Un cas molt ben estudiat de regulació tèrmica en les condicions del desert és el del camell. Aquest animal no panteixa, encara que excepcionalment ho pot fer quan ha de baixar la temperatura corporal, però té una extraordinària capacitat de variar aquesta temperatura i la utilitza quan no pot beure aigua regularment. La fluctuació de la seva temperatura corporal és de 6 °C. A la nit, quan la temperatura exterior és baixa, la seva temperatura corporal baixa per sota dels 35 °C; de dia, la pot incrementar fins per sobre dels 40 °C. Estalvia, així, l’aigua que perdria amb la calor i aconsegueix mantenir la seva temperatura dins d’uns límites. Altres factors que intervenen en aquest manteniment són la tendència a mantenir-se agrupats en els moments de descans, que fa que la superfície exposada al sol sigui menor, i també la seva resistència a la set (aguanta deshidratacions del 27%).
De vegades, però, les condicions del medi esdevenen tan desfavorables per excès de fred o de calor que els organismes han d’aturar la seva activitat. Enfront del fred, presenten dos tipus de reaccions: l’endormiscament (els óssos) o la hivernació autèntica (els rats-penats). Enfront de la calor, els esquirols de les nostres latituds presenten una reacció particular; a l’època de màxima calor, el seu metabolisme és tan intens que difícilment podrien mantenir la seva temperatura corporal i entren en un endormiscament del qual no surten fins que han passat els màxims tèrmics.
L’estratègia biològica
La reproducció
El zel i l’aparellament dels mamífers és, sempre, discret. No es manifesta amb el cant i l’esplendor de colors que es dona en els ocells. Ara bé, és possible que, en canvi, sigui espectacular en el món de les olors i que l’home no sigui capaç d’interpretar tots aquests fets, ja que, com a primat, el seu comportament —excepcional entre els mamífers— es dirigeix per la visió i no per l’olfacte. L’home interpreta millor el món dels ocells, ja que aquests també es relacionen visualment. En els mamífers, no hi ha brillants danses nupcials ni cants melòdics, però sí que poden haver-hi certes veus com el bramul del cérvol o els ultrasons dels rats-penats, a més de tot el llenguatge mímic i el moviment del pèl i de la cua.
Excepte en els primats superiors i en algun altre grup, les femelles dels mamífers presenten amplis períodes en què no són sexualment receptives. El període d’activitat sexual que es dona en les femelles és l’estre, calor, moguda o zel. En l’estre, es realitza la posta de l’òvul gràcies a una coordinada secreció hormonal. L’òvul recollit per les trompes de Fal·lopi és fecundat en el primer terç de l’oviducte abans d’alliberar el segon corpuscle polar i s’implanta en l’úter. L’estre pròpiament dit és precedit d’un preestre, en què creixen els fol·licles ovàrics, i seguit d’un metaestre, en què es degrada el cos groc; per fi, la femella entra en la fase de diestre o calma sexual. En les tres primeres fases, hi ha tota una sèrie de manifestacions fisiològiques i comportamentals.
El nombre de cicles anuals és variable. Les espècies monoèstriques en tenen un a l’any; altres, les polièstriques permanents, com els primats superiors, presenten cicles periòdics continus durant tot l’any. Les polièstriques estacionals tenen més d’un cicle per any, però presenten llargues fases d’inactivitat, algunes en cada equinocci, com el gos; altres poden tenir dues ventrades seguides a la primavera, com l’eriçó. La còpula és dorsiventral com a norma, encara que pot ésser ventral en grups molt distants filèticament, com l’hàmster, l’home i els cetacis.
Alguns mamífers son monògams i les seves parelles es mantenen després d’haver criat els petits (llops, primats), però el més freqüent és la promiscuïtat total, tant en els típicament socials (ramats) com en els estrictament solitaris. La poligàmia amb formació d’harems es presenta en els cérvols i en els pinnípedes, però no en els fòcids.
Tipus de placentes: epicorial i difusa, pròpia de paquiderms, cetacis, cavalls i porcs; mesocorial i cotiledonal, pròpia dels remugants; endocorial i zonal, pròpia dels carnívors; hemocorial i discoïdal, pròpia de l’home i altres primats. A les placentes indecídues (a dalt) la mucosa uterina participa poc en la formació de la placenta i pràcticament no hi ha hemorràgia quan es desprèn. A les placentes decídues (a baix) l’endometri i el còrion estableixen un contacte íntim i quan es desprèn la placenta hi ha hemorràgia. Hom ha indicat en el dibuix: 1 endometri, 2 còrion 3 capil·lar uterí 4 epiteli uterí 5 sinciti corial 6 capil·lar al·lantoïdeu 7 mucosa uterina. 8 llacuna de sang materna 9 mesoderma al·lantoïdeu.
Amadeu Blasco, segons Houillon.
La característica més important del desenvolupament dels mamífers és l’existència d’una placenta que possibilita l’intercanvi de materials entre la mare i el fetus i resol la poca reserva nutrititva de l’ou, que es consumeix entre els quatre i els deu primers dies, segons les espècies. L’aparell placentari és el conjunt resultant de la placenta fetal originada a partir del blastòcit i la placenta materna sorgida de la mucosa uterina. La mucosa uterina pot participar en diferent grau en la formació de la placenta. Segons aquest grau de participació, es distingeixen quatre tipus de placenta: les placentes epicorial i mesocorial, indecídues, i les placentes endocorial i hemocorial, decídues.
La placenta epicorial o epitelicorial presenta l’epiteli uterí no modificat. Entre les vellositats corials i les vellositats maternes, hi ha una secreció de les glàndules uterines que és absorbida pel sincici corial. Com sigui que els annexos fetals i l’úter estan només en contacte durant el part, no hi ha hemorràgia. Aquesta placenta es denomina també difusa, perquè les vellositats placentàries no tenen una localització preferent. És la placenta dels cetacis, les eugues i les truges.
A la placenta mesocorial, l’epiteli uterí es destrueix a l’alçada de les vellositats corials. El sincici corial, situat directament en contacte amb el teixit conjuntiu matern, permet un intercanvi més íntim. Les vellositats es concentren en determinades àrees o cotilèdons; per això, la placenta es diu cotiledònia. És típica dels remugants.
A la placenta endocorial, l’epiteli uterí i el conjuntiu subjacent són destruïts pel sincici corial i els capil·lars sanguinis guarden el seu endoteli, però queden immersos en el sincici corial que és molt desenvolupat. A causa del fet que els teixits materns i fetals estan més íntimament relacionats, en l’expulsió de la placenta es provoca una hemorràgia. Les vellositats no ocupen més que una zona anular i, per això, la placenta es diu zonal. És la placenta pròpia dels carnívors.
A la placenta hemocorial, el sincici corial ocupa una gran extensió i destrueix no sols l’epiteli uterí, sinó també l’endoteli dels capil·lars uterins. Els vasos sanguinis materns dilatats formen llacunes, però es mantenen els vasos sanguinis d’origen fetal. Les vellositats es localitzen en una regió que té aspecte de disc i, per això, la placenta es diu discoïdal. Aquest tipus de placenta és pròpia dels insectívors, els quiròpters i els primats i, per tant, també, de l’home.
A l’embrió, apareixen també els annexos embrionaris típics dels amniotes, l’amni i l’allantoide, encara que aquesta última redueix la seva importància.

Fetus de rorqual comú (Balaenoptera physalus) allotjat en una de les branques uterines de la mare (l’altra branca de l’úter, que és bicorne en els cetacis, és ocupada per l’al·lantoide); en aquest cas, en què no sembla que hi hagi implantació diferida, el temps de desenvolupament embrionari és d’onze mesos, i la cria neix amb més de 6 m de llargada.
Esteve Grau
El període de gestació és molt variable i és relacionat amb el tipus de placenta, amb la relació mare-fetus i, a més, amb la mida de l’espècie. En alguns casos, es veuen clares oscil·lacions, que són conseqüència de fenòmens d’implantació diferida: l’embrió, en estat de blastòcit, queda en les vies genitals femenines fins el moment en què la femella és en condicions fisiològiques de poder aportar-li aliment. Es dona en un ampli nombre de mamífers, com, per exemple, els elefants, els carnívors, els rosegadors, etc.
En el naixement, s’han de diferenciar dos processos: el part pròpiament dit i l’alliberació de la placenta, que és ingerida en la majoria de les espècies per la mare, que l’aprofita com a aliment, ingereix hormones i no deixa rastres que avisin els possibles depredadors. En els marsupials, les estructures que relacionen la mare i els embrions són variables segons les espècies, però sols permeten un curt període de desenvolupament a l’interior del claustre matern. Els monotremes són ovípars i el seu desenvolupament sembla més el d’un ocell.
El comportament en el part ofereix curioses adaptacions. Algunes espècies pareixen tombades, altres a peu dret; els rats-penats ho fan penjats de tres potes, mentre que amb l’altra estiren l’uropatagi per recollir el nadís. En els parts normals, la presentació és de cap, però en els cetacis la normal és de cua per tal que el jove pugui anar ràpidament a respirar fora de l’aigua. Les mares acostumen a parir en llocs amagats i de manera discreta i, en néixer el petit, el llepen immediatament per assecarlo i netejar-lo del líquid amniòtic. Després, el petit mama el calostre (líquid que secreta la glàndula mamària abans de la llet) i la primera llet.

Un dels trets conductuals rellevants dels mamífers és la cura de les cries a les primeres fases de la vida extrauterina. Entre les dependències dels fills respecte la mare cal destacar les derivades de la protecció. A la fotografia il·lustrem el cas d’una lleona que transporta la seva cria, cosa freqüent entre els fèlids.
Prensalcor/Jacana.
El transport dels petits és, de vegades, molt especial: en alguns casos les mares els agafen amb la boca quan els han de canviar de cau (cas del gat), en d’altres, les cries van en processó mossegant cadascuna la cua de la del davant i la primera la de la seva mare (les musaranyes) o van muntades sobre la mare i agafades a la seva cua (sarigues), etc. Les mares ensenyen els petits a distingir l’aliment o les possibles preses fins que progressivament deixen d’ésser alletats i s’espavilen sols. El temps d’alletament varia segons les espècies.
Els cicles d’activitat
L’activitat diària dels individus és influïda pel fotoperíode. Els mamífers diürns inicien la seva activitat a la sortida del sol i l’acaben al crepuscle. A l’estiu, sol haver-hi un descans en els moments de calor més forta mentre que, a l’hivern, els individus no hivernants acostumen a prendre el sol al migdia. Hi ha espècies crepusculars o nocturnes, com els rats-penats o el teixó, que inicien la seva activitat al crepuscle. La lluminositat i la humitat afecten molt la intensitat de la seva activitat. Dediquen la major part del dia a la cerca i captura d’aliment, bé que hi ha una gran variació en les hores d’activitat segons l’època de l’any i el temps atmosfèric. També varia molt segons les espècies; és molt constant en els herbívors i oscil·la molt en els carnívors. Els jocs, les passejades, la neteja, la defensa contra els depredadors, el manteniment de la jerarquia social i la defensa del territori propi consumeixen la resta del temps. Les estones d’inactivitat les dediquen al repòs, a la remugació, si n’hi ha, i a dormir.
Les migracions i la letargia
Davant de condicions adverses, com el fred o la falta d’aliment, els mamífers reaccionen, segons les espècies, de dues maneres diferents: hivernant o desplaçant-se, tot fent migracions periòdiques. En aquest cas, hi ha un àrea d’hivernada i una de reproducció. Els cetacis, els pinnípedes i els grans herbívors són mamífers migratoris típics. Els rats-penats fan migracions, però també són capaços d’hivernar. Migració i hivernació es relacionen amb les condicions ambientals. Els herbívors emigren per buscar indrets adequats per a viure; els pinnípedes, per buscar les àrees de cria. Els cetacis, sobretot les grans balenes, migren seguint àrees riques en aliment (les zones fredes), però cerquen aigües càlides per tenir la descendència: les balenes neixen sense la capa de greix protectora, per la qual cosa la difusió de la calor és massa gran per a néixer i viure en aigües fredes; la llet de la mare, molt rica en greix, fa que s’estructuri ràpidament la capa de greix al voltant del cos i, per tant, al cap de poc temps poden iniciar la migració cap a les aigües fredes riques en aliment.
Les particularitats anatòmiques i fisiològiques de cada espècie, així com la seva posició en la xarxa tròfica, influeixen enormement en la mida de les poblacions respectives. Les espècies que tenen a l’abast una important quantitat d’aliment són les més abundoses; les que se situen al final de les cadenes tròfiques formen, lògicament, poblacions més reduïdes. Influeixen també les condicions ambientals, que expliquen les oscil·lacions de les poblacions en el temps. És molt coneguda la dels lèmmings, l’èxit reproductor dels quals depèn de la quantitat d’aliment que troben i, en última instància, del flux d’energia incorporat pels productors primaris. Els lèmmings, en els anys bons, fan autèntiques explosions de població i exhaureixen l’aliment en aquestes àrees, de manera que han de realitzar impressionants migracions col·lectives en cerca de noves àrees on poder viure. És en aquestes migracions que travessen els rius, on formen autèntics ponts vivents passant els uns sobre els altres; fins i tot quan arriben a la mar, utilitzen la mateixa tàctica, però mai no arriben a terra ferma; aquest intent migratori és el que la gent interpreta com un suïcidi en massa. (L’any 1868 es va observar a Trondeheim un banc de lèmmings tan gran que un vaixell a vapor va tardar quinze minuts a travessar-lo.) Hi ha moltes altres espècies que pateixen aquestes oscil·lacions en el nombre d’individus. Són molt conegudes les dels esquirols, ratolins de camp, guineus, etc.
És lògic que una variació en la vegetació influeixi enormement en la resta de la cadena, tant per qüestions alimentàries com per les de refugi que suposa; per aquest motiu, en tots els intents de protecció d’una espècie animal s’ha de vigilar que es protegeixi l’entorn. La profunda alteració de la fauna australiana, en molts casos, més que el producte d’una acció directa de l’home sobre ella, ha estat una conseqüència de l’alteració del medi.
No hem d’oblidar que la majoria dels mamífers de petita mida i extraordinàriament nombrosos són font d’aliment de molts altres vertebrats: ocells, rèptils, alguns amfibis i, fins i tot, de certs peixos. Fins i tot aquesta visió de la importància dels mamífers com a aportació alimentària seria simplista si no es tingués en compte la importància dels mamífers en el reciclatge de la matèria, sobretot per la seva aportació de materials residuals. L’impacte dels mamífers sobre la vegetació és, en molts casos, selectiva, ja que prefereixen alimentar-se de determinades espècies vegetals.
La locomoció
Els mamífers han aconseguit en el seu procés d’evolució dominar tots els medis, no sols el terrestre, ja que certes formes han colonitzat amb èxit el medi aquàtic i l’aeri. Bàsicament es troben, doncs, tres medis de locomoció: la marxa, la natació i el vol.
La marxa

El desplaçament dels mamífers presenta característiques pròpies en funció del tipus de marxa. Les variacions de la morfologia dels peus són característiques dels diferents grups i, per això, les petjades permeten determinar, en molts casos, les espècies. A l’esquerra veiem un rastre de llebre i a la dreta, de marta, deixats pel seu pas sobre la neu.
Jaume Orta i Jordi Ruiz Olmo.
El mètode més típic de desplaçament dels mamífers és el que es produeix per mitjà del recolzament sobre les quatre potes, que és el que es dona a la immensa majoria de les formes terrestres. Quan el mamífer va al pas, es recolza en les extremitats situades en diagonal i desplaça endavant les altres dues, que passen a ésser les extremitats en què es recolza en el següent moviment. Algunes espècies, com el camell, la girafa i l’ós, coordinen de diferent manera el moviment dels seus membres i desplacen primer i de manera simultània els d’una banda, i després els de l’altra (són ambladors): l’individu en moviment va com si caigués, desplaçant el seu cos de manera alternativa a un costat i a l’altre, al mateix temps que progressa cap endavant. En el trot, l’animal llança el seu cos amb les extremitats en diagonal, de manera que, en canviar el suport d’unes extremitats en diagonal a les altres, hi ha un moment en què està en suspensió. El galop es pot dir que consisteix en una sèrie de salts i caigudes alternades i concatenades: l’animal salta amb les extremitats posteriors i frena la caiguda amb les anteriors; les petjades que deixa a terra acostumen a disposar-se segons un model quadrangular.
En cadascun d’aquests passos, l’animal augmenta la seva velocitat de desplaçament a traves del moviment més ràpid de les extremitats i de l’augment de la distància que les separa. Les velocitats màximes varien segons les espècies. Però no sols és important la velocitat màxima d’una espècie, sinó el temps que és capaç de mantenir-la i el temps que triga en arribar a adquirir-la; és a dir, l’acceleració. Ens trobem, per exemple, que els herbívors, que per norma general són més lents que els seus depredadors (els carnívors), són capaços de mantenir la seva velocitat màxima durant més temps; aquest fet els permet evitar, sempre que mantinguin una distància prudencial respecte dels carnívors (l’anomenada distància de fugida) que aquests els puguin atrapar. Només els atrapen quan són més lents en la seva fugida, és a dir, en casos de malaltia, vellesa, etc.
Alguns quadrúpedes poden mantenir-se en posició bípeda durant un cert temps: alguns cérvols, l’antílop girafa i la mateixa cabra, són capaços de mantenir-se en posició vertical per menjar fulles d’arbre. El ximpanzé i el gorila es recolzen en el dors de les falanges de les mans i, en curts trajectes, insinuen el bipedisme, que es fa normal en l’espècie humana. Alguns mamífers tenen una manera d’avançar molt especial, un salt en què intervé la cua; és el desplaçament típic dels cangurs, per mitjà del qual arriben a velocitats de fins 50 km/h i a salts rècords de 7 a 12 m. Entre els rosegadors, els jerbus avancen de la mateixa manera i, entre els primats, els del gènere Tarsius, encara que arborícoles, sols es desplacen a salts. La braquiació es dona en certes mones arborícoles, la més representativa de les quals pot ésser la mona aranya (gènere Ateles), que, si bé és cert que es pot desplaçar amb els membres posteriors, són els membres anteriors els que actuen normalment com a locomotors. El gibó, quan va per terra, utilitza les extremitats posteriors, però quan va pels arbres, es desplaça per braquiació, i en un moment determinat sols s’agafa per una mà a una branca, però, aleshores, impulsa el cos a tall de pèndul i arriba amb l’altra a una nova posició. Algunes espècies utilitzen, a més, la cua.
Velocitats màximes assolides per diferents espècies de mamífers
| ESPÈCIE | VELOCITAT Km/h |
| Camell | 14-16 |
| Ovella | 24 |
| Conill | 38 |
| Elefant | 25-40 |
| Llop | 45 |
| Girafa | 45-50 |
| Llebre | 55-70 |
| Zebra | 65 |
| Gasela | 80 |
| Lleó | 80 |
| Guepard | 104-112 |
| Dades elaborades per l’autor | |
La natació
La majoria dels mamífers estrictament terrestres són, en determinades ocasions, capaços de nedar. Entre ells es troben la llebre, la majoria de les rates i els mustèlids; aquests darrers són mamífers nedadors. Els ungulats i els carnívors neden perfectament; els senglars fins i tot són capaços de travessar l’aigua salada, i alguns rats-penats, que s’alimenten dels insectes que volen prop de l’aigua o de peixos, són capaços d’entrar en contacte i estar durant cert temps en l’aigua per reprendre immediatament el vol. Hi ha mamífers en què és normal el contacte amb l’aigua, com els castors, les llúdries i les rates d’aigua. La llúdria de mar rarament va a la costa i per descansar es lliga amb la vegetació per evitar ésser arrossegada pels corrents.
Les foques, les morses i les otàries es mouen per l’aigua gràcies als moviments coordinats de les seves extremitats; tant les otàries com les morses, a terra ferma, es desplacen doblegant les extremitats posteriors alternativament cap endavant i cap enrere. Les foques no tenen les extremitats posteriors reversibles, la qual cosa confirma l’origen distint d’aquest grup. Els cetacis i els sirenis avancen per moviments de la cua, mentre que les extremitats fan d’estabilitzadors; els cercles que es veuen a l’aigua, que indiquen la presència de cetacis, són fets per les columnes d’aigua que generen els cops de cua.
El vol

L’únic grup de mamífers que presenta autèntiques adaptacions al vol actiu és el dels quiròpters, els quals disposen d’una membrana, el patagi, que els faculta per al vol batut, alhora que tenen la capacitat d’orientar-se per un sistema d’ecolocalització. La figura ens mostra una rata-pinyada pipistrel·la (Pipistrellus) en ple vol.
Ricard Gutiérrez.
Entre els mamífers, es troben dos tipus diferents de vol: el planatge, que realitzen els esquirols voladors, tots els dermòpters i algun marsupial, i el vol batut, que és típic dels ratspenats. El planatge és un tipus de desplaçament secundari; en canvi, el vol batut dels rats-penats els fa estrictament aeris (el seu desplaçament per terra ferma és molt feixuc).
Els esquirols voladors del gènere Glaucomys presenten ungles ben desenvolupades que els permeten grimpar entre les branques dels arbres. Tenen una àmplia membrana en els costats del cos que enllaça les extremitats anteriors amb les posteriors; a més, a les anteriors, hi tenen un esperó cartilaginós, que és un suport addicional del plec cutani. Aquesta estructura, juntament amb la cua, els permet amplis vols planats. La tendència al vol s’observa, ja, en els nostres esquirols, que són capaços de deixar-se caure d’un arbre estenent les extremitats i la cua.
Els quiròpters o rats-penats són els mamífers voladors per excel·lència, que, per adaptar-se al vol, han hagut de modificar molt la seva extremitat anterior, en especial els metacarpians i les falanges dels dits, estructures òssies que suporten una doble membrana cutània que recobreix internament i externa aquestes formacions. La irrigació del patagi és especial i evita la pèrdua de calor, però no ha pogut evitar la pèrdua d’aigua. Aquesta és la raó per la qual els quiròpters són crepusculars o nocturns, ja que la humitat de la nit fa que hi hagi menys pèrdua d’aigua per la membrana del patagi; només en llocs amb un elevat índex d’humitat poden portar una vida activa durant el dia, com per exemple al delta de l’Ebre. A més d’aquest quiropatagi, presenten un prepatagi a la part anterior de les extremitats toràciques i un uropatagi entre les extremitats posteriors i la mà. En aquest uropatagi, recullen la cria en el moment del part i, a més, els serveix per a capturar alguns insectes. Els macroquiròpters o guineus voladores són animals fungívors d’Africa i Austràlia, d’ulls grans i en els quals no s’ha demostrat l’ecolocalització. Treballs recents (1986), basats en l’estructura del tracte òptic, demostren que aquesta és molt semblant a la dels primats, molt característica, per cert, i diferent de la dels altres mamífers. Els microquiròpters la tenen, en canvi, com la majoria dels mamífers. Sempre s’havia parlat del possible difiletisme entre els microquiròpters i els macroquiròpters, però no deixa de ser sorprenent que es puguin arribar a interpretar com una soca que s’ha escindit dels primers primats i s’ha adaptat al vol. En aquest cas, el patagi dels microquiròpters i macroquiròpters s’hauria d’interpretar com una convergència. L’ull ben desenvolupat dels macroquiròpters pot lligar amb el seu origen, ja que la vida arborícola dels primats requereix una excel·lent capacitat d’anàlisi visual de la distància i de la possible fragilitat de les branques, per assegurar el seu desplaçament pel món aeri.
L’estructura social

Molts mamífers presenten organització social, de vegades d’un grau molt complex, que pot ser temporal i variable segons les fases del seu cicle biològic. Hi ha espècies en què els mascles i les femelles només estan junts durant l’època de la reproducció i, un cop acabada aquesta, les femelles i les cries constitueixen petits harems apartats; és el cas del senglar. A la fotografia una femella amb les seves cries, ja crescudes i per tant gairebé desproveïdes de les franges brunes que les caracteritzen en els primers mesos de vida.
Antonio Bea.
L’existència d’una jeraquia s’observa, fins i tot, en grups de ratolins de laboratori (artificialment establerts): no es comporten anàrquicament, sinó amb unes relacions de dominància i subordinació ben establertes. Entre els herbívors, la jerarquització s’estableix de manera que hi ha una ordenació entre els mascles, i aquests dominen les femelles, que també són ordenades jeràrquicament; els joves formen el tercer grup, en el qual hi ha una seriació jeràrquica, per bé que són dominats pels adults, tant pels mascles com per les femelles. Entre els cèrvids, la jerarquia s’estableix per lluita i el mascle dominant és el que presenta la cornamenta més desenvolupada, el més fort físicament; expulsats de l’entorn els altres masces, cobrirà les femelles. Hi ha molts altres exemples clàssics, com el dels babuïns, els ximpanzés o els dofins. En els grups familiars, hi ha també una notable jerarquització que es veu clarament reflectida en el moment en què els seus membres ingereixen el menjar després de la caça. En el cas dels lleons, per exemple, el primer en alimentar-se és el mascle (per aquest motiu es parla de la "part del lleó" quan es fa referència a la part més bona en un repartiment).
Hi ha espècies estrictament solitàries, però, almenys en l’època d’alletament, la femella va amb les cries. Altres espècies viuen en societat de manera permanent, formant grups més o menys importants i ramats que tenen una estructura espacial típica. Ocupen un territori que defensen enfront d’altres individus de la mateixa espècie.
La dimensió d’una població és subjecta a canvis cíclics. Són típics els dels lèmmings, que influeixen en tota la xarxa tròfica: els anys en què es reprodueixen bé també ho fan els seus depredadors. Això ens indica que, malgrat haver-hi una fluctuació poblacional, hi ha un cicle determinat. El nombre d’individus d’una espècie no passa de certs límits; la natura té mecanismes de regulació que limiten la població.
Una organització social ben coneguda és la dels cérvols. A l’hivern, mascles i femelles formen ramats diferents amb un domini vital propi. El més propi sembla ésser el de les femelles, melles, que és on es dona el part; en canvi, el dels mascles es pot considerar un refugi d’hivern. A l’estiu, diferents ramats de cérvols poden reunir-se per pasturar junts i, per tant, perden la seva individualitat. A la tardor, els mascles s’introdueixen en el territori de les femelles i lluiten entre ells per formar un harem. En aquesta època, sembla que els mascles manifestin un domini sobre les femelles, però això sols és així en l’aspecte sexual, ja que, enfront d’un perill, les femelles es reuneixen en un grup compacte conduït pel seu líder femení habitual, tal com s’esdevenia abans de la formació dels harems. L’organització social dels bòvids és més o menys semblant; en canvi, la dels èquids és molt diferent: moltes zebres van en ramats de mascles i femelles, que es desfan a l’època de zel quan s’estructuren les parelles, i no sembla que hi hagi un animal dominant en el ramat. En els ases de Núbia, hi ha ramats de femelles i joves per una banda i de mascles per l’altra. Els mascles només es reuneixen amb les femelles a l’època de reproducció. En el cas dels hemions asiàtics (Equus hemionus), a l’època de zel, els mascles s’asseguren la possesió de tres o quatre femelles amb els seus petits. En els cavalls salvatges, els ramats són conduïts per un animal dominant. Sembla observable una transició des dels grups laxos dirigits de les zebres fins a les societats patriarcals dels cavalls autèntics, passant per les societats matriarcals dels ases i dels hemions.
Pràcticament tots els primats viuen en grups d’elevada complexitat social. La unitat sol ésser una família, composta per un mascle dominant i algunes femelles que formen el seu harem acompanyats de cries i joves sexualment no madurs. Poden haver-hi altres grups com els formats només per mascles. Un grup gran de mones és format per un nombre variable d’associacions menors que acostumen a anar juntes en cerca d’aliment. Els territoris de cada grup coincideixen, però les diverses agrupacions eviten el contacte directe en els seus desplaçaments per terrenys comuns. També hi ha espècies de primats que són estrictament territorials.
La composició d’un grup pel que fa a l’edat i al sexe pot variar en les diferents èpoques de l’any i del cicle reproductor. En això, hi ha una gran diferència segons grups i condicions de vida. El comportament social dels mamífers és, doncs, en conjunt, extraordinàriament variat.
És característic de diversos grups de mamífers la construcció de caus o cataus, galeries subterrànies que ells mateixos excaven i on resideixen habitualment; en tractar les diferents espècies separadament, farem referències concretes a cadascun dels tipus. Altres mamífers construeixen habitacles aeris més o menys sofisticats o, fins i tot, aquàtics, com és el cas del castor.
Les relacions tròfiques
L’alimentació

El règim alimentari dels mamífers és molt variat: pràcticament aprofiten tots els recursos orgànics existents. Si bé d’una manera general podem parlar de mamífers insectívors, carnívors, herbívors i omnívors, val a dir que en molts casos es fa difícil establir una separació neta entre ells, especialment en el camp dels petits mamífers, que sovint introdueixen restes animals en la seva dieta. La figura il·lustra un ratolí de bosc (Apodemus sylvaticus) en actitud de rosegar un bolet, bé que no és aquest un aliment molt freqüent en la seva dieta.
Carme Borràs.
Les necessitats metabòliques dels mamífers fan que, excepte aquelles espècies que poden hivernar, les altres hagin de tenir aliment a la seva disposició durant tot l’any. Normalment, són poc exigents quant a l’aliment, però també és cert que hi ha una sèrie d’espècies molt estenòfagues; el porc formiguer africà, el panda gegant —que menja brots de bambú— i el coala —que menja fulles d’eucaliptus— en són bons exemples. A més, en aquesta darrera espècie, la mare ha de donar defecacions pròpies a la seva descendència per provocar-li una sembra intestinal de bacteris, que fan possible la seva alimentació a partir de les fulles d’eucaliptus. Els artiodàctils remugants presenten unes especialitzacions, que ja hem descrit, en el seu tub digestiu que els permeten la digestió de la cel·lulosa ingerida i el seu aprofitament, gràcies als protozous amb bacteris que presenten en el rumen. Els herbívors no remugants digereixen la cel·lulosa en un cec de mida gran.
La dentició és una de les estructures característiques en els mamífers. Tant el nombre i la forma com la disposició de les peces dentàries, així com l’organització general de la boca, obeeixen a adaptacions relacionades amb el règim alimentari del grup. Els carnívors, als quals correspon el crani de l’esquerra, presenten una dentició heterodonta típica, amb incisives que els serveixen per a esquinçar la presa, ullals amb els quals la maten, i premolars i molars, que els serveixen per a tallar la carn i trencar els ossos; és característica la dent carnissera, una molariforme molt més desenvolupada que les altres (P4 de la mandíbula superior i M1 de la inferior) que actua com una veritable cisalla en tallar la carn. En els herbívors hi ha una disposició totalment diferent (crani de la dreta, d’un remugant): desapareixen les incisives i les canines de la mandíbula superior i les molariformes tendeixen a uniformitzar-se. Entre les incisives i les canines apareix un ampli espai, anomenat diastema, pel qual circula l’herba tallada entre les parts labial i bucal. Els herbívors tenen un moviment mandibular rotatori, de manera que trenquen l’herba per abrasió sobre les corones dentàries i s’estableix una circulació de l’aliment per la boca, gràcies al diastema.
Jordi Vidal
Els fruits són l’aliment de molts mamífers, especialment dels tropicals, com els macroquiròpters i els primats en general. Les llavors són l’aliment exclusiu d’un bon nombre de rosegadors (Dipodomys i Perognathus, del desert del SW dels Estats Units, no ingereixen ni aigua); en canvi, les rates blanques alimentades amb aquestes llavors s’amagreixen i moren al cap de pocs dies. Les escorces són l’aliment de cèrvids, esquirols i ratolins de camp; el porc espí americà es nodreix, a l’hivern, quasi exclusivament d’escorces; l’ant i el castor també s’alimenten d’escorces, principalment a l’hivern; els esquirols s’alimentan també de bolets i sembla que, fins i tot, aprofiten espècies tòxiques per a l’home. Productes tan sorprenents com el pol·len són aprofitats per alguns rats-penats, que intervenen en processos de pol·linització.
Podem considerar que una forma especial d’alimentació és la ingestió de sang, pròpia del quiròpters coneguts —fabulacions literàries a banda— com a vampirs (Desmodus, Diphylla, Diaemus), espècies americanes de petita mida que solen alimentar-se de la sang del bestiar: quan mosseguen i xuclen la sang, hom se n’adona, malgrat que la mosegada és extraordinàriament fina i delicada, perquè l’animal perd força. En l’home, la mossegada més freqüent sol ésser en els dits dels peus, deixats inadvertidament al descobert mentre hom dorm.
L’alimentació càrnia és característica de grups complets com els cànids i els fèlids; les grans espècies cacen, sobretot, ungulats, mentre que les petites es conformen amb preses més modestes, segons la ja clàssica relació depredador-presa. La caça es pot fer a l’aguait o perseguint la presa; algunes vegades, com fa —a l’hivern— el llop, diversos individus col·laboren en la persecució. Alguns mamífers s’alimenten de restes alimentàries deixades per grans caçadors. Es diu, i no deixa d’ésser cert, que les hienes s’alimenten de les restes dels grans banquets dels lleons, però moltes vegades s’han de conformar amb els petits mamífers que agafen; d’altra banda, els grans mamífers no acostumen a deixar abandonades grans exquisideses. Hi ha dades concretes que un lleopard enfilà 60 kg de carn d’un jove búfal a les branques d’una acàcia a més de 4 m del sòl per fer una bona reserva d’aliment. És molt coneguda l’afecció de les mangostes per les serps; no són molt resistents al verí dels ofidis, però coneixen molt bé la seva distància efectiva d’atac i no entren en el cercle d’acció de les serps si no és per matar-les, quan aquestes, nervioses, han errat diverses vegades en els seus intents d’atac. Els eriçons s’alimenten d’escurçons. Els peixos són la font d’aliment de les llúdries, els gats pescadors i els rats-penats pescaires. És molt típic que l’ós aprofiti el viatge migratori dels salmons per capturar-los i alimentar-se d’ells. També, com a recurs desesperat, els joves fèlids, quan encara no tenen prou pràctica de caçar grans preses, recorren als peixos per alimentar-se. Els insectes constitueixen una presa típica de molts mamífers, com tots els insectívors i molts rats-penats. És l’aliment complementari d’alguns primats i algunes espècies se n’alimenten exclusivament, com els pangolins, els formiguers i els porcs formiguers.
El plàncton, especialment els crustacis, és la font d’aliment de les balenes, que, gràcies a aparells especials de filtració —que ja hem descrit—, en explotar un recurs tan abundós i poder-se saltar molts graus de l’escala tròfica, poden arribar a tenir grans dimensions.
Un gran nombre de mamífers són molt eurífags. Molts rosegadors, els úrsids i el mateix home són omnívors, la qual cosa, evidentment, és un avantatge enfront d’unes condicions variables del medi.
Els recursos defensius
Els mamífers són preses d’altres mamífers o, fins i tot, d’altres classes de vertebrats. Hi ha amfibis anurs capaços d’ingerir petits mamífers; alguns rèptils i molts ocells també els depreden. La majoria dels mamífers fugen davant d’un enemic, però n’hi ha que romanen en la immobilitat més absoluta; els mateixos cérvols, en els primers dies de vida, es posen contra el terra per passar desapercebuts. Per entendre aquest comportament, cal pensar que l’estímul de caça que es desencadena en el depredador el dona el moviment de la presa (fins i tot l’home caçador no acostuma a disparar la seva escopeta si la presa no és en moviment). No obstant això, la majoria no fugen davant l’enemic fins que aquest no arriba a certa distància, l’anomenada distància de fugida. Per això, els depredadors sols acostumen a caçar els animals dèbils o malalts, la qual cosa no vol dir que quan aquests ja no poden escapar del depredador se’n defensin com puguin.
L’existència de depredadors també ens fa entendre determinades associacions, com la dels estruços amb alguns grans herbívors africans. L’extraordinària vista dels ocells, en aquest cas dels estruços, fa que detectin abans la presència del deprededor, que, amb el seu nerviosisme, posa alerta tot el ramat mixt; d’altra banda, el moviment constant dels herbívors posa al descobert els petits mamífers de què s’alimenten els estruços. Els mamífers que viuen en grups, estan, ordinàriament, molt ben organitzats per a detectar un eventual perill; un exemple clàssic és el dels bous mesquers, que es disposen en cercle, tancant al mig els individus joves, quan se senten amenaçats i fan front a l’enemic amb les seves fortes defenses. En altres casos, pot no haver-hi, pròpiament, un únic animal destinat a la vigilància, que esdevé una funció col·lectiva. Això es dona en les marmotes dels Alps, en les quals l’existència d’un perill, moltes vegades una àliga que les sobrevola, és avisada per la primera que el detecta amb un xiulet.
Alguns mamífers, proveïts d’una armadura completa o bé de fortes formacions defensives, es fan una bola, de manera que els possibles depredadors han de desistir de donarlos caça. Una altra manera de fer desistir el caçador és enviar-li una forta descàrrega pudenta de les glàndules anals, acte conegut i típic de les mofetes. Algunes espècies de rosegadors deixen fàcilment la beina de la seva cua i aconsegueixen fugir del possible deprededor; posteriorment, els cau l’eix ossi, la columna vertebral, que havia quedat al descobert. En altres casos, la capa del pèl és homocròmica respecte al medi en què viuen. Aquest fenomen és típic de molts mamífers àrtics —en els quals el pèl és o es torna blanc a l’hivern— i d’aquells que viuen en zones altes i, per tant, amb hiverns rigorosos. S’interpreta de diferent manera, encara que la causa fisiològica de l’acció dels repressors sigui la mateixa. Si són rosegadors passen més desapercebuts als depredadors i, en canvi, si són carnívors, poden aproximar-se més fàcilment a la presa. Hom pot interpretar-ho com una conseqüència comportamental de la falta de síntesi del pigment.